 Ana Paula Daga1*
Ana Paula Daga1* Vanessa Lumi Koga2
Vanessa Lumi Koga2 João Gabriel Material Soncini1Caroline Martins de Matos1
João Gabriel Material Soncini1Caroline Martins de Matos1 Marcia Regina Eches Perugini1Marsileni Pelisson1
Marcia Regina Eches Perugini1Marsileni Pelisson1 Renata Katsuko T. Kobayashi2Eliana Carolina Vespero1
Renata Katsuko T. Kobayashi2Eliana Carolina Vespero1- 1Laboratory of Clinical Microbiology, Department of Pathology, Clinical and Toxicological Analysis, Health Sciences Center, State University of Londrina, Londrina, Brazil
- 2Laboratory of Microbiology, Department of Microbiology, State University of Londrina, Londrina, Brazil
Extraintestinal pathogenic Escherichia coli (ExPEC) isolates are responsible for many bloodstream infections. The aim of this study was to characterize E. coli isolated from the bloodstreams of patients (n = 48) at the University Hospital in Brazil. Epidemiological data were obtained through the analysis of medical records and laboratory tests. By PCR analysis, we investigated the presence of virulence factors (VFs), pathogenicity islands (PAIs), extended-spectrum β-lactamase (ESBL), phylogenetic classifications (A, B1, B2, C, D, E, and F) and molecular genotype by enterobacterial repetitive intergenic consensus-polymerase chain reaction (ERIC-PCR). The mortality analysis showed that 33.3% of the deaths were associated with bacteraemia due to E. coli infections; in addition, an age between 60 and 75 years (p < 0.001; OR = 6.3[2.1–18.9]) and bacteraemia with an abdominal origin (p = 0.02; OR = 5[1.2–20.5]) were risk factors for the severity of the infection. Additionally, the presence of the afa gene was associated with mortality due to E. coli bacteraemia (p = 0.027; OR = 11.4[1.5–85.7]). Immunosuppression (27.1%), intestinal diseases (25.0%) and diabetes (18.8%), were prevalent among patients, and most of the bacteraemia cases were secondary to urinary tract infections (50.0%). The serum resistance gene traT was present in 77.1% of isolates, group capsular 2 (kpsMT II) was present in 45.8% and the K5 capsule was present in 20.8% of isolates. The isolates also showed a high prevalence for the siderophore yersiniabactina (fyuA) (70.8%) and PAI IV536 (77.1%). Phylogenetic analysis showed that group B2 (45.8%) was the most prevalent, and was the phylogroup that had a higher prevalence of VFs and PAIs. However, in this study, a considerable number of isolated bacteria were classified as group B1 (18.8%) and as group E (14.6%). Eight (16.7%) isolates were resistant to third and fourth generation cephalosporin and group CTX-M-1 (CTX-M-15) was the most prevalent ESBL type. The molecular genotyping showed two clonal lineages and several isolates that were not related to each other. This study provides additional information on the epidemiological and molecular characteristics of E. coli bloodstream infections in Brazil.
Introduction
Escherichia coli is the gram-negative organism most frequently isolated in adult patients with bacteraemia (Mora-Rillo et al., 2015) and in severe cases it may lead to death (Owrangi et al., 2018). The rates of bacteraemia have increased steadily in recent years (De Kraker et al., 2013; Miajlovic et al., 2016). In general, E. coli are a part of the normal commensal gut microbiota of healthy human populations (Köhler and Dobrindt, 2011; Micenková et al., 2017). However, some strains can cause intestinal or extraintestinal infections due to specific virulence factors (VFs) (Burdet et al., 2014; Usein et al., 2016).
Isolates that are capable of gaining access to and surviving in the bloodstream are known as extraintestinal pathogenic E. coli (ExPEC) (Russo and Johnson, 2003) and cause a variety of infections, including urinary tract infections (UTI), sepsis, and neonatal meningitis (Ron, 2010; Mora-Rillo et al., 2015). The most common extra-intestinal site colonized by these bacteria is the urinary tract, which in turn, is a common source for bloodstream infections (Micenková et al., 2017).
According to the phylogenetic classification, E. coli was divided into 7 groups (A, B1, B2, C, D, E, and F). The pathogenic groups of the ExPEC strains generally belong to the groups B2 and D, and the commensal strains that survive in the intestines, i.e., non-pathogenic strains, are generally included in groups A or B1 (Clermont et al., 2013).
ExPEC strains have several virulence factors (VFs) that may play a role in infection by enabling the bacterial cells to colonize the host and disseminate. VFs are either encoded on the bacterial chromosome, where they are usually located within pathogenicity islands (PAIs), or on plasmids (Dale and Woodford, 2015); these VFs include adhesion molecules, iron acquisition systems, host defense-subverting mechanisms, and toxins. Several VFs have been associated with bloodstream infections (Lefort et al., 2011; Mora-Rillo et al., 2015). However, predictions of the initial severity and outcome based on bacterial VFs alone are not entirely accurate, and the consideration of host determinants, including underlying diseases, facilitates such predictions (Jauréguy et al., 2007).
When the infection occurs, the host immune system responds to eliminate the infectious agents (Diacovich and Gorvel, 2010). The progression of infection is mainly associated with the bacterial capability to survive such defenses. In addition, bacterial virulence properties influence the severity and extent of the infection (Lefort et al., 2011; Owrangi et al., 2018). The presence of E. coli in the bloodstream can result in the induction of a vigorous host inflammatory response that lead to sepsis, which is associated with high morbidity and mortality (Russo and Johnson, 2003; Miajlovic and Smith, 2014).
In addition, the rates of multidrug resistant (MDR) E. coli infections are on the rise. In particular, the predominant ExPEC global lineage sequence type (ST) 131 is frequently associated with fluoroquinolone resistance and the production of extended-spectrum β-lactamases (ESBLs) (Shaik et al., 2017), and the treatment of infections caused by ExPEC has become very challenging due to the emergence of resistance to the first-line and the last-resort antibiotics (Usein et al., 2016; Shaik et al., 2017).
In developed countries, E. coli is a very important pathogen in infections of the bloodstream. According to Laupland (2013), E. coli was ranked first or second in the incidence of bloodstream infections in countries such as Australia, Canada, Denmark, Finland, Iceland, New Zealand, Sweden, and the USA. However, analyzing the frequency of E. coli bloodstream infection at the University Hospital in Brazil, showed that in recent years, E. coli was responsible for 4.7% of bloodstream infections. Other studies also showed a low prevalence of E. coli bloodstream infections in Brazilian hospitals (Marra et al., 2011; Yokota et al., 2016; Taveira et al., 2017). For this reason, there has been little research regarding E. coli bloodstream infection in Brazil. However, knowledge of the impact of VFs, antimicrobial resistance, and host determinants on the severity of E. coli bloodstream infections is important for the determination of the epidemiological profiles of bloodstream infections; this could provide targets for specific intervention in the future. The aim of this study was to characterize the VFs, antimicrobial sensitivity profile, and epidemiological data of E. coli isolates from the blood of patients at the University Hospital of Londrina.
