 Huilan Su
Huilan Su Liqun Huang2†
Liqun Huang2† Jianjun Zhou
Jianjun Zhou Guosheng Yang
Guosheng Yang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol. , 08 August 2024
Sec. Stem Cell Research
Volume 12 - 2024 | https://doi.org/10.3389/fcell.2024.1410102
This article is part of the Research Topic Advances in Translational and Applied Stem Cell Biology View all 6 articles
Prostate cancer (PCa) is the most common malignancy among men worldwide. Through androgen receptor signaling inhibitor (ARSI) treatment, patients eventually succumb to castration-resistant prostate cancer (CRPC). For this, the prostate cancer stem cells (PCSCs), as a minor population of tumor cells that can promote tumor relapse, ARSI resistance, and disease progression, are gaining attention. Therefore, specific therapy targeting PCSCs has momentum. This study reviewed the identification and characterization of PCSCs and PCSC-based putative biomarkers and summarized their mechanisms of action. We further discussed clinical trials of novel therapeutic interventions focused on PCSC-related pathways, the PCSC microenvironment, cutting-edge miRNA therapy, and immunotherapy approaches from a mechanistic standpoint. This review provides updated insights into PCSC plasticity, identifying new PCSC biomarkers and optimized treatments for patients with advanced PCa.
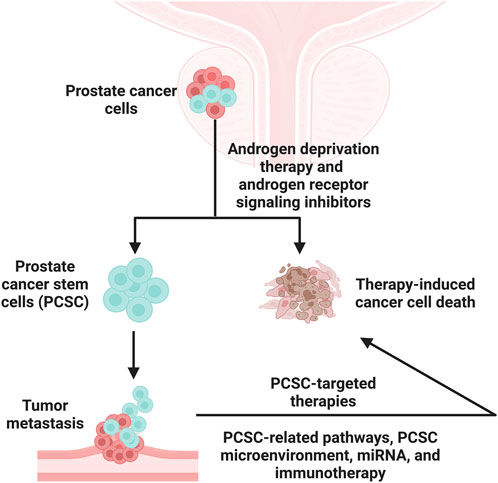
Prostate cancer (PCa) is the most common malignancy among men, accounting for 29% of estimated new cancer cases (2023) and a substantial burden to the public health system (Porcacchia et al., 2022; Siegel et al., 2023). Worldwide, PCa ranks second in terms of mortality (400,000 deaths annually), and this rate is expected to be 2-fold higher by 2040 (Sandhu et al., 2021). Even in an East Asian country like China, where the incidence rate of prostate cancer is generally low, the number of PCa patients is continually on the rise, making it a front-runner of urinary tumor-related disease (Qiu et al., 2021). As the disease progresses, PCa might undergo a transition phase from hormone-sensitive prostate cancer (HSPC) to castration-resistant prostate cancer (CRPC) and from localized disease to metastatic castration-resistant prostate cancer (mCRPC) (Terrisse et al., 2022; Yehya et al., 2022). When diagnosed with an advanced stage, men would have a considerably diminished 5-year overall survival (OS) (30%), making advanced PCa a threat to patients (Gao et al., 2020).
The onset and progression of PCa are driven by androgen receptor (AR) signaling (Zheng et al., 2022). However, despite being initially effective and durable for localized and advanced prostate tumors, androgen deprivation therapy (ADT) and AR-directed strategies (e.g., enzalutamide) will move to a stage characterized by the inevitable emergence of resistance (Storck et al., 2022; Zheng et al., 2022; Zhu et al., 2022). At this point, the heterogeneous progenies containing enriched PCSCs from advanced PCa become the predominant population and are almost all negative in prostate-specific antigen (PSA) and AR levels (Zhang et al., 2015).
How does the unique mechanism contribute to treatment resistance? The inherent properties of prostate cancer stem cells (PCSC) may provide new insights into this puzzle. We identified that AR−/lo cells were linked to increased PCSC populations, which are proven to promote tumor relapse and disease progression (Tang, 2022). Accordingly, understanding the molecular features of the PCSCs that drive the phenotypic transition from ADT-sensitivity to CRPC could help provide more meaningful results for ongoing research and designing more appropriate treatment strategies in the clinic.
A prerequisite to tracking PCSCs is enumerating normal human prostate (NHP) cell lineages in full detail. Given their histological appearance and specific antigen expression, the epithelial cells of NHP are composed of basal and luminal layers and scarce neuroendocrine (NE) cells (Tang, 2022). The basal layer contains a small population of multipotent stem cells (SCs) (<5%), whereas the number in the luminal layer is less than 1% (Abate-Shen and Shen, 2000). Investigating the unique properties of layers would be meaningful for understanding and laying a foundation for novel therapeutics targeting PCSCs.
Owing to the relative undifferentiation and survival priority of AR ablation, the basal population of epithelial cells tends to possess characterizations of PCSCs (Goldstein et al., 2008). In addition, basal cells preferentially express cell adhesion/cytoskeleton and extracellular matrix remodeling-related genes (Zhang D. et al., 2016). Lineage-tracing studies revealed that PTEN deletion and deacetylated Klf5 contributed to rapid differentiation of luminal progeny by controlling basal progenitor cell fate (Zhang J. et al., 2018; Zhang et al., 2020). Interestingly, acute prostatitis mediated the differentiation of basal cells into luminal cells via a specific program in the microenvironment (Toivanen et al., 2016). Meanwhile, Kwon et al. (2014) described a mouse model where tissue repair in the prostate epithelium was regulated partly by basal-to-luminal differentiation. In addition, basal cells functionally revealing neurogenic properties brought out the underlying hypothesis of cells-or-origin for neuroendocrine prostate cancer (NEPC) (Verma et al., 2023). Of clinical relevance, Zhang et al. revealed the contribution of basal cells to promote castration-resistant and metastatic PCa (Zhang D. et al., 2016). Multiple crucial molecules such as CK14, B-cell lymphoma-2 (Bcl-2), and human telomerase reverse transcriptase (hTERT) have been well documented to preferentially localize in the basal layer (Wang et al., 2009; Banerjee et al., 2023). Thus, it is thought that basal cells share SC characteristics. On the other hand, a growing body of evidence has increasingly linked SC-like cells to the basal cells due to their co-expressed markers. In vitro and in vivo prostate assays have exhibited SC-enhanced global transcription and rRNA transcription activity in the basal layer (Zhang D. et al., 2016). Many other protein markers associated with CSC phenotype, such as spinocerebellar ataxia 1 (Sca-1), CD133, CD44, CD117, CD49f, α2 integrin, C-X-C motif chemokine receptor type 4 (CXCR4), epithelial cell adhesion molecule (EpCaM), CD54, and sex-determining region Y-box 2 (SOX2) et al., have also been detected in basal cells (details are shown in Table1) (Goldstein et al., 2008; Wang et al., 2009; Hoogland et al., 2014; Galoczova et al., 2021; Verma et al., 2023). Tumor samples derived from Sca-1, CD133, CD44, and CD117-positive cells basal cells possessed the self-renewal ability and reconstituted the prostatic ducts (Wang et al., 2009). However, Hoogland et al. (2014) raised doubt about the reliability of CD117, CD133, and OCT3/4 to label PCSC characteristics because these markers were not detected in clinical tissue. For specific treatment, ADT-treated PCa tended to develop into NEPC, accompanied by high levels of stem- (SOX2) and basal cell markers (KRT5; TP63) (Verma et al., 2023). This could provide a novel platform for screening drug candidates in a clinical situation via monitoring the ADT-resistant stem cell-like population.
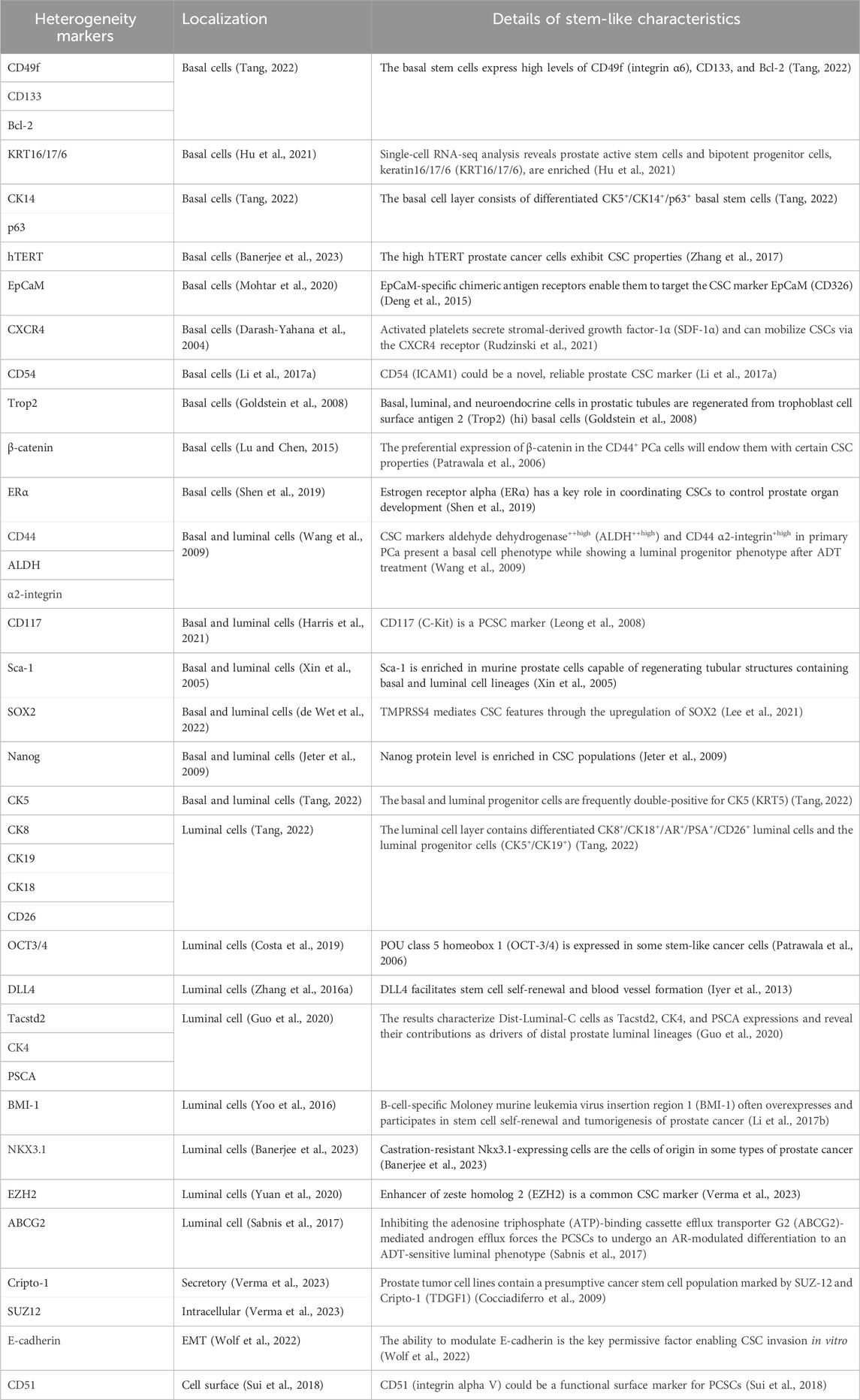
Table 1. List of putative biomarkers for CSCs.
