 Zhanguo Gao1
Zhanguo Gao1 Yulin Dai
Yulin Dai Zhongming Zhao
Zhongming Zhao Kristin Eckel-Mahan
Kristin Eckel-Mahan Mikhail G. Kolonin
Mikhail G. Kolonin
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
DATA REPORT article
Front. Cell Dev. Biol. , 07 December 2023
Sec. Molecular and Cellular Pathology
Volume 11 - 2023 | https://doi.org/10.3389/fcell.2023.1295072
This article is part of the Research Topic The Emerging Role of Endothelial Cells in Vascular and Metabolic Disorders; Endothelium Regeneration and Vascular Repair is the Future for Therapeutics View all 6 articles
Endothelial cells (EC), lining the vasculature, serve essential functions fundamental for normal physiology of every organ and the organism as a whole. EC are the first line of exposure to toxic xenobiotics as well as endogenously generated molecules causing cell damage and aging. Cell senescence, the irreversible arrest of cell proliferation caused by organelle damage, is an underpinning of aging responsible for tissue changes leading to age-related diseases (Fossel et al., 2022). Cell senescence is characterized by senescence-associated gene expression, and pro-inflammatory senescence-associated secretory phenotype (SASP), which negatively affects the function of non-senescent cells (Gorgoulis et al., 2019; Yousefzadeh et al., 2021; Fossel et al., 2022). The role of EC senescence in aging and disease has remained insufficiently understood. Even in mice it has not been established how EC senescence underlies the pathophysiology of aging by affecting other components of the vascular system, the perivascular/smooth muscle cells, as well as the parenchyma. In distinct organs, EC are exposed to different microenvironmental pressures and may have different rates of replication and self-renewal. In organs undergoing constant remodeling, such as adipose tissues (AT) and skeletal muscle, high proliferative pressure on EC is expected to result in telomere attrition occurring sooner than in other organs. However, reactive oxygen species and other metabolism byproducts may also expedite senescence of EC irrespective of their proliferation in other organs, such as the brain. The resulting dysfunction of EC leads to conduit vessel disease and obstruction to flow, as well as rarefaction of the microvasculature. This may set the stage for cardiovascular diseases and neurodegeneration (Xu et al., 2022). Understanding the changes taking place in EC undergoing senescence in the brain and other organs is essential for the development of new approaches to intervene in metabolic and degenerative diseases.
A key protein protecting cells from senescence is Telomerase, an enzyme coded for by the TERT gene. TERT is required for telomere maintenance, protection from genotoxic stress, and mitochondrial function (Sahin and Depinho, 2010). TERT is active in stem cells but is turned off in somatic cells, which permits telomere erosion and cell aging. Telomere-independent functions of TERT have also surfaced and remain incompletely understood (Stewart et al., 2002; Romaniuk et al., 2019). In addition to global effects on nuclear transcriptome and physiology (Park et al., 2009), recent reports have revealed an important function of TERT in the mitochondria (Ale-Agha et al., 2021; Ait-Aissa et al., 2022). Evidence is accumulating that re-activation of telomerase can have beneficial anti-aging effects (Nazari-Shafti and Cooke, 2015). In mice, TERT gene therapy delays aging and increases longevity (Bernardes de Jesus et al., 2012; Jaijyan et al., 2022). In a clinically relevant study, induction of TERT decreased DNA damage activation and inflammatory signaling in human colon organoids from patients with inflammatory bowel disease (Chakravarti et al., 2021). Moreover, TERT gene therapy enhances learning pathway networks in human neurons (Shim et al., 2021).
