 Grant Cox
Grant Cox Michihiro Kobayashi
Michihiro Kobayashi Brian D. Rudd
Brian D. Rudd Momoko Yoshimoto
Momoko Yoshimoto
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol., 12 March 2025
Sec. Stem Cell Research
Volume 13 - 2025 | https://doi.org/10.3389/fcell.2025.1555877
This article is part of the Research TopicAdvancements in Hematopoietic Stem Cell Proliferation and Self-Renewal MaintenanceView all 6 articles
Hematopoietic stem cells (HSCs) provide all kinds of blood cells for life while maintaining self-renewal ability. During development, HSCs are first produced in the mouse embryo around embryonic day (E) 11. At this time, only one or two transplantable HSCs can be detected per embryo. Then, HSCs migrate to the fetal liver, where the number of HSCs rapidly increases, showing enhanced self-renewal ability. After birth, a transition occurs from the rapidly proliferating fetal HSCs to the more slowly dividing adult HSCs, which ends by 3–4 weeks of age. It is known that fetal HSCs express distinct surface markers and transcriptomes and produce a variety of distinct immune cells that are not made by adult HSCs. Accumulating evidence indicates that the ontogeny of the hematopoietic system is driven by a highly conserved and developmentally regulated RNA binding protein known as Lin28b. Lin28b is predominantly expressed in the fetal hematopoietic stem and progenitor cells (HSPCs) and regulates the developmental switch from fetal to adult HSCs. In this review, we will provide an overview of how Lin28b regulates the expansion and differentiation of HSCs in early life. These insights can be taken into consideration when developing ex vivo HSC expansion utilizing such physiological characteristics of HSCs.
Ex vivo HSC expansion is a longstanding desire in the hematology field, since it would enable more efficient transplantation therapy with larger numbers of HSCs. However, it is challenging to expand HSCs. First, HSCs seldomly divide. In fact, the fetal to neonatal stage is the only time when HSCs expand in number. Second, once HSCs are transferred to an in vitro environment, they either quickly differentiate and lose the ability to self-renew or die without the appropriate signals or environment. Starting from traditional stromal cell cocultures and cytokine combinations, recent advancements using small molecules (such as UM171) and albumin alternatives (such as polyvinyl alcohol) have improved strategies for HSC expansion (Ueda et al., 2000; Fares et al., 2014; Bastani et al., 2023; Rubio-Lara et al., 2023; Sakurai et al., 2023; Meaker and Wilkinson, 2024). However, these methods also induce HSC differentiation, and it is not clear that long-term engrafting HSCs are really expanded (Bastani et al., 2023). The mechanisms of HSC expansion and the potential long-term effects of the cytokines and molecules on HSC function are not well understood. Therefore, it is critically important to better understand the normal physiological settings in which HSCs expand. Notably, self-renewing HSCs expand considerably in the fetal liver (Ema and Nakauchi, 2000), while their differentiation is minimal (Yokomizo et al., 2022; Kobayashi et al., 2023). Thus, understanding the mechanism of HSC expansion in the fetus is key for translating ex-vivo HSC expansion. In this paper, we will provide an overview of the mechanisms by which HSCs expand in early life and explain the biological differences between fetal and adult HSCs. Also, we will address how manipulation of HSCs into “fetal-type HSCs” impacts on hematopoiesis and their differentiation capability, specifically focusing on Lin28b expression, which regulates the function of fetal HSCs. A better understanding of these mechanisms will provide us with key information needed to improve HSC expansion strategies.
HSCs have the ability to self-renew and differentiate into multiple lineages, allowing them to produce all types of blood cells and maintain blood homeostasis for life. While HSCs are first produced from special endothelial cells (ECs), referred to as hemogenic ECs, in the aortic region of the embryo around E11.5, multiple waves of hematopoiesis proceed before HSC emergence (Medvinsky and Dzierzak, 1996; Chen et al., 2009; Kobayashi and Yoshimoto, 2023). As early as E7.5, primitive type erythroid and myeloid cell production occurs in the extraembryonic yolk sac (YS), followed by definitive type erythro-myeloid progenitor (EMP) production (Lux et al., 2008). At E8.5, lymphoid and multipotent progenitor potentials can be detected in both the YS and the para-aortic region (Godin et al., 1995; Cumano et al., 1996; Nishikawa et al., 1998; Yoshimoto et al., 2011; Yoshimoto et al., 2012). The precursors of HSCs (pre-HSCs) that can become transplantable after aggregation culture are detectable around E10.5–11.5 (Rybtsov et al., 2011; Kobayashi et al., 2019). At this time, the pool of pre-HSCs is comprised of various hematopoietic progenitors that arise from hemogenic ECs, including HSC-independent multipotent progenitors (Dignum et al., 2021; Kobayashi et al., 2023). Some of the pre-HSCs mature into HSCs in the fetal liver, where they proliferate and then migrate to the spleen and bone marrow just before birth. After birth, HSCs are maintained in the bone marrow to provide blood homeostasis for life.
HSCs are defined by transplantation assays, specifically as cells that repopulate the hematopoietic system in irradiated mice. Using transplantation assays, several groups have examined the numbers of HSCs found throughout gestation. Kumaravelu et al. reported that there was only one HSC in the YS, aorta-gonad-mesonephros (AGM) region, and fetal liver at E11.5 (Kumaravelu et al., 2002). At E12.5, they detected approximately two to 3 HSCs in the YS, AGM region, and circulation. However, interestingly, 53 HSCs were detected in the fetal liver at E12.5, indicating a rapid increase in total HSC numbers per embryo. At E13.5, the numbers of HSCs in the YS and AGM region diminished, but the number of HSCs in the fetal liver increased to 260, which is an astonishing 5-fold increase from E12.5. Additionally, the placenta has been reported to contain significant numbers of HSCs at E12.5 – E13.5 (Gekas et al., 2005). Ema et al. reported that HSCs further proliferate in the fetal liver during E12.5 to E16.5 (Ema and Nakauchi, 2000). Based on all these findings, we can estimate that HSCs undergo 1.8 divisions from E12.5 to 14.5, and 3.3 divisions from E12.5 to 16.5.
As mentioned above, while most adult HSCs in the bone marrow are quiescent, which divide only once every 2–5 months, HSCs in the fetus are in cell cycle, which allows them to expand during the transition from the aortic region to the fetal liver, and from the fetal liver to the bone marrow (Cheshier et al., 1999; Wilson et al., 2008). When pre-HSCs arise from ECs at E10.5, half of them are cycling in the G2M phase (Batsivari et al., 2017). Cycling HSCs can be found until ∼3 weeks of age. However, by 4 weeks of age, there is a rapid decrease in proliferation and most of the HSCs are in the resting (G0) phase (Bowie et al., 2006; Copley et al., 2013). This cell cycle change is accompanied by characteristic changes in surface markers (CD34, CD38) (Matsuoka et al., 2001) and gene expression profiles (Sox17, Lin28b) (Kim et al., 2007; Yuan et al., 2012; Copley et al., 2013). How do these cell-cycling statuses and engraftment capabilities correlate?
Batsivari et al. examined the engraftment capabilities of HSCs in different phases of cell-cycling in mid-gestation of the mouse embryo (Batsivari et al., 2017). At E11.5, both cycling and non-cycling (G0/G1) pre-HSCs can engraft in the recipient mice after aggregation culture. In the fetal liver, until E14, the cycling HSCs possess the ability to engraft after transplantation, but after that time, only G0/G1 HSCs exhibit engraftment capability (Batsivari et al., 2017). A report by Bowie et al. precisely examined the repopulating capability of HSPCs based on their cell cycle from E14 fetal liver to up to 10 weeks bone marrow (Bowie et al., 2006). They found a clear transition of HSPC repopulation capability from 3-week bone marrow cells in the G1/S/G2/M phases to 4-week bone marrow cells in the G0 phase. The different results in timing for engraftable cycling HSCs (until E14 vs 3 weeks of age) may be based on which donor population included the G1 phase cells (because G1 phase cells are supposed to engraft efficiently). In any case, what these data suggest is that HSCs enter cell cycle in early life in order to expand but later must go back to G1/G0 status to maintain the ability to engraft.
Another aspect of the developmental changes in engraftment capability may relate to differences in the environment. CXCL12, for example, is a niche factor in the bone marrow that maintains the self-renewal ability of HSCs (Sugiyama et al., 2006). CXCL12 antagonist is usually used to harvest donor peripheral blood stem cells mobilized from the bone marrow (Broxmeyer et al., 2005). Interestingly, Bowie et al. showed that when recipient mice were treated with CXCL12 antagonist, the engraftment capacity of these cycling HSCs from 4 week-old bone marrow was rescued (Bowie et al., 2006), suggesting that it may be advantageous for cycling HSCs not to interact with CXCL12 in the adult bone marrow. However, CXCL12 expression is indispensable for HSC homing to neonatal bone marrow (Ara et al., 2003), and most HSCs are cycling at neonatal stage. It is possible that the structure of neonatal bone marrow is different from the structure of adult bone marrow; thus, the interaction between cycling HSCs and CXCL12 could be different between neonates and adults. Indeed, fetal- and adult-derived HSPCs prefer to engraft in environments of similar ages: fetal HSCs engraft more efficiently in neonatal recipient mice, while adult HSPCs engraft more efficiently in adult recipient mice (Arora et al., 2014).
Consistent with the idea that certain environmental factors may support cycling HSCs, Sigurdsson et al. have shown that bile duct acid can protect cycling HSCs in the fetal liver (Sigurdsson et al., 2016). These studies suggest that the environment plays a pivotal role in protecting cycling HSCs, but further investigation of the role of the environment for HSPCs of different ages is necessary.
Up until now, we have described developmental changes in HSCs that occur during the fetal and neonatal stages of life, including their cell cycle and engraftment capabilities. An important question is how HSC expansion during the fetal and neonatal period is maintained. A number of studies have suggested that the key molecule that distinguishes fetal from adult HSCs is Lin28b.
