 Steven Higgins
Steven Higgins Stephan Heeb
Stephan Heeb Giordano Rampioni
Giordano Rampioni Mathew P. Fletcher
Mathew P. Fletcher Paul Williams
Paul Williams Miguel Cámara
Miguel Cámara
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 23 July 2018
Sec. Molecular Bacterial Pathogenesis
Volume 8 - 2018 | https://doi.org/10.3389/fcimb.2018.00252
This article is part of the Research Topic Quorum Network (Sensing/Quenching) in Multidrug-Resistant Pathogens View all 18 articles
The Pseudomonas aeruginosa quorum sensing (QS) network plays a key role in the adaptation to environmental changes and the control of virulence factor production in this opportunistic human pathogen. Three interlinked QS systems, namely las, rhl, and pqs, are central to the production of pyocyanin, a phenazine virulence factor which is typically used as phenotypic marker for analysing QS. Pyocyanin production in P. aeruginosa is a complex process involving two almost identical operons termed phzA1B1C1D1E1F1G1 (phz1) and phzA2B2C2D2E2F2G2 (phz2), which drive the production of phenazine-1-carboxylic acid (PCA) which is further converted to pyocyanin by two modifying enzymes PhzM and PhzS. Due to the high sequence conservation between the phz1 and phz2 operons (nucleotide identity > 98%), analysis of their individual expression by RNA hybridization, qRT-PCR or transcriptomics is challenging. To overcome this difficulty, we utilized luminescence based promoter fusions of each phenazine operon to measure in planktonic cultures their transcriptional activity in P. aeruginosa PAO1-N genetic backgrounds impaired in different components of the las, rhl, and pqs QS systems, in the presence or absence of different QS signal molecules. Using this approach, we found that all three QS systems play a role in differentially regulating the phz1 and phz2 phenazine operons, thus uncovering a higher level of complexity to the QS regulation of PCA biosynthesis in P. aeruginosa than previously appreciated.
IMPORTANCE
The way the P. aeruginosa QS regulatory networks are intertwined creates a challenge when analysing the mechanisms governing specific QS-regulated traits. Multiple QS regulators and signals have been associated with the production of phenazine virulence factors. In this work we designed experiments where we dissected the contribution of specific QS switches using individual mutations and complementation strategies to gain further understanding of the specific roles of these QS elements in controlling expression of the two P. aeruginosa phenazine operons. Using this approach we have teased out which QS regulators have either indirect or direct effects on the regulation of the two phenazine biosynthetic operons. The data obtained highlight the sophistication of the QS cascade in P. aeruginosa and the challenges in analysing the control of phenazine secondary metabolites.
Pseudomonas aeruginosa is a highly adaptable bacterium, which can be found in a range of challenging environments, including the human host. This is achieved in great part by the ability of this opportunistic pathogen to finely control the expression of a wide range of genes, including those involved in the production of virulence determinants, in response to environmental and metabolic stimuli (Lee et al., 2006; Balasubramanian et al., 2013; Sun et al., 2016). The expression of many virulence genes in P. aeruginosa is also controlled in a cell density dependent manner by quorum sensing (QS) (Smith and Iglewski, 2003; Bjarnsholt and Givskov, 2007).
P. aeruginosa has a sophisticated QS network consisting of three separate but interwoven systems, namely las, rhl, and pqs and their cognate QS signal molecules (QSMs). The QSMs N-3-oxo-dodecanoyl-homoserine lactone (3OC12-HSL) produced by LasI, and N-butanoyl-homoserine lactone (C4-HSL) produced by RhlI interact with their cognate transcriptional regulators LasR and RhlR respectively, leading to the activation or repression of multiple genes including the genes coding for their cognate signal synthases (Schuster et al., 2013). The LasR/3OC12-HSL complex also induces the transcription of rsaL, a gene integrated in the las QS system coding for the global transcriptional regulator RsaL (de Kievit et al., 1999). This protein directly represses the transcription of multiple genes, including lasI, hence exerting a homeostatic effect on 3OC12-HSL production, and conferring robustness to the expression of a sub-set of genes of the las regulon with respect to fluctuations in LasR levels (Rampioni et al., 2006, 2007; Bertani et al., 2007).
The pqs QS system is more complex than the las and rhl systems, since multiple enzymes encoded by the pqsABCDE operon are required for the synthesis of 2-alkyl-4(1H)-quinolones (AQs) including the QSMs 2-heptyl-4-hydroxyquinoline (HHQ), which in turn is converted to 2-heptyl-3-hydroxy-4-quinolone (PQS) by the monooxygenase PqsH. Both HHQ and PQS can bind to and activate the transcriptional regulator PqsR (also known as MvfR). The PqsR/HHQ and PqsR/PQS complexes bind the PpqsA promoter region and increase the transcription of the pqsABCDE operon, thus generating a feedback loop that accelerates AQ biosynthesis and increasing production of PqsE, coded by the last gene of the pqsABCDE operon (Heeb et al., 2011; Dulcey et al., 2013). PqsE is a thioesterase involved in AQ biosynthesis (Drees and Fetzner, 2015) but this protein also controls indirectly the expression of multiple virulence factors even in the absence of AQs. The molecular mechanism by which PqsE impacts on QS target gene expression remains unknown (Hazan et al., 2010; Rampioni et al., 2010, 2016).
The QS circuit of P. aeruginosa has been widely reported to have a hierarchal structure. Under growth conditions using rich media, it is generally accepted that the las QS system is the first to become active leading to the activation of the rhl and pqs systems (Pesci et al., 1997; de Kievit et al., 2002; Gallagher et al., 2002; Xiao et al., 2006). However it has been reported that RhlR can in part overcome the absence of the las system in late stationary phase (Dekimpe and Deziel, 2009). RhlR is required for production of certain virulence factors but has a negative impact on the pqs system by repressing PQS signal production through interference with the expression of pqsR and pqsABCDE (McKnight et al., 2000; Wade et al., 2005; Xiao et al., 2006; Brouwer et al., 2014). In turn the pqs system has a positive effect upon the rhl system, as addition of PQS to a P. aeruginosa culture has been shown to increase the levels of RhlR and the rhl QS signal C4-HSL (McKnight et al., 2000; Diggle et al., 2003). The interactions of the QS systems are detailed in Figure S1.
QS has been shown to affect the transcription of hundreds of downstream genes (Schuster et al., 2003; Wagner et al., 2003; Rampioni et al., 2007, 2010) with some of these specifically controlled by distinct QS systems, while others are induced or repressed by multiple QS regulators (Schuster and Greenberg, 2007; Farrow et al., 2008; Cornforth et al., 2014; Rampioni et al., 2016).
The production of pyocyanin (PYO), a key virulence factor produced by P. aeruginosa, has been linked to multiple QS systems. This particular phenazine is often used as a marker to assess QS behavior as it is easily measurable and contributes significantly toward the green color of P. aeruginosa cultures (Frank and Demoss, 1959). Although PYO is the most studied phenazine in P. aeruginosa, this organism is capable of producing up to 5 different phenazine derivatives (Mavrodi et al., 2001, 2010). Phenazine biosynthesis begins with the conversion of chorismic acid to phenazine-1-carboxylic acid (PCA) by the action of the enzymes encoded by the biosynthetic operon phzABCDEFG, which is conserved across the fluorescent Pseudomonad species (Mavrodi et al., 2006, 2010). Interestingly P. aeruginosa has 2 functional copies of this operon designated phz1 and phz2. Both operons produce PCA, which can be further converted to phenazine-1-carboxamide by the action of PhzH and to 1-hydroxyphenazine by PhzS. The action of PhzM is to convert PCA to 5-methylphenazine-1-carboxylic acid betaine, which can be further converted to PYO by PhzS (Mavrodi et al., 2001, 2006, 2010).
PYO production has been linked to QS in many reported studies and to date LasR, RhlR, RsaL, PqsE, PqsR and both AQ signal molecules HHQ and PQS have been found to play a role in the control of its production (Whiteley and Greenberg, 2001; Gallagher et al., 2002; Diggle et al., 2003; Schuster et al., 2003; Wagner et al., 2003; Rampioni et al., 2007, 2010; Farrow et al., 2008; Lu et al., 2009; Liang et al., 2011; Recinos et al., 2012; Cabeen, 2014; Sun et al., 2017). Although QS controls PYO production, the high sequence conservation between the two phenazine producing operons phz1 and phz2 have made analysing their individual expression by DNA hybridization techniques challenging (Schuster et al., 2003; Wagner et al., 2003; Rampioni et al., 2007, 2010).
