
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell Dev. Biol., 21 November 2024
Sec. Molecular and Cellular Reproduction
Volume 12 - 2024 | https://doi.org/10.3389/fcell.2024.1493872
 Andrea Gómez-Zúñiga1
Andrea Gómez-Zúñiga1 Daniel Adrián Landero-Huerta2
Daniel Adrián Landero-Huerta2 Julio César Rojas-Castañeda2
Julio César Rojas-Castañeda2 Karla Sánchez-Huerta3
Karla Sánchez-Huerta3 Itzel Jatziri Contreras-García2
Itzel Jatziri Contreras-García2 Rafael Reynoso-Robles4
Rafael Reynoso-Robles4 Marcela Arteaga-Silva5
Marcela Arteaga-Silva5 Rosa María Vigueras-Villaseñor2*
Rosa María Vigueras-Villaseñor2*Background: Congenital hypothyroidism (CH) is a pathology that affects various organs, including the testicles. The mechanisms by which this condition alters fertility is unknown. This study aimed at determining if experimental CH affects gonocyte differentiation and arrests meiosis; and the possible role of the Sertoli cell (SC) in this condition.
Material and Methods: Two groups of rats consisting of Control group and Methimazole (MMI) induced CH rats were formed. The induction of CH was achieved by the administration of MMI starting on day 16 postcoitum and continued until euthanized. Euthanasia was performed at 5, 8, 10, 16 and 64 days of age. Following this, the testicular tissue of each animal was extracted and processed for histopathological and ultrastructural analysis. In addition, the tissue was used for the determination of proteins and their transcriptions, events which are characteristics of gonocyte differentiation. The SC functionality proteins was determined immunohistochemically, while sperm parameters of the cauda epididymis were verified.
Results: CH caused a delay in the gonocyte differentiation, and arrested meiosis and spermiogenesis. These events had long-term repercussions on the quality of the seminiferous epithelium. The results show that CH induces alterations in the functional state of SCs that may have led to the deficiency in the synthesis and/or in the release of molecules necessary for gonocyte differentiation; as well as disorders in the process of meiosis that resulted in sperm absence.
Conclusion: These results suggest that CH affects gonocyte differentiation and arrests meiosis, possibly through altering the functional status of SCs.
Congenital hypothyroidism (CH) is one of the most common endocrine disorders (Grüters and Krude, 2011). Its prevalence in Europe is put to one in every 2,000 to 3,000 newborns (van Trotsenburg et al., 2021). It is a pathology of great importance for its effects on metabolism (Pan et al., 2019); and on different organs, such as the skin, heart, kidneys, among others (Maciel et al., 2013; Yousefichaijan et al., 2015; Omidi et al., 2020). Induced CH in experimental rodents has also been associated with fertility problems as a result of decreased concentrations of circulating testosterone, progesterone, prolactin, and negative effect on sperm quality or inhibition of spermatogenesis (Umezu et al., 2004; Anbalagan et al., 2010; Aiceles et al., 2017).
It is known that thyroid hormones (TH) are necessary for the maturation of Sertoli cells (SCs) (Palmero et al., 1989; Bunick et al., 1994; de França et al., 1995), as well as for their ultrastructural, biochemical and molecular maintenance. Hence, they are essential for adequate spermatogenesis (Cooke et al., 1994; de França et al., 1995; Palmero et al., 1995; Zanatta et al., 2013). It has been reported that in rdw rats with CH due to missense mutation of the thyroglobulin gene, the SCs are unable to detain their proliferations, which is a necessary process for the maturation of these cells (Sakai et al., 2004). In the rat, the receptor for TH (TR) α1 (TRα1) isoform is the one expressed in immature, proliferating and adult SCs. In turn, TRα and TR β1 isoforms are expressed in spermatogonia, spermatocytes, round and elongated spermatids (Buzzard et al., 2000; Fadlalla et al., 2017).
In addition, the expression of TRs, in rats, is high at birth with a peak at 16 days postpartum (dpp) and decreases in adulthood (Buzzard et al., 2000; Jannini et al., 2000). During this period of maximum expression, gonocytes differentiate into spermatogonia, and the first spermatogenic wave begins with meiosis (Culty, 2009).
Gonocytes synthesize proteins as POU domain class 5 transcription factor 1 (POU5F1), (Zhao and Garbers, 2002); transcription factor AP2 (TFAP2C) (Chazaud et al., 1996); sal-like protein 4 (SALL4) (Eildermann et al., 2012); that confer to them pluripotent ability, undifferentiated characteristic, and survival potential. During their differentiation, these proteins are downregulated and the cells migrate towards the basal membrane where they change shape and begin the expression of Stra8 protein, events that are purely characteristics of spermatogonia (Culty, 2009). The Stra8 protein is necessary for the initiation and continuity of meiosis (Koubova et al., 2006). Simorangkir et al. (1997) reported a delay in the differentiation of gonocytes in experimental neonatal hypothyroidism induced by the administration of propylthiouracil (PTU).
Therefore, the mechanisms of damage generated by CH on gonocyte differentiation, the inhibition of spermatogenesis and sperm quality previously reported are not known (Umezu et al., 2004; Anbalagan et al., 2010; Aiceles et al., 2017). Hence, the evidences here exposed, depict the possibility that TH regulate the function of SCs for the production of paracrine molecules that are necessary in the differentiation of gonocytes and in the initiation and continuation of meiosis (Cooke et al., 1994; de França et al., 1995; Palmero et al., 1995; Simorangkir et al., 1997; Zanatta et al., 2013; Meroni et al., 2019). Therefore, the objective of the present work was to determine the mechanisms by which experimental CH affects gonocyte differentiation and stops germ cell meiosis, as well as the role of the SC in this affectation.
The animals were handled in accordance with the official Mexican standard (SAGARPA NOM-062-Z00-1999) “Technical specifications for the production, care and use of laboratory animals” and with the internal regulation on ethical principles and treatment of animals of the Institutional Committee for the Care and Use of Laboratory Animals of the National Institute of Pediatrics, Mexico City, Mexico, with registration number INP019/2019.
Wistar rats were bred and maintained in the vivarium of the National Institute of Pediatrics. The animals were allowed free access to food (Rodent Lab Chow 5001, Purina Inc.) and water, and maintained at a temperature of 21°C ± 2°C, relative humidity 50% ± 10% and a 12-h light/dark cycles (light on 06:00). At night, the male rats (∼250 g body weight) were paired with females (∼200 g body weight) in estrus to have pups. The next morning, vaginal smears were obtained from the females to confirm the presence of sperm. If the vaginal smear was positive, this day was recorded as day 0 postcoitum. Subsequently, the pregnant animals were divided into two groups: the Control (without administration of any drug in the drinking water) and the exposed to methimazole (MMI). The administration of MMI (1-methyl-3H-imidazole-2-thione Sigma-Aldrich, Mexico City, Mexico) was via drinking water containing the substance (23 mg/kg body weight/day) administered to the mothers from 16 days postcoitum (dpc) until the pups were euthanized. This dose was calculated daily based on water consumption the day before its preparation. In rats, the formation of thyroid gland is completed on the day 16 of gestation, and the synthesis of TH is begun. MMI acts by inhibiting the activity of thyroid peroxidase, which participates in the iodination of tyrosine residues, preventing the synthesis of TH (Awosika et al., 2023).
At birth, the litters were adjusted to 8 pups and all the males were maintained and kept in the vivarium until euthanized on 5, 8, 10, 16 and 64 dpp. The sacrifice of the animals was carried out by an overdose of pentobarbital administered intraperitoneally in saline solution (120–150 mg/kg) after sedation with xylazine (10 mg/kg) (Pfizer, Mexico City, Mexico). Also, the body and testicular weight of each animal was recorded (Figure 1).
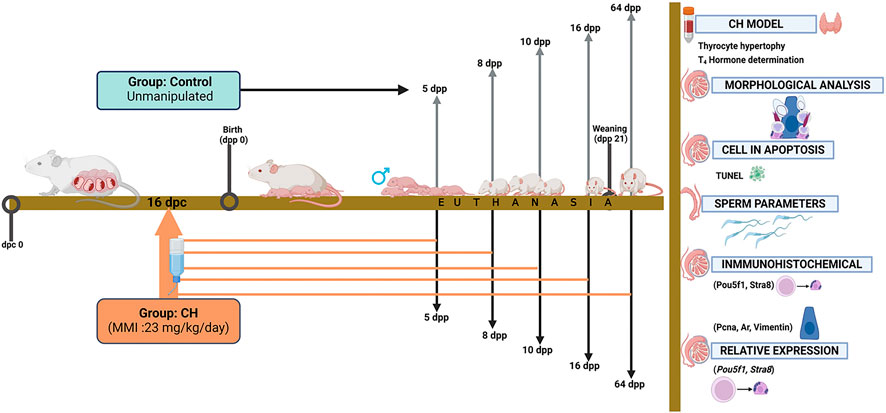
Figure 1. Experimental design and sample processing. The Control group received no drug in the drinking water and the CH group received MMI from 16 dpc. Euthanasia was carried out in both groups at 5, 8, 10, 16 and 64 dpp. Blood and thyroid samples were obtained for model validation. The testes were analyzed for morphology, ultrastructure; and apoptosis and sperm parameters were determined. Specific proteins were analyzed by immunohistochemistry and relative expression to determine gonocyte differentiation and Sertoli cell functionality. CH, congenital hypothyroidism; MMI, methimazole; dpc, days postcoitum; dpp, days postpartum.