The lack of studies on organ specificity of EC senescence mechanisms and repercussions is in part due to laboratory mice being inappropriate as a model to study replicative senescence (Eckel-Mahan et al., 2020). Laboratory mice are not an ideal model to access consequences of replicative senescence. Humans are born with telomeres in a 10–15 kb range and TERT is inactivated in humans postnatally. In contrast, mice of the commonly used C57BL/6 background are born with telomeres of over 50 kb and continue to express TERT in somatic cells (Kipling and Cooke, 1990). Thus, laboratory mice are more resistant to replicative senescence and stem cell depletion. Indeed, disruption of telomere function in EC has been shown to induce premature senescence (Barinda et al., 2020; Bloom et al., 2023). As we have previously reported, knockout (KO) of TERT in perivascular cells of Pdgfra+ or Pdgfrb+ lineage accelerates the onset of cell senescence in adipose tissue (AT) of mice fed high-calorie diet (HCD), which predisposes them to type-2 diabetes (Gao et al., 2020). Here, we have generated mice with TERT gene knock-out (KO) specifically in EC. We performed genomic analysis of EC from AT and skeletal muscle by both total RNA and single cell (sc) RNA sequencing (RNAseq). We also challenged the mice with HCD to determine its effect on EC senescence and function. Preliminary analysis of data deposited online is presented. Our results suggest that TERT has genome-wide telomere-independent effects on cell transcriptome and physiology (Fossel et al., 2022).
We crossed mice expressing Cre under the control of EC-specific Tie2e promoter (Kano et al., 2003) with TERTfl/fl mice also carrying the mTmG reporter (Muzumdar et al., 2007) to generate mice with TERT gene knock-out (KO) specifically in EC (Figure 1A). Tie2e-cre; TERTfl/fl; mTmG (TERT-EC-KO) and control Tie2e-cre; TERT+/+; mTmG (WT) mice were fed HCD (D12451, 45 kcal% Fat, Research Diets) from 1 to 8 months of age. Cells isolated from subcutaneous AT (SAT), intraperitoneal visceral AT (VAT), as well as combined quadricep and gastrocnemius skeletal muscles (M), were subjected to FACS sorting to isolate mG+ (GFP+) cells (EC) for mRNA extraction by using protocols we described previously (Gao et al., 2018; Gao et al., 2021; Daquinag et al., 2022; Gao et al., 2022). Data quality control confirmed 2.3 × 107 or more reads for all samples. At least 91.5% of reads for all samples were mapped to the mouse genome sequence database. To compare gene expression levels in the tissues, the distribution of gene expression levels and expected number of fragments per kilobase of transcript sequence per millions base pairs sequenced (FPKM) was assessed. Gene expression distribution was found comparable among all samples (Figure 1B). RNA-seq confirmed TERT gene expression loss in mG+ cells of TERT-EC-KO mice. In another experiment, we fed cohorts of TERT-EC-KO and control WT male mice with high-calorie diet from 2 to 7 months of age. Cells were then isolated from SAT and visceral VAT and subjected to single cell RNA sequencing (scRNA-seq) using methodology that we previously described (Gao et al., 2020). Single cell capture and library construction were performed with the Chromium Single Cell 3ʹ Reagent Kit v3.1. Barcoded single-cell gel beads were loaded onto Chromium Next GEM ChipG (PN-1000120). After running on 10X Chromium Single Cell Controller, gel beads-in-emulsion (GEMs) were generated. The barcoded and full-length cDNAs were produced after incubation of the GEMs and amplified via PCR. Library was qualified by Agilent Bioanalyzer 2,100 and quantified by real-time PCR on QuantStudio3. Sequencing was done with Illumina NextSeq 550 System using High Output Kit v2.5 (50,000 reads per cell). The Cell Ranger™ Single Cell Software Suite v.3.1.0 was used to perform bioinformatic analysis. The reads were aligned to the mouse transcriptome reference (mm10, Ensembl 93) with STAR (Dobin et al., 2013). Raw read count tables were analyzed using the Seurat (v3.1.1) pipeline (Butler et al., 2018) on R platform (3.5.2). FindVariableGenes was used to calculate the principal components. Cell clusters were identified using the Shared Nearest Neighbor (SNN) algorithm with a resolution parameter 0.8. UMAP clusters of cells were identified based on the first 10 principal components and feature plots were displayed with the log (raw read count +1) of gene/cell on UMAP.