Materials and Methods
Bacterial Isolates
This study included 48 E. coli samples that were isolated from the bloodstreams of patients admitted to the University Hospital of Londrina from 2015 to 2017. Haemoculture positive samples were detected by BACTEC™ FX (Becton Dickinson, USA), and aetiological agents were previously identified by the VITEK® 2 system (bioMérieux, USA). Following identification, the isolates were kept in Tryptic Soy Broth with 15% glycerol (−20°C).
Collected Data
Demographic data were collected from all patients, including age, gender, and comorbidities of medical records provided for consultation by the Medical Archive and Statistics Service (SAME) of the University Hospital of Londrina. The study was approved by the Ethics and Research Committee of the State University of Londrina CAAE 43013315.8.0000.5231.
Antimicrobial Susceptibility
For the identification of the isolates, the VITEK® 2 GN ID card and the VITEK® 2 AST 239 card were used for antibiogram analyses, which was complemented with the diffusion disk method. All cards are from bioMérieux. The interpretation was performed according to the CLSI 2017 (Clinical and Laboratory Standards Institute) criteria.
β-Lactamase Characterization
The detection of the blaCTX−M−1, blaCTX−M−2, blaCTX−M−8, blaCTX−M−9, blaCTX−M−25 genes was performed by multiplex PCR, as described by Woodford et al. (2006) and blaCTX−M−15 according to Leflon-Guibout et al. (2004). The PCR reaction was enhanced using the TopTaq® Master Mix Kit (QIAGEN). PCR for the screening of the blaKPC gene was performed according to Bradford et al. (2004) (Supplementary Material).
Phylogenetic Classification
The E. coli isolates were classified into 7 phylogenetic groups (A, B1, B2, C, D, E, and F), based on the presence of the genes chuA, yjaA, arpA, and trpA, and a DNA fragment (TSPE4.C2), which were detected by the PCR method (Clermont et al., 2013) (Supplementary Material).
Detection of Virulence Factor Genes
The genotypic identification of the main virulence factors in ExPEC was performed using the PCR method. The genes selected are the most frequent in ExPEC and included haemolysins (hlyA and hlyF), cytotoxic necrotizing factors (cnf 1 and cnf 2), colicin V (cvaC), aerobactin (iutA), yersiniabactin (fyuA), salmochelin (iroN), type 1 fimbrial adhesin (fimH), P-fimbriae (papC and papG), S-fimbrial adhesin (sfaA and sfaS), afimbrial adhesin (afa), serum resistance (iss and traT), brain microvascular endothelium invasion (ibeA), capsules (kpsMT K1, kpsMT K5, kpsMT II, and kpsMT III), and an outer membrane protein (ompT) (Johnson and Stell, 2000; Koga et al., 2014) (Supplementary Material).
Detection of Pathogenicity Islands
The presence of sequences associated with seven different PAIs, previously characterized in uropathogenic E. coli, was determined (PAI I536, II536, III536, IV536, ICFT073, IICFT073, IJ96, and IIJ96) (Sabaté et al., 2006; Koga et al., 2014) (Supplementary Material).
Molecular Genotyping
Enterobacterial repetitive intergenic consensus (ERIC-PCR) was performed as previously described by Versalovic et al. (1991). Analysis of genomic fingerprinting was performed using GelJ v.2.0 software by the Dice similarity method (Heras et al., 2015). Strains were considered genetically related if the similarity index was ≥85%.
Statistical Analysis
Categorical data are shown frequencies and percentages. The analysis was performed by logistic regression, associated to the selection of stepwise variables, Nagelkerke test to determine the value of Pseudo-R2, and Odds Ratio test associated with the Likelihood test. In addition, the Odds Ratio test associated with the Chi-square test was used. The alpha significance level was 0.05. Data analysis was performed using Statistical Package for Social Sciences (SPSS—IBM Corp., New York, USA), version 20 for Windows.
Results
Demographic and Clinical Characteristics
The median age of the 48 patients was 63 years old (range 0–90 years) and 56.2% were male. Twenty-three (47.9%) were aged over 65 years and 4 (8.3%) of patients were aged under 1 year. In this study, the most prevalent comorbidity was systemic arterial hypertension (43.8%), followed by intestinal disease (25.0%), diabetes (18.8%), coronary disease, solid tumor, and renal failure at (12.5%) each, hematological malignancy and cirrhosis at 10.4% each, immunologic disease and viral hepatitis at 8.3% each, and HIV at 6.3% (Table 1).
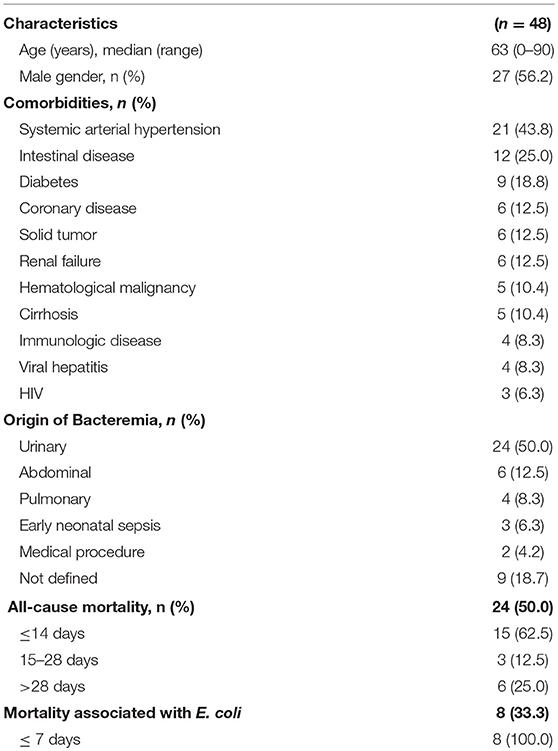
Table 1. Characteristics of patients with Escherichia coli bacteremia.
According to the data from the medical records and laboratory data, 24 (50.0%) patients had bacteraemia from a urinary origin, 6 (12.5%) had abdominal sepsis, 4 (8.3%) had pulmonary sepsis, 3 (6,3%) had early-onset neonatal sepsis, 2 (4.2%) had bacteraemia after a medical procedure, one patient had bacteraemia after amputation of the lower limb, another patient had bacteraemia after a prostate biopsy, and 9 (18.7%) cases of bacteraemia did not have the origin defined. Of the 48 patients studied, half (50.0%) died, and 8 (33.3%) of these patient deaths were related to E. coli bloodstream infections (Table 1).
In this study, the univariate statistical analysis showed that ages between 60 and 75 years old (p < 0.001; OR = 6.3[2.1–18.9]) and abdominal origin bacteraemia (p = 0.02; OR = 5[1.2–20.5]) were risk factors for the severity of the infection (Table 2).
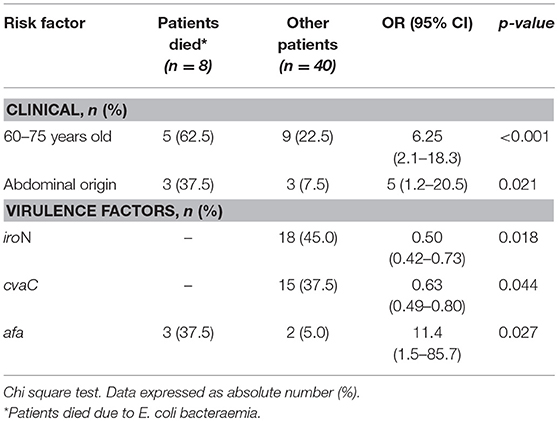
Table 2. Risk factor death from E. coli bacteremia identified by univariate analyses.
