 Giampaolo Placidi1,2
Giampaolo Placidi1,2 Clara Mattu
Clara Mattu Gianluca Ciardelli
Gianluca Ciardelli Carlo C. Campa
Carlo C. Campa- 1Italian Institute for Genomic Medicine, Candiolo, Italy
- 2Department of Mechanical and Aerospace Engineering, Politecnico di Torino, Turin, Italy
- 3Chemical-Physical Processes, National Research Council (CNR-IPCF), Pisa, Italy
- 4Candiolo Cancer Institute, FPO-IRCCS, Candiolo, Italy
Over the past years a growing number of studies highlighted the pivotal role of intracellular trafficking in cell physiology. Among the distinct transport itineraries connecting the endocytic system, both internalization (endocytosis) and recycling (endocytic recycling) pathways were found fundamental to ensure cellular sensing, cell-to-cell communication, cellular division, and collective cell migration in tissue specific-contexts. Consistently, the dysregulation of endocytic trafficking pathways is correlated with several human diseases including both cancers and neurodegeneration. Aimed at suppress specific intracellular trafficking routes involved in disease onset and progression, huge efforts have been made to identify small molecule inhibitors with suitable pharmacological properties for in vivo administration. Here, we review most used drugs and recently discovered small molecules able to block endocytosis and endocytic recycling pathways. We characterize such pharmacological inhibitors by emphasizing their target specificity, molecular affinity, biological activity and efficacy in both in vitro and in vivo experimental models.
Introduction
Endocytic membrane trafficking plays an essential role in delivering both solute molecules and membrane components (e.g., lipids and proteins) to various intracellular destinations (Doherty and McMahon, 2009; Haucke, 2015). Conceptually, endocytic trafficking routes are relatively simple, with the main pathways carrying either to degradation in lysosomes or to recycle back to the plasma membrane. However, genetic redundancy and pleiotropy profoundly impact on molecular organization of membrane trafficking, thus limiting identification of both specialized trafficking itineraries and pivotal protein interactions (Yarwood et al., 2020).
Endocytic membrane trafficking is a ubiquitous process in eucaryotic organisms. Non-etheless, not all cells respond to perturbation of membrane trafficking machinery in the same way (De Matteis and Luini, 2011; Yarwood et al., 2020; García-Cazorla et al., 2022). This is posited to depend on: 1) the abundance and the degree of functional redundancy of membrane trafficking components, and 2) the relevance of the transported cargo for cellular functions and tissue homeostasis. For instance, the nervous system is susceptible to disruption of endocytic genes involved in both endocytic recycling and autophagy. This is due to both the low proliferating activity and the elevated speed of neurotransmitters secretion/internalization that characterize neuronal cells (Schreij et al., 2016). Consequently, impairment in endocytic trafficking pathways significantly affect behaviour and function of the nervous tissue, often resulting in neurological disorders including frontotemporal dementia (FTD), Alzheimer and Parkinson diseases (Harold et al., 2009; DeJesus-Hernandez et al., 2011; Vilariño-Güell et al., 2011; Vilariño-Güell et al., 2014). Similarly, alteration in endocytic recycling significantly impact in cells with elevated secretory activity, such as pancreatic beta cells. In particular, mutations in AS160, an endocytic recycling regulator, impairs the translocation of the glucose transporter GLUT4, thus leading to defective glucose blood clearance and hence muscle insulin-resistance and type 2 diabetes (Karlsson et al., 2005; Mîinea et al., 2005; Moltke et al., 2014). Despite these considerations, the employment of membrane trafficking alterations to predict both type and status of a disease is far from being achieved.
The therapeutic targeting of membrane trafficking pathways might be used to increase both delivery and efficacy of currently employed therapeutics for cancer, respiratory disorders and, concomitantly, to provide novel strategies for the treatment of over 300 Mendelian genetic diseases (Chew et al., 2020; Kang et al., 2020; Riva et al., 2020; García-Cazorla et al., 2022). Unfortunately, the complex organization of the endomembrane system limits the identification of membrane trafficking inhibitors using phenotype-based screening strategies. In parallel, it remains still difficult to identify druggable molecular mechanisms pivotal for transport of specific molecular cargoes (De Matteis et al., 2013; Mishev et al., 2013).
This review provides an overview of main drugs controlling regulatory functions of endocytic proteins. This is particularly important in view of recent progress into endocytic trafficking field. To get insights into mechanisms characterizing novel transport itineraries that are not covered by this review, we remand readers to these excellent works (Redpath et al., 2020; Renard and Boucrot, 2021; Sigismund et al., 2021; Gilleron and Zeigerer, 2023).
Targets for endocytic recycling
The internalization and their transport back to the plasma membrane of both solute molecules and membrane components (e.g., lipids and proteins) is carryout by both the endocytic and the recycling membrane trafficking machineries, respectively. Every transport itinerary requires the budding, scission and transport of vesicles from the donor compartment and their subsequent targeting, tethering, docking and fusion to the acceptor membrane. Notably, these trafficking steps are controlled by protein subsets that, according to their roles during the transport process, can be organized into specific functional modules. Below, we provide description of building blocks used by both endocytic and recycling pathways. This list encompass both adaptors, coats, shaping, fission, small GTPase, kinase, tethering and fusion proteins involved in endocytic recycling (Liberali et al., 2008).
Adaptor module
Both AP180, β-arrestin, NUMB, HIP1, DAB2, ARH, EPSIN1, EPS-15 proteins and components of the adaptor complexes AP1, AP2, AP3 play a central role in cargo recognition and membrane-coat complex assembly during endocytic recycling (Reider and Wendland, 2011). These adaptors are composed by different protein domains which mediate interactions with membrane lipids (e.g., phosphatidylinositol 4,5-bisphosphate), coats (e.g., clathrin) and cargo proteins (e.g., G-protein coupled receptors, tyrosine kinase receptors). Notably, adaptor recruitment to cargoes can be controlled by post-translational modifications (e.g., phosphorylation), thus ensuring internalization of signalling receptors following agonist stimulation.
Coat module
This functional module includes clathrin (both light and heavy chains), caveolin and flotillin, each of which is recruited to both plasma membrane and endosomes by several endocytic adaptor subsets. Specific protein-binding motifs enable both clathrin polymerization and caveolin/flotillin oligomerization into higher-order structures (i.e., basket, disk-shaped or tetramers, respectively), thus helping the budding of vesicles from donor membranes (Van Jaarsveld et al., 1981; Chaudhary et al., 2014; Kononenko et al., 2014; Watanabe et al., 2014; Shvets et al., 2015; Han et al., 2020; Porta et al., 2022; Singh et al., 2022).
Shaping module
Bin/amphiphysin/Rvs (BAR) domain-containing proteins play a prominent role in membrane remodelling in response to protein surface density, membrane tension, or membrane shape alterations. Arfaptin, Amphiphysin, Sortin Nexins, Endophilin, FCHo proteins are the most pervasive membrane-shaping regulators controlling the invagination of both cell surface and endocytic membranes. In addition to the BAR domain, such membrane remodelling proteins can encode a small GTPase-regulatory region. For instance, ASAP and ACAP proteins display an Arf-GAP domain which allow functional coupling between BAR-mediated membrane sculpting and actin polymerization, specifically. Lastly, while the mentioned proteins control endocytic vesicle formation on both endosome and plasma membrane, a different BAR domain-containing proteins family (e.g., MIM, IRTKS, IRSp53) evolved to direct the generation of plasma membrane protrusion (Simunovic et al., 2015; Renard et al., 2018; Simunovic et al., 2019).
Fission module
The membrane fission module relies on mechanoenzymatic machineries that bind phosphatidylinositol-enriched membranes to favour membrane constriction through a GTP-dependent mechanism. Dynamin proteins (e.g., DNM1, DNM2, DNM3) are GTPase enzymes that hydrolyse GTP to prompt vesicle fissions in endocytic membrane-bound compartments. DNM1 and DNM3 are enriched in brain and they both mediate retrieval of synaptic vesicle membranes, an event that occurs in concomitance with exocytic fusion and cargo recycling to the cell surface. In contrast, DNM2 is ubiquitously expressed and mediates endocytic uptake (Ferguson and De Camilli, 2012; Wu et al., 2014; Lee et al., 2016; Renard et al., 2018; Jimah and Hinshaw, 2019; Imoto et al., 2022).
Rab GTPase modules
The RabGTPase module comprises over 60 distinct genes in the human genome. Rab GTPases are molecular switches that, by cycling between an active and inactive state, they recruit, on membranes, specific molecular effectors. Through their effectors, Rab GTPases regulate vesicle formation, vesicle movement along actin and tubulin networks, and membrane fusion. Notably, specific Rab GTPases subsets are localized to different membrane-bound compartments and hence they act as signpost to recognize transport itineraries in mammalian cells. In this context, RAB4 family (A, B, C), RAB5 family (A, B, C), RAB13, RAB 20, RAB21, RAB22 family (A, B), and RAB23 are restricted to early endosomes, while the recycling compartment stained positive for RAB3 family (A, B, C, D), RAB8 family (A, B), RAB10, RAB11 family (A, B, C/RAB25), RAB14, RAB15, RAB17, RAB35 (Wandinger-Ness and Zerial, 2014).
Kinase module
This module comprises both protein and lipid kinases controlling phosphorylation, and hence activation, of either protein or lipid-signalling, in response to cell’s environmental changes. As an example, the phosphorylation of Rab GDP dissociation inhibitor (GDI) by p38 (MAPK11, -13, -14), a stress-activated protein kinase, causes Rab5 cytosolic sequestration and endocytosis blockage. As a further example, dynamins are substrates for both receptor-activated kinases (e.g., SRC, casein kinase, GSK3) and cell cycle regulated kinases (e.g., CDK5, DYRK1), thus providing inducible control in vesicle fission. In parallel, the lipid kinases-mediated generation of signalling lipids promotes maturation of endocytic membranes through the recruitment of both membrane-associated and cytosolic proteins. In this context, phosphatidylinositol 4-phosphate 5-kinase (PI4P5K) and phosphatidylinositol 5-phosphate 4-kinase (PI5P4K) enzymes generates phosphatidylinositol 4,5 biphosphate [PtdIns(4,5)P2], the major signalling lipid involved in endocytosis and essential for plasma membrane association of AP2 complex (Cavalli et al., 2001; Clayton et al., 2010; Liberali et al., 2014; Wallroth and Haucke, 2018; Perez Verdaguer et al., 2021).
Tethering module
Two distinct molecular classes control long-distance vesicle recognition to acceptors compartment: coiled-coil proteins and multi-subunit complexes. EEA1 and Rabphilin-3A are prototypical coiled-coil proteins involved in fusion of endocytic vesicles to early endosomes and plasma membrane, respectively. In parallel, multi-subunit complexes such as CORVET promotes tethering and fusion of Rab5/Vps21-positive membranes, while Exocyst, EARP, FERARI control the fusion of endocytic vesicles to plasma membrane (Peplowska et al., 2007; Balderhaar et al., 2013; Schindler et al., 2015; Ahmed et al., 2018; Solinger et al., 2022).