It is unlikely that both phenazine biosynthesis operons are controlled in the same manner as they are located some distance apart on the PAO1 chromosome and have very different promoter regions (Mavrodi et al., 2001; Whiteley and Greenberg, 2001; Rampioni et al., 2007; Winsor et al., 2011). The phz1 operon (from PA4210 to PA4216) is flanked by phzM upstream (PA4209) and phzS downstream (PA4217), both of which are required to produce PYO. The phz2 operon (from PA1899 to PA1905) is flanked upstream by the qscR gene (PA1898), coding for the orphan QS receptor QscR, and downstream by the PA1906 gene, coding for a hypothetical protein of unknown function. The phzH gene (PA0051) is unlinked to the other phenazine biosynthetic genes (Figure S2). It would appear by looking at the positions of the operons on the chromosome of P. aeruginosa PAO1 that the phz1 operon is clustered with the genes required to produce PYO, and hence could be more closely associated with PYO production than phz2. That said, the phz2 operon has been shown to contribute significantly to the production of PYO, especially under non-planktonic growth conditions (Recinos et al., 2012; Dietrich et al., 2013).
There is a greater quantity of available information about the control of phz1 than phz2, and a lux box, for LasR or RhlR binding, has been predicted upstream of the −10 region of the phz1 promoter (PphzA1) (Whiteley and Greenberg, 2001). The QS repressor RsaL has also been shown to bind to this promoter in an electrophoretic mobility shift assay (EMSA) at the downstream end of the −10 promoter region, thus acting as a repressor of phz1 transcription (Rampioni et al., 2007). Moreover, RsaL exerts an indirect negative effect on phz1 transcription by increasing the production of the phz1 repressor protein CdpR (Sun et al., 2017).
Less is known about the regulation of the phz2 promoter (PphzA2). The intergenic region between qscR and phz2 was probed for RsaL binding by different groups with negative results (Rampioni et al., 2007; Sun et al., 2017), hence RsaL appears to control PphzA1 but not PphzA2. Recinos and colleagues found that phz2 transcription is induced by HHQ under anaerobic conditions (Recinos et al., 2012). Identification of a predicted ANR/DNR binding site within the PphzA2 promoter supports the notion that phz2 transcription is increased in anaerobic environments (Trunk et al., 2010). The orphan luxR QS regulator, QscR, which is encoded directly upstream of phzA2 has been reported to be a repressor of phzA2 (Ledgham et al., 2003; Lequette et al., 2006). The qscR and phz2 intergenic region was probed for QscR binding with a negative result (Lee et al., 2006) suggesting the effect of QscR on phzA2 is indirect due to the ability of QscR to form inactive heterodimers with LasR and RhlR (Chugani et al., 2001).
To gain a further understanding of the control of each phenazine biosynthesis operon by QS, lux-based promoter fusions for each operon were created and tested in a range of QS mutants. We identified an RsaL dependent switch, which can move PCA production from one operon to the other, and vice versa. This switching mechanism was confirmed by modification of the QS network activity in selected mutants with the addition of QS signal molecules and specific QS regulator genes expressed from plasmids. This allowed us to confirm the hierarchal structure of QS in rich media under planktonic conditions and to develop a more in depth model of how QS controls phz1 and phz2 transcription in P. aeruginosa.
The bacterial strains and plasmids used in this study are detailed in Table S1. They were routinely grown in Lysogeny Broth (LB) at 37°C with shaking at 200 rpm, with the exception of P. aeruginosa conjugation recipient strains, which were incubated at 42°C. When required, LB was supplemented with the following antibiotics: for E. coli, 10 μg ml−1 tetracycline (Tc), 30 μg ml−1 chloramphenicol (Cm), or 100 μg ml−1 ampicillin (Ap); for P. aeruginosa, 150 μg ml−1 Tc, 375 μg ml−1 Cm or 800 μg ml−1 streptomycin (Sm). Media were supplemented with 1 mM (final concentration) isopropyl β-D-1-thiogalactopyranoside (IPTG) for inducible strains where required, unless otherwise stated. Synthetic signal molecules PQS and 2-methyl-3-hydroxy-4-quinolone (mPQS) were added to cultures at a final concentration of 100 μM where required. To select for P. aeruginosa after mating experiments LB agar plates were supplemented with 15 μg ml−1 nalidixic acid (Nal).
All plasmids generated and/or used in this study are listed in Table S1. Routine DNA manipulations including extraction, restriction, ligation, electroporation, conjugation and agarose gel electrophoresis were performed using standard molecular methods (Sambrook and Russell, 2001). Plasmid extraction was completed using a Qiagen™ QiaQuick miniprep kit following the manufacturer's instructions. The TcR marker of pMINI-CTX1 derived constructs integrated into the chromosome of P. aeruginosa was removed using the Flp recombinase system as previously described (Hoang et al., 1998, 2000). All primers used for DNA amplification by PCR are detailed in Table S2. DNA sequencing was conducted at the University of Nottingham's DNA sequencing facility.
pMINI-lux was generated by cloning the BamHI-EcoRI fragment of pBluelux (Atkinson et al., 2008), containing the luxCDABE operon, into similarly digested mini-CTX1, using standard molecular methods (Sambrook and Russell, 2001). The PphzA1 and PphzA2 promoter regions were PCR amplified from P. aeruginosa PAO1 chromosomal DNA using primer pairs FWPphzA1-RVPphzA1, and FWPphzA2-RVPphzA2, respectively (Table S2). The PCR products were independently cloned into the pMINI-lux construct between the EcoRI and XhoI restriction sites, resulting in the plasmids pPphzA1-lux and pPphzA1-lux, respectively.
The rsaL coding region was amplified by PCR from P. aeruginosa PAO1 chromosomal DNA using primers FWrsaL and RVrsaL (Table S2). The resulting PCR product was cloned into pME6032 between the EcoRI and XhoI restriction sites using standard molecular techniques. This plasmid was introduced to P. aeruginosa strains by electroporation (Choi et al., 2006).
All cloned fragments obtained by PCR were verified by DNA sequencing to match the reference sequences (Winsor et al., 2011).
To generate the double mutant strain P. aeruginosa pqsEind ΔlasR, the lasR gene was deleted from the chromosome of the PAO1 pqsEind strain (Rampioni et al., 2010) by using the pME3087-ΔlasR plasmid (Harrison et al., 2014).
Briefly, the pME3087-ΔlasR plasmid was mobilized by conjugation into the P. aeruginosa pqsEind recipient strain using E. coli S17.1 λpir as a donor. Exconjugants were selected on LB plates supplemented with 150 μg ml−1 Tc and 15 μg ml−1 Nal. Strains were re-streaked twice on LB lacking antibiotic and then subjected to 1 round of Tc sensitivity enrichment to select for double cross-over events (Voisard et al., 1994). Five colonies which were TcS were then tested by PCR for loss of the lasR coding region.
To generate a P. aeruginosa PAO1 mutant strain with an rsaL deletion (ΔrsaL), allelic exchange was obtained by using the pDM4-ΔrsaL plasmid, derived from the suicide vector pDM4 (Milton et al., 1996). The upstream and the downstream DNA regions of rsaL were PCR amplified from P. aeruginosa PAO1 chromosomal DNA using primer pairs FWrsaLUP + RVrsaLUP and FWrsaLDOWN + RVrsaLDOWN, respectively (Table S2). The upstream and downstream PCR fragments were subsequently cloned in pDM4 by XhoI-BamHI and BamHI-XbaI restriction, respectively. The resulting pDM4-ΔrsaL plasmid was verified by restriction analysis and sequencing. Allelic exchange in P. aeruginosa PAO1 following conjugal mating with the E. coli S17.1 λpir (pDM4-ΔrsaL) donor strain and sucrose counter selection was performed as previously described (Westfall et al., 2004). The resulting PAO1 ΔrsaL mutant strain was confirmed by PCR.
Three independent single colonies of P. aeruginosa strains carrying reporter constructs were grown overnight in LB (separate tubes) at 37°C with shaking at 200 rpm. One-milliliter of overnight culture was washed in 1 ml of fresh LB to remove secreted bacterial products and QS signal molecules. Twenty-microliters aliquots were inoculated into 1 ml of fresh LB, and 300 μl of the resulting cultures were dispensed into wells of a 96-well black flat-transparent-bottom microtiter plate. When needed, strains with inducible genes were grown with or without 1 mM IPTG unless otherwise stated. Microtiter plates were incubated at 37°C in a TECAN GENios automated luminometer-spectrophotometer with which luminescence and turbidity were recorded every 30 min. Promoter activity per cell is given as relative light units divided by absorbance at 600 nm wavelength (A600).
Standard deviation of the mean of the three biological replicates is reported. A paired t-test was used to compare each mutant with the relevant control for reach experiment. A P-value of ≤0.05 was considered significant.
To ascertain how the phenazine operons phz1 and phz2 are regulated by the different elements of the QS circuit in P. aeruginosa PAO1-N, the pPphzA1-lux and pPphzA2-lux reporter plasmids, containing transcriptional fusions between the PphzA1 and PphzA2 promoter regions and the luxCDABE operon for bioluminescence, respectively, were generated and inserted in the chromosome of strain PAO1-N and different mutants derived from it. In detail, in the pPphzA1-lux plasmid a 727-bp DNA fragment comprising the entire intergenic region between phzM and phzA1 was cloned upstream of the luxCDABE operon, while in pPphzA2-lux a 497-bp DNA fragment including the entire intergenic region between qscR and phzA2 was cloned instead. By including these relatively large promoter regions the likelihood of missing any key regulatory element upstream of the known unique transcription start sites of PphzA1 and PphzA2 (Dötsch et al., 2012) was minimized. Cloned promoter regions include the first two codons of phzA1 and phzA2, respectively. Since the vast majority of studies on the QS circuit of P. aeruginosa have been undertaken in rich media, LB was used in this work so that predictions about the behavior of the QS network in specific QS mutants could be made and the results obtained compared with previous studies.