Antibiotic Susceptibility
The E. coli isolates exhibited a high rate of resistance to ampicillin (64.6%), ampicillin-sulbactam (56.3%), sulfamethoxazole-trimethoprim (45.8%), and ciprofloxacin (35.4%). Nine (18.8%) isolates were resistant to gentamicin, 1 (2.1%) was resistant to amikacin, 7 (14.6%) were resistant to piperacillin-tazobactam and only 1 (2.1%) isolate was resistant to carbapenems and was positive for blaKPC (Figure 1).

Figure 1. ERIC-PCR molecular fingerprint profiles of 48 Escherichia coli bloodstream infection isolates, virulence determinants and antimicrobial resistance profiles. Amikacin (AMK), gentamicin (GEN), ampicillin (AMP), ampicillin-sulbactam (SAN), aztreonam (ATM), cephalothin (CEP), cefuroxime (CXM), cefoxitin (FOX), ceftazidime (CAZ), ceftriaxone (CRO), cefepime (FEP), imipenem (IMP), meropem (MEM), ertapenem (ERT), piperacillin-tazobactan (PTZ), ciprofloxacin (CIP), and sulfamethoxazole.
The CTX-M enzymes are important β-lactamases described in worldwide. In this study, 48 isolates 8 (16.7%) were resistant to third and fourth generation cephalosporin and were producing ESBL type CTX-M. Seven (14.6%) isolates were positive for CTX-M-1, and five isolates were positive for CTX-M-15. Other variants were also found, including CTX-M-9, CTX-M-2, and CTX-M-25. All ExPEC-producing CTX-M showed a combination of more than one enzyme except for 1 isolate (Figure 1).
Phylogenetic Analysis
The phylogenetic group B2 (45.8%) was predominant among the ExPEC strains. Groups B1 (18.8%), E (14.6%), F (6.3%), A (4.2%), and D (2.1%) exhibited the lowest frequencies, and 4 (8.2%) isolates were not grouped.
Distribution of Virulence Factors and Pathogenicity Islands
In the present study, the most predominant virulence genes were fimH (95.8%), traT (77.1%), fyuA (70.8%), iutA (64.3%), kpsMT II (45.8%), iroN (37.5%), papC (35.4%), cvaC (31.3%), iss (20.8%), kpsMT K5 (20.8%), ompT (20.8%), hlyF (20.8%), hlyA (14.6%), cnf 1 (12.5%), afa (10.4%), kpsMT K1 (8.3%), sfaS (6.3%), sfaA (4.2%), ibeA (4.2%), and kpsMT III (2.1%). No isolates were positive for the genes papG and cnf 2 (Table 3).
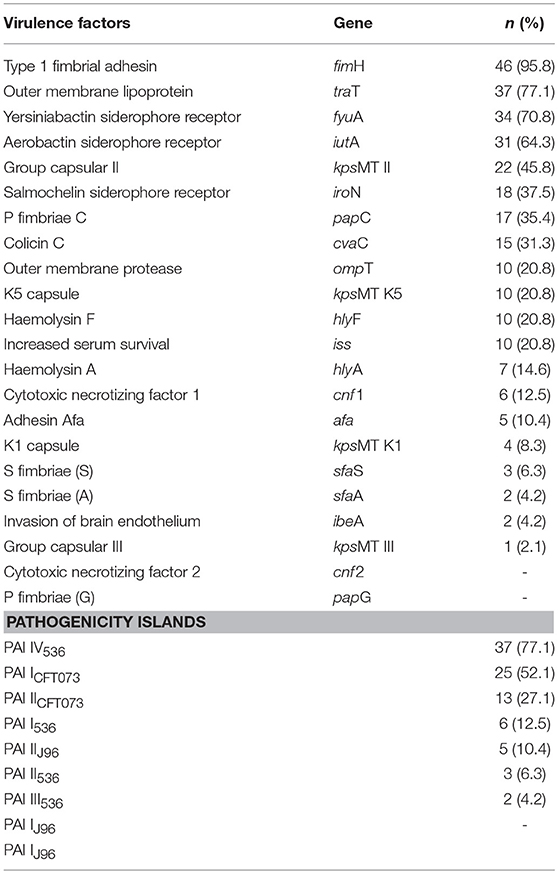
Table 3. Prevalence of virulence factors, islands of pathogenicity of 48 blood Escherichia coli isolates, 2015–2018.
The presence of the afa gene was associated with mortality due to bacteraemia caused by E. coli (p = 0.027; OR = 11.4[1.5–85.7]). This is unlike the iroN (p = 0.018; OR = 0.50[0.42–0.73]) and cvaC genes (p = 0.044; OR = 0.63[0.49–0.80]), which were not present in the same isolates (Table 2).
Ten (20.8%) isolates were positive for the cvaC, hlyF, iroN, iss, and ompT genes. These genes have been described as the conserved virulence plasmidic (CVP) region (having 8 genes or operons: iro, iuc, sit, ompTp, cva, hlyF, ets, and iss). Six of these isolates were classified into 4 different phylogenetic groups (B2, B1, F, and E), and two isolates were not grouped.
The most prevalent PAI was PAI IV536 (77.1%); this was followed by PAI ICFT073 (52.1%), PAI IICFT073 (27.1%), PAI I536 (12.5%), PAI IIJ96 (10.4%), PAI II536 (6.3%), and PAI III536 (4.2%). PAI IJ96 was not detected in this study (Table 2). The bloodstream isolates contained at least one of the investigated PAIs, and the most frequent combination pattern was related to those strains with PAI IV536 and PAI ICFT073 (47.9%). No strain presented all the PAIs.
Comparison of Virulence Factors and Pathogenicity Islands With Phylogroups
The phylogenetic group B2 had the highest frequency regarding both the virulence genes and PAI markers compared those in the other phylogenetic groups. In this investigation, there was a significant association between the phylogenetic group B2 and the presence of virulence genes such as papC, fyuA, kpsMT K1, kpsMT K5, kpsMT II, hlyA, and cnf 1 (Table 4).
