
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol., 23 December 2022
Sec. Epigenomics and Epigenetics
Volume 10 - 2022 | https://doi.org/10.3389/fcell.2022.1065837
This article is part of the Research TopicEpigenetic Factors in Neurodevelopment and Neurodegenerative DiseasesView all 6 articles
 Nasrin Ahangar Davoodi1Sajad Najafi2Zari Naderi Ghale-Noie3Ashkan Piranviseh4Samaneh Mollazadeh5Sahar Ahmadi Asouri6Zatollah Asemi6Mohammadamin Morshedi7,8Seyed Saeed Tamehri Zadeh9
Nasrin Ahangar Davoodi1Sajad Najafi2Zari Naderi Ghale-Noie3Ashkan Piranviseh4Samaneh Mollazadeh5Sahar Ahmadi Asouri6Zatollah Asemi6Mohammadamin Morshedi7,8Seyed Saeed Tamehri Zadeh9 Michael R. Hamblin10Amirhossein Sheida7,8*
Michael R. Hamblin10Amirhossein Sheida7,8* Hamed Mirzaei6*
Hamed Mirzaei6*Retinoblastoma (RB) is a rare aggressive intraocular malignancy of childhood that has the potential to affect vision, and can even be fatal in some children. While the tumor can be controlled efficiently at early stages, metastatic tumors lead to high mortality. Non-coding RNAs (ncRNAs) are implicated in a number of physiological cellular process, including differentiation, proliferation, migration, and invasion, The deregulation of ncRNAs is correlated with several diseases, particularly cancer. ncRNAs are categorized into two main groups based on their length, i.e. short and long ncRNAs. Moreover, ncRNA deregulation has been demonstrated to play a role in the pathogenesis and development of RB. Several ncRNAs, such as miR-491-3p, miR-613,and SUSD2 have been found to act as tumor suppressor genes in RB, but other ncRNAs, such as circ-E2F3, NEAT1, and TUG1 act as tumor promoter genes. Understanding the regulatory mechanisms of ncRNAs can provide new opportunities for RB therapy. In the present review, we discuss the functional roles of the most important ncRNAs in RB, their interaction with the genes responsible for RB initiation and progression, and possible future clinical applications as diagnostic and prognostic tools or as therapeutic targets.
Retinoblastoma (RB) accounts for 3% of all pediatric malignancies, and is the most prevalent intraocular malignant tumor (Rao and Honavar, 2017; Dimaras and Corson, 2019). There are no racial, regional, or gender differences in the incidence of retinoblastoma, which is roughly 1/15,000 across the world (Maheshwari and Finger, 2018). For unilateral retinoblastoma, the average age of diagnosis is 24 months, compared to 12 months for bilateral retinoblastoma. Bilateral tumors occur in between 30% and 40% of retinoblastoma cases. Most cases have been reported to be sporadic (94%), while only 6% of new cases were determined to be familial (Bishop and Madson, 1975; Maheshwari and Finger, 2018). This malignant tumor accounts for 1% of all infant deaths, and can be extremely malignant owing to intracranial metastasis (He et al., 2014). Retinoblastoma typically progresses rapidly without treatment, and damages the structure of the eyeball resulting in blindness (Aerts et al., 2015). Moreover, the tumor can directly invade the brain via the optic nerve, or spread to other parts of body via the blood, including bones, lungs, and other distant organs (Correa-Acosta et al., 2018). It imposes a heavy emotional, financial, and medical burden on patients and society, as well as affecting their long-term health and the quality of life. The survival rate of patients with retinoblastoma is low, despite some recent significant advances in treatment. One reason is that early disease detection seems to be difficult to achieve (Park et al., 2014). Hence, early diagnosis and prompt treatment are crucial in order to prevent loss of vision and metastasis of retinoblastoma. Around 75% of the entire human genome can be transcribed into RNAs, although only 3% can be transcribed into protein-coding mRNAs (Kimura, 2020). Non-coding RNAs (ncRNAs) are classified into distinct types based on their length, structure, and location. The three main types of ncRNA are, circular RNAs (circRNA), long ncRNAs (lncRNA), and microRNAs (miRNA). MiRNAs are small RNAs with a typical length of 22 nucleotides (nt). After binding of miRNAs to the complementary base sequence in the target mRNAs, they are is degraded by the RNA-induced silencing complex (RISC) (Vos et al., 2019). LncRNAs and circRNAs are both longer than 200 nt, however circRNAs are closed circles whereas lncRNAs are linear. LncRNAs and circRNAs can both arise by transcription of exons, introns, intergenic regions, or 5′/3′-untranslated regions of various genes. However, they can fold to produce complex secondary structures which can interact with proteins, DNA, or RNA (Invalid Citationa). LncRNAs and circRNAs can regulate expression of genes via various mechanisms. To prevent the targeted mRNA from being degraded, they can act as miRNA decoys or sponges. Furthermore, they can alter the affinity of transcription factors for binding to their promoters, in order to regulate the expression of specific genes (Zhao et al., 2014). In the present review, we discuss the role of various non-coding RNAs in retinoblastoma.
MicroRNAs (miRNAs) are a group of endogenous non-coding small RNAs with the ability to regulate the expression of their target genes (Lytle et al., 2007). They directly interact with the 3′-untranslated region of mRNA, and then negatively affect the expression of the target genes, either by mRNA degradation or suppression of translation (Guarnieri and DiLeone, 2008). Based on the miRBase database, the human genome includes 4,469 genes which encode miRNAs, consisting of 1,881 precursor and 2,588 mature miRNAs. Various types of miRNAs have been found to be abnormally expressed in RB samples. These altered miRNAs can affect many cancer-related processes, including the epithelial-mesenchymal transition, cell proliferation and survival, apoptosis, metastasis, and angiogenesis (Delsin et al., 2019). MiR-98 (Reese et al., 2005) and miR-186 (Davidson et al., 2004) are two examples of miRNAs that are overexpressed in RB and contribute to the malignancy of the tumor, whereas miR-98 (Reese et al., 2005), miR-186(18), and miR-106b (Reese et al., 2006) are expressed only at low levels in RB and are therefore expected to act as tumor inhibitors.
Stannin (SNN), which has a single transmembrane helix, an unstructured linker domain, and a cytoplasmic domain, is a highly conserved protein in vertebrate animals. It has previously been reported to be strongly associated with the toxicity of trimethyltin (TMT) and has a significant effect on apoptosis (Reese et al., 2005). It has been demonstrated that SNN plays a critical role in the toxic effect of organotin (Davidson et al., 2004) and endosomal maturation (Pueyo et al., 2016). Many investigations have also suggested the role of SNN in tumor growth (Reese et al., 2006). In a study by Reese et al. it was demonstrated that protein kinase C could regulate tumor necrosis factor-alpha (TNF-α) leading to increased expression of SNN (Reese et al., 2006). SNN plays a crucial role in MAPK signaling pathways (Huang et al., 2016), and the MAPK signaling pathway is connected to a number of cancers. The expression of SNN may be affected by certain miRNAs. The effects of miR-491-3p on proliferation, migration, invasion, and apoptosis in RB cells was examined in a study by Hu et al. (Hu et al., 2021). Primary RB tissue samples from 15 patients were collected as well as paired adjacent non-cancerous tissues. Quantitative real-time PCR (qRT-PCR) was used to examine the expression profile of miR-491-3p. In order to investigate the expression profiles of epithelial–mesenchymal transition (EMT)-related proteins (vimentin, E-cadherin, and N-cadherin) in RB tissues and RB cell lines they performed qRT-PCR, Western blotting and in situ immunocytochemistry. Cell proliferation was measured using MTS and colony formation assays. Apoptosis was determined using FACS, while cell migration and invasion were examined using transwell chambers. Target gene prediction databases were used to predict the genes that miR-491-3p would bind to. To evaluate the relationship between miR-491-3p and SNN, dual luciferase reporter assays were carried out. In 15 pairs of Rb tissues as well as RB cell lines it was found that miR-491-3p was noticeably downregulated.
Artificial overexpression of miR-491-3p dramatically suppressed RB cell proliferation, migration, and invasion, while it increased apoptosis. By contrast, treatment with a miR-491-3p inhibitor produced the opposite results, leading to less apoptosis, and increased proliferation in ARPE-19 cells. Additionally, miR-491-3p altered the morphology of RB cell lines resulting in cells that were more adherent, and it significantly reduced the expression of vimentin and N-cadherin while increasing E-cadherin suggesting it could inhibit the EMT. The SNN was identified as a miR-491-3p base-pairing target, and the two molecules could interact with each other. They also discovered that the overexpression of SNN almost completely neutralized the effects of miR-491-3p in RB cells. In total, their findings suggested that miR-491-3p acts as a tumor suppressor gene in RB, and could inhibit the growth and metastasis of tumors as a novel therapeutic target in RB (Hu et al., 2021).
Wnt-inhibitory factor-1 (WIF1) acts as a secreted repressor protein that can directly bind to various ligands of the Wnt signaling pathway, and inhibit their activity (Hsieh et al., 1999; Poggi et al., 2018). In previous studies, WIF1 was shown to be involved in a variety of biological processes including tumor growth, cell cycle, metastasis, apoptosis, and stemness (Wu et al., 2012a; Ramachandran et al., 2012; Ramachandran et al., 2014; Wang et al., 2015; Huang et al., 2016). In numerous human cancers, WIF1 promoter hypermethylation was linked to a poor prognosis (Lee et al., 2013; Roperch et al., 2013; Zhao et al., 2017a; Lin et al., 2017; Zhou et al., 2018). Additionally, a number of ncRNAs such as HOTAIR, HOXC6, miR-181a, miR-552, miR-590-3p, and miR-603, have been shown to regulate WIF1 expression at the transcriptional level (Ji et al., 2014; Guo et al., 2015; Jiang et al., 2016; Feng et al., 2017a; Li et al., 2018; Yan et al., 2018). Gastric tumors have been linked to hypermethylation of the WIF1 promoter, which inhibits its expression and abnormally activates the Wnt signaling pathway (Poggi et al., 2018). Moreover, WIF1 hypermethylation has been shown to be correlated with poor survival rate in some cancer patients, such as non-small-cell lung cancer (Guo et al., 2017), esophageal squamous cell carcinoma (Guo et al., 2016), and chondrosarcoma (Liu et al., 2017).
Using the GEO datasets, Li et al. set up an interaction network involving numerous distinct genes and miRNAs that were selectively expressed in retinoblastoma (Fuchs et al., 2001). They found that RB cell lines and tissues highly expressed miR-340, and up-regulation of miR-340 was associated with a poor prognosis in patients. The Gene Expression Omnibus (GEO) repository was used to retrieve differentially expressed miRNAs (DEmiRs) and genes (DEGs) in retinoblastoma. Besides, qRT-PCR was used to determine the expression of WIF1 and miR-340 in RB tissues and cell lines. Gain-of-function and loss-of-function assays were used to examine the effects of miR-340 on cell proliferation, migration, and invasion. Bioinformatics analysis and luciferase reporter experimental assays were used to investigate the interaction between miR-340 and WIF. Blood and tissue samples from retinoblastoma patients displayed a total of 11 DEmiRs. They confirmed that miR-340 was the most abundantly expressed miRNA, and was associated with ICRB stage, tumor size, and optic nerve invasion. They proposed that miR-340 could promote RB cell proliferation, migration, and invasion. They subsequently set up a regulatory network of miRNA-mRNA pairs after identifying 26 DEGs from three retinoblastoma GEO datasets. Further research revealed that miR-340 directly targeted WIF1. Furthermore, WIF1 overexpression might inhibit retinoblastoma progression induced by miR-340 in vitro and in vivo. Overall, miR-340 modulates WIF1 and then acts as an oncomiRNA to enhance RB cell proliferation, migration, and invasion. They also identified multiple miRNAs and genes that could help researchers better understand retinoblastoma pathogenesis (Fuchs et al., 2001).
E2F5 is a transcription factor and a member of the E2 promoter binding factor (E2F) family, involved in cell proliferation (Dimova and Dyson, 2005; Cao et al., 2010; Wan et al., 2010). Basically, E2F5 links the cell cycle to post-transcriptional pathways, so it is essential for regulating cell growth, and also affects biological processes involved in cancer development (Ishimoto et al., 2013; Xie et al., 2020). Inhibition of E2F5 may be able to reduce metastasis and growth of gastric cancer, pointing to a possible role for this protein in new treatments (Yao and Yan, 2018). E2F5 may be involved in the development of other cancers, such as hepatocellular carcinoma, colon, breast, ovarian, osteogenic sarcoma, and esophageal squamous cell carcinoma (Fuchs et al., 2001; Lassmann et al., 2007; Umemura et al., 2009; Kothandaraman et al., 2010; Jiang et al., 2011). E2F5 expression was found to be increased in RB where it affected the cell cycle, growth and proliferation (Zhang et al., 2017a).
Zhang et al. measured the expression of miR-613 in human RB samples and investigated its function in RB cells (Zhang et al., 2017a). They found that the level of miR-613 was lower in RB tissues and cell lines. In vitro studies showed that overexpression of miR-613 led to suppression of RB cell proliferation, migration, and invasion, as well as producing cell cycle arrest. Besides, miR-613 overexpression prevented retinoblastoma cells from developing tumors in vivo. They also showed that E2F5 was a direct target of miR-613. The introduction of E2F5 mRNA without a 3′-untranslated region abrogated the effects miR-613 on proliferation and invasion. Their findings showed that miR-613 acts as a tumor suppressor in retinoblastoma by downregulating E2F5. Their data supported the idea of targeting the miR-613/E2F5 axis as a potential therapeutic approach for retinoblastoma (Table 1) (Zhang et al., 2017a).
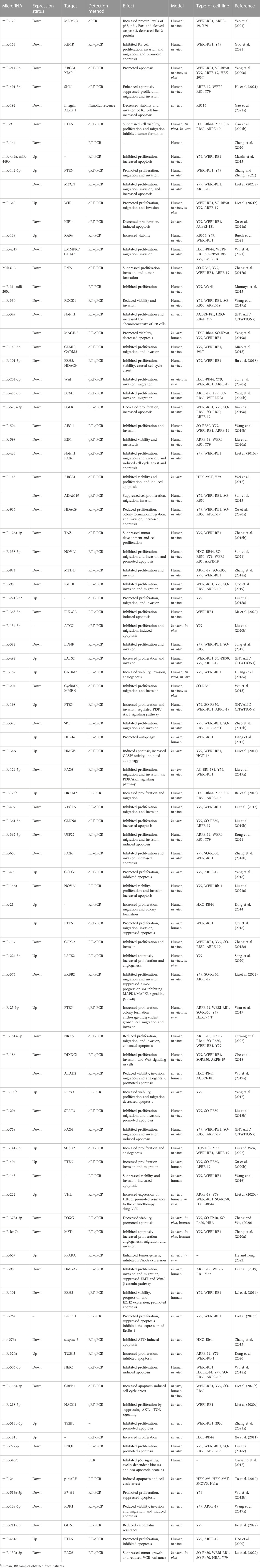
TABLE 1. The role of miRNAs in retinoblastoma initiation and progression.