The determination of T4 concentrations was performed to verify the presence of CH in the animals. For this, blood sample was obtained by cardiac puncture from each of the animals between 1:00 p.m. and 2:00 p.m. to avoid circadian fluctuations. The samples were centrifuged at 12,000 revolutions per minute for 10 min to obtain serum. T4 determination was performed using an ELISA kit (Calbiotech, Cordell Court, El Cajon, CA) following the manufacturer’s instructions.
Half of the right testicular sample plus the thyroid gland tissue were fixed in Karnovsky solution for 2 h. Later, they were postfixed in 1% osmium tetroxide (OsO4) (Sigma Aldrich, St Louis MO. United States), dehydrated and processed for inclusion in Epon 812 (Ted Pella, Inc., Redding, CA, United States). Sectioning of the samples were made at 1 µm thickness using an ultramicrotome (Ultracut UCT, Leica, Vienna, Austria) and were stained with 0.5% toluidine blue. Histological analysis was performed using a BX 51 Olympus optical microscope (Olympus Corp., Tokyo, Japan). Fifteen to twenty cross sections of the seminiferous cords/tubes were evaluated for each animal. The area of the seminiferous epithelium was determined using an image analysis system (Image-Pro Plus 7.0, Media Cybernetics, INC. MD, United States). The number of Sertoli cells, gonocytes, spermatogonia, spermatocytes, round and elongated spermatids per cords/tubes seminiferous was obtained. The degree of maturation of the seminiferous epithelium (MI), called the Johnsen Index (Johnsen, 1970) and the histopathological index (HI) (Vigueras-Villaseñor et al., 2009) were also determined.
The gonocyte differentiation index (number of spermatogonia/number of gonocytes) was obtained. Finally, the thyroid gland was analyzed to determine thyrocyte hypertrophy in the CH model. All histological evaluations were performed by a single observer.
To determine apoptosis, the terminal deoxynucleotidyl transferase-mediated dUDP nick-end labelling (TUNEL) assay was used. The testicular portions were fixed in 4% paraformaldehyde for 18 h. These tissue samples were dehydrated, classified and embedded in paraffin. Afterwards, each tissue sample was cut at 4 μm thickness and mounted on a slide coated with poly-L-lysine (Sigma Aldrich). Subsequently, it was deparaffinized and dehydrated in a graded series of alcohols. Then, the tissue sections were treated with Triton X-100 solution (Sigma) at 0.1% for 2 min. They were later incubated with TUNEL solution (50 µL terminal deoxynucleotidyl transferase and 450 µL nucleotide mixture, in situ Cell Death Detection Kit, Roche Diagnostic Corporation, Indianapolis, IN, United States) for 1 h at 37°C. The tissue sections were then mounted with fluorescence medium (Fluoroshield Mounting Medium with DAPI, ABCAM) for observation using an Olympus fluorescence microscope (Olympus BX51). The number of apoptotic cells per seminiferous cords/tubes was calculated and the results were expressed as the number of apoptotic cells per cross section. 15 to 20 transverse sections of the seminiferous cords/tubes were evaluated for each of the animals. All dilutions and washes between steps were performed using PBS (0.1 M). Negative control sections were processed identically, but the enzyme solution (terminal deoxynucleotidyl transferase) was omitted. All histological examinations were performed by a single observer.
The epididymis of each rat was dissected, and the adjacent associated tissue was removed, and placed in saline (154 mM NaCl at 38°C). The caudal portion of the epididymis was sectioned by radial cuts using fine-point scissors. This section was incubated for 5 min in 2 mL of BTS (Beltsville Thawing Solution, BTS™, Minitub™, Hauptstrasse, Tiefenbach, Germany) with occasional gentle shakes to allow the release of sperm into the medium. Subsequently, 10 µL of the medium with sperm were taken and placed on slides that were stained with hematoxylin-eosin to determine the percentage of normal sperm, discarding morpho-abnormalities (alterations in the head, midpiece, tail) per 100 sperm (Vigueras-Villaseñor et al., 2011). Sperm concentration was determined in a Neubauer chamber (BRANDGMBH, Wertheim, Germany) using a solution diluted in saline solution (1:20), and the results were expressed x106/mL. The percentage of vitality was obtained by evaluating 100 spermatozoa stained with Coomassie brilliant blue (BBC) (0.22% Coomassie blue G-250, in 50% methanol, 10% glacial acetic acid and 40% distilled water). All the analysis were performed in triplicate with Olympus BX51 microscope.
Tissue samples from the half of the right testicular of each of the animals were fixed in 4% paraformaldehyde (Sigma-Aldrich) and processed for inclusion in paraffin. Sections of 4 μm thickness were made and mounted on slides covered with poly-L-lysine (Sigma-Aldrich, St. Louis, MO, United States). Tissue sections were deparaffinized with xylene and hydrated with graded ethanol solutions. They were subsequently incubated with citrate buffer (0.01M, pH 7.6) for 5 min in a microwave oven at 800 W. Then, the sections were washed three times each for 5 min with phosphate buffer (PBS)/0.1% Tween 20 (Sigma-Aldrich) and blocked with 1% bovine serum albumin in PBS for 2 h in a humid chamber. Subsequently, they were incubated overnight at room temperature with the corresponding antibody. The following mouse monoclonal primary antibodies were used: 1) Anti-Pou5f1 antibody, pluripotency protein used to identify gonocytes (sc-5279, Santa Cruz, Biotechnology, CA, United States), at a 1:25 dilution; 2) Anti-proliferating-cell nuclear antigen (Pcna) antibody, Sertoli cell proliferation protein (sc-56, Santa Cruz, Biotechnology) at a 1:50 dilution; 3) Anti-Vimentin antibody, structural protein that allows the determination of the functionality of Sertoli cells, (sc-6260, Santa Cruz Biotechnology) at a 1:30 dilution. In addition, rabbit polyclonal primary antibodies were used as follows: 1) Anti-Stra8 antibody, protein for the initiation of meiosis, characteristic of spermatogonia (ab49602, ABCAM, Cambridge, United Kingdom) at a 1:50 dilution and 2) Anti-Androgen Receptor (Ar) antibody, a protein that allows the determination of the functionality of Sertoli cells (sc-816, Santa Cruz Biotechnology) at a 1:30 dilution. This was followed by another incubation with biotinylated secondary antibodies (Horse anti-mouse IgG Biotinylated BA 2000, lot ZJ1212 Vectastain Vector Labs, Burlington, ON, Canada, and with Goat anti-rabbit IgG Biotinylated, cat BA1000, Lot ZE1218, Vectastain Vector, respectively), at a dilution of 1:200 for 2 h at room temperature. The samples were then exposed to Avidin-Biotin System conjugate (ABC kit, Vectastain Vector Labs) for 1 h at room temperature. The reaction was evidenced using diaminobenzidine (DAB Chromogen Kit 901-DB801-032118, Biocare, Concord, CA, United States). Subsequently, tissue sections were counterstained with Gill’s hematoxylin, and coverslipped with entellan. The number of immunoreactive cells per transversely cut seminiferous cords/tubes was quantified. 15 to 20 seminiferous tubes per animal were evaluated. The distribution pattern of vimentin immunoreactivity in the cytoplasm of SCs was also determined.
For the animals of 5–16 dpp, total RNA was extracted from the tissue obtained from the halves of both testes of each animal, while for the animals of 64 dpp, the extraction of total RNA was from the testicular tissue obtained from the central part of each testis using the Qiagen® RNeasy Lipid Tissue Mini Kit (Cat. 74804, Hilden, Germany). Subsequently, relative expression was determined by RT-qPCR assays. For reverse transcription, the TaqMan Reverse Transcription Reagents kit (Life Technologies, United States) was used to obtain cDNA from aliquots of 100 ng of RNA for each sample. Subsequently, qPCR assays were performed using the TaqMan Universal Master Mix kit (Thermo Fisher, UK). 5 μL of Taqman Universal PCR Master Mix, 0.5 µL of the test probe as appropriate, Stra8 (RN01747849_m1) and Pou5f1 (Rn06413993_s1), 3.5 µL of nuclease-free water and 1 µL of cDNA were used for a total volume of 10 µL. Reactions were performed in triplicate in a thermocycler (Applied Biosystem Step One TM, Foster City, CA, United States) under the following conditions: 50°C for 2 min, 95°C for 10 min, 40 cycles at 95°C for 15 s and 60°C for 1 min. Expression levels were determined using the 2−ΔΔCT method. For data normalization, the probe (Gapdh, RN01775763_g1) was used as an endogenous control.