Lin28 was originally identified in Caenorhabditis elegans as a heterochronic gene, which regulates a variety of developmental events, including patterns of cell division, the lengths of specific cell cycles, and stage-specific terminal differentiation events (Ambros and Horvitz, 1984). Lin28b encodes an RNA binding protein that binds and inhibits the Let-7 miRNA family; thus, Lin28b expression diminishes Let-7 expression and absence of Lin28b expression increases Let-7 expression. (Ambros and Horvitz, 1984; Moss et al., 1997; Yuan et al., 2012). Lin28 plays a critical role during development through the regulation of miRNA biogenesis, transcriptional level DNA regulation, and post-transcriptional direct mRNA binding in various tissues (Polesskaya et al., 2007; Newman et al., 2008; Oshima et al., 2016; Wang et al., 2019). Lin28-let7 axis is known to regulate glucose metabolism via insluin-PI3K-mTOR pathway (Zhu et al., 2011). Lin28 regulates proliferation and self-renew of human and mouse embryonic stem cells (Xu et al., 2009). LIN28B can function with NANOG, OCT4, and SOX2 and both LIN28A and LIN28B are required for efficient reprograming of human inducible Pluripotent Stem Cells (iPSCs) (Zhang et al., 2016). As such, Lin28b is expressed in stem cells in various tissues and cancer stem cells (Shyh-Chang and Daley, 2013). In hematopoietic lineages, with several partners, Lin28b acts as a heterochronic regulator of the switch from fetal to adult gene expression by their diminished expression within the context of HSC self-renewal, lineage commitment, and differentiation (Yuan et al., 2012).
Lin28b has attracted the attention of the hematology field due to its expression in fetal liver HSCs and its ability to enhance self-renewal (Yuan et al., 2012; Copley et al., 2013). Importantly, when Lin28b is overexpressed in adult bone marrow lineage−sca-1+c-kit+ HSPCs (LSK cells) and transplanted into lethally irradiated congenic mice, these Lin28b-expressing HSCs differentiate into fetal-type immune cells, including B-1a cells, γδT cells, and NKT cells, which are not usually produced by adult bone marrow HSCs (Yuan et al., 2012; Wang et al., 2019). Yuan et al. showed that overexpression of Lin28b reprogrammed adult HSCs into fetal-type HSCs (Figure 1) (Yuan et al., 2012). Wang et al. further examined whether overexpressing Lin28b in vivo could generate fetal-like HSCs in adult mice (Wang et al., 2019). For these studies, they used doxycycline inducible Lin28b transgenic mice (iLin28b mice) (Zhu et al., 2011). Gene expression profiles of common lymphoid progenitors (CLPs) from the fetal liver, adult bone marrow, and adult iLin28b bone marrow revealed that Lin28b overexpression alone might not be sufficient to completely reprogram HSPCs into fetal-type HSCs. The Wang group also identified another RNA binding protein, Igf2bp3, that interacts with Lin28b, and these two proteins share multiple target mRNA binding sites. When Igf2bp3 was retrovirally transduced into iLin28 HSPCs (LSK cells) in the adult bone marrow and transplanted into Rag1−/− mice, better repopulation of fetal-type B-1a and marginal zone B cells resulted, indicating the important role of Igf2bp3 in HSPC reprogramming (Wang et al., 2019).
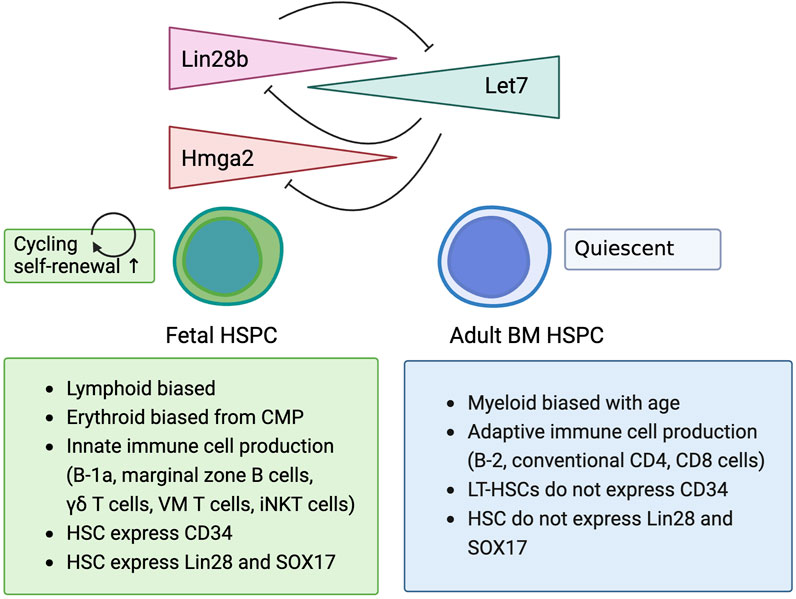
Figure 1. Lin28b regulates biological characteristics of fetal hematopoietic stem/progenitor cell.
Among the Let-7 target genes (Hmga2, Lin28b, Igf2bp3, Igf2bp2, Igf2bp1, and Slc31a1), Hmga2 has been found to follow the expression pattern of Lin28b over the course of development. The HMGA2 protein is a member of the HMGA family of nonhistone chromatin proteins. HMGA2 proteins have been reported to play critical roles in cell proliferation, cell-cycle progression, apoptosis, senescence, and cancer development (Ikeda et al., 2011). Interestingly, when Lin28b levels decrease, Hmga2 levels in HSCs also decrease, along with their self-renewal ability (Copley et al., 2013). When Lin28b is overexpressed in CD45+EPCR+CD48−CD150+ (ESLAM) HSCs from the adult bone marrow, the expression of Hmga2 is also increased, and the overexpression of Lin28b or Hmga2 in adult bone marrow ESLAM HSCs significantly increases their self-renewal ability, resulting in higher engraftment in the recipient mice (Copley et al., 2013). This result also indicates that Lin28b overexpression in adult bone marrow HSCs can induce cell autonomous expansion without the fetal liver environment that supports fetal HSC expansion (Agrawal et al., 2024). In addition, Hmga2 overexpression in human cord blood-derived CD34+ HSPCs also enhances long-term engraftment ability by serial transplantation assays (although with erythroid-biased) (Kumar et al., 2019). Conversely, HSCs from Hgma2 knockout mice exhibit lower self-renewal ability than their counterparts from wildtype mice. These findings indicate that the Lin28b-let7-Hgma2 axis regulates the self-renewal ability of HSCs in both mice and humans.
However, the mechanism by which Hmga2 increases the self-renewal ability of HSCs is still not well known. To address this question, Sashida’s group generated Rosa locus Hmga2 conditional knock-in (KI)-GFP mice to induce Hmga2 by tamoxifen injection (Sun et al., 2022). They induced Hmga2 expression in adult hematopoietic cells by tamoxifen injection and transplanted LSK cells from the Hmga2-GFP-induced and control (YFP-conditional KI) bone marrow cells into the lethally irradiated congenic mice. Interestingly, 10–12 months after transplantation, whereas the frequency and number of donor-derived HSPC populations and spleen lymphocytes showed similar numbers, Hmga2-GFP+ proportions were significantly increased compared to control YFP-KI mice, indicating that Hmga2 expression increased HSPC population while still allowing for differentiation to the mature cells. Furthermore, these Hmga2-overexpressed HSCs showed elevated Igf2bp2 expression at a similar level as fetal liver HSCs. The Sashida group also generated mutant Hmga2-KI mouse lines that lack the N- and C-terminal domains and found that Hmga2 requires its three AT-hooks and the C-terminal linker region to activate the transcription of Igf2bp2 and promote the self-renewal function of HSCs (Sun et al., 2022).
The Sashida group further examined HSC function in mice transplanted with Hgma2-KI or control bone marrow cells under stress conditions caused by 5-FU (Kubota et al., 2024). After 5-FU injection, they found increased frequency of Hmga2-KI lin-EPCR+CD48−CD150+ HSCs and megakaryocyte progenitors. They also showed that Hmga2-KI HSCs went into cell cycle more in the S/G2M phase and divided more compared to WT HSCs in the recipient mice. Three days after 5-FU injection, Hmga2-KI HSCs highly expressed Igf2bp2 and increased cell cycle regulators, but maintained stem cell genes, while repressing genes involved in inflammation and differentiation. Moreover, the Hmga2 protein was phosphorylated by casein kinase 2 (CK2) and promoted to its access and binding to chromatin transcription of anti-inflammatory target genes under stress conditions. These data indicate that Hmga2 can regulate hematopoiesis under stress conditions. However, Hmga2 is not expressed in adult HSCs in the physiological setting, so it is not clear if this Hmga2 function occurs during fetal development, or if Hmga2 is upregulated and functions under stress conditions even in adult wildtype mice. Indeed, the Hmga2 protein was upregulated in WT HSCs 3 days after 5-FU injection.