The findings regarding cells-of-origin for NEPC are intriguing. Ci et al., 2020. established a patient-derived xenograft (PDX) model of adenocarcinoma (LTL331)-to-NEPC (LTL331R) transdifferentiation to support a basal progenitor cell model (Ci et al., 2020). Dong et al. (2020) employed the single-cell RNA sequencing detecting transcriptomes of six CRPC needle biopsies, which provided direct evidence of the cellular states underlying luminal–neuroendocrine transdifferentiation. Notably, this transdifferentiation has never been revealed in normal prostate development. In addition, basal cell marker p63 was considered indispensable for prostate development (Goldstein et al., 2008). The next year, explants from p63 null mice could form prostate tissue in the absence of basal cells, supporting the necessity of luminal progenitor cells (Wang et al., 2009). Furthermore, a growing body of evidence indicated that cancers could be driven by tumorigenic luminal cells without initiating basal cells, and murine lineage-tracing experiments also presented luminal-to-basal differentiation (Karthaus et al., 2014).
The controversies are worth pondering. If basal stem cells could represent a cell type of origin, one must wonder why basal or squamous cell carcinomas account for a small proportion of PCa phenotypes (Ali and Epstein, 2007). Given culture condition scarcity, prostatic gland architecture could not be realistically reconstituted. It has remained challenging to determine whether these transitions apply to humans in the absence of a 3D culture system. In terms of organoids of luminal and basal cells, Karthaus et al. (2014) proved that luminal-derived organoids more closely resemble prostate glands. Tang’s research also confirmed luminal progenitor cell (LP) as a preferred cell of origin for PCa (Tang, 2022). Furthermore, Gao’s group brought out a novel insight into tracking of cells-or-origin for mouse prostate. Briefly, they characterized Dist-Luminal-C cells as tumor-associated calcium signal transducer 2 (Tacstd2), CK4, and prostate stem cell antigen (PSCA) expression and revealed its contribution as the driver of distal prostate luminal lineages (Guo et al., 2020). In addition, not only basal compartment but also luminal markers such as NKX3.1, CK18, CK8, CD26, OCT3/4, and delta-like ligand 4 (DLL4) et al. have been demonstrated to be co-expressed with the CSCs (details are shown in Table1) (Wang et al., 2009; Zhang D. et al., 2016; Park et al., 2016; Costa et al., 2019). Most notably, basal stem-like cells have been suggested to be the cell of origin in primary prostatic tumors, while only stem-like cells with luminal phenotype reinitiated CRPC deterioration after androgen ablation (Wang et al., 2009; Germann et al., 2012). For example, CSC markers (ALDH++high CD44 α2-integrin+high) in primary PCa presented a basal cell phenotype while showing a luminal progenitor phenotype after ADT treatment. One explanation is that primary treatment-induced lower AR level results in an ARlow stem-like luminal cell (Zhang B. et al., 2016). Meanwhile, luminal progenitor cell plays a significant role in treatment resistance and poor outcomes. Over the course of CRPC progression, significant increases in PSA−/lo PCa cells with LP characteristics and human LP markers (i.e., CD38low and ALDHhi CD44 α2β1) have been demonstrated (Zhang D. et al., 2018; Verma et al., 2023; Gorodetska et al., 2024). We propose that low-grade prostate tumors are driven by basal cells, but tumorigenic luminal and LP cells rapidly expand in CRPC.
Several studies have shown that PCSCs could originate from cancerous cells (i.e., inflammatory cells and stromal cells) (Banerjee et al., 2023). This viewpoint could explain why PCSCs have the renewal capacity to achieve malignant transformation where the differentiated cells present accumulative mutations (Banerjee et al., 2023). Herein, inflammation-induced alterations not only cause epithelial lineage differentiation but also promote oncogenic signaling to induce tumor initiation. Many studies have shown that the stem phenotype of advanced PCa was intimately associated with epithelial–mesenchymal transition (EMT), which was derived from stromal cells in the tumor microenvironment (Chen et al., 2020). In addition, observations suggested enhancer of zeste homolog (EZH2)- and cancer-associated fibroblasts (CAF)-mediated EMT resulted in the enrichment of CSC-like properties (Giannoni et al., 2010; Yamada and Beltran, 2021). Additionally, various signaling pathways involved in the progression and therapy resistance, such as Notch, Wingless (Wnt)/β-Catenin, Hedgehog, Hippo, Ras/mitogen-activated protein kinase (MAPK), Janus kinase (JAK)/signal transducer and activator of transcription (STAT), phosphoinositide 3 kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR), epidermal growth factor receptor (EGFR), and hypoxia-inducible factor (HIF), have been reported to drive CSC emergence (details are shown in Table 2) (Meisel et al., 2020; Wu et al., 2020; Yang et al., 2020; Ramesh et al., 2023; Verma et al., 2023).
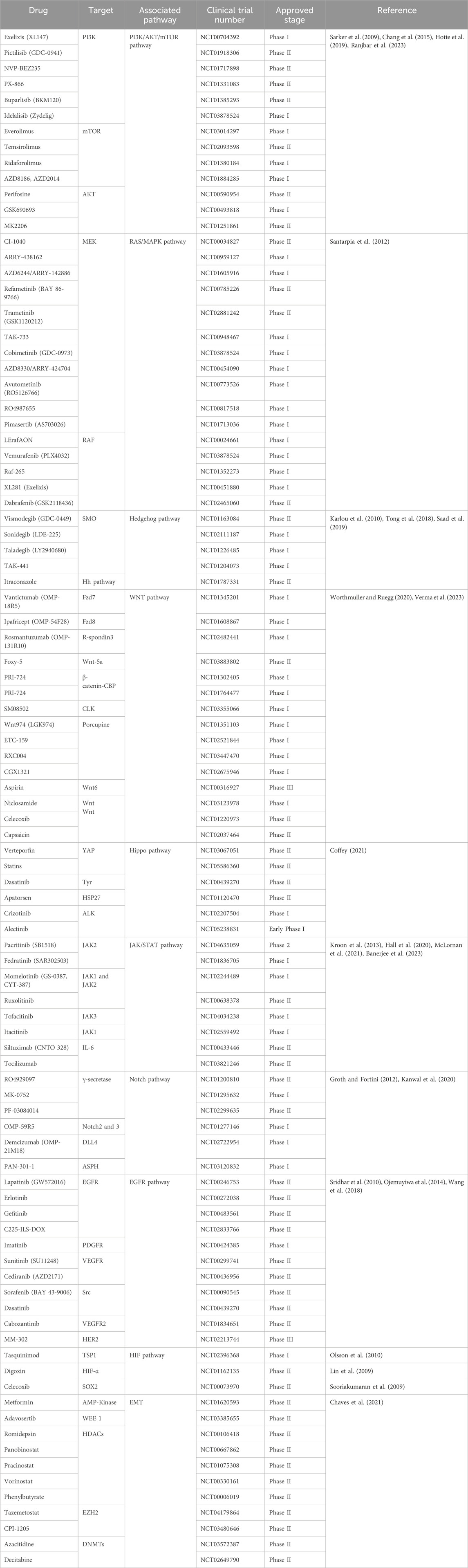
Table 2. List of drugs for CSC targeted therapy under clinical trials.
Isolating cells with tumor-initiating and stem-like properties like PCSCs presents undeniable challenges. Utilizing specific markers expressed by PCSCs can offer solutions. Techniques such as fluorescence-activated cell sorting (FACS) and magnetic-activated cell sorting (MACS) can effectively isolate and purify PCSCs based on known surface markers (Banerjee et al., 2023). For instance, selecting for CD44+α2β1-/lo cells has been proposed as a representation of PCSCs (Patrawala et al., 2007). Additionally, nuclear staining dyes like Hoechst 33,342 and Rhodamine 123 can aid in isolating PCSCs. It is reported that strategically repeated chemotherapy and radiotherapy could maintain cell populations of therapy-resistant phenotypes and provide favorable conditions for PCSC proliferation. The sphere formation assay has been suggested as another option. Spheres derived from PCSCs can be further characterized (Verma et al., 2023). Thus, PCSCs can be isolated either by selecting marker-based populations or by inducing cell de-differentiation.
Current treatments for PCa, such as ADT, chemotherapy, and radiation, are designed to eliminate large numbers of conventional tumor cells but do not appear to be effective against drug-resistant PCSCs. Therefore, therapies targeting PCSCs are emerging as promising approaches. These approaches focus on PCSC-related pathways, the PCSC microenvironment, miRNA, and immunotherapy. In this context, several inhibitors have been reported in clinical trials or are undergoing clinical trial evaluation (details are shown in Table 2).
PI3K, frequently activated in PCa, stimulates mTOR through activated AKT. Recent discoveries indicated that the intricate crosstalk within the PI3K/AKT/mTOR pathway could facilitate tumor formation, enhance CSC properties, and increase therapeutic resistance (Verma et al., 2023). To date, several inhibitors targeting the PI3K/AKT/mTOR pathway have been evaluated in phase I or II clinical trials (details are shown in Table 2). These inhibitors could also be used with chemo- or radiotherapy to restore the sensitivity of CRPC patients to traditional treatments (Bitting and Armstrong, 2013). In PTEN-loss models, the inhibition of AR could activate the PI3K/AKT pathway and vice-versa. To address the problem, a PI3K inhibitor (such as PX-866) was designed to target CRPC patients, which had a beneficial effect and overcame resistance (Hotte et al., 2019). However, the dual PI3K and mTOR inhibition might cause unpredictable toxicity in patients with mCRPC (Wei et al., 2017).
MAPK signaling is reported to be responsible for stem characteristics in PCSCs, and phosphorylation events play critical parts in tumorigenesis (Santarpia et al., 2012). Hindering MAPK via targeted inhibitors has been an applicable model for cancer therapeutics. Abnormal activation of the RAS-RAF-MEK-ERK-MAPK (RAS-MAPK) pathway promotes CSC self-propelling and poses a second hit to an alteration of the PTEN/PI3K/AKT axis (Santarpia et al., 2012). MAPK kinase inhibitor PD098059 restored the growth inhibitory role of TGF-β1 in PCa, which carried an oncogenic mutation in RAS (Park et al., 2000). Although PD098059 and PD325901 have been demonstrated to be effective in mouse studies, they have not been targeted for clinical development (Mukhopadhyay et al., 2007; Mulholland et al., 2012). PD184352 (CI-1040) has been evaluated in phase I clinical trials but not yet verified in phase II trials (Papatsoris et al., 2007). In addition, drugs that obstruct the RAS/MAPK pathway might exhibit widespread mechanism-induced toxicities.