HDAC9 is a member of the histone deacetylase (HDAC) family, involved in transcriptional regulation, cell death, and especially in carcinogenesis and cancer progression (Marks et al., 2004; Dokmanovic and Marks, 2005; Singh et al., 2017). The level of HDAC9 was higher in RB samples, and this upregulation was correlated with tumor size, regional lymph node metastasis, and poor tumor differentiation (Zhang et al., 2016a). Patients with RB tunors with high levels of HDAC9 had shorter overall survival and progression-free survival, compared to patients who expressed low levels of HDAC9 (Zhang et al., 2016a). HDAC9 has been hypothesized to act as an oncogene in the onset of RB and its progression by affecting a range of pathophysiological pathways (Mohammad et al., 2017; Jin et al., 2018). HDAC9 has the ability to reduce EGFR expression and hence inhibit the activation of the downstream PI3K/AKT signaling pathway, which leads to the development of cancer (Watson et al., 2013).
MiR 936 expression was quantified in RB by Xu et al. Importantly, they identified the contribution of miR-936 to RB oncogenesis, and suggested some underlying molecular pathways (Sheets et al., 2020). Reverse-transcription quantitative PCR was used to assess miR-936 expression in RB tissues and cell lines. Various tests including cell counting kit 8, flow cytometry, invasion, migration, and colony formation assays, as well as subcutaneous heterotopic xenografts were used to assess cell proliferation, colony formation, apoptosis, migration, and invasion in vitro, and tumor growth in vivo. Bioinformatic analysis was used to determine the possible targets of miR-936, and RT-qPCR, luciferase reporter assays and Western blotting were used to confirm the targets. Low levels of miR-936 in RB were correlated with lymph node metastasis, differentiation, and TNM stage, in both RB tissues and cells. Exogenous miR-936 inhibited RB cell proliferation, colony formation, migration, and invasion in vitro, and reduced tumor growth in vivo, because overexpression of miR-936 increased apoptosis. It was found that the histone deacetylase 9 (HDAC9) mRNA was a direct target of miR-936. HDAC9 depletion produced effects on RB cells that were comparable to those of miR-936 overexpression. The restoration of HDAC9 expression led to a decrease in the tumor-suppressor effects of miR-936, and increased the oncogenicity of RB cells. Exogenous expression of miR-936 inhibited the activity of the PI3K/AKT pathway both in vitro and in vivo, by suppressing the expression of HDAC9 in RB cells. A poor prognosis in RB patients was correlated with downregulation of miR-936, and its overexpression decreased RB aggressiveness by reducing HDAC9 mRNA and subsequently inactivating the PI3K/AKT pathway (Xu et al., 2020a).
The polycomb repressor complex 2 catalytic core protein, also known as enhancer of zeste homolog 2 (EZH2), is a highly conserved histone methyltransferase (Kuzmichev et al., 2004). By catalyzing the trimethylation of histone H3 K27, EZH2 suppressed the expression of its target genes (Cheng et al., 2016). EZH2 has been found to be upregulated in a variety of cancers, including colorectal, prostate, and breast cancer (Varambally et al., 2002; Kleer et al., 2003; Fluge et al., 2009). According to a study by Khan et al. (Khan et al., 2015), the level of EZH2 was increased in RB specimens. It is also well known that EZH2 stimulates the growth of cancer cells. Lian et al. (Gao et al., 2021a) demonstrated that EZH2 modulated Runt-related transcription factor 3 expression in order to promote the proliferation of laryngeal cancer cells. In addition, small-molecule EZH2 inhibitors suppressed tumor growth by inducing the tumor-suppressor protein p16INK4A (Mohammad et al., 2017).
Jin et al. investigated the role of miR-101-3p in retinoblastoma and tested the hypothesis that miR-101-3p targeted histone deacetylase 9 (HDAC9) as well as EZH2 (Jin et al., 2018). Retinoblastoma specimens had been shown to have downregulated expression of miR-101-3p. MTT and flow cytometry assays were used to show that exogenous overexpression of miR-101-3p significantly inhibited viability and caused cell cycle arrest in WERI-Rb-1 and Y79 cells. In vivo mouse studies supported the anti-tumor activity of miR-101-3p in retinoblastoma. Furthermore, predictions from TargetScan software suggested that miR-101-3p would target the 3′-untranslated regions of both HDAC9 and EZH2 mRNAs. The results obtained from the dual luciferase reporter assay showed that miR-101-3p directly targeted EZH2 and HDAC9 to inhibit the proliferation of RB cells. The anti-proliferative effects of miR-101-3p in WERI-RB-1 and Y79 cells were abrogated by the restoration of EZH2 or HDAC9 expression. These findings underline the importance of miR-101-3p in the growth of RB tumors, and suggest a potential new therapeutic target (Zheng et al., 2020).
Sushi domain-containing 2 (SUSD2) is a type I transmembrane protein that contains an AMOP (adhesion-associated domain present in MUC4 and other proteins) domain. The AMOP domain is also found in somatomedin B and von Willebrand factor type D, which are proteins involved in cell-cell and cell-matrix adhesion (Watson et al., 2013). Previous studies have shown that SUSD2 can function as a tumor suppressor in a wide range of cancers. It was found that SUSD2 could play both a negative and a positive role in high-grade serous ovarian cancer. SUSD2 was negatively associated with metastasis, but was positively associated with a longer survival time (Sheets et al., 2020). Additionally, lung adenocarcinoma patients with low SUSD2 expression levels have shorter survival, so SUSD2 may function as an independent prognostic indicator (Guo et al., 2020). In lung cancer and RCC, lower SUSD2 expression also acted as a tumor suppressor (Cheng et al., 2016).
Liu et al. performed both in vitro and in vivo studies on the development of retinoblastoma and the involvement of angiogenesis. In these studies, they investigated the putative function and mechanism of the miR-141-3p/SUSD2 axis (Liu and Wen, 2022). They used the Gene Expression Omnibus (GEO) datasets, to identify the expression levels of SUSD2 and miR-141-3p in retinoblastoma samples compared to healthy controls. The biological role and molecular mechanism of the miR-141-3p/SUSD2 axis in RB development and progression were investigated using bioinformatics analysis, a dual-luciferase reporter assay, and loss, gain, and rescue of function assays. Their findings demonstrated that RB tissues and cells expressed significantly lower levels of SUSD2. In vitro, an increased level of SUSD2 inhibited RB cell viability, promoted apoptosis, and prevented primary human umbilical vein endothelial cells (HUVECs) from forming tubes. The results of the dual-luciferase reporter assay and bioinformatics analysis demonstrated that miR-141-3p directly regulated SUSD2. MiR-141-3p overexpression accelerated angiogenesis, whereas miR-141-3p suppression inhibited RB development. Both in vivo and in vitro overexpression of SUSD2 partially reversed these effects. They concluded that SUSD2 acts as a RB tumor suppressor, and the miR-141-3p/SUSD2 axis could regulate both retinoblastoma development and angiogenesis, making it a new target for RB treatment (Liu and Wen, 2022).
Long non-coding RNAs (lncRNAs) are more than 200 nucleotides in length (Silva et al., 2019; Wang et al., 2021a). LncRNAs are involved in many pathways that affect tumor biology, and have major roles in controlling oncogenes as well as tumor suppressor genes (Huang et al., 2017). For instance, LINC00504 was increased in breast cancer, where it promotes tumor cell proliferation and migration (Hou et al., 2021). By acting as a competing endogenous RNA (ceRNA) or sponge, lncRNAs can affect gene transcription. CeRNAs can specifically sponge their matching miRNA to control expression of the mRNAs of the relevant target genes (Chen et al., 2021a; Wang et al., 2021b). These regulated target genes can play a role in several signaling pathways, almost all of which are strongly linked to tumors (Gao et al., 2021c). It has been confirmed that certain lncRNAs are involved in the progression of human tumors (Tsai et al., 2011; Tang et al., 2013), including retinoblastoma (Wang et al., 2018a). Numerous human cancers, such as hepatocellular carcinoma (HCC) (Li and Zheng, 2017), colorectal cancer (CRC) (Yan et al., 2020), and prostate cancer have been shown to be affected by the oncogenic roles of the actin-binding LIM and SH3 domain protein 1 (LASP1) (Wang et al., 2017c). In an investigation into oral cancer, it was revealed that silencing of LASP1 resulted in cell cycle arrest in G2 phase (Shimizu et al., 2013). Additionally, in clear cell renal cell carcinoma, LASP1 silencing inhibited cell migration (Yang et al., 2014). MiRNAs have been hypothesized to regulate LASP1 in the initiation and progression of various cancers (INVALID CITATIONa; Hu et al., 2017a). The development of oral squamous cell carcinoma was affected by LASP1, a miR-342-3p downstream target, according to a study by Song et al. (Nordlund and Reichard, 2006). Yang et al. (Yang et al., 2019b) also showed that lncRNA SNHG16 control led LASP1 to promote RB cell invasion and migration.
Xu et al. examined the regulatory mechanism and potential role of the lncRNA, myocardial infarction-associated transcript (MIAT) in RB progression (Xu et al., 2021b). The expression of MIAT, miR-665, LASP1, and other proteins were measured using RT-qPCR in RB cells or patient tissues. The dual-luciferase reporter assay was used to confirm the interactions between miR-665 and MIAT/LASP1. Western blotting, MTT and transwell assays were used in vitro to investigate the effects of the MIAT/miR-665/LASP1 axis on RB development. Their findings showed that MIAT specifically targeted miR-665. High MIAT expression was found in cell lines and RB tissues, while miR-665 was downregulated in RB tissues. In addition, either miR-665 overexpression or MIAT downregulation led to reduced proliferation, migration, and invasion of RB cells, HXO-RB44 and Y79. LASP1 was also suggested to be a target gene for miR-665. Both downregulation of miR-665 and upregulation of LASP1 reversed the suppressive effects of MIAT knockdown on the proliferation, migration, and invasion of Y79 cells. Additionally, MIAT silencing prevented RB proliferation by regulating the miR-665/LASP1 axis. They suggested that MIAT might be a promising target for RB treatment (Grossi et al., 2015).
Brain-derived neurotrophic factor (BDNF) is involved in the survival, differentiation, growth, and development of neurons (INVALID CITATIONa; McAllister, 2001). In cancer biology, the expression of BDNF was found to be elevated in gliomas, breast cancer, colorectal cancer, gastric cancer, bladder cancer, and other human tumors (Colucci-D’Amato et al., 2020). In colon cancer, BDNF promoted cancer progression by increasing the expression of HO-1 and transcription of VEGF, as well as the activation of the MAPK signaling pathway (Huang et al., 2015). BDNF promoted cancer development by activating tropomyosin receptor kinase B (TrkB) in triple-negative breast cancer (Tsai et al., 2017). High levels of BDNF increased the proliferation and invasion of RB cells by triggering the PI3K/AKT signaling pathway (INVALID CITATIONa).
Xu et al. investigated the function of the lncRNA XIST, and its underlying mechanism in RB (Xu et al., 2021c). In this investigation, RT-qPCR and Western blotting were used to measure the levels of XIST, miR-191-5p, BDNF mRNA, and BDNF protein in RB tissues and cell lines. In order to carry out gain-of-function and loss-of-function experiments, they transfected pcDNA3.1-XIST, XIST siRNA, and miR-191-5p mimics and inhibitors into SO-Rb50 and Y79 cells. Several methods including CCK-8, transwell, and terminal deoxynucleotide transferase UTP nick-end labeling (TUNEL) experiments were employed to measure RB cell proliferation, invasion, migration, and apoptosis. The use of luciferase reporter assays, RT-qPCR, Western blotting, and bioinformatics analysis, allowed the regulatory relationships between BDNF, XIST, and miR-191-5p to be elucidated. They found that XIST expression was significantly increased in RB tissues and cells. High expression of XIST promoted proliferation, invasion, migration, and inhibited apoptosis in RB cells, but miR-191-5p showed the opposite effects. Furthermore, miR-191-5p inhibited the expression of BDNF at both mRNA and protein levels. XIST indirectly increased the expression of BDNF by acting as a ceRNA and inhibiting miR-191-5p expression. They concluded that the expression level of XIST was increased in RB tissues, and XIST could control proliferation, invasion, migration, and apoptosis through regulating the miR-191-5p/BDNF axis (Xu et al., 2021c).
The hypoxia-inducible factor-1α (HIF-1α) gene is located on chromosome 14q23.2. HIF-1α is a crucial subunit of hypoxia-inducible factor-1 (HIF-1). Numerous studies have confirmed the oncogenic role of HIF-1α. For example, an interaction between HIF-1α and LOXL2 (lysyl oxidase like 2) in hepatocellular carcinoma promoted cancer development, and increased angiogenesis and the EMT (Wang et al., 2017b). Elevated expression of HIF-1α in thyroid follicular carcinoma was associated with distant metastasis (Klaus et al., 2018). HIF-1α also acts as an oncoprotein in RB, and its knockdown led to increased expression of pro-apoptotic proteins, including Bax, caspase-9, and caspase-3, thus triggering apoptosis in cancer cells (Gao et al., 2014). In addition, HIF-1α increased invasion of RB cells by inducing MMP-9 expression. (Li and Zheng, 2017). Besides, it has been shown that activation of HIF-1α can increase resistance to various cancer therapies (Liang et al., 2013; Wang et al., 2017c). Investigations by Dong et al. (Wang et al., 2017c) and Greco and Scott (Greco and Scott, 2007) both showed that HIF-1α plays a role in the increased survival of tumor cells in response to chemotherapy and/or radiotherapy.
Yan et al. (Yan et al., 2022) investigated the role of the lncRNA MIR17HG (miR-17-92a-1 cluster host gene) and its interaction with miR-155-5p and HIF-1α pathway in RB development. qRT-PCR showed that up-regulation of MIR17HG was negatively associated with miR-155-5p expression. CCK-8 and transwell assays showed that overexpression of MIR17HG increased proliferation, migration, and invasion of RB cells. MiR-155-5p was a MIR17HG target which could inhibit growth, migration, and invasion of RB cells. Furthermore, MIR17HG enhanced the gene and protein expression of HIF-1α in RB cells. Taken together, this study suggested the oncogenic effects of MIR17HG in RB mediated through the miR-155-5p/HIF-1α axis (Yan et al., 2022).
Ribonucleotide reductase M2 subunit (RRM2) catalyzes the rate-limiting step of DNA synthesis and repair, and has important effects on multiple cell processes, including proliferation, migration, invasion, and senescence (Nordlund and Reichard, 2006). In addition, upregulation of RRM2 acted as a cancer driver in different malignancies (Grossi et al., 2015; Grolmusz et al., 2016). In adrenocortical cancer, RRM2 was strongly associated with Ki67 expression (Grolmusz et al., 2016). In pancreatic cancer, RRM2 increased the expression of Bcl-2, an anti-apoptotic protein, and decreased cleaved caspase-3 (Xia et al., 2017). In RB, enhanced expression of RRM2 affected RB cell cycle progression (Nie et al., 2021).