As noted above, Igf2bp2 is an important co-factor of Lin28 b (Zhu et al., 2011; Shyh-Chang and Daley, 2013). Overexpression of Hmga2 upregulates Igf2bp2 expression in mouse bone marrow HSCs (Sun et al., 2022; Kubota et al., 2024). The role of Igf2bp2 in HSCs has recently been emphasized by several papers (Suo et al., 2022; Yin et al., 2022), especially as it relates to self-renewal ability of HSCs. Since N6-methyladenosine (m6A) is the most abundant modification on mRNA, Zhang’s group extensively investigated its role in adult hematopoiesis (Yin et al., 2022). They found higher levels of m6A in long-term (LT)-HSCs compared to other progenitor populations. While m6A and mRNA levels were negatively correlated across hematopoietic progenitor populations, LT-HSCs displayed positive correlations between m6A and mRNA levels. It has been reported that Igf2bp1/2/3 stabilize m6A-tagged mRNAs (Huang et al., 2018). As Igf2bp2 was highly expressed in LT-HSCs, Zhang’s group examined the effects of Igf2bp2 deletion on HSCs. In Igf2bp2−/− mice, the frequency and numbers of LT-HSCs and short-term (ST)-HSCs were significantly decreased. The cell cycle of these Igf2bp2 knockout HSCs was shifted from G0/G1 quiescent status to cycling. The repopulation ability of Igf2bp2−/− bone marrow was significantly reduced over serial transplantation (Yin et al., 2022). The Zhang group also showed that Igf2bp2 directly binds to its targets, Bmi1, Cdk13, and cbfb in m6A sites. Although loss of Igf2bp2 did not affect any transcripts of Bmi1, an essential polycomb repressive complex (PRC) one protein gene for HSC self-renewal, it substantially impaired mRNA stability in HSCs, resulting in reduced Bmi1 expression, which then derepressed mitochondrial genes and induced loss of HSCs. Thus, Igf2bp2 seems to play a critical role in stabilizing HSC self-renewal genes.
The role of Igf2bp2 in aging of HSCs has also been reported (Suo et al., 2022). Suo et al. found that expression of Lin28b, Hmga2, and Igf2bp2 in HSCs (CD150+CD34−LSK cells) was decreased with age, while the expression of Hmga2 and Igf2bp2 was maintained in the multi-potent progenitor (MPP) population with age. HSCs from Igf2bp2−/− young mice had significantly reduced repopulation ability after transplantation, whereas HSCs from Igf2bp2−/− old mice displayed the same repopulation ability as WT HSCs. Igf2bp2−/− young HSCs showed lower mitochondrial respiration compared to Igf2bp2−/− old HSCs. Suo et al. also demonstrated that Igf2bp2 deletion significantly reduced the age-related HSC expansion generally seen in WT aged mice (Yamamoto et al., 2018). These data indicate that Igf2bp2 stabilizes HSC self-renewal genes in young mice. One caveat is that this report by Suo et al. used 3-6-month-old mice and defined them as “young mice,” though 6-month-old mice may be considered middle aged. Since the expression level of Lin28b in adult mice is significantly less than it is in fetal liver HSPCs by 3 months, it is not clear if Igf2bp2 is really downstream of Lin28b in adult mice.
While Lin28b overexpression in adult HSCs induces fetal-type hematopoiesis in the recipient mice, Hmga2 overexpression in adult HSCs does not (Copley et al., 2013). These data raise the question of what other Lin28b co-factors are involved in inducing fetal-like hematopoiesis. Iwama’s group found that the polycomb protein gene Exh2 represses fetal gene expression in adult bone marrow HSPCs (Oshima et al., 2016). They transplanted E14.5 fetal liver cells from CreERT2 control, Cre-ERT2: Ezh2f/f, and Cre-ERT2: Ezh2f/f: Tet2f/f embryos into lethally irradiated congenic mice and deleted the Ezh2 gene by tamoxifen injection 1 month after transplantation. Ezh2 deletion upregulated Lin28b, Hgma2, and Igf2bp3 in LSK HSPC populations in the recipient mice. Ezh2 deletion also induced more fetal-type hematopoiesis compared to control WT donor cells, including more B-1a, marginal zone B, and TCRgd T cell production in the adult recipient mice. Iwama’s group also conducted an analysis of H3K27me3 levels in WT fetal liver and bone marrow LSK cells and found that fetal HSPC genes are direct targets of Ezh2 in the adult bone marrow, with Ezh2 silencing these genes in the adult bone marrow via H3K27me3 modification (Oshima et al., 2016).
Rowe’s group found that Cbx2, a component of PRC1 and a target of Let-7, is also highly expressed in E14.5 fetal liver HSCs and common myeloid progenitors (Wang et al., 2022). Cbx2 deletion results in diminishment of ST-HSC and MPP2 populations in the fetal liver and smaller numbers of B cells in the neonatal spleen. Conversely, activation of Cbx2 in adult HSPCs induces erythroid skewing in granulocyte-erythroid-macrophage-monocyte (GEMM) colony-forming cells, similar to the phenotype of fetal GEMM colonies. These results indicate that Cbx2 plays a role in the differentiation process of fetal HSPCs, possibly under the regulation of Lin28b (Figure 2).
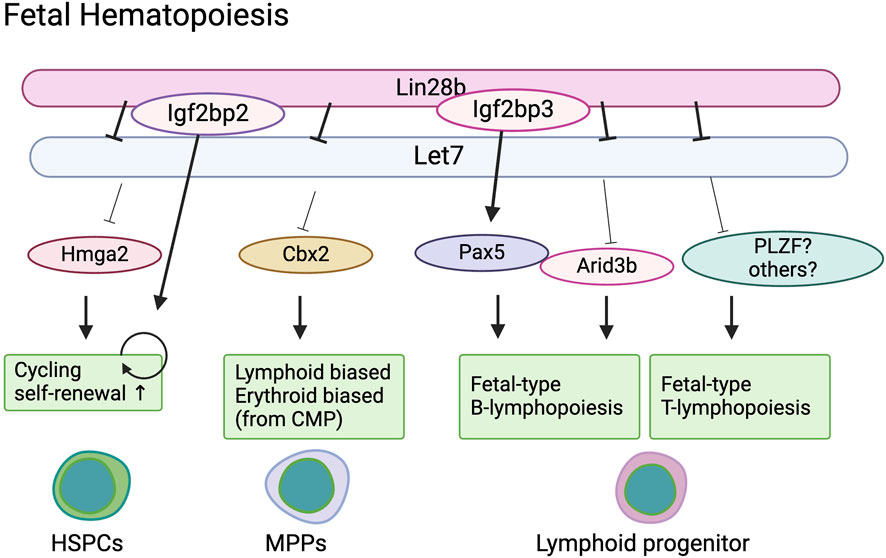
Figure 2. Lin28b regulate fetal hematopoiesis in a stage specific manner.
As mentioned above, Lin28b expression in adult HSPCs induces fetal-type hematopoiesis upon transplantation. Lin28b+ HSCs efficiently repopulate peritoneal B-1a cells and marginal zone B cells, TCRγδ T cells, and iNKT cells, which is similar to the engraftment phenotype seen in yolk sac transplantation or “transient HSC” transplantation (Yoshimoto et al., 2011; Yuan et al., 2012; Beaudin et al., 2016). Although it would be of interest to increase Lin28b expression to enhance HSC self-renewal for clinical applications, it is crucial to first obtain a complete understanding of how Lin28b expression in adult HSCs alters hematopoiesis. Below, we outline some of the key studies that describe how Lin28b changes the output of immune cells generated from adult HSCs.
The developmental switch of B cells has been studied for decades (Hardy and Hayakawa, 1991). It was originally described within the context of a layered immune system in which distinct types of stem cells produced different types of lymphoid cells with age (Herzenberg, 1989). Recent data have validated the layered immune system and shown that in the fetal period, rather than different types of stem cells, different types of progenitors exist. Among B cell subsets, B-1 cells and a portion of marginal zone (MZ) B cells are exclusively derived from fetal progenitors (Montecino-Rodriguez et al., 2006; Yoshimoto et al., 2011; Kobayashi et al., 2014; Ghosn et al., 2019). B-1 cells are innate-like B cells that secrete natural IgM antibodies without T cell help and are considered to have the ability to self-renew, maintaining themselves for life without being replenished by bone marrow progenitors. B-1 cells and MZ B cells originate from the yolk sac and embryo as early as E8.25 (Yoshimoto et al., 2011).
When adult HSPCs that are retrovirally overexpressed with Lin28b are transplanted into lethally irradiated mice, the Lin28b-expressing HSPCs predominantly repopulate B-1a cells and MZ B cells, similar to fetal liver HSPCs (Yuan et al., 2012; Wang et al., 2019). These effects are accompanied by Let-7 downregulation. Adult HSCs show high levels of Let-7 and no expression of Lin28b. Based on these studies, Zou et al. overexpressed Let-7 in B progenitors in the fetal liver and converted their fate into adult-type B-2 cells (Zhou et al., 2015). They also overexpressed Lin28b in B progenitors in the adult bone marrow and switched their fate into fetal-type B-1a cells. Furthermore, they identified Arid3a as a target of Let-7 that regulates fetal B cell development. Arid3a is part of the AT-rich interaction domain (ARID) super family of DNA-binding proteins and is known to enhance Ig heavy chain expression (Schmidt et al., 2009). Arid3a deletion results in embryonic lethality with hematopoietic defects at E12.5 (Webb et al., 2011). Zhou et al. found that the 3′ untranslated region (UTR) of the Arid3a mRNA contains several Let-7 target sites and that Arid3a knockdown reprograms fetal liver B progenitors to differentiate into adult type B-2 cells. Thus, Lin28b regulates B cell fate via the let-7-Arid3a axis in early lymphoid/B progenitor stages.
Lin28b also plays a role in positive selection of B cells (Vanhee et al., 2019). Vanhee et al. found a correlation between Lin28b expression levels and CD5 expression in B-1 cells. By using doxycycline inducible Lin28b transgenic mice (iLin28b), they found that inducing Lin28 in adult B cell progenitors can generate fully functional B-1 cells via positive selection.