Fusion module
The fusion module comprises SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor)-motif containing proteins that mediate vesicles fusion to acceptor membranes. Previously named as v- and t- SNARE, due to their localization to either vesicle or target membranes, while currently classified into Qa-, Qb, Qc- and R-SNARE based on their sequence features, SNARE proteins bind to each other to form a parallel four-helix bundle. This helix structure, bridges and ultimately fuses vesicle membranes. The endocytic fusion machinery requires Syntaxin (Stx), VAMP and SNAP-25 protein homologues such as: Stx13, Stx6, Stx2, Stx4, Vti1a, VAMP4, and VAMP7. Targeting specificity is determined by the combinatorial assembly of the over 35 different SNAREs. Nonetheless, recent reports indicate that some SNAREs can functionally substitute for each other if they belong to the same subfamily (Jahn and Scheller, 2006; Koike and Jahn, 2017; Koike and Jahn, 2019).
Small molecules targeting the endocytic machinery
Internalization and delivery of both protein and lipids to intracellular membrane-bound compartments are fundamental for cell physiology (Holter, 1959; Harding et al., 1983; Sigismund et al., 2012; Shafaq-Zadah et al., 2020). Such endocytic itineraries are classified based on the molecular machinery employed, principally either clathrin or dynamin -based. To date, five major endocytic trafficking routes are identified: 1) clathrin-mediated endocytosis (CME; clathrin and dynamin dependent), 2) caveolae-mediated endocytosis (CavMe, clathrin independent and dynamin dependent), 3) fast endophilin-mediated endocytosis (FEME, a clathrin-independent but dynamin-dependent pathway), 4) clathrin-independent carrier (CLIC)/glycosylphosphatidylinositol-anchored protein enriched early endocytic compartment (GEEC) endocytosis (clathrin and dynamin independent), 5) macropinocytosis and 6) phagocytosis (Cardoso et al., 2021). Due to the fundamental role of both coat and pinch-off proteins, most efforts have been directed towards the discovery of small molecules targeting both clathrin and dynamin pathways. However, beside these rationally-designed compounds, many drugs, with unknown target specificity and able to interfere with trafficking pathways, are still employed in membrane trafficking filed. For instance, Genistein (4′,5,7-trihydroxyisoflavone), a molecule that belong to tyrosine kinase inhibitors, hamper caveolae-mediated endocytosis and partially clathrin-mediated endocytosis by an unknown mechanism (DeLouise, 2012). In addition, Methyl-b-cyclodextrin (MbCD), a cyclic heptasaccharid, depletes cholesterol from cell membrane causing lipid-rafts/caveolin depletion and the subsequent perturbation of both clathrin- and caveolae-mediated pathways (Vercauteren et al., 2010).
In the section below, we describe main endocytic inhibitors characterized by elevated target specificity. In this summary, we not include either molecules or treatment, such as potassium depletion or sucrose treatment (Rennick et al., 2021), responsible for internalization defects by unknown mechanisms of function (Supplementary Figure S1; Table 1).
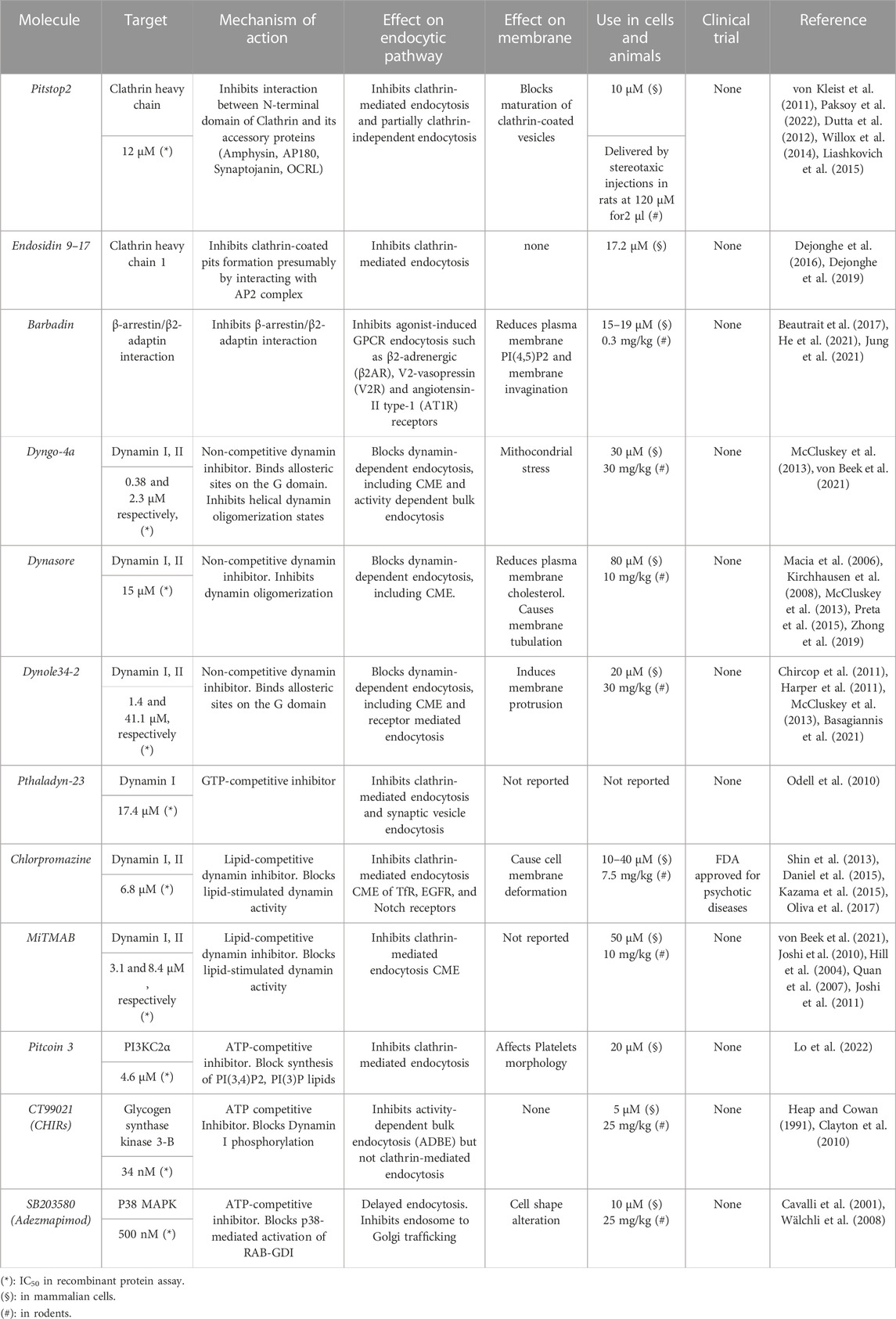
TABLE 1. Small molecules inhibitors targeting endocytic molecules.
Clathrin heavy chain inhibitors
Clathrin heavy chain (CHC) is a self-assembling protein that coats transport vesicles during their endocytic sorting. The assembly of clathrin coat requires interaction between CHC and clathrin-associated sorting proteins (CLASPs) through the CHC terminal domain (TD) (Schmid and McMahon, 2007; Ranjan et al., 2017). CLASPs comprise many endocytic proteins such as the AP2 complex, AP180, CALM, Eps15, Amphiphysin involved in internalization of many growth factor receptors (Lemmon and Traub, 2012). CLASPs localization guides clathrin basket formation at distinct cellular compartments, allowing directionality in membrane trafficking flow. As an example, clathrin-coated vesicles (CCVs), programmed for being delivered from cell surface and internal organelles, are assembled on both endosomes and plasma membrane by specific adaptor proteins such as AP1B1 (designated β1) and AP2B2 (β2), respectively. Based on the important role played by CHC and CLASPs binding for CCVs assembly, huge efforts were made to identify compounds able to block the interaction between CHC’s TD and CLASPs, of which Pitstop2 and Endosidin9-17 are the most noticeable.
Pitstop2 associates with the clathrin TD and obstructs binding of accessory proteins involved in both maturation and disassembly of CCP such as Amphysin, AP180, Synaptojanin, OCRL (von Kleist et al., 2011). In cells, Pitstop2 blocks clathrin-mediated endocytosis and generates enlarged endosomes when used in rodents and specifically at the calyx of Held, a synapse optimized for high frequency synaptic transmission in the auditory brainstem (Paksoy et al., 2022). Nonetheless, to date several clathrin-independent effects were reported for Pitstop2 (Dutta et al., 2012).
A similar mechanism of function is employed by Endosidin9 (ES9), a small molecule that induces, after short exposure (∼30 min), both clathrin and AP2 mislocalization in human, plants and fruit-fly cells (Dejonghe et al., 2016). Notably, ES9-17, a ES9 more potent derivative, was demonstrated to block endocytosis in Arabidopsis Thaliana, a model system resistant to Pitstop2 inhibition due to an aminoacidic substitution (residue 80) in plant’s clathrin heavy chain isoform.
Caveolin inhibitors
Caveolins are integral membrane proteins that play an important structural role in caveolae-mediated endocytosis, an endocytic process mediating the internalization of Cholera toxin, Ebola, Hepatitis B, Japanese encephalitis, human coronaviruses -229E and -OC43 (Xing et al., 2020; Parton et al., 2021). Three distinct caveolin isoforms were identified (Cav1, Cav2, Cav3) and found differentially expressed in human tissues. In particular, Cav1 is enriched in brain, skeletal muscle, liver, stomach, lung, kidney and heart. Cav2 is predominantly expressed in endothelial cells, smooth muscle cells, and fibroblasts, while Cav three is found in muscular tissue (Scherer et al., 1997; McMahon et al., 2009). Caveolins are composed by a transmembrane region flanked by two (N-, C-) terminal domains, each of them is exposed to cell’s cytoplasmic side and involved in caveolin post-transaltional regulation. The transmembrane portion comprises an oligomerization domain and a scaffolding domain which control, respectively, caveolin oligomerization and cholesterol binding (Parton and del Pozo, 2013; Porta et al., 2022). Both these protein-protein and protein-lipid interactions are required for caveolin-mediated caveolae formation. The oligomerization domain mediates the aggregation of caveolin in 14–16-unit oligomers (Porta et al., 2022), while the scaffolding portion allows both membrane insertion into cholesterol-enriched membrane regions and interaction with cholesterol-binding proteins (Krishna and Sengupta, 2019).
Currently, two distinct strategies are available to block caveolin activity during endocytosis: cholesterol inhibition and caveolin-oligomer disruption. The first employs Methyl-b-cyclodextrin, FilipinIII and Nyastatin to deplete cholesterol and ergosterol from cellular membranes, while the second uses WL47, a synthetic peptide, to reduce in-vitro caveolin oligomers assembly (Plummer and Manchester, 2013; Gilliam et al., 2016).
CLASPs inhibitors
The interaction between clathrin-associated sorting proteins (CLASPs) and clathrin guides the internalization of cell surface proteins. Both β-arrestins and AP2 complex bind clathrin and both are required to induce internalization of cell surface receptors. Specifically, β-arrestins are activated downstream G-protein coupled receptors (GPCRs). Following sustained agonist stimulation, β-arrestin are recruited at the plasma membrane by phosphorylated-activated GPCRs. This event promotes functionally uncoupling of the activated receptors from their heterotrimeric G proteins (Lohse et al., 1990). Subsequently, the association of β-arrestins to AP2 induces GPCR internalization and trafficking towards endosomes for their degradation/recycling.