Firstly, the impact of QS elements, previously identified as key players in the regulation of PYO production, on PphzA1 and PphzA2 activity was investigated in PAO1-N, since there have been some strain-specific differences shown in the regulation of these operons by QS (Sun et al., 2016). Figure 1, shows that under the planktonic conditions studied the activity of PphzA1 is several fold higher than that of PphzA2 which is in line with what has been detected in P. aeruginosa PA14 (Recinos et al., 2012). LasR, RhlR, and PqsE showed a positive effect on the activation of the PphzA1 and PphzA2 promoters. PphzA1 activity was completely abrogated in the ΔlasR and ΔrhlR mutants, and strongly decreased (90% reduction) in the non-induced pqsE conditional mutant strain, pqsEind (P < 0.01). The effect of LasR, RhlR, and PqsE on PphzA2 appear to be milder although still significant (P < 0.05), with reporter activities reduced by 78, 71, and 52% in the ΔlasR, ΔrhlR and non-induced pqsEind strains, respectively. When PqsE was fully induced with 1 mM IPTG in the pqsEind strain, a 3.5-fold increase in promoter activity of both PphzA1 and PphzA2 was observed relative to the PAO1-N wild type (P < 0.01). Conversely, RsaL had an opposite effect on the two promoters, since PphzA1 activity is significantly increased (298% increase) and PphzA2 activity decreased (80% reduction) (P < 0.01), in the ΔrsaL mutant compared to PAO1-N wild type. These results are in accordance with published data for other P. aeruginosa strains, showing a positive effect of LasR, RhlR and PqsE on PCA biosynthesis in the human pathogen P. aeruginosa PA14 and in the rhizosphere bacterium P. aeruginosa PA1201 (Recinos et al., 2012; Sun et al., 2016, 2017). Also the dual effect of RsaL on PphzA1 and PphzA2 is in line with what was previously observed in P. aeruginosa PA1201 (Sun et al., 2017). The growth data for this experiment is shown in Figure S3.
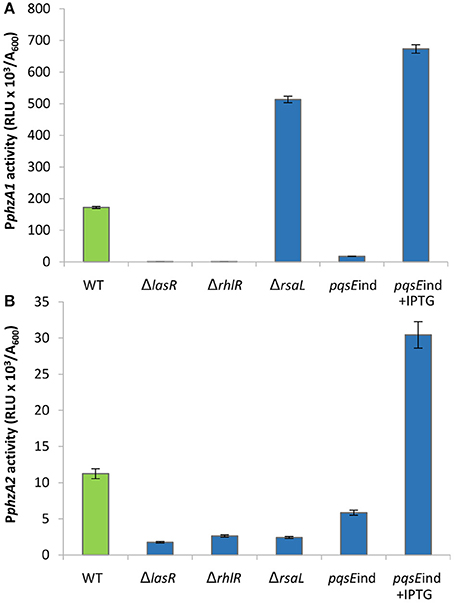
Figure 1. Effect of QS elements on PphzA1 and PphzA2 activity. Maximal promoter activity per cell measured in the indicated strains derived from P. aeruginosa PAO1-N carrying the transcriptional fusions PphzA1-lux (A) or PphzA2-lux (B). Bioluminescence (relative light units, RLU) and cell density (A600) were recorded after 6 h incubation at 37°C. Mean of three independent experiments is shown with standard deviation.
To further validate the regulation of PphzA2 by RsaL, the rsaL deletion was complemented via the IPTG inducible pRsaL plasmid. Some partial restoration of PphzA2 activity was observed in the ΔrsaL strain in the presence of pRsaL, likely as a consequence of basal rsaL transcription from the tac promoter (Guzman et al., 1995), while in the presence of 0.1 mM IPTG PphzA2 activity was restored to wild type levels (P < 0.05) (Figure 2). Overall, these data confirm that in PAO1-N RsaL is a repressor of phz1 transcription and has a positive effect upon PphzA2, the latter likely mediated by an ancillary PphzA2-regulator under the control of RsaL, since purified RsaL has not been shown to directly bind to PphzA2 in EMSA studies (Rampioni et al., 2007; Sun et al., 2017). The growth data for this experiment is shown in Figure S4.
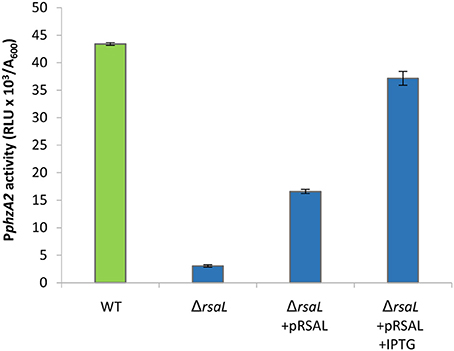
Figure 2. Effect of pRsaL on PphzA2 activity. Maximal promoter activity of PphzA2-lux in the wild type PAO1-N and the rsaL mutant background, with and without complementation. Promoter activities are normalized by cell density (A600).
High levels of PqsE resulted in an increase in promoter activity for both phenazine biosynthesis operons (Figure 1). Since a lux-box is present in the PphzA1 promoter region and PqsE does not act as a transcriptional regulator, it can be hypothesized that PqsE exerts a positive effect on PphzA1 activity via the LasR and/or RhlR transcriptional regulators. This hypothesis was tested by analysing PphzA1 activity in the double mutants pqsEind ΔlasR and pqsEind ΔrhlR respectively in which pqsE expression can be restored in the presence of IPTG. Figure 3 reveals that while PqsE induction with IPTG resulted in high PphzA1 activity in the pqsEind strain, the activity of this promoter under induced conditions was reduced by 80% in the pqsEind ΔlasR mutant and a 2 h delay in activation of PphzA1 relative to the pqsEind strain induced with IPTG was observed. PphzA1 activity was completely abrogated in the pqsEind ΔrhlR background. The growth data for this experiment is shown in Figure S5.
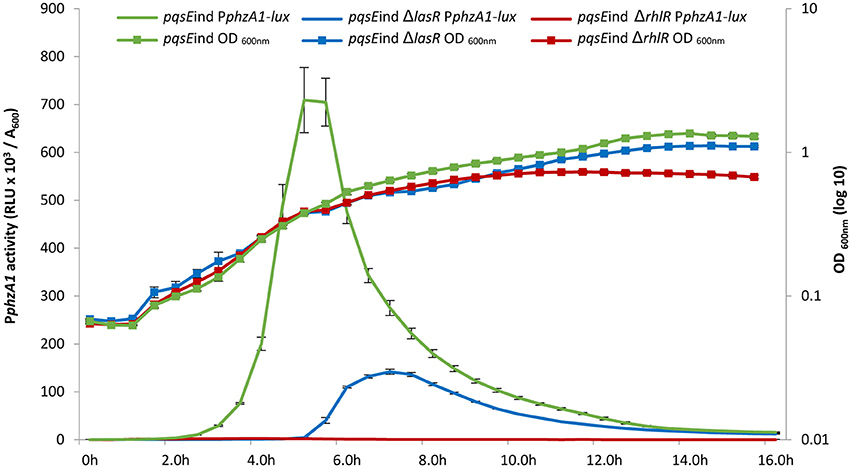
Figure 3. A delay in the timing of PphzA1 activation was observed when lasR was mutated in the pqsEind strain. The pqsEind strain increased the activity of PphzA1-lux (green line) but when lasR is mutated a 2 h delay in promoter activation is observed (blue line) compared with the pqsEind. When rhlR is deleted from the pqsEind strain PphzA1-lux activity is abolished (red line). The growth curves of the 3 mutants are also shown and this data is plotted on the Z axis. All strains were induced with 1 mM IPTG and promoter activities are normalized by cell density (A600).
The las QS system has a positive effect on the activity of both the rhl and pqs QS systems (Pesci et al., 1997; Medina et al., 2003; Xiao et al., 2006). Moreover, a study by McKnight et al. (2000) showed that addition of exogenous PQS positively regulates the rhl system, and Diggle et al. (2003) showed that addition of exogenous PQS advances and enhances pyocyanin production and increases RhlR levels. We therefore hypothesized that the reduction of transcriptional activity of PphzA1 in the pqsEind ΔlasR mutant could be caused by reduced activity of the rhl and pqs systems in this mutant background, and hence exogenous provision of PQS should compensate for a las mutation. To test this, PphzA1 activity was analyzed in the ΔlasR and ΔrhlR strains in the presence of 100 μM exogenous PQS. To discard any effects related to the iron chelating properties of PQS, the non-signaling quinolone molecule methyl PQS (mPQS) was used as a control, since this molecule is capable of binding iron like PQS, but is unable to trigger gene expression via PqsR (Diggle et al., 2007). Addition of PQS was found to compensate for a lasR deletion (P < 0.05), while an rhlR deletion resulted in no activation of the PphzA1 promoter, irrespective of the absence or presence of PQS (Figure 4). The addition of PQS also impacted on the timing of PphzA1 gene expression in both wild type and lasR deletion strains with the activation of this promoter triggered 1 h earlier than in the absence of this signal molecule (data not shown). The growth data for this experiment is shown in Figure S6.