Emerging studies have demonstrated that the abnormal involvement of Hedgehog signaling was accountable for PCSC maintenance. Recently, preclinical studies showed that PCSCs were subjected to Hedgehog-related inhibition (Banerjee et al., 2023). One such Hedgehog receptor smoothened (SMO) inhibitor is GDC-0449, which promotes PCSC apoptosis via GLI-dependent regulation (Tong et al., 2018). A randomized phase I/II trial study explored antihormone therapy together with GDC-0449 to see how well they work in advanced PCa patients, and the results were highly anticipated (NCT01163084). Sonidegib, an SMO inhibitor, underwent a phase I clinical trial in patients with high-risk localized PCa and caused a 2-fold reduction in GLI1 levels (Tong et al., 2018). Other inhibitors of GLI1, such as IPI-269609, GANT61, GANT58, zerumbone, physalin F and physalin B, and SMO inhibitor CUR61414, have not yet been tested in clinical trials (Karlou et al., 2010). Identifying the stages of PCa may provide the most clinical benefit.
The Kjd Wnt/β-catenin signaling pathway is one of the vital mechanisms responsible for PCa self-renewal ability, and dysregulation of Wnt signaling increases the proportion of PCSCs (Karlou et al., 2010). An in vitro study suggested that capsaicin could be a potential chemotherapeutic drug for CRPC via blocking the Wnt/β-catenin pathway (Ramesh et al., 2023). Accordingly, a phase II trial was designed to determine the chemopreventive properties of capsaicin in PCa patients enrolled in the active surveillance program or patients scheduled to undergo radical prostatectomy (NCT02037464). In addition, agents like aspirin, which has been approved by the FDA, are applied in the clinics (Verma et al., 2023). Consequently, Wnt-related research has been a significant field for the development and application of targeted drugs. The inhibitors targeting the Wnt/β-catenin pathway are classified into non-steroidal anti-inflammatory drugs (ibuprofen and aspirin) and CBP/β antagonists (ICG-001 and NSC668036) (Verma et al., 2023). Meanwhile, Worthmuller and Ruegg (2020) divide Wnt-related agents into ligand/receptor level (vantictumab, ipafricept, etc.), transcriptional level (CWP232291, PRI-724, etc.), and Wnt secretion (WNT974, ETC-15, etc).
The Hippo pathway and its core downstream effectors, Yes-associated protein (YAP) and paralog, a transcriptional coactivator with the PDZ-binding motif (TAZ), are crucial for tissue regeneration through the regulation of stem cells (Messina et al., 2023). Inhibition of Hippo remains challenging owing to its complicated regulation and crossing with other pathways. Although the YAP/TAZ targeted therapeutic drug, verteporfin, has been approved by the FDA, its future use for cancer treatment appears to be multimodal, relying on the cellular background (Coffey, 2021). In addition, several FAK inhibitors have been measured in clinical trials with prospective results in PCa. One is apatorsen (OGX427), which could induce tumor regression in preclinical models of metastatic CRPC and has shown encouraging preliminary results in phase II clinical trials (Coffey, 2021).
Gene expression profiling of CD44+/α2β1hi/CD133+ primary cancer cells reveals a significant over-representation of the JAK-STAT signaling pathway, indicating aberrant alterations of this pathway in CSCs could accelerate the tumor load (Kroon et al., 2013). Wang et al. (2024) demonstrated that blocking STAT3 via berbamine resulted in downregulation of CSC level and increased drug sensitivity to cabazitaxel. However, there are not yet any clinical trials for berbamine. The blockade of activated STAT3 by another anti-IL-6 antibody, tocilizumab, suppressed the activity of the TAM-stimulated CD44+ cells in high-grade diseases (Wan et al., 2014). A phase I trial was aimed to evaluate the safety and efficacy of CC-1 (a dual mode of anticancer action) with prophylactic IL-6R blockade using tocilizumab in CRPC patients after failure of third-line therapy (NCT04104607). The research would help better define the action of CC-1 and identify biomarkers for further clinical development.
The Notch pathway, which regulates cell fate determination, metastasis, and chemoresistance, has been found to be dysregulated in PCa (Banerjee et al., 2023). One approach involves the exploration of antibodies to obstruct specific Notch receptors, their activating ligands, or other targets of the Notch signaling in tumors (Han et al., 2021). Chemotherapy combined with Notch1 inhibitors is proved to reduce the chemotherapy-enriched CSC population in a complementary manner (Banerjee et al., 2023). Recently, Cui et al. (2015) suggested that Notch blocking via a γ-secretase inhibitor (GSI) named PF-03084014 could slow the growth of tumor cells and reinforce the anti-metastatic effect of docetaxel in PCa in vivo and in vitro. In contrast, PF-03084014 failed to produce a clinical benefit to CRPC patients owing to its systemic toxicity and off-target effects (Banerjee et al., 2023). Another small-molecule inhibitor of aspartate β-hydroxylase (ASPH), PAN-301-1 vaccine against ASPH has been tested in a phase I clinical trial in PCa patients, indicating that ASPH is a promising target (Kanwal et al., 2020).
Rybak et al. (2013) have presented evidence that EGFR signaling promoted maintenance of PCSC-like characteristics, in part by stimulating the MEK-ERK pathway. Inhibition of ERK activation by U0126 treatment and ERK1/ERK2 knockdown could account for a rapid reduction in PCSC propagation (Rybak et al., 2013). Clinically, modulation of the EGFR pathway is correlated with therapeutic efficiency. Recently, there has been a trend in evaluating tyrosine kinase inhibitors (TKIs) that impede angiogenic growth factor targets. A phase II trial tested sorafenib, an oral inhibitor of EGFR, in metastatic CRPC patients. This agent works by blocking radiological progression and, in part, promoting the regression of bone metastases (Antonarakis et al., 2010). Erlotinib is also a selective TKI of EGFR and has moderate activity in chemotherapy-naïve CRPC in combination with chemotherapy (Nabhan et al., 2009). In addition, PCa has upregulation of platelet-derived growth factor receptor (PDGFR), cooperating with the PI3K/AKT pathway. However, the antitumor effect of PDGFR inhibitor imatinib has been disappointing (Antonarakis et al., 2010).
HIF signaling is activated in PCa in response to hypoxic conditions within the tumor microenvironment. O’Reilly et al. (2019) demonstrated that HIF-2α interacted with SOX2 under long-term hypoxia, promoting stem cell renewal and metastasis of PCSCs. Taken together, these identify HIF and associated pathways as novel cancer drug targets, as well as inhibitors of the hypoxia-response pathway, that are being developed. A phase II clinical trial using oral tasquinimod exhibited moderate activity against mCRPC via upregulation of TSP1, accounting for the downregulation of HIF-1α (Olsson et al., 2010). In addition, camptothecin (CPT), a potent inhibitor of HIF-1α, failed to produce a clinical benefit owing to significant toxicity. Schmidt et al. (2020) designed a nanoparticle–drug conjugate (NDC) of CPT named NLG207 to facilitate drug delivery to tumors. Work on this is ongoing at the National Cancer Institute.
Tumor cells undergo EMT, wherein they lose their epithelial surface markers, most notably E-cadherin, and obtain mesenchymal markers, including vimentin and N-cadherin (Banerjee et al., 2023). Drivers (such as Snail, Twist, and STAT3) and abundant signaling pathways are activated in EMT (Chaves et al., 2021). Given their vital roles in the EMT process, treatments aimed at suppressing specific regulations could provide an approach to achieve the antineoplastic effect. It is already confirmed that miRNAs affected the proportion of PCSCs indirectly via the EMT process. Zhang et al. proved that metformin prevented EMT via microRNA-30a-modulated SOX4 expression (Zhang et al., 2014). However, a phase II trial named “castration compared to castration plus metformin as first-line treatment for patients with advanced PCa” yielded no clinical benefit of adding metformin (NCT01620593). In addition, the abnormality of miR-205 could impede CAF-mediated EMT in vitro and in vivo (Ramesh et al., 2023).
Anticancer strategies have been developed for CAF, varying from metronomic chemotherapy to immune-based therapies. For instance, a GPR77-neutralizing antibody is demonstrated to be valid for restoring tumor sensitivity to chemotherapy in a PDX model (Fiori et al., 2019). Moreover, tazemetostat (EZH2 inhibitor) hitting the PRC2-mediated EMT is designed to determine the recommended dose of tazemetostat in combination with either enzalutamide or abiraterone/prednisone. This approach is being evaluated in a phase II clinical trial enrolling advanced PCa patients (NCT04179864). Presently, efforts to develop therapeutic agents targeting EMT are in progress, and promising results are within reach.
Some miRNAs that are related to good prognosis have been downregulated in CRPC patients. miR-34a, miR-708, miR-143, and miR-145 are negative regulators of CD44 in PCSCs and thus have the potential to serve as therapeutic drugs for advanced PCa patients (Ramesh et al., 2023). In addition, it has been suggested that overexpression of miR-let-7c, miR-101-3p, and miR-138-5p could block the stemness of PCSCs by suppressing EZH2 (Kong et al., 2012; Rizzo, 2021; Ramesh et al., 2023). Subsequently, BR-DIM (metabolite 3,3′-diindolylmethane) is applied to reduce PCSC percentages through EZH2 downregulation (Kong et al., 2012). Mechanistically, miR-7, miR-100, miR-143/miR-145, miR-218, miR-199a-3p, miR-141, and miR-320 suppress PCSCs by targeting the KLF4/PI3K/AKT/p21 pathways, oncogene argonaute 2 (AGO2), OCT4, GLI1, EGFR, actin related protein 2/3 complex subunit 5 (ARPC5), and Wnt/β-catenin, respectively (Rizzo, 2021; Ramesh et al., 2023). For chemotherapy resistance, the expression of miR-125a-3p, miR-34a-5p, miR-204, miR-205, and miR-3 could hamper the enrichment of stem cells and strengthen docetaxel sensitivity in PCa samples, making them ideal therapeutic targets (Rizzo, 2021). In particular, miR-205 also increases radiation sensitivity (El Bezawy et al., 2019). In summary, new therapeutic approaches based on miRNAs might be a good prospect.
Recently, increasing numbers of clinical trials have addressed immunotherapy incorporating vaccine-based therapies, immune checkpoint inhibitors (ICIs), and chimeric antigen receptor (CAR)-modified T-cell therapy, which targets CSC-associated tumor antigens. These products are emerging as new therapeutic approaches for advanced PCa patients (Bansal et al., 2021).