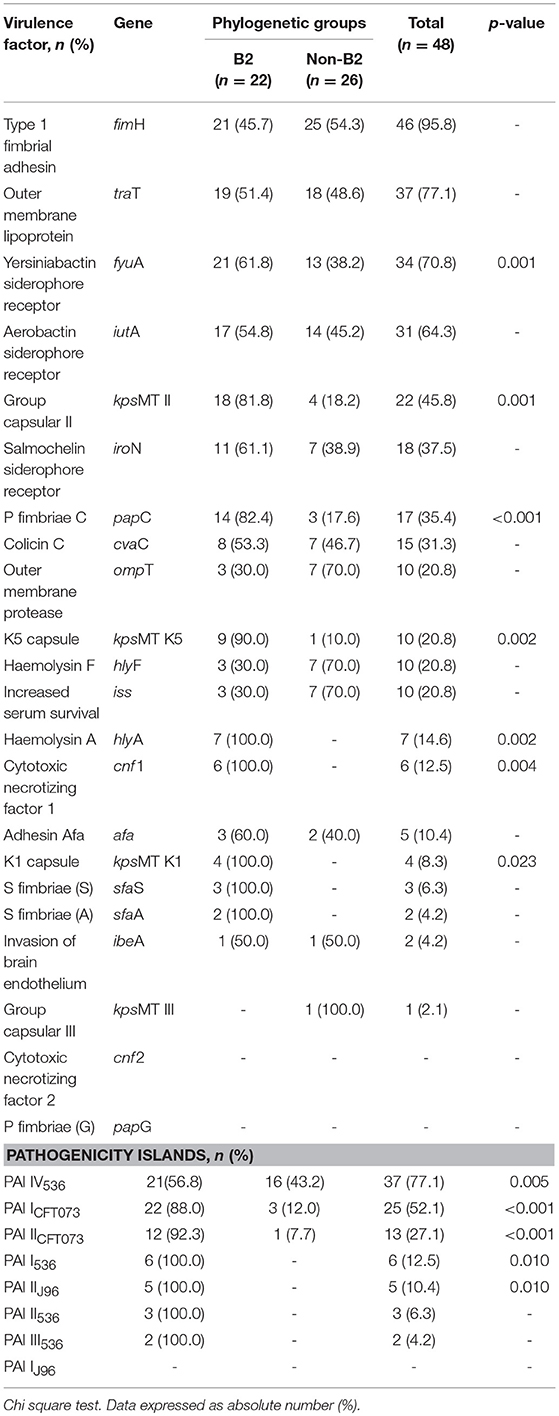
Table 4. Phylogenetic group relation with virulence factors and islands of pathogenicity of 48 blood Escherichia coli isolates, 2015–2018.
An increased presence of PAI markers was observed in the phylogenetic group B2, while the lowest presence of PAI markers was related to the other phylogroups. The high frequency of the B2 group was statistically significant in PAI I536, PAI IV536, PAI ICFT073, PAI IICFT073, and PAI IIJ96 (Table 4).
Genetic Correlation Between E. coli Blood Strains
Analysis of molecular genotyping using the ERIC-PCR technique demonstrated the presence of two major clonal lineages and several isolates that were not related to each other (Figure 1). In the data analysis, a cutoff of 85% was used due to the high diversity of the isolates. A more detailed analysis verified that the two lineages were significantly different in relation to the antimicrobial resistance profile (Table 5). The differences were significant for ESBL production and for the presence of the blaCTX−M−1, blaCTX−M−15, and blaCTX−M−9 genes. In addition, the two clonal groups showed significant differences in a single VF, the kpsMT K5 gene. The most prevalent phylogenetic group remained the B2 group in the two clonal groups, and presented a similar prevalence in the other phylogroups (Table 6).
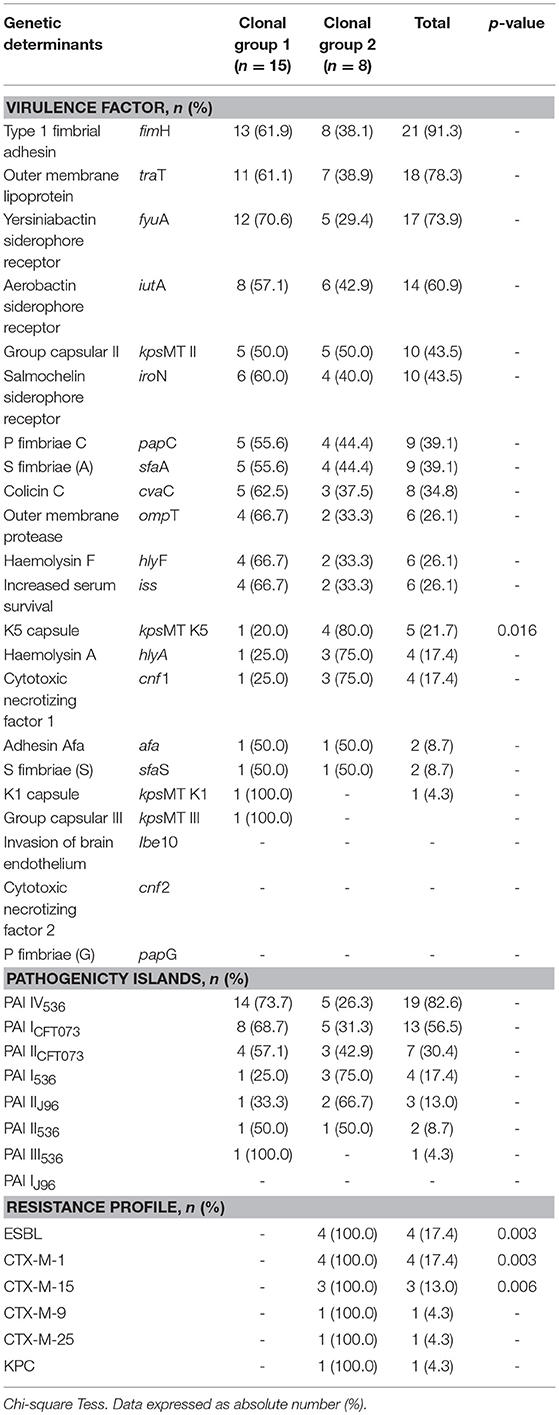
Table 5. Comparison of virulence factors, pathogenicity islands and resistance profile between two clonal groups.
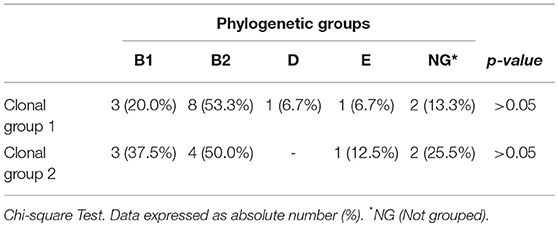
Table 6. Phylogenetic classification among isolates belonging to clonal groups.
Discussion
Demographic and Clinical Characteristics
ExPEC is a very important bacterium that is involved in bloodstream infections, and in severe cases, it may lead to death (Owrangi et al., 2018). For the characterization of this infection, it is necessary to consider the bacterial determinants, such as virulence factors, as well as antimicrobial susceptibility and host determinants (Jauréguy et al., 2007; Lefort et al., 2011). The understanding and characterization of these factors may facilitate the development of new strategies for the treatment of E. coli bloodstream infections.
The presence of intestinal diseases was very prevalent among patients (25.0%). It is possible for ExPEC to initiate systemic infections by causing sepsis or bacteraemia through abdominal translocation in patients with some intestinal disease (Vaishnavi, 2013). Diabetes was also a common comorbidity and was observed with an 18.8% prevalence. Studies have reported an association between diabetes and pyelonephritis by E. coli and the increased risk of complications such as renal abscess and necrosis (Alves et al., 2012).
These infections usually occur as complications of infections of the urinary or gastrointestinal tract or as complications of other infections (Micenková et al., 2017). In this study, 24 (50.0%) of the bloodstream infections had a urinary origin and 6 (12.5%) had an abdominal origin. Five of 6 patients with bactaeremia of abdominal origin had some abdominal disease and one had hepatic cirrhosis. Patients with hepatic cirrhosis had a risk of bacteraemia due to an altered immune system and an increased bacterial translocation (Lefort et al., 2011), and 8 of the 9 patients with diabetes had bacteraemia of a urinary origin.