All data were expressed as the mean ± SEM. The Student’s T-test was used for statistical analysis in the SPSS package and values p < 0.05 was considered significant.
Animals with CH showed a significant reduction in body and testicular weight at dpp 16 and 64 compared to the Control, as well as in T4 concentrations (Table 1, p < 0.05), with thyrocyte hypertrophy (complementary Figure).
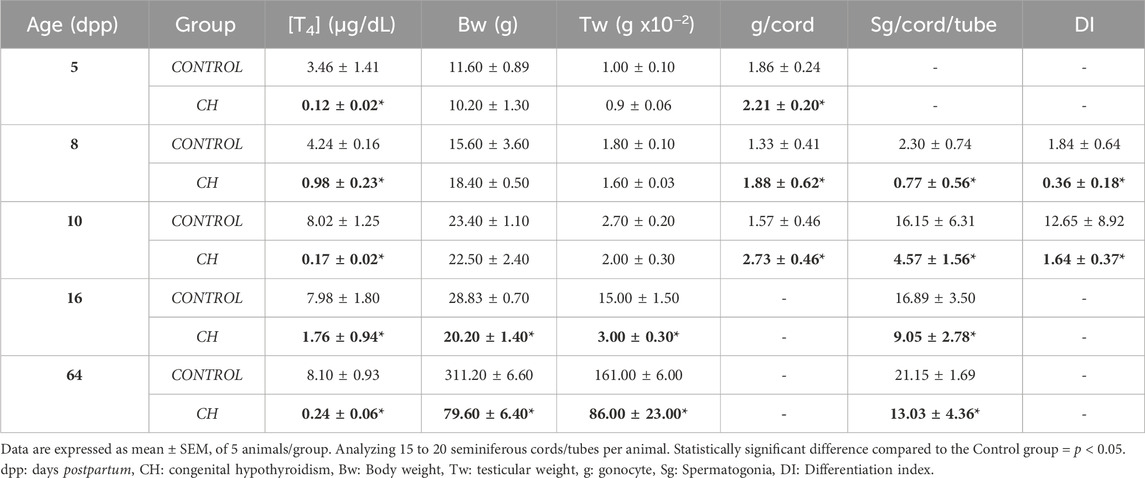
Table 1. Parameters evaluated in Control and CH animals.
The seminiferous cords/tubes of the animals in the Control group showed a normal cytoarchitecture, in accordance to their age. At 5 dpp, the gonocytes were found in migration phase towards the basement membrane, and occasionally, in the center of the cords (Figure 2A). At 8 dpp, the first spermatogonia were observed (Figure 2B). At 10 dpp, the first spermatocytes emerged (Figure 2C) and at 16 dpp, an increase in their proliferation was observed (Figure 2D). At 64 dpp, the seminiferous epithelium showed a complete spermatogenic process from spermatogonia to elongated spermatids (Figure 2E). In the seminiferous cords/tubes of animals with CH, as compared to the Control, a greater number of gonocytes was observed at 5, 8 and 10 dpp with a decrease in the number of spermatogonia starting at 8 dpp (Table 1; Figures 2F–H p < 0.05); thus, generating a reduction in the gonocyte differentiation index (Table 1, p < 0.05) at 8 and 10 dpp. Spermatocytes were evident until 16 dpp (Figure 2I); although, they were rarely present in some cords at 10 dpp. Germ cell populations were lower at all ages (Table 2, p < 0.05). The population of SCs was significantly reduced (p < 0.05) at 8 and 10 dpp; however, at 16 and 64 dpp, these cells were present in greater numbers (Table 2, p < 0.05). The MI was reduced starting from 8 dpp, while the HI increased (p < 0.05) at all ages, characterized by desquamation, vacuolization, folding of the basement membrane and pyknosis (Table 2, Figures 2G–J). In the animals with CH at 64 dpp, spermatogenesis was arrested, and spermatogonia, spermatocytes and in some cases round spermatids were observed in the seminiferous tubes (Figure 2J).
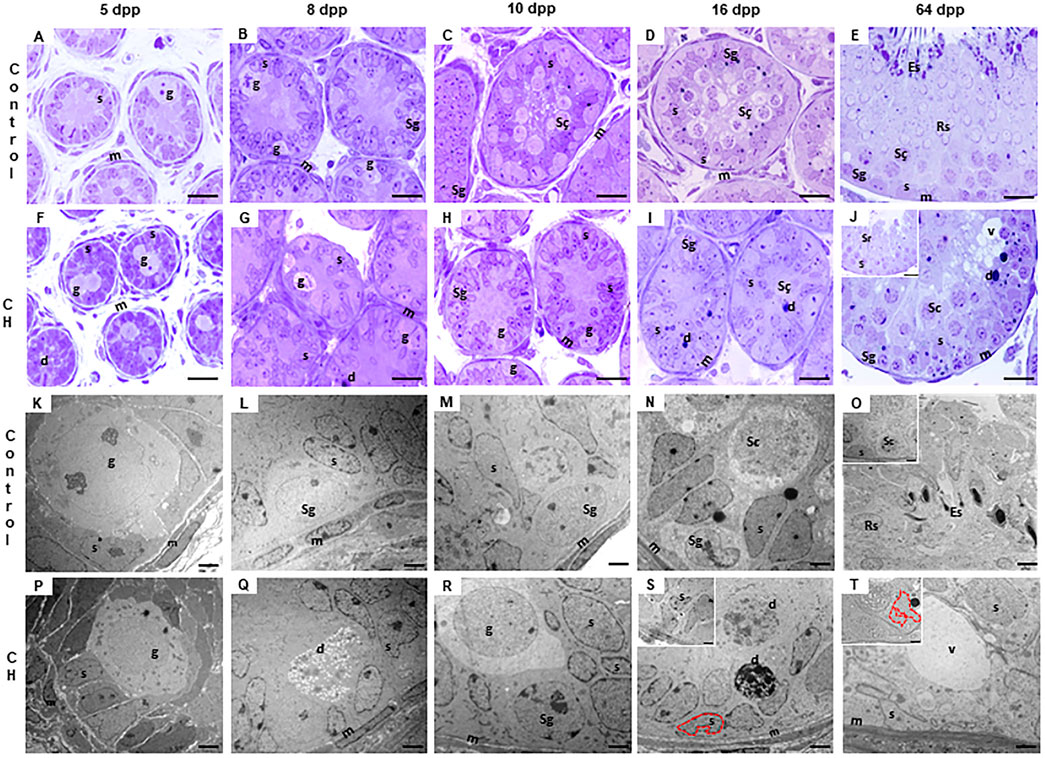
Figure 2. Cords/tubes seminiferous of different ages. It is observed in the group with CH (F–J, P–T), unlike the Control (A–E, K–O), the permanence of gonocytes until 10 dpp (H); delay in the emergence of spermatocytes at 16 dpp (I) and arrest of spermatogenesis at 64 dpp (J) with histological alterations, such as cellular degeneration and epithelial vacuolization. In the CH group, electron microscopy showed a delay in the migration of gonocytes until 10 dpp (P–R), cellular degeneration, vacuolization and Sertoli cell nuclei with deep indentations (S, T and image insert T) and displaced towards the central part of the cord (S, insert). Congenital hypothyroidism (CH), Gonocytes (g), Spermatogonia (Sg), spermatocytes (Sc), round (Rs) and elongated spermatids (Es), nuclei of Sertoli cells (s), myoid cells (m), degenerating cells (d), vacuoles (v). Toluidine Blue (A–J). Scale bar: 20 μm. Electron microscopy (K–T). Scale bar 2 μm.
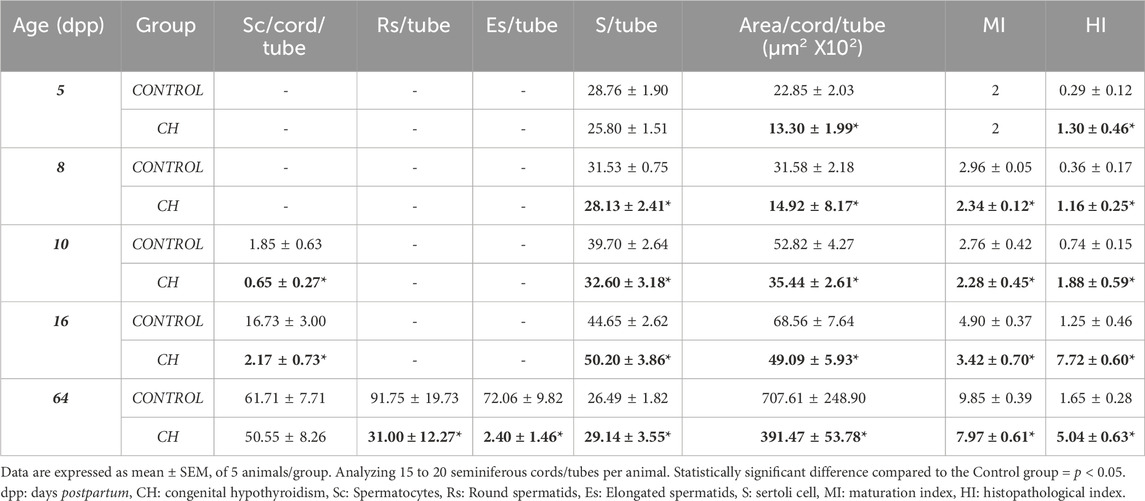
Table 2. Testicular histological parameters in Control and CH animals.