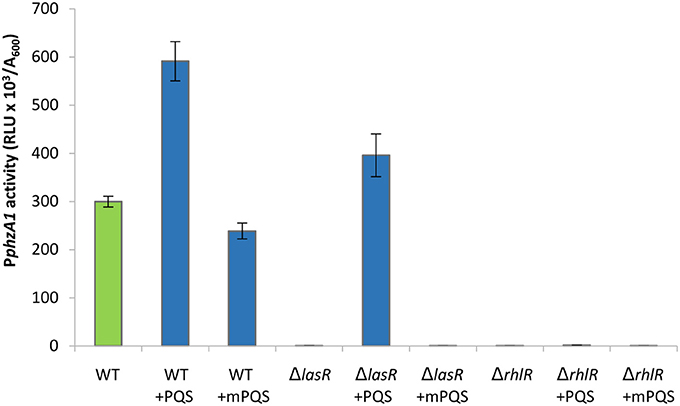
Figure 4. The addition of exogenous PQS compensates for a mutation in lasR by increasing activity of PphzA1. Maximal promoter activity of PphzA1-lux in the wild type PAO1-N, lasR and rhlR deletion backgrounds in the presence of PQS or methyl PQS (mPQS). Promoter activities are normalized by cell density (A600).
These data suggest that RhlR is a direct positive regulator of the phz1 operon, as loss of this element abolishes PphzA1 activity, while LasR acts as an indirect PphzA1 regulator.
Addition of PQS in the previous experiment would be expected to increase the expression of the pqsABCDE operon and hence PqsE production through the activation of PqsR. The data presented here suggests that PqsE is required to activate PphzA1 and PqsE would be present in high levels after the addition of exogenous PQS to the culture. To further investigate the importance of RhlR and LasR in the PqsE-mediated activation of PphzA1 the above experiments were repeated using the pqsEind conditional mutant with additional lasR and rhlR mutations.
Addition of PQS to the un-induced pqsEind strain resulted in no significant increase in PphzA1 activity, compared to the un-induced pqsEind strain (P < 0.05). This result confirms that PqsE rather than PQS on its own or through PqsR activation is required to reach high levels of PphzA1 transcription (Figure 5). Since PqsR has been reported to directly bind to rhlI/R resulting in some increased expression of these genes (Maura et al., 2016) this result suggests that activated RhlR in combination with PqsR are unable to activate PphzA1 transcription. In the pqsEind strain the expression of pqsE is decoupled from that of pqsABCD, due to a transcriptional terminator introduced downstream of pqsD, hence PqsE production is not under the control of PqsR (Rampioni et al., 2010). Addition of PQS when pqsEind was induced by IPTG slightly increased PphzA1 activity compared with the non-induced pqsEind without PQS, this may be due to some PqsR direct activation of rhlI/R. Again PQS was unable to trigger reporter gene expression in the pqsEind ΔlasR strain when pqsE was not induced. Interestingly, when pqsE was fully induced and PQS was added to the pqsEind ΔlasR mutant, a significant increase in PphzA1 activity (P < 0.05) of approximately 50% was observed compared with the fully induced pqsEind mutant. No PphzA1 expression was detected in the pqsEind ΔrhlR strain under any of the conditions tested confirming the importance of RhlR in activating phz1 transcription (Figure 5). The growth data for this experiment is shown in Figure S6.
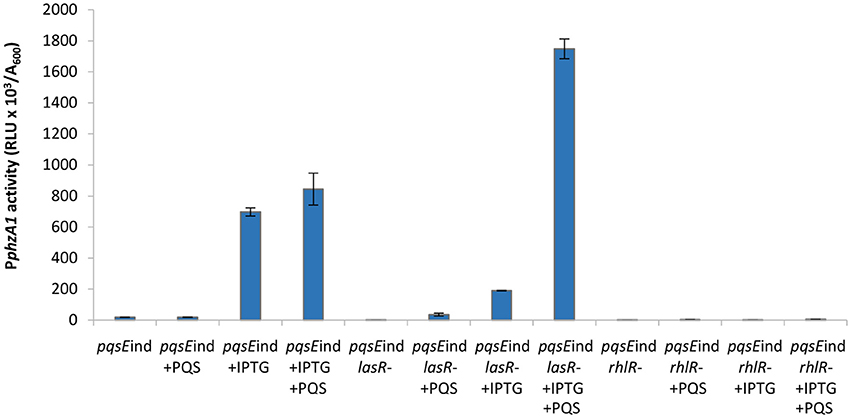
Figure 5. PQS is unable to induce high levels of PphzA1 transcription in the absence of PqsE or RhlR. Maximal promoter activity of PphzA1-lux in the pqsEind strain and pqsEind strain carrying additional lasR or rhlR mutations in the presence or absence of IPTG and PQS. Promoter activities are normalized by cell density (A600).
The high levels of PphzA1 activity observed when both, PQS is added and PqsE expression is induced with IPTG in the pqsEind ΔlasR strain, could be due to low levels of RsaL, which is a repressor of PphzA1 (Figure 1). Since LasR activates the rsaL promoter (PrsaL), the ΔlasR mutant is expected to express low levels of RsaL. Overall, these data are consistent with PqsE and RhlR as the key activators of the phz1 operon with RsaL acting as a repressor.
In Figure 1 we show that both PqsE and RsaL exert positive control over the expression of PphzA2 but the influence the las and rhl systems may have on this regulation is not clear. To investigate this further, the expression of PphzA2 was studied in pqsEind and pqsEind with a lasR or a rhlR deletion. When pqsE was induced in either strain, PphzA2 activity could only achieve 5% and 10% of the pqsEind strain (Figure 6), growth data Figure S7. This differed from the result obtained for PphzA1, as a lasR deletion in the pqsEind induced strain decreased but did not abrogate PphzA1 activity (Figure 5). To ascertain whether addition of exogenous PQS could compensate for lasR and rhlR deletions, PphzA2 activity was evaluated in the ΔlasR and ΔrhlR mutants supplemented with exogenous PQS or mPQS, the latter molecule used as an iron-binding negative control as before. Unlike PphzA1, where PQS restored promoter activity in the ΔlasR mutant, no significant increase (P < 0.05) in PphzA2 activity was observed when PQS was added to the ΔlasR and ΔrhlR strains (Figure 7), growth data Figure S8.
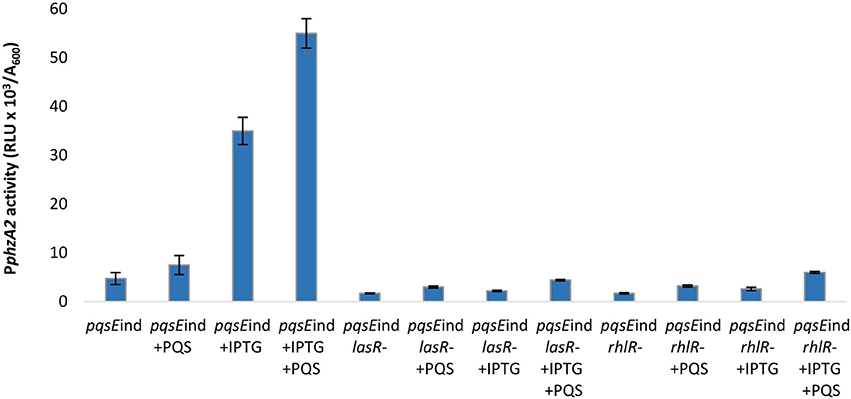
Figure 6. PQS is unable to induce PphzA2 transcription in the absence of PqsE, LasR, and RhlR. Maximal promoter activity of PphzA1-lux in the pqsEind strain and pqsEind strain carrying additional lasR or rhlR mutations in the presence or absence of IPTG and PQS. Promoter activities are normalized by cell density (A600).
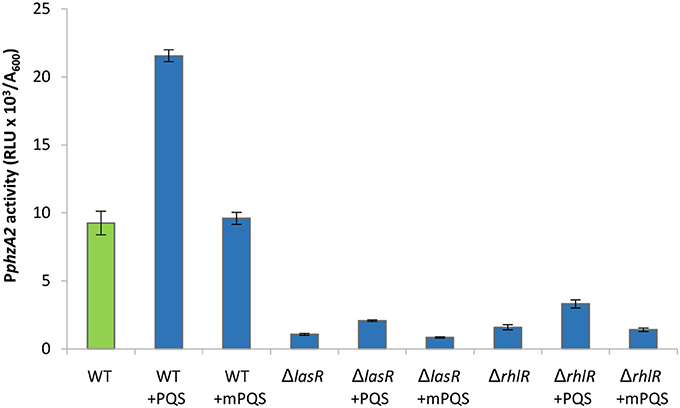
Figure 7. PphzA2 activity cannot be restored by exogenous PQS in lasR or rhlR mutants. Maximal promoter activity of PphzA2-lux in the wild type PAO1-N, lasR and rhlR deletion backgrounds in the presence of PQS or methyl PQS (mPQS). Promoter activities are normalized by cell density (A600).