ICIs present antitumor activities by targeting the dysfunctional immune system, where a T-cell antitumor response is generated (Lentz et al., 2021). Ipilimumab is a humanized anti-CTLA-4 antibody that is expressed on the surface of T lymphocytes (Chen, 2004). Its use to treat PCa is investigational. An early phase I clinical trial is aimed at studying the impact of ipilimumab on the immune system of patients receiving hormone therapy, but subsequent results have not yet been presented (NCT02113657). Examples of other immune checkpoint protein PD-1 inhibitors are nivolumab and pembrolizumab, which restore T cells’ ability to eradicate cancer cells (Bansal et al., 2021). A recent update on a phase II clinical trial confirmed the antitumor activity of pembrolizumab with an acceptable safety and encouraging OS evaluation (NCT02787005) (Antonarakis et al., 2020).
Anti-PD-L1 immunotherapies, such as avelumab and atezolizumab, are also being studied (Bansal et al., 2021). In 2021, an ongoing phase II clinical trial of avelumab was designed to evaluate its effects against PICK-NEPC (NCT03179410). Evaluations of monotherapy and the strategies cooperating ICIs with chemotherapy, radiation, PARP inhibitors, adenosine receptor antagonists, IL-2 agonists, and CD11b agonists are in progress (Bansal et al., 2021). For instance, an investigational immunotherapy of nivolumab in combination with rucaparib, docetaxel, or enzalutamide in mCRPC patients is ongoing (NCT03338790). Fizazi et al. (2022) reported results from cohorts A1 and A2 of CheckMate 9KD that nivolumab plus rucaparib were active in HRD-positive postchemotherapy or chemotherapy-naïve mCRPC groups. Notably, a further step is needed to reveal whether nivolumab supplementary incrementally improves OS versus rucaparib alone (Fizazi et al., 2022). Additionally, an immunosuppressive TME and impaired cellular immunity may impede ICI application in advanced PCa (Bansal et al., 2021).
A vaccine based on tumor-associated antigen (TAA) could activate a particular immune response to cancer cells. PCa could express substantial TAA involving PSA, prostate-specific membrane antigen (PSMA), prostatic acid phosphatase (PAP), and PSCA (Bansal et al., 2021). To target these antigens, different forms of PCa vaccines have been developed, such as cellular vaccines, viral vector-based vaccines, polypeptide vaccines, nucleic acid vaccines, and mRNA-based vaccines (Wang et al., 2023). Sipuleucel-T, an FDA-approved autologous cell vaccine, is designed to induce a T-cell-mediated immune response to recombinant PAP (Cha et al., 2020). Currently, related clinical trials have been completed. Phase III (NCT00065442, NCT00005947, and NCT01133704) suggested that sipuleucel-T treatment induced a 3-fold increase in activated T cells from prostatectomy specimens (Ju et al., 2022). It is worth mentioning that the sipuleucel-T treatment can help patients stay where they are rather than fully recovering works to block further deterioration of advanced PCa tumors, not subside.
PROSTVAC has undergone tests in numerous clinical trials. In a phase II clinical trial (TBC-PRO-002), PROSTVAC was associated with a longer median survival time of 9.9 months in men with mCRPC (Kantoff et al., 2010). Conversely, in low- or intermediate-risk PCa, no differences in postvaccination peripheral T-cell responses were observed (NCT02326805) (Parsons et al., 2023). For cellular vaccines, Wang et al. developed an immunogenic peptide-sensitized dendritic cell (DC)-cytokine-induced killer cell (CIK)-based cell, which manifested an antitumor effect against PCa xenografts derived from the PCSC-enriched prostatospheroids. This therapeutic platform is expected to apply to immunotherapy (Wang et al., 2020).
PCVAC/PCa is another cellular cancer vaccine. Regrettably, the combination therapy of DCVAC/PCa, docetaxel, and prednisone was deemed ineffective in extending OS in patients with mCRPC (NCT02111577) (Vogelzang et al., 2022). In addition, individualized polypeptide vaccine (PPV) stands out, bypassing immune diversity and evading immune tolerance (Wang et al., 2023). Yoshimura et al. (2016) compared clinical outcomes of the treatment with PPV, adding dexamethasone versus dexamethasone alone in 2016, where the PPV group presented longer median OS and progression-free survival (PFS). Of note, the recruited patients in this study were diagnosed in the early stage of CRPC (Yoshimura et al., 2016). Another prostate cancer vaccine, GVAX, has been shown to induce infiltrating immune cells that may promote PD-L1 upregulation (Palicelli et al., 2021). However, the exact efficacy remains to be unveiled.
DNA vaccines could evoke antitumor immune response by changing the sequence of plasmid DNA (Wang et al., 2023). An example is the pTVG-HP vaccine, which encodes the human PAP antigens and is being evaluated in mCRPC trials. Given their instability and inefficiency, the development of mRNA-based vaccines is still in slow progress (Bansal et al., 2021).
CAR-T cell therapy targeting PCSC-associated antigens emerges as a promising therapeutic approach. Despite no results, some phase I clinical trials with PSCA are ongoing to assess the immune activity of PSCA-specific CAR-T cells in patients with mCRPC (NCT03927573 and NCT03873805). Subsequently, BPX-601 acted as a PSCA-directed CAR-T cell and was applied in the clinical trial I/II, in which feasibility, safety, and clinical activity were measured at the recommended dose (NCT02744287). Both BPX-601 and 4-1BB are designed to enhance the immune response of patients with PSCA+ mCRPC (NCT03873805). By targeting another well-known antigen (EpCAM), EpCAM-specific CAR-T cell is introduced into human peripheral blood lymphocytes (PBLs) with the strategy of substantially preventing PC-3 growth in vitro and in vivo (Deng et al., 2015).
Owing to PCSC resistance to fractionated irradiation, which is characterized by high B7-H3 levels, B7-H3 CAR-T cells are demonstrated to support radiation therapy against PCSCs (Zhang et al., 2021). In addition, the CAR-T cell strategy targeting PSMA with lutetium-177 (177Lu-J591) has proven a clinical benefit in phase II clinical trial testing (Tagawa et al., 2013). The results from Frieling et al. (2023) revealed that γδ CAR-T cells targeting PSCA caused a robust regression of established tumors in a preclinical murine model of bone mCRPC. Another novel cell therapy, the tumor-infiltrating lymphocytes (TILs) strategy, has gained striking momentum. Recently, Gao’s group overcame sorafenib resistance to liver cancer by targeting stem-like CCR4+ regulatory T cells and inhibiting the maintenance of the TIL-Treg pool (Gao et al., 2022). However, obtaining TILs from PCa patients with poor immunogenicity remains challenging. In 2019, Yunger et al. (2019) managed to expand TILs from eight PCa patients under ADT treatment, supporting the development of prostate-TIL therapy. Experiments based on PC3-bearing humanized immunodeficiency IL2Rγ null (hNSG) mice with an intravenous injection of human CD34+ hematopoietic stem cells indicated that the N-cadherin antagonist ADH-1 promoted TIL antitumor responses (Sun et al., 2021). Elevated density of CD8+ TILs was demonstrated to improve clinical outcomes from PCa patients undergoing radical prostatectomy (Yang et al., 2021). Although no phase III data have been reported for prostate-TIL products, some clinical trials are recruiting patients. Collectively, CAR-T cells targeting PCSCs and TILs represent promising therapeutic options in the future.
PCSCs are the cancer-initiating cells that play a pivotal role in tumor relapse and therapy resistance. Identifying the characteristics and presence of PCSCs is important to reveal their mechanism and develop targeted therapies against CSC. The establishment of a 3D culture system provides general support for the point that basal stem-like cells are suggested to be the cells of origin in primary prostatic tumors, while stem-like cells with luminal phenotypes reinitiate CRPC relapse after ADT. Additionally, PCSCs could also exist in reprogrammed non-epithelial cancerous cells (i.e., inflammatory and stromal cells). As Figure 1A shows, putative biomarkers for PCSCs from basal (KRT16/17/6, Bcl-2, CK5, CK14, p63, hTERT, Trop2, β-catenin, ERα, ALDH, Sca-1, CD133, CD44, CD117, CD49f, Nanog, α2 integrin, CXCR4, EpCaM, CD54, and SOX2) and luminal (Tacstd2, NKX3-1, CK18, CK19, CK8, CK4, CD26, OCT3/4, DLL4, PSCA, BMI-1, EZH2, ABCG2, etc.) are listed. Of note, PCSC-related therapies concentrating on PCSC-related pathways, the PCSC microenvironment, miRNA, and immunotherapy (see Figure 1B) are valid goals to aim for and also have massive hurdles to overcome. Collectively, based on this review of PCSC characteristics and accessible clinical trials, it is clear that a great need exists for further testing of these targeted therapies.
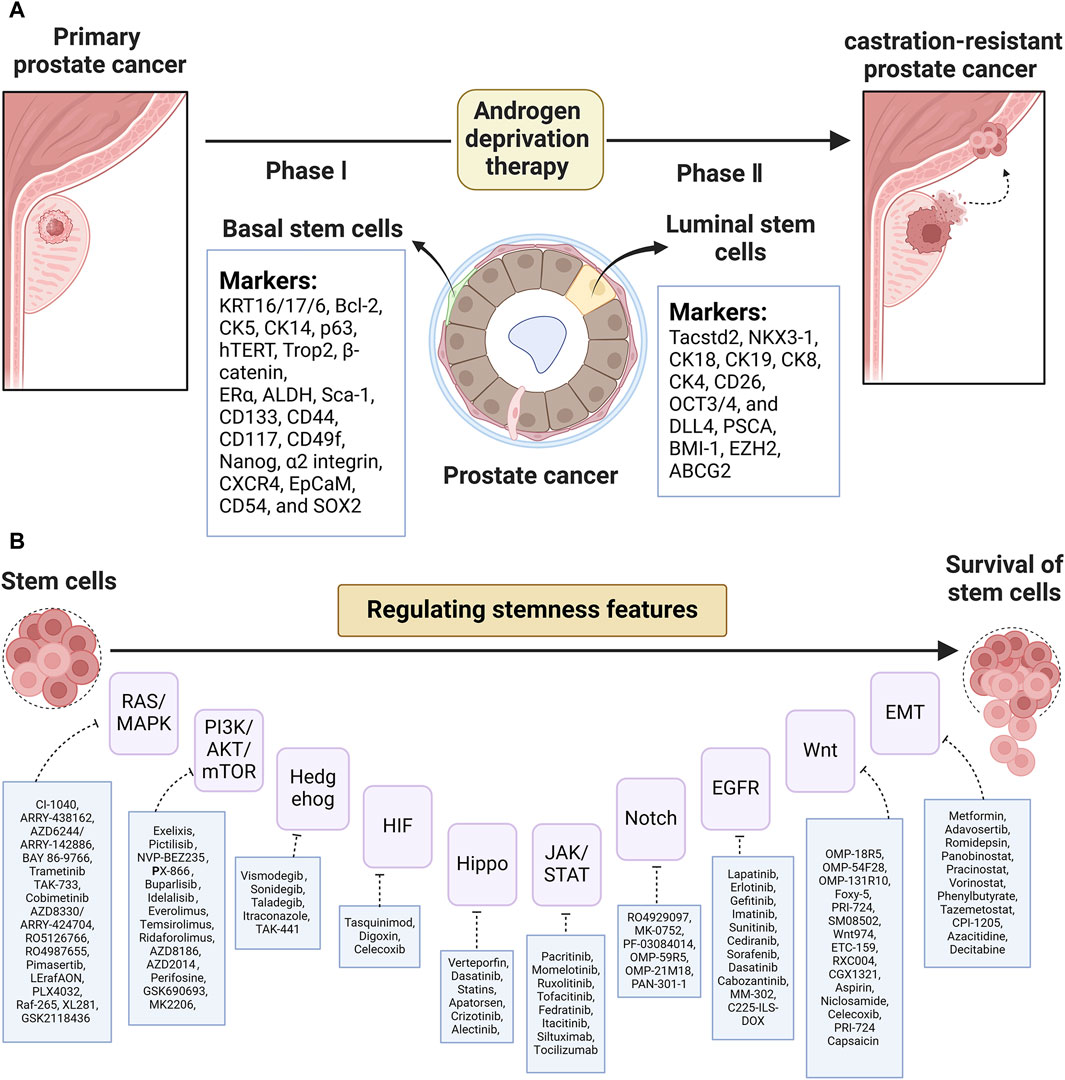
Figure 1. (A) List of putative biomarkers for CSCs based on basal and luminal layers and (B) CSC-related pathway targeted agents in PCa.