To investigate the underlying mechanism of the lncRNA HOTAIR in RB development, Fu et al., (Fu et al., 2022), showed that HOTAIR was the upregulated in RB cells (ORB50, Y79, HXO-RB44, and WERI-RB) in comparison with normal retinal cells (ARPE-19 and RPE-1). HXO-RB44 and Y79 cells had relatively higher expression of this marker. In the next step, delivery of sh-HOTAIR into RB cells (Y79 and HXO-RB44) arrested the cell-cycle, inhibited proliferation, and increased apoptosis, as detected by CCK-8 assay and flow cytometry. Dual-luciferase assay reveled that HOTAIR acted as a ceRNA of miR-20b-5p and could also increase the expression of RRM2. Besides, functional rescue experiments showed that downregulation of miR-20b-5p or upregulation of RRM2 could promote proliferation and the RB cell-cycle, inhibit apoptosis, and reverse the effects of sh-HOTAIR on inhibition of RB cells. Treatment of a xenograft tumor model with sh-HOTAIR not only decreased tumor development and the number of Ki67 + cells, but also inactivated the PI3K/AKT axis. LncRNA HOTAIR competitively bound to miR-20b-5p, and therefore upregulated RRM2 and activated the PI3K/AKT pathway to enhance RB cell proliferation and suppress apoptosis (Fu et al., 2022).
High-mobility group box 1protein (HMGB1) is a nuclear protein with cytokine-like activity, which was secreted by neurons following ethanol exposure (Crews et al., 2013; Zou and Crews, 2014). HMGB1 can activate immune responses to TLR7 agonists (Yanai et al., 2009), and functions as a chaperone for cytokines or DNA, and improves their interactions with their normal receptors (Boonyaratanakornkit et al., 1998; Sha et al., 2008; Bianchi, 2009). HMGB1 is released inside microvesicles (MVs) from macrophages and its expression and active secretion in the brain are increased in response to ethanol (Ardoin and Pisetsky, 2008; Zou and Crews, 2014). The expression of the HMGB1 gene located chromosomal region 8q22, is increased in RB, and is correlated with poorly differentiated tumor and invasion of the optic nerve (Singh et al., 2015). HMGB1 was found to stimulate RB tumorigenesis and potentiate its malignant properties (Liu et al., 2014; Wang et al., 2017d; Chai et al., 2018; Liu et al., 2018d).
Zhang et al., (Zhang et al., 2020b), examined the expression of LINC00205 in RB specimens to uncover its exact role in RB tumorigenesis. They found that high expression of LINC00205 in RB cells and tissues was associated with unfavorable clinicopathological properties and shorter overall survival times in RB patients. LINC00205 knockdown in vitro inhibited proliferation and stimulated apoptosis in RB cells, while it slowed RB tumor development in vivo. Mechanistically, LINC00205 increased the expression of the miR-665 target mRNA HMGB1, because it acted as a ceRNA to sponge miR-665 in RB cells. If the miR-665-HMGB1 pathway was activated, it weakened the effects of LINC00205 depletion in RB cells. Therefore, the LINC00205/miR-665/HMGB1 axis could be a useful target for RB prognosis, diagnosis, and even therapy (Zhang et al., 2020b).
E2F transcription factor 3 (E2F3) is a member of the E2F family of transcription factors which are classified into three main groups; atypical inhibitors (E2F7 and E2F8), canonical inhibitors (E2F3b-E2F6), and activators (E2F1, E2F2, and E2F3a) of gene transcription (Kent and Leone, 2019). The atypical and the canonical inhibitors prevent gene transcription and act as tumor suppressors (Kent and Leone, 2019). There are lines of evidence showing that miRNAs, such as miR-145-5p can regulate E2F3 (Zehavi et al., 2015; Hu et al., 2017b). E2F3 is involved in various human tumors as a tumor promoter (Wang et al., 2019c). E2F3 was found to be upregulated in RB (Madhavan et al., 2009; Zhao et al., 2020a), while its downregulation was associated with the inhibition of RB cell proliferation (Zhao et al., 2020a).
Zhang et al., (Zhang et al., 2020c), investigated the effects of the lncRNA CASC9 on RB cells malignant properties, such as proliferation, invasion, EMT, and apoptosis. CASC9 overexpression noticeably promoted the malignant phenotype of RB cells. By contrast, silencing of CASC9 suppressed the malignant potential of RB cells and increased apoptosis. RNA immunoprecipitation and dual-luciferase reporter assays demonstrated that CASC9 could regulate E2F3 via sponging miR-145-5p. In more detail, the effects of CASC9 knockdown could be reversed in part by inhibition of miR-145-5p or overexpression of E2F3. Moreover, the overexpression of miR-145-5p could further promote properties already stimulated by CASC9 silencing. All of which suggest that CASC9 could be a therapeutic target to control RB (Table 2) (Zhang et al., 2020c).
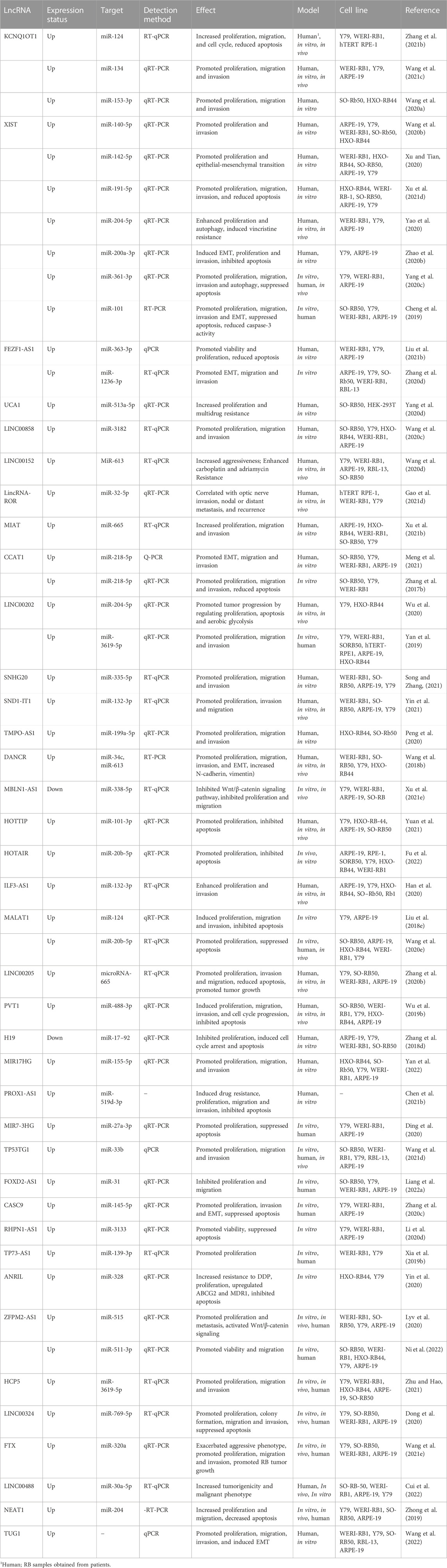
TABLE 2. The role of lncRNAs in retinoblastoma.
CircRNAs are a subclass of endogenous ncRNAs, which were first misinterpreted as by-products of splicing errors (Sanger et al., 1976). Recently however, it has been shown that circRNAs are derived from intronic or exonic sequences by back-splicing, and then form a stable covalently closed circular loop without any 5′ end caps or 3′ end poly(A) tails (Wang et al., 2017e). The majority of circRNAs are evolutionarily conserved, widespread, abundant, and stable, and they show tissue or developmental specificity in eukaryotes (Bahn et al., 2015; Wang et al., 2017f). In contrast to linear RNAs, circRNAs are not sensitive to exonuclease digestion, and can resist the degradation suffered by linear RNA due to their distinctive single-stranded closed circlular loop. Their presence in different samples such as serum, tissues, and urine makes them useful as biomarkers for various human cancers, as well as age-related disorders (Memczak et al., 2013; Fang et al., 2019). It is accepted that circRNAs are involved in several human diseases such as cancer, because of their ability to alter the malignant properties of cancer cells and their response to chemotherapy agents (Li et al., 2015). Therefore, circRNAs could be novel therapeutic targets to manage various tumors (Chen, 2016; Shao et al., 2018; Drula et al., 2020; Li et al., 2020e; Luo et al., 2020) including pancreatic cancer (Sharma et al., 2021).
WNT3A is an important element in the Wnt/β-catenin pathway, which contributes to proliferation, differentiation, and carcinogenesis (Yun et al., 2005; He et al., 2015). WNT3A expression levels were associated with cancer cell proliferation and drug-resistance via Wnt/β-catenin signaling pathway (INVALID CITATIONb). The overexpression of miR-15a-5p could target WNT3A mRNA leading to inhibition of proliferation and stemness of human endometrial adenocarcinoma (HEC-1-A) cells (Zhang et al., 2007). Moreover, miR-485 could inhibit WNT3A in RB cells and further inhibit Wnt/β-catenin signaling in these cells (INVALID CITATIONb).
Wang et al. (Wang et al., 2020f) reported that circDHDDS was upregulated in RB tissues and cells (Y-79, RPCs, and WERI-RB1) when compared to retinal pigment epithelial cells and normal retinal tissues. Colony formation, transwell, and flow cytometry assays showed that knockdown of circDHDDS inhibited the malignant potential of RB cells, and caused cell cycle arrest. Similar results were found in a RB xenograft model following silencing of circDHDDS. To elucidate the molecular mechanism by which circDHDDS promoted RB progression, bioinformatics databases and a dual-luciferase reporter assay revealed that circDHDDS could sponge miR-361-3p which in turn targeted WNT3A. Consequently, miR-361-3p overexpression inhibited WNT3A expression resulting in inhibition of RB progression. Thus, the circDHDDS/miR-361-3p/WNT3A pathway promoted RB progression and increased proliferation, migration, invasion, and the cell cycle in RB cells (Wang et al., 2020f).
It was found that Syntaxin 17 (STX17) could bind to vesicle-associated membrane protein 8 (VAMP8) and synaptosome-associated protein 29 (SNAP29) to promote the fusion of autophagosomes with lysosomes by modulating the autophagosome membrane (Itakura et al., 2012; Uematsu et al., 2017). STX17 is a localized endoplasmic reticulum membrane protein, which affects cell survival via interactions with Fis1, ATG14L, and BAP31 to form a functional complex (Wang et al., 2008; Hamasaki et al., 2013; Machihara and Namba, 2019). Huang et al. investigated the effect of STX1 on RB cell autophagy via the lncRNA MALAT1/miR-124 axis (Huang et al., 2018b). Upregulation of STX17 in RB cells increased autophagy (Huang et al., 2018b).
Liu et al. reported the increased expression of circ_0000034 and STX17 as well as the reduced expression of miR-361-3p in RB cells and tissues. Silencing of circ_0000034 inhibited proliferation, migration, invasion, autophagy, and tumor growth, and induced death in RB cells. The use of dual-luciferase reporter assays and RNA immunoprecipitation demonstrated an interaction between circ_0000034 and miR-361-3p, as well as an interaction between miR-361-3p and STX17. Circ_0000034 overexpression and miR-361-3p depletion both resulted in increased expression of STX17, and promoted the progression of RB. On the other hand, circ_0000034 knockdown inhibited RB progression via modulating the miR-361-3p/STX17 axis (Liu et al., 2020c).
A Disintegrin and Metalloproteinase 19 (ADAM19) is a member of the ADAM family, which is highly expressed in different tumors (Wildeboer et al., 2006; Chan et al., 2008; Zhang et al., 2015a; Zhang et al., 2019), including NSCLC cells (Wang et al., 2019d). ADAM19 is a transmembrane protein which contributes to tumor development, such as glioma and colorectal cancer (Qi et al., 2009). ADAM19 was found to affect the proliferation and invasion of RB cells (Sun et al., 2015). Notably, miR-145 overexpression could target ADAM19 mRNA and reverse its effects on RB development (Sun et al., 2015).
Jiang et al. (Jiang et al., 2021a) used qRT-PCR to show that circ_0000034 expression was increased in RB tissues and cells. Silencing of circ_0000034 not only inhibited proliferation, migration, invasion, and EMT of RB cells as shown by CCK-8 and transwell assays, but also stimulated apoptosis as shown by flow cytometry. RB growth in an animal model was inhibited by silencing of circ_0000034. Using dual-luciferase reporter, RIP, and RNA pull-down assays, it was shown that circ_0000034 could sponge miR-361-3p, and thereby reverse its effects on ADAM19 in RB cells. Moreover, miR-361-3p suppression abolished the effects of silencing circ_0000034 on the malignant properties of RB cells. Overexpression of ADAM19 reversed the effects of the miR-361-3p mimic on the survival, migration, invasion, apoptosis, and EMT of RB cells. They concluded that circ_0000034 promoted RB tumorigenesis via the miR-361-3p/ADAM19 axis, which could therefore be a target for RB therapy (Jiang et al., 2021a).
It has been shown that Rho-associated protein kinase 1 (ROCK1) affects tumor development by regulating cellular processes, including proliferation, migration, invasion, apoptosis, and the EMT (Abe et al., 2014; Zhang et al., 2015b; Xiang et al., 2015; Leonel et al., 2017). Wang et al. reported that ROCK1 knockdown could inhibit the adhesion and invasion of RB cells (Wang et al., 2014). ROCK1 could also increase proliferation and metastasis of RB cells, and inhibit apoptosis (Wu et al., 2018b; Wang et al., 2019a). In terms of function, ROCK1 suppression could decrease the oncogenic potential of RB cells (Wang et al., 2014).
Huang et al. (Huang et al., 2021) investigated the mechanism of circ-E2F3 in RB progression. They used qRT-PCR analysis to show that circ-E2F3 was overexpressed in RB tissues and cells. Circ-E2F3 silencing inhibited proliferation, migration, and invasion of RB cells as shown by MTT, transwell, colony formation, and scratch wound healing assays. In addition, circ-E2F3 knockdown increased apoptosis in RB cells as shown by flow cytometry. Depletion of circ-E2F3 in xenograft models of RB decreased tumor growth. A dual-luciferase reporter assay showed that circ-E2F3 could sponge miR-204-5p, which in turn targeted ROCK1 mRNA. Inhibition of miR-204-5p could promote the stimulatory effects of circ-E2F3 on RB progression. Taken together, circ-E2F3 could promote RB progression via the miR-204-5p/ROCK1 axis (Huang et al., 2021).
The L-type amino acid transporter (LAT1) can regulate cancer cell functions, such as apoptosis, proliferation, and drug-resistance by activation of the downstream AKT/mTOR pathway (Rosilio et al., 2015; Grzes et al., 2017). He et al. demonstrated that upregulation of miR-184 reduced the expression of solute carrier family 7 member 5 (SLC7A5, another term for LAT1) and reduced the proliferation, migration and invasion of RB cells (He et al., 2019). SLC7A5 expression levels tended to be higher in RB tissue samples (He et al., 2019).
Zheng et al., examined the effects and functional mechanism of the circRNA ER membrane protein complex subunit 9 (circ-FAM158A) in RB cells (Zheng et al., 2021). They first analyzed the expression levels of miR-138-5p, circ-FAM158A and SLC7A5 in RB samples by qRT-PCR. They found that circ-FAM158A and SLC7A5 were both upregulated, while miR-138-5p was downregulated in RB tissues. Knockdown of circ-FAM158A reduced the oncogenic properties of RB cells as shown by by CCK-8, colony formation, and transwell assays. Depletion of circ-FAM158A also stimulated apoptosis in RB cells as shown by by flow cytometry. A mouse xenograft model was used to confirm the role of circ-FAM158A in RB progression. By using StarBase and a dual-luciferase reporter assay, the molecular interplay between circ-FAM158A, miR-138-5p, and SLC7A5 was confirmed. They showed that circ-FAM158A could not only sponge miR-138-5p, but also upregulate its target SLC7A5. Functionally, miR-138-5p inhibition could reverse the anti-cancer effects of the silencing of circ-FAM158A on RB progression. Also, SLC7A5 overexpression blocked the anti-cancer effects of miR-138-5p in RB cells. Therefore, the anti-tumor effects of circ-FAM158A knockdown in RB cells occurred via the miR-138-5p/SLC7A5 axis (Zheng et al., 2021).