Barbadin is a small molecule that targets the contact interface between β-arrestin and AP2. Barbadin blocks GPCR internalization without affecting trafficking of Transferrin receptor (TfR), a non-GPCR that interacts directly with AP2 in a β-arrestin independent manner (Jing et al., 1990). In a preclinical experiment, Barbadin was demonstrated to potentiate the effects of Lorcaserin, a serotonin 2C receptor (5-HT2CR) selective agonist. Barbadin treatment inhibits 5-HT2CR internalization after lorcaserin stimulation, maintaining proper proopiomelanocortin (POMC) neuron responses to serotonin-agonist challenge in vivo, ultimately leading to appetite reduction and weight gain in mouse models (He et al., 2021).
Dynamin inhibitors
Dynamins are large GTPase proteins that mediate membrane fission and fusion during endocytosis, recycling, and organelle biogenesis (Jimah and Hinshaw, 2019). Three distinct dynamin isoforms were identified and found differentially expressed in human tissues. Dynamin I is mostly expressed in brain-related tissues, Dynamin II is ubiquitous, whereas Dynamin III is enriched in brain, lungs, and testis (Ferguson and De Camilli, 2012). All dynamin isoforms contain a GTPase (G) domain involved in GTP binding and hydrolyses, a pleckstrin-homology (PH) domain which recognizes PtdIns(4,5)P2, a membrane lipid, and a GTPase effector domain (GED) that allows oligomerization and stimulation of Dynamin GTPase activity (Jimah and Hinshaw, 2019). Three distinct classes of small molecule inhibitors were developed to differentially target either dynamin’s oligomerization, GTPase activity or lipid-stimulated GTPase activity (Table 1).
Dyngo-4A and Dynasore are small molecules that, by binding allosteric sites in the Dynamin G domain, block dynamin oligomerization into either ring-like or helical structures, thereby impeding cooperative GTP hydrolysis, membrane tubulation and hence clathrin-mediated endocytosis, as demonstrated by reduction of TfR uptake (Tuma and Collins, 1994; Hinshaw and Schmid, 1995; Macia et al., 2006; McCluskey et al., 2013). A different mechanism is adopted by Chlorpromazine, a phenothiazine-derived antipsychotic drug and MiTMAB. These compounds block lipid-stimulated Dynamin GTPase activity by competing with lipid binding (Daniel et al., 2015). In parallel, competitive inhibition of GTP nucleotide binding is used by Pthaladyn-23 (Odell et al., 2010).
One of the potential employments of dynamin inhibitors is the boost of antibody-dependent cellular cytotoxicity (ADCC) response for cancer treatment. By blocking internalization of cell surface antigens, dynamin inhibitors increase target receptor availability for antibody binding. Thus, retention of antigen-antibody complexes enhances recognition of cancer cells by natural killer (NK) cells. These findings were corroborated in preclinical experiments by using Dyngo-4a and prochloropyrazine, a less toxic chlorpromazine analogue. These dynamin inhibitors were found effective in combination with cetuximab, trastuzumab and avelumab, three distinct clinical-approved antibody-based treatment targeting EGFR, HER2 and PD-L1 receptors, respectively (Chew et al., 2020). Although impressive results were obtained by these compounds, dynamin inhibitors are not isoform-selective and hence not able to block specific trafficking pathways. Since dynamin isoforms act on distinct cellular districts (Gray et al., 2003), major concerns regarding their on-target toxicity during endocytic recycling are present.
Kinase inhibitors
Kinases are enzymes that catalyse the transfer of phosphate groups to either proteins or lipids. As a result, kinases activity is rarely limited to endocytic pathways, often resulting in perturbation of several cellular processes (Roskoski, 2015). Nonetheless, the kinase-induced phosphorylation mechanism permits the endocytic machinery to react to changing cellular demands and hence ensuring appropriate cell’s response to a fluctuating environment (Roskoski, 2015).
CT99021 is an inhibitor of Glycogen synthase kinase (GSK3B), a serine-threonine kinase regulating the phosphorylation of more than 100 substrates including both cell proliferation and membrane trafficking pathway components (Chen et al., 2007; Beurel et al., 2015; Hermida et al., 2017; Zheng and Conner, 2018a). CT99021 blocks GSK3B-mediated phosphorylation of Dynamin I at Ser-774 resulting in activity-dependent bulk endocytosis (ADBE) defects (Clayton et al., 2010). Notably, CT99021-mediated GSK3B inhibition is currently used in preclinical models to investigate the potential of endocytosis blockade in depression treatment, spatial learning and memory amelioration (Smillie et al., 2013; Lee et al., 2021). In this context, synaptic injury and cognitive decline can be rescued by SB203850, a p38 MAP kinases (MAPK11-14) inhibitor, that delay Rab5-mediated endocytosis through RAB-GDI inactivation (Cavalli et al., 2001; Wälchli et al., 2008; Yu et al., 2018).
Differently to other protein kinases, lipid kinases catalyse the ATP-dependent transferase reaction on lipid membranes, thereby localizing signalling reactions on membrane surfaces (Feng and Yu, 2021; Yoshioka, 2021; Burke et al., 2022). Therefore, alteration of lipid kinases activity is rarely associated with either inhibition or mistargeting of specific trafficking routes (Ronan et al., 2014). Nonetheless, Pitcoin3, an ATP-competitive PI3KC2α inhibitor, was recently developed to block clathrin-mediated endocytosis. By reducing plasma membrane and endosomal phosphoinositide’s content (i.e., PtdIns(3,4)P2 and PtdIns(3)P) Pitcoin3 mimicks the effects of enzyme’s genetic loss (Posor et al., 2013; Campa et al., 2018; Yoshioka, 2021; Lo et al., 2022).
Small molecules targeting the endocytic recycling machinery
Delivery of endocytosed plasma membrane components to various intracellular compartments is achieved by a multitude of trafficking pathways. Based on the final cargo’s destination, such intracellular trafficking routes are classified in the degradative, retrograde and recycling routes (Scott et al., 2014). In this section, we describe lysosomotropic agents (e.g., bafilomycin A1, NH4Cl, chloroquine, ionomycin, nigericin, monensin) and small molecules affecting endocytic recycling (Johnson et al., 1993; Scott and Gruenberg, 2011) (Supplementary Figure S2; Table 2).
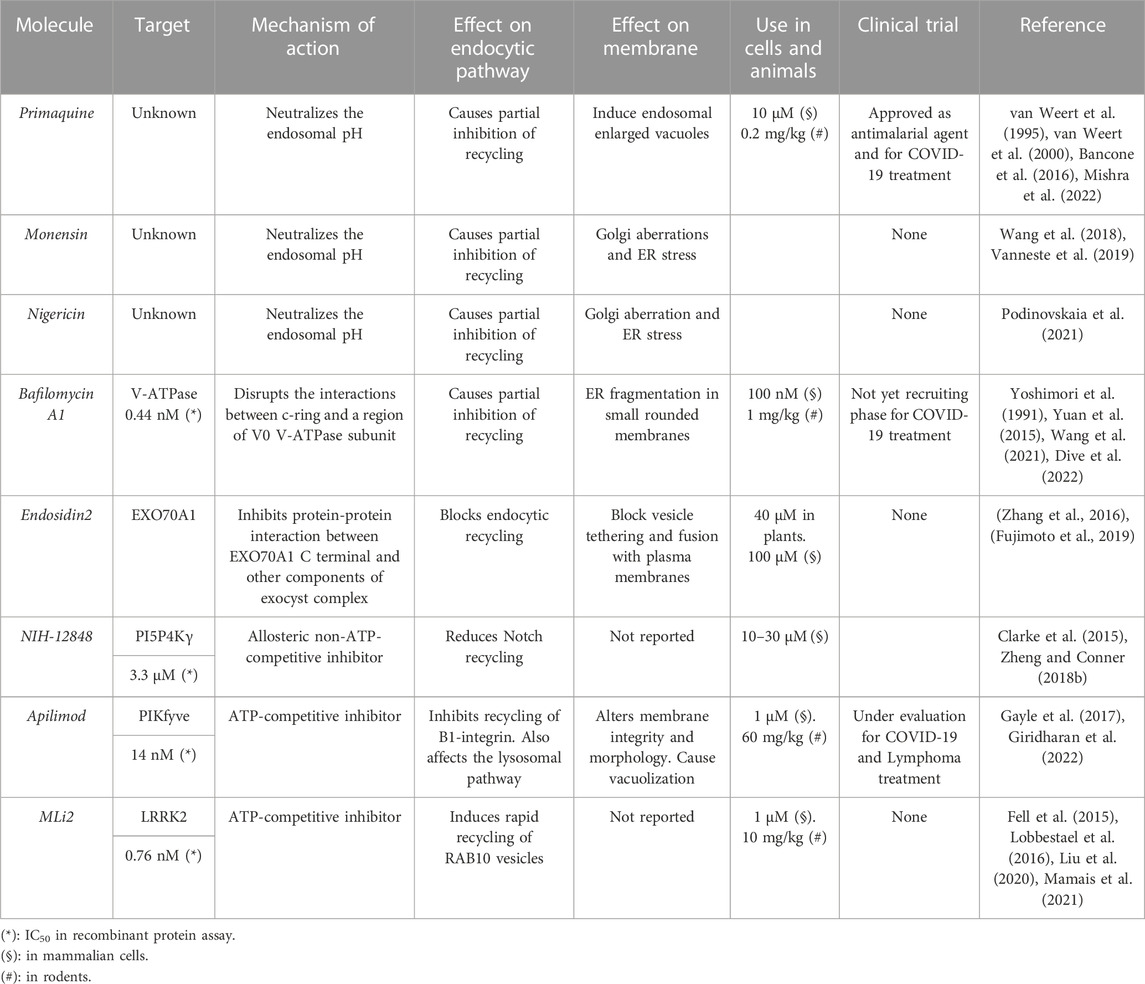
TABLE 2. Small molecules inhibitors targeting specific recycling modules.
Lysosomotropic inhibitors
Many compounds belonging to the family of antimalarial agents, such as chloroquine and primaquine, were introduced as recycling inhibitors (van Weert et al., 1995; van Weert et al., 2000). These small molecules are weak bases, that once protonated, accumulate in endosomes causing both neutralization and deacidification of endosomal pH, thus resulting in endosomal recycling inhibition.
Primaquine and chloroquine are antiviral and antiparasite compounds used to block Zika, Ebola and malaria-related infections (Naghipour et al., 2020; Persoons et al., 2021). Moreover, primaquine and Chloroquine were recently under evaluation for therapeutic treatment of autoimmune diseases (Rainsford et al., 2015). As generic medications, huge efforts have been made to repurpose Chloroquine and Primaquine (and all others lysosomotropic drug) for cancer treatment. However, results coming from these studies are puzzling, thus limiting the employment of lysosomotropic drugs in both basic and clinical research (Ashley et al., 2014). Similar consideration must be made for BafilomycinA1, a macrolide antibiotic that blocks receptor recycling by inhibiting V-ATPase proton pump (Yoshimori et al., 1991). The V-ATPase consists of two main multisubunit complexes named V0 and V1. Structural studies and biochemical experiments defined that BafilomycinA1 causes steric hindrance between elements of V0 subunit, resulting in inhibition of V-ATPase activity (Wang et al., 2021).