The work has several critical limitations:
1. The description of the isolation and enrichment of PCSCs is limited and warrants a more thorough examination to provide greater insights.
2. Apart from PCSCs, drug resistance in PCa involves factors such as hypoxia, oxidative regulation, EMT, and autophagy. A more extensive discussion is needed.
3. Although numerous clinical studies are underway, their outcomes remain inconclusive. Further monitoring and statistical analysis are warranted.
4. While this work predominantly focuses on the role of signaling pathways in PCSC development, the significance of PCa-related metabolism should also be explored.
HS: writing–original draft. LH: investigation, resources, and writing–review and editing. JZ: resources, validation, and writing–review and editing. GY: funding acquisition and writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Scientific Research Foundation for Advanced Talents of Shanghai East Hospital (DFRC2020003 to GY) and the Excellent Young Medical Talents Training Project of Pudong New Area Health Committee (PWRq2020-43).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abate-Shen, C., and Shen, M. M. (2000). Molecular genetics of prostate cancer. Genes. Dev. 14 (19), 2410–2434. doi:10.1101/gad.819500
Ali, T. Z., and Epstein, J. I. (2007). Basal cell carcinoma of the prostate: a clinicopathologic study of 29 cases. Am. J. Surg. Pathol. 31 (5), 697–705. doi:10.1097/01.pas.0000213395.42075.86
Antonarakis, E. S., Carducci, M. A., and Eisenberger, M. A. (2010). Novel targeted therapeutics for metastatic castration-resistant prostate cancer. Cancer Lett. 291 (1), 1–13. doi:10.1016/j.canlet.2009.08.012
Antonarakis, E. S., Piulats, J. M., Gross-Goupil, M., Goh, J., Ojamaa, K., Hoimes, C. J., et al. (2020). Pembrolizumab for treatment-refractory metastatic castration-resistant prostate cancer: multicohort, open-label phase II KEYNOTE-199 study. J. Clin. Oncol. 38 (5), 395–405. doi:10.1200/JCO.19.01638
Banerjee, P., Kapse, P., Siddique, S., Kundu, M., Choudhari, J., Mohanty, V., et al. (2023). Therapeutic implications of cancer stem cells in prostate cancer. Cancer Biol. Med. 20 (6), 401–420. doi:10.20892/j.issn.2095-3941.2022.0714
Bansal, D., Reimers, M. A., Knoche, E. M., and Pachynski, R. K. (2021). Immunotherapy and immunotherapy combinations in metastatic castration-resistant prostate cancer. Cancers (Basel) 13 (2), 334. doi:10.3390/cancers13020334
Bitting, R. L., and Armstrong, A. J. (2013). Targeting the PI3K/Akt/mTOR pathway in castration-resistant prostate cancer. Endocr. Relat. Cancer 20 (3), R83–R99. doi:10.1530/ERC-12-0394
Cha, H. R., Lee, J. H., and Ponnazhagan, S. (2020). Revisiting immunotherapy: a focus on prostate cancer. Cancer Res. 80 (8), 1615–1623. doi:10.1158/0008-5472.CAN-19-2948
Chang, L., Graham, P. H., Ni, J., Hao, J., Bucci, J., Cozzi, P. J., et al. (2015). Targeting PI3K/Akt/mTOR signaling pathway in the treatment of prostate cancer radioresistance. Crit. Rev. Oncol. Hematol. 96 (3), 507–517. doi:10.1016/j.critrevonc.2015.07.005
Chaves, L. P., Melo, C. M., Saggioro, F. P., Reis, R. B. D., and Squire, J. A. (2021). Epithelial-mesenchymal transition signaling and prostate cancer stem cells: emerging biomarkers and opportunities for precision therapeutics. Genes. (Basel) 12 (12), 1900. doi:10.3390/genes12121900
Chen, L. (2004). Co-inhibitory molecules of the B7-CD28 family in the control of T-cell immunity. Nat. Rev. Immunol. 4 (5), 336–347. doi:10.1038/nri1349
Chen, L., Boleslaw Olszewski, M., Kruithof-de Julio, M., and Snaar-Jagalska, B. E. (2020). Zebrafish microenvironment elevates EMT and CSC-like phenotype of engrafted prostate cancer cells. Cells 9 (4), 797. doi:10.3390/cells9040797
Ci, X., Hao, J., Dong, X., Xue, H., Wu, R., Choi, S. Y. C., et al. (2020). Conditionally reprogrammed cells from patient-derived xenograft to model neuroendocrine prostate cancer development. Cells 9 (6), 1398. doi:10.3390/cells9061398
Cocciadiferro, L., Miceli, V., Kang, K. S., Polito, L. M., Trosko, J. E., and Carruba, G. (2009). Profiling cancer stem cells in androgen-responsive and refractory human prostate tumor cell lines. Ann. N. Y. Acad. Sci. 1155, 257–262. doi:10.1111/j.1749-6632.2009.03696.x
Coffey, K. (2021). Targeting the Hippo pathway in prostate cancer: what's new? Cancers (Basel) 13 (4), 611. doi:10.3390/cancers13040611
Cortes, J. E., Gutzmer, R., Kieran, M. W., and Solomon, J. A. (2019). Hedgehog signaling inhibitors in solid and hematological cancers. Cancer Treat. Rev. 76, 41–50. doi:10.1016/j.ctrv.2019.04.005
Costa, C. D., Justo, A. A., Kobayashi, P. E., Story, M. M., Palmieri, C., Laufer Amorim, R., et al. (2019). Characterization of OCT3/4, Nestin, NANOG, CD44 and CD24 as stem cell markers in canine prostate cancer. Int. J. Biochem. Cell. Biol. 108, 21–28. doi:10.1016/j.biocel.2019.01.002
Cui, D., Dai, J., Keller, J. M., Mizokami, A., Xia, S., and Keller, E. T. (2015). Notch pathway inhibition using PF-03084014, a gamma-secretase inhibitor (GSI), enhances the antitumor effect of docetaxel in prostate cancer. Clin. Cancer Res. 21 (20), 4619–4629. doi:10.1158/1078-0432.CCR-15-0242
Darash-Yahana, M., Pikarsky, E., Abramovitch, R., Zeira, E., Pal, B., Karplus, R., et al. (2004). Role of high expression levels of CXCR4 in tumor growth, vascularization, and metastasis. FASEB J. 18 (11), 1240–1242. doi:10.1096/fj.03-0935fje
Deng, Z., Wu, Y., Ma, W., Zhang, S., and Zhang, Y. Q. (2015). Adoptive T-cell therapy of prostate cancer targeting the cancer stem cell antigen EpCAM. BMC Immunol. 16 (1), 1. doi:10.1186/s12865-014-0064-x
de Wet, L., Williams, A., Gillard, M., Kregel, S., Lamperis, S., Gutgesell, L. C., et al. (2022). SOX2 mediates metabolic reprogramming of prostate cancer cells. Oncogene 41 (8), 1190–1202. doi:10.1038/s41388-021-02157-x
Dong, B., Miao, J., Wang, Y., Luo, W., Ji, Z., Lai, H., et al. (2020). Single-cell analysis supports a luminal-neuroendocrine transdifferentiation in human prostate cancer. Commun. Biol. 3 (1), 778. doi:10.1038/s42003-020-01476-1
El Bezawy, R., Tinelli, S., Tortoreto, M., Doldi, V., Zuco, V., Folini, M., et al. (2019). miR-205 enhances radiation sensitivity of prostate cancer cells by impairing DNA damage repair through PKCε and ZEB1 inhibition. J. Exp. Clin. Cancer Res. 38 (1), 51. doi:10.1186/s13046-019-1060-z
Fiori, M. E., Di Franco, S., Villanova, L., Bianca, P., Stassi, G., and De Maria, R. (2019). Cancer-associated fibroblasts as abettors of tumor progression at the crossroads of EMT and therapy resistance. Mol. Cancer 18 (1), 70. doi:10.1186/s12943-019-0994-2
Fizazi, K., Retz, M., Petrylak, D. P., Goh, J. C., Perez-Gracia, J., Lacombe, L., et al. (2022). Nivolumab plus rucaparib for metastatic castration-resistant prostate cancer: results from the phase 2 CheckMate 9KD trial. J. Immunother. Cancer 10 (8), e004761. doi:10.1136/jitc-2022-004761
Frieling, J. S., Tordesillas, L., Bustos, X. E., Ramello, M. C., Bishop, R. T., Cianne, J. E., et al. (2023). γδ-Enriched CAR-T cell therapy for bone metastatic castrate-resistant prostate cancer. Sci. Adv. 9 (18), eadf0108. doi:10.1126/sciadv.adf0108
Galoczova, M., Nenutil, R., Pokorna, Z., Vojtesek, B., and Coates, P. J. (2021). TAp63 and ΔNp63 (p40) in prostate adenocarcinomas: ΔNp63 associates with a basal-like cancer stem cell population but not with metastasis. Virchows Arch. 478 (4), 627–636. doi:10.1007/s00428-020-02944-z
Gao, Y., Wang, Y. T., Chen, Y., Wang, H., Young, D., Shi, T., et al. (2020). Proteomic tissue-based classifier for early prediction of prostate cancer progression. Cancers (Basel) 12 (5), 1268. doi:10.3390/cancers12051268
Gao, Y., You, M., Fu, J., Tian, M., Zhong, X., Du, C., et al. (2022). Intratumoral stem-like CCR4+ regulatory T cells orchestrate the immunosuppressive microenvironment in HCC associated with hepatitis B. J. Hepatol. 76 (1), 148–159. doi:10.1016/j.jhep.2021.08.029
Germann, M., Wetterwald, A., Guzmán-Ramirez, N., van der Pluijm, G., Culig, Z., Cecchini, M. G., et al. (2012). Stem-like cells with luminal progenitor phenotype survive castration in human prostate cancer. Stem Cells 30 (6), 1076–1086. doi:10.1002/stem.1087
Giannoni, E., Bianchini, F., Masieri, L., Serni, S., Torre, E., Calorini, L., et al. (2010). Reciprocal activation of prostate cancer cells and cancer-associated fibroblasts stimulates epithelial-mesenchymal transition and cancer stemness. Cancer Res. 70 (17), 6945–6956. doi:10.1158/0008-5472.CAN-10-0785
Goldstein, A. S., Lawson, D. A., Cheng, D., Sun, W., Garraway, I. P., and Witte, O. N. (2008). Trop2 identifies a subpopulation of murine and human prostate basal cells with stem cell characteristics. Proc. Natl. Acad. Sci. U. S. A. 105 (52), 20882–20887. doi:10.1073/pnas.0811411106
Gorodetska, I., Offermann, A., Püschel, J., Lukiyanchuk, V., Gaete, D., Kurzyukova, A., et al. (2024). ALDH1A1 drives prostate cancer metastases and radioresistance by interplay with AR- and RAR-dependent transcription. Theranostics 14 (2), 714–737. doi:10.7150/thno.88057
Groth, C., and Fortini, M. E. (2012). Therapeutic approaches to modulating Notch signaling: current challenges and future prospects. Semin. Cell. Dev. Biol. 23 (4), 465–472. doi:10.1016/j.semcdb.2012.01.016
