 Junhong Wang1,2,3,4†
Junhong Wang1,2,3,4† Chunwei Shi1,2,3,4†Mingyang Cheng1,2,3,4Yiyuan Lu1,2,3,4Xiaoyu Zhang1,2,3,4Fengdi Li1,2,3,4Yu Sun1,2,3,4Xiaoxu Li1,2,3,4Xinyang Li1,2,3,4Yan Zeng1,2,3,4*
Chunwei Shi1,2,3,4†Mingyang Cheng1,2,3,4Yiyuan Lu1,2,3,4Xiaoyu Zhang1,2,3,4Fengdi Li1,2,3,4Yu Sun1,2,3,4Xiaoxu Li1,2,3,4Xinyang Li1,2,3,4Yan Zeng1,2,3,4* Chunfeng Wang1,2,3,4*
Chunfeng Wang1,2,3,4* Xin Cao1,2,3,4*
Xin Cao1,2,3,4*- 1College of Veterinary Medicine, Jilin Agricultural University, Changchun, China
- 2Jilin Provincial Key Laboratory of Animal Microecology and Healthy Breeding, Jilin Agricultural University, Changchun, China
- 3Jilin Provincial Engineering Research Center of Animal Probiotics, Jilin Agricultural University, Changchun, China
- 4Key Laboratory of Animal Production and Product Quality Safety of Ministry of Education, Jilin Agricultural University, Changchun, China
Zbtb1 (zinc finger and BTB domain containing 1) is a member of mammalian zbtb gene family. A series of bioinformatics analysis was carried out for the EL4 cell and the Zbtb1-deficient EL4 cell by Hi-C, ATAC-seq and RNA-seq techniques. Finally, Hi-C results showed that the intensity of chromatin interaction in the deletion group decreased with distance, the degree of chromosome interaction decreased significantly, the AB division region changed significantly, and the compactness of TAD structure decreased; The results of ATAC-seq showed that the open area and degree of chromatin in the deletion group decreased; 7778 differentially expressed mRNAs were found by RNA-seq. Our experimental results for the first time expounded the significance of Zbtb1 gene for T cell development, lymphocyte production and apoptosis from the aspects of chromosome spatial structure and chromatin opening degree, and provided relevant theoretical basis and data support for the in-depth study of related Zbtb1 genes in the future.
Introduction
Zbtb1 plays a key role in T cell development and lymphocyte development, mRNA encoding Zbtb1 is most highly expressed in hematopoietic stem cells, thymocytes and pre-B cells, In addition to its role in T cell development, it was also demonstrated to be involved in the differentiation of B cells and NK cells, homozygous knockout of the Zbtb1 gene leads to severe combined immune deficiency in mice (Punwani et al., 2012; Lu et al., 2017). In other areas, acts as a transcriptional repressor (Matic et al., 2010); Represses cAMP-responsive element (CRE)-mediated transcriptional activation (Liu et al., 2011); Has a role in translesion DNA synthesis. Requires for UV-inducible RAD18 loading, PCNA monoubiquitination, POLH recruitment to replication factories and efficient translesion DNA synthesis (Kim et al., 2014). Our previous experimental results showed that Zbtb1 gene deletion slowed the growth rate of EL4 cells (Wang et al., 2021a).
Hi-C technology, derived from (Chromosome Conformation Capture—3C) technology, uses high-throughput sequencing technology, using proximity ligation combined with high-throughput sequencing, to study the interaction of the entire chromatin DNA on a genome-wide scale, taking the entire cell nucleus as the object of study (van Berkum et al., 2010). The formation of chromatin interactions is essential for the normal function of cells for the normal function of cells (Lafontaine et al., 2021). Hi-C data analysis is able to obtain information on interactions between genomic loci, divide the genome into bins of a specific size, and thus measure the strength of the interaction between two genomic loci (bins) (Fortin and Hansen, 2015).