The mortality associated with bacteraemia due to E. coli was 33.3%. According to Russo and Johnson (2003), E. coli bacteraemia has a case-fatality rate of 5 to 30% and represents an increasingly important endemic problem, accounting for hundreds of thousands of lives lost and billions of health care dollars spent each year. In agreement with the results of Lefort et al. (2011), a patient age between 60 and 75 years old (p < 0.001) was considered a risk factor for mortality. Additionally, as in this study, Lefort et al. (2011) found that the rate of death was higher among those whose sepsis originated from the digestive tract than it was for those whose sepsis originated from the urinary tract.
Phylogenetic Classification, Virulence Factors and Pathogenicity Islands Distribution
Bacterial determinants were also important to characterize bacteraemia. The phylogenetic classification of ExPEC isolates showed that B2 (45.8%) and B1 (18.8%) were the most prevalent groups. In the literature, B2 was reported to be the most common phylogenetic group in ExPEC (Koga et al., 2014; Micenková et al., 2017). The presence of B1 isolates in bloodstream infections shows that this group is also capable of causing systemic infections; however, literature reports have stated that isolates that belong to the groups A and B1 are more often strictly commensal strains from the intestinal microbiota (Skjøt-Rasmussen et al., 2012; Usein et al., 2016).
According to Fratamico et al. (2016), groups B2 and D have a higher virulence in humans that allows them to induce extraintestinal infections in both healthy and immunocompromised hosts. However, in this study, a lower prevalence of group D was found. The low prevalence of E. coli from group D can be explained because in the old classification of Clermont et al. (2000), there was no distinction between groups D, E and F, and all strains were classified as group D.
One of the mechanisms responsible for the survival of E. coli strains in the bloodstream is the escape of the complement system through serum resistance (Miajlovic and Smith, 2014; Micenková et al., 2017). Multiple virulence factors have been shown to be involved in serum survival. In this study, 41 (85.4%) isolates of E. coli had one or more genes related to serum resistance, and the traT gene presented a high prevalence among isolates (77.1%). Similar results were found in studies on E. coli bloodstream infections (Koga et al., 2014; Miajlovic et al., 2016; Bozcal et al., 2018). Among the capsules researched, group 2 (kpsMT II) was the most prevalent (45.8%) and encodes the K1 and K5 capsules (Russo et al., 1998); in addition, 10 (20.8%) isolates presented the K5-specific capsule. In total, 23 (47.9%) of the E. coli isolates showed at least one capsule.
These polysaccharide antigens have been shown to be important for resistance to host immune responses in systemic infections (Miajlovic and Smith, 2014). K5 is involved in the pathogenesis of extraintestinal infections, such as bacteraemia, urinary tract infections, and neonatal meningitis (Blundell et al., 2009). In addition, the K5 capsule confers resistance to both the human innate and adaptive immune system. The chemical structure is similar to heparan sulfate and is probably what contributes to the ability to escape from the immune system, thus, conferring high virulence to the E coli isolates (Blundell et al., 2009).
Iron is a necessary element in virtually all microorganisms and is utilized to catalyze a wide variety of indispensable enzymatic reactions, but iron is also essential to host cells. However, the affinities of bacterial siderophores to iron are generally much higher than those of host proteins/molecules, allowing pathogens to outcompete the host in iron acquisition (Wilson et al., 2017). These siderophore systems are commonly associated with ExPEC strains and those isolated from bacteraemia (Koga et al., 2014; Bozcal et al., 2018). In the present study, yersiniabactin (fyuA) was the most prevalent, and was present in 70.8% of the isolates tested. The fyuA gene product is involved in the efficient uptake of iron from the bloodstream (Ananias and Yano, 2008) and in invasion of the bloodstream from the urinary tract (Johnson and Stell, 2000). Lefort et al. (2011) reported results regarding fyuA and mortality, and found that fyuA was present more frequently in survivors (78.09 vs. 66.18%, p = 0.0025).
In this study, genes related to adhesins were researched and included type 1 fimbria (fimH), P fimbria (papC and papG), Dr binding adhesin (afa), and S fimbria (sfaA and sfaS). The most prevalent was fimH (95.8%). FimH is a crucial factor for the virulence of uropathogenic E. coli strains by mediating adhesion to uroepithelial proteins (Dale and Woodford, 2015) and bacteraemia usually occurs as a complication of an infections in urinary tract (Laupland, 2013). The second most prevalent adhesin gene was papC, which is also involved in urinary tract infections and subsequent bacteraemia (Lefort et al., 2011; Subashchandrabose and Mobley, 2015). Ananias and Yano (2008) reported that the association of papC with fyuA could be the minimal prerequisite for bacterial passage from a renal focus of infection into the bloodstream of non-compromised patients, attached to kpsMT II or to another capsule or protectin.
The genes related to toxin and haemolysin production were researched and included cnf 2, cnf 1, hlyA, hlyF, cvaC, and ibeA. The cvaC and hlyA genes are frequently detected in ExPEC strains (Koga et al., 2014). The prevalence of colicin V (cvaC) was 31.3%. Its role in the pathogenesis of bloodstream infections is not well-understood, but cvaC inhibits the cell growth of other bacteria by, reducing competition for nutrients in conditions of scarcity or stress (Gérard et al., 2005); in addition, cvaC is present on the ColV plasmid, which is known to encode the aerobactin iron uptake system as well as serum resistance factors (Mokady et al., 2005). According to Nedialkova et al. (2014), colicin production is a common trait in E. coli populations and on average, 30% of natural E. coli populations produce one or more colicins.
The toxin α-haemolysin (hlyA) was present in 14.6% of isolates. This toxin incites a noxious inflammatory response, thereby causing clinical disease. The E. coli protein HlyA is a pore-forming bacterial exotoxin that may contribute to the virulence of bacteria during bloodstream infections and sepsis (Sonnen and Henneke, 2013). Bonacorsi et al. (2006) found that in infants younger than 90 days, the presence of hlyA and/or iroN, similar to the presence of hlyA and/or antigen K1, was associated with strains causing urinary tract infections that lead to bacteraemia.
Virulence genes are mainly organized in large clusters called PAIs or are plasmids that have integrated into the genome and, by horizontal gene transfer, explain the notable plasticity of the E. coli genome (Phillips-Houlbracq et al., 2018). Thirty-nine (81.3%) of the isolates were positive for one or more PAI, and the most prevalent was PAI IV536 (77.1%). This PAI is considered an island of high pathogenicity and is associated with the synthesis and transport of the yersiniabactin siderophore, one of the main bacterial iron uptake systems (Dobrindt et al., 2002). This information was confirmed by the presence of the fyuA gene in 70.8% of isolates, and all the isolates that were positive for fyuA were also positive for PAI IV536. PAIs were found primarily in the strains of uropathogenic E. coli, and the high prevalence among the E. coli strains isolated from bloodstream infections can be associated with the fact that urinary tract infections are very common type of infection, and bacteraemia is often a complication of this infection.