Exocyst complex inhibitor
The Exocyst complex mediates the tethering of secretory vesicles to plasma membrane, a trafficking step that anticipate the SNARE-mediated vesicle fusion (Mei and Guo, 2018). This multisubunit complex is composed by Sec3, Sec5, Sec6, Sec8, Exo84, and Exo70 (Martin-Urdiroz et al., 2016). Endosidin2, an exocyst complex inhibitor, interacts with Exo70 causing reduction of transferrin recycling in mammalian cells (Zhang et al., 2016).
Kinase inhibitors
LRRK2 is a serine/threonine-protein kinase activated by oxidative, endolysosomal and autophagic stressors. LRRK2 phosphorylates a broad range of endocytic recycling regulators including the small GTPase Rab8A and Rab10 (Steger et al., 2017; Dhekne et al., 2018; Bonet-Ponce et al., 2020; Herbst et al., 2020; Kuwahara et al., 2020). Selective inhibition of LRRK2 by MLi2, an ATP-competitive inhibitor induces rapid recycling of RAB10 vesicles, thus reducing endocytic defects caused by hyperactivation of LRRK2 signalling in a murine model of Parkinson disease (Scott et al., 2017).
In contrast to the protein kinases, members of the lipid kinase family have less structural similarity in enzyme’s active site, simplifying generation of isoform-selective inhibitors (Roskoski, 2016). As an example, reduction of 5′ phosphorylated phosphoinositide signalling can be obtained using both competitive and non-competitive ATP inhibitors. Apilimod, an ATP competitive inhibitor, and NIH-12848, a non-competitive ATP inhibitor, reduce endocytic recycling of internalized cargoes by blocking the activity of PIKfyve and PI5P4Kγ lipid kinases, respectively (Clarke et al., 2015; Sbrissa et al., 2018; Baranov et al., 2020). Despite these findings, most studies are currently focused on the role of both Apilimod and NIH-12848 in endolysosomal regulation and its impact in immune-regulatory associated diseases (Gayle et al., 2017; Poli et al., 2021). Whether the endocytic recycling and the lysosomal trafficking pathways are linked by the 5′ phosphorylated phosphoinositide signalling is still unclear.
Small GTPase inhibitors
Small GTPases are enzymes that catalyse the hydrolysis of guanosine triphosphate (GTP) and the subsequent generation of guanosine diphosphate (GDP). Activation of small GTPase is controlled by guanine nucleotide exchange factors (GEFs) which promote the release of GDP and the subseqeunt GTP loading. Conversely, GTPase-activating proteins (GAPs) bind to activated small GTPase to stimulate their GTPase activity. Small GTPase targeting might be used to control main steps involved in membrane trafficking including vesicle generation, transport, and fusion. Historically, the evidence of small GTPase targeting potential in membrane transport was demonstrated by Brefeldin A (BFA), a fungal toxin able to block ARF small GTPase activity (Helms and Rothman, 1992; Reiner and Lundquist, 2018). BFA is a protein-protein interaction inhibitor that disrupts the interaction between Arf1 and GBF1, an Arf GEF (Rouhana et al., 2013). Optimization of this protein-protein inhibitor has led to the development of LG186. LG186 is a selective Arf1-GBF1 interaction inhibitor, used to block, in addition to common Golgi-localized ARF1 trafficking pathways, CLIC/GEEC internalization (Klausner et al., 1992; Boal et al., 2010; Sathe et al., 2018).
However, the targeting of small GTPase by small molecule inhibitors remains difficult (Gray et al., 2020). At this regard, novel approaches, that use synthetic peptides and covalent small molecules, are now employed to prevent the interaction between small GTPase and their effector proteins and hence small GTPase signalling (Ali et al., 2019). As an example, the stapled peptide RFP14 blocks the interaction between Rab25 and FIP2, a Rab25 effector protein, causing a decrease of RAB25-driven cell proliferation (Mitra et al., 2017). A similar approach was employed to achieve RAB27 pathway inhibition, a key player in exosome secretion. Nexinhib20, a small molecule targeting the interaction between Rab27a and its effector JFC1, was found to regulate exocytosis-dependent neutrophil’s function and exosome secretion both in vitro and in vivo models of inflammation, a context in which exosome secretion and recycling are pivotal pathways for cancer disease progression (Johnson et al., 2016). Lastly, by taking advantage of two residues, that are unique to Rab27A and Rab27B, among the over 60 Rab family proteins (i.e., C123 and C188), a recent report identifies two covalent ligands (A01 and B01) that react preferentially with these cysteines, paving the way for future development of covalent RAB27A signalling inhibitors (Jamshidiha et al., 2022).
Conclusion
The pharmacological targeting of both endocytosis and endosomal recycling pathways has the potential to improve therapeutic treatment of Mendelian genetic diseases, parasites/virus infections, neurodegeneration and cancers. However, due to the pleiotropic behaviour of endocytic genes, it remains difficult to hijack specific transport itineraries, as observed for Endosidin 2, a Exo70 small molecule inhibitor, that affects both constitutive exocytosis and receptor recycling. In parallel, drug repurposing has been explored to evaluate the therapeutic potential of membrane trafficking targeting in human diseases. For instance, Chlorpromazine, a GPCR antagonist used as an antipsychotic medication, was found to block dynamins activity and hence endocytosis of surface proteins. Despite these considerations, the druggability of many trafficking molecules is still unexplored.
In recent years, the development of both virtual screenings and cell-based functional assays have been employed for identification of novel trafficking inhibitors with improved target specificity and pathways selectivity. In this context, synthetic peptides, or covalent inhibitors able to block Rab small GTPases function could address selectivity issues in membrane trafficking inhibition.
Author contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
The research leading to these results has received funding from AIRC under MFAG 2020-ID. 24897 project–P.I. CC.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2023.1125801/full#supplementary-material
References
Ahmed, S. M., Nishida-Fukuda, H., Li, Y., McDonald, W. H., Gradinaru, C. C., and Macara, I. G. (2018). Exocyst dynamics during vesicle tethering and fusion. Nat. Commun. 9 (1), 5140. doi:10.1038/s41467-018-07467-5
Ali, A. M., Atmaj, J., Van Oosterwijk, N., Groves, M. R., and Dömling, A. (2019). Stapled peptides inhibitors: A new window for target drug discovery. Comput. Struct. Biotechnol. J. 17, 263–281. doi:10.1016/j.csbj.2019.01.012
Ashley, E. A., Recht, J., and White, N. J. (2014). Primaquine: The risks and the benefits. Malar. J. 13, 418. doi:10.1186/1475-2875-13-418
Balderhaar, H. J., Lachmann, J., Yavavli, E., Bröcker, C., Lürick, A., and Ungermann, C. (2013). The CORVET complex promotes tethering and fusion of Rab5/Vps21-positive membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (10), 3823–3828. doi:10.1073/pnas.1221785110
Bancone, G., Chowwiwat, N., Somsakchaicharoen, R., Poodpanya, L., Moo, P. K., Gornsawun, G., et al. (2016). Single low dose primaquine (0.25 mg/kg) does not cause clinically significant haemolysis in G6PD deficient subjects. PLoS One 11 (3), e0151898. doi:10.1371/journal.pone.0151898
Baranov, M. V., Bianchi, F., and van den Bogaart, G. (2020). The PIKfyve inhibitor apilimod: A double-edged sword against COVID-19. Cells 10 (1), 30. doi:10.3390/cells10010030
Basagiannis, D., Zografou, S., Goula, E., Gkeka, D., Kolettas, E., and Christoforidis, S. (2021). Chemical inhibitors of dynamin exert differential effects in VEGF signaling. Cells 10 (5), 997. doi:10.3390/cells10050997
Beautrait, A., Paradis, J. S., Zimmerman, B., Giubilaro, J., Nikolajev, L., Armando, S., et al. (2017). A new inhibitor of the β-arrestin/AP2 endocytic complex reveals interplay between GPCR internalization and signalling. Nat. Commun. 8, 15054. doi:10.1038/ncomms15054
Beurel, E., Grieco, S. F., and Jope, R. S. (2015). Glycogen synthase kinase-3 (GSK3): Regulation, actions, and diseases. Pharmacol. Ther. 148, 114–131. doi:10.1016/j.pharmthera.2014.11.016
Boal, F., Guetzoyan, L., Sessions, R. B., Zeghouf, M., Spooner, R. A., Lord, J. M., et al. (2010). LG186: An inhibitor of GBF1 function that causes Golgi disassembly in human and canine cells. Traffic 11 (12), 1537–1551. doi:10.1111/j.1600-0854.2010.01122.x
Bonet-Ponce, L., Beilina, A., Williamson, C. D., Lindberg, E., Kluss, J. H., Saez-Atienzar, S., et al. (2020). LRRK2 mediates tubulation and vesicle sorting from lysosomes. Sci. Adv. 6 (46), eabb2454. doi:10.1126/sciadv.abb2454
Burke, J. E., Triscott, J., Emerling, B. M., and Hammond, G. R. V. (2022). Beyond PI3Ks: Targeting phosphoinositide kinases in disease. Nat. Rev. Drug Discov. 14, 1–30. doi:10.1038/s41573-022-00582-5
Campa, C. C., Margaria, J. P., Derle, A., Del Giudice, M., De Santis, M. C., Gozzelino, L., et al. (2018). Rab11 activity and PtdIns(3)P turnover removes recycling cargo from endosomes. Nat. Chem. Biol. 14 (8), 801–810. doi:10.1038/s41589-018-0086-4