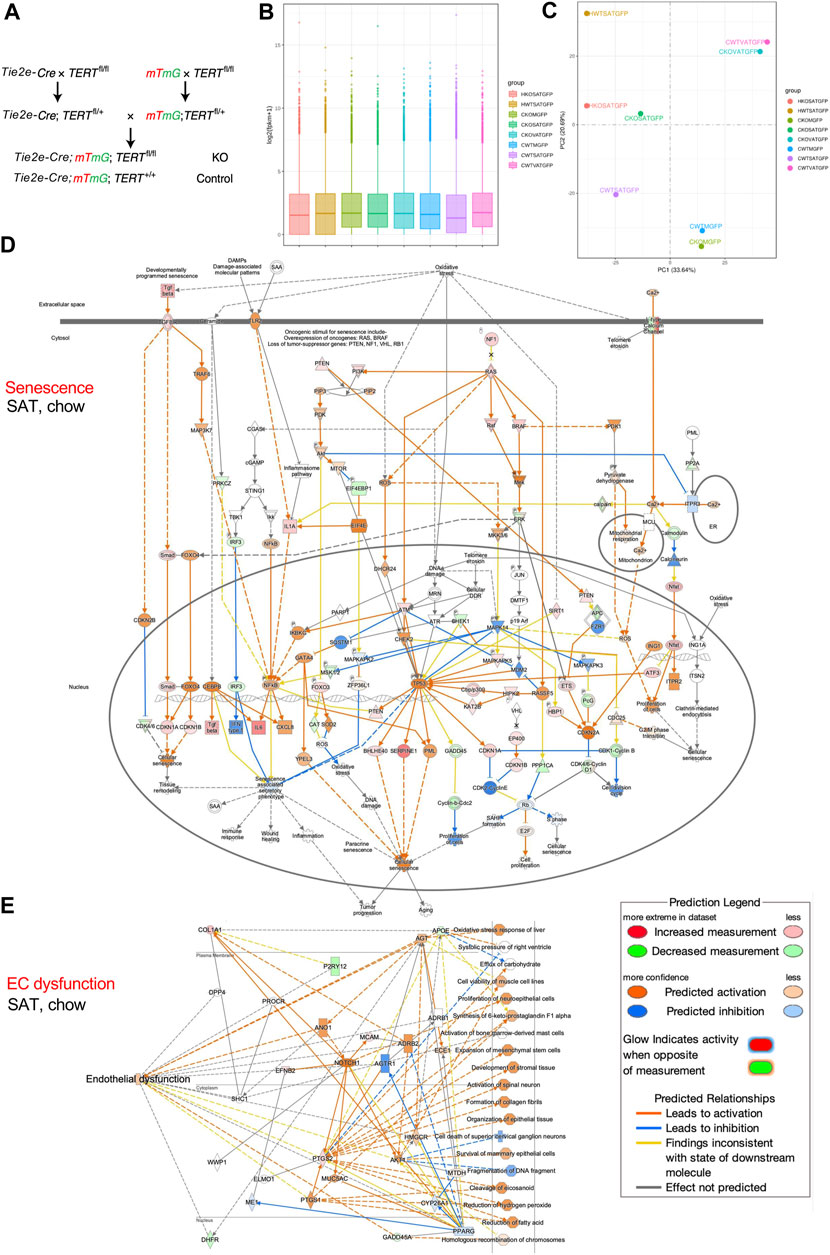
FIGURE 1. TERT knockout in mouse endothelial cells (EC). (A), Breeding scheme to generate mice with mG+ and TERT-EC-KO (fl/fl) or WT (+/+) Tie2+ cells (EC) and other cells mT+. EC senescence and dysfunction caused by TERT loss was assessed in 8-month-old female mice fed chow (C) or HCD (H). Cells isolated from subcutaneous AT (SAT), intraperitoneal visceral AT (VAT), quadricep and gastrocnemius skeletal muscle (M) were subjected to FACS sorting to isolate mG+ cells for mRNA extraction. (B), Gene expression distribution. X axis: mouse groups (also shown on the right). Parameters of box plots are indicated, including maximum, upper quartile, mid-value, lower quartile and minimum. (C), Principal component analysis result (mouse groups shown on the right). (D), IPA analysis focusing on senescence-related pathways identifies genes upregulated in mG+ cells from SAT of TERT-EC-KO mice fed chow compared to mG+ cells from SAT of WT mice fed chow. (E), IPA analysis focusing on EC dysfunction-related pathways identifies genes upregulated in mG+ cells from SAT of TERT-EC-KO mice fed chow compared to mG+ cells from SAT of WT mice fed chow.