Ten (20.8%) isolates were positivite for the cvaC, hlyF, iroN, iss, and ompT genes. The presence of these genes and of the genes encoding salmochelin, aerobactin, and the iron–uptake system SitABC was a signature of conserved virulence plasmidic (CVP) (Lemaître et al., 2013). The phylogenetic classification of these isolates showed that commensal strains also have virulence determinates that cause extraintestinal infections. Only 3 out of the 10 isolates were classified in the B2 group.
Our results demonstrate that the group B2 had the highest frequency regarding both virulence genes and PAI markers compared to those in the non-B2 group. Three of the 4 polysaccharide capsules (kpsMT K1, kpsMT K5, and kpsMT II) studied were more prevalent in the isolates of group B2. The genes papC, fyuA, hlyA, and cnf 1 were also statistically more prevalent in group B2 isolates. Likewise, 5 out of the 8 PAIs studied were statistically more prevalent in the isolates of group B2. These results represent the ability of group B2 isolates to cause extraintestinal infections, such as infections of the bloodstream (Johnson and Russo, 2002; Köhler and Dobrindt, 2011).
Molecular Genotyping and Resistance Profile
The analysis of molecular genotyping demonstrated the presence of two clonal lineages and several isolates not related to each other. A lineage composed of 8 isolates, of which, 7 were positive for the presence of the blaCTX−M gene, and another lineage composed of 15 non-ESBL producing isolates. The results show no significant difference between the two strains in relation to the other antimicrobials tested or in relation to virulence determinates, except for KpsMT K5 which was more prevalent in the clonal lineage that presented a resistance profile to β-lactams. However, there was no relationship between clonality and clinical outcome (mortality due to bacteraemia caused by E. coli).
In this study, 16.7% (n = 8) of isolates were ESBL producers and were positive for the presence of the blaCTXM gene. CTX-M enzymes have become the predominant ESBLs encountered in the clinic, and the rapid dissemination of ESBL is an alarming trend and is considered to be one of the world's main health threats (Yair and Gophna, 2018). It was previously reported that CTX-M-producing E. coli isolates often carry resistance to additional antibiotic classes, which can include co-trimoxazole, aminoglycosides, and fluoroquinolones (Pitout and Laupland, 2008). Six of the 8 isolates showed resistance to sulfamethoxazole-trimethoprim and aminoglycosides, and all isolates showed resistance to ciprofloxacin.
The prevalence of antimicrobial resistance is lower among strains belonging to the B2 phylogenetic group, suggesting a trade-off between resistance and virulence (Clermont et al., 2008). Our results show that only 2 of the 22 isolates classified into group B2 were ESBL producers and that 9 of 14 isolates, sensitive to all antibiotics, belonged to group B2. The rate of E. coli isolation from the bloodstream and the production of ESBL is quite variable. Miajlovic et al. (2016) found a prevalence of 40% of ESBL producers in Ireland; however, Burdet et al. (2014) and Namikawa et al. (2017) both found a lower prevalence: at 3.6% and at <10.0%, respectively. In Brazil, Seki et al. (2013) found a prevalence of 15% of ESBL producers, and 55% were producers of CTX-M-15. We found a similar prevalence: 16.6% were producers of ESBLs and 62.5% were producer of CTX-M-15.
In summary, these results indicate that the bloodstream infection process can be mediated by several alternative VFs, and each strain may have a unique combination of these factors. This variety of virulence genes can be explained by several genetic factors that contribute to genomic plasticity, such as plasmids, phages, and transposable elements. Multiple VFs were found however most of them in relatively small number of isolates, thus, the study did not have enough power to determine their effect on mortality. Although the phylogenetic group B2 corresponds with the virulence potential, our results show that isolates of the B1 (commensal) and E groups are also capable of causing extraintestinal infections, and the VFs carried by these strains appear to be more important in determining the pathogenic potential than is the organism's phylogenetic background. Other important factors to be considered in the pathogenesis of E. coli bloodstream infections are host characteristics, such as the portal of entry and comorbidities. Likewise, the resistance profile is important information for the management and treatment of patients with E. coli bloodstream infections. Among ESBL-producing E. coli isolates, the CTX-M-1 group (CTX-M-15) had a high prevalence, and some of the more recent resistance mechanisms, such as CTM-M-15, arose from high-risk clones that facilitate the persistence and spread of resistance around the world. This study is important because it shows the clinical and molecular characteristics of E. coli bloodstream infections from Brazilian isolates.
Limitation
The work was performed using the PCR and ERIC-PCR techniques. Further sequencing of E. coli strains causing bacteraemia could aid in the finding and description of genes or gene clusters that are peculiar to the E. coli strains that cause bacteremia.
Data Availability
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
AD, EV, and RK: conceived and designed the experiments. AD and JS: performed the experiments. AD, EV, and RK: analyzed the data. RK and VK: contributed reagents and materials. MP and MREP: reviewed the study. CdM: performed confirmation experiments.
Funding
The authors would like to thank the financial support of the Master's program Clinical and Laboratory Pathophysiology, the Research and Postgraduate Pro-Rector's Office, and the support office for the researcher of the State University of Londrina and the Araucária Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00191/full#supplementary-material
References
Alves, C., Casqueiro, J., and Casqueiro, J. (2012). Infections in patients with diabetes mellitus: a review of pathogenesis. Ind J. Endocrinol. Metab. 16, S27–S36. doi: 10.4103/2230-8210.94253
Ananias, M., and Yano, T. (2008). Serogroups and virulence genotypes of Escherichia coli isolated from patients with sepsis. Braz. J. Med. Biol. Res. 4, 877–883. doi: 10.1590/S0100-879X2008001000008
Blundell, C. D., Roberts, I. S., Sheehan, J. K., and Almond, A. (2009). Investigating the molecular basis for the virulence of Escherichia coli K5 by nuclear magnetic resonance analysis of the capsule polysaccharide. J. Mol. Microbiol. Biotechnol. 17, 71–82. doi: 10.1159/000215933