Cardoso, D. A., Chau, N., and Robinson, P. J. (2021). High-content drug discovery screening of endocytosis pathways. Methods Mol. Biol. 2233, 71–91. doi:10.1007/978-1-0716-1044-2_5
Cavalli, V., Vilbois, F., Corti, M., Marcote, M. J., Tamura, K., Karin, M., et al. (2001). The stress-induced MAP kinase p38 regulates endocytic trafficking via the GDI:Rab5 complex. Mol. Cell. 7 (2), 421–432. doi:10.1016/s1097-2765(01)00189-7
Chaudhary, N., Gomez, G. A., Howes, M. T., Lo, H. P., McMahon, K. A., Rae, J. A., et al. (2014). Endocytic crosstalk: Cavins, caveolins, and caveolae regulate clathrin-independent endocytosis. PLoS Biol. 12 (4), e1001832. doi:10.1371/journal.pbio.1001832
Chen, P., Gu, Z., Liu, W., and Yan, Z. (2007). Glycogen synthase kinase 3 regulates N-methyl-D-aspartate receptor channel trafficking and function in cortical neurons. Mol. Pharmacol. 72 (1), 40–51. doi:10.1124/mol.107.034942
Chew, H. Y., De Lima, P. O., Gonzalez Cruz, J. L., Banushi, B., Echejoh, G., Hu, L., et al. (2020). Endocytosis inhibition in humans to improve responses to ADCC-mediating antibodies. Cell 180 (5), 895–914. doi:10.1016/j.cell.2020.02.019
Chircop, M., Perera, S., Mariana, A., Lau, H., Ma, M. P. C., Gilbert, J., et al. (2011). Inhibition of dynamin by dynole 34-2 induces cell death following cytokinesis failure in cancer cells. Mol. Cancer Ther. 10 (9), 1553–1562. doi:10.1158/1535-7163.Mct-11-0067
Clarke, J. H., Giudici, M. L., Burke, J. E., Williams, R. L., Maloney, D. J., Marugan, J., et al. (2015). The function of phosphatidylinositol 5-phosphate 4-kinase γ (PI5P4Kγ) explored using a specific inhibitor that targets the PI5P-binding site. Biochem. J. 466 (2), 359–367. doi:10.1042/bj20141333
Clayton, E. L., Sue, N., Smillie, K. J., O'Leary, T., Bache, N., Cheung, G., et al. (2010). Dynamin I phosphorylation by GSK3 controls activity-dependent bulk endocytosis of synaptic vesicles. Nat. Neurosci. 13 (7), 845–851. doi:10.1038/nn.2571
Daniel, J. A., Chau, N., Abdel-Hamid, M. K., Hu, L., von Kleist, L., Whiting, A., et al. (2015). Phenothiazine-derived antipsychotic drugs inhibit dynamin and clathrin-mediated endocytosis. Traffic 16 (6), 635–654. doi:10.1111/tra.12272
De Matteis, M. A., and Luini, A. (2011). Mendelian disorders of membrane trafficking. N. Engl. J. Med. 365 (10), 927–938. doi:10.1056/NEJMra0910494
De Matteis, M. A., Vicinanza, M., Venditti, R., and Wilson, C. (2013). Cellular assays for drug discovery in genetic disorders of intracellular trafficking. Annu. Rev. Genomics Hum. Genet. 14, 159–190. doi:10.1146/annurev-genom-091212-153415
DeJesus-Hernandez, M., Mackenzie, I. R., Boeve, B. F., Boxer, A. L., Baker, M., Rutherford, N. J., et al. (2011). Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron 72 (2), 245–256. doi:10.1016/j.neuron.2011.09.011
Dejonghe, W., Kuenen, S., Mylle, E., Vasileva, M., Keech, O., Viotti, C., et al. (2016). Mitochondrial uncouplers inhibit clathrin-mediated endocytosis largely through cytoplasmic acidification. Nat. Commun. 7, 11710. doi:10.1038/ncomms11710
Dejonghe, W., Sharma, I., Denoo, B., De Munck, S., Lu, Q., Mishev, K., et al. (2019). Disruption of endocytosis through chemical inhibition of clathrin heavy chain function. Nat. Chem. Biol. 15 (6), 641–649. doi:10.1038/s41589-019-0262-1
DeLouise, L. A. (2012). Applications of nanotechnology in dermatology. J. Invest. Dermatol. 132 (2), 964–975. doi:10.1038/jid.2011.425
Dhekne, H. S., Yanatori, I., Gomez, R. C., Tonelli, F., Diez, F., Schule, B., et al. (2018). A pathway for Parkinson's Disease LRRK2 kinase to block primary cilia and Sonic hedgehog signaling in the brain. Elife. 6, e40202. doi:10.7554/eLife.40202
Dive, I., Klann, K., Michaelis, J. B., Heinzen, D., Steinbach, J. P., Munch, C., et al. (2022). Inhibition of mTOR signaling protects human glioma cells from hypoxia-induced cell death in an autophagy-independent manner. Cell Death Discov. 8 (1), 409. doi:10.1038/s41420-022-01195-y
Doherty, G. J., and McMahon, H. T. (2009). Mechanisms of endocytosis. Annu. Rev. Biochem. 78, 857–902. doi:10.1146/annurev.biochem.78.081307.110540
Dutta, D., Williamson, C. D., Cole, N. B., and Donaldson, J. G. (2012). Pitstop 2 is a potent inhibitor of clathrin-independent endocytosis. PLoS One 7 (9), e45799. doi:10.1371/journal.pone.0045799
Fell, M. J., Mirescu, C., Basu, K., Cheewatrakoolpong, B., DeMong, D. E., Ellis, J. M., et al. (2015). MLi-2, a potent, selective, and centrally active compound for exploring the therapeutic potential and safety of LRRK2 kinase inhibition. J. Pharmacol. Exp. Ther. 355 (3), 397–409. doi:10.1124/jpet.115.227587
Feng, Z., and Yu, C. H. (2021). PI(3,4)P(2)-mediated membrane tubulation promotes integrin trafficking and invasive cell migration. Proc. Natl. Acad. Sci. U. S. A. 118 (19), e2017645118. doi:10.1073/pnas.2017645118
Ferguson, S. M., and De Camilli, P. (2012). Dynamin, a membrane-remodelling GTPase. Nat. Rev. Mol. Cell Biol. 13 (2), 75–88. doi:10.1038/nrm3266
Fujimoto, B. A., Young, M., Carter, L., Pang, A. P. S., Corley, M. J., Fogelgren, B., et al. (2019). The exocyst complex regulates insulin-stimulated glucose uptake of skeletal muscle cells. Am. J. Physiol. Endocrinol. Metab. 317 (6), E957–E972. doi:10.1152/ajpendo.00109.2019
García-Cazorla, A., Oyarzábal, A., Saudubray, J. M., Martinelli, D., and Dionisi-Vici, C. (2022). Genetic disorders of cellular trafficking. Trends Genet. Jul 38 (7), 724–751. doi:10.1016/j.tig.2022.02.012
Gayle, S., Landrette, S., Beeharry, N., Conrad, C., Hernandez, M., Beckett, P., et al. (2017). Identification of apilimod as a first-in-class PIKfyve kinase inhibitor for treatment of B-cell non-Hodgkin lymphoma. Blood 129 (13), 1768–1778. doi:10.1182/blood-2016-09-736892
Gilleron, J., and Zeigerer, A. (2023). Endosomal trafficking in metabolic homeostasis and diseases. Nat. Rev. Endocrinol. Jan. 19 (1), 28–45. doi:10.1038/s41574-022-00737-9
Gilliam, A. J., Smith, J. N., Flather, D., Johnston, K. M., Gansmiller, A. M., Fishman, D. A., et al. (2016). Affinity-guided design of caveolin-1 ligands for deoligomerization. J. Med. Chem. 59 (8), 4019–4025. doi:10.1021/acs.jmedchem.5b01536
Giridharan, S. S. P., Luo, G., Rivero-Rios, P., Steinfeld, N., Tronchere, H., Singla, A., et al. (2022). Lipid kinases VPS34 and PIKfyve coordinate a phosphoinositide cascade to regulate retriever-mediated recycling on endosomes. Elife 11, e69709. doi:10.7554/eLife.69709
Gray, J. L., von Delft, F., and Brennan, P. E. (2020). Targeting the small GTPase superfamily through their regulatory proteins. Angew. Chem. Int. Ed. Engl. 59 (16), 6342–6366. doi:10.1002/anie.201900585
Gray, N. W., Fourgeaud, L., Huang, B., Chen, J., Cao, H., Oswald, B. J., et al. (2003). Dynamin 3 is a component of the postsynapse, where it interacts with mGluR5 and Homer. Curr. Biol. 13 (6), 510–515. doi:10.1016/s0960-9822(03)00136-2
Han, B., Porta, J. C., Hanks, J. L., Peskova, Y., Binshtein, E., Dryden, K., et al. (2020). Structure and assembly of CAV1 8S complexes revealed by single particle electron microscopy. Sci. Adv. 6 (49), eabc6185. doi:10.1126/sciadv.abc6185
Harding, C., Heuser, J., and Stahl, P. (1983). Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J. Cell Biol. 97 (2), 329–339. doi:10.1083/jcb.97.2.329
Harold, D., Abraham, R., Hollingworth, P., Sims, R., Gerrish, A., Hamshere, M. L., et al. (2009). Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat. Genet. 41 (10), 1088–1093. doi:10.1038/ng.440
Harper, C. B., Martin, S., Nguyen, T. H., Daniels, S. J., Lavidis, N. A., Popoff, M. R., et al. (2011). Dynamin inhibition blocks botulinum neurotoxin type A endocytosis in neurons and delays botulism. J. Biol. Chem. 286 (41), 35966–35976. doi:10.1074/jbc.M111.283879
Haucke, V. (2015). Cell biology: On the endocytosis rollercoaster. Nature. 517 (7535), 446–447. doi:10.1038/nature14081
He, Y., Liu, H., Yin, N., Yang, Y., Wang, C., Yu, M., et al. (2021). Barbadin potentiates long-term effects of lorcaserin on POMC neurons and weight loss. J. Neurosci. 41 (26), 5734–5746. doi:10.1523/JNEUROSCI.3210-20.2021
Heap, B. J., and Cowan, G. O. (1991). The epidemiology of snake bite presenting to British Military Hospital Dharan during 1989. J. R. Army Med. Corps 137 (3), 123–125. doi:10.1136/jramc-137-03-03
Helms, J. B., and Rothman, J. E. (1992). Inhibition by brefeldin A of a Golgi membrane enzyme that catalyses exchange of guanine nucleotide bound to ARF. Nature 360 (6402), 352–354. doi:10.1038/360352a0
Herbst, S., Campbell, P., Harvey, J., Bernard, E. M., Papayannopoulos, V., Wood, N. W., et al. (2020). LRRK2 activation controls the repair of damaged endomembranes in macrophages. Embo J. 39 (18), e104494. doi:10.15252/embj.2020104494
Hermida, M. A., Dinesh Kumar, J., and Leslie, N. R. (2017). GSK3 and its interactions with the PI3K/AKT/mTOR signalling network. Adv. Biol. Regul. 65, 5–15. doi:10.1016/j.jbior.2017.06.003
Hill, T. A., Odell, L. R., Quan, A., Abagyan, R., Ferguson, G., Robinson, P. J., et al. (2004). Long chain amines and long chain ammonium salts as novel inhibitors of dynamin GTPase activity. Bioorg Med. Chem. Lett. 14 (12), 3275–3278. doi:10.1016/j.bmcl.2004.03.096