In the Control group, electron microscopy showed few cells in degeneration and with compact to slightly diffuse basement membranes at all ages, intact nuclei of the SCs moved to the base of the seminiferous epithelium from 16 dpp (Figures 2K–O). On the contrary, electron microscopy of the groups with CH showed germ cells in the process of degeneration and diffused basal membranes at all ages. SC nuclei were displaced towards the apical part of the seminiferous epithelium from 16 dpp, and these were observed with deep indentations of the nuclear envelope (Figures 2P–T).
In the CH group, apoptosis was found to be significantly increased (p < 0.05) at 16 and 64 dpp when compared to the Control group (Table 3, Figures 3A–D).
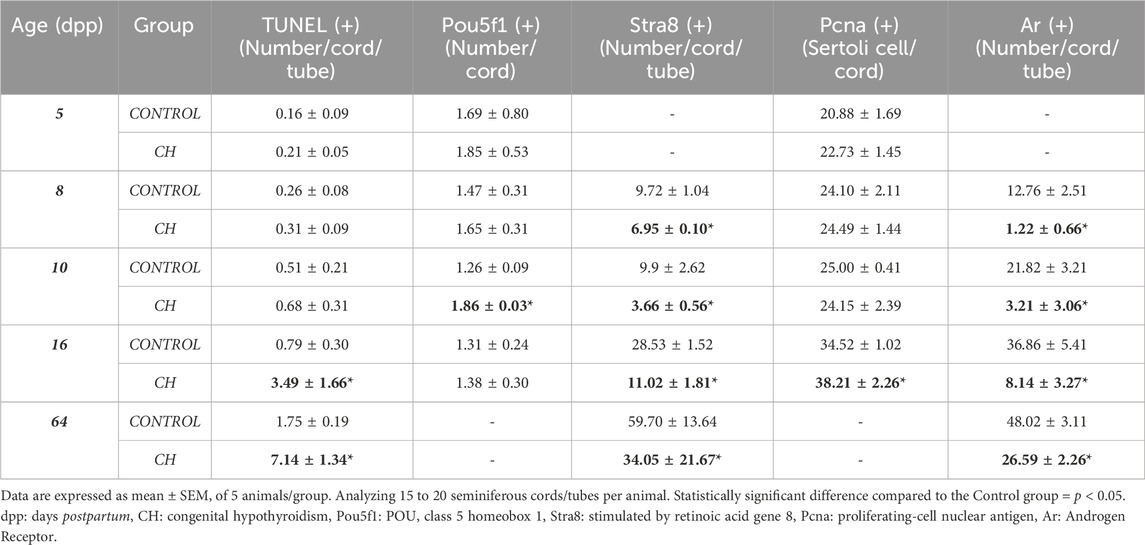
Table 3. Cell apoptosis and immunoreactivity.
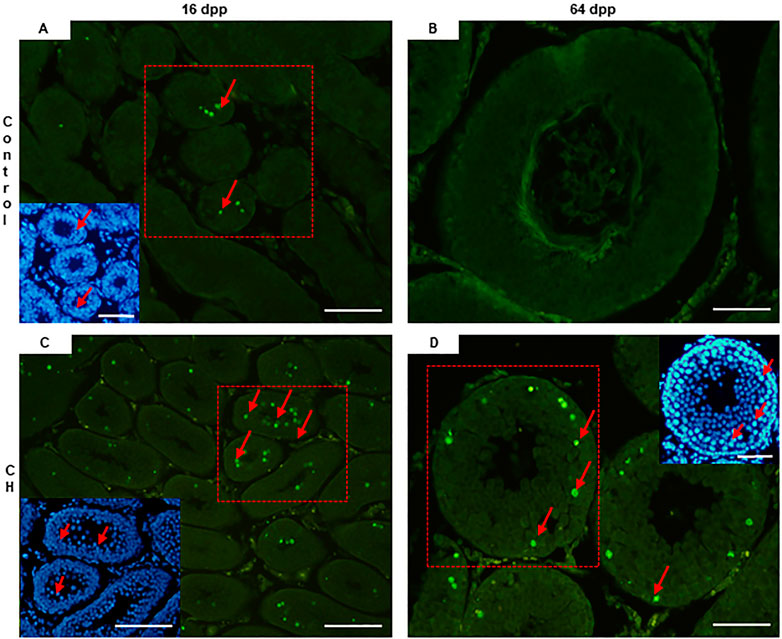
Figure 3. Seminiferous tubes with cells in apoptosis. (A) 16 dpp Control; (C) CH of 16 dpp; (B) 64 dpp Control; (D) CH of 64 dpp. At both ages, a greater number of apoptotic spermatocytes were seen in the CH group. Congenital hypothyroidism (CH). TUNEL. Inserts DAPI. Scale bar 20 µm.
Therefore, it was not possible to determine the sperm parameters in the CH group since no sperm were observed in the epididymis cauda, a situation contrary to the Control group where the sperm parameters showed a vitality of 73.2% ± 8.21%, concentration of 14.7 ± 1.19 × 106/mL and normal morphology of 83.8% ± 3.96%.
In the control group, immunoreactivity of Pou5f1 on day 5 and 8 was observed in gonocytes located in the center and periphery of the seminiferous cords and at 10 in gonocytes located at the periphery of the cords (Figures 4A–C). Immunoreactivity to Stra8, characteristic of spermatogonia, was observed from 8 dpp (Figures 4J–L). In the CH group at 5 and 8 dpp, Pou5f1 immunoreactivity was observed in gonocytes located mainly in the center of the seminiferous cords (Figures 4E, F) and at 10 dpp, unlike the Control, the number of gonocytes immunoreactive to the Pou5f1 protein increased (p < 0.05) (Table 3; Figure 4G). Starting from 8 dpp, the Stra8 protein was significantly reduced (p < 0.05, Figures 4N–P) with respect to the Control group (Table 3; Figures 4J–L).
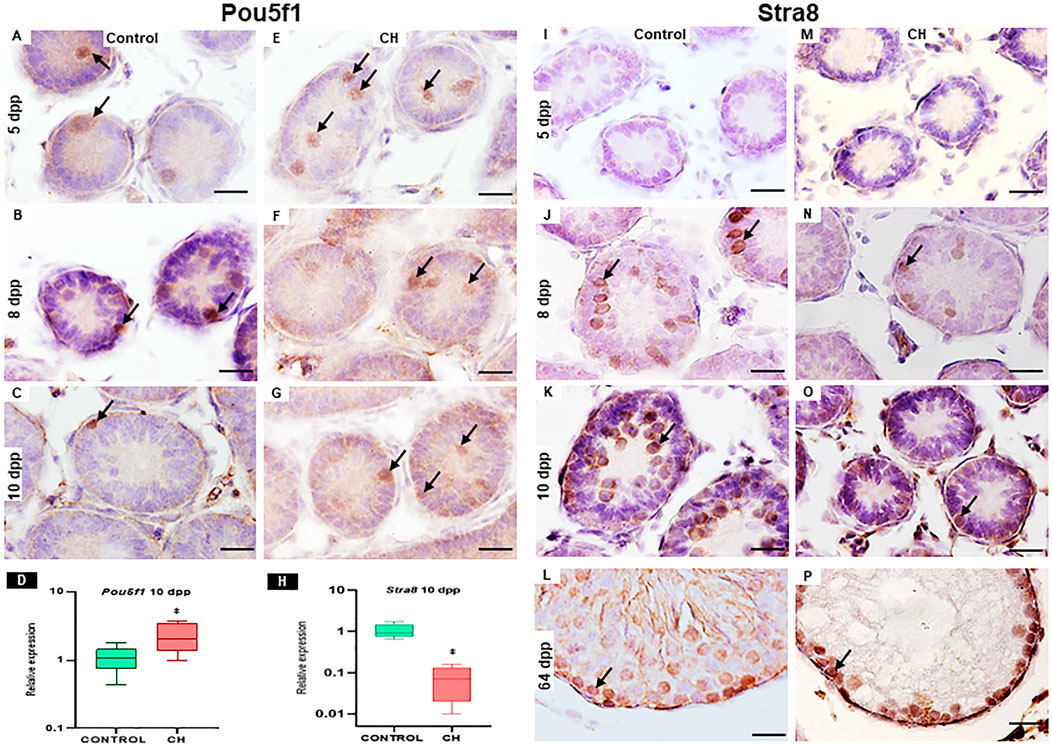
Figure 4. Cords and seminiferous tubes from Control and CH animals where immunoreactivity to Pou5f1 (arrow (A–G) and Stra8 (arrow (I–P)) can be seen. Greater immunoreactivity to Pou5f1 was observed at 10 dpp in the CH group (G) unlike the Control group (C). The Stra8 protein becomes evident from 8 dpp, with greater immunoreactivity in the Control group at all ages (I–L) compared to the CH group (M–P). Figures D and H show the relative expression of Pou5f1 and Stra8 at 10 dpp. Congenital hypothyroidism (CH). Scale Bar 20 μm.