Interestingly, Lin28b has also been shown to regulate B cell fate in a Let-7 independent manner. Wang et al. identified a possible Lin28b co-factor in the context of fetal B cell development (Wang et al., 2019). They found that Igf2bp3, another RNA binding protein, is highly expressed in fetal liver HSCs, common lymphoid progenitors (CLPs), and pro-B cells, but not in their adult counterparts. Gene expression profiling has identified Pax5 as a common target of Lin28 and Igf2bp3. Pax5 mRNA harbors a prominent binding site shared by LIN28 and IGF2BP3 in its 3′UTR near the stop codon and is not predicted to be a Let-7 target. Wang et al. demonstrated that co-binding of Lin28b and IGF2BP3 to Pax5 3′UTR is necessary to fully increase Pax5 expression (Wang et al., 2019). Pax5 is highly expressed in fetal CLPs compared to adult BM CLPs and has previously been shown to be differentially required for B cell development in the fetal liver and adult bone marrow. Pax5 deletion results in the absence of B lymphopoiesis in the fetal liver, whereas B progenitors (but not mature B cells) are present in the adult bone marrow (Nutt et al., 1997). Therefore, while Pax5 is indispensable for fetal B cell lineage commitment, in adult bone marrow, Pax5 is dispensable for B cell lineage commitment but essential for B cell maturation. These results support the recent notion that fetal B progenitors originate from different progenitors than adult HSCs (Ghosn et al., 2016; Kobayashi et al., 2023). Taken together, Pax5 is indispensable for fetal-derived B cell development under the control of Lin28b and Igf2bp3 (Figure 2).
One caveat of these studies is whether Lin28b overexpression in adult HSC really change the fate of their differentiation at HSC level. The report by Wang et al. showed that Lin28 and Igf2bp3 cooperatively stabilize their shared target (Pax5) in a Let-7 independent manner and indue fetal-type B cells, whereas Lin28b expand HSCs via Let-7 downregulation. Therefore, it is plausible that Lin28b overexpression induce fetal B-lymphopoiesis in CLP and may not change the differentiation characteristics of HSC itself. For example, if Lin28b is transiently expressed only in LT-HSCs, the progenies may not show fetal-type hematopoietic phenotype. Since many of these studies are conducted using HSPCs, not purified LT-HSCs, further investigation would be required to understand how Lin28b works in each early HSPC population such as LT-HSCs, MPPs, CLP, and pro-B cells.
Lin28b can also be used to shape the production and function of T cells. Similar to the ontogeny of B cells, Lin28b enables adult progenitors to give rise to certain subsets of T cells that are typically only made by fetal and neonatal progenitors. For example, γδT cells are primarily made in the fetal thymus prior to birth. However, Lin28b can reprogram human cord blood CD34+ HSPCs to make fetal gamma delta T cells (Tieppo et al., 2020). Similar studies have been performed in mice. However, overexpression of Lin28b in adult HSPCs leads to increased numbers of some but not all subsets of murine γδT cells (Yuan et al., 2012). Specifically, adult bone marrow HSPCs transduced with Lin28b give rise to the perinatally-derived Vγ1+Vδ6.3+ T cells but not the fetal-derived Vγ5 dendritic epidermal T cells (DETC) or Vγ6+ IL-17 producing γδT cells (Heilig and Tonegawa, 1986; Itohara et al., 1990; Haas et al., 2012; Yuan et al., 2012). Although it is not known why Lin28b only permits the production of certain subsets of gamma delta T cells, it is possible that the earliest waves of fetal gamma delta T cells require not only fetal programming by Lin28b, but also a fetal thymic environment (Havran and Allison, 1990).
Another subset of T cells that is primarily made in early life is the invariant natural killer T (iNKT) cells, which are an innate subset of T cells that recognize lipids. Previous work has shown that iNKT cells are more abundantly made by fetal and neonatal progenitors in the murine thymus (Lee et al., 2010; Watarai et al., 2010). However, overexpression of Lin28b in adult HSPCs facilitates the production of larger numbers of iNKT cells, presumably by enhancing the expression of a key transcription factor, PLZF (promyelocytic leukemia zinc finger), which is required for their development (Yuan et al., 2012; Pobezinsky et al., 2015). Notably, PLZF is also required for the production of Vγ1+Vδ6.3+ T cells in early life but is targeted by Let-7 during later stages of development (Kreslavsky et al., 2009; Alonzo et al., 2010; Lu et al., 2015). Thus, overexpression of Lin28b may block the repression of PLZF by Let-7, allowing for the outgrowth of both Vγ1+Vδ6.3+ T cells and iNKT cells by adult progenitors (Figure 2).
Alternatively, Lin28b may bias T cell output towards the more innate-like lineages (e.g., Vγ1+Vδ6.3+ T cells and iNKT cells) by altering the T cell receptor (TCR) repertoire. In early life, T cells are made in the absence of terminal deoxynucleotidyl transferase (TdT), which is responsible for the insertion of random nucleotides in the junctional regions of the TCRs (Gilfillan et al., 1993; Komori et al., 1993; Gavin and Bevan, 1995; Cabaniols et al., 2001). As a result, the T cells produced during fetal and neonatal stages of development are less diverse and comprised of shorter and more germline-encoded TCRs. Importantly, recent studies have found that overexpression of Lin28 b in adult progenitors leads to a decrease in TdT expression (Wang et al., 2019; Tieppo et al., 2020). It is possible that innate lymphocytes with germline-configured TCRs are more efficiently made in Lin28b-expressing progenitors because the addition of random nucleotides is suppressed (Aono et al., 2000).
In addition to altering the types of T cells made, Lin28b also changes the functions of conventional CD8+ and CD4+ T cells. Interestingly, inducing Lin28b in adult progenitors allows them to give rise to CD8+ T cells that are phenotypically and functionally analogous to neonatal CD8+ T cells (Wang et al., 2016; Wells et al., 2017; Smith et al., 2018; Tabilas et al., 2019). For example, prior to infection, both neonatal and adult Lin28b transgenic mice contain a large proportion of CD8+ T cells that exhibit a virtual memory (VM) phenotype (CD44+CD122+), whereas only a small fraction of adult CD8+ T cells show this phenotype (Wang et al., 2016). Importantly, intrathymic injections and mixed bone chimeras indicate that the accumulation of VM cells in neonatal mice and Lin28b transgenic mice does not come as a result of changes in the thymic environment, but instead is driven by cell-autonomous changes in the progenitors (Wang et al., 2016). Also, neonatal and Lin28b transgenic CD8+ T cells preferentially give rise to short-lived effectors after infection, while adult CD8+ T cells are more efficient at generating memory precursor cells (Wang et al., 2016; Tabilas et al., 2019). More recently, neonatal and Lin28b Tg CD8+ T cells were found to have an enhanced ability to respond to inflammation and protect the host against unrelated infections in a TCR-independent manner, which corresponded to their unique ability to rapidly undergo chromatin remodeling (Watson et al., 2024). Thus, Lin28b can be employed to generate CD8+ T cells with more fast-acting innate-like properties.
In the case of CD4+ T cells, Lin28b may bias CD4+ T cells towards becoming Tregs in early life (Mold et al., 2010; Wang et al., 2010; Jain, 2020). Notably, inhibition of Lin28b in human fetal CD4+ T cells impairs their ability to differentiate into regulatory T cells (Bronevetsky et al., 2016). Although the underlying mechanisms are poorly understood, some evidence indicates that Lin28b promotes a fetal Treg bias by enhancing TGF-β signaling, which is repressed by let-7 during later stages of development (Bronevetsky et al., 2016). Alternatively, it is possible that Lin28b enhances the propensity for naïve T cells to become Tregs by altering thymic selection. Earlier studies have shown that fetal and neonatal T cells in both mice and humans exhibit higher self-reactivity than their adult counterparts, which is a key feature of regulatory T cells (Dong et al., 2017). It will be important to determine whether Lin28b alters the threshold of self-reactivity in T cells, similar to how Lin28b enhances the selection of more self-reactive B cells (Vanhee et al., 2019; Xu et al., 2020).
Lin28b also regulates erythro-myeloid differentiation (Rowe et al., 2016). Lin28b is highly expressed in common myeloid progenitors (CMPs) in the fetal liver. Hematopoietic colonies produced by fetal CMPs are biased towards erythroid outputs, whereas colonies produced by adult BM CMPs are biased towards myeloid outputs with much higher expression of Let-7. Hmg2a induction in adult CMPs recapitulated erythroid output, suggesting that the Lin28b-let7-Hmg2a axis regulates CMP differentiation in the fetal liver.
Lin28b and Hmg2a are also expressed in megakaryocyte progenitors in the fetus. Fetal-activated platelets express low levels of P-selectin, which is phenocopied when Lin28b is overexpressed in adult HSPCs, indicating that the Lin28b-let7-Hmg2a axis also controls platelet maturation from fetal megakaryocytes (Stolla et al., 2019).
There is also data to suggest that Lin28b induces fetal-type erythropoiesis in humans. Similar to what has been observed in mice, Let-7 has been associated with a fetal-to-adult switch of globin chains in the human embryo. Specifically, erythrocytes produce a fetal γ-globin-based HbF hemoglobin and transition to an adult β-globin-based HbA hemoglobin prior to birth, and this phenotype is maintained into adulthood (Noh et al., 2009). During the transition, Lin28b is highly expressed in erythrocytes through fetal maturity and exerts the same downregulatory effect on Let-7, resulting in increased expression of HbF (Yuan et al., 2012; Lee et al., 2013). In contrast, Lin28b knockdown in cord blood CD34+ cells has been shown to decrease HbF and increase HbA. Retroviral overexpression of LIN28b in human adult erythroblasts forces increased production of fetal γ-globin mRNA, with a corresponding decrease in adult β-globin (Lee et al., 2013). These changes seem to be controlled by the inhibition of Let-7 and its effect on BCL11A, a regulator of fetal HbF and adult HbA production (Sankaran et al., 2008; Lee et al., 2013).