Hinshaw, J. E., and Schmid, S. L. (1995). Dynamin self-assembles into rings suggesting a mechanism for coated vesicle budding. Nature. 374 (6518), 190–192. doi:10.1038/374190a0
Imoto, Y., Raychaudhuri, S., Ma, Y., Fenske, P., Sandoval, E., Itoh, K., et al. (2022). Dynamin is primed at endocytic sites for ultrafast endocytosis. Neuron. 110 (17), 2815–2835.e13. doi:10.1016/j.neuron.2022.06.010
Jahn, R., and Scheller, R. H. (2006). SNAREs--engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 7 (9), 631–643. doi:10.1038/nrm2002
Jamshidiha, M., Lanyon-Hogg, T., Sutherell, C. L., Craven, G. B., Tersa, M., De Vita, E., et al. (2022). Identification of the first structurally validated covalent ligands of the small GTPase RAB27A. RSC Med. Chem. 13 (2), 150–155. doi:10.1039/d1md00225b
Jimah, J. R., and Hinshaw, J. E. (2019). Structural insights into the mechanism of dynamin superfamily proteins. Trends Cell Biol. 29 (3), 257–273. doi:10.1016/j.tcb.2018.11.003
Jing, S. Q., Spencer, T., Miller, K., Hopkins, C., and Trowbridge, I. S. (1990). Role of the human transferrin receptor cytoplasmic domain in endocytosis: Localization of a specific signal sequence for internalization. J. Cell Biol. 110 (2), 283–294. doi:10.1083/jcb.110.2.283
Johnson, J. L., Ramadass, M., He, J., Brown, S. J., Zhang, J., Abgaryan, L., et al. (2016). Identification of neutrophil exocytosis inhibitors (nexinhibs), small molecule inhibitors of neutrophil exocytosis and inflammation: DRUGGABILITY of the small GTPase Rab27a. J. Biol. Chem. 291 (50), 25965–25982. doi:10.1074/jbc.M116.741884
Johnson, L. S., Dunn, K. W., Pytowski, B., and McGraw, T. E. (1993). Endosome acidification and receptor trafficking: Bafilomycin A1 slows receptor externalization by a mechanism involving the receptor's internalization motif. Mol. Biol. Cell. 4 (12), 1251–1266. doi:10.1091/mbc.4.12.1251
Joshi, S., Braithwaite, A. W., Robinson, P. J., and Chircop, M. (2011). Dynamin inhibitors induce caspase-mediated apoptosis following cytokinesis failure in human cancer cells and this is blocked by Bcl-2 overexpression. Mol. Cancer 10, 78. doi:10.1186/1476-4598-10-78
Joshi, S., Perera, S., Gilbert, J., Smith, C. M., Mariana, A., Gordon, C. P., et al. (2010). The dynamin inhibitors MiTMAB and OcTMAB induce cytokinesis failure and inhibit cell proliferation in human cancer cells. Mol. Cancer Ther. 9 (7), 1995–2006. doi:10.1158/1535-7163.Mct-10-0161
Jung, S. R., Jiang, Y., Seo, J. B., Chiu, D. T., Hille, B., and Koh, D. S. (2021). β-arrestin-dependent PI(4,5)P(2) synthesis boosts GPCR endocytosis. Proc. Natl. Acad. Sci. U. S. A. 118 (17), e2011023118. doi:10.1073/pnas.2011023118
Kang, Y. L., Chou, Y. Y., Rothlauf, P. W., Liu, Z., Soh, T. K., Cureton, D., et al. (2020). Inhibition of PIKfyve kinase prevents infection by Zaire ebolavirus and SARS-CoV-2. Proc. Natl. Acad. Sci. U. S. A. 117 (34), 20803–20813. doi:10.1073/pnas.2007837117
Karlsson, H. K., Zierath, J. R., Kane, S., Krook, A., Lienhard, G. E., and Wallberg-Henriksson, H. (2005). Insulin-stimulated phosphorylation of the Akt substrate AS160 is impaired in skeletal muscle of type 2 diabetic subjects. Diabetes 54 (6), 1692–1697. doi:10.2337/diabetes.54.6.1692
Kazama, I., Ejima, Y., Endo, Y., Toyama, H., Matsubara, M., Baba, A., et al. (2015). Chlorpromazine-induced changes in membrane micro-architecture inhibit thrombopoiesis in rat megakaryocytes. Biochim. Biophys. Acta 1848 (11), 2805–2812. doi:10.1016/j.bbamem.2015.08.013
Kirchhausen, T., Macia, E., and Pelish, H. E. (2008). Use of dynasore, the small molecule inhibitor of dynamin, in the regulation of endocytosis. Methods Enzymol. 438, 77–93. doi:10.1016/s0076-6879(07)38006-3
Klausner, R. D., Donaldson, J. G., and Lippincott-SchwartzBrefeldin, J. A. (1992). Brefeldin A: Insights into the control of membrane traffic and organelle structure. J. Cell Biol. 116 (5), 1071–1080. doi:10.1083/jcb.116.5.1071
Koike, S., and Jahn, R. (2017). Probing and manipulating intracellular membrane traffic by microinjection of artificial vesicles. Proc. Natl. Acad. Sci. U. S. A. 114 (46), E9883–e9892. doi:10.1073/pnas.1713524114
Koike, S., and Jahn, R. (2019). SNAREs define targeting specificity of trafficking vesicles by combinatorial interaction with tethering factors. Nat. Commun. 10 (1), 1608. doi:10.1038/s41467-019-09617-9
Kononenko, N. L., Puchkov, D., Classen, G. A., Walter, A. M., Pechstein, A., Sawade, L., et al. (2014). Clathrin/AP-2 mediate synaptic vesicle reformation from endosome-like vacuoles but are not essential for membrane retrieval at central synapses. Neuron 82 (5), 981–988. doi:10.1016/j.neuron.2014.05.007
Krishna, A., and Sengupta, D. (2019). Interplay between membrane curvature and cholesterol: Role of palmitoylated caveolin-1. Biophys. J. 116 (1), 69–78. doi:10.1016/j.bpj.2018.11.3127
Kuwahara, T., Funakawa, K., Komori, T., Sakurai, M., Yoshii, G., Eguchi, T., et al. (2020). Roles of lysosomotropic agents on LRRK2 activation and Rab10 phosphorylation. Neurobiol. Dis. 145, 105081. doi:10.1016/j.nbd.2020.105081
Lee, J. E., Westrate, L. M., Wu, H., Page, C., and Voeltz, G. K. (2016). Multiple dynamin family members collaborate to drive mitochondrial division. Nature 540 (7631), 139–143. doi:10.1038/nature20555
Lee, Y., Bortolotto, Z. A., Bradley, C. A., Sanderson, T. M., Zhuo, M., Kaang, B. K., et al. (2021). The GSK-3 inhibitor CT99021 enhances the acquisition of spatial learning and the accuracy of spatial memory. Front. Mol. Neurosci. 14, 804130. doi:10.3389/fnmol.2021.804130
Lemmon, S. K., and Traub, L. M. (2012). Getting in touch with the clathrin terminal domain. Traffic 13 (4), 511–519. doi:10.1111/j.1600-0854.2011.01321.x
Liashkovich, I., Pasrednik, D., Prystopiuk, V., Rosso, G., Oberleithner, H., and Shahin, V. (2015). Clathrin inhibitor Pitstop-2 disrupts the nuclear pore complex permeability barrier. Sci. Rep. 5, 9994. doi:10.1038/srep09994
Liberali, P., Rämö, P., and Pelkmans, L. (2008). Protein kinases: Starting a molecular systems view of endocytosis. Annu. Rev. Cell Dev. Biol. 24, 501–523. doi:10.1146/annurev.cellbio.041008.145637
Liberali, P., Snijder, B., and Pelkmans, L. (2014). A hierarchical map of regulatory genetic interactions in membrane trafficking. Cell 157 (6), 1473–1487. doi:10.1016/j.cell.2014.04.029
Liu, Z., Xu, E., Zhao, H. T., Cole, T., and West, A. B. (2020). LRRK2 and Rab10 coordinate macropinocytosis to mediate immunological responses in phagocytes. Embo J. 39 (20), e104862. doi:10.15252/embj.2020104862
Lo, W. T., Belabed, H., Kücükdisli, M., Metag, J., Roske, Y., Prokofeva, P., et al. (2022). Development of selective inhibitors of phosphatidylinositol 3-kinase C2α. Nat. Chem. Biol. 19, 18–27. doi:10.1038/s41589-022-01118-z
Lobbestael, E., Civiero, L., De Wit, T., Taymans, J. M., Greggio, E., and Baekelandt, V. (2016). Pharmacological LRRK2 kinase inhibition induces LRRK2 protein destabilization and proteasomal degradation. Sci. Rep. 6, 33897. doi:10.1038/srep33897
Lohse, M. J., Benovic, J. L., Codina, J., Caron, M. G., and Lefkowitz, R. J. (1990). beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science. 248 (4962), 1547–1550. doi:10.1126/science.2163110
Macia, E., Ehrlich, M., Massol, R., Boucrot, E., Brunner, C., and Kirchhausen, T. (2006). Dynasore, a cell-permeable inhibitor of dynamin. Dev. Cell. 10 (6), 839–850. doi:10.1016/j.devcel.2006.04.002
Mamais, A., Kluss, J. H., Bonet-Ponce, L., Landeck, N., Langston, R. G., Smith, N., et al. (2021). Mutations in LRRK2 linked to Parkinson disease sequester Rab8a to damaged lysosomes and regulate transferrin-mediated iron uptake in microglia. PLoS Biol. Dec 19 (12), e3001480. doi:10.1371/journal.pbio.3001480
Martin-Urdiroz, M., Deeks, M. J., Horton, C. G., Dawe, H. R., and Jourdain, I. (2016). The exocyst complex in health and disease. Front. Cell Dev. Biol. 4, 24. doi:10.3389/fcell.2016.00024
McCluskey, A., Daniel, J. A., Hadzic, G., Chau, N., Clayton, E. L., Mariana, A., et al. (2013). Building a better dynasore: The dyngo compounds potently inhibit dynamin and endocytosis. Traffic 14 (12), 1272–1289. doi:10.1111/tra.12119
McMahon, K. A., Zajicek, H., Li, W. P., Peyton, M. J., Minna, J. D., Hernandez, V. J., et al. (2009). SRBC/cavin-3 is a caveolin adapter protein that regulates caveolae function. EMBO J. 28 (8), 1001–1015. doi:10.1038/emboj.2009.46
Mei, K., and Guo, W. (2018). The exocyst complex. Curr. Biol. 28 (17), R922–R925. doi:10.1016/j.cub.2018.06.042
Mîinea, C. P., Sano, H., Kane, S., Sano, E., Fukuda, M., Peranen, J., et al. (2005). AS160, the Akt substrate regulating GLUT4 translocation, has a functional Rab GTPase-activating protein domain. Biochem. J. 391 (1), 87–93. doi:10.1042/bj20050887
Mishev, K., Dejonghe, W., and Russinova, E. (2013). Small molecules for dissecting endomembrane trafficking: A cross-systems view. Chem. Biol. 20 (4), 475–486. doi:10.1016/j.chembiol.2013.03.009
Mishra, A., Hourigan, D., and Lindsay, A. J. (2022). Inhibition of the endosomal recycling pathway downregulates HER2 activation and overcomes resistance to tyrosine kinase inhibitors in HER2-positive breast cancer. Cancer Lett. 529, 153–167. doi:10.1016/j.canlet.2022.01.003