These results suggests that both LasR and RhlR are required to activate PphzA2. Since addition of exogenous PQS increases the levels of PqsE (Heeb et al., 2011) we next investigated whether PqsE or PqsR were responsible for the increase in PphzA2 activity. The same experimental strategy as for PphzA1 analysis was used and activity of PphzA2 was assayed in the pqsEind mutant strain and the pqsEind strains with additional lasR and rhlR deletions, in the presence of exogenously added PQS and IPTG.
The result of this experiment showed that addition of PQS to the un-induced pqsEind strain resulted in no significant increase in PphzA2 activity, compared to the un-induced pqsEind strain (P < 0.05). When pqsE was induced with IPTG, PphzA2 activity was triggered and further increased by addition of exogenous PQS (P < 0.05) (Figure 6). These data suggest that PqsE rather than PqsR is required to positively regulate PphzA2, as it was for PphzA1. Hardly any increase in the expression of the PphzA2 promoter was observed in the pqsEind strains carrying additional lasR and rhlR deletions, either in the absence or presence of IPTG and/or PQS, suggesting that LasR, RhlR and PqsE are all key for PphzA2 transcription.
We have shown that RsaL has a positive effect on PphzA2 (Figures 1, 2). The rsaL promoter is positively regulated by LasR, hence in the lasR deletion mutant low levels of RsaL would be expected, which in turn should have a negative impact on PphzA2 activity. Therefore, we hypothesized that LasR is an indirect activator of PphzA2 acting via RsaL, and to test this we transformed the lasR mutant strain with the inducible pRsaL plasmid. As LasR affects the activity of the rhl and pqs QS systems, exogenous PQS was also added to increase the activity of the rhl and pqs QS systems.
The result of these experiments suggest that LasR is an indirect activator of PphzA2, since an increase in PphzA2 activity in the lasR mutant carrying pRsaL was observed, compared with the lasR mutant. Addition of exogenous PQS further increased PphzA2 activity to the level of the wild type PAO1-N level when pRsaL was induced with IPTG (P < 0.05) (Figure 8), growth data Figure S9.
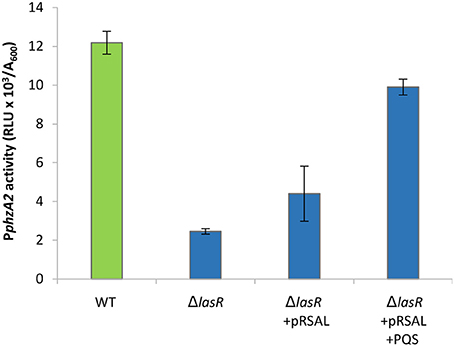
Figure 8. PphzA2 activity in a lasR mutant is restored by overexpressing RsaL and supplementing with exogenous PQS. Maximal promoter activity of PphzA2-lux in the wild type PAO1-N, lasR and lasR deletion carrying pRsaL in the presence of PQS. Promoter activities are normalized by cell density (A600).
These data suggest that RsaL alone is unable to induce the PphzA2 promoter to wild type levels and must be working in conjunction with other regulatory elements. To gain further evidence that LasR is an indirect activator of PphzA2 and investigate the requirement of PqsE and RhlR to activate PphzA2, we introduced pRsaL in the pqsEind and pqsEind strains with additional lasR and rhlR deletions. When RsaL and PqsE expression was induced in these strains with IPTG, PphzA2 activity of the pqsEind ΔlasR strain carrying pRsaL was significantly increased (P < 0.05) compared to the pqsEind ΔlasR strain and PphzA2 activity was comparable to the induced pqsEind strain. No PphzA2 activity was observed in the pqsEind strain carrying an additional rhlR deletion, confirming that RsaL, RhlR, and PqsE are all required to trigger transcription of the phz2 operon (Figure 9). The growth data for this experiment is shown in Figure S10.
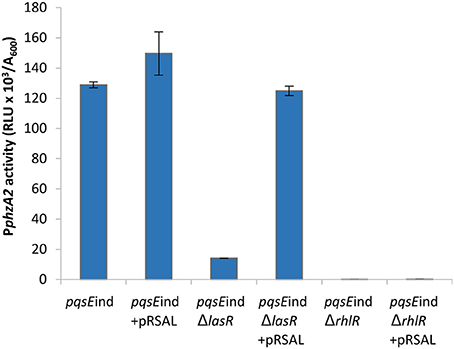
Figure 9. RsaL can activate PphzA2 in the absence of LasR when pqsE is induced and RhlR is present. Maximal promoter activity of PphzA2-lux in the pqsEind strain and pqsEind strain with additional lasR or rhlR mutations and in the presence or absence of pRsaL. All strains were induced with 0.1 mM IPTG. Promoter activities are normalized by cell density (A600).
Here it has been demonstrated that the QS regulators LasR, RhlR, RsaL and the enzyme PqsE are all involved in controlling the expression of both phenazine operons phz1 and phz2 in P. aeruginosa with some differences. Initially it was unclear which regulators played a direct role in activating each operon and which were indirect because of the hierarchal structure of the QS network in rich media (Figure 1). A combination of the deletion of specific genes, the inducible expression of specific QS regulators and/or exogenous provision of QS signal molecules has allowed us to tease out which regulators are direct activators and which can be considered indirect because of their effect upon the QS network (Figures 3–9). Although rich media is not representative of the natural environment in which P. aeruginosa is found our experiments have closed an unanswered question of which QS regulators directly control each phz operon.
The results obtained allow us to postulate a model by which the QS cascade interacts and controls phenazine production in planktonic cultures (Figure 10).
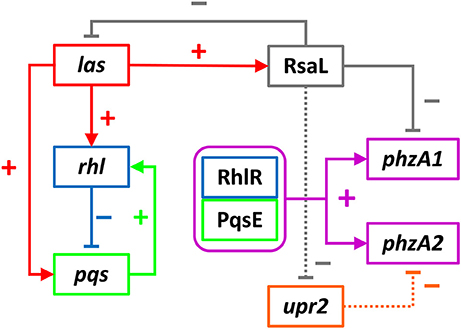
Figure 10. Interactions of the QS cascade to control PphzA1 and PphzA2 activity. The auto-inducing las system positively regulates the rhl and pqs system via its regulator LasR, as well as the QS repressor RsaL. The rhl system represses the pqs system, which in turn has a positive effect on the rhl system. These two systems interact via RhlR and PqsE (purple box) which are both required to induce PphzA1 and PphzA2. The QS repressor RsaL is a repressor of PphzA1 but an indirect activator of PphzA2. We hypothesize that this is achieved by its ability to repress the promoter of an Unknown Phenazine Regulator of phzA2 (orange box), that remains to be identified. Through this mechanism production of PCA from phz1 can be switched to phz2 and vice versa. Positive interactions are indicated by arrows whereas negative interactions are indicated by T-bars.
Evidence has been presented showing that RhlR is a positive regulator for both operons and also that PqsE must be present to induce each operon. This is not surprising as it has been previously reported that PqsE and RhlR are both required for pyocyanin production (Farrow et al., 2008). It could be the case that all genes in the rhl regulon may be co-dependent upon PqsE as the production of the RhlR controlled genes lasB and rhlA are enhanced in the presence of PqsE (Farrow et al., 2008; Rampioni et al., 2010). The data presented show that PqsE rather than PqsR is required to activate the transcription of both phenazine biosynthesis operons. It was demonstrated by Recinos et al. (2012) that HHQ plays a role in activation of PphzA2 under anaerobic conditions. HHQ would inevitably increase the levels of PqsE as pqsABCDE is a direct target for PqsR when bound to either HHQ or PQS (Fletcher et al., 2007; Rampioni et al., 2016). As molecular oxygen is required to convert HHQ to PQS (Schertzer et al., 2010) it would appear that under anaerobic conditions PqsE can still be produced when the signal HHQ binds PqsR, emphasizing that PqsE is important for activating the phenazine operons under both aerobic and anaerobic conditions.