The results of the relative expression at 10 dpp showed a significant increase in Pou5f1 (p < 0.05) and a significant reduction in Stra8 (p < 0.05) (Figures 4D, H) when compared with the Control group.
In the Control group, cell proliferation determined by Pcna immunoreactivity in SCs was observed from 5 to 16 dpp. (Figures 5A–D). The presence of Ar was evident from 8 dpp and the number of cells immunoreactive to this receptor increased reaching its maximum at 64 dpp (Figures 5J–M). The distributed of vimentin in the basal cytoplasm of the SCs took place from 5 to 10 dpp, when it was appreciated to surround the nucleus of these cells (Figures 5S–U). At 16 and 64 dpp, the distribution was perpendicular to the basal membrane; thus, facilitating the support of germ cells (Figures 5V, W). In the CH group, Pcna immunoreactivity presented a distribution pattern similar to the Control group, except at 16 dpp when it became significantly increased (p < 0.05) (Table 3; Figures 5E–H). The number of Ar-immunoreactive cells in the CH group was observed to reduced significantly (p < 0.05) from 8 dpp onwards (Table 3; Figures 5N–R) when compared with the Control group. In the groups with CH, vimentin distribution until 10 dpp was similar to that of the Control group; however, at 16 dpp, it remained concentrated at the base of the cytoplasm of the SC, which became more evident at 64 dpp when it was observed disorganized (Figures 5X–Ab).
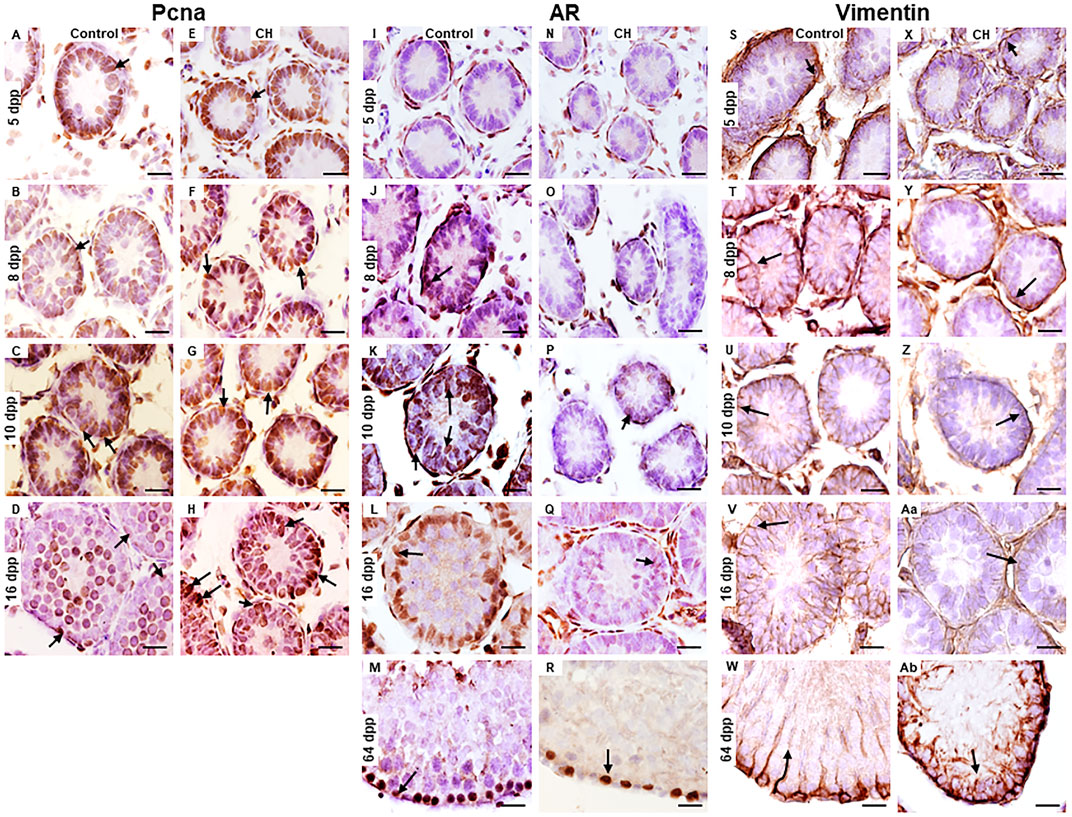
Figure 5. Cells in the cords and seminiferous tubes of Control and CH animals immunoreactive to the Pcna, Ar and Vimentin proteins. Sertoli cells immunoreactive for Pcna at 16 dpp were higher in the CH group (arrows E-H) unlike the Control (A–D). At 64 dpp, no Pcna immunoreactivity was observed in Sertoli cells. Immunoreactivity for Ar (I–R) in Sertoli cells was reduced in the CH group (N–R) unlike the Control (I–M) from 8 dpp. Immunoreactivity for vimentin was greater in the Control group (S–W), when compared the CH group (X-Ab) where its distribution was disorganized. Congenital hypothyroidism (CH). Scale bar 20 μm.
CH generated during gestation with MMI administration and using rdw rats with CH due to missense mutation of the thyroglobulin gene is associated with alterations in fertility of their male offspring through its negative effect on sperm quality or inhibition of spermatogenesis (Umezu et al., 2004; Anbalagan et al., 2010; Aiceles et al., 2017).
Considering that in early postnatal stages, there is a maximum expression of TRs in testicular cells (Jannini et al., 1990; Canale et al., 2001; Holsberger and Cooke, 2005), it was interesting to study how CH affected the early processes of gonocyte differentiation and meiosis of the first spermatogenic waves.
The CH model achieved by the administration of MMI during pregnancy was confirmed in our study by the low concentrations of T4 plus the thyrocyte hypertrophy that occurred at all ages, as well as the reduction in body and testicular weight after 16 dpp.
In the present study, a reduction in the differentiation index of gonocytes in the CH group was observed. This reduction was demonstrated by the increase in the number of gonocytes, the reduction in the number of spermatogonia and in the number of immunoreactive cells to Stra8 at 8, 10 and 16 dpp, added to an increase in the number of immunoreactive cells to Pou5f1 at 10 dpp supported by the expression of the messengers of these proteins at this same age. This event resulted in a lower population of germ cells, reduced area of the cords/tubes seminiferous with histological alterations. Furthermore, once spermatogenesis began, it did not continue beyond pachytene spermatocytes and on rare occasions, to round spermatids. This led to poor maturation of the seminiferous epithelium with azoospermia.
It has been reported that neonatal hypothyroidism delays the differentiation of gonocytes, since its population is increased, possibly due to arrest of their migration to the basement membrane (Simorangkir et al., 1997). However, this report did not point out a possible cause of this event. The administration of T3 in healthy neonatal rats tends to increase the differentiation of gonocytes and the number of spermatogonia (Faraone-Mennella et al., 2009; Marchlewska et al., 2013; Marchlewska et al., 2011). This may have resulted from the increase in platelet-derived growth factor beta receptor (PDGFR-β) protein, which is known to be involved in gonocyte differentiation (Basciani et al., 2008; Basciani et al., 2010). T3 treatment increases PDGFR-β levels in the heart of mice with PTU-induced hypothyroidism (Chen et al., 2012).
In line with the above, other authors have also reported a delay or an arrest of spermatogenesis in spermatocyte or spermatid using different CH models (Francavilla et al., 1991; Sakai et al., 2004; Yildirim et al., 2017). Furthermore, the presence of TRα has been demonstrated in intermediate spermatogonia and even in pachytene spermatocytes, which coincides with the low amount of these cell types due to the absence of the hormone (Jannini et al., 1999; Buzzard et al., 2000). Sarkar and Singh (2017) mentioned that TRα1 immunostaining is intense in the acrosomal region of elongated spermatids and when the neonatal hypothyroidism model with PTU is developed, it leads to the absence of elongated spermatids. This demonstrates the importance of THs in cell differentiation during spermiogenesis.