Mitra, S., Montgomery, J. E., Kolar, M. J., Li, G., Jeong, K. J., Peng, B., et al. (2017). Stapled peptide inhibitors of RAB25 target context-specific phenotypes in cancer. Nat. Commun. 8 (1), 660. doi:10.1038/s41467-017-00888-8
Moltke, I., Grarup, N., Jørgensen, M. E., Bjerregaard, P., Treebak, J. T., Fumagalli, M., et al. (2014). A common Greenlandic TBC1D4 variant confers muscle insulin resistance and type 2 diabetes. Nature 512 (7513), 190–193. doi:10.1038/nature13425
Naghipour, S., Ghodousi, M., Rahsepar, S., and Elyasi, S. (2020). Repurposing of well-known medications as antivirals: Hydroxychloroquine and chloroquine - from HIV-1 infection to COVID-19. Expert Rev. Anti Infect. Ther. 18 (11), 1119–1133. doi:10.1080/14787210.2020.1792291
Odell, L. R., Howan, D., Gordon, C. P., Robertson, M. J., Chau, N., Mariana, A., et al. (2010). The pthaladyns: GTP competitive inhibitors of dynamin I and II GTPase derived from virtual screening. J. Med. Chem. 53 (14), 5267–5280. doi:10.1021/jm100442u
Oliva, C. R., Zhang, W., Langford, C., Suto, M. J., and Griguer, C. E. (2017). Repositioning chlorpromazine for treating chemoresistant glioma through the inhibition of cytochrome c oxidase bearing the COX4-1 regulatory subunit. Oncotarget 8 (23), 37568–37583. doi:10.18632/oncotarget.17247
Paksoy, A., Hoppe, S., Dorflinger, Y., Horstmann, H., Satzler, K., and Korber, C. (2022). Effects of the clathrin inhibitor Pitstop-2 on synaptic vesicle recycling at a central synapse in vivo. Front. Synaptic Neurosci. 14, 1056308. doi:10.3389/fnsyn.2022.1056308
Parton, R. G., and del Pozo, M. A. (2013). Caveolae as plasma membrane sensors, protectors and organizers. Nat. Rev. Mol. Cell Biol. 14 (2), 98–112. doi:10.1038/nrm3512
Parton, R. G., Tillu, V., McMahon, K. A., and Collins, B. M. (2021). Key phases in the formation of caveolae. Curr. Opin. Cell Biol. 71, 7–14. doi:10.1016/j.ceb.2021.01.009
Peplowska, K., Markgraf, D. F., Ostrowicz, C. W., Bange, G., and Ungermann, C. (2007). The CORVET tethering complex interacts with the yeast Rab5 homolog Vps21 and is involved in endo-lysosomal biogenesis. Dev. Cell. 12 (5), 739–750. doi:10.1016/j.devcel.2007.03.006
Perez Verdaguer, M., Zhang, T., Paulo, J. A., Gygi, S., Watkins, S. C., Sakurai, H., et al. (2021). Mechanism of p38 MAPK-induced EGFR endocytosis and its crosstalk with ligand-induced pathways. J. Cell Biol. 220 (7), e202102005. doi:10.1083/jcb.202102005
Persoons, L., Vanderlinden, E., Vangeel, L., Wang, X., Do, N. D. T., Foo, S. Y. C., et al. (2021). Broad spectrum anti-coronavirus activity of a series of anti-malaria quinoline analogues. Antivir. Res. 193, 105127. doi:10.1016/j.antiviral.2021.105127
Plummer, E. M., and Manchester, M. (2013). Endocytic uptake pathways utilized by CPMV nanoparticles. Mol. Pharm. 10 (1), 26–32. doi:10.1021/mp300238w
Podinovskaia, M., Prescianotto-Baschong, C., Buser, D. P., and Spang, A. (2021). A novel live-cell imaging assay reveals regulation of endosome maturation. Elife 30. doi:10.7554/elife.70982
Poli, A., Abdul-Hamid, S., Zaurito, A. E., Campagnoli, F., Bevilacqua, V., Sheth, B., et al. (2021). PIP4Ks impact on PI3K, FOXP3, and UHRF1 signaling and modulate human regulatory T cell proliferation and immunosuppressive activity. Proc. Natl. Acad. Sci. U. S. A. 118 (31), e2010053118. doi:10.1073/pnas.2010053118
Porta, J. C., Han, B., Gulsevin, A., Chung, J. M., Peskova, Y., Connolly, S., et al. (2022). Molecular architecture of the human caveolin-1 complex. Sci. Adv. 8 (19), eabn7232. doi:10.1126/sciadv.abn7232
Posor, Y., Eichhorn-Gruenig, M., Puchkov, D., Schoneberg, J., Ullrich, A., Lampe, A., et al. (2013). Spatiotemporal control of endocytosis by phosphatidylinositol-3,4-bisphosphate. Nature 499 (7457), 233–237. doi:10.1038/nature12360
Preta, G., Cronin, J. G., and Sheldon, I. M. (2015). Dynasore - not just a dynamin inhibitor. Cell Commun. Signal 13, 24. doi:10.1186/s12964-015-0102-1
Quan, A., McGeachie, A. B., Keating, D. J., van Dam, E. M., Rusak, J., Chau, N., et al. (2007). Myristyl trimethyl ammonium bromide and octadecyl trimethyl ammonium bromide are surface-active small molecule dynamin inhibitors that block endocytosis mediated by dynamin I or dynamin II. Mol. Pharmacol. 72 (6), 1425–1439. doi:10.1124/mol.107.034207
Rainsford, K. D., Parke, A. L., Clifford-Rashotte, M., and Kean, W. F. (2015). Therapy and pharmacological properties of hydroxychloroquine and chloroquine in treatment of systemic lupus erythematosus, rheumatoid arthritis and related diseases. Inflammopharmacology. 23 (5), 231–269. doi:10.1007/s10787-015-0239-y
Ranjan, R., Dwivedi, H., Baidya, M., Kumar, M., and Shukla, A. K. (2017). Novel structural insights into GPCR-β-arrestin interaction and signaling. Trends Cell Biol. 27 (11), 851–862. doi:10.1016/j.tcb.2017.05.008
Redpath, G. M. I., Betzler, V. M., Rossatti, P., and Rossy, J. (2020). Membrane heterogeneity controls cellular endocytic trafficking. Front. Cell Dev. Biol. 8, 757. doi:10.3389/fcell.2020.00757
Reider, A., and Wendland, B. (2011). Endocytic adaptors--social networking at the plasma membrane. J. Cell Sci. 124 (10), 1613–1622. doi:10.1242/jcs.073395
Reiner, D. J., and Lundquist, E. A. (2018). Small GTPases. WormBook. 2018, 1–65. doi:10.1895/wormbook.1.67.2
Renard, H. F., and Boucrot, E. (2021). Unconventional endocytic mechanisms. Curr. Opin. Cell Biol. 71, 120–129. doi:10.1016/j.ceb.2021.03.001
Renard, H. F., Johannes, L., and Morsomme, P. (2018). Increasing diversity of biological membrane fission mechanisms. Trends Cell Biol. 28 (4), 274–286. doi:10.1016/j.tcb.2017.12.001
Rennick, J. J., Johnston, A. P. R., and Parton, R. G. (2021). Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 16 (3), 266–276. doi:10.1038/s41565-021-00858-8
Riva, L., Yuan, S., Yin, X., Martin-Sancho, L., Matsunaga, N., Pache, L., et al. (2020). Discovery of SARS-CoV-2 antiviral drugs through large-scale compound repurposing. Nature 586 (7827), 113–119. doi:10.1038/s41586-020-2577-1
Ronan, B., Flamand, O., Vescovi, L., Dureuil, C., Durand, L., Fassy, F., et al. (2014). A highly potent and selective Vps34 inhibitor alters vesicle trafficking and autophagy. Nat. Chem. Biol. 10 (12), 1013–1019. doi:10.1038/nchembio.1681
Roskoski, R. (2015). A historical overview of protein kinases and their targeted small molecule inhibitors. Pharmacol. Res. 100, 1–23. doi:10.1016/j.phrs.2015.07.010
Roskoski, R. (2016). Classification of small molecule protein kinase inhibitors based upon the structures of their drug-enzyme complexes. Pharmacol. Res. 103, 26–48. doi:10.1016/j.phrs.2015.10.021
Rouhana, J., Hoh, F., Estaran, S., Henriquet, C., Boublik, Y., Kerkour, A., et al. (2013). Fragment-based identification of a locus in the Sec7 domain of Arno for the design of protein-protein interaction inhibitors. J. Med. Chem. 56 (21), 8497–8511. doi:10.1021/jm4009357
Sathe, M., Muthukrishnan, G., Rae, J., Disanza, A., Thattai, M., Scita, G., et al. (2018). Small GTPases and BAR domain proteins regulate branched actin polymerisation for clathrin and dynamin-independent endocytosis. Nat. Commun. 9 (1), 1835. doi:10.1038/s41467-018-03955-w
Sbrissa, D., Naisan, G., Ikonomov, O. C., and Shisheva, A. (2018). Apilimod, a candidate anticancer therapeutic, arrests not only PtdIns(3,5)P2 but also PtdIns5P synthesis by PIKfyve and induces bafilomycin A1-reversible aberrant endomembrane dilation. PLoS One 13 (9), e0204532. doi:10.1371/journal.pone.0204532
Scherer, P. E., Lewis, R. Y., Volonte, D., Engelman, J. A., Galbiati, F., Couet, J., et al. (1997). Cell-type and tissue-specific expression of caveolin-2. Caveolins 1 and 2 co-localize and form a stable hetero-oligomeric complex in vivo. J. Biol. Chem. 272 (46), 29337–29346. doi:10.1074/jbc.272.46.29337
Schindler, C., Chen, Y., Pu, J., Guo, X., and Bonifacino, J. S. (2015). EARP is a multisubunit tethering complex involved in endocytic recycling. Nat. Cell Biol. 17 (5), 639–650. doi:10.1038/ncb3129
Schmid, E. M., and McMahon, H. T. (2007). Integrating molecular and network biology to decode endocytosis. Nature 448 (7156), 883–888. doi:10.1038/nature06031
Schreij, A. M., Fon, E. A., and McPherson, P. S. (2016). Endocytic membrane trafficking and neurodegenerative disease. Cell Mol. Life Sci. 73 (8), 1529–1545. doi:10.1007/s00018-015-2105-x
Scott, C. C., and Gruenberg, J. (2011). Ion flux and the function of endosomes and lysosomes: pH is just the start: The flux of ions across endosomal membranes influences endosome function not only through regulation of the luminal pH. Bioessays. 33 (2), 103–110. doi:10.1002/bies.201000108
Scott, C. C., Vacca, F., and Gruenberg, J. (2014). Endosome maturation, transport and functions. Semin. Cell Dev. Biol. 31, 2–10. doi:10.1016/j.semcdb.2014.03.034
Scott, J. D., DeMong, D. E., Greshock, T. J., Basu, K., Dai, X., Harris, J., et al. (2017). Discovery of a 3-(4-pyrimidinyl) indazole (MLi-2), an orally available and selective leucine-rich repeat kinase 2 (LRRK2) inhibitor that reduces brain kinase activity. J. Med. Chem. 60 (7), 2983–2992. doi:10.1021/acs.jmedchem.7b00045