The SMAD family member 2 (SMAD2) gene is located on 18q21.1, and is a crucial signal transducer of the TGF-β pathway. Inhibition of SMAD2 resulted in the prevention of EMT progress, and reduced the proliferation and invasion stimulated by the TGF-β signaling pathway (Isselbacher et al., 2016; Tang et al., 2018). SMAD2 upregulation could promote the growth and metastasis of RB cells (Asnaghi et al., 2019). By contrast, SMAD2 inhibition reduced RB cell proliferation and invasion (Asnaghi et al., 2019).
To explore the function and mechanism of circ_0000527 in RB, Liang et al. (Liang et al., 2022b), measured the expression levels of circ_0000527, miR-1236-3p and SMAD2 using qRT-PCR. Their findings showed that upregulation of circ_0000527 in RB tissue samples was linked to clinicopathological features such as advanced TNM stage, and choroidal or optic nerve invasion. Also, circ_0000527 knockdown had anti-tumor effects in RB cells, with reduced proliferation, migration, invasion, and angiogenesis as shown by CCK-8, EdU, colony formation, scratch wound healing, transwell, and endothelial cell tube formation assays, respectively. In addition, flow cytometry provided evidence that silencing of circ_0000527 increased apoptosis in RB cells. Dual-luciferase reporter as well as RIP assays showed that circ_0000527 could sponge miR-1236-3p, which targeted SMAD2. Inhibition of miR-1236-3p reversed the inhibitory effects of circ_0000527 knockdown on RB malignant properties. Furthermore, overexpression of miR-1236-3p abrogated the effects of SMAD2 on RB progression. Moreover, in vivo experiments showed that circ_0000527 knockdown inhibited tumor formation. This study showed the role of the circ_0000527/miR-1236-3p/SMAD2 axis in RB progression (Table 3) (Liang et al., 2022b).
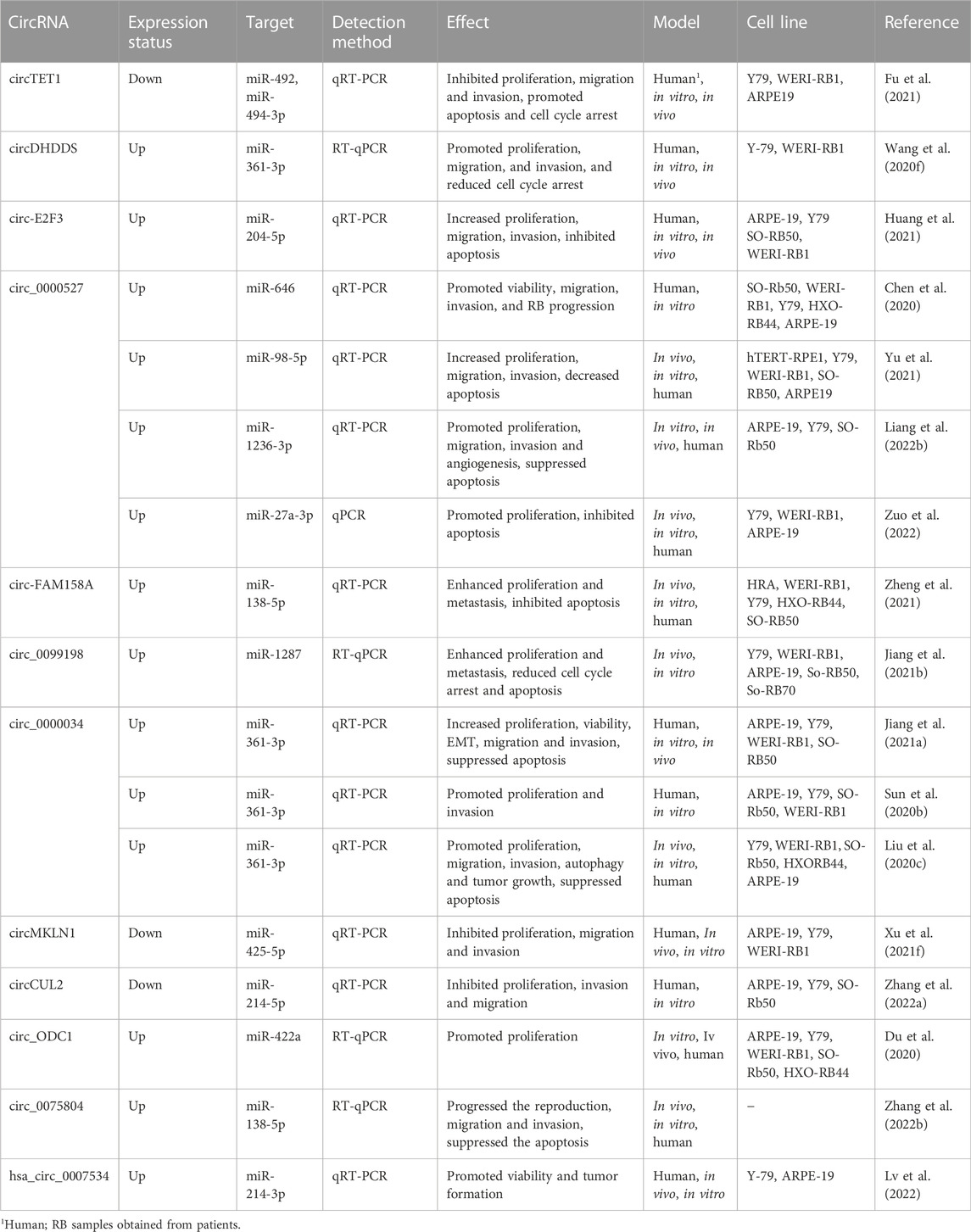
TABLE 3. The role of circRNAs in retinoblastoma.
Exosomes are nanosized vesicles 40–100 nm in diameter, which are released by many types of cells and can carry various biomolecules, such as lipids, proteins, mRNAs, and microRNAs. These extracellular vesicles participate in signaling pathways and in cellular communication between cells (Thébaud and Stewart, 2012; Rashed et al., 2017). Exosomes can fuse with the plasma membrane of recepient cells to release their genetic contents into the cytosol. Tumor cells release exosomes which are involved in various steps of angiogenesis (Feng et al., 2017b), immunosuppression (Chen et al., 2018), and tumor progression (Boyiadzis and Whiteside, 2017). In this content, WERI-RB1 cell-derived exosomes could infiltrate into the RB tumor microenvironment to promote tumor development (Chen et al., 2021c) by an unknown mechanism. However, the microRNAs contained in exosomes are known to exert a powerful effect upon tumor pathogenesis. For example, miR-210 contained inside hepatocellular carcinoma (HCC)-derived exosomes can stimulate angiogenesis by direct inhibition of SMAD4 and STAT6 (Lin et al., 2018). Furthermore, the miRNA-25-3p contained inside colorectal cancer (CRC)-derived exosomes can promote vascular leakage and increase the CRC metastatic phenotype (Zeng et al., 2018).
Krüppel-like factor 2 (KLF2) is an important member of the KLF family, a subclass of zinc-finger-containing transcription factors with DNA-binding domains, which can regulate tumor proliferation, metastasis, and affect the microenvironment (Tetreault et al., 2013). This molecule could inhibit the VEGFR2 promoter activity leading to reduced angiogenesis (Bhattacharya et al., 2005). KLF2 was downregulated by exosomal miR-25-3p to increase angiogenesis and raise vascular permeability in CRC (Zeng et al., 2018). Interestingly, KLF2 was found to be downregulated in breast (Zhang et al., 2015c), colorectal (Wang et al., 2017g), and lung cancer (Li et al., 2016c). Despite its downregulation in multiple cancer types with different stages and grades, KLF2 is highly expressed in normal epithelial cells (Wang et al., 2005).
Chen et al., identified the function and potential mechanism of exosomes released from WERI-RB1cells in RB tumor angiogenesis using biochemical approaches and animal experiments (Chen et al., 2021d). The in vitro findings showed that the exosomes were possibly taken up by human vascular endothelial cells (HUVECs). These exosomes led to increased survival and an inflammatory response in HUVECs by upregulation of genes, such as VCAM1, ICAM1, IL-1, IL-6, IL-8, and MCP-1. Additionally, cell migration and tube formation were significantly increased in HUVECs treated with RB cell-derived exosomes. Furthermore, in vivo data showed that tumors could absorb exosomes which were injected near the tumors. Compared to control tissues, the number of blood vessels and endothelial cells was increased in tumor tissues incubated with exosomes. The mechanism of stimulation of angiogenesis by the RB-derived exosomes was propsed to be miR-92a-3p contained within the exosomes. When HUVECs were treated with these exosomes they showed an increased expression of this miR. Treatment of HUVECs with a miR-92a-3p inhibitor significantly abrogated the effect of exosomes on tube formation and migration, as well as downregulation of the angiogenesis-related genes. The opposite results were obtained after treatment of HUVECs with a miR-92a-3p mimic. Bioinformatics analysis showed that KLF2 mRNA might be targeted by miR-92a-3p, which was confirmed both in vitro and in vivo. Therefore exosomal miR-92a-3p could be a promising therapeutic option for RB (Chen et al., 2021d).
Plousiou et al., examined the regulatory role of miRNAs in RB using a zebrafish model, (Plousiou et al., 2022). They first showed that co-culture of monocytes with RB cell lines significantly reduced proliferation, and the monocyte conditioned medium was added to RB cells to investigate the effect of the supernatant on RB progression. The found that miR-142-3p was upregulated in RB cells as well as in the medium used for their culture, compared with controls. Next they showed that monocytes secreted exosomes, which carried miR-142-3p into the co-culture medium and were then taken up by RB cells, leading to cell cycle arrest and inhibition of proliferation. They proposed that miR-142-3p could target the mRNA for TGFβR1 (transforming growth factor β receptor). Therefore, exosomal miR-142-3p could be a new miR-based treatment strategy to control RB tumor growth (Table 4) (Plousiou et al., 2022).

TABLE 4. The role of exosomal ncRNAs in retinoblastoma.
The importance of ncRNAs in affecting many normal physiological processes in humans, as well as pathological pathways is becoming increasingly appreciated. These include cell differentiation, proliferation, apoptosis, and migration. It has been widely accepted that deregulation of ncRNAs is linked to various diseases, particularly different cancers. RB is no exception, and several ncRNAs have now been identified as either tumor suppressor genes or tumor promoter genes. However it is known that several ncRNAs can have opposite functions across various cancer types, and even within the same tumor depending on stage, underlining the importance of characterizing the particular ncRNAs, and their mechanisms of action. Future research should focus on identifying the regulatory roles of ncRNAs and their related mechanisms in RB. Furthermore, the clinical applications of ncRNAs are yet to be evaluated. In addition to their value as prognostic or diagnostic biomarkers of disease, more studies are warranted to assess whether ncRNAs can be used in therapeutic regimens for RB, and whether exosomal ncRNAs may be more useful.
HM involved in conception, design, statistical analysis and drafting of the manuscript. NAD, SN, ZNGH, AP, SM, SAA. ZA, MM, SSTZ, MRH, AS contributed in data collection and manuscript drafting. All authors approved the final version for submission.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abe, H., Kamai, T., Hayashi, K., Anzai, N., Shirataki, H., Mizuno, T., et al. (2014). The Rho-kinase inhibitor HA-1077 suppresses proliferation/migration and induces apoptosis of urothelial cancer cells. BMC cancer 14 (1), 412–12. doi:10.1186/1471-2407-14-412
Aerts, I., Lumbroso-Le Rouic, L., Gauthier-Villars, M., Brisse, H., and Doz, F. (2015). Retinoblastoma update. Arch. Pediatr. 23 (1), 112–116. doi:10.1016/j.arcped.2015.09.025
Ardoin, S. P., and Pisetsky, D. S. (2008). The role of cell death in the pathogenesis of autoimmune disease: HMGB1 and microparticles as intercellular mediators of inflammation. Mod. Rheumatol. 18 (4), 319–326. doi:10.1007/s10165-008-0054-z
Asnaghi, L., White, D. T., Key, N., Choi, J., Mahale, A., Alkatan, H., et al. (2019). ACVR1C/SMAD2 signaling promotes invasion and growth in retinoblastoma. Oncogene 38 (12), 2056–2075. doi:10.1038/s41388-018-0543-2
Bahn, J. H., Zhang, Q., Li, F., Chan, T-M., Lin, X., Kim, Y., et al. (2015). The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin. Chem. 61 (1), 221–230. doi:10.1373/clinchem.2014.230433
Bai, S., Tian, B., Li, A., Yao, Q., Zhang, G., and Li, F. (2016). MicroRNA-125b promotes tumor growth and suppresses apoptosis by targeting DRAM2 in retinoblastoma. Eye Lond. Engl. 30 (12), 1630–1638. doi:10.1038/eye.2016.189
Bhattacharya, R., SenBanerjee, S., Lin, Z., Mir, S., Hamik, A., Wang, P., et al. (2005). Inhibition of vascular permeability factor/vascular endothelial growth factor-mediated angiogenesis by the Kruppel-like factor KLF2. J. Biol. Chem. 280 (32), 28848–28851. doi:10.1074/jbc.C500200200
Bianchi, M. E. (2009). HMGB1 loves company. J. Leukoc. Biol. 86 (3), 573–576. doi:10.1189/jlb.1008585
Bishop, J. O., and Madson, E. C. (1975). Retinoblastoma. Review of the current status. Surv. Ophthalmol. 19 (6), 342–366.