ATAC-seq (Assay for Transposase-Accessible Chromatin with high-throughput sequencing) uses the preference for of open region chromatin to transposase for open region identification, and uses modified Tn5 transposase to directly introduce sequencing junctions into the open chromatin region by transposition reaction, and amplifies and sequences the open chromatin to finally obtain a genome-wide open chromatin map (Buenrostro et al., 2015).
Hi-C interaction data were analyzed jointly with ATAC-seq and transcriptome data, which can elucidate the mechanisms involved in organismal trait formation in terms of gene regulatory networks and epigenetic networks.
Results
Basic Quality-Related Data
Our sequencing analysis of Hi-C, ATAC-seq and RNA-seq was performed with the assistance of Annoroad Gene Technology (Beijing, China), and the related experimental methods are shown in (Supplementary Material S1), the preliminary quality statistics and extensive basic data data are shown in (Supplementary Material S2, S3, S4).
Analysis of Hi-C Data Results
The results of the Hi-C experiment showed that the interaction ratio of CIS and trans chromosomes in the Zbtb1-deficient EL4 cell (KO group) and the EL4 cell (WT group) was approximately 75:25 (Supplementary Figure S1A). Chromatin interaction frequencies (IFS) attenuated with increasing distance in a given range, Decay of chromatin opening with distance in KO and WT groups (Figures 1A,B) and the interaction decay exponents (IDEs) of the KO group samples increased (Figure 1C). The interaction degree of chromosomes changed significantly, including CIS interaction changes as shown in Figure 1D (taking the first chromosome as an example) and trans interaction changes (shown in Supplementary Figure S1B). A/B components have cell specificity and can be transformed into different tissues and cells. This transformation is related to gene expression regulation (Chiliński et al., 2021). A comparison of the A/B component transformation between KO group and WT group is shown in Figure 1E, Also chromatin opening peaks and transcription of RNA were altered (taking the first chromosome as an example). In Hi-C interaction thermograms of mammals at around 40 kb resolution, we were able to observe that the thermograms show distinct triangular structures (Dixon et al., 2012). These triangular structures are named as Topologically associated domains (TAD), and these structural domains have distinct boundaries between them. The TAD structure is conserved in different times and spaces (organization, development stage, etc.), and there were also some dynamic changes. We found that the TAD structure of the cells without the Zbtb1 gene changed significantly, and the boundary was blurred or even disappeared, For example, the TAD change at position 7160000–7200000 on the first chromosome (Figure 1F). TAD display on the first chromosome of the WT and KO groups in Figure 1G. The TAD score, an index used to measure the tightness of the TAD structure, is shown in Figure 1H (Wang et al., 2021b).