Shafaq-Zadah, M., Dransart, E., and Johannes, L. (2020). Clathrin-independent endocytosis, retrograde trafficking, and cell polarity. Curr. Opin. Cell Biol. 65, 112–121. doi:10.1016/j.ceb.2020.05.009
Shin, S. Y., Lee, K. S., Choi, Y. K., Lim, H. J., Lee, H. G., Lim, Y., et al. (2013). The antipsychotic agent chlorpromazine induces autophagic cell death by inhibiting the Akt/mTOR pathway in human U-87MG glioma cells. Carcinogenesis 34 (9), 2080–2089. doi:10.1093/carcin/bgt169
Shvets, E., Bitsikas, V., Howard, G., Hansen, C. G., and Nichols, B. J. (2015). Dynamic caveolae exclude bulk membrane proteins and are required for sorting of excess glycosphingolipids. Nat. Commun. 6, 6867. doi:10.1038/ncomms7867
Sigismund, S., Confalonieri, S., Ciliberto, A., Polo, S., Scita, G., and Di Fiore, P. P. (2012). Endocytosis and signaling: Cell logistics shape the eukaryotic cell plan. Physiol. Rev. 92 (1), 273–366. doi:10.1152/physrev.00005.2011
Sigismund, S., Lanzetti, L., Scita, G., and Di Fiore, P. P. (2021). Endocytosis in the context-dependent regulation of individual and collective cell properties. Nat. Rev. Mol. Cell Biol. 22 (9), 625–643. doi:10.1038/s41580-021-00375-5
Simunovic, M., Evergren, E., Callan-Jones, A., and Bassereau, P. (2019). Curving cells inside and out: Roles of BAR domain proteins in membrane shaping and its cellular implications. Annu. Rev. Cell Dev. Biol. 35, 111–129. doi:10.1146/annurev-cellbio-100617-060558
Simunovic, M., Voth, G. A., Callan-Jones, A., and Bassereau, P. (2015). When physics takes over: BAR proteins and membrane curvature. Trends Cell Biol. 25 (12), 780–792. doi:10.1016/j.tcb.2015.09.005
Singh, J., Elhabashy, H., Muthukottiappan, P., Stepath, M., Eisenacher, M., Kohlbacher, O., et al. (2022). Cross-linking of the endolysosomal system reveals potential flotillin structures and cargo. Nat. Commun. 13 (1), 6212. doi:10.1038/s41467-022-33951-0
Smillie, K. J., Pawson, J., Perkins, E. M., Jackson, M., and Cousin, M. A. (2013). Control of synaptic vesicle endocytosis by an extracellular signalling molecule. Nat. Commun. 4, 2394. doi:10.1038/ncomms3394
Solinger, J. A., Rashid, H. O., and Spang, A. (2022). FERARI and cargo adaptors coordinate cargo flow through sorting endosomes. Nat. Commun. 13 (1), 4620. doi:10.1038/s41467-022-32377-y
Steger, M., Diez, F., Dhekne, H. S., Lis, P., Nirujogi, R. S., Karayel, O., et al. (2017). Systematic proteomic analysis of LRRK2-mediated Rab GTPase phosphorylation establishes a connection to ciliogenesis. Elife. 10, e31012. doi:10.7554/eLife.31012
Tuma, P. L., and Collins, C. A. (1994). Activation of dynamin GTPase is a result of positive cooperativity. J. Biol. Chem. 269 (49), 30842–30847. doi:10.1016/s0021-9258(18)47358-9
Van Jaarsveld, P. P., Nandi, P. K., Lippoldt, R. E., Saroff, H., and Edelhoch, H. (1981). Polymerization of clathrin protomers into basket structures. Biochemistry 20 (14), 4129–4135. doi:10.1021/bi00517a028
van Weert, A. W., Dunn, K. W., Geuze, H. J., Maxfield, F. R., and Stoorvogel, W. (1995). Transport from late endosomes to lysosomes, but not sorting of integral membrane proteins in endosomes, depends on the vacuolar proton pump. J. Cell Biol. 130 (4), 821–834. doi:10.1083/jcb.130.4.821
van Weert, A. W., Geuze, H. J., Groothuis, B., and Stoorvogel, W. (2000). Primaquine interferes with membrane recycling from endosomes to the plasma membrane through a direct interaction with endosomes which does not involve neutralisation of endosomal pH nor osmotic swelling of endosomes. Eur. J. Cell Biol. 79 (6), 394–399. doi:10.1078/0171-9335-00062
Vanneste, M., Huang, Q., Li, M., Moose, D., Zhao, L., Stamnes, M. A., et al. (2019). High content screening identifies monensin as an EMT-selective cytotoxic compound. Sci. Rep. 9 (1), 1200. doi:10.1038/s41598-018-38019-y
Vercauteren, D., Vandenbroucke, R. E., Jones, A. T., Rejman, J., Demeester, J., De Smedt, S. C., et al. (2010). The use of inhibitors to study endocytic pathways of gene carriers: Optimization and pitfalls. Mol. Ther. 18 (3), 561–569. doi:10.1038/mt.2009.281
Vilariño-Güell, C., Rajput, A., Milnerwood, A. J., Shah, B., Szu-Tu, C., Trinh, J., et al. (2014). DNAJC13 mutations in Parkinson disease. Hum. Mol. Genet. 23 (7), 1794–1801. doi:10.1093/hmg/ddt570
Vilariño-Güell, C., Wider, C., Ross, O. A., Dachsel, J. C., Kachergus, J. M., Lincoln, S. J., et al. (2011). VPS35 mutations in Parkinson disease. Am. J. Hum. Genet. 89 (1), 162–167. doi:10.1016/j.ajhg.2011.06.001
von Beek, C., Alriksson, L., Palle, J., Gustafson, A. M., Grujic, M., Melo, F. R., et al. (2021). Dynamin inhibition causes context-dependent cell death of leukemia and lymphoma cells. PLoS One 16 (9), e0256708. doi:10.1371/journal.pone.0256708
von Kleist, L., Stahlschmidt, W., Bulut, H., Gromova, K., Puchkov, D., Robertson, M. J., et al. (2011). Role of the clathrin terminal domain in regulating coated pit dynamics revealed by small molecule inhibition. Cell 146 (3), 471–484. doi:10.1016/j.cell.2011.06.025
Wälchli, S., Skånland, S. S., Gregers, T. F., Lauvrak, S. U., Torgersen, M. L., Ying, M., et al. (2008). The Mitogen-activated protein kinase p38 links Shiga Toxin-dependent signaling and trafficking. Mol. Biol. Cell. 19 (1), 95–104. doi:10.1091/mbc.e07-06-0565
Wallroth, A., and Haucke, V. (2018). Phosphoinositide conversion in endocytosis and the endolysosomal system. J. Biol. Chem. 293 (5), 1526–1535. doi:10.1074/jbc.R117.000629
Wandinger-Ness, A., and Zerial, M. (2014). Rab proteins and the compartmentalization of the endosomal system. Cold Spring Harb. Perspect. Biol. 6 (11), a022616. doi:10.1101/cshperspect.a022616
Wang, R., Wang, J., Hassan, A., Lee, C. H., Xie, X. S., and Li, X. (2021). Molecular basis of V-ATPase inhibition by bafilomycin A1. Nat. Commun. 12, 1782. doi:10.1038/s41467-021-22111-5
Wang, X., Wu, X., Zhang, Z., Ma, C., Wu, T., Tang, S., et al. (2018). Monensin inhibits cell proliferation and tumor growth of chemo-resistant pancreatic cancer cells by targeting the EGFR signaling pathway. Sci. Rep. 8 (1), 17914. doi:10.1038/s41598-018-36214-5
Watanabe, S., Trimbuch, T., Camacho-Pérez, M., Rost, B. R., Brokowski, B., Sohl-Kielczynski, B., et al. (2014). Clathrin regenerates synaptic vesicles from endosomes. Nature 515 (7526), 228–233. doi:10.1038/nature13846
Willox, A. K., Sahraoui, Y. M., and Royle, S. J. (2014). Non-specificity of Pitstop 2 in clathrin-mediated endocytosis. Biol. Open 3 (5), 326–331. doi:10.1242/bio.20147955
Wu, Y., O'Toole, E. T., Girard, M., Ritter, B., Messa, M., Liu, X., et al. (2014). A dynamin 1-dynamin 3- and clathrin-independent pathway of synaptic vesicle recycling mediated by bulk endocytosis. Elife 3, e01621. doi:10.7554/eLife.01621
Xing, Y., Wen, Z., Gao, W., Lin, Z., Zhong, J., and Jiu, Y. (2020). Multifaceted functions of host cell caveolae/caveolin-1 in virus infections. Viruses 12 (5), 487. doi:10.3390/v12050487
Yarwood, R., Hellicar, J., Woodman, P. G., and Lowe, M. (2020). Membrane trafficking in health and disease. Dis. Model Mech. 13 (4), dmm043448. doi:10.1242/dmm.043448
Yoshimori, T., Yamamoto, A., Moriyama, Y., Futai, M., and Tashiro, Y. (1991). Bafilomycin A1, a specific inhibitor of vacuolar-type H(+)-ATPase, inhibits acidification and protein degradation in lysosomes of cultured cells. J. Biol. Chem. 266 (26), 17707–17712. doi:10.1016/s0021-9258(19)47429-2
Yoshioka, K. (2021). Class II phosphatidylinositol 3-kinase isoforms in vesicular trafficking. Biochem. Soc. Trans. 49 (2), 893–901. doi:10.1042/bst20200835
Yu, Q., Wang, Y., Du, F., Yan, S., Hu, G., Origlia, N., et al. (2018). Overexpression of endophilin A1 exacerbates synaptic alterations in a mouse model of Alzheimer's disease. Nat. Commun. 9 (1), 2968. doi:10.1038/s41467-018-04389-0
Yuan, N., Song, L., Zhang, S., Lin, W., Cao, Y., Xu, F., et al. (2015). Bafilomycin A1 targets both autophagy and apoptosis pathways in pediatric B-cell acute lymphoblastic leukemia. Haematologica. 100 (3), 345–356. doi:10.3324/haematol.2014.113324
Zhang, C., Brown, M. Q., van de Ven, W., Zhang, Z. M., Wu, B., Young, M. C., et al. (2016). Endosidin2 targets conserved exocyst complex subunit EXO70 to inhibit exocytosis. Proc. Natl. Acad. Sci. U. S. A. 113 (1), E41–E50. doi:10.1073/pnas.1521248112
Zheng, L., and Conner, S. D. (2018). Glycogen synthase kinase 3β inhibition enhances Notch1 recycling. Mol. Biol. Cell. 29 (4), 389–395. doi:10.1091/mbc.E17-07-0474
Zheng, L., and Conner, S. D. (2018). PI5P4Kγ functions in DTX1-mediated Notch signaling. Proc. Natl. Acad. Sci. U. S. A. 115 (9), E1983–e1990. doi:10.1073/pnas.1712142115
Keywords: endocytosis, endocytic recycling, small molecules, inhibitors, mechanism of action
Citation: Placidi G, Mattu C, Ciardelli G and Campa CC (2023) Small molecules targeting endocytic uptake and recycling pathways. Front. Cell Dev. Biol. 11:1125801. doi: 10.3389/fcell.2023.1125801
Received: 16 December 2022; Accepted: 23 February 2023;
Published: 10 March 2023.
Edited by:
Pero Lucin, University of Rijeka, CroatiaReviewed by:
Inmaculada Navarro-Lérida, Autonomous University of Madrid, SpainCopyright © 2023 Placidi, Mattu, Ciardelli and Campa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carlo C. Campa, Y2FybG9jb3NpbW8uY2FtcGFAaWlnbS5pdA==