Total RNA-seq was performed on mRNA from mG+ cells (EC) of TERT-EC-KO and control WT mice. Principal component analysis (PCA) was used to evaluate inter-sample differences. As expected, this revealed close similarity of gene expression in AT of WT and TERT-EC-KO mice and in skeletal muscle of WT and TERT-EC-KO mice (Figure 1C). HCD feeding resulted in a marked change of gene expression (Figure 1C). Notably, gene expression in SAT from TERT-EC-KO mice fed chow, compared to WT mice fed chow, was found to be more similar to that in SAT of mice fed HCD (Figure 1C).
We then performed Ingenuity Pathway Analysis (QIAGEN IPA) on FACS-sorted AT and muscle EC. Analysis of genes upregulated in mG+ cells of TERT-EC-KO mice demonstrated that a number of key genes implicated in cell senescence were induced in SAT (Figure 1D). This included CDKN1A, CDKN1B, CDKN2A, CDKN2B, TP53, as well as SASP genes IL6 and CXCL8. Induction of pathways mediating senescence was also observed in VAT and skeletal muscle of TERT-EC-KO mice (data not shown). IPA analysis also revealed upregulation of genes specifically implicated in EC dysfunction, including APOE, AGT, ANO1, MYC5AC, and NOTCH1 (Figure 1E). This was also apparent for VAT, and less so for skeletal muscle, of TERT-EC-KO mice (data not shown). IPA analysis of SAT from mice fed HCD revealed a markedly higher level of upregulation of senescence effectors, including CHEK1, p19Arf, and CDKN2A (Supplementary Figure S1A) in additional to upregulation of endothelial dysfunction pathways (Supplementary Figure S1B).
Finally, we assessed scRNAseq data from AT of TERT-EC-KO and WT mice fed HCD. This analysis identified two sub-populations of EC, as well as smooth muscle cells (SMC), adipose stromal cells (ASC), as well as the distinct types of blood cells (Figures 2A, B). UMAP cluster analysis revealed that SAT of TERT-EC-KO mice had a markedly increased presence of certain T-cell subtypes, while the presence of one B-cell subtype was reduced (Supplementary Figure S2). IPA analysis of EC revealed higher expression of senescence-associated genes in EC of TERT-EC-KO AT (Figure 2C). These included ATM, CHEK1, CHEK2, TP53, CDKN2A, as well as SASP genes IL6 and CXCL8.

FIGURE 2. Single cell transcriptomics of adipose cells from EC Tert KO mice. TERT-EC-KO and WT 7-month-old male mice were fed HCD for 5 months prior to cells isolation from SAT and VAT and RNAseq. (A), Integrated heatmap of combined RNAseq data with genes (left) identifying cell clusters designated on top. ACS, adipose stromal cells; RBC, red blood cells; SMC, smooth muscle cells. (B), Regression UMAP clusters of combined KO and WT cells from SAT and VAT generated based on the first 10 principal components displayed with the log (raw read count +1) of gene/cell. (C), Pathways upregulated in SAT EC of EC-TERT KO mice fed chow identified in scRNAseq data by IPA analysis focusing on senescence-related pathways.
The data, being consistent between total RNAseq and scRNAseq experiments, demonstrate that the TERT-EC-KO mice are an appropriate model of EC senescence. These mice can be used to further characterize consequences of endothelial senescence and develop models of aging/disease by subjecting these models to vascular injury insults.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: Total RNA-seq: GEO database via GSE239686 and single-cell RNA-seq: GSE239687.
The animal study was approved by the UTHealth Animal Care and Use Committee. The study was conducted in accordance with the local legislation and institutional requirements.
ZG: Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing–review and editing. YY: Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing–review and editing. YD: Data curation, Formal Analysis, Methodology, Software, Writing–review and editing. ZZ: Data curation, Formal Analysis, Methodology, Software, Writing–review and editing. KE-M: Conceptualization, Funding acquisition, Writing–review and editing. MK: Conceptualization, Data curation, Funding acquisition, Project administration, Resources, Supervision, Writing–original draft.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by the Bovay Foundation, the Levy-Longenbaugh Fund, and grant 1R01DK125922 from the NIH, grant RP180734 from CPRIT.