In rich media, LasR drives expressions of the rhl and pqs systems, which then interact through RhlR and PqsE to activate the phenazine operon promoters. A deletion of lasR caused a reduction in PphzA1 activity, which further demonstrates that in rich media the QS cascade has a hierarchal structure. This is in accordance with the work of others who have demonstrated that the loss of LasR results in a delay in production of pyocyanin (Dekimpe and Deziel, 2009; Cabeen, 2014). LasR also drives expression from PrsaL and in turn RsaL represses PphzA1, PlasI and its own production. In vitro protein-DNA interaction experiments revealed that RsaL binds to the PphzA1 promoter at a region encompassing the −10 sequence (Rampioni et al., 2007; Sun et al., 2017), hence RsaL directly exerts a repressive effect on phz1 expression. Moreover, RsaL was shown to exert an indirect repressive effect on PphzA1 by increasing the expression of the PphzA1-repressor CdpR (Sun et al., 2017). It has been hypothesized by Rampioni et al. (2007) that RsaL maintains signal homeostasis by repressing PlasI and in the context of phenazine production could provide a similar feature (Bondi et al., 2017). RsaL could act to keep PphzA1 inactive until the rhl and pqs systems are interacting before commencing transcription, thereby creating a checkpoint in the system. The QS signal molecule PQS has multiple functions as it can bind iron and also act as an anti-oxidant (Diggle et al., 2007; Häussler and Becker, 2008). In the presence of oxygen, pyocyanin generates reactive oxygen species (ROS) (Rada and Leto, 2013). Hence this checkpoint could function to allow adequate PQS to be produced and reduce deleterious effects of ROS produced by pyocyanin before triggering transcription of phz1.
It is thought that RsaL has a secondary function and strong evidence that RsaL can repress PphzA1 but indirectly induce PphzA2 has been presented. Previous studies have failed to demonstrate an interaction between RsaL and a DNA probe encompassing the PphzA2 promoter region (Rampioni et al., 2007; Sun et al., 2017), suggesting that the positive effect exerted by RsaL on phz2 expression is not direct. RsaL increases the expression of the PphzA1-repressor CdpR, but a ChIP-seq assay did not show any interaction between CdpR and the PphzA2 promoter region in strain PA1201 (Zhao et al., 2016), suggesting that CdpR is not involved in the RsaL-mediated activation of PphzA2. The positive effect of RsaL on PphzA2 is probably achieved via an unidentified phenazine biosynthesis gene regulator, which we termed Unidentified Phenazine Regulator of phzA2 (Upr2). Although this regulator has not been identified, data presented thus far strongly imply the existence of this additional regulator, which in turn is controlled by the QS repressor RsaL.
We hypothesize that Upr2 is a PphzA2 repressor and its expression could be repressed by RsaL. If Upr2 had a positive effect upon PphzA2 we would expect that in an rhlR mutant some PphzA2 activity would be observed. We show that when RsaL and PqsE are present but rhlR is mutated, no PphzA2 activity was observed (Figure 9) making it unlikely that Upr2 is a positive regulator. It is likely that when the QS network is activated by LasR, that RsaL represses the promoter of upr2 and as Upr2 is turned over and diluted through cell division that PphzA2 can be triggered by RhlR in conjunction with PqsE, since both the rhl and pqs systems are positively regulated by LasR. In these experiments we observed significantly less activity from PphzA2 compared with PphzA1, further suggesting that the PphzA2 promoter is tightly controlled by a repressor. Upr2 could have its own regulon which may have significant overlap with that of the las regulon. We found that phz2 transcription can be triggered in a lasR mutant when the rsaL deletion is complemented and PQS added exogenously to stimulate RhlR and PqsE production (Figure 8). Hence it is likely that some of the genes identified as lasR- or rsaL- specific in comparative transcriptome studies could belong to the upr2 regulon.
We hypothesize that P. aeruginosa can switch PCA production from phz1 to phz2 when RsaL levels are elevated and from phz2 to phz1 when RsaL is absent. This switch could be related to a reduction in oxygen availability and an increase in oxidative stress as the population size increases, however, this remains to be investigated.
PCA is converted to PYO via the action of PhzM and PhzS. The phzM gene is located directly upstream of phzA1 and the intergenic region between these has two predicted lux boxes (Whiteley and Greenberg, 2001). The lux boxes are flanked by two rsaL binding sites (Rampioni et al., 2007) which suggests that phz1 and phzM are controlled in a similar manner. In the study by Rampioni et al. (2007) a microarray assay was used to identify the rsaL regulon. In that work it was discovered that both phzM and phzS were up regulated in the rsaL mutant compared with the wild type, suggesting that RsaL represses both genes. Here we present evidence that RsaL also represses PphzA1. As phzM, phzS, and phz1 are all required to produce PYO, which in turn contributes toward oxidative stress. It is conceivable that when oxidative stress is high, RsaL can repress phz1, phzM and phzS but maintain PCA production by indirectly activating phz2. Evidence to support this hypothesis was provided when the oxidative stress response regulator OxyR was shown to bind the promoter of rsaL (Wei et al., 2012). A previous study of the OxyR regulon showed that when this regulator is mutated, pyocyanin levels increase, suggesting that OxyR can repress pyocyanin production and this could be achieved through RsaL (Vinckx et al., 2010).
One of the proposed main functions of phenazines is to cycle electrons which allows P. aeruginosa to continue respiration in microaerophilic environments by controlling the intracellular redox state (Dietrich et al., 2013). Switching off PYO production would therefore cause a problem under these conditions. Unlike PYO, PCA can donate electrons to iron (III) rather than oxygen, hence maintaining redox homeostasis without producing ROS (Wang and Newman, 2008; Wang et al., 2011). PYO may be used in addition to PCA to cycle electrons as O2 is a better electron accepter than iron. Through this switch, phenazine production may continue while lowering oxidative stress on the bacterium and maintaining redox balance.
SHi, SHe, GR, MF, PW, and MC designed the study and analyzed the data. SHi, GR, and MF conducted the experiments. SHi, SHe, GR, and MC wrote the manuscript. All authors reviewed the manuscript.
This work was supported by the Biotechnology and Biological Sciences Research Council [grant number BB/F014392/1] and the Engineering and Physical Sciences Research Council [grant number 977836].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2018.00252/full#supplementary-material
Atkinson, S., Chang, C. Y., Patrick, H. L., Buckley, C. M., Wang, Y., Sockett, R. E., et al. (2008). Functional interplay between the Yersinia pseudotuberculosis YpsRI and YtbRI quorum sensing systems modulates swimming motility by controlling expression of flhDC and fliA. Mol. Microbiol. 69, 137–151. doi: 10.1111/j.1365-2958.2008.06268.x
Balasubramanian, D., Schneper, L., Kumari, H., and Mathee, K. (2013). A dynamic and intricate regulatory network determines Pseudomonas aeruginosa virulence. Nucleic Acids Res. 41, 1–20. doi: 10.1093/nar/gks1039
Bertani, I., Rampioni, G., Leoni, L., and Venturi, V. (2007). The Pseudomonas putida Lon protease is involved in N-acyl homoserine lactone quorum sensing regulation. BMC Microbiol. 7:71. doi: 10.1186/1471-2180-7-71
Bjarnsholt, T., and Givskov, M. (2007). The role of quorum sensing in the pathogenicity of the cunning aggressor Pseudomonas aeruginosa. Anal. Bioanal. Chem. 387, 409–414. doi: 10.1007/s00216-006-0774-x
Bondí, R., Longo, F., Messina, M., D'Angelo, F., Visca, P., Leoni, L., et al. (2017). The multi-output incoherent feedforward loop constituted by the transcriptional regulators LasR and RsaL confers robustness to a subset of quorum sensing genes in Pseudomonas aeruginosa. Mol. Biosyst. 13, 1080–1089. doi: 10.1039/c7mb00040e
Brouwer, S., Pustelny, C., Ritter, C., Klinkert, B., Narberhaus, F., and Häussler, S. (2014). The PqsR and RhlR transcriptional regulators determine the level of Pseudomonas quinolone signal synthesis in Pseudomonas aeruginosa by producing two different pqsABCDE mRNA isoforms. J. Bacteriol. 196, 4163–4171. doi: 10.1128/jb.02000-14
Cabeen, M. T. (2014). Stationary phase-specific virulence factor overproduction by a lasR mutant of Pseudomonas aeruginosa. PLoS ONE 9:e88743. doi: 10.1371/journal.pone.0088743
Choi, K. H., Kumar, A., and Schweizer, H. P. (2006). A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 64, 391–397. doi: 10.1016/j.mimet.2005.06.001
Chugani, S. A., Whiteley, M., Lee, K. M., D'Argenio, D., Manoil, C., and Greenberg, E. P. (2001). QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci U.S.A. 98, 2752–2757. doi: 10.1073/pnas.051624298.