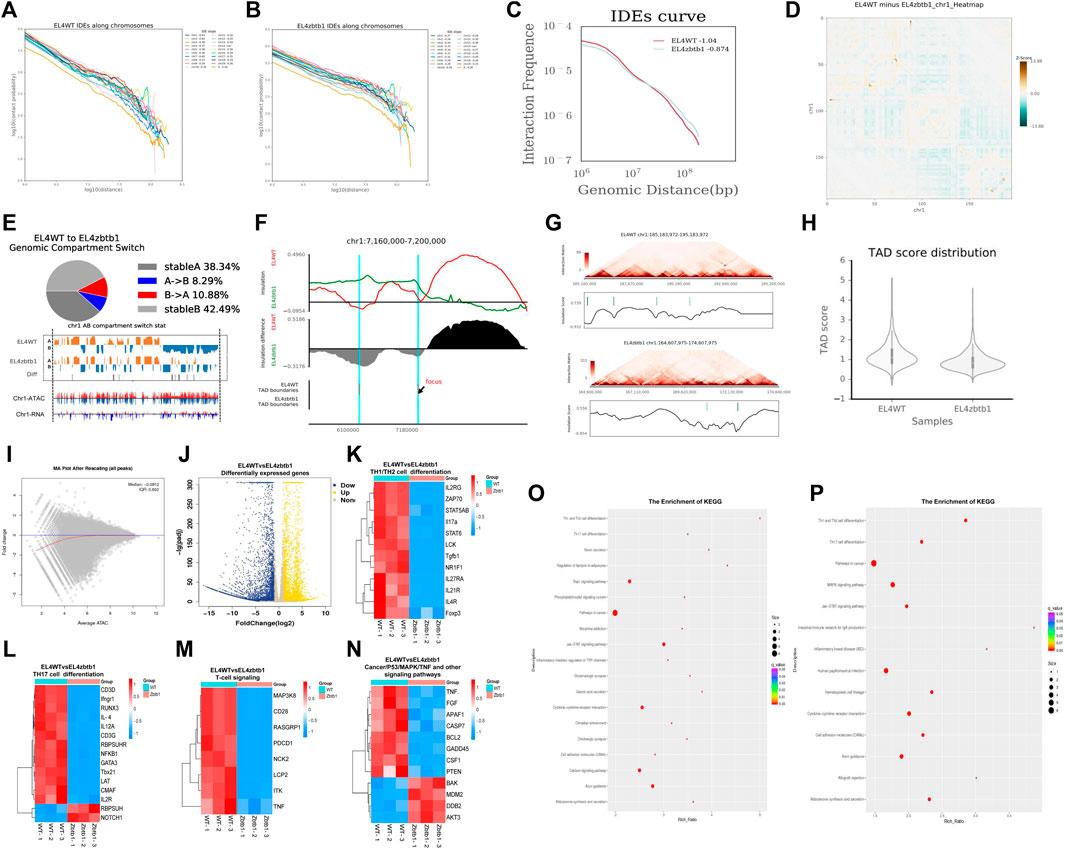
FIGURE 1. The analysis results of EL4 lacking zbtb1 were displayed by Hi-C, RNA-seq and ATAC-seq.
Analysis of ATAC-Seq Data Results
ATAC-seq results showed that the open peak regions in the deletion group cells were seriously reduced, and the openness of many regions was weakened. The chromatin open region map at the chromosome level in the whole genome is shown in Figure 1I visually shows the difference in the openness of all peaks, which is consistent with our RNA-seq verification results. There was a certain correlation between the differentially open region and the expression of differentially expressed mRNAs, the correlation was especially consistent between the open weakened region and the differentially downregulated mRNAs (Supplementary Figure S1C). The binding sites of transcription factors and other DNA sequences have certain characteristics, which are called Motifs, and therefore the detection of these Motifs in open regions of the whole genome can help to discover new transcription factors and annotate new functions of known transcription factors. We found that the Motifs of the KO group differed significantly from those of the WT group (Supplementary Material S5).
Analysis of RNA-Seq Data Results
The RNA-seq results showed that 3185 differentially expressed mRNAs were upregulated and 4593 were downregulated. The volcanic map of the differentially expressed genes is shown in Figure 1J. We clustered the differentially expressed mRNA and found significant differences in the expression of key genes in Th1 and Th2 cell differentiation, Th17 cell differentiation and other pathways (Figures 1K–N). KEGG enrichment of genes annotated in chromosome differentially open regions and differentially expressed mRNAs (Figures 1O,P) found they were significantly enriched in lymphatic development-related pathways such as Th1 and Th2 cell differentiation, Th17 cell differentiation, etc. The signaling pathways related to cell growth, apoptosis and damage repair, such as MAPK, cancer pathways and TNF, were significantly enriched. According to the GO enrichment results (Supplementary Figure S1D), the differentially expressed mRNAs were enriched in 58 GO terms and significantly enriched in organelles, cell parts, cellular processes, biological regulation and binding.