Guo, W., Li, L., He, J., Liu, Z., Han, M., Li, F., et al. (2020). Single-cell transcriptomics identifies a distinct luminal progenitor cell type in distal prostate invagination tips. Nat. Genet. 52 (9), 908–918. doi:10.1038/s41588-020-0642-1
Hall, W. A., Sabharwal, L., Udhane, V., Maranto, C., and Nevalainen, M. T. (2020). Cytokines, JAK-STAT signaling and radiation-induced DNA repair in solid tumors: novel opportunities for radiation therapy. Int. J. Biochem. Cell. Biol. 127, 105827. doi:10.1016/j.biocel.2020.105827
Han, Q., Xie, Q. R., Li, F., Cheng, Y., Wu, T., Zhang, Y., et al. (2021). Targeted inhibition of SIRT6 via engineered exosomes impairs tumorigenesis and metastasis in prostate cancer. Theranostics 11 (13), 6526–6541. doi:10.7150/thno.53886
Harris, K. S., Shi, L., Foster, B. M., Mobley, M. E., Elliott, P. L., Song, C. J., et al. (2021). CD117/c-kit defines a prostate CSC-like subpopulation driving progression and TKI resistance. Sci. Rep. 11 (1), 1465. doi:10.1038/s41598-021-81126-6
Hoogland, A. M., Verhoef, E. I., Roobol, M. J., Schröder, F. H., Wildhagen, M. F., van der Kwast, T. H., et al. (2014). Validation of stem cell markers in clinical prostate cancer: α6-integrin is predictive for non-aggressive disease. Prostate 74 (5), 488–496. doi:10.1002/pros.22768
Hotte, S. J., Chi, K. N., Joshua, A. M., Macfarlane, R. J., Gregg, R. W., et al. (2019). A phase II study of PX-866 in patients with recurrent or metastatic castration-resistant prostate cancer: Canadian cancer trials group study IND205. Clin. Genitourin. Cancer 17 (3), 201–208.e1. doi:10.1016/j.clgc.2019.03.005
Hu, W. Y., Hu, D. P., Xie, L., Nonn, L., Lu, R., Abern, M., et al. (2021). Keratin profiling by single-cell RNA-sequencing identifies human prostate stem cell lineage hierarchy and cancer stem-like cells. Int. J. Mol. Sci. 22 (15), 8109. doi:10.3390/ijms22158109
Iyer, A. K., Singh, A., Ganta, S., and Amiji, M. M. (2013). Role of integrated cancer nanomedicine in overcoming drug resistance. Adv. Drug Deliv. Rev. 65 (13-14), 1784–1802. doi:10.1016/j.addr.2013.07.012
Jeter, C. R., Badeaux, M., Choy, G., Chandra, D., Patrawala, L., Liu, C., et al. (2009). Functional evidence that the self-renewal gene NANOG regulates human tumor development. Stem Cells 27 (5), 993–1005. doi:10.1002/stem.29
Ju, M., Fan, J., Zou, Y., Yu, M., Jiang, L., Wei, Q., et al. (2022). Computational recognition of a regulatory T-cell-specific signature with potential implications in prognosis, immunotherapy, and therapeutic resistance of prostate cancer. Front. Immunol. 13, 807840. doi:10.3389/fimmu.2022.807840
Kantoff, P. W., Schuetz, T. J., Blumenstein, B. A., Glode, L. M., Bilhartz, D. L., Wyand, M., et al. (2010). Overall survival analysis of a phase II randomized controlled trial of a Poxviral-based PSA-targeted immunotherapy in metastatic castration-resistant prostate cancer. J. Clin. Oncol. 28 (7), 1099–1105. doi:10.1200/JCO.2009.25.0597
Kanwal, M., Smahel, M., Olsen, M., Smahelova, J., and Tachezy, R. (2020). Aspartate β-hydroxylase as a target for cancer therapy. J. Exp. Clin. Cancer Res. 39 (1), 163. doi:10.1186/s13046-020-01669-w
Karlou, M., Tzelepi, V., and Efstathiou, E. (2010). Therapeutic targeting of the prostate cancer microenvironment. Nat. Rev. Urol. 7 (9), 494–509. doi:10.1038/nrurol.2010.134
Karthaus, W. R., Iaquinta, P. J., Drost, J., Gracanin, A., van Boxtel, R., Wongvipat, J., et al. (2014). Identification of multipotent luminal progenitor cells in human prostate organoid cultures. Cell. 159 (1), 163–175. doi:10.1016/j.cell.2014.08.017
Kong, D., Heath, E., Chen, W., Cher, M. L., Powell, I., Heilbrun, L., et al. (2012). Loss of let-7 up-regulates EZH2 in prostate cancer consistent with the acquisition of cancer stem cell signatures that are attenuated by BR-DIM. PLoS One 7 (3), e33729. doi:10.1371/journal.pone.0033729
Kroon, P., Berry, P. A., Stower, M. J., Rodrigues, G., Mann, V. M., Simms, M., et al. (2013). JAK-STAT blockade inhibits tumor initiation and clonogenic recovery of prostate cancer stem-like cells. Cancer Res. 73 (16), 5288–5298. doi:10.1158/0008-5472.CAN-13-0874
Kwon, O. J., Zhang, L., Ittmann, M. M., and Xin, L. (2014). Prostatic inflammation enhances basal-to-luminal differentiation and accelerates initiation of prostate cancer with a basal cell origin. Proc. Natl. Acad. Sci. U. S. A. 111 (5), E592–E600. doi:10.1073/pnas.1318157111
Lee, Y., Yoon, J., Ko, D., Yu, M., Lee, S., and Kim, S. (2021). TMPRSS4 promotes cancer stem-like properties in prostate cancer cells through upregulation of SOX2 by SLUG and TWIST1. J. Exp. Clin. Cancer Res. 40 (1), 372. doi:10.1186/s13046-021-02147-7
Lentz, R. W., Colton, M. D., Mitra, S. S., and Messersmith, W. A. (2021). Innate immune checkpoint inhibitors: the next breakthrough in medical oncology? Mol. Cancer Ther. 20 (6), 961–974. doi:10.1158/1535-7163.MCT-21-0041
Leong, K. G., Wang, B. E., Johnson, L., and Gao, W. Q. (2008). Generation of a prostate from a single adult stem cell. Nature 456 (7223), 804–808. doi:10.1038/nature07427
Li, C., Liu, S., Yan, R., Han, N., Wong, K. K., and Li, L. (2017a). CD54-NOTCH1 axis controls tumor initiation and cancer stem cell functions in human prostate cancer. Theranostics 7 (1), 67–80. doi:10.7150/thno.16752
Li, Y., Wang, L., Liu, J., Zhang, P., An, M., Han, C., et al. (2017b). O-GlcNAcylation modulates Bmi-1 protein stability and potential oncogenic function in prostate cancer. Oncogene 36 (45), 6293–6305. doi:10.1038/onc.2017.223
Lin, J., Denmeade, S., and Carducci, M. A. (2009). HIF-1alpha and calcium signaling as targets for treatment of prostate cancer by cardiac glycosides. Curr. Cancer Drug Targets 9 (7), 881–887. doi:10.2174/156800909789760249
Lu, T. L., and Chen, C. M. (2015). Differential requirements for β-catenin in murine prostate cancer originating from basal versus luminal cells. J. Pathol. 236 (3), 290–301. doi:10.1002/path.4521
McLornan, D. P., Pope, J. E., Gotlib, J., and Harrison, C. N. (2021). Current and future status of JAK inhibitors. Lancet 398 (10302), 803–816. doi:10.1016/S0140-6736(21)00438-4
Meisel, C. T., Porcheri, C., and Mitsiadis, T. A. (2020). Cancer stem cells, quo vadis? The Notch signaling pathway in tumor initiation and progression. Cells 9 (8), 1879. doi:10.3390/cells9081879
Messina, B., Lo Sardo, F., Scalera, S., Memeo, L., Colarossi, C., Mare, M., et al. (2023). Hippo pathway dysregulation in gastric cancer: from Helicobacter pylori infection to tumor promotion and progression. Cell. Death Dis. 14 (1), 21. doi:10.1038/s41419-023-05568-8
Mohtar, M. A., Syafruddin, S. E., Nasir, S. N., and Low, T. Y. (2020). Revisiting the roles of pro-metastatic EpCAM in cancer. Biomolecules 10 (2), 255. doi:10.3390/biom10020255
Mukhopadhyay, N. K., Cinar, B., Mukhopadhyay, L., Lutchman, M., Ferdinand, A. S., Kim, J., et al. (2007). The zinc finger protein ras-responsive element binding protein-1 is a coregulator of the androgen receptor: implications for the role of the Ras pathway in enhancing androgenic signaling in prostate cancer. Mol. Endocrinol. 21 (9), 2056–2070. doi:10.1210/me.2006-0503
Mulholland, D. J., Kobayashi, N., Ruscetti, M., Zhi, A., Tran, L. M., Huang, J., et al. (2012). Pten loss and RAS/MAPK activation cooperate to promote EMT and metastasis initiated from prostate cancer stem/progenitor cells. Cancer Res. 72 (7), 1878–1889. doi:10.1158/0008-5472.CAN-11-3132
Nabhan, C., Lestingi, T. M., Galvez, A., Tolzien, K., Kelby, S. K., Tsarwhas, D., et al. (2009). Erlotinib has moderate single-agent activity in chemotherapy-naive castration-resistant prostate cancer: final results of a phase II trial. Urology 74 (3), 665–671. doi:10.1016/j.urology.2009.05.016
Ojemuyiwa, M. A., Madan, R. A., and Dahut, W. L. (2014). Tyrosine kinase inhibitors in the treatment of prostate cancer: taking the next step in clinical development. Expert Opin. Emerg. Drugs 19 (4), 459–470. doi:10.1517/14728214.2014.969239
Olsson, A., Björk, A., Vallon-Christersson, J., Isaacs, J. T., and Leanderson, T. (2010). Tasquinimod (ABR-215050), a quinoline-3-carboxamide anti-angiogenic agent, modulates the expression of thrombospondin-1 in human prostate tumors. Mol. Cancer 9, 107. doi:10.1186/1476-4598-9-107
O’Reilly, D., Johnson, P., and Buchanan, P. J. (2019). Hypoxia induced cancer stem cell enrichment promotes resistance to androgen deprivation therapy in prostate cancer. Steroids 152, 108497. doi:10.1016/j.steroids.2019.108497
Palicelli, A., Bonacini, M., Croci, S., Magi-Galluzzi, C., Cañete-Portillo, S., Chaux, A., et al. (2021). What do we have to know about PD-L1 expression in prostate cancer? A systematic literature review. Part 2: clinic-pathologic correlations. Cells 10 (11), 3165. doi:10.3390/cells10113165
Papatsoris, A. G., Karamouzis, M. V., and Papavassiliou, A. G. (2007). The power and promise of "rewiring" the mitogen-activated protein kinase network in prostate cancer therapeutics. Mol. Cancer Ther. 6 (3), 811–819. doi:10.1158/1535-7163.MCT-06-0610
Park, B. J., Park, J. I., Byun, D. S., Park, J. H., and Chi, S. G. (2000). Mitogenic conversion of transforming growth factor-beta1 effect by oncogenic Ha-Ras-induced activation of the mitogen-activated protein kinase signaling pathway in human prostate cancer. Cancer Res. 60 (11), 3031–3038.