Boonyaratanakornkit, V., Melvin, V., Prendergast, P., Altmann, M., Ronfani, L., Bianchi, M. E., et al. (1998). High-mobility group chromatin proteins 1 and 2 functionally interact with steroid hormone receptors to enhance their DNA binding in vitro and transcriptional activity in mammalian cells. Mol. Cell. Biol. 18 (8), 4471–4487. doi:10.1128/mcb.18.8.4471
Boyiadzis, M., and Whiteside, T. (2017). The emerging roles of tumor-derived exosomes in hematological malignancies. Leukemia 31 (6), 1259–1268. doi:10.1038/leu.2017.91
Busch, M., Miroschnikov, N., Dankert, J. T., Wiesehöfer, M., Metz, K., Stephan, H., et al. (2021). Impact of RARα and miR-138 on retinoblastoma etoposide resistance. Tumour Biol. 43 (1), 11–26. doi:10.3233/TUB-200072
Cao, L., Peng, B., Yao, L., Zhang, X., Sun, K., Yang, X., et al. (2010). The ancient function of RB-E2F pathway: Insights from its evolutionary history. Biol. Direct 5 (1), 55–21. doi:10.1186/1745-6150-5-55
Carvalho, I. N., Reis, A. H., Dos Santos, A. C., and Vargas, F. R. (2017). A polymorphism in mir-34b/c as a potential biomarker for early onset of hereditary retinoblastoma. Cancer Biomark. 18 (3), 313–317. doi:10.3233/CBM-160248
Chai, Y., Xiao, J., Zhang, S., Du, Y., Luo, Z., Zhou, X., et al. (2018). High-mobility group protein B1 silencing promotes susceptibility of retinoblastoma cells to chemotherapeutic drugs through downregulating nuclear factor-κB. Int. J. Mol. Med. 41 (3), 1651–1658. doi:10.3892/ijmm.2018.3379
Chan, M. W., Huang, Y. W., Hartman-Frey, C., Kuo, C. T., Deatherage, D., Qin, H., et al. (2008). Aberrant transforming growth factor beta1 signaling and SMAD4 nuclear translocation confer epigenetic repression of ADAM19 in ovarian cancer. Neoplasia (New York, NY) 10 (9), 908–919. doi:10.1593/neo.08540
Che, X., Qian, Y., and Li, D. (2018). Suppression of disheveled-axin domain containing 1 (DIXDC1) by MicroRNA-186 inhibits the proliferation and invasion of retinoblastoma cells. J. Mol. Neurosci. 64 (2), 252–261. doi:10.1007/s12031-017-1017-7
Chen, G., Huang, A. C., Zhang, W., Zhang, G., Wu, M., Xu, W., et al. (2018). Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 560 (7718), 382–386. doi:10.1038/s41586-018-0392-8
Chen, K., Hou, Y., Liao, R., Li, Y., Yang, H., and Gong, J. (2021). LncRNA SNHG6 promotes G1/S-phase transition in hepatocellular carcinoma by impairing miR-204-5p-mediated inhibition of E2F1. Oncogene 40 (18), 3217–3230. doi:10.1038/s41388-021-01671-2
Chen, L-L. (2016). The biogenesis and emerging roles of circular RNAs. Nat. Rev. Mol. Cell Biol. 17 (4), 205–211. doi:10.1038/nrm.2015.32
Chen, N. N., Chao, D. L., and Li, X. G. (2020). Circular RNA has_circ_0000527 participates in proliferation, invasion and migration of retinoblastoma cells via miR-646/BCL-2 axis. Cell biochem. Funct. 38 (8), 1036–1046. doi:10.1002/cbf.3535
Chen, S., Chen, X., Luo, Q., Liu, X., Wang, X., Cui, Z., et al. (2021). Retinoblastoma cell-derived exosomes promote angiogenesis of human vesicle endothelial cells through microRNA-92a-3p. Cell Death Dis. 12 (7), 695. doi:10.1038/s41419-021-03986-0
Chen, S., Chen, X., Qiu, J., Chen, P., Han, X., Wu, Y., et al. (2021). Exosomes derived from retinoblastoma cells enhance tumour deterioration by infiltrating the microenvironment. Oncol. Rep. 45 (1), 278–290. doi:10.3892/or.2020.7858
Chen, Y., Lu, B., Liu, L., Pan, X., Jiang, C., and Xu, H. (2021). Long non-coding RNA PROX1-AS1 knockdown upregulates microRNA-519d-3p to promote chemosensitivity of retinoblastoma cells via targeting SOX2. Cell Cycle 20 (20), 2149–2159. doi:10.1080/15384101.2021.1971352
Cheng, Y., Chang, Q., Zheng, B., Xu, J., Li, H., and Wang, R. (2019). LncRNA XIST promotes the epithelial to mesenchymal transition of retinoblastoma via sponging miR-101. Eur. J. Pharmacol. 843, 210–216. doi:10.1016/j.ejphar.2018.11.028
Cheng, Y., Wang, X., Wang, P., Li, T., Hu, F., Liu, Q., et al. (2016). SUSD2 is frequently downregulated and functions as a tumor suppressor in RCC and lung cancer. Tumour Biol. 37 (7), 9919–9930. doi:10.1007/s13277-015-4734-y
Colucci-D’Amato, L., Speranza, L., and Volpicelli, F. (2020). Neurotrophic factor BDNF, physiological functions and therapeutic potential in depression, neurodegeneration and brain cancer. Int. J. Mol. Sci. 21 (20), 7777. doi:10.3390/ijms21207777
Correa-Acosta, A., González-Alviar, M. E., and Gaviria-Bravo, M. L. (2018). Retinoblastoma and optic nerve enhancement in a brain magnetic resonance scan: Is it always a metastasis? Arch. Soc. Esp. Oftalmol. 93 (5), 251–254. doi:10.1016/j.oftal.2017.10.010
Crews, F. T., Qin, L., Sheedy, D., Vetreno, R. P., and Zou, J. (2013). High mobility group box 1/Toll-like receptor danger signaling increases brain neuroimmune activation in alcohol dependence. Biol. Psychiatry 73 (7), 602–612. doi:10.1016/j.biopsych.2012.09.030
Cui, X., Liang, T., Ji, X., Shao, Y., Zhao, P., and Li, X. (004882022). LINC00488 induces tumorigenicity in retinoblastoma by regulating microRNA-30a-5p/EPHB2 Axis. Ocul. Immunol. Inflamm., 1–9. doi:10.1080/09273948.2022.2037659
Davidson, C. E., Reese, B. E., Billingsley, M. L., and Yun, J. K. (2004). Stannin, a protein that localizes to the mitochondria and sensitizes NIH-3T3 cells to trimethyltin and dimethyltin toxicity. Mol. Pharmacol. 66 (4), 855–863. doi:10.1124/mol.104.001719
Delsin, L. E. A., Salomao, K. B., Pezuk, J. A., and Brassesco, M. S. (2019). Expression profiles and prognostic value of miRNAs in retinoblastoma. J. Cancer Res. Clin. Oncol. 145 (1), 1–10. doi:10.1007/s00432-018-2773-7
Dimaras, H., and Corson, T. W. (2019). Retinoblastoma, the visible CNS tumor: A review. J. Neurosci. Res. 97 (1), 29–44. doi:10.1002/jnr.24213
Dimova, D. K., and Dyson, N. J. (2005). The E2F transcriptional network: Old acquaintances with new faces. Oncogene 24 (17), 2810–2826. doi:10.1038/sj.onc.1208612
Ding, F., Jiang, K., Sheng, Y., Li, C., and Zhu, H. (2020). LncRNA MIR7-3HG executes a positive role in retinoblastoma progression via modulating miR-27a-3p/PEG10 axis. Exp. Eye Res. 193, 107960. doi:10.1016/j.exer.2020.107960
Ding, Y., Wu, M., Liu, J., Wu, C., Huang, R., Zhu, R., et al. (2014). Seed-targeting anti-miR-21 inhibiting malignant progression of retinoblastoma and analysis of their phosphorylation signaling pathways. Exp. Eye Res. 122, 1–8. doi:10.1016/j.exer.2014.02.017
Dokmanovic, M., and Marks, P. A. (2005). Prospects: Histone deacetylase inhibitors. J. Cell. Biochem. 96 (2), 293–304. doi:10.1002/jcb.20532
Dong, Y., Wan, G., Yan, P., Qian, C., Li, F., and Peng, G. (2020). Long noncoding RNA LINC00324 promotes retinoblastoma progression by acting as a competing endogenous RNA for microRNA-769-5p, thereby increasing STAT3 expression. Aging 12 (9), 7729–7746. doi:10.18632/aging.103075
Drula, R., Braicu, C., Harangus, A., Nabavi, S. M., Trif, M., Slaby, O., et al. (2020). Critical function of circular RNAs in lung cancer. Wiley Interdiscip. Rev. RNA 11 (5), e1592. doi:10.1002/wrna.1592
Du, S., Wang, S., Zhang, F., and Lv, Y. (2020). SKP2, positively regulated by circ_ODC1/miR-422a axis, promotes the proliferation of retinoblastoma. J. Cell. Biochem. 121 (1), 322–331. doi:10.1002/jcb.29177
Fang, X., Wen, J., Sun, M., Yuan, Y., and Xu, Q. (2019). CircRNAs and its relationship with gastric cancer. J. Cancer 10 (24), 6105–6113. doi:10.7150/jca.32927
Feng, Q., Zhang, C., Lum, D., Druso, J. E., Blank, B., Wilson, K. F., et al. (2017). A class of extracellular vesicles from breast cancer cells activates VEGF receptors and tumour angiogenesis. Nat. Commun. 8 (1), 14450–14517. doi:10.1038/ncomms14450
Feng, Z., Xu, X., Cen, D., Luo, C., and Wu, S. (2017). miR-590-3p promotes colon cancer cell proliferation via Wnt/β-catenin signaling pathway by inhibiting WIF1 and DKK1. Eur. Rev. Med. Pharmacol. Sci. 21 (21), 4844–4852.
Fluge, Ø., Gravdal, K., Carlsen, E., Vonen, B., Kjellevold, K., Refsum, S., et al. (2009). Expression of EZH2 and Ki-67 in colorectal cancer and associations with treatment response and prognosis. Br. J. Cancer 101 (8), 1282–1289. doi:10.1038/sj.bjc.6605333
Fu, C., Wang, S., Jin, L., Zhang, M., and Li, M. (2021). CircTET1 inhibits retinoblastoma progression via targeting miR-492 and miR-494-3p through wnt/β-catenin signaling pathway. Curr. Eye Res. 46 (7), 978–987. doi:10.1080/02713683.2020.1843685
Fu, K., Zhang, K., and Zhang, X. (2022). LncRNA HOTAIR facilitates proliferation and represses apoptosis of retinoblastoma cells through the miR-20b-5p/RRM2/PI3K/AKT axis. Orphanet J. Rare Dis. 17 (1), 119. doi:10.1186/s13023-022-02206-y
Fuchs, B., Zhang, K., Schabel, A., Bolander, M. E., and Sarkar, G. (2001). Identification of twenty-two candidate markers for human osteogenic sarcoma. Gene 278 (1-2), 245–252. doi:10.1016/s0378-1119(01)00731-4
Gao, J., Dai, C., Yu, X., Yin, X. B., and Zhou, F. (2021). Long noncoding RNA LEF1-AS1 acts as a microRNA-10a-5p regulator to enhance MSI1 expression and promote chemoresistance in hepatocellular carcinoma cells through activating AKT signaling pathway. J. Cell. Biochem. 122 (1), 86–99. doi:10.1002/jcb.29833
Gao, M., Cui, Z., Zhao, D., Zhang, S., and Cai, Q. (2021). MicroRNA-9 inhibits proliferation and progression in retinoblastoma cells by targeting PTEN. Genes Genomics 43 (9), 1023–1033. doi:10.1007/s13258-021-01043-w
Gao, Y., Jing, M., Ge, R., Zhou, Z., and Sun, Y. (2014). Inhibition of hypoxia inducible factor 1α by siRNA-induced apoptosis in human retinoblastoma cells. J. Biochem. Mol. Toxicol. 28 (9), 394–399. doi:10.1002/jbt.21576
Gao, Y., Luo, X., and Zhang, J. (2021). LincRNA-ROR is activated by H3K27 acetylation and induces EMT in retinoblastoma by acting as a sponge of miR-32 to activate the Notch signaling pathway. Cancer Gene Ther. 28 (1-2), 42–54. doi:10.1038/s41417-020-0181-z
Gao, Y., Zhang, H., Zhao, S., He, D., and Gu, C. (2021). Nanofluorescence probes to detect miR-192/integrin alpha 1 and their correlations with retinoblastoma. J. Biomed. Nanotechnol. 17 (11), 2176–2185. doi:10.1166/jbn.2021.3185
Greco, O., and Scott, S. (2007). Tumor hypoxia and targeted gene therapy. Int. Rev. Cytol. 257, 181–212. doi:10.1016/S0074-7696(07)57005-1
Grolmusz, V. K., Karászi, K., Micsik, T., Tóth, E. A., Mészáros, K., Karvaly, G., et al. (2016). Cell cycle dependent RRM2 may serve as proliferation marker and pharmaceutical target in adrenocortical cancer. Am. J. Cancer Res. 6 (9), 2041–2053.
Grossi, F., Dal Bello, M. G., Salvi, S., Puzone, R., Pfeffer, U., Fontana, V., et al. (2015). Expression of ribonucleotide reductase subunit-2 and thymidylate synthase correlates with poor prognosis in patients with resected stages I–III non-small cell lung cancer. Dis. Markers 2015, 302649. doi:10.1155/2015/302649
Grzes, K. M., Swamy, M., Hukelmann, J. L., Emslie, E., Sinclair, L. V., and Cantrell, D. A. (2017). Control of amino acid transport coordinates metabolic reprogramming in T-cell malignancy. Leukemia 31 (12), 2771–2779. doi:10.1038/leu.2017.160
Guarnieri, D. J., and DiLeone, R. J. (2008). MicroRNAs: A new class of gene regulators. Ann. Med. 40 (3), 197–208. doi:10.1080/07853890701771823
Gui, F., Hong, Z., You, Z., Wu, H., and Zhang, Y. (2016). MiR-21 inhibitor suppressed the progression of retinoblastoma via the modulation of PTEN/PI3K/AKT pathway. Cell Biol. Int. 40 (12), 1294–1302. doi:10.1002/cbin.10678
Guo, H., Zhou, S., Tan, L., Wu, X., Wu, Z., and Ran, R. (2017). Clinicopathological significance of WIF1 hypermethylation in NSCLC, a meta-analysis and literature review. Oncotarget 8 (2), 2550–2557. doi:10.18632/oncotarget.13707
Guo, L., Bai, Y., Ji, S., and Ma, H. (2019). MicroRNA-98 suppresses cell growth and invasion of retinoblastoma via targeting the IGF1R/k-Ras/Raf/MEK/ERK signaling pathway. Int. J. Oncol. 54 (3), 807–820. doi:10.3892/ijo.2019.4689
Guo, L., Bai, Y., Ni, T., Li, Y., Cao, R., Ji, S., et al. (2021). MicroRNA‑98 suppresses cell growth and invasion of retinoblastoma via targeting the IGF1R/k‑Ras/Raf/MEK/ERK signaling pathway. Int. J. Oncol. 59 (1), 807–820. doi:10.3892/ijo.2019.4689
Guo, M., Zhang, X., Wang, G., Sun, J., Jiang, Z., Khadarian, K., et al. (2015). miR-603 promotes glioma cell growth via Wnt/β-catenin pathway by inhibiting WIF1 and CTNNBIP1. Cancer Lett. 360 (1), 76–86. doi:10.1016/j.canlet.2015.02.003
Guo, Q., Wang, H-B., Li, Y-H., Li, H-F., Li, T-T., Zhang, W-X., et al. (2016). Correlations of promoter methylation in WIF-1, RASSF1A, and CDH13 genes with the risk and prognosis of esophageal cancer. Med. Sci. Monit. 22, 2816–2824. doi:10.12659/msm.896877
Guo, W., Shao, F., Sun, S., Song, P., Guo, L., Xue, X., et al. (2020). Loss of SUSD2 expression correlates with poor prognosis in patients with surgically resected lung adenocarcinoma. J. Cancer 11 (7), 1648–1656. doi:10.7150/jca.39319
Hamasaki, M., Furuta, N., Matsuda, A., Nezu, A., Yamamoto, A., Fujita, N., et al. (2013). Autophagosomes form at ER–mitochondria contact sites. Nature 495 (7441), 389–393. doi:10.1038/nature11910
Han, S., Song, L., Chen, Y., Hou, M., Wei, X., and Fan, D. (2020). The long non-coding RNA ILF3-AS1 increases the proliferation and invasion of retinoblastoma through the miR-132-3p/SMAD2 axis. Exp. Cell Res. 393 (2), 112087. doi:10.1016/j.yexcr.2020.112087
Hao, B., Shi, A., Li, X., Li, J., Liu, Z., and Yuan, H. (2020). miR-4516 inhibits the apoptosis of RB tumor cells by targeting the PTEN/AKT signaling pathway. Exp. Eye Res. 200, 108224. doi:10.1016/j.exer.2020.108224
He, M. Y., An, Y., Gao, Y. J., Qian, X. W., Li, G., and Qian, J. (2014). Screening of RB1 gene mutations in Chinese patients with retinoblastoma and preliminary exploration of genotype-phenotype correlations. Mol. Vis. 20, 545–552.