Combined Analysis of Hi-C, ATAC-Seq and RNA-Seq
Previous studies have shown that Zbtb1 is a key determinant of T cell development and lymphocyte production and it affects cell apoptosis (Cao et al., 2016; Cheng et al., 2021). Through the combined analysis of Hi-C, ATAC-seq and RNA-seq, we found that KO group lost the expression of multiple key genes involved in Th1, Th2 and Th17 cell differentiation, T cell signaling and p53 signaling. Combined with the results of TAD, AB compartment and chromatin openness in the region where the genes are located, it was found that the degree of chromosome interaction at the location of the genes not being transcribed was generally reduced, the TAD boundary of the gene location became blurred, the open chromatin region was reduced, and the openness was weakened. Therefore, some mRNA cannot be transcribed normally and the transcriptional restriction of these genes might be the reason why mice with deletion of Zbtb1 gene cannot survive normally. The genes that were not being transcribed included regulatory genes important for lymphocyte development and differentiation, including GATA3 (Jiang et al., 2021) (Figure 2A), PDCD1 (Supplementary Figure S1E), RASGRP1 (Supplementary Figure S1F) and others, such as signal transduction-related genes lat (Supplementary Figure S1G), JAK1 and LCK (Figure 2B); cell receptor-related genes, such as CD3 (including CD3d, CD3g, CD3e and other genes) (Figure 2C), IL7R (Bevington et al., 2020) (Supplementary Figure S1H), IL2RG (Supplementary Figure S1I), and CD69 (Supplementary Figure S1J); regulatory factor genes, such as IL17a, IL17f (Supplementary Figure S1K) and RUNX3 (Supplementary Figure S1L); and isopathways of apoptosis, such as BCL2 (Supplementary Figure S1M), LCP2 (Supplementary Figure S1N), the target gene PERP of p53/p63 and the antitumor key gene IFNGR1 (Roberts and Paraoan, 2020) (Figure 2D).
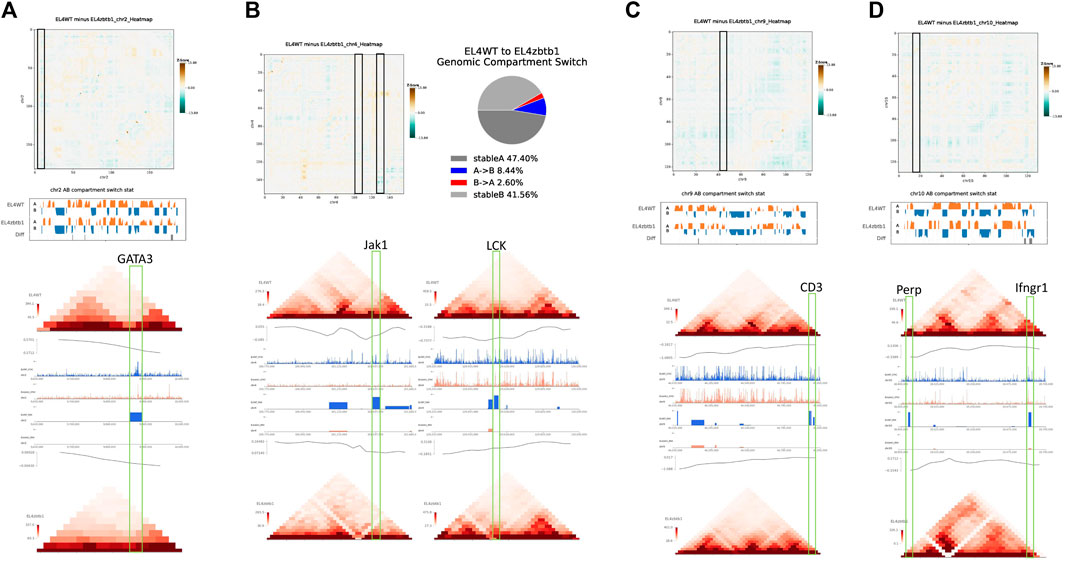
FIGURE 2. For the deletion of Zbtb1 gene the results of the association analysis demonstrated.
Discussion
Our results showed that the deletion of the Zbtb1 gene in EL4 cells led to the downregulation of the expression of many genes and great changes in the spatial structure of the chromatin. As a transcriptional repressor gene, Zbtb1 deletion also led to the upregulation of some genes. However, the ATAC results showed that the open chromatin region of many upregulated genes did not change significantly, which may be due to the restriction of gene expression due to transcriptional inhibition. After the deletion of the Zbtb1 gene, the inhibition was relieved, resulting in the upregulation of related gene expression, but it is difficult to say whether it is direct or indirect regulation.