Park, J. W., Lee, J. K., Phillips, J. W., Huang, P., Cheng, D., Huang, J., et al. (2016). Prostate epithelial cell of origin determines cancer differentiation state in an organoid transformation assay. Proc. Natl. Acad. Sci. U. S. A. 113 (16), 4482–4487. doi:10.1073/pnas.1603645113
Parsons, J. K., Pinto, P. A., Pavlovich, C. P., Uchio, E., Nguyen, M. N., Kim, H. L., et al. (2023). A phase 2, double-blind, randomized controlled trial of PROSTVAC in prostate cancer patients on active surveillance. Eur. Urol. Focus 9 (3), 447–454. doi:10.1016/j.euf.2022.12.002
Patrawala, L., Calhoun, T., Schneider-Broussard, R., Li, H., Bhatia, B., Tang, S., et al. (2006). Highly purified CD44+ prostate cancer cells from xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells. Oncogene 25 (12), 1696–1708. doi:10.1038/sj.onc.1209327
Patrawala, L., Calhoun-Davis, T., Schneider-Broussard, R., and Tang, D. G. (2007). Hierarchical organization of prostate cancer cells in xenograft tumors: the CD44+alpha2beta1+ cell population is enriched in tumor-initiating cells. Cancer Res. 67 (14), 6796–6805. doi:10.1158/0008-5472.CAN-07-0490
Porcacchia, A. S., Pires, G. N., Ortiz, V., Andersen, M. L., and Tufik, S. (2022). Prostate cancer mortality and costs of prostate surgical procedures in the Brazilian public health system. Int. Braz J. Urol. 48 (3), 583–590. doi:10.1590/S1677-5538.IBJU.2021.0781
Qiu, H., Cao, S., and Xu, R. (2021). Cancer incidence, mortality, and burden in China: a time-trend analysis and comparison with the United States and United Kingdom based on the global epidemiological data released in 2020. Cancer Commun. (Lond) 41 (10), 1037–1048. doi:10.1002/cac2.12197
Ramesh, S., Selvakumar, P., Ameer, M. Y., Lian, S., Abdullah Alzarooni, A. I. M., Ojha, S., et al. (2023). State-of-the-art therapeutic strategies for targeting cancer stem cells in prostate cancer. Front. Oncol. 13, 1059441. doi:10.3389/fonc.2023.1059441
Ranjbar, A., Soltanshahi, M., Taghiloo, S., and Asgarian-Omran, H. (2023). Glucose metabolism in acute myeloid leukemia cell line is regulated via combinational PI3K/AKT/mTOR pathway inhibitors. Iran. J. Pharm. Res. 22 (1), e140507. doi:10.5812/ijpr-140507
Rizzo, M. (2021). Mechanisms of docetaxel resistance in prostate cancer: the key role played by miRNAs. Biochim. Biophys. Acta Rev. Cancer 1875 (1), 188481. doi:10.1016/j.bbcan.2020.188481
Rudzinski, J. K., Govindasamy, N. P., Asgari, A., Saito, M. S., Lewis, J. D., and Jurasz, P. (2021). Preferential interaction of platelets with prostate cancer cells with stem cell markers. Thromb. Res. 206, 42–51. doi:10.1016/j.thromres.2021.08.008
Rybak, A. P., Ingram, A. J., and Tang, D. (2013). Propagation of human prostate cancer stem-like cells occurs through EGFR-mediated ERK activation. PLoS One 8 (4), e61716. doi:10.1371/journal.pone.0061716
Saad, F., Shore, N., Zhang, T., Sharma, S., Cho, H. K., and Jacobs, I. A. (2019). Emerging therapeutic targets for patients with advanced prostate cancer. Cancer Treat. Rev. 76, 1–9. doi:10.1016/j.ctrv.2019.03.002
Sabnis, N. G., Miller, A., Titus, M. A., and Huss, W. J. (2017). The efflux transporter ABCG2 maintains prostate stem cells. Mol. Cancer Res. 15 (2), 128–140. doi:10.1158/1541-7786.MCR-16-0270-T
Sandhu, S., Moore, C. M., Chiong, E., Beltran, H., Bristow, R. G., and Williams, S. G. (2021). Prostate cancer. Lancet 398 (10305), 1075–1090. doi:10.1016/S0140-6736(21)00950-8
Santarpia, L., Lippman, S. M., and El-Naggar, A. K. (2012). Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy. Expert Opin. Ther. Targets 16 (1), 103–119. doi:10.1517/14728222.2011.645805
Sarker, D., Reid, A. H. M., Yap, T. A., and de Bono, J. S. (2009). Targeting the PI3K/AKT pathway for the treatment of prostate cancer. Clin. Cancer Res. 15 (15), 4799–4805. doi:10.1158/1078-0432.CCR-08-0125
Schmidt, K. T., Peer, C. J., Huitema, A. D. R., Williams, M. D., Wroblewski, S., Schellens, J. H. M., et al. (2020). Measurement of NLG207 (formerly CRLX101) nanoparticle-bound and released camptothecin in human plasma. J. Pharm. Biomed. Anal. 181, 113073. doi:10.1016/j.jpba.2019.113073
Shen, Y., Cao, J., Liang, Z., Lin, Q., Wang, J., Yang, X., et al. (2019). Estrogen receptor α-NOTCH1 axis enhances basal stem-like cells and epithelial-mesenchymal transition phenotypes in prostate cancer. Cell. Commun. Signal 17 (1), 50. doi:10.1186/s12964-019-0367-x
Siegel, R. L., Miller, K. D., Wagle, N. S., and Jemal, A. (2023). Cancer statistics, 2023. CA Cancer J. Clin. 73 (1), 17–48. doi:10.3322/caac.21763
Sooriakumaran, P., Coley, H. M., Fox, S. B., Macanas-Pirard, P., Lovell, D. P., Henderson, A., et al. (2009). A randomized controlled trial investigating the effects of celecoxib in patients with localized prostate cancer. Anticancer Res. 29 (5), 1483–1488.