He, S., Lu, Y., Liu, X., Huang, X., Keller, E. T., Qian, C-N., et al. (2015). Wnt3a: Functions and implications in cancer. Chin. J. Cancer 34 (3), 554–562. doi:10.1186/s40880-015-0052-4
He, T. G., Xiao, Z. Y., Xing, Y. Q., Yang, H. J., Qiu, H., and Chen, J. B. (2019). Tumor suppressor miR-184 enhances chemosensitivity by directly inhibiting SLC7A5 in retinoblastoma. Front. Oncol. 9, 1163. doi:10.3389/fonc.2019.01163
He, X., and Feng, Y. (2022). MicroRNA hsa-miR-657 promotes retinoblastoma malignancy by inhibiting peroxisome proliferator-activated receptor alpha expression. Anticancer. Drugs 33 (5), 478–488. doi:10.1097/CAD.0000000000001308
Hou, T., Ye, L., and Wu, S. (2021). Knockdown of LINC00504 inhibits the proliferation and invasion of breast cancer via the downregulation of miR-140-5p. Onco. Targets. Ther. 14, 3991–4003. doi:10.2147/OTT.S294965
Hsieh, J-C., Kodjabachian, L., Rebbert, M. L., Rattner, A., Smallwood, P. M., Samos, C. H., et al. (1999). A new secreted protein that binds to Wnt proteins and inhibits their activities. Nature 398 (6726), 431–436. doi:10.1038/18899
Hu, C. E., Du, P. Z., Zhang, H. D., and Huang, G. J. (2017). Long noncoding RNA CRNDE promotes proliferation of gastric cancer cells by targeting miR-145. Cell. Physiol. biochem. 42 (1), 13–21. doi:10.1159/000477107
Hu, S., Ran, Y., Chen, W., Zhang, Y., and Xu, Y. (2017). MicroRNA-326 inhibits cell proliferation and invasion, activating apoptosis in hepatocellular carcinoma by directly targeting LIM and SH3 protein 1. Oncol. Rep. 38 (3), 1569–1578. doi:10.3892/or.2017.5810
Hu, Y., Zhao, M., Li, L., Ding, J., Gui, Y. M., and Wei, T. W. (2021). miR-491-3p is downregulated in retinoblastoma and inhibit tumor cells growth and metastasis by targeting SNN. Biochem. Genet. 59 (2), 453–474. doi:10.1007/s10528-020-10007-w
Huang, G-Q., Ke, Z-P., Hu, H-B., and Gu, B. (2017). Co-expression network analysis of long noncoding RNAs (IncRNAs) and cancer genes reveals SFTA1P and CASC2 abnormalities in lung squamous cell carcinoma. Cancer Biol. Ther. 18 (2), 115–122. doi:10.1080/15384047.2017.1281494
Huang, J., Yang, Y., Fang, F., and Liu, K. (2018). MALAT1 modulates the autophagy of retinoblastoma cell through miR-124-mediated stx17 regulation. J. Cell. Biochem. 119 (5), 3853–3863. doi:10.1002/jcb.26464
Huang, S-M., Lin, C., Lin, H-Y., Chiu, C-M., Fang, C-W., Liao, K-F., et al. (2015). Brain-derived neurotrophic factor regulates cell motility in human colon cancer. Endocr. Relat. Cancer 22 (3), 455–464. doi:10.1530/ERC-15-0007
Huang, Y., Du, Q., Wu, W., She, F., and Chen, Y. (2016). Rescued expression of WIF-1 in gallbladder cancer inhibits tumor growth and induces tumor cell apoptosis with altered expression of proteins. Mol. Med. Rep. 14 (3), 2573–2581. doi:10.3892/mmr.2016.5532
Huang, Y., Xue, B., Pan, J., and Shen, N. (2021). Circ-E2F3 acts as a ceRNA for miR-204-5p to promote proliferation, metastasis and apoptosis inhibition in retinoblastoma by regulating ROCK1 expression. Exp. Mol. Pathol. 120, 104637. doi:10.1016/j.yexmp.2021.104637
Huang, Y. X., Nie, X. G., Li, G. D., Fan, D. S., Song, L. L., and Zhang, X. L. (2018). Downregulation of microRNA-182 inhibits cell viability, invasion and angiogenesis in retinoblastoma through inhibition of the PI3K/AKT pathway and CADM2 upregulation. Int. J. Oncol. 53 (6), 2615–2626. doi:10.3892/ijo.2018.4587
Ishimoto, T., Shiozaki, A., Ichikawa, D., Fujiwara, H., Konishi, H., Komatsu, S., et al. (2013). E2F5 as an independent prognostic factor in esophageal squamous cell carcinoma. Anticancer Res. 33 (12), 5415–5420.
Isselbacher, E. M., Lino Cardenas, C. L., and Lindsay, M. E. (2016). Hereditary influence in thoracic aortic aneurysm and dissection. Circulation 133 (24), 2516–2528. doi:10.1161/CIRCULATIONAHA.116.009762
Itakura, E., Kishi-Itakura, C., and Mizushima, N. (2012). The hairpin-type tail-anchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell 151 (6), 1256–1269. doi:10.1016/j.cell.2012.11.001
Ji, D., Chen, Z., Li, M., Zhan, T., Yao, Y., Zhang, Z., et al. (2014). MicroRNA-181a promotes tumor growth and liver metastasis in colorectal cancer by targeting the tumor suppressor WIF-1. Mol. Cancer 13 (1), 86–18. doi:10.1186/1476-4598-13-86
Jiang, Y., Li, Z., Zheng, S., Chen, H., Zhao, X., Gao, W., et al. (2016). The long non-coding RNA HOTAIR affects the radiosensitivity of pancreatic ductal adenocarcinoma by regulating the expression of Wnt inhibitory factor 1. Tumour Biol. 37 (3), 3957–3967. doi:10.1007/s13277-015-4234-0
Jiang, Y., Xiao, F., Wang, L., Wang, T., and Chen, L. (2021). Circular RNA has_circ_0000034 accelerates retinoblastoma advancement through the miR-361-3p/ADAM19 axis. Mol. Cell. Biochem. 476 (1), 69–80. doi:10.1007/s11010-020-03886-5
Jiang, Y., Xiao, F., Wang, L., Wang, T., and Chen, L. (2021). Hsa_circ_0099198 facilitates the progression of retinoblastoma by regulating miR-1287/LRP6 axis. Exp. Eye Res. 206, 108529. doi:10.1016/j.exer.2021.108529
Jiang, Y., Yim, S-H., Xu, H-D., Jung, S-H., Yang, S. Y., Hu, H-J., et al. (2011). A potential oncogenic role of the commonly observed E2F5 overexpression in hepatocellular carcinoma. World J. Gastroenterol. 17 (4), 470–477. doi:10.3748/wjg.v17.i4.470
Jin, Q., He, W., Chen, L., Yang, Y., Shi, K., and You, Z. (2018). MicroRNA-101-3p inhibits proliferation in retinoblastoma cells by targeting EZH2 and HDAC9. Exp. Ther. Med. 16 (3), 1663–1670. doi:10.3892/etm.2018.6405
Ke, N., Chen, L., Liu, Q., Xiong, H., Chen, X., and Zhou, X. (2022). Downregulation of miR-211-5p promotes carboplatin resistance in human retinoblastoma Y79 cells by affecting the GDNF-LIF interaction. Front. Oncol. 12, 848733. doi:10.3389/fonc.2022.848733
Kent, L. N., and Leone, G. (2019). The broken cycle: E2F dysfunction in cancer. Nat. Rev. Cancer 19 (6), 326–338. doi:10.1038/s41568-019-0143-7
Khan, M., Walters, L. L., Li, Q., Thomas, D. G., Miller, J. M., Zhang, Q., et al. (2015). Characterization and pharmacologic targeting of EZH2, a fetal retinal protein and epigenetic regulator, in human retinoblastoma. Lab. Invest. 95 (11), 1278–1290. doi:10.1038/labinvest.2015.104
Kimura, T. (2020). Non-coding natural antisense RNA: Mechanisms of action in the regulation of target gene expression and its clinical implications. Yakugaku Zasshi 140 (5), 687–700. doi:10.1248/yakushi.20-00002
Klaus, A., Fathi, O., Tatjana, T-W., Bruno, N., and Oskar, K. (2018). Expression of hypoxia-associated protein HIF-1α in follicular thyroid cancer is associated with distant metastasis. Pathol. Oncol. Res. 24 (2), 289–296. doi:10.1007/s12253-017-0232-4
Kleer, C. G., Cao, Q., Varambally, S., Shen, R., Ota, I., Tomlins, S. A., et al. (2003). EZH2 is a marker of aggressive breast cancer and promotes neoplastic transformation of breast epithelial cells. Proc. Natl. Acad. Sci. U. S. A. 100 (20), 11606–11611. doi:10.1073/pnas.1933744100
Kong, L., Sun, Y., Chen, M., Dai, Y., and Liu, Z. (2020). Downregulation of microRNA-320a inhibits proliferation and induces apoptosis of retinoblastoma cells via targeting TUSC3. Exp. Ther. Med. 20 (5), 9. doi:10.3892/etm.2020.9137
Kothandaraman, N., Bajic, V. B., Brendan, P. N., Huak, C. Y., Keow, P. B., Razvi, K., et al. (2010). E2F5 status significantly improves malignancy diagnosis of epithelial ovarian cancer. BMC cancer 10 (1), 64–13. doi:10.1186/1471-2407-10-64
Kuzmichev, A., Jenuwein, T., Tempst, P., and Reinberg, D. (2004). Different EZH2-containing complexes target methylation of histone H1 or nucleosomal histone H3. Mol. Cell 14 (2), 183–193. doi:10.1016/s1097-2765(04)00185-6
Lassmann, S., Weis, R., Makowiec, F., Roth, J., Danciu, M., Hopt, U., et al. (2007). Array CGH identifies distinct DNA copy number profiles of oncogenes and tumor suppressor genes in chromosomal-and microsatellite-unstable sporadic colorectal carcinomas. J. Mol. Med. 85 (3), 293–304. doi:10.1007/s00109-006-0126-5
Lee, S. M., Park, J. Y., and Kim, D. S. (2013). Wif1 hypermethylation as unfavorable prognosis of non-small cell lung cancers with EGFR mutation. Mol. Cells 36 (1), 69–73. doi:10.1007/s10059-013-0060-7
Lei, Q., Shen, F., Wu, J., Zhang, W., Wang, J., and Zhang, L. (2014). MiR-101, downregulated in retinoblastoma, functions as a tumor suppressor in human retinoblastoma cells by targeting EZH2. Oncol. Rep. 32 (1), 261–269. doi:10.3892/or.2014.3167
Leonel, C., Carvalho Ferreira, L., Ferraz Borin, T., Gobbe Moschetta, M., Scavacini Freitas, G., Raineri Haddad, M., et al. (2017). Inhibition of epithelial-mesenchymal transition in response to treatment with metformin and Y27632 in breast cancer cell lines. Anticancer. Agents Med. Chem. 17 (8), 1113–1125. doi:10.2174/1871520617666170102153954
Li, C., Wang, Z., Chen, S., Zhang, J., Qu, K., and Liu, C. (2018). MicroRNA-552 promotes hepatocellular carcinoma progression by downregulating WIF1. Int. J. Mol. Med. 42 (6), 3309–3317. doi:10.3892/ijmm.2018.3882
Li, C., Zhao, J., and Sun, W. (2020). microRNA-222-Mediated VHL downregulation facilitates retinoblastoma chemoresistance by increasing HIF1α expression. Invest. Ophthalmol. Vis. Sci. 61 (10), 9. doi:10.1167/iovs.61.10.9
Li, H., Fan, D., Wang, W., Zhang, X., Song, L., and Huang, Y. (2021). MiR-142-5p serves as a tumor suppressor in retinoblastoma cells by regulating MYCN. Biochem. Biophys. Res. Commun. 574, 20–26. doi:10.1016/j.bbrc.2021.07.099
Li, J., Liu, X., Wang, W., and Li, C. (2020). miR-133a-3p promotes apoptosis and induces cell cycle arrest by targeting CREB1 in retinoblastoma. Arch. Med. Sci. 16 (4), 941–956. doi:10.5114/aoms.2019.86901
Li, J., Ma, M., Yang, X., Zhang, M., Luo, J., Zhou, H., et al. (2020). Circular HER2 RNA positive triple negative breast cancer is sensitive to Pertuzumab. Mol. Cancer 19 (1), 142–218. doi:10.1186/s12943-020-01259-6
Li, J., Yang, J., Zhou, P., Le, Y., Zhou, C., Wang, S., et al. (2015). Circular RNAs in cancer: Novel insights into origins, properties, functions and implications. Am. J. Cancer Res. 5 (2), 472–480.