Retinoic acid is known to play an important role in gonocyte differentiation and the initiation of meiosis, inducing the expression of the Stra8 protein in A1 spermatogonia, preleptotene and pachytene spermatocytes (Koubova et al., 2006; Green et al., 2018). The presence of retinoic acid receptors in Sertoli and germ cells has been demonstrated, and their alteration generates fertility failures (Griswold, 2016). CH has been documented to affect the expression of retinoic acid receptor β in the liver (Pallet et al., 1994). It is likely that the deficiency of this receptor is related to the decrease in the immunoreactivity to the Stra8 protein and its expression at 10 dpp, which alters the differentiation of gonocytes and the continuation of meiosis. This suggests the possibility of Sertoli cell dysfunction, without ruling out that CH may directly affect post-gonocyte germ cells, since these cell types also present TRs (Buzzard et al., 2000; Fadlalla et al., 2017). TRs in germ cells have been associated with their survival (Buzzard et al., 2000). CH generates a reduction or absence in the expression of TRs (Fadlalla et al., 2017; Sarkar and Singh, 2017). This probably led to apoptosis of testicular cells as seen in this work.
In the present study, it was observed that from 8 dpp through all the ages, the number of SCs immunoreactive to Ar in animals with CH was lower. It has been documented that T3 upregulates the mRNA for Ar in SCs (Arambepola et al., 1998) since this steroid is important to support their differentiation and survival (Walczak-Jedrzejowska et al., 2007). Panno et al. (1996) mentioned that T3 increases Ar in peripubertal SCs. When T3 and androgens are administered together, they generate a positive regulation of Ar, which could support the role of both hormones in the responsiveness of SCs to androgens during spermatogenesis (Panno et al., 1996). It was reported that Ar deficiency is associated to immaturity of SCs (Arambepola et al., 1998). This immaturity has also been demonstrated in neonatal hypothyroidism due to estrogen production, characteristic of immature SCs (Sarkar and Singh, 2017). In addition, this situation can be accentuated by the continued proliferation of SCs beyond 15 dpp (Gaytan et al., 1986; Picut et al., 2015), that brought about an increase in the population of these cells, as was observed in the present work at 16 dpp in the CH group. Therefore, these facts support the theory of a delay in maturation or dysfunction of SCs that affects the synthesis and/or transmission of molecules necessary for the maintenance and differentiation of germ cells.
Alterations in the transmission of maintenance and differentiation molecules may be due to the effects on connexin 43, which has been shown to be regulated by THs (Gilleron et al., 2006; Rodrigues et al., 2022). The expression of this connexin is reduced in PTU induced neonatal hypothyroidism (Gilleron et al., 2006). Connexin 43 is a component of gap junctions that allows communication among SCs, and between these cells and germ cells (Decrouy et al., 2004); thus, it helps to maintain the ultrastructure and maturation of SCs. The seminiferous epithelium of Sertoli cell-specific connexin 43 knockout mice (SCCx43KO−/−) has been reported to show vacuolization caused by the absence of germ cells with disorganization of the cytoplasmic processes of SCs. These cells lose polarity, generating nuclei with deep indentations of their nuclear envelope associated with vimentin and their displacement towards the central region of the tubes (Aumüller et al., 1992; Staggenborg et al., 2022), as observed in the present work on animals with CH. The gonocytes, the spermatogonia and the spermatocytes that failed to receive molecules for their maintenance and differentiation degenerated by apoptosis as observed at 16 and 64 dpp, which generated a reduction in the population of germ cells and the absence of spermatozoa. A reduction in the number of germ cells and live sperm has been reported in rats with persistent hypothyroidism from birth (Sahoo et al., 2008; La Vignera and Vita, 2018). It is mentioned that this reduction may be caused by an imbalance between the increase in free radicals and the reduction of the endogenous antioxidant system, such as catalase and superoxide dismutase (Choudhury et al., 2003; Sahoo et al., 2008). In addition, Zamoner et al. (2008) reported that superoxide dismutase, glutathione S-transferase and Glutathione reductase enzymes were affected by PTU-induced CH. In this work, however, the endogenous antioxidant system enzymes were not determined.
Furthermore, alterations in vimentin distribution in the CH group backs up the idea of Sertoli cell dysfunction. Vimentin is an intermediate microfilament that is expressed in SCs (Romeo et al., 1995; Show et al., 2003; Franke et al., 2004). It plays several roles in cell architecture, such as maintaining the cytoplasmic structural integrity that gives support to developing germ cells, surrounding the nucleus and keeping it fixed in its location, being deposited in the indentations of the nuclear envelope, and forming binding structures that are necessary for the transmission of molecules for spermatogenesis (Russell and Peterson, 1985; Aumüller et al., 1992; Tanemura et al., 1994; Show et al., 2003). Zamoner et al. (2008) reported that CH generates hyperphosphorylation of vimentin without affecting its expression, but leads to accumulation of insoluble vimentin. Cell junction and communication proteins are important for the migration of gonocytes from the center of the cord to the basal membrane and for the movement of spermatocytes from the basal to the apical compartment, as well as for the transmission of molecules from the SCs for their maintenance and differentiation. Hence, the disorganization of vimentin, as observed in this study, could have interfered with the migration of gonocytes and their differentiation to spermatogonia in the CH rat model.
It would be important to analyze whether the administration of T4 in CH model, favors the functioning of Sertoli cells by generating differentiation of gonocytes, completion of meiosis and conclusion of spermiogenesis, reducing cell death and restoring sperm parameters.
CH generated a delay in the differentiation of gonocytes, arrest of meiosis and conclusion of spermiogenesis. This may be caused by alterations in the SCs functions as was demonstrated by the deficiency in immunoreactivity to Ar, vimentin and the presence of altered nuclei with deep indentations that probably generated a deficiency in the synthesis and/or transmission of proteins and factors necessary for differentiation, which could have had an impact on poor cell proliferation and differentiation with sperm absence.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
The animal study was approved by Institutional Committee for the Care and Use of Laboratory Animals of the National Institute of Pediatrics, Mexico City, Mexico. The study was conducted in accordance with the local legislation and institutional requirements.
AG-Z: Data curation, Investigation, Methodology, Writing–original draft, Writing–review and editing. DL-H: Methodology, Supervision, Writing–review and editing. JR: Data curation, Supervision, Writing–review and editing, Investigation. KS-H: Conceptualization, Methodology, Supervision, Writing–review and editing. IC-G: Methodology, Writing–review and editing. RR-R: Investigation, Methodology, Writing–review and editing. MA-S: Investigation, Methodology, Supervision, Writing–review and editing. RV-V: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing–original draft, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by Instituto Nacional de Pediatria (grant number INP: 19/2019, E-022). This work was supported by a CONAHCYT (814684) fellowship to Andrea Gómez Zúñiga.
The authors are grateful to Pedro Medina Granados and Edgar Daniel Cervantes Arias for their invaluable support in processing the samples. We thank Cyril Ndidi Nwoye Nnamezie (MD), a native English speaker and an expert translator for his help in preparing the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2024.1493872/full#supplementary-material
Aiceles, V., Gombar, F., and da Fonte Ramos, C. (2017). Hormonal and testicular changes in rats submitted to congenital hypothyroidism in early life. Mol. Cell Endocrinol. 439, 65–73. doi:10.1016/j.mce.2016.10.026
Anbalagan, J., Sashi, A. M., Vengatesh, G., Stanley, J. A., Neelamohan, R., and Aruldhas, M. M. (2010). Mechanism underlying transient gestational-onset hypothyroidism-induced impairment of posttesticular sperm maturation in adult rats. Fertil. Steril. 93, 2491–2497. doi:10.1016/j.fertnstert.2010.02.005
Arambepola, N. K., Bunick, D., and Cooke, P. S. (1998). Thyroid hormone effects on androgen receptor messenger RNA expression in rat Sertoli and peritubular cells. J. Endocrinol. 156, 43–50. doi:10.1677/joe.0.1560043
Aumüller, G., Schulze, C., and Viebahn, C. (1992). Intermediate filaments in Sertoli cells. Microsc. Res. Tech. 20, 50–72. doi:10.1002/jemt.1070200107
Awosika, A. O., Singh, G., and Correa, R. (2023). “Methimazole,” in StatPearls (Treasure Island (FL): StatPearls Publishing).