Our experimental results for the first time explained the important effects of the Zbtb1 gene on T cell development, lymphocyte production and apoptosis from the aspects of chromosome structure and chromatin spatial changes, which provided a relevant theoretical basis and data support for a future in-depth study of Zbtb1.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
XC, CW and YZ conceived and designed research. JW and CS conducted experiments. YZ analyzed data. JW wrote the manuscript. YL, MC, XZ, FL, YS, XAL, XNL contributed to the work. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (81760287, 31941018, and 32072888) and the Science and Technology Development Program of Jilin Province (20190301042NY, 20200402041NC and YDZJ202102CXJD029), Science and Technology Project of the Education Department of Jilin Province during the 13th Five-year Plan (JJKH20200360KJ), China Agriculture Research System of MOF and MARA.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank XC and CW for their support in the experiments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2022.874525/full#supplementary-material
References
Bevington, S. L., Keane, P., Soley, J. K., Tauch, S., Gajdasik, D. W., Fiancette, R., et al. (2020). IL-2/IL-7-inducible Factors pioneer the Path to T Cell Differentiation in advance of Lineage-Defining Factors. EMBO J. 39 (22), e105220. doi:10.15252/embj.2020105220
Buenrostro, J. D., Wu, B., Chang, H. Y., and Greenleaf, W. J. (2015). ATAC-seq: A Method for Assaying Chromatin Accessibility Genome-wide. Curr. Protoc. Mol. Biol. 109, 21–29. doi:10.1002/0471142727.mb2129s109
Cao, X., Lu, Y., Zhang, X., and Kovalovsky, D. (2016). Zbtb1 Safeguards Genome Integrity and Prevents P53-Mediated Apoptosis in Proliferating Lymphoid Progenitors. J. Immunol. 197. 1199–1211. doi:10.4049/jimmunol.1600013
Cheng, Z.-Y., He, T.-T., Gao, X.-M., Zhao, Y., and Wang, J. (2021). ZBTB Transcription Factors: Key Regulators of the Development, Differentiation and Effector Function of T Cells. Front. Immunol. 12, 713294. doi:10.3389/fimmu.2021.713294
Chiliński, M., Sengupta, K., and Plewczynski, D. (2021). From DNA Human Sequence to the Chromatin Higher Order Organisation and its Biological Meaning: Using Biomolecular Interaction Networks to Understand the Influence of Structural Variation on Spatial Genome Organisation and its Functional Effect. Semin. Cel. Dev. Biol.
Dixon, J. R., Selvaraj, S., Yue, F., Kim, A., Li, Y., Shen, Y., et al. (2012). Topological Domains in Mammalian Genomes Identified by Analysis of Chromatin Interactions. Nature 485 (7398), 376–380. doi:10.1038/nature11082
Fortin, J.-P., and Hansen, K. D. (2015). Reconstructing A/B Compartments as Revealed by Hi-C Using Long-Range Correlations in Epigenetic Data. Genome Biol. 16, 180. doi:10.1186/s13059-015-0741-y
Jiang, M., Cai, R., Wang, J., Li, Z., Xu, D., Jing, J., et al. (2021). ILC2 Cells Promote Th2 Cell Differentiation in AECOPD through Activated Notch-GATA3 Signaling Pathway. Front. Immunol. 12, 685400. doi:10.3389/fimmu.2021.685400