Another way in which Lin28b exerts its effects on fetal hemoglobin production is through the previously discussed IGF2 family. When overexpressed via retroviral transduction into human CD34+ HSPCs derived from adult bone marrow, IGF2BP1 triggered a robust switch in production of β-globin mRNA to γ-globin mRNA, which coincided with posttranscriptional repression of BCL11A (de Vasconcellos et al., 2017). This effect was independent of any increase in Lin28b or inhibition of Let-7. The post-transcriptional regulation of BCL11A demonstrates a different mechanism of regulation than the transcriptional regulation of Lin28b-IGF2BP3 binding of the Pax5 3′ UTR (Wang et al., 2019), further demonstrating the diverse ways that Lin28b fine-tunes gene expression.
Lin28b also regulate mast cell expansion. Using doxycycline-inducible Lin28b transgenic mice, Wang et al. induced ectopic Lin28b expression and found increased numbers of mast cells and their progenitors (Wang et al., 2015). They also found that Lin28b expression was upregulated in the pathogenic mast cells from patients with systemic mastocytosis. These results are supported by the recent papers showing that mast cells are fetal-derived (Gentek et al., 2018; Li et al., 2018; Yoshimoto et al., 2022).
We introduced the roles of Lin28b and their downstream factors in HSC self-renewal and various lineage specifications. Especially, the Lin28-let7-Hmga2-Igf2bp2 axis is important for enhancing the ability of HSCs to self-renew. These genes push HSCs to enter cell cycle and proliferate, while still maintaining both stem cell and differentiation abilities via epigenetic modification or m6A modifications. However, to utilize the physiological expansion ability of HSCs for clinical applications, thorough consideration is required. For example, all these studies testing the role of Lin28b and downstream factors in HSCs use adult bone marrow HSPCs in inducible or knockout mouse models or overexpression of genes by virus transduction. Therefore, it is not clear that Lin28-let7-Hmga2-Igf2bp2 axis induce HSC expansion in the fetal liver in the physiological setting by the mechanisms described in those reports.
A promising consideration when inducing Lin28b expression in adult HSCs is that ectopic transient expression of Lin28b will lead to fetal-like hematopoiesis. Since adult HSCs lack the differentiation capability of innate immune cells that are primarily made in early life, induction of Lin28b may help to restore or elevate the presence of these immune cells in patients undergoing conventional blood stem cell transplantation therapy or in older people, later in life, who have lost such cells. For example, fetal-derived B-1 cells are known to produce anti-PC antibodies that can prevent pneumococcal invasion; streptococcus pneumoniae are among the pathogens that can cause severe infection in older people. Thus, expanding stem cell numbers and regulating immune cell production via Lin28 could improve the outcomes of both older patients and those receiving transplantation therapy.
A possible negative outcome of Lin28b induction into adult HSCs is the potential for developing leukemia, as Lin28 is highly expressed in some pediatric leukemias (Helsmoortel et al., 2016a; Helsmoortel et al., 2016b). It has also been reported that ectopic Hmga2 expression in adult HSPCs led to the development of acute myeloid leukemias (AML) in a Tet2-deficient context (Bai et al., 2021). Therefore, if one overexpresses Lin28b in HSCs, transient expression would be desired. On the other hand, Lin28 expression has been reported to promote differentiation of MLL-fusion-driven AML (Eldeeb et al., 2023; Li et al., 2024). Silencing Hmga2 has also been proposed as a potential therapeutic option for AML (Tan et al., 2018; Yang et al., 2019). As such, the role of Lin28b is context dependent, therefore, a better understanding of the mechanisms and roles of the Lin28b/Hmga2-Igf2bp2/3 axis in different HSPC population at different ages will open the door to more therapeutic opportunities.
In addition, although transplantable HSC numbers are considered to increase during development, a recent report has suggested that HSCs do not robustly expand, as previously thought (Ganuza et al., 2022). It has been postulated that pre-HSCs mature into transplantable HSCs between E11 and E12 (and possibly later) in the fetal liver. However, the mechanisms and processes behind maturation of pre-HSCs into transplantable HSCs are still largely unknown. Therefore, more precise elucidation of HSC numbers and behaviors during fetal life will be required to understand the biological mechanisms of HSC expansion in a physiological setting.
Lastly, there are limitations to studying human fetal HSCs due to different legal allowances for the use of human fetal tissues in each country. Because of this, it may be challenging to understand the mechanisms that support HSC expansion in the human fetal liver or possibly other fetal tissues.
All of these studies are expected to contribute to our basic understanding of hematopoiesis and immune development at different stages of life.
GC: Writing–original draft, Writing–review and editing. MK: Writing–review and editing. BR: Writing–original draft, Writing–review and editing. MY: Conceptualization, Writing–original draft, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work is supported by National Institute of Health awards R01AI165798 (to MY), R01AG084752 (to MY), R21AI178153 (to MY), R01AI110613 (to B.D.R), R01HD107798 (to B.D.R), and R01AI105265 (to B.D.R).
Figures were created using BioRender.com.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Agrawal, H., Mehatre, S. H., and Khurana, S. (2024). The hematopoietic stem cell expansion niche in fetal liver: current state of the art and the way forward. Exp. Hematol. 136, 104585. doi:10.1016/j.exphem.2024.104585
Alonzo, E. S., Gottschalk, R. A., Das, J., Egawa, T., Hobbs, R. M., Pandolfi, P. P., et al. (2010). Development of promyelocytic zinc finger and ThPOK-expressing innate gamma delta T cells is controlled by strength of TCR signaling and Id3. J. Immunol. 184 (3), 1268–1279. doi:10.4049/jimmunol.0903218
Ambros, V., and Horvitz, H. R. (1984). Heterochronic mutants of the nematode Caenorhabditis elegans. Science 226 (4673), 409–416. doi:10.1126/science.6494891
Aono, A., Enomoto, H., Yoshida, N., Yoshizaki, K., Kishimoto, T., and Komori, T. (2000). Forced expression of terminal deoxynucleotidyl transferase in fetal thymus resulted in a decrease in gammadelta T cells and random dissemination of Vgamma3Vdelta1 T cells in skin of newborn but not adult mice. Immunology 99 (4), 489–497. doi:10.1046/j.1365-2567.2000.00987.x
Ara, T., Tokoyoda, K., Sugiyama, T., Egawa, T., Kawabata, K., and Nagasawa, T. (2003). Long-term hematopoietic stem cells require stromal cell-derived factor-1 for colonizing bone marrow during ontogeny. Immunity 19 (2), 257–267. doi:10.1016/s1074-7613(03)00201-2
Arora, N., Wenzel, P. L., McKinney-Freeman, S. L., Ross, S. J., Kim, P. G., Chou, S. S., et al. (2014). Effect of developmental stage of HSC and recipient on transplant outcomes. Dev. Cell 29 (5), 621–628. doi:10.1016/j.devcel.2014.04.013
Bai, J., Yokomizo-Nakano, T., Kubota, S., Sun, Y., Kanai, A., Iimori, M., et al. (2021). Overexpression of Hmga2 activates Igf2bp2 and remodels transcriptional program of Tet2-deficient stem cells in myeloid transformation. Oncogene 40 (8), 1531–1541. doi:10.1038/s41388-020-01629-w
Bastani, S., Staal, F. J. T., and Cante-Barrett, K. (2023). The quest for the holy grail: overcoming challenges in expanding human hematopoietic stem cells for clinical use. Stem Cell Investig. 10, 15. doi:10.21037/sci-2023-016
Batsivari, A., Rybtsov, S., Souilhol, C., Binagui-Casas, A., Hills, D., Zhao, S., et al. (2017). Understanding hematopoietic stem cell development through functional correlation of their proliferative status with the intra-aortic cluster architecture. Stem Cell Rep. 8 (6), 1549–1562. doi:10.1016/j.stemcr.2017.04.003
Beaudin, A. E., Boyer, S. W., Perez-Cunningham, J., Hernandez, G. E., Derderian, S. C., Jujjavarapu, C., et al. (2016). A transient developmental hematopoietic stem cell gives rise to innate-like B and T cells. Cell Stem Cell 19 (6), 768–783. doi:10.1016/j.stem.2016.08.013