Basciani, S., De Luca, G., Dolci, S., Brama, M., Arizzi, M., Mariani, S., et al. (2008). Platelet-derived growth factor receptor beta-subtype regulates proliferation and migration of gonocytes. Endocrinol 149, 6226–6235. doi:10.1210/en.2008-0349
Basciani, S., Mariani, S., Spera, G., and Gnessi, L. (2010). Role of platelet-derived growth factors in the testis. Endocr. Rev. 31, 916–939. doi:10.1210/er.2010-0004
Bunick, D., Kirby, J., Hess, R. A., and Cooke, P. S. (1994). Developmental expression of testis messenger ribonucleic acids in the rat following propylthiouracil-induced neonatal hypothyroidism. Biol. Reprod. 51, 706–713. doi:10.1095/biolreprod51.4.706
Buzzard, J. J., Morrison, J. R., O'Bryan, M. K., Song, Q., and Wreford, N. G. (2000). Developmental expression of thyroid hormone receptors in the rat testis. Biol. Reprod. 62, 664–669. doi:10.1095/biolreprod62.3.664
Canale, D., Agostini, M., Giorgilli, G., Caglieresi, C., Scartabelli, G., Nardini, V., et al. (2001). Thyroid hormone receptors in neonatal, prepubertal, and adult rat testis. J. Androl. 22, 284–288. doi:10.1002/j.1939-4640.2001.tb02182.x
Chazaud, C., Oulad-Abdelghani, M., Bouillet, P., Décimo, D., Chambon, P., and Dollé, P. (1996). AP-2.2, a novel gene related to AP-2, is expressed in the forebrain, limbs and face during mouse embryogenesis. Mech. Dev. 54, 83–94. doi:10.1016/0925-4773(95)00463-7
Chen, J., Ortmeier, S. B., Savinova, O. V., Nareddy, V. B., Beyer, A. J., Wang, D., et al. (2012). Thyroid hormone induces sprouting angiogenesis in adult heart of hypothyroid mice through the PDGF-Akt pathway. J. Cell Mol. Med. 16, 2726–2735. doi:10.1111/j.1582-4934.2012.01593.x
Choudhury, S., Chainy, G. B., and Mishro, M. M. (2003). Experimentally induced hypo- and hyper-thyroidism influence on the antioxidant defence system in adult rat testis. Andrologia 35, 131–140. doi:10.1046/j.1439-0272.2003.00548.x
Cooke, P. S., Zhao, Y. D., and Bunick, D. (1994). Triiodothyronine inhibits proliferation and stimulates differentiation of cultured neonatal Sertoli cells: possible mechanism for increased adult testis weight and sperm production induced by neonatal goitrogen treatment. Biol. Reprod. 51, 1000–1005. doi:10.1095/biolreprod51.5.1000
Culty, M. (2009). Gonocytes, the forgotten cells of the germ cell lineage. Birth Defects Res. C Embryo Today 87, 1–26. doi:10.1002/bdrc.20142
Decrouy, X., Gasc, J. M., Pointis, G., and Segretain, D. (2004). Functional characterization of Cx43 based gap junctions during spermatogenesis. J. Cell Physiol. 200, 146–154. doi:10.1002/jcp.10473
De França, L. R., Hess, R. A., Cooke, P. S., and Russell, L. D. (1995). Neonatal hypothyroidism causes delayed Sertoli cell maturation in rats treated with propylthiouracil: evidence that the Sertoli cell controls testis growth. Anat. Rec. 242, 57–69. doi:10.1002/ar.1092420108
Eildermann, K., Aeckerle, N., Debowski, K., Godmann, M., Christiansen, H., Heistermann, M., et al. (2012). Developmental expression of the pluripotency factor sal-like protein 4 in the monkey, human and mouse testis: restriction to premeiotic germ cells. Cells Tissues Organs 196, 206–220. doi:10.1159/000335031
Fadlalla, M. B., Wei, Q., Fedail, J. S., Mehfooz, A., Mao, D., and Shi, F. (2017). Effects of hyper- and hypothyroidism on the development and proliferation of testicular cells in prepubertal rats. Anim. Sci. J. 88, 1943–1954. doi:10.1111/asj.12883
Faraone-Mennella, M. R., Ferone, A., Marino, L., Cardone, A., Comitato, R., Venditti, P., et al. (2009). Poly (ADP-ribosyl) ation of proteins and germ cell development in hyperthyroid rat testes. Mol. Cell Biochem. 323, 119–129. doi:10.1007/s11010-008-9970-7
Francavilla, S., Cordeschi, G., Properzi, G., Di Cicco, L., Jannini, E. A., Palmero, S., et al. (1991). Effect of thyroid hormone on the pre- and post-natal development of the rat testis. J. Endocrinol. 129, 35–42. doi:10.1677/joe.0.1290035
Franke, F. E., Pauls, K., Rey, R., Marks, A., Bergmann, M., and Steger, K. (2004). Differentiation markers of Sertoli cells and germ cells in fetal and early postnatal human testis. Anat. Embryol. Berl. 209, 169–177. doi:10.1007/s00429-004-0434-x
Gaytan, F., Pinilla, L., Aguilar, R., Lucena, M. C., and Paniagua, R. (1986). Effects of neonatal estrogen administration on rat testis development with particular reference to Sertoli cells. J. Androl. 7, 112–121. doi:10.1002/j.1939-4640.1986.tb00890.x
Gilleron, J., Nebout, M., Scarabelli, L., Senegas-Balas, F., Palmero, S., Segretain, D., et al. (2006). A potential novel mechanism involving connexin 43 gap junction for control of sertoli cell proliferation by thyroid hormones. J. Cell Physiol. 209, 153–161. doi:10.1002/jcp.20716
Green, C. D., Ma, Q., Manske, G. L., Shami, A. N., Zheng, X., Marini, S., et al. (2018). A comprehensive roadmap of murine spermatogenesis defined by single-cell RNA-seq. Dev. Cell 46, 651–667. doi:10.1016/j.devcel.2018.07.025
Griswold, M. D. (2016). Spermatogenesis: the commitment to meiosis. Physiol. Rev. 96, 1–17. PMID: 26537427. doi:10.1152/physrev.00013.2015
Grüters, A., and Krude, H. (2011). Detection and treatment of congenital hypothyroidism. Nat. Rev. Endocrinol. 8, 104–113. doi:10.1038/nrendo.2011.160
Holsberger, D. R., and Cooke, P. S. (2005). Understanding the role of thyroid hormone in Sertoli cell development: a mechanistic hypothesis. Cell Tissue Res. 322, 133–140. doi:10.1007/s00441-005-1082-z
Jannini, E. A., Carosa, E., Rucci, N., Screponi, E., and D'Armiento, M. (1999). Ontogeny and regulation of variant thyroid hormone receptor isoforms in developing rat testis. J. Endocrinol. Invest 22, 843–848. doi:10.1007/BF03343656
Jannini, E. A., Crescenzi, A., Rucci, N., Screponi, E., Carosa, E., de Matteis, A., et al. (2000). Ontogenetic pattern of thyroid hormone receptor expression in the human testis. J. Clin. Endocrinol. Metab. 85, 3453–3457. doi:10.1210/jcem.85.9.6803
Jannini, E. A., Olivieri, M., Francavilla, S., Gulino, A., Ziparo, E., and D'Armiento, M. (1990). Ontogenesis of the nuclear 3,5,3'-triiodothyronine receptor in the rat testis. Endocrinology 126, 2521–2526. doi:10.1210/endo-126-5-2521
Johnsen, S. G. (1970). Testicular biopsy score count--a method for registration of spermatogenesis in human testes: normal values and results in 335 hypogonadal males. Horm 1, 2–25. doi:10.1159/000178170
Koubova, J., Menke, D. B., Zhou, Q., Capel, B., Griswold, M. D., and Page, D. C. (2006). Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc. Natl. Acad. Sci. U. S. A. 103, 2474–2479. doi:10.1073/pnas.0510813103
La Vignera, S., and Vita, R. (2018). Thyroid dysfunction and semen quality. Int. J. Immunopathol. Pharmacol. 32, 2058738418775241. doi:10.1177/2058738418775241
Maciel, L. M., Kimura, E. T., Nogueira, C. R., Mazeto, G. M., Magalhães, P. K., Nascimento, M. L., et al. (2013). Congenital hypothyroidism: recommendations of the thyroid department of the Brazilian society of endocrinology and metabolism. Arq. Bras. Endocrinol. Metabol. 57, 184–192. doi:10.1590/s0004-27302013000300004
Marchlewska, K., Kula, K., Walczak-Jedrzejowska, R., Kula, W., Oszukowska, E., Filipiak, E., et al. (2013). Maturational changes in connexin 43 expression in the seminiferous tubules may depend on thyroid hormone action. Arch. Med. Sci. 9, 139–145. doi:10.5114/aoms.2013.33074
Marchlewska, K., Kula, K., Walczak-Jedrzejowska, R., Oszukowska, E., Orkisz, S., and Slowikowska-Hilczer, J. (2011). Triiodothyronine modulates initiation of spermatogenesis in rats depending on treatment timing and blood level of the hormone. Mol. Cell Endocrinol. 341, 25–34. doi:10.1016/j.mce.2011.04.022
Meroni, S. B., Galardo, M. N., Rindone, G., Gorga, A., Riera, M. F., and Cigorraga, S. B. (2019). Molecular mechanisms and signaling pathways involved in sertoli cell proliferation. Front. Endocrinol. (Lausanne) 10, 224. doi:10.3389/fendo.2019.00224
Omidi, N., Khorgami, M., Tajrishi, F. Z., Seyedhoseinpour, A., and Pasbakhsh, P. (2020). The role of thyroid diseases and their medications in cardiovascular disorders: a review of the literature. Curr. Cardiol. Rev. 16, 103–116. doi:10.2174/1573403X15666191008111238
Pallet, V., Audouin-Chevallier, I., Verret, C., Garcin, H., and Higueret, P. (1994). Retinoic acid differentially modulates triiodothyronine and retinoic acid receptors in rat liver according to thyroid status. Eur. J. Endocrinol. 131, 377–384. doi:10.1530/eje.0.1310377
Palmero, S., de Marchis, M., Gallo, G., and Fugassa, E. (1989). Thyroid hormone affects the development of Sertoli cell function in the rat. J. Endocrinol. 123, 105–111. doi:10.1677/joe.0.1230105
Palmero, S., Prati, M., Bolla, F., and Fugassa, E. (1995). Tri-iodothyronine directly affects rat Sertoli cell proliferation and differentiation. J. Endocrinol. 145, 355–362. doi:10.1677/joe.0.1450355
Pan, Y. W., Tsai, M. C., Yang, Y. J., Chen, M. Y., Chen, S. Y., and Chou, Y. Y. (2019). The relationship between nonalcoholic fatty liver disease and pediatric congenital hypothyroidism patients. Kaohsiung J. Med. Sci. 35, 778–786. doi:10.1002/kjm2.12118
Panno, M. L., Sisci, D., Salerno, M., Lanzino, M., Pezzi, V., Morrone, E. G., et al. (1996). Thyroid hormone modulates androgen and oestrogen receptor content in the Sertoli cells of peripubertal rats. J. Endocrinol. 148, 43–50. doi:10.1677/joe.0.1480043
Picut, C. A., Remick, A. K., de Rijk, E. P., Simons, M. L., Stump, D. G., and Parker, G. A. (2015). Postnatal development of the testis in the rat: morphologic study and correlation of morphology to neuroendocrine parameters. Toxicol. Pathol. 43, 326–342. doi:10.1177/0192623314547279
Rodrigues, M. S., Tovo-Neto, A., Rosa, I. F., Doretto, L. B., Fallah, H. P., Habibi, H. R., et al. (2022). Thyroid hormones deficiency impairs male germ cell development: a cross talk between hypothalamic-pituitary-thyroid, and-gonadal axes in zebrafish. Front. Cell Dev. Biol. 10, 865948. doi:10.3389/fcell.2022.865948
Romeo, R., Castorina, S., and Marcello, M. F. (1995). Intermediate filaments of human Sertoli cells in germinal alterations. Ital. J. Anat. Embryol. 100, 75–81.