Bonacorsi, S., Houdouin, V., Mariani-Kurkdjian, P., Mahjoub-Messai, F., and Bingen, E. (2006). Comparative prevalence of virulence factors in Escherichia coli causing urinary tract infection in male infants with and without bacteremia. J. Clin. Microbiol. 44, 1156–1158. doi: 10.1128/JCM.44.3.1156-1158.2006
Bozcal, E., Eldem, V., Aydemir, S., and Skurnik, M. (2018). The relationship between phylogenetic classification, virulence and antibiotic resistance of extraintestinal pathogenic Escherichia coli in Izmir province, Turkey. PeerJ. 6:e5470. doi: 10.7717/peerj.5470
Bradford, P. A., Bratu, S., Urban, C., Visalli, M., Mariano, N., Landman, D., et al. (2004). Emergence of carbapenem-resistant klebsiella species possessing the class A carbapenem-hydrolyzing KPC-2 and inhibitor-resistant TEM-30-lactamases in New York City. Clin. Infect. Dis. 39, 55–60. doi: 10.1086/421495
Burdet, C., Clermont, O., Bonacorsi, S., Laouénan, C., Bingen, E., Aujard, Y., et al. (2014). Escherichia coli bacteremia in children age and portal of entry are the main predictors of severity. Pediatr. Infect. Dis. J. 33, 872–879. doi: 10.1097/INF.0000000000000309
Clermont, O., Bonacorsi, S., and Bingen, E. (2000). Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 66, 4555–4558. doi: 10.1128/AEM.66.10.4555-4558.2000
Clermont, O., Christenson, J. K., Denamur, E., and Gordon, D. M. (2013). The clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 5, 58–65. doi: 10.1111/1758-2229.12019
Clermont, O., Lavollay, M., Vimont, S., Deschamps, C., Forestier, C., Branger, C., et al. (2008). The CTX-M-15-producing Escherichia coli diffusing clone belongs to a highly virulent B2 phylogenetic subgroup. J. Antimicrob. Chemother. 61, 1024–1028. doi: 10.1093/jac/dkn084
Dale, A. P., and Woodford, N. (2015). Extra-intestinal pathogenic Escherichia coli (ExPEC): disease, carriage and clones. J. Infect. 71, 615–626. doi: 10.1016/j.jinf.2015.09.009
De Kraker, M. E. A., Jarlier, V., Monen, J. C. M., Heuer, O. E., Van de Sande, N., and Grundmann, H. (2013). The changing epidemiology of bacteraemias in Europe: trends from the European antimicrobial resistance surveillance system. Clin. Microbiol. Infect. 19, 860–868. doi: 10.1111/1469-0691.12028
Diacovich, L., and Gorvel, J. P. (2010). Bacterial manipulation of innate immunity to promote infection. Nat. Rev. Microbiol. 8, 117–128. doi: 10.1038/nrmicro2295
Dobrindt, U., Blum-Oehler, G., Nagy, G., Schneider, G., Johann, A., Gottschalket, G., et al. (2002). Genetic structure and distribution of four pathogenicity islands (PAI I536 to PAI IV536) of uropathogenic Escherichia coli strain 536. Infect. Immun. 70, 6365–6372. doi: 10.1128/IAI.70.11.6365-6372.2002
Fratamico, P. M., DebRoy, C., Liu, Y., Needleman, D. S., Baranzoni, G. M., and Feng, P. (2016). Advances in molecular serotyping and subtyping of Escherichia coli. Front Microbiol. 3:644. doi: 10.3389/fmicb.2016.00644
Gérard, F., Pradel, N., and Wu, L. F. (2005). Bactericidal activity of colicin V is mediated by an inner membrane protein, SdaC, of Escherichia coli. J. Bacteriol. 187:1945–1950. doi: 10.1128/JB.187.6.1945-1950.2005
Heras, J., Domínguez, C., Mata, E., and Pascual, V. (2015). GelJ – a tool for analyzing DNA fingerprint gel images. BMC Bioinformat. 16:270. doi: 10.1186/s12859-015-0703-0
Jauréguy, F., Carbonnelle, E., Bonacorsi, S., Clech, C., Casassus, P., Bingen, E., et al. (2007). Host and bacterial determinants of initial severity and outcome of Escherichia coli sepsis. Clin. Microbiol. Infect. 13, 854–62. doi: 10.1111/j.1469-0691.2007.01775.x
Johnson, J. R., and Russo, T. A. (2002). Extraintestinal pathogenic Escherichia coli: “The other bad E coli.” J. Lab. Clin. Med. 139, 155–162. doi: 10.1067/mlc.2002.121550
Johnson, J. R., and Stell, A. L. (2000). Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise james. J. Infect. Dis. 181, 261–272. doi: 10.1086/315217
Koga, V. L., Tomazetto, G., Cyoia, P. S., Neves, M. S., Vidotto, M. C., Nakazato, G., et al. (2014). Molecular screening of virulence genes in extraintestinal pathogenic Escherichia coli isolated from human blood culture in Brazil. Biomed. Res. Int. 2014:465054. doi: 10.1155/2014/465054
Köhler, C. D., and Dobrindt, U. (2011). What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 301, 642–647. doi: 10.1016/j.ijmm.2011.09.006
Laupland, K. B. (2013). Incidence of bloodstream infection: a review of population-based studies. Clin. Microbiol. Infect. 19, 492–500. doi: 10.1111/1469-0691.12144
Leflon-Guibout, V., Jurand, C., Bonacorsi, S., Espinasse, F., Guelfi, M. C., Duportail, F., et al. (2004). Emergence and spread, of three clonally related virulent isolates of CTX-M-15-producing Escherichia coli with variable resistance to aminoglycosides and tetracycline in a French geriatric hospital. Antimicrob. Agents Chemother. 48, 3736–3742. doi: 10.1128/AAC.48.10.3736-3742.2004
Lefort, A., Panhard, X., Clermont, O., Woerther, P. L., Branger, C., Mentré, F., et al. (2011). Host factors and portal of entry outweigh bacterial determinants to predict the severity of Escherichia coli bacteremia. J. Clin. Microbiol. 49, 777–783. doi: 10.1128/JCM.01902-10
Lemaître, C., Mahjoub-Messai, F., Dupont, D., Caro, V., Diancourt, L., Bingen, E., et al. (2013). A conserved virulence plasmidic region contributes to the virulence of the multiresistant Escherichia coli meningitis strain S286 belonging to phylogenetic group C. PLoS ONE. 8:074423. doi: 10.1371/journal.pone.0074423
Marra, A. R., Camargo, L. F. A., Pignatari, A. C. C., Sukiennik, T., Behar, P. R. P., Medeiros, E. A. S., et al. (2011). Nosocomial bloodstream infections in Brazilian hospitals: analysis of 2,563 cases from a prospective nationwide surveillance study. J. Clin. Microbiol. 49, 1866–1871. doi: 10.1128/JCM.00376-11
Miajlovic, H., Mac Aogáin, M., Collins, C. J., Rogers, T. R., and Smith, S. G. J. (2016). Characterization of Escherichia coli bloodstream isolates associated with mortality. J. Med. Microbiol. 65, 71–79. doi: 10.1099/jmm.0.000200
Miajlovic, H., and Smith, S. G. (2014). Bacterial self-defence: how Escherichia coli evades serum killing. FEMS Microbiol. Lett. 354, 1–9. doi: 10.1111/1574-6968.12419
Micenková, L., Benováa, A., Frankovicováb, L., Bosáka, J., Vrbac, M., Sevcíkovác, A., et al. (2017). Human Escherichia coli isolates from hemocultures: septicemia linked to urogenital tract infections is caused by isolates harboring more virulence genes than bacteraemia linked to other conditions. Int. J. Med. Microbiol. 307, 182–189. doi: 10.1016/j.ijmm.2017.02.003