Bowie, M. B., McKnight, K. D., Kent, D. G., McCaffrey, L., Hoodless, P. A., and Eaves, C. J. (2006). Hematopoietic stem cells proliferate until after birth and show a reversible phase-specific engraftment defect. J. Clin. Invest. 116 (10), 2808–2816. doi:10.1172/JCI28310
Bronevetsky, Y., Burt, T. D., and McCune, J. M. (2016). Lin28b regulates fetal regulatory T cell differentiation through modulation of TGF-β signaling. J. Immunol. 197 (11), 4344–4350. doi:10.4049/jimmunol.1601070
Broxmeyer, H. E., Orschell, C. M., Clapp, D. W., Hangoc, G., Cooper, S., Plett, P. A., et al. (2005). Rapid mobilization of murine and human hematopoietic stem and progenitor cells with AMD3100, a CXCR4 antagonist. J. Exp. Med. 201 (8), 1307–1318. doi:10.1084/jem.20041385
Cabaniols, J. P., Fazilleau, N., Casrouge, A., Kourilsky, P., and Kanellopoulos, J. M. (2001). Most alpha/beta T cell receptor diversity is due to terminal deoxynucleotidyl transferase. J. Exp. Med. 194 (9), 1385–1390. doi:10.1084/jem.194.9.1385
Chen, M. J., Yokomizo, T., Zeigler, B. M., Dzierzak, E., and Speck, N. A. (2009). Runx1 is required for the endothelial to haematopoietic cell transition but not thereafter. Nature 457 (7231), 887–891. doi:10.1038/nature07619
Cheshier, S. H., Morrison, S. J., Liao, X., and Weissman, I. L. (1999). In vivo proliferation and cell cycle kinetics of long-term self-renewing hematopoietic stem cells. Proc. Natl. Acad. Sci. U. S. A. 96 (6), 3120–3125. doi:10.1073/pnas.96.6.3120
Copley, M. R., Babovic, S., Benz, C., Knapp, D. J., Beer, P. A., Kent, D. G., et al. (2013). The Lin28b-let-7-Hmga2 axis determines the higher self-renewal potential of fetal haematopoietic stem cells. Nat. Cell Biol. 15 (8), 916–925. doi:10.1038/ncb2783
Cumano, A., Dieterlen-Lievre, F., and Godin, I. (1996). Lymphoid potential, probed before circulation in mouse, is restricted to caudal intraembryonic splanchnopleura. Cell 86 (6), 907–916. doi:10.1016/s0092-8674(00)80166-x
de Vasconcellos, J. F., Tumburu, L., Byrnes, C., Lee, Y. T., Xu, P. C., Li, M., et al. (2017). IGF2BP1 overexpression causes fetal-like hemoglobin expression patterns in cultured human adult erythroblasts. Proc. Natl. Acad. Sci. U. S. A. 114 (28), E5664-E5672–E5672. doi:10.1073/pnas.1609552114
Dignum, T., Varnum-Finney, B., Srivatsan, S. R., Dozono, S., Waltner, O., Heck, A. M., et al. (2021). Multipotent progenitors and hematopoietic stem cells arise independently from hemogenic endothelium in the mouse embryo. Cell Rep. 36 (11), 109675. doi:10.1016/j.celrep.2021.109675
Dong, M., Artusa, P., Kelly, S. A., Fournier, M., Baldwin, T. A., Mandl, J. N., et al. (2017). Alterations in the thymic selection threshold skew the self-reactivity of the TCR repertoire in neonates. J. Immunol. 199 (3), 965–973. doi:10.4049/jimmunol.1602137
Eldeeb, M., Yuan, O., Guzzi, N., Thi Ngoc, P. C., Konturek-Ciesla, A., Kristiansen, T. A., et al. (2023). A fetal tumor suppressor axis abrogates MLL-fusion-driven acute myeloid leukemia. Cell Rep. 42 (2), 112099. doi:10.1016/j.celrep.2023.112099
Ema, H., and Nakauchi, H. (2000). Expansion of hematopoietic stem cells in the developing liver of a mouse embryo. Blood 95 (7), 2284–2288. doi:10.1182/blood.v95.7.2284
Fares, I., Chagraoui, J., Gareau, Y., Gingras, S., Ruel, R., Mayotte, N., et al. (2014). Cord blood expansion. Pyrimidoindole derivatives are agonists of human hematopoietic stem cell self-renewal. Science 345 (6203), 1509–1512. doi:10.1126/science.1256337
Ganuza, M., Hall, T., Myers, J., Nevitt, C., Sanchez-Lanzas, R., Chabot, A., et al. (2022). Murine foetal liver supports limited detectable expansion of life-long haematopoietic progenitors. Nat. Cell Biol. 24 (10), 1475–1486. doi:10.1038/s41556-022-00999-5
Gavin, M. A., and Bevan, M. J. (1995). Increased peptide promiscuity provides a rationale for the lack of N regions in the neonatal T cell repertoire. Immunity 3 (6), 793–800. doi:10.1016/1074-7613(95)90068-3
Gekas, C., Dieterlen-Lievre, F., Orkin, S. H., and Mikkola, H. K. (2005). The placenta is a niche for hematopoietic stem cells. Dev. Cell 8 (3), 365–375. doi:10.1016/j.devcel.2004.12.016
Gentek, R., Ghigo, C., Hoeffel, G., Bulle, M. J., Msallam, R., Gautier, G., et al. (2018). Hemogenic endothelial fate mapping reveals dual developmental origin of mast cells. Immunity 48 (6), 1160–1171. doi:10.1016/j.immuni.2018.04.025
Ghosn, E., Yoshimoto, M., Nakauchi, H., Weissman, I. L., and Herzenberg, L. A. (2019). Hematopoietic stem cell-independent hematopoiesis and the origins of innate-like B lymphocytes. Development 146 (15), dev170571. doi:10.1242/dev.170571
Ghosn, E. E., Waters, J., Phillips, M., Yamamoto, R., Long, B. R., Yang, Y., et al. (2016). Fetal hematopoietic stem cell transplantation fails to fully regenerate the B-lymphocyte compartment. Stem Cell Rep. 6 (1), 137–149. doi:10.1016/j.stemcr.2015.11.011
Gilfillan, S., Dierich, A., Lemeur, M., Benoist, C., and Mathis, D. (1993). Mice lacking TdT: mature animals with an immature lymphocyte repertoire. Science 261 (5125), 1175–1178. doi:10.1126/science.8356452
Godin, I., Dieterlen-Lievre, F., and Cumano, A. (1995). Emergence of multipotent hemopoietic cells in the yolk sac and paraaortic splanchnopleura in mouse embryos, beginning at 8.5 days postcoitus. Proc. Natl. Acad. Sci. U. S. A. 92 (3), 773–777. doi:10.1073/pnas.92.3.773
Haas, J. D., Ravens, S., Duber, S., Sandrock, I., Oberdorfer, L., Kashani, E., et al. (2012). Development of interleukin-17-producing γδ T cells is restricted to a functional embryonic wave. Immunity 37 (1), 48–59. doi:10.1016/j.immuni.2012.06.003
Hardy, R. R., and Hayakawa, K. (1991). A developmental switch in B lymphopoiesis. Proc. Natl. Acad. Sci. U. S. A. 88 (24), 11550–11554. doi:10.1073/pnas.88.24.11550
Havran, W. L., and Allison, J. P. (1990). Origin of Thy-1+ dendritic epidermal cells of adult mice from fetal thymic precursors. Nature 344 (6261), 68–70. doi:10.1038/344068a0
Heilig, J. S., and Tonegawa, S. (1986). Diversity of murine gamma genes and expression in fetal and adult T lymphocytes. Nature 322 (6082), 836–840. doi:10.1038/322836a0
Helsmoortel, H. H., Bresolin, S., Lammens, T., Cave, H., Noellke, P., Caye, A., et al. (2016a). LIN28B overexpression defines a novel fetal-like subgroup of juvenile myelomonocytic leukemia. Blood 127 (9), 1163–1172. doi:10.1182/blood-2015-09-667808
Helsmoortel, H. H., De Moerloose, B., Pieters, T., Ghazavi, F., Bresolin, S., Cave, H., et al. (2016b). LIN28B is over-expressed in specific subtypes of pediatric leukemia and regulates lncRNA H19. Haematologica 101 (6), e240–e244. doi:10.3324/haematol.2016.143818
Herzenberg, L. A. (1989). Toward a layered immune system. Cell 59 (6), 953–954. doi:10.1016/0092-8674(89)90748-4
Huang, H., Weng, H., Sun, W., Qin, X., Shi, H., Wu, H., et al. (2018). Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 20 (3), 285–295. doi:10.1038/s41556-018-0045-z
Ikeda, K., Mason, P. J., and Bessler, M. (2011). 3'UTR-truncated Hmga2 cDNA causes MPN-like hematopoiesis by conferring a clonal growth advantage at the level of HSC in mice. Blood 117 (22), 5860–5869. doi:10.1182/blood-2011-02-334425
Itohara, S., Farr, A. G., Lafaille, J. J., Bonneville, M., Takagaki, Y., Haas, W., et al. (1990). Homing of a gamma delta thymocyte subset with homogeneous T-cell receptors to mucosal epithelia. Nature 343 (6260), 754–757. doi:10.1038/343754a0
Jain, N. (2020). The early life education of the immune system: moms, microbes and (missed) opportunities. Gut Microbes 12 (1), e1824564–1-20. doi:10.1080/19490976.2020.1824564
Kim, I., Saunders, T. L., and Morrison, S. J. (2007). Sox17 dependence distinguishes the transcriptional regulation of fetal from adult hematopoietic stem cells. Cell 130(3), 470–483. doi:10.1016/j.cell.2007.06.011
Kobayashi, M., Shelley, W. C., Seo, W., Vemula, S., Lin, Y., Liu, Y., et al. (2014). Functional B-1 progenitor cells are present in the hematopoietic stem cell-deficient embryo and depend on Cbfβ for their development. Proc. Natl. Acad. Sci. U. S. A. 111 (33), 12151–12156. doi:10.1073/pnas.1407370111
Kobayashi, M., Tarnawsky, S. P., Wei, H., Mishra, A., Azevedo Portilho, N., Wenzel, P., et al. (2019). Hemogenic endothelial cells can transition to hematopoietic stem cells through a B-1 lymphocyte-biased state during maturation in the mouse embryo. Stem Cell Rep. 13 (1), 21–30. doi:10.1016/j.stemcr.2019.05.025