Li, J., Zhang, Y., Wang, X., and Zhao, R. (2017). microRNA-497 overexpression decreases proliferation, migration and invasion of human retinoblastoma cells via targeting vascular endothelial growth factor A. Oncol. Lett. 13 (6), 5021–5027. doi:10.3892/ol.2017.6083
Li, K., Han, F., Wu, Y., and Wang, X. (2021). miR-340 promotes retinoblastoma cell proliferation, migration and invasion through targeting WIF1. Onco. Targets. Ther. 14, 3635–3648. doi:10.2147/OTT.S302800
Li, L., Yu, H., and Ren, Q. (2020). MiR-218-5p suppresses the progression of retinoblastoma through targeting NACC1 and inhibiting the AKT/mTOR signaling pathway. Cancer Manag. Res. 12, 6959–6967. doi:10.2147/CMAR.S246142
Li, M., Chen, X. M., Wang, D. M., Gan, L., and Qiao, Y. (2016). Effects of miR-26a on the expression of Beclin 1 in retinoblastoma cells. Genet. Mol. Res. 15 (2). doi:10.4238/gmr.15028193
Li, W., Sun, M., Zang, C., Ma, P., He, J., Zhang, M., et al. (2016). Upregulated long non-coding RNA AGAP2-AS1 represses LATS2 and KLF2 expression through interacting with EZH2 and LSD1 in non-small-cell lung cancer cells. Cell Death Dis. 7 (5), e2225–e. doi:10.1038/cddis.2016.126
Li, W., Wang, J., Zhang, D., Zhang, X., Xu, J., and Zhao, L. (2019). MicroRNA-98 targets HMGA2 to inhibit the development of retinoblastoma through mediating Wnt/β-catenin pathway. Cancer Biomark. 25 (1), 79–88. doi:10.3233/CBM-182315
Li, X., Yang, L., Shuai, T., Piao, T., and Wang, R. (2016). MiR-433 inhibits retinoblastoma malignancy by suppressing Notch1 and PAX6 expression. Biomed. Pharmacother. = Biomedecine Pharmacother. 82, 247–255. doi:10.1016/j.biopha.2016.05.003
Li, Y., and Zheng, Y. (2017). Hypoxia promotes invasion of retinoblastoma cells in vitro by upregulating HIF-1α/MMP9 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 21 (23), 5361–5369. doi:10.26355/eurrev_201712_13921
Li, Z. N., Ge, M. X., Cao, L. J., and Yuan, Z. F. (2020). lncRNA RHPN1-AS1 serves as a sponge for miR-3133 modulating the cell proliferation of retinoblastoma through JAK2. Biomed. Res. Int. 2020, 3502981. doi:10.1155/2020/3502981
Liang, T., Fan, M., Meng, Z., Sun, B., Mi, S., and Gao, X. (2022). Circ_0000527 drives retinoblastoma progression by regulating miR-1236-3p/SMAD2 pathway. Curr. Eye Res. 47 (4), 624–633. doi:10.1080/02713683.2021.2007535
Liang, Y., Chen, X., and Liang, Z. (2017). MicroRNA-320 regulates autophagy in retinoblastoma by targeting hypoxia inducible factor-1α. Exp. Ther. Med. 14 (3), 2367–2372. doi:10.3892/etm.2017.4779
Liang, Y., Wang, H., Song, R., and Yin, X. (2022). lncRNA FOXD2-AS1 promotes the retinoblastoma cell viability and migration by sponging miR-31. Biomed. Res. Int. 2022, 7723425. doi:10.1155/2022/7723425
Liang, Y., Zheng, T., Song, R., Wang, J., Yin, D., Wang, L., et al. (2013). Hypoxia-mediated sorafenib resistance can be overcome by EF24 through Von Hippel-Lindau tumor suppressor-dependent HIF-1α inhibition in hepatocellular carcinoma. Hepatology 57 (5), 1847–1857. doi:10.1002/hep.26224
Lin, B., Hong, H., Jiang, X., Li, C., Zhu, S., Tang, N., et al. (2017). WNT inhibitory factor 1 promoter hypermethylation is an early event during gallbladder cancer tumorigenesis that predicts poor survival. Gene 622, 42–49. doi:10.1016/j.gene.2017.04.034
Lin, X-J., Fang, J-H., Yang, X-J., Zhang, C., Yuan, Y., Zheng, L., et al. (2018). Hepatocellular carcinoma cell-secreted exosomal microRNA-210 promotes angiogenesis in vitro and in vivo. Mol. Ther. Nucleic Acids 11, 243–252. doi:10.1016/j.omtn.2018.02.014
Liu, B., Lu, B., Wang, X., Jiang, H., and Kuang, W. (2019). MiR-361-5p inhibits cell proliferation and induces cell apoptosis in retinoblastoma by negatively regulating CLDN8. Childs Nerv. Syst. 35 (8), 1303–1311. doi:10.1007/s00381-019-04199-9
Liu, F., Zhang, Q., and Liang, Y. (2020). MicroRNA-598 acts as an inhibitor in retinoblastoma through targeting E2F1 and regulating AKT pathway. J. Cell. Biochem. 121 (3), 2294–2302. doi:10.1002/jcb.29453
Liu, H., Cao, B., Zhao, Y., Liang, H., and Liu, X. (2018). Upregulated miR-221/222 promotes cell proliferation and invasion and is associated with invasive features in retinoblastoma. Cancer Biomark. 22 (4), 621–629. doi:10.3233/CBM-170721
Liu, H., Yuan, H. F., Xu, D., Chen, K. J., Tan, N., and Zheng, Q. J. (2020). Circular RNA circ_0000034 upregulates STX17 level to promote human retinoblastoma development via inhibiting miR-361-3p. Eur. Rev. Med. Pharmacol. Sci. 24 (23), 12080–12092. doi:10.26355/eurrev_202012_23997
Liu, K., Huang, J., Xie, M., Yu, Y., Zhu, S., Kang, R., et al. (2014). MIR34A regulates autophagy and apoptosis by targeting HMGB1 in the retinoblastoma cell. Autophagy 10 (3), 442–452. doi:10.4161/auto.27418
Liu, L., Xiao, C., and Sun, Q. (2022). MiRNA-375 inhibits retinoblastoma progression through targeting ERBB2 and inhibiting MAPK1/MAPK3 signalling pathway. Cutan. Ocul. Toxicol. 41 (1), 1–10. doi:10.1080/15569527.2021.1994587
Liu, M., Wang, S. M., Jiang, Z. X., Lauren, H., and Tao, L. M. (2018). Effects of miR-22 on viability, migration, invasion and apoptosis in retinoblastoma Y79 cells by targeting high-mobility group box 1. Int. J. Ophthalmol. 11 (10), 1600–1607. doi:10.18240/ijo.2018.10.05
Liu, P., Shen, J. K., Hornicek, F. J., Liu, F., and Duan, Z. (2017). Wnt inhibitory factor 1 (WIF1) methylation and its association with clinical prognosis in patients with chondrosarcoma. Sci. Rep. 7 (1), 1580–1611. doi:10.1038/s41598-017-01763-8
Liu, S., and Wen, C. (2022). miR-141-3p promotes retinoblastoma progression via inhibiting sushi domain-containing protein 2. Bioengineered 13 (3), 7410–7424. doi:10.1080/21655979.2022.2048770
Liu, S., Yan, G., Zhang, J., and Yu, L. (2018). Knockdown of long noncoding RNA (lncRNA) metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) inhibits proliferation, migration, and invasion and promotes apoptosis by targeting miR-124 in retinoblastoma. Oncol. Res. 26 (4), 581–591. doi:10.3727/096504017X14953948675403
Liu, S., Zhang, X., Hu, C., Wang, Y., and Xu, C. (2018). miR-29a inhibits human retinoblastoma progression by targeting STAT3. Oncol. Rep. 39 (2), 739–746. doi:10.3892/or.2017.6144
Liu, X., Li, X., and Li, J. (2021). Long non-coding RNA FEZF1-AS1 promotes growth and reduces apoptosis through regulation of miR-363-3p/PAX6 Axis in retinoblastoma. Biochem. Genet. 59 (3), 637–651. doi:10.1007/s10528-020-10026-7
Liu, X. M., Li, X. F., and Li, J. C. (2021). MiR-146a functions as a potential tumor suppressor in retinoblastoma by negatively regulate neuro-oncological ventral antigen-1. Kaohsiung J. Med. Sci. 37 (4), 286–293. doi:10.1002/kjm2.12337
Liu, Y., Li, H., Liu, Y., and Zhu, Z. (2018). MiR-22-3p targeting alpha-enolase 1 regulates the proliferation of retinoblastoma cells. Biomed. Pharmacother. = Biomedecine Pharmacother. 105, 805–812. doi:10.1016/j.biopha.2018.06.038
Liu, Y., Liang, G., Wang, H., and Liu, Z. (2019). MicroRNA-129-5p suppresses proliferation, migration and invasion of retinoblastoma cells through PI3K/AKT signaling pathway by targeting PAX6. Pathol. Res. Pract. 215 (12), 152641. doi:10.1016/j.prp.2019.152641
Liu, Z., Huang, Y., Zhang, F., Tang, H., and Wang, Y. (2020). Autophagy-related gene 7 deficiency caused by miR-154-5p overexpression suppresses the cell viability and tumorigenesis of retinoblastoma by increasing cell apoptosis. Ann. Transl. Med. 8 (21), 1451. doi:10.21037/atm-20-6009
Lu, X., Tu, H., Tang, D., Huang, X., and Sun, F. (2022). miR-130a-3p enhances the chemosensitivity of Y79 retinoblastoma cells to vincristine by targeting PAX6 expression. Curr. Eye Res. 47 (3), 418–425. doi:10.1080/02713683.2021.1984537
Luo, Z., Rong, Z., Zhang, J., Zhu, Z., Yu, Z., Li, T., et al. (2020). Circular RNA circCCDC9 acts as a miR-6792-3p sponge to suppress the progression of gastric cancer through regulating CAV1 expression. Mol. Cancer 19 (1), 86–21. doi:10.1186/s12943-020-01203-8
Lv, X., Yang, H., Zhong, H., He, L., and Wang, L. (2022). Osthole exhibits an antitumor effect in retinoblastoma through inhibiting the PI3K/AKT/mTOR pathway via regulating the hsa_circ_0007534/miR-214-3p axis. Pharm. Biol. 60 (1), 417–426. doi:10.1080/13880209.2022.2032206
Lytle, J. R., Yario, T. A., and Steitz, J. A. (2007). Target mRNAs are repressed as efficiently by microRNA-binding sites in the 5′ UTR as in the 3′ UTR. Proc. Natl. Acad. Sci. U. S. A. 104 (23), 9667–9672. doi:10.1073/pnas.0703820104
Lyv, X., Wu, F., Zhang, H., Lu, J., Wang, L., and Ma, Y. (2020). Long noncoding RNA ZFPM2-AS1 knockdown restrains the development of retinoblastoma by modulating the MicroRNA-515/HOXA1/wnt/β-catenin Axis. Invest. Ophthalmol. Vis. Sci. 61 (6), 41. doi:10.1167/iovs.61.6.41
Ma, X., Jin, L., Lei, X., Tong, J., and Wang, R. (2020). MicroRNA-363-3p inhibits cell proliferation and induces apoptosis in retinoblastoma cells via the Akt/mTOR signaling pathway by targeting PIK3CA. Oncol. Rep. 43 (5), 1365–1374. doi:10.3892/or.2020.7544
Machihara, K., and Namba, T. (2019). BAP31 inhibits cell adaptation to ER stress conditions, negatively regulating autophagy induction by interaction with STX17. Cells 8 (11), 1350. doi:10.3390/cells8111350
Madhavan, J., Mitra, M., Mallikarjuna, K., Pranav, O., Srinivasan, R., Nagpal, A., et al. (2009). KIF14 and E2F3 mRNA expression in human retinoblastoma and its phenotype association. Mol. Vis. 15, 235–240.
Maheshwari, A., and Finger, P. T. (2018). Cancers of the eye. Cancer Metastasis Rev. 37 (4), 677–690. doi:10.1007/s10555-018-9762-9
Marks, P. A., Richon, V. M., Kelly, W. K., Chiao, J. H., and Miller, T. (Editors) (2004). “Histone deacetylase inhibitors: Development as cancer therapy,” Reversible protein acetylation: Novartis foundation symposium 259 (Wiley Online Library).
Martin, A., Jones, A., Bryar, P. J., Mets, M., Weinstein, J., Zhang, G., et al. (2013). MicroRNAs-449a and -449b exhibit tumor suppressive effects in retinoblastoma. Biochem. Biophys. Res. Commun. 440 (4), 599–603. doi:10.1016/j.bbrc.2013.09.117
McAllister, A. (2001). Neurotrophins and neuronal differentiation in the central nervous system. Cell. Mol. Life Sci. 58 (8), 1054–1060. doi:10.1007/PL00000920
Memczak, S., Jens, M., Elefsinioti, A., Torti, F., Krueger, J., Rybak, A., et al. (2013). Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 495 (7441), 333–338. doi:10.1038/nature11928
Meng, X., Zhang, Y., Hu, Y., Zhong, J., Jiang, C., and Zhang, H. (2021). LncRNA CCAT1 sponges miR-218-5p to promote EMT, cellular migration and invasion of retinoblastoma by targeting MTF2. Cell. Signal. 86, 110088. doi:10.1016/j.cellsig.2021.110088
Miao, X., Wang, Z., Chen, B., Chen, Y., Wang, X., Jiang, L., et al. (2018). miR-140-5p suppresses retinoblastoma cell proliferation, migration, and invasion by targeting CEMIP and CADM3. Cell. Mol. Biol. 64 (6), 42–47. doi:10.14715/cmb/2018.64.6.8
Mohammad, F., Weissmann, S., Leblanc, B., Pandey, D. P., Højfeldt, J. W., Comet, I., et al. (2017). EZH2 is a potential therapeutic target for H3K27M-mutant pediatric gliomas. Nat. Med. 23 (4), 483–492. doi:10.1038/nm.4293
Montoya, V., Fan, H., Bryar, P. J., Weinstein, J. L., Mets, M. B., Feng, G., et al. (2015). Novel miRNA-31 and miRNA-200a-mediated regulation of retinoblastoma proliferation. PloS one 10 (9), e0138366. doi:10.1371/journal.pone.0138366
Ni, W., Li, Z., and Ai, K. (2022). lncRNA ZFPM2-AS1 promotes retinoblastoma progression by targeting microRNA miR-511-3p/paired box protein 6 (PAX6) axis. Bioengineered 13 (1), 1637–1649. doi:10.1080/21655979.2021.2021346
Nie, C., Ma, H., Gao, Y., Li, J., Tang, Z., Chen, Y., et al. (2021). RNA sequencing and bioinformatic analysis on retinoblastoma revealing that cell cycle deregulation is a key process in retinoblastoma tumorigenesis. Ophthalmologica. 244 (1), 51–59. doi:10.1159/000506993
Nordlund, P., and Reichard, P. (2006). Ribonucleotide reductases. Annu. Rev. Biochem. 75, 681–706. doi:10.1146/annurev.biochem.75.103004.142443
Ouyang, M., Liu, G., Xiong, C., and Rao, J. (2022). microRNA-181a-5p impedes the proliferation, migration, and invasion of retinoblastoma cells by targeting the NRAS proto-oncogene. Clinics 77, 100026. doi:10.1016/j.clinsp.2022.100026
Park, S. J., Woo, S. J., and Park, K. H. (2014). Incidence of retinoblastoma and survival rate of retinoblastoma patients in Korea using the Korean National Cancer Registry database (1993-2010). Invest. Ophthalmol. Vis. Sci. 55 (5), 2816–2821. doi:10.1167/iovs.14-14078
Peng, X., Yan, J., and Cheng, F. (2020). LncRNA TMPO-AS1 up-regulates the expression of HIF-1α and promotes the malignant phenotypes of retinoblastoma cells via sponging miR-199a-5p. Pathol. Res. Pract. 216 (4), 152853. doi:10.1016/j.prp.2020.152853
Plousiou, M., De Vita, A., Miserocchi, G., Bandini, E., Vannini, I., Melloni, M., et al. (2022). Growth inhibition of retinoblastoma cell line by exosome-mediated transfer of miR-142-3p. Cancer Manag. Res. 14, 2119–2131. doi:10.2147/CMAR.S351979
Poggi, L., Casarosa, S., and Carl, M. (2018). An eye on the Wnt inhibitory factor Wif1. Front. Cell Dev. Biol. 6, 167. doi:10.3389/fcell.2018.00167
Pueyo, J. I., Magny, E. G., Sampson, C. J., Amin, U., Evans, I. R., Bishop, S. A., et al. (2016). Hemotin, a regulator of phagocytosis encoded by a small ORF and conserved across metazoans. PLoS Biol. 14 (3), e1002395. doi:10.1371/journal.pbio.1002395
Qi, B., Newcomer, R. G., and Sang, Q. X. (2009). ADAM19/adamalysin 19 structure, function, and role as a putative target in tumors and inflammatory diseases. Curr. Pharm. Des. 15 (20), 2336–2348. doi:10.2174/138161209788682352
Ramachandran, I., Ganapathy, V., Gillies, E., Fonseca, I., Sureban, S., Houchen, C., et al. (2014). Wnt inhibitory factor 1 suppresses cancer stemness and induces cellular senescence. Cell Death Dis. 5 (5), e1246–e. doi:10.1038/cddis.2014.219
Ramachandran, I., Thavathiru, E., Ramalingam, S., Natarajan, G., Mills, W., Benbrook, D., et al. (2012). Wnt inhibitory factor 1 induces apoptosis and inhibits cervical cancer growth, invasion and angiogenesis in vivo. Oncogene 31 (22), 2725–2737. doi:10.1038/onc.2011.455
Rao, R., and Honavar, S. G. (2017). Indian J. Pediatr. 84 (12), 937–944. doi:10.1007/s12098-017-2395-0
Rashed, M. H., Bayraktar, E., Helal, G. K., Abd-Ellah, , Amero, P., Chavez-Reyes, A., et al. (2017). Exosomes: From garbage bins to promising therapeutic targets. Int. J. Mol. Sci. 18 (3), 538. doi:10.3390/ijms18030538
Reese, B. E., Davidson, C., Billingsley, M. L., and Yun, J. (2005). Protein kinase cϵ regulates tumor necrosis factor-α-induced stannin gene expression. J. Pharmacol. Exp. Ther. 314 (1), 61–69. doi:10.1124/jpet.105.084236
Reese, B. E., Krissinger, D., Yun, J. K., and Billingsley, M. L. (2006). Elucidation of stannin function using microarray analysis: Implications for cell cycle control. Gene Expr. 13 (1), 41–52. doi:10.3727/000000006783991944
Rong, J., Li, Z., Xu, L., Lang, L., and Zheng, G. (2021). microRNA-362-3p targets USP22 to retard retinoblastoma growth via reducing deubiquitination of LSD1. Cell Cycle 20 (3), 298–307. doi:10.1080/15384101.2021.1874685
Roperch, J-P., Incitti, R., Forbin, S., Bard, F., Mansour, H., Mesli, F., et al. (2013). Aberrant methylation of NPY, PENK, and WIF1 as a promising marker for blood-based diagnosis of colorectal cancer. BMC cancer 13 (1), 566–610. doi:10.1186/1471-2407-13-566
Rosilio, C., Nebout, M., Imbert, V., Griessinger, E., Neffati, Z., Benadiba, J., et al. (2015). L-Type amino-acid transporter 1 (LAT1): A therapeutic target supporting growth and survival of T-cell lymphoblastic lymphoma/T-cell acute lymphoblastic leukemia. Leukemia 29 (6), 1253–1266. doi:10.1038/leu.2014.338
Sanger, H. L., Klotz, G., Riesner, D., Gross, H. J., and Kleinschmidt, A. K. (1976). Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. U. S. A. 73 (11), 3852–3856. doi:10.1073/pnas.73.11.3852
Sha, Y., Zmijewski, J., Xu, Z., and Abraham, E. (2008). HMGB1 develops enhanced proinflammatory activity by binding to cytokines. J. Immunol. 180 (4), 2531–2537. doi:10.4049/jimmunol.180.4.2531
Shao, F., Huang, M., Meng, F., and Huang, Q. (2018). Circular RNA signature predicts gemcitabine resistance of pancreatic ductal adenocarcinoma. Front. Pharmacol. 9, 584. doi:10.3389/fphar.2018.00584
Sharma, G. G., Okada, Y., Von Hoff, D., and Goel, A. (Editors) (2021). “Non-coding RNA biomarkers in pancreatic ductal adenocarcinoma,” Seminars in cancer biology (Elsevier).