Mokady, D., Gophna, U., and Ron, E. Z. (2005). Virulence factors of septicemic Escherichia coli strains. Int. J. Med. Microbiol. 295, 455–462. doi: 10.1016/j.ijmm.2005.07.007
Mora-Rillo, M., Fernández-Romero, N., Navarro-San Francisco, C., Díez-Sebastián, J., Romero-Gómez, M. P., Fernández, F. A., et al. (2015). Impact of virulence genes on sepsis severity and survival in Escherichia coli bacteremia. Virulence 6, 93–100. doi: 10.4161/21505594.2014.991234
Namikawa, H., Yamada, K., Fujimoto, H., Oinuma, K. I., Tochino, Y., Takemoto, Y., et al. (2017). Clinical characteristics of bacteremia caused by extended-spectrum beta-lactamase-producing Escherichia coli at a Tertiary Hospital. Intern. Med. 56, 1807–1815. doi: 10.2169/internalmedicine.56.7702
Nedialkova, L. P., Denzler, R., Koeppel, M. B., Diehl, M., Ring, D., Wille, T., et al. (2014). Inflammation fuels colicin Ib-dependent competition of Salmonella Serovar Typhimurium and E. coli in Enterobacterial Blooms. PLoS Pathog. 10:1003844. doi: 10.1371/journal.ppat.1003844
Owrangi, B., Masters, N., Kuballa, A., O'Dea, C., Vollmerhausen, T. L., and Katouli, M. (2018). Invasion and translocation of uropathogenic Escherichia coli isolated from urosepsis and patients with community-acquired urinary tract infection. Eur. J. Clin. Microbiol. Infect. Dis. 37, 833–839. doi: 10.1007/s10096-017-3176-4
Phillips-Houlbracq, M., Ricarda, J. D., Foucriera, A., Yoder-Himesc, D., Gaudrya, S., Bexa, J., et al. (2018). Pathophysiology of Escherichia coli pneumonia: respective contribution of pathogenicity islands to virulence. Int. J. Med. Microbiol. 308, 290–296. doi: 10.1016/j.ijmm.2018.01.003
Pitout, J. D., and Laupland, K. B. (2008). Extended spectrum β-lactamase producing Enterobacteriaceae: an emerging public-health concern. Lancet. Infect. Dis. 8, 159–166. doi: 10.1016/S1473-3099(08)70041-0
Ron, E. Z. (2010). Distribution and evolution of virulence factors in septicemic Escherichia coli. Int. J. Med. Microbiol. 300, 367–370. doi: 10.1016/j.ijmm.2010.04.009
Russo, T. A., and Johnson, J. R. (2003). Medical and economic impact of extraintestinal infections due to Escherichia coli: focus on an increasingly important endemic problem. Microbes Infect. 5, 449–56. doi: 10.1016/S1286-4579(03)00049-2
Russo, T. A., Wenderoth, S., Carlino, U. B., Merrick, J. M., and Lesse, A. J. (1998). Identification, genomic organization, and analysis of the group III capsular polysaccharide genes kpsD, kpsM, kpsT, and kpsE from an extraintestinal isolate of Escherichia coli (CP9, O4/K54/H5). J. Bacteriol. 180, 338–49.
Sabaté, M., Moreno, E., Pérez, T., Andreu, A., and Prats, G. (2006). Pathogenicity island markers in commensal and uropathogenic Escherichia coli isolates. Clin. Microbiol. Infect. 12, 880–6. doi: 10.1111/j.1469-0691.2006.01461.x
Seki, L. M., Pereira, P. S., Conceição, M. S., Souza, M. J., Marques, E. A., and Carballido, J. M. (2013). Molecular epidemiology of CTX-M producing Enterobacteriaceae isolated from bloodstream infections in Rio de Janeiro, Brazil: Emergence of CTX-M-15. Braz. J. Infect. Dis. 17, 640–646. doi: 10.1016/j.bjid.2013.03.012
Shaik, S., Ranjan, A., Tiwari, S. K., Hussain, A., Nandanwar, N., Kumar, N., et al. (2017). Comparative genomic analysis of globally dominant ST131 clone with other epidemiologically successful. MBio 8:5. doi: 10.1128/mBio.01596-17
Skjøt-Rasmussen, L., Ejrnæs, K., Lundgren, B., Hammerum, A. M., and Frimodt-Møller, N. (2012). Virulence factors and phylogenetic grouping of Escherichia coli isolates from patients with bacteraemia of urinary tract origin relate to sex and hospital- vs. community-acquired origin. Int. J. Med. Microbiol. 302, 129–34. doi: 10.1016/j.ijmm.2012.03.002
Sonnen, A. F. P., and Henneke, P. (2013). Role of pore-forming toxins in neonatal sepsis. Clin. Dev. Immunol. 2013:608456. doi: 10.1155/2013/608456
Subashchandrabose, S., and Mobley, H. L. T. (2015). Virulence and fitness determinants of uropathogenic Escherichia coli. Microbiol. Spectr. 3:4. doi: 10.1128/microbiolspec.UTI-0015-2012
Taveira, M. R. V., Lima, L. S., Araújo, C. C., and Mello, M. J. G. (2017). Risk factors for central line–associated bloodstream infection in pediatric oncology patients with a totally implantable venous access port: a cohort study. Pediatr. Blood Cancer. 64, 336–342. doi: 10.1002/pbc.26225
Usein, C. R., Papagheorghe, R., Oprea, M., Conde,I, M., and Strãut, M. (2016). Molecular characterization of bacteremic Escherichia coli isolates in Romania. Folia Microbiol. 6:6. doi: 10.1007/s12223-015-0427-6
Vaishnavi, C. (2013). Translocation of gut flora and its role in sepsis. Ind. J. Med. Microbiol. 31, 334–342. doi: 10.4103/0255-0857.118870
Versalovic, J., Koeuth, T., and Lupski, J. R. (1991). Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 19, 6823–6831. doi: 10.1093/nar/19.24.6823
Wilson, B. R., Bogdan, A. R., Miyazawa, M., Hashimoto, K., and Tsuji, Y. (2017). Siderophores in iron metabolism: from mechanism to therapy potential. Trends Mol. Med. 22, 1077–1090. doi: 10.1016/j.molmed.2016.10.005
Woodford, N., Fagan, E. J., and Ellington, M. J. (2006). Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 57:154–155. doi: 10.1093/jac/dki412
Yair, Y., and Gophna, U. (2018). Pandemic Bacteremic Escherichia coli Strains: evolution and emergence of drug-resistant pathogens. Curr. Top. Microbiol. Immunol. 416, 163–180. doi: 10.1007/82_2018_109
Yokota, P. K. O., Marra, A. R., Belucci, T. R., Victor, E. S., Santos, O. F. P., and Edmond, M. B. (2016). Outcomes and predictive factors associated with adequacy of antimicrobial therapy in patients with central line-associated bloodstream infection. Front. Public Health. 4:284 doi: 10.3389/fpubh.2016.00284
Keywords: ExPEC, bloodstream infection, phylogenetic group, virulence factors, ESBL, CTX- M-15
Citation: Daga AP, Koga VL, Soncini JGM, de Matos CM, Perugini MRE, Pelisson M, Kobayashi RKT and Vespero EC (2019) Escherichia coli Bloodstream Infections in Patients at a University Hospital: Virulence Factors and Clinical Characteristics. Front. Cell. Infect. Microbiol. 9:191. doi: 10.3389/fcimb.2019.00191
Received: 22 February 2019; Accepted: 20 May 2019;
Published: 06 June 2019.
Edited by:
Jean-Ralph Zahar, Hôpital Avicenne, FranceReviewed by:
Yehuda Carmeli, Tel Aviv Sourasky Medical Center, IsraelAdriana Bentancor, University of Buenos Aires, Argentina
Copyright © 2019 Daga, Koga, Soncini, de Matos, Perugini, Pelisson, Kobayashi and Vespero. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Paula Daga, YW5hX2RnYUBob3RtYWlsLmNvbQ==