Kobayashi, M., Wei, H., Yamanashi, T., Azevedo Portilho, N., Cornelius, S., Valiente, N., et al. (2023). HSC-independent definitive hematopoiesis persists into adult life. Cell Rep. 42 (3), 112239. doi:10.1016/j.celrep.2023.112239
Kobayashi, M., and Yoshimoto, M. (2023). Multiple waves of fetal-derived immune cells constitute adult immune system. Immunol. Rev. 315 (1), 11–30. doi:10.1111/imr.13192
Komori, T., Okada, A., Stewart, V., and Alt, F. W. (1993). Lack of N regions in antigen receptor variable region genes of TdT-deficient lymphocytes. Science 261 (5125), 1171–1175. doi:10.1126/science.8356451
Kreslavsky, T., Savage, A. K., Hobbs, R., Gounari, F., Bronson, R., Pereira, P., et al. (2009). TCR-inducible PLZF transcription factor required for innate phenotype of a subset of gammadelta T cells with restricted TCR diversity. Proc. Natl. Acad. Sci. U. S. A. 106 (30), 12453–12458. doi:10.1073/pnas.0903895106
Kubota, S., Sun, Y., Morii, M., Bai, J., Ideue, T., Hirayama, M., et al. (2024). Chromatin modifier Hmga2 promotes adult hematopoietic stem cell function and blood regeneration in stress conditions. EMBO J. 43 (13), 2661–2684. doi:10.1038/s44318-024-00122-4
Kumar, P., Beck, D., Galeev, R., Thoms, J. A. I., Talkhoncheh, M. S., de Jong, I., et al. (2019). HMGA2 promotes long-term engraftment and myeloerythroid differentiation of human hematopoietic stem and progenitor cells. Blood Adv. 3 (4), 681–691. doi:10.1182/bloodadvances.2018023986
Kumaravelu, P., Hook, L., Morrison, A. M., Ure, J., Zhao, S., Zuyev, S., et al. (2002). Quantitative developmental anatomy of definitive haematopoietic stem cells/long-term repopulating units (HSC/RUs): role of the aorta-gonad-mesonephros (AGM) region and the yolk sac in colonisation of the mouse embryonic liver. Development 129 (21), 4891–4899. doi:10.1242/dev.129.21.4891
Lee, Y. J., Jeon, Y. K., Kang, B. H., Chung, D. H., Park, C. G., Shin, H. Y., et al. (2010). Generation of PLZF+ CD4+ T cells via MHC class II-dependent thymocyte-thymocyte interaction is a physiological process in humans. J. Exp. Med. 207 (1), 237–246. doi:10.1084/jem.20091519
Lee, Y. T., de Vasconcellos, J. F., Yuan, J., Byrnes, C., Noh, S. J., Meier, E. R., et al. (2013). LIN28B-mediated expression of fetal hemoglobin and production of fetal-like erythrocytes from adult human erythroblasts ex vivo. Blood 122 (6), 1034–1041. doi:10.1182/blood-2012-12-472308
Li, Y., Mendoza-Castrejon, J., Patel, R. M., Casey, E. B., Denby, E., Bryder, D., et al. (2024). LIN28B promotes differentiation of fully transformed AML cells but is dispensable for fetal leukemia suppression. Leukemia 38 (3), 648–651. doi:10.1038/s41375-024-02167-0
Li, Z., Liu, S., Xu, J., Zhang, X., Han, D., Liu, J., et al. (2018). Adult connective tissue-resident mast cells originate from late erythro-myeloid progenitors. Immunity 49 (4), 640–653. doi:10.1016/j.immuni.2018.09.023
Lu, Y., Cao, X., Zhang, X., and Kovalovsky, D. (2015). PLZF controls the development of fetal-derived IL-17+vγ6+ γδ T cells. J. Immunol. 195 (9), 4273–4281. doi:10.4049/jimmunol.1500939
Lux, C. T., Yoshimoto, M., McGrath, K., Conway, S. J., Palis, J., and Yoder, M. C. (2008). All primitive and definitive hematopoietic progenitor cells emerging before E10 in the mouse embryo are products of the yolk sac. Blood 111 (7), 3435–3438. doi:10.1182/blood-2007-08-107086
Matsuoka, S., Ebihara, Y., Xu, M., Ishii, T., Sugiyama, D., Yoshino, H., et al. (2001). CD34 expression on long-term repopulating hematopoietic stem cells changes during developmental stages. Blood 97 (2), 419–425. doi:10.1182/blood.v97.2.419
Meaker, G. A., and Wilkinson, A. C. (2024). Ex vivo hematopoietic stem cell expansion technologies: recent progress, applications, and open questions. Exp. Hematol. 130, 104136. doi:10.1016/j.exphem.2023.12.001
Medvinsky, A., and Dzierzak, E. (1996). Definitive hematopoiesis is autonomously initiated by the AGM region. Cell 86 (6), 897–906. doi:10.1016/s0092-8674(00)80165-8
Mold, J. E., Venkatasubrahmanyam, S., Burt, T. D., Michaelsson, J., Rivera, J. M., Galkina, S. A., et al. (2010). Fetal and adult hematopoietic stem cells give rise to distinct T cell lineages in humans. Science 330 (6011), 1695–1699. doi:10.1126/science.1196509
Montecino-Rodriguez, E., Leathers, H., and Dorshkind, K. (2006). Identification of a B-1 B cell-specified progenitor. Nat. Immunol. 7 (3), 293–301. doi:10.1038/ni1301
Moss, E. G., Lee, R. C., and Ambros, V. (1997). The cold shock domain protein LIN-28 controls developmental timing in C. elegans and is regulated by the lin-4 RNA. Cell 88 (5), 637–646. doi:10.1016/s0092-8674(00)81906-6
Newman, M. A., Thomson, J. M., and Hammond, S. M. (2008). Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing. RNA 14 (8), 1539–1549. doi:10.1261/rna.1155108
Nishikawa, S. I., Nishikawa, S., Kawamoto, H., Yoshida, H., Kizumoto, M., Kataoka, H., et al. (1998). In vitro generation of lymphohematopoietic cells from endothelial cells purified from murine embryos. Immunity 8 (6), 761–769. doi:10.1016/s1074-7613(00)80581-6
Noh, S. J., Miller, S. H., Lee, Y. T., Goh, S. H., Marincola, F. M., Stroncek, D. F., et al. (2009). Let-7 microRNAs are developmentally regulated in circulating human erythroid cells. J. Transl. Med. 7, 98. doi:10.1186/1479-5876-7-98
Nutt, S. L., Urbanek, P., Rolink, A., and Busslinger, M. (1997). Essential functions of Pax5 (BSAP) in pro-B cell development: difference between fetal and adult B lymphopoiesis and reduced V-to-DJ recombination at the IgH locus. Genes Dev. 11 (4), 476–491. doi:10.1101/gad.11.4.476
Oshima, M., Hasegawa, N., Mochizuki-Kashio, M., Muto, T., Miyagi, S., Koide, S., et al. (2016). Ezh2 regulates the Lin28/let-7 pathway to restrict activation of fetal gene signature in adult hematopoietic stem cells. Exp. Hematol. 44 (4), 282–296. doi:10.1016/j.exphem.2015.12.009
Pobezinsky, L. A., Etzensperger, R., Jeurling, S., Alag, A., Kadakia, T., McCaughtry, T. M., et al. (2015). Let-7 microRNAs target the lineage-specific transcription factor PLZF to regulate terminal NKT cell differentiation and effector function. Nat. Immunol. 16 (5), 517–524. doi:10.1038/ni.3146
Polesskaya, A., Cuvellier, S., Naguibneva, I., Duquet, A., Moss, E. G., and Harel-Bellan, A. (2007). Lin-28 binds IGF-2 mRNA and participates in skeletal myogenesis by increasing translation efficiency. Genes Dev. 21 (9), 1125–1138. doi:10.1101/gad.415007
Rowe, R. G., Wang, L. D., Coma, S., Han, A., Mathieu, R., Pearson, D. S., et al. (2016). Developmental regulation of myeloerythroid progenitor function by the Lin28b-let-7-Hmga2 axis. J. Exp. Med. 213 (8), 1497–1512. doi:10.1084/jem.20151912
Rubio-Lara, J. A., Igarashi, K. J., Sood, S., Johansson, A., Sommerkamp, P., Yamashita, M., et al. (2023). Expanding hematopoietic stem cell ex vivo: recent advances and technical considerations. Exp. Hematol. 125-126, 6–15. doi:10.1016/j.exphem.2023.07.006
Rybtsov, S., Sobiesiak, M., Taoudi, S., Souilhol, C., Senserrich, J., Liakhovitskaia, A., et al. (2011). Hierarchical organization and early hematopoietic specification of the developing HSC lineage in the AGM region. J. Exp. Med. 208 (6), 1305–1315. doi:10.1084/jem.20102419
Sakurai, M., Ishitsuka, K., Ito, R., Wilkinson, A. C., Kimura, T., Mizutani, E., et al. (2023). Chemically defined cytokine-free expansion of human haematopoietic stem cells. Nature 615 (7950), 127–133. doi:10.1038/s41586-023-05739-9
Sankaran, V. G., Menne, T. F., Xu, J., Akie, T. E., Lettre, G., Van Handel, B., et al. (2008). Human fetal hemoglobin expression is regulated by the developmental stage-specific repressor BCL11A. Science 322 (5909), 1839–1842. doi:10.1126/science.1165409
Schmidt, C., Kim, D., Ippolito, G. C., Naqvi, H. R., Probst, L., Mathur, S., et al. (2009). Signalling of the BCR is regulated by a lipid rafts-localised transcription factor, Bright. EMBO J. 28 (6), 711–724. doi:10.1038/emboj.2009.20
Shyh-Chang, N., and Daley, G. Q. (2013). Lin28: primal regulator of growth and metabolism in stem cells. Cell Stem Cell 12 (4), 395–406. doi:10.1016/j.stem.2013.03.005
Sigurdsson, V., Takei, H., Soboleva, S., Radulovic, V., Galeev, R., Siva, K., et al. (2016). Bile acids protect expanding hematopoietic stem cells from unfolded protein stress in fetal liver. Cell Stem Cell 18 (4), 522–532. doi:10.1016/j.stem.2016.01.002