Russell, L. D., and Peterson, R. N. (1985). Sertoli cell junctions: morphological and functional correlates. Int. Rev. Cytol. 94, 177–211. doi:10.1016/s0074-7696(08)60397-6
Sahoo, D. K., Roy, A., Bhanja, S., and Chainy, G. B. (2008). Hypothyroidism impairs antioxidant defence system and testicular physiology during development and maturation. Gen. Comp. Endocrinol. 156, 63–70. doi:10.1016/j.ygcen.2007.11.007
Sakai, Y., Yamashina, S., and Furudate, S. (2004). Developmental delay and unstable state of the testes in the rdw rat with congenital hypothyroidism. Dev. Growth Differ. 46, 327–334. doi:10.1111/j.1440-169x.2004.00748.x
Sarkar, D., and Singh, S. K. (2017). Effect of neonatal hypothyroidism on prepubertal mouse testis in relation to thyroid hormone receptor alpha 1 (THRα1). Gen. Comp. Endocrinol. 251, 109–120. doi:10.1016/j.ygcen.2016.08.001
Show, M. D., Anway, M. D., Folmer, J. S., and Zirkin, B. R. (2003). Reduced intratesticular testosterone concentration alters the polymerization state of the Sertoli cell intermediate filament cytoskeleton by degradation of vimentin. Endocrinol 144, 5530–5536. doi:10.1210/en.2003-0735
Simorangkir, D. R., Wreford, N. G., and De Kretser, D. M. (1997). Impaired germ cell development in the testes of immature rats with neonatal hypothyroidism. J. Androl. 18, 186–193. doi:10.1002/j.1939-4640.1997.tb01899.x
Staggenborg, S., Koch, R., Rode, K., Hüneke, H., Tiedje, L., Wirth, G., et al. (2022). Connexin43 represents an important regulator for Sertoli cell morphology, Sertoli cell nuclear ultrastructure, and Sertoli cell maturation. Sci. Rep. 12, 12898. doi:10.1038/s41598-022-16919-4
Tanemura, K., Kurohmaru, M., Kuramoto, K., Matsumoto, M., and Hayashi, Y. (1994). Age-related changes in cytoskeletal components of the BDF1 mouse Sertoli cell. Tissue Cell 26, 447–455. doi:10.1016/0040-8166(94)90028-0
Umezu, M., Kagabu, S., Jiang, J. Y., Niimura, S., and Sato, E. (2004). Developmental hormonal profiles in rdw rats with congenital hypothyroidism accompanying increased testicular size and infertility in adulthood. J. Reprod. Dev. 50, 675–684. doi:10.1262/jrd.50.675
van Trotsenburg, P., Stoupa, A., Léger, J., Rohrer, T., Peters, C., Fugazzola, L., et al. (2021). Congenital hypothyroidism: a 2020-2021 consensus guidelines update-an endo-European reference network initiative endorsed by the European society for pediatric endocrinology and the European society for endocrinology. Thyroid 31, 387–419. doi:10.1089/thy.2020.0333
Vigueras-Villaseñor, R. M., Molina-Ortiz, D., Reyes-Torres, G., del Angel, D. S., Moreno-Mendoza, N. A., Cruz, M. E., et al. (2009). Effect of allopurinol on damage caused by free radicals to cryptorchid testes. Acta histochem. 111, 127–137. doi:10.1016/j.acthis.2008.05.004
Vigueras-Villaseñor, R. M., Ojeda, I., Gutierrez-Pérez, O., Chavez-Saldaña, M., Cuevas, O., Maria, D. S., et al. (2011). Protective effect of α-tocopherol on damage to rat testes by experimental cryptorchidism. Int. J. Exp. Pathol. 92, 131–139. doi:10.1111/j.1365-2613.2010.00757.x
Walczak-Jedrzejowska, R., Slowikowska-Hilczer, J., Marchlewsk, K., Oszukowska, E., and Kula, K. (2007). During seminiferous tubule maturation testosterone and synergistic action of FSH with estradiol support germ cell survival while estradiol alone has pro-apoptotic effect. Folia Histochem Cytobiol. 45, S59–S64.
Yildirim, A. B., Ozdamar, S., and Kaymak, E. (2017). The effects of maternal hypothyroidism on the immunoreactivity of cytochrome p450 aromatase in the postnatal rat testicles. Acta Endocrinol. (Buchar) 13, 7–16. doi:10.4183/aeb.2017.7
Yousefichaijan, P., Dorreh, F., Rafeie, M., Sharafkhah, M., Safi, F., Amiri, M., et al. (2015). Congenital anomalies of kidney and upper urinary tract in children with congenital hypothyroidism; a case-control study. J. Ren. Inj. Prev. 4, 120–126. doi:10.12861/jrip.2015.26
Zamoner, A., Barreto, K. P., Filho, D. W., Sell, F., Woehl, V. M., Guma, F. C., et al. (2008). Propylthiouracil-induced congenital hypothyroidism upregulates vimentin phosphorylation and depletes antioxidant defenses in immature rat testis. J. Mol. Endocrinol. 40, 125–135. doi:10.1677/JME-07-0089
Zanatta, A. P., Zanatta, L., Gonçalves, R., Zamoner, A., and Silva, F. R. (2013). Rapid responses to reverse TRapid responses to reverse T₃ hormone in immature rat Sertoli cells: calcium uptake and exocytosis mediated by integrinhormone in immature rat Sertoli cells: calcium uptake and exocytosis mediated by integrin. PLoS One. 8, e77176. doi:10.1371/journal.pone.0077176
Keywords: congenital hypothyroidism, Sertoli cells, gonocyte, meiosis, methimazole
Citation: Gómez-Zúñiga A, Landero-Huerta DA, Rojas-Castañeda JC, Sánchez-Huerta K, Contreras-García IJ, Reynoso-Robles R, Arteaga-Silva M and Vigueras-Villaseñor RM (2024) Methimazole-induced congenital hypothyroidism affects gonocytes differentiation and arrests meiosis: role of Sertoli cells. Front. Cell Dev. Biol. 12:1493872. doi: 10.3389/fcell.2024.1493872
Received: 09 September 2024; Accepted: 05 November 2024;
Published: 21 November 2024.
Edited by:
Maria Eugenia Teves, Virginia Commonwealth University, United StatesReviewed by:
Atsushi P. Kimura, Hokkaido University, JapanCopyright © 2024 Gómez-Zúñiga, Landero-Huerta, Rojas-Castañeda, Sánchez-Huerta, Contreras-García, Reynoso-Robles, Arteaga-Silva and Vigueras-Villaseñor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosa María Vigueras-Villaseñor, cm12aWd1ZXJhc0B5YWhvby5jb20ubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.