Cornforth, D. M., Popat, R., McNally, L., Gurney, J., Scott-Phillips, T. C., Ivens, A., et al. (2014). Combinatorial quorum sensing allows bacteria to resolve their social and physical environment. Proc. Nat. Acad. Sci. U.S.A. 111, 4280–4284. doi: 10.1073/pnas.1319175111.
de Kievit, T., Kakai, Y., Register, J. K., Pesci, E. C., and Iglewski, B. H. (2002). Role of the Pseudomonas aeruginosa las and rhl quorum-sensing systems in rhlI regulation. Fems Microbiol. Lett. 212, 101–106. doi: 10.1016/S0378-1097(02)00735-8
de Kievit, T., Seed, P. C., Nezezon, J., Passador, L., and Iglewski, B. H. (1999). RsaL, a novel repressor of virulence gene expression in Pseudomonas aeruginosa. J. Bacteriol. 181, 2175–2184
Dekimpe, V., and Déziel, E. (2009). Revisiting the quorum-sensing hierarchy in Pseudomonas aeruginosa: the transcriptional regulator RhlR regulates LasR-specific factors. Microbiology 155, 712–723. doi: 10.1099/mic.0.022764-0
Dietrich, L. E., Okegbe, C., Price-Whelan, A., Sakhtah, H., Hunter, R. C., and Newman, D. K. (2013). Bacterial community morphogenesis is intimately linked to the intracellular redox state. J. Bacteriol. 195, 1371–1380. doi: 10.1128/jb.02273-12
Diggle, S. P., Matthijs, S., Wright, V. J., Fletcher, M. P., Chhabra, S. R., Lamont, I. L., et al. (2007). The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem. Biol. 14, 87–96. doi: 10.1016/j.chembiol.2006.11.014
Diggle, S. P., Winzer, K., Chhabra, S. R., Worrall, K. E., Cámara, M., and Williams, P. (2003). The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol. Microbiol. 50, 29–43. doi: 10.1046/j.1365-2958.2003.03672.x
Dötsch, A., Eckweiler, D., Schniederjans, M., Zimmermann, A., Jensen, V., Scharfe, M., et al. (2012). The Pseudomonas aeruginosa transcriptome in planktonic cultures and static biofilms using RNA sequencing. PLoS ONE 7:e31092. doi: 10.1371/journal.pone.0031092
Drees, S. L., and Fetzner, S. (2015). PqsE of Pseudomonas aeruginosa acts as pathway-specific thioesterase in the biosynthesis of alkylquinolone signaling molecules. Chem. Biol. 22, 611–618. doi: 10.1016/j.chembiol.2015.04.012
Dulcey, C. E., Dekimpe, V., Fauvelle, D. A., Milot, S., Groleau, M. C., Doucet, N., et al. (2013). The end of an old hypothesis: the pseudomonas signaling molecules 4-hydroxy-2-alkylquinolines derive from fatty acids, not 3-ketofatty acids. Chem. Biol. 20, 1481–1491. doi: 10.1016/j.chembiol.2013.09.021
Farrow, J. M., Sund, Z. M., Ellison, M. L., Wade, D. S., Coleman, J. P., and Pesci, E. C. (2008). PqsE functions independently of PqsR-pseudomonas quinolone signal and enhances the rhl quorum-sensing system. J. Bacteriol. 190, 7043–7051. doi: 10.1128/jb.00753-08
Fletcher, M. P., Diggle, S. P., Crusz, S. A., Chhabra, S. R., Cámara, M., and Williams, P. (2007). A dual biosensor for 2-alkyl-4-quinolone quorum-sensing signal molecules. Environ. Microbiol. 9, 2683–2693. doi: 10.1111/j.1462-2920.2007.01380.x
FRANK, L. H., and DEMOSS, R. D. (1959). On the biosynthesis of pyocyanine. J. Bacteriol. 77, 776–782.
Gallagher, L. A., McKnight, S. L., Kuznetsova, M. S., Pesci, E. C., and Manoil, C. (2002). Functions required for extracellular quinolone signaling by Pseudomonas aeruginosa. J. Bacteriol. 184, 6472–6480. doi: 10.1128/jb.184.23.6472-6480.2002
Guzman, L. M., Belin, D., Carson, M. J., and Beckwith, J. (1995). Tight regulation, modulation, and high-level expression by vectors containing the arabinose P-bad promoter. J. Bacteriol. 177, 4121–4130.
Harrison, F., Muruli, A., Higgins, S., and Diggle, S. P. (2014). Development of an ex vivo porcine lung model for studying growth, virulence, and signaling of Pseudomonas aeruginosa. Infect. Immun. 82, 3312–3323. doi: 10.1128/iai.01554-14
Häussler, S., and Becker, T. (2008). The Pseudomonas quinolone signal (PQS) balances life and death in Pseudomonas aeruginosa populations. PLoS Pathog. 4:e1000166. doi: 10.1371/journal.ppat.1000166
Hazan, R., He, J., Xiao, G., Dekimpe, V., Apidianakis, Y., Lesic, B., et al. (2010). Homeostatic interplay between bacterial cell-cell signaling and iron in virulence. PLoS Pathog. 6:e1000810. doi: 10.1371/journal.ppat.1000810
Heeb, S., Fletcher, M. P., Chhabra, S. R., Diggle, S. P., Williams, P., and Cámara, M. (2011). Quinolones: from antibiotics to autoinducers. FEMS Microbiol. Rev. 35, 247–274. doi: 10.1111/j.1574-6976.2010.00247.x
Hoang, T. T., Karkhoff-Schweizer, R. R., Kutchma, A. J., and Schweizer, H. P. (1998). A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 212, 77–86. doi: 10.1016/s0378-1119(98)00130-9
Hoang, T. T., Kutchma, A. J., Becher, A., and Schweizer, H. P. (2000). Integration-proficient plasmids for Pseudomonas aeruginosa: site-specific integration and use for engineering of reporter and expression strains. Plasmid 43, 59–72 doi: 10.1006/plas.1999.1441
Ledgham, F., Ventre, I., Soscia, C., Foglino, M., Sturgis, J. N., and Lazdunski, A. (2003). Interactions of the quorum sensing regulator QscR: interaction with itself and the other regulators of Pseudomonas aeruginosa LasR and RhlR. Mol. Microbiol. 48, 199–210 doi: 10.1046/j.1365-2958.2003.03423.x
Lee, J. H., Lequette, Y., and Greenberg, E. P. (2006). Activity of purified QscR, a Pseudomonas aeruginosa orphan quorum-sensing transcription factor. Mole. Microbiol. 59, 602–609. doi: 10.1111/j.1365-2958.2005.04960.x
Lequette, Y., Lee, J. H., Ledgham, F., Lazdunski, A., and Greenberg, E. P. (2006). A distinct QscR regulon in the Pseudomonas aeruginosa quorum-sensing circuit. J. Bacteriol. 188, 3365–3370. doi: 10.1128/jb.188.9.3365-3370.2006
Liang, H., Duan, J., Sibley, C. D., Surette, M. G., and Duan, K. (2011). Identification of mutants with altered phenazine production in Pseudomonas aeruginosa. J. Med. Microbiol. 60, 22–34. doi: 10.1099/jmm.0.022350-0
Lu, J., Huang, X., Li, K., Li, S., Zhang, M. Y., Wang, Y., et al. (2009). LysR family transcriptional regulator PqsR as repressor of pyoluteorin biosynthesis and activator of phenazine-1-carboxylic acid biosynthesis in Pseudomonas sp M18. J. Biotechnol. 143, 1–9. doi: 10.1016/j.jbiotec.2009.06.008
Maura, D., Hazan, R., Kitao, T., Ballok, A. E., and Rahme, L. G. (2016). Evidence for direct control of virulence and defense gene circuits by the Pseudomonas aeruginosa Quorum Sensing Regulator, MvfR. Sci. Rep. 6:34083. doi: 10.1038/srep34083
Mavrodi, D. V., Blankenfeldt, W., and Thomashow, L. S. (2006). Phenazine compounds in fluorescent Pseudomonas spp. biosynthesis and regulation. Ann. Rev. Phytopathol. 44, 417–445. doi: 10.1146/annurev.phyto.44.013106.145710
Mavrodi, D. V., Bonsall, R. F., Delaney, S. M., Soule, M. J., Phillips, G., and Thomashow, L. S. (2001). Functional analysis of genes for biosynthesis of pyocyanin and phenazine-1-carboxamide from Pseudomonas aeruginosa PAO1. J. Bacteriol. 183, 6454–6465. doi: 10.1128/jb.183.21.6454-6465.2001
Mavrodi, D. V., Peever, T. L., Mavrodi, O. V., Parejko, J. A., Raaijmakers, J. M., Lemanceau, P., et al. (2010). Diversity and evolution of the phenazine biosynthesis pathway. Appl. Environ. Microbiol. 76, 866–879. doi: 10.1128/aem.02009-09
McKnight, S. L., Iglewski, B. H., and Pesci, E. C. (2000). The Pseudomonas quinolone signal regulates rhl quorum sensing in Pseudomonas aeruginosa. J. Bacteriol. 182, 2702–2708. doi: 10.1128/jb.182.10.2702-2708.2000
Medina, G., Juárez, K., Díaz, R., and Sóberon-Chaávez, G. (2003). Transcriptional regulation of Pseudomonas aeruginosa rhlR, encoding a quorum-sensing regulatory protein. Microbiology 149, 3073–3081. doi: 10.1099/mic.0.26282-0
Milton, D. L., O'Toole, R., Horstedt, P., and Wolf-Watz, H. (1996). Flagellin A is essential for the virulence of Vibrio anguillarum. J. Bacteriol. 178, 1310–1319.