Smith, N. L., Patel, R. K., Reynaldi, A., Grenier, J. K., Wang, J., Watson, N. B., et al. (2018). Developmental origin governs CD8(+) T cell fate decisions during infection. Cell 174 (1), 117–130. doi:10.1016/j.cell.2018.05.029
Stolla, M. C., Catherman, S. C., Kingsley, P. D., Rowe, R. G., Koniski, A. D., Fegan, K., et al. (2019). Lin28b regulates age-dependent differences in murine platelet function. Blood Adv. 3 (1), 72–82. doi:10.1182/bloodadvances.2018020859
Sugiyama, T., Kohara, H., Noda, M., and Nagasawa, T. (2006). Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 25 (6), 977–988. doi:10.1016/j.immuni.2006.10.016
Sun, Y., Kubota, S., Iimori, M., Hamashima, A., Murakami, H., Bai, J., et al. (2022). The acidic domain of Hmga2 and the domain's linker region are critical for driving self-renewal of hematopoietic stem cell. Int. J. Hematol. 115 (4), 553–562. doi:10.1007/s12185-021-03274-9
Suo, M., Rommelfanger, M. K., Chen, Y., Amro, E. M., Han, B., Chen, Z., et al. (2022). Age-dependent effects of Igf2bp2 on gene regulation, function, and aging of hematopoietic stem cells in mice. Blood 139 (17), 2653–2665. doi:10.1182/blood.2021012197
Tabilas, C., Wang, J., Liu, X., Locasale, J. W., Smith, N. L., and Rudd, B. D. (2019). Cutting edge: elevated glycolytic metabolism limits the formation of memory CD8(+) T cells in early life. J. Immunol. 203 (10), 2571–2576. doi:10.4049/jimmunol.1900426
Tan, L., Xu, H., Chen, G., Wei, X., Yu, B., Ye, J., et al. (2018). Silencing of HMGA2 reverses retardance of cell differentiation in human myeloid leukaemia. Br. J. Cancer 118 (3), 405–415. doi:10.1038/bjc.2017.403
Tieppo, P., Papadopoulou, M., Gatti, D., McGovern, N., Chan, J. K. Y., Gosselin, F., et al. (2020). The human fetal thymus generates invariant effector γδ T cells. J. Exp. Med. 217 (3), 20190580. doi:10.1084/jem.20190580
Ueda, T., Tsuji, K., Yoshino, H., Ebihara, Y., Yagasaki, H., Hisakawa, H., et al. (2000). Expansion of human NOD/SCID-repopulating cells by stem cell factor, Flk2/Flt3 ligand, thrombopoietin, IL-6, and soluble IL-6 receptor. J. Clin. Invest. 105 (7), 1013–1021. doi:10.1172/JCI8583
Vanhee, S., Akerstrand, H., Kristiansen, T. A., Datta, S., Montano, G., Vergani, S., et al. (2019). Lin28b controls a neonatal to adult switch in B cell positive selection. Sci. Immunol. 4 (39), eaax4453. doi:10.1126/sciimmunol.aax4453
Wang, D., Tanaka-Yano, M., Meader, E., Kinney, M. A., Morris, V., Lummertz da Rocha, E., et al. (2022). Developmental maturation of the hematopoietic system controlled by a Lin28b-let-7-Cbx2 axis. Cell Rep. 39 (1), 110587. doi:10.1016/j.celrep.2022.110587
Wang, G., Miyahara, Y., Guo, Z., Khattar, M., Stepkowski, S. M., and Chen, W. (2010). Default generation of neonatal regulatory T cells. J. Immunol. 185 (1), 71–78. doi:10.4049/jimmunol.0903806
Wang, J., Wissink, E. M., Watson, N. B., Smith, N. L., Grimson, A., and Rudd, B. D. (2016). Fetal and adult progenitors give rise to unique populations of CD8+ T cells. Blood 128 (26), 3073–3082. doi:10.1182/blood-2016-06-725366
Wang, L. D., Rao, T. N., Rowe, R. G., Nguyen, P. T., Sullivan, J. L., Pearson, D. S., et al. (2015). The role of Lin28b in myeloid and mast cell differentiation and mast cell malignancy. Leukemia 29 (6), 1320–1330. doi:10.1038/leu.2015.19
Wang, S., Chim, B., Su, Y., Khil, P., Wong, M., Wang, X., et al. (2019). Enhancement of LIN28B-induced hematopoietic reprogramming by IGF2BP3. Genes Dev. 33 (15-16), 1048–1068. doi:10.1101/gad.325100.119
Watarai, H., Rybouchkin, A., Hongo, N., Nagata, Y., Sakata, S., Sekine, E., et al. (2010). Generation of functional NKT cells in vitro from embryonic stem cells bearing rearranged invariant Valpha14-Jalpha18 TCRalpha gene. Blood 115 (2), 230–237. doi:10.1182/blood-2009-04-217729
Watson, N. B., Patel, R. K., Kean, C., Veazey, J., Oyesola, O. O., Laniewski, N., et al. (2024). The gene regulatory basis of bystander activation in CD8(+) T cells. Sci. Immunol. 9 (92), eadf8776. doi:10.1126/sciimmunol.adf8776
Webb, C. F., Bryant, J., Popowski, M., Allred, L., Kim, D., Harriss, J., et al. (2011). The ARID family transcription factor bright is required for both hematopoietic stem cell and B lineage development. Mol. Cell Biol. 31 (5), 1041–1053. doi:10.1128/MCB.01448-10
Wells, A. C., Daniels, K. A., Angelou, C. C., Fagerberg, E., Burnside, A. S., Markstein, M., et al. (2017). Modulation of let-7 miRNAs controls the differentiation of effector CD8 T cells. Elife 6, e26398. doi:10.7554/eLife.26398
Wilson, A., Laurenti, E., Oser, G., van der Wath, R. C., Blanco-Bose, W., Jaworski, M., et al. (2008). Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair. Cell 135 (6), 1118–1129. doi:10.1016/j.cell.2008.10.048
Xu, B., Zhang, K., and Huang, Y. (2009). Lin28 modulates cell growth and associates with a subset of cell cycle regulator mRNAs in mouse embryonic stem cells. RNA 15 (3), 357–361. doi:10.1261/rna.1368009
Xu, X., Deobagkar-Lele, M., Bull, K. R., Crockford, T. L., Mead, A. J., Cribbs, A. P., et al. (2020). An ontogenetic switch drives the positive and negative selection of B cells. Proc. Natl. Acad. Sci. U. S. A. 117 (7), 3718–3727. doi:10.1073/pnas.1915247117
Yamamoto, R., Wilkinson, A. C., Ooehara, J., Lan, X., Lai, C. Y., Nakauchi, Y., et al. (2018). Large-scale clonal analysis resolves aging of the mouse hematopoietic stem cell compartment. Cell Stem Cell 22 (4), 600–607. doi:10.1016/j.stem.2018.03.013
Yang, S., Gu, Y., Wang, G., Hu, Q., Chen, S., Wang, Y., et al. (2019). HMGA2 regulates acute myeloid leukemia progression and sensitivity to daunorubicin via Wnt/β-catenin signaling. Int. J. Mol. Med. 44 (2), 427–436. doi:10.3892/ijmm.2019.4229
Yin, R., Chang, J., Li, Y., Gao, Z., Qiu, Q., Wang, Q., et al. (2022). Differential m(6)A RNA landscapes across hematopoiesis reveal a role for IGF2BP2 in preserving hematopoietic stem cell function. Cell Stem Cell 29 (1), 149–159.e7. doi:10.1016/j.stem.2021.09.014
Yokomizo, T., Ideue, T., Morino-Koga, S., Tham, C. Y., Sato, T., Takeda, N., et al. (2022). Independent origins of fetal liver haematopoietic stem and progenitor cells. Nature 609 (7928), 779–784. doi:10.1038/s41586-022-05203-0
Yoshimoto, M., Kosters, A., Cornelius, S., Valiente, N., Cheng, H., Latorre, A., et al. (2022). Mast cell repopulating ability is lost during the transition from pre-HSC to FL HSC. Front. Immunol. 13, 896396. doi:10.3389/fimmu.2022.896396
Yoshimoto, M., Montecino-Rodriguez, E., Ferkowicz, M. J., Porayette, P., Shelley, W. C., Conway, S. J., et al. (2011). Embryonic day 9 yolk sac and intra-embryonic hemogenic endothelium independently generate a B-1 and marginal zone progenitor lacking B-2 potential. Proc. Natl. Acad. Sci. U. S. A. 108 (4), 1468–1473. doi:10.1073/pnas.1015841108
Yoshimoto, M., Porayette, P., Glosson, N. L., Conway, S. J., Carlesso, N., Cardoso, A. A., et al. (2012). Autonomous murine T-cell progenitor production in the extra-embryonic yolk sac before HSC emergence. Blood 119 (24), 5706–5714. doi:10.1182/blood-2011-12-397489
Yuan, J., Nguyen, C. K., Liu, X., Kanellopoulou, C., and Muljo, S. A. (2012). Lin28b reprograms adult bone marrow hematopoietic progenitors to mediate fetal-like lymphopoiesis. Science 335 (6073), 1195–1200. doi:10.1126/science.1216557
Zhang, J., Ratanasirintrawoot, S., Chandrasekaran, S., Wu, Z., Ficarro, S. B., Yu, C., et al. (2016). LIN28 regulates stem cell metabolism and conversion to primed pluripotency. Cell Stem Cell 19 (1), 66–80. doi:10.1016/j.stem.2016.05.009
Zhou, Y., Li, Y. S., Bandi, S. R., Tang, L., Shinton, S. A., Hayakawa, K., et al. (2015). Lin28b promotes fetal B lymphopoiesis through the transcription factor Arid3a. J. Exp. Med. 212 (4), 569–580. doi:10.1084/jem.20141510
Keywords: fetal and adult hematopoietic stem cells, HSC expansion, LIN28B, Let7, HMGA2, IGF2BP2
Citation: Cox G, Kobayashi M, Rudd BD and Yoshimoto M (2025) Regulation of HSC development and function by Lin28b. Front. Cell Dev. Biol. 13:1555877. doi: 10.3389/fcell.2025.1555877
Received: 05 January 2025; Accepted: 24 February 2025;
Published: 12 March 2025.
Edited by:
Pawan Kumar Raghav, University of California, San Francisco, United StatesReviewed by:
Thierry Jaffredo, Centre National de la Recherche Scientifique (CNRS), FranceCopyright © 2025 Cox, Kobayashi, Rudd and Yoshimoto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Momoko Yoshimoto, TW9tb2tvLllvc2hpbW90b0B3bWVkLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.