Sridhar, S. S., Hotte, S. J., Chin, J. L., Hudes, G. R., Gregg, R., Trachtenberg, J., et al. (2010). A multicenter phase II clinical trial of lapatinib (GW572016) in hormonally untreated advanced prostate cancer. Am. J. Clin. Oncol. 33 (6), 609–613. doi:10.1097/COC.0b013e3181beac33
Storck, W. K., May, A. M., Westbrook, T. C., Duan, Z., Morrissey, C., Yates, J. A., et al. (2022). The role of epigenetic change in therapy-induced neuroendocrine prostate cancer lineage plasticity. Front. Endocrinol. (Lausanne) 13, 926585. doi:10.3389/fendo.2022.926585
Sui, X., Cai, J., Li, H., He, C., Zhou, C., Dong, Y., et al. (2018). p53-dependent CD51 expression contributes to characteristics of cancer stem cells in prostate cancer. Cell. Death Dis. 9 (5), 523. doi:10.1038/s41419-018-0541-x
Sun, Y., Jing, J., Xu, H., Xu, L., Hu, H., Tang, C., et al. (2021). N-cadherin inhibitor creates a microenvironment that protect TILs from immune checkpoints and Treg cells. J. Immunother. Cancer 9 (3), e002138. doi:10.1136/jitc-2020-002138
Tagawa, S. T., Milowsky, M. I., Morris, M., Vallabhajosula, S., Christos, P., Akhtar, N. H., et al. (2013). Phase II study of Lutetium-177-labeled anti-prostate-specific membrane antigen monoclonal antibody J591 for metastatic castration-resistant prostate cancer. Clin. Cancer Res. 19 (18), 5182–5191. doi:10.1158/1078-0432.CCR-13-0231
Tang, D. G. (2022). Understanding and targeting prostate cancer cell heterogeneity and plasticity. Semin. Cancer Biol. 82, 68–93. doi:10.1016/j.semcancer.2021.11.001
Terrisse, S., Zitvogel, L., and Kroemer, G. (2022). Effects of the intestinal microbiota on prostate cancer treatment by androgen deprivation therapy. Microb. Cell. 9 (12), 202–206. doi:10.15698/mic2022.12.787
Toivanen, R., Mohan, A., and Shen, M. M. (2016). Basal progenitors contribute to repair of the prostate epithelium following induced luminal anoikis. Stem Cell. Rep. 6 (5), 660–667. doi:10.1016/j.stemcr.2016.03.007
Tong, W., Qiu, L., Qi, M., Liu, J., Hu, K., Lin, W., et al. (2018). GANT-61 and GDC-0449 induce apoptosis of prostate cancer stem cells through a GLI-dependent mechanism. J. Cell. Biochem. 119 (4), 3641–3652. doi:10.1002/jcb.26572
Verma, P., Shukla, N., Kumari, S., Ansari, M. S., Gautam, N. K., and Patel, G. K. (2023). Cancer stem cell in prostate cancer progression, metastasis and therapy resistance. Biochim. Biophys. Acta Rev. Cancer 1878 (3), 188887. doi:10.1016/j.bbcan.2023.188887
Vogelzang, N. J., Beer, T. M., Gerritsen, W., Oudard, S., Wiechno, P., Kukielka-Budny, B., et al. (2022). Efficacy and safety of autologous dendritic cell-based immunotherapy, docetaxel, and prednisone vs placebo in patients with metastatic castration-resistant prostate cancer: the VIABLE phase 3 randomized clinical trial. JAMA Oncol. 8 (4), 546–552. doi:10.1001/jamaoncol.2021.7298
Wan, S., Zhao, E., Kryczek, I., Vatan, L., Sadovskaya, A., Ludema, G., et al. (2014). Tumor-associated macrophages produce interleukin 6 and signal via STAT3 to promote expansion of human hepatocellular carcinoma stem cells. Gastroenterology 147 (6), 1393–1404. doi:10.1053/j.gastro.2014.08.039
Wang, D., Sun, Y., Liu, Y., Meng, F., and Lee, R. J. (2018). Clinical translation of immunoliposomes for cancer therapy: recent perspectives. Expert Opin. Drug Deliv. 15 (9), 893–903. doi:10.1080/17425247.2018.1517747
Wang, J., Zhou, K., Zhu, H., Wei, F., Ma, S., Kan, Y., et al. (2023). Current status and progress of the development of prostate cancer vaccines. J. Cancer 14 (5), 835–842. doi:10.7150/jca.80803
Wang, L., Lyu, C., Stadlbauer, B., Buchner, A., Nößner, E., and Pohla, H. (2024). Berbamine targets cancer stem cells and reverses cabazitaxel resistance via inhibiting IGF2BP1 and p-STAT3 in prostate cancer. Prostate 84 (2), 131–147. doi:10.1002/pros.24632
Wang, X., Kruithof-de Julio, M., Economides, K. D., Walker, D., Yu, H., Halili, M. V., et al. (2009). A luminal epithelial stem cell that is a cell of origin for prostate cancer. Nature 461 (7263), 495–500. doi:10.1038/nature08361
Wang, Z., Li, Y., Wang, Y., Wu, D., Lau, A. H. Y., Zhao, P., et al. (2020). Targeting prostate cancer stem-like cells by an immunotherapeutic platform based on immunogenic peptide-sensitized dendritic cells-cytokine-induced killer cells. Stem Cell. Res. Ther. 11 (1), 123. doi:10.1186/s13287-020-01634-6
Wei, X. X., Hsieh, A. C., Kim, W., Friedlander, T., Lin, A. M., Louttit, M., et al. (2017). A phase I study of abiraterone acetate combined with BEZ235, a dual PI3K/mTOR inhibitor, in metastatic castration resistant prostate cancer. Oncologist 22 (5), 503–e43. doi:10.1634/theoncologist.2016-0432
Wolf, I., Gratzke, C., and Wolf, P. (2022). Prostate cancer stem cells: clinical aspects and targeted therapies. Front. Oncol. 12, 935715. doi:10.3389/fonc.2022.935715
Worthmuller, J., and Ruegg, C. (2020). The crosstalk between FAK and Wnt signaling pathways in cancer and its therapeutic implication. Int. J. Mol. Sci. 21 (23), 9107. doi:10.3390/ijms21239107
Wu, J., Cang, S., Liu, C., Ochiai, W., and Chiao, J. W. (2020). Development of human prostate cancer stem cells involves epigenomic alteration and PI3K/AKT pathway activation. Exp. Hematol. Oncol. 9, 12. doi:10.1186/s40164-020-00168-0
Xin, L., Lawson, D. A., and Witte, O. N. (2005). The Sca-1 cell surface marker enriches for a prostate-regenerating cell subpopulation that can initiate prostate tumorigenesis. Proc. Natl. Acad. Sci. U. S. A. 102 (19), 6942–6947. doi:10.1073/pnas.0502320102
Yamada, Y., and Beltran, H. (2021). Clinical and biological features of neuroendocrine prostate cancer. Curr. Oncol. Rep. 23 (2), 15. doi:10.1007/s11912-020-01003-9
Yang, L., Shi, P., Zhao, G., Xu, J., Peng, W., Zhang, J., et al. (2020). Targeting cancer stem cell pathways for cancer therapy. Signal Transduct. Target Ther. 5 (1), 8. doi:10.1038/s41392-020-0110-5
Yang, Y., Attwood, K., Bshara, W., Mohler, J. L., Guru, K., Xu, B., et al. (2021). High intratumoral CD8(+) T-cell infiltration is associated with improved survival in prostate cancer patients undergoing radical prostatectomy. Prostate 81 (1), 20–28. doi:10.1002/pros.24068
Yehya, A., Ghamlouche, F., Zahwe, A., Zeid, Y., Wakimian, K., Mukherji, D., et al. (2022). Drug resistance in metastatic castration-resistant prostate cancer: an update on the status quo. Cancer Drug Resist 5 (3), 667–690. doi:10.20517/cdr.2022.15
Yoo, Y. A., Roh, M., Naseem, A. F., Lysy, B., Desouki, M. M., Unno, K., et al. (2016). Bmi1 marks distinct castration-resistant luminal progenitor cells competent for prostate regeneration and tumour initiation. Nat. Commun. 7, 12943. doi:10.1038/ncomms12943
Yoshimura, K., Minami, T., Nozawa, M., Kimura, T., Egawa, S., Fujimoto, H., et al. (2016). A phase 2 randomized controlled trial of personalized peptide vaccine immunotherapy with low-dose dexamethasone versus dexamethasone alone in chemotherapy-naive castration-resistant prostate cancer. Eur. Urol. 70 (1), 35–41. doi:10.1016/j.eururo.2015.12.050
Yuan, H., Han, Y., Wang, X., Li, N., Liu, Q., Yin, Y., et al. (2020). SETD2 restricts prostate cancer metastasis by integrating EZH2 and AMPK signaling pathways. Cancer Cell. 38 (3), 350–365. doi:10.1016/j.ccell.2020.05.022
Yunger, S., Bar El, A., Zeltzer, L. A., Fridman, E., Raviv, G., Laufer, M., et al. (2019). Tumor-infiltrating lymphocytes from human prostate tumors reveal anti-tumor reactivity and potential for adoptive cell therapy. Oncoimmunology 8 (12), e1672494. doi:10.1080/2162402X.2019.1672494
Zhang, B., Ci, X., Tao, R., Ni, J. J., Xuan, X., King, J. L., et al. (2020). Klf5 acetylation regulates luminal differentiation of basal progenitors in prostate development and regeneration. Nat. Commun. 11 (1), 997. doi:10.1038/s41467-020-14737-8
Zhang, B., Kwon, O. J., Henry, G., Malewska, A., Wei, X., Zhang, L., et al. (2016b). Non-cell-autonomous regulation of prostate epithelial homeostasis by androgen receptor. Mol. Cell. 63 (6), 976–989. doi:10.1016/j.molcel.2016.07.025
Zhang, D., Park, D., Zhong, Y., Lu, Y., Rycaj, K., Gong, S., et al. (2016a). Stem cell and neurogenic gene-expression profiles link prostate basal cells to aggressive prostate cancer. Nat. Commun. 7, 10798. doi:10.1038/ncomms10798
Zhang, D., Zhao, S., Li, X., Kirk, J. S., and Tang, D. G. (2018b). Prostate luminal progenitor cells in development and cancer. Trends Cancer 4 (11), 769–783. doi:10.1016/j.trecan.2018.09.003
Zhang, J., Shen, C., Wang, L., Ma, Q., Xia, P., Qi, M., et al. (2014). Metformin inhibits epithelial-mesenchymal transition in prostate cancer cells: involvement of the tumor suppressor miR30a and its target gene SOX4. Biochem. Biophys. Res. Commun. 452 (3), 746–752. doi:10.1016/j.bbrc.2014.08.154
Zhang, J., Zhang, Y., Cheng, L., Li, C., Dai, L., Zhang, H., et al. (2018a). Enrichment and characterization of cancer stem-like cells in ultra-low concentration of serum and non-adhesive culture system. Am. J. Transl. Res. 10 (5), 1552–1561.
Zhang, K., Guo, Y., Wang, X., Zhao, H., Ji, Z., Cheng, C., et al. (2017). WNT/β-Catenin directs self-renewal symmetric cell division of hTERThigh prostate cancer stem cells. Cancer Res. 77 (9), 2534–2547. doi:10.1158/0008-5472.CAN-16-1887
Zhang, W., Meng, Y., Liu, N., Wen, X. F., and Yang, T. (2015). Insights into chemoresistance of prostate cancer. Int. J. Biol. Sci. 11 (10), 1160–1170. doi:10.7150/ijbs.11439
Zhang, Y., He, L., Sadagopan, A., Ma, T., Dotti, G., Wang, Y., et al. (2021). Targeting radiation-resistant prostate cancer stem cells by B7-H3 CAR T cells. Mol. Cancer Ther. 20 (3), 577–588. doi:10.1158/1535-7163.MCT-20-0446
Zheng, Z., Li, J., Liu, Y., Shi, Z., Xuan, Z., Yang, K., et al. (2022). The crucial role of AR-V7 in enzalutamide-resistance of castration-resistant prostate cancer. Cancers (Basel) 14 (19), 4877. doi:10.3390/cancers14194877
Keywords: prostate cancer, prostate cancer stem cells, basal progenitor cells, luminal progenitor cells, targeted therapy
Citation: Su H, Huang L, Zhou J and Yang G (2024) Prostate cancer stem cells and their targeted therapies. Front. Cell Dev. Biol. 12:1410102. doi: 10.3389/fcell.2024.1410102
Received: 31 March 2024; Accepted: 23 July 2024;
Published: 08 August 2024.
Edited by:
Antal Nógrádi, University of Szeged, HungaryReviewed by:
Kshitiz Raj Shrestha, Independent Researcher, Kathmandu, NepalCopyright © 2024 Su, Huang, Zhou and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianjun Zhou, emhvdWpqXzIwMDBAYWxpeXVuLmNvbQ==; Guosheng Yang, MjAwOHlhbmdndW9zaGVuZ0BzaW5hLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.