Kim, H., Dejsuphong, D., Adelmant, G., Ceccaldi, R., Yang, K., Marto, J. A., et al. (2014). Transcriptional Repressor ZBTB1 Promotes Chromatin Remodeling and Translesion DNA Synthesis. Mol. Cel. 54 (1), 107–118. doi:10.1016/j.molcel.2014.02.017
Lafontaine, D. L., Yang, L., Dekker, J., and Gibcus, J. H. (2021). Hi-C 3.0: Improved Protocol for Genome-wide Chromosome Conformation Capture. Curr. Protoc. 1 (7), e198. doi:10.1002/cpz1.198
Liu, Q., Yao, F., Wang, M., Zhou, B., Cheng, H., Wang, W., et al. (2011). Novel Human BTB/POZ Domain-Containing Zinc finger Protein ZBTB1 Inhibits Transcriptional Activities of CRE. Mol. Cel Biochem 357, 405–414. doi:10.1007/s11010-011-0911-5
Lu, Y., Zhang, X., Bouladoux, N., Kaul, S. N., Jin, K., Sant’Angelo, D., et al. (2017). Zbtb1 Controls NKp46+ ROR-Gamma-T+ Innate Lymphoid Cell (ILC3) Development. Oncotarget 8 (34), 55877–55888. doi:10.18632/oncotarget.19645
Matic, I., Schimmel, J., Hendriks, I. A., van Santen, M. A., van de Rijke, F., van Dam, H., et al. (2010). Site-specific Identification of SUMO-2 Targets in Cells Reveals an Inverted SUMOylation Motif and a Hydrophobic Cluster SUMOylation Motif. Mol. Cel. 39 (4), 641–652. doi:10.1016/j.molcel.2010.07.026
Punwani, D., Simon, K., Choi, Y., Dutra, A., Gonzalez-Espinosa, D., Pak, E., et al. (2012). Transcription Factor Zinc finger and BTB Domain 1 Is Essential for Lymphocyte Development. J. Immunol. 189 (3), 1253–1264. doi:10.4049/jimmunol.1200623
Roberts, O., and Paraoan, L. (2020). PERP-ing into Diverse Mechanisms of Cancer Pathogenesis: Regulation and Role of the P53/p63 Effector PERP. Biochim. Biophys. Acta (Bba) - Rev. Cancer 1874 (1), 188393. doi:10.1016/j.bbcan.2020.188393
van Berkum, N. L., Lieberman-Aiden, E., Williams, L., Imakaev, M., Gnirke, A., Mirny, L. A., et al. (2010). Hi-C: a Method to Study the Three-Dimensional Architecture of Genomes. J. Vis. Exp. (39). doi:10.3791/1869
Wang, J.-H., Shi, C.-W., Lu, Y.-Y., Zeng, Y., Cheng, M.-Y., Wang, R.-Y., et al. (2021). MicroRNA and circRNA Expression Analysis in a Zbtb1 Gene Knockout Monoclonal EL4 Cell Line. Front. Cel. Infect. Microbiol. 11, 706919. doi:10.3389/fcimb.2021.706919
Keywords: Zbtb1, EL4, Hi-C, ATAC-seq, RNA-seq
Citation: Wang J, Shi C, Cheng M, Lu Y, Zhang X, Li F, Sun Y, Li X, Li X, Zeng Y, Wang C and Cao X (2022) Effects of the Zbtb1 Gene on Chromatin Spatial Structure and Lymphatic Development: Combined Analysis of Hi-C, ATAC-Seq and RNA-Seq. Front. Cell Dev. Biol. 10:874525. doi: 10.3389/fcell.2022.874525
Received: 15 February 2022; Accepted: 30 March 2022;
Published: 25 April 2022.
Edited by:
Ann-Kristin Östlund Farrants, Stockholm University, SwedenReviewed by:
Peter Krijger, Hubrecht Institute (KNAW), NetherlandsCopyright © 2022 Wang, Shi, Cheng, Lu, Zhang, Li, Sun, Li, Li, Zeng, Wang and Cao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yan Zeng, emVuZ3lhbkBqbGF1LmVkdS5jbg==; Chunfeng Wang, d2FuZ2NodW5mZW5nQGpsYXUuZWR1LmNu; Xin Cao, eGluY0BqbGF1LmVkdS5jbg==
†These authors have contributed equally to this work