Pesci, E. C., Pearson, J. P., Seed, P. C., and Iglewski, B. H. (1997). Regulation of las and rhl quorum sensing in Pseudomonas aeruginosa. J. Bacteriol. 179, 3127–3132.
Rada, B., and Leto, T. L. (2013). Pyocyanin effects on respiratory epithelium: relevance in Pseudomonas aeruginosa airway infections. Trends Microbiol. 21, 73–81. doi: 10.1016/j.tim.2012.10.004
Rampioni, G., Bertani, I., Zennaro, E., Polticelli, F., Venturi, V., and Leoni, L. (2006). The quorum-sensing negative regulator RsaL of Pseudomonas aeruginosa binds to the lasI promoter. J. Bacteriol. 188, 815–819. doi: 10.1128/jb.188.2.815-819.2006
Rampioni, G., Falcone, M., Heeb, S., Frangipani, E., Fletcher, M. P., Dubern, J. F., et al. (2016). Unravelling the genome-wide contributions of specific 2-Alkyl-4-quinolones and PqsE to quorum sensing in Pseudomonas aeruginosa. PLOS Pathog. 12:e1006029. doi: 10.1371/journal.ppat.1006029
Rampioni, G., Pustelny, C., Fletcher, M. P., Wright, V. J., Bruce, M., Rumbaugh, K. P., et al. (2010). Transcriptomic analysis reveals a global alkyl-quinolone-independent regulatory role for PqsE in facilitating the environmental adaptation of Pseudomonas aeruginosa to plant and animal hosts. Environ. Microbiol. 12, 1659–1673. doi: 10.1111/j.1462-2920.2010.02214.x
Rampioni, G., Schuster, M., Greenberg, E. P., Bertani, I., Grasso, M., Venturi, V., et al. (2007). RsaL provides quorum sensing homeostasis and functions as a global regulator of gene expression in Pseudomonas aeruginosa. Mol. Microbiol. 66, 1557–1565. doi: 10.1111/j.1365-2958.2007.06029.x
Recinos, D. A., Sekedat, M. D., Hernandez, A., Cohen, T. S., Sakhtah, H., Prince, A. S., et al. (2012). Redundant phenazine operons in Pseudomonas aeruginosa exhibit environment-dependent expression and differential roles in pathogenicity. Proc. Nat. Acad. Sci. U.S.A. 109, 19420–19425. doi: 10.1073/pnas.1213901109
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Schertzer, J. W., Brown, S. A., and Whiteley, M. (2010). Oxygen levels rapidly modulate Pseudomonas aeruginosa social behaviours via substrate limitation of PqsH. Mol. Microbiol. 77, 1527–1538. doi: 10.1111/j.1365-2958.2010.07303.x
Schuster, M., and Greenberg, E. P. (2007). Early activation of quorum sensing in Pseudomonas aeruginosa reveals the architecture of a complex regulon. BMC Genomics 8:287. doi: 10.1186/1471-2164-8-287
Schuster, M., Lostroh, C. P., Ogi, T., and Greenberg, E. P. (2003). Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: a transcriptome analysis. J. Bacteriol. 185, 2066–2079. doi: 10.1128/jb.185.7.2066-2079.2003
Schuster, M., Sexton, D. J., Diggle, S. P., and Greenberg, E. P. (2013). Acyl-homoserine lactone quorum sensing: from evolution to application. Annu. Rev. Microbiol. 67, 43–63. doi: 10.1146/annurev-micro-092412-155635
Smith, R. S., and Iglewski, B. H. (2003). Pseudomonas aeruginosa quorum sensing as a potential antimicrobial target. J. Clin. Invest. 112, 1460–1465. doi: 10.1172/jci20364
Sun, S., Chen, B., Jin, Z. J., Zhou, L., Fang, Y. L., Thawai, C., et al. (2017). Characterization of the multiple molecular mechanisms underlying RsaL control of phenazine-1-carboxylic acid biosynthesis in the rhizosphere bacterium Pseudomonas aeruginosa PA1201. Mol. Microbiol. 104, 931–947. doi: 10.1111/mmi.13671
Sun, S., Zhou, L., Jin, K., Jiang, H., and He, Y. W. (2016). Quorum sensing systems differentially regulate the production of phenazine-1-carboxylic acid in the rhizobacterium Pseudomonas aeruginosa PA1201. Sci. Rep. 6:30352. doi: 10.1038/srep30352
Trunk, K., Benkert, B., Quäck, N., Münch, R., Scheer, M., Garbe, J., et al. (2010). Anaerobic adaptation in Pseudomonas aeruginosa: definition of the Anr and Dnr regulons. Environ. Microbiol. 12, 1719–1733. doi: 10.1111/j.1462-2920.2010.02252.x
Vinckx, T., Wei, Q., Matthijs, S., and Cornelis, P. (2010). The Pseudomonas aeruginosa oxidative stress regulator OxyR influences production of pyocyanin and rhamnolipids: protective role of pyocyanin. Microbiology 156, 678–686. doi: 10.1099/mic.0.031971-0
Voisard, C., Bull, C., Keel, C., Laville, J., Maurhofer, M., Schnider, U., et al. (1994). “Biocontrol of root diseases by Pseudomonas fluorescens CHA0: current concepts and experimental approaches,” in Molecular Ecology of Rhizosphere Microorganisms, eds F. O'Gara, D. N. Dowling, and B. Boesten (Weinheim: VCH Publishers), 67–89. doi: 10.1002/9783527615810.ch6
Wade, D. S., Calfee, M. W., Rocha, E. R., Ling, E. A., Engstrom, E., Coleman, J. P., et al. (2005). Regulation of Pseudomonas quinolone signal synthesis in Pseudomonas aeruginosa. J. Bacteriol. 187, 4372–4380. doi: 10.1128/JB.187.13.4372-4380.2005
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I., and Iglewski, B. H. (2003). Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J. Bacteriol. 185, 2080–2095. doi: 10.1128/jb.185.7.2080-2095.2003
Wang, Y., and Newman, D. K. (2008). Redox reactions of phenazine antibiotics with ferric (hydr)oxides and molecular oxygen. Environ. Sci. Tech. 42, 2380–2386. doi: 10.1021/es702290a
Wang, Y., Wilks, J. C., Danhorn, T., Ramos, I., Croal, L., and Newman, D. K. (2011). Phenazine-1-Carboxylic acid promotes bacterial biofilm development via ferrous iron acquisition. J. Bacteriol. 193, 3606–3617. doi: 10.1128/jb.00396-11
Wei, Q., Minh, P. N., Dötsch, A., Hildebrand, F., Panmanee, W., Elfarash, A., et al. (2012). Global regulation of gene expression by OxyR in an important human opportunistic pathogen. Nucleic Acids Res. 40, 4320–4333. doi: 10.1093/nar/gks017
Westfall, L. W., Luna, A. M., San Francisco, M., Diggle, S. P., Worrall, K. E., Williams, P., et al. (2004). The Pseudomonas aeruginosa global regulator MvaT specifically binds to the ptxS upstream region and enhances ptxS expression. Microbiology 150(Pt 11), 3797–3806. doi: 10.1099/mic.0.27270-0
Whiteley, M., and Greenberg, E. P. (2001). Promoter specificity elements in Pseudomonas aeruginosa quorum-sensing-controlled genes. J. Bacteriol. 183, 5529–5534. doi: 10.1128/jb.183.19.5529-5534.2001
Winsor, G. L., Lam, D. K., Fleming, L., Lo, R., Whiteside, M. D., Yu, N. Y., et al. (2011). Pseudomonas genome database: improved comparative analysis and population genomics capability for Pseudomonas genomes. Nucl. Acids Res. 39(Suppl 1), D596–D600. doi: 10.1093/nar/gkq869
Xiao, G., He, J., and Rahme, L. G. (2006). Mutation analysis of the Pseudomonas aeruginosa mvfR and pqsABCDE gene promoters demonstrates complex quorum-sensing circuitry. Microbiology 152, 1679–1686. doi: 10.1099/mic.0.28605-0
Keywords: Pseudomonas aeruginosa, phenazines, pyocyanin, quorum sensing, LasR, RhlR, RsaL, PqsE
Citation: Higgins S, Heeb S, Rampioni G, Fletcher MP, Williams P and Cámara M (2018) Differential Regulation of the Phenazine Biosynthetic Operons by Quorum Sensing in Pseudomonas aeruginosa PAO1-N. Front. Cell. Infect. Microbiol. 8:252. doi: 10.3389/fcimb.2018.00252
Received: 10 April 2018; Accepted: 03 July 2018;
Published: 23 July 2018.
Edited by:
Maria Tomas, Complexo Hospitalario Universitario A Coruña, SpainReviewed by:
Rodolfo García-Contreras, Universidad Nacional Autónoma de México, MexicoCopyright © 2018 Higgins, Heeb, Rampioni, Fletcher, Williams and Cámara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel Cámara, bWlndWVsLmNhbWFyYUBub3R0aW5naGFtLmFjLnVr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.