We are grateful to Xian Chen for the help with RNA sequencing analysis. We thank Joseph Rupert, Alex Daquinag, Rafael Bravo Santos, and John P. Cooke for many helpful discussions.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2023.1295072/full#supplementary-material
Ait-Aissa, K., Norwood-Toro, L. E., Terwoord, J., Young, M., Paniagua, L. A., Hader, S. N., et al. (2022). Noncanonical role of telomerase in regulation of microvascular redox environment with implications for coronary artery disease. Funct. (Oxf) 3, zqac043. doi:10.1093/function/zqac043
Ale-Agha, N., Jakobs, P., Goy, C., Zurek, M., Rosen, J., Dyballa-Rukes, N., et al. (2021). Mitochondrial telomerase reverse transcriptase protects from myocardial ischemia/reperfusion injury by improving complex I composition and function. Circulation 144, 1876–1890. doi:10.1161/CIRCULATIONAHA.120.051923
Barinda, A. J., Ikeda, K., Nugroho, D. B., Wardhana, D. A., Sasaki, N., Honda, S., et al. (2020). Endothelial progeria induces adipose tissue senescence and impairs insulin sensitivity through senescence associated secretory phenotype. Nat. Commun. 11, 481. doi:10.1038/s41467-020-14387-w
Bernardes De Jesus, B., Vera, E., Schneeberger, K., Tejera, A. M., Ayuso, E., Bosch, F., et al. (2012). Telomerase gene therapy in adult and old mice delays aging and increases longevity without increasing cancer. EMBO Mol. Med. 4, 691–704. doi:10.1002/emmm.201200245
Bloom, S. I., Liu, Y., Tucker, J. R., Islam, M. T., Machin, D. R., Abdeahad, H., et al. (2023). Endothelial cell telomere dysfunction induces senescence and results in vascular and metabolic impairments. Aging Cell 22, e13875. doi:10.1111/acel.13875
Butler, A., Hoffman, P., Smibert, P., Papalexi, E., and Satija, R. (2018). Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420. doi:10.1038/nbt.4096
Chakravarti, D., Lee, R., Multani, A. S., Santoni, A., Keith, Z., Hsu, W. H., et al. (2021). Telomere dysfunction instigates inflammation in inflammatory bowel disease. Proc. Natl. Acad. Sci. U. S. A. 118, e2024853118. doi:10.1073/pnas.2024853118
Daquinag, A. C., Gao, Z., Yu, Y., and Kolonin, M. G. (2022). Endothelial TrkA coordinates vascularization and innervation in thermogenic adipose tissue and can be targeted to control metabolism. Mol. Metab. 63, 101544. doi:10.1016/j.molmet.2022.101544
Dobin, A., Davis, C. A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., et al. (2013). STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21. doi:10.1093/bioinformatics/bts635
Eckel-Mahan, K., Ribas Latre, A., and Kolonin, M. G. (2020). Adipose stromal cell expansion and exhaustion: mechanisms and consequences. Cells 9, 863. doi:10.3390/cells9040863
Fossel, M., Bean, J., Khera, N., and Kolonin, M. G. (2022). A unified model of age-related cardiovascular disease. Biol. (Basel) 11, 1768. doi:10.3390/biology11121768
Gao, Z., Daquinag, A. C., Fussell, C., Zhao, Z., Dai, Y., Rivera, A., et al. (2020). Age-associated telomere attrition in adipocyte progenitors predisposes to metabolic disease. Nat. Metab. 2, 1482–1497. doi:10.1038/s42255-020-00320-4
Gao, Z., Daquinag, A. C., Su, F., Snyder, B., and Kolonin, M. G. (2018). PDGFRα/PDGFRβ signaling balance modulates progenitor cell differentiation into white and beige adipocytes. Development 145, 1–13. doi:10.1242/dev.155861
Gao, Z., Daquinag, A. C., Yu, Y., and Kolonin, M. G. (2022). Endothelial prohibitin mediates Bi-directional long chain fatty acid transport in white and Brown adipose tissues. Diabetes 71, 1400–1409. In press. doi:10.2337/db21-0972
Gao, Z., Lu, A., Daquinag, A. C., Yu, Y., Huard, M., Tseng, C., et al. (2021). Partial ablation of non-myogenic progenitor cells as a therapeutic approach to duchenne muscular dystrophy. Biomolecules 11, 1519. doi:10.3390/biom11101519
Gorgoulis, V., Adams, P. D., Alimonti, A., Bennett, D. C., Bischof, O., Bishop, C., et al. (2019). Cellular senescence: defining a path forward. Cell 179, 813–827. doi:10.1016/j.cell.2019.10.005
Jaijyan, D. K., Selariu, A., Cruz-Cosme, R., Tong, M., Yang, S., Stefa, A., et al. (2022). New intranasal and injectable gene therapy for healthy life extension. Proc. Natl. Acad. Sci. U. S. A. 119, e2121499119. doi:10.1073/pnas.2121499119
Kano, A., Wolfgang, M. J., Gao, Q., Jacoby, J., Chai, G. X., Hansen, W., et al. (2003). Endothelial cells require STAT3 for protection against endotoxin-induced inflammation. J. Exp. Med. 198, 1517–1525. doi:10.1084/jem.20030077
Kipling, D., and Cooke, H. J. (1990). Hypervariable ultra-long telomeres in mice. Nature 347, 400–402. doi:10.1038/347400a0
Muzumdar, M. D., Tasic, B., Miyamichi, K., Li, L., and Luo, L. (2007). A global double-fluorescent Cre reporter mouse. Genesis 45, 593–605. doi:10.1002/dvg.20335
Nazari-Shafti, T. Z., and Cooke, J. P. (2015). Telomerase therapy to reverse cardiovascular senescence. Methodist Debakey Cardiovasc J. 11, 172–175. doi:10.14797/mdcj-11-3-172
Park, J. I., Venteicher, A. S., Hong, J. Y., Choi, J., Jun, S., Shkreli, M., et al. (2009). Telomerase modulates Wnt signalling by association with target gene chromatin. Nature 460, 66–72. doi:10.1038/nature08137
Romaniuk, A., Paszel-Jaworska, A., Toton, E., Lisiak, N., Holysz, H., Krolak, A., et al. (2019). The non-canonical functions of telomerase: to turn off or not to turn off. Mol. Biol. Rep. 46, 1401–1411. doi:10.1007/s11033-018-4496-x
Sahin, E., and Depinho, R. A. (2010). Linking functional decline of telomeres, mitochondria and stem cells during ageing. Nature 464, 520–528. doi:10.1038/nature08982
Shim, H. S., Horner, J. W., Wu, C. J., Li, J., Lan, Z. D., Jiang, S., et al. (2021). Telomerase reverse transcriptase preserves neuron survival and cognition in alzheimer's disease models. Nat. Aging 1, 1162–1174. doi:10.1038/s43587-021-00146-z
Stewart, S. A., Hahn, W. C., O'connor, B. F., Banner, E. N., Lundberg, A. S., Modha, P., et al. (2002). Telomerase contributes to tumorigenesis by a telomere length-independent mechanism. Proc. Natl. Acad. Sci. U. S. A. 99, 12606–12611. doi:10.1073/pnas.182407599
Xu, Q., Mojiri, A., Boulahouache, L., Morales, E., Walther, B. K., and Cooke, J. P. (2022). Vascular senescence in progeria: role of endothelial dysfunction. Eur. Heart J. Open 2, oeac047. doi:10.1093/ehjopen/oeac047
Keywords: endothelial, senescence, telomerase, TERT, knockout, mouse, gene expression
Citation: Gao Z, Yu Y, Dai Y, Zhao Z, Eckel-Mahan K and Kolonin MG (2023) Gene expression in mice with endothelium-specific telomerase knockout. Front. Cell Dev. Biol. 11:1295072. doi: 10.3389/fcell.2023.1295072
Received: 15 September 2023; Accepted: 28 November 2023;
Published: 07 December 2023.
Edited by:
Chunsik Lee, Sun Yat-sen University, ChinaReviewed by:
Erica Salvati, National Research Council (CNR), ItalyCopyright © 2023 Gao, Yu, Dai, Zhao, Eckel-Mahan and Kolonin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mikhail G. Kolonin, bWlraGFpbC5nLmtvbG9uaW5AdXRoLnRtYy5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.