Sheets, J. N., Patrick, M. E., and Egland, K. A. (2020). SUSD2 expression correlates with decreased metastasis and increased survival in a high-grade serous ovarian cancer xenograft murine model. Oncotarget 11 (24), 2290–2301. doi:10.18632/oncotarget.27626
Shimizu, F., Shiiba, M., Ogawara, K., Kimura, R., Minakawa, Y., Baba, T., et al. (2013). Overexpression of LIM and SH3 Protein 1 leading to accelerated G2/M phase transition contributes to enhanced tumourigenesis in oral cancer. PloS one 8 (12), e83187. doi:10.1371/journal.pone.0083187
Silva, A. M., Moura, S. R., Teixeira, J. H., Barbosa, M. A., Santos, S. G., and Almeida, M. I. (2019). Long noncoding RNAs: A missing link in osteoporosis. Bone Res. 7 (1), 10–16. doi:10.1038/s41413-019-0048-9
Singh, A., Patel, P., Patel, K. V., Jain, K. D., Veerasamy, R., Sharma, C. P., et al. (2017). Histone deacetylase inhibitors for the treatment of colorectal cancer: Recent progress and future prospects. Curr. Cancer Drug Targets 17 (5), 456–466. doi:10.2174/1568009617666170109150134
Singh, M. K., Singh, L., Pushker, N., Sen, S., Sharma, A., Chauhan, F. A., et al. (2015). Correlation of high mobility group box-1 protein (HMGB1) with clinicopathological parameters in primary retinoblastoma. Pathol. Oncol. Res. 21 (4), 1237–1242. doi:10.1007/s12253-015-9951-6
Song, D., Diao, J., Yang, Y., and Chen, Y. (2017). MicroRNA-382 inhibits cell proliferation and invasion of retinoblastoma by targeting BDNF-mediated PI3K/AKT signalling pathway. Mol. Med. Rep. 16 (5), 6428–6436. doi:10.3892/mmr.2017.7396
Song, J., and Zhang, Z. (2021). Long non-coding RNA SNHG20 promotes cell proliferation, migration and invasion in retinoblastoma via the miR-335-5p/E2F3 axis. Mol. Med. Rep. 24 (2), 543. doi:10.3892/mmr.2021.12182
Song, L., Huang, Y., Zhang, X., Han, S., Hou, M., and Li, H. (2020). Downregulation of microRNA-224-3p hampers retinoblastoma progression via activation of the hippo-YAP signaling pathway by increasing LATS2. Invest. Ophthalmol. Vis. Sci. 61 (3), 32. doi:10.1167/iovs.61.3.32
Sun, Q. X., Wang, R. R., Liu, N., and Liu, C. (2020). Dysregulation of miR-204-3p driven by the viability and motility of retinoblastoma via wnt/β-catenin pathway in vitro and in vivo. Pathol. Oncol. Res. 26 (3), 1549–1558. doi:10.1007/s12253-019-00722-0
Sun, S., Wang, R., Yi, S., Li, S., Wang, L., and Wang, J. (2021). Roles of the microRNA-338-3p/NOVA1 axis in retinoblastoma. Mol. Med. Rep. 23 (5), 394. doi:10.3892/mmr.2021.12033
Sun, Z., Zhang, A., Hou, M., and Jiang, T. (2020). Circular RNA hsa_circ_0000034 promotes the progression of retinoblastoma via sponging microRNA-361-3p. Bioengineered 11 (1), 949–957. doi:10.1080/21655979.2020.1814670
Sun, Z., Zhang, A., Jiang, T., Du, Z., Che, C., and Wang, F. (2015). MiR-145 suppressed human retinoblastoma cell proliferation and invasion by targeting ADAM19. Int. J. Clin. Exp. Pathol. 8 (11), 14521–14527.
Tang, J., Gifford, C. C., Samarakoon, R., and Higgins, P. J. (2018). Deregulation of negative controls on TGF-β1 signaling in tumor progression. Cancers 10 (6), 159. doi:10.3390/cancers10060159
Tang, J-Y., Lee, J-C., Chang, Y-T., Hou, M-F., Huang, H-W., Liaw, C-C., et al. (2013). Long noncoding RNAs-related diseases, cancers, and drugs. Sci. World J. 2013, 943539. doi:10.1155/2013/943539
Tetreault, M-P., Yang, Y., and Katz, J. P. (2013). Krüppel-like factors in cancer. Nat. Rev. Cancer 13 (10), 701–713. doi:10.1038/nrc3582
Thébaud, B., and Stewart, D. J. (2012). Exosomes: Cell garbage can, therapeutic carrier, or trojan horse? Am Heart Assoc, 2553–2555.
To, K. H., Pajovic, S., Gallie, B. L., and Thériault, B. L. (2012). Regulation of p14ARF expression by miR-24: A potential mechanism compromising the p53 response during retinoblastoma development. BMC cancer 12, 69. doi:10.1186/1471-2407-12-69
Tsai, M-C., Spitale, R. C., and Chang, H. Y. (2011). Long intergenic noncoding RNAs: New links in cancer progression. Cancer Res. 71 (1), 3–7. doi:10.1158/0008-5472.CAN-10-2483
Tsai, Y-F., Tseng, L-M., Hsu, C-Y., Yang, M-H., Chiu, J-H., and Shyr, Y-M. (2017). Brain-derived neurotrophic factor (BDNF)-TrKB signaling modulates cancer-endothelial cells interaction and affects the outcomes of triple negative breast cancer. PLoS One 12 (6), e0178173. doi:10.1371/journal.pone.0178173
Uematsu, M., Nishimura, T., Sakamaki, Y., Yamamoto, H., and Mizushima, N. (2017). Accumulation of undegraded autophagosomes by expression of dominant-negative STX17 (syntaxin 17) mutants. Autophagy 13 (8), 1452–1464. doi:10.1080/15548627.2017.1327940
Umemura, S., Shirane, M., Takekoshi, S., Kusakabe, T., Itoh, J., Egashira, N., et al. (2009). Overexpression of E2F-5 correlates with a pathological basal phenotype and a worse clinical outcome. Br. J. Cancer 100 (5), 764–771. doi:10.1038/sj.bjc.6604900
Varambally, S., Dhanasekaran, S. M., Zhou, M., Barrette, T. R., Kumar-Sinha, C., Sanda, M. G., et al. (2002). The polycomb group protein EZH2 is involved in progression of prostate cancer. Nature 419 (6907), 624–629. doi:10.1038/nature01075
Vos, P. D., Leedman, P. J., Filipovska, A., and Rackham, O. (2019). Modulation of miRNA function by natural and synthetic RNA-binding proteins in cancer. Cell. Mol. Life Sci. 76 (19), 3745–3752. doi:10.1007/s00018-019-03163-9
Wan, W., Wan, W., Long, Y., Li, Q., Jin, X., Wan, G., et al. (2019). MiR-25-3p promotes malignant phenotypes of retinoblastoma by regulating PTEN/Akt pathway. Biomed. Pharmacother. 118, 109111. doi:10.1016/j.biopha.2019.109111
Wan, Z., Zhi, N., Wong, S., Keyvanfar, K., Liu, D., Raghavachari, N., et al. (2010). Human parvovirus B19 causes cell cycle arrest of human erythroid progenitors via deregulation of the E2F family of transcription factors. J. Clin. Invest. 120 (10), 3530–3544. doi:10.1172/JCI41805
Wang, B., Heath-Engel, H., Zhang, D., Nguyen, N., Thomas, D. Y., Hanrahan, J. W., et al. (2008). BAP31 interacts with Sec61 translocons and promotes retrotranslocation of CFTRDeltaF508 via the derlin-1 complex. Cell 133 (6), 1080–1092. doi:10.1016/j.cell.2008.04.042
Wang, F., Zhu, Y., Huang, Y., McAvoy, S., Johnson, W. B., Cheung, T. H., et al. (2005). Transcriptional repression of WEE1 by Kruppel-like factor 2 is involved in DNA damage-induced apoptosis. Oncogene 24 (24), 3875–3885. doi:10.1038/sj.onc.1208546
Wang, G., Wang, J-J., Fu, X-L., Guang, R., and To, S-S. T. (2017). Advances in the targeting of HIF-1α and future therapeutic strategies for glioblastoma multiforme (Review). Oncol. Rep. 37 (2), 657–670. doi:10.3892/or.2016.5309
Wang, H., Li, M., Cui, H., Song, X., and Sha, Q. (2020). CircDHDDS/miR-361-3p/WNT3A Axis promotes the development of retinoblastoma by regulating proliferation, cell cycle, migration, and invasion of retinoblastoma cells. Neurochem. Res. 45 (11), 2691–2702. doi:10.1007/s11064-020-03112-0
Wang, H., Zhang, Z., Zhang, Y., and Li, L. (2022). Knockdown of the long noncoding RNA TUG1 suppresses retinoblastoma progression by disrupting the epithelial-mesenchymal transition. Cell Transpl. 31, 9636897221078026. doi:10.1177/09636897221078026
Wang, H., Zhang, Z., Zhang, Y., Liu, S., and Li, L. (2021). Long non-coding RNA TP53TG1 upregulates SHCBP1 to promote retinoblastoma progression by sponging miR-33b. Cell Transpl. 30, 9636897211025223. doi:10.1177/09636897211025223
Wang, H-G., Cao, B., Zhang, L-X., Song, N., Li, H., Zhao, W-Z., et al. (2017). KLF2 inhibits cell growth via regulating HIF-1α/Notch-1 signal pathway in human colorectal cancer HCT116 cells. Oncol. Rep. 38 (1), 584–590. doi:10.3892/or.2017.5708
Wang, J., Liu, X. H., Yang, Z. J., Xie, B., and Zhong, Y. S. (2014). The effect of ROCK-1 activity change on the adhesive and invasive ability of Y79 retinoblastoma cells. BMC Cancer 14, 89. doi:10.1186/1471-2407-14-89
Wang, J. X., Yang, Y., and Li, K. (2018). Long noncoding RNA DANCR aggravates retinoblastoma through miR-34c and miR-613 by targeting MMP-9. J. Cell. Physiol. 233 (10), 6986–6995. doi:10.1002/jcp.26621
Wang, L., Lyu, X., Ma, Y., Wu, F., and Wang, L. (2019). MicroRNA-504 targets AEG-1 and inhibits cell proliferation and invasion in retinoblastoma. Mol. Med. Rep. 19 (4), 2935–2942. doi:10.3892/mmr.2019.9923
Wang, L., Wang, L., Li, L., Zhang, H., and Lyu, X. (2019). MicroRNA-330 is downregulated in retinoblastoma and suppresses cell viability and invasion by directly targeting ROCK1. Mol. Med. Rep. 20 (4), 3440–3447. doi:10.3892/mmr.2019.10545
Wang, L., Wang, L., and Zhang, X. (2019). Knockdown of lncRNA HOXA-AS2 inhibits viability, migration and invasion of osteosarcoma cells by miR-124-3p/E2F3. Onco. Targets. Ther. 12, 10851–10861. doi:10.2147/OTT.S220072
Wang, L., Yi, S., Wang, R., and Wang, J. (2021). Long non-coding RNA KCNQ1OT1 promotes proliferation, migration and invasion via targeting miR-134 in retinoblastoma. J. Gene Med. 23 (6), e3336. doi:10.1002/jgm.3336
Wang, L., Zhang, Y., and Xin, X. (2020). Long non-coding RNA MALAT1 aggravates human retinoblastoma by sponging miR-20b-5p to upregulate STAT3. Pathol. Res. Pract. 216 (6), 152977. doi:10.1016/j.prp.2020.152977
Wang, L. L., Feng, Y. Q., and Cheng, Y. H. (2017). Effect on proliferation and apoptosis of retinoblastoma cell by RNA inhibiting high mobility group protein box-1 expression. Int. J. Ophthalmol. 10 (1), 30–34. doi:10.18240/ijo.2017.01.05
Wang, L. L., Hu, H. F., and Feng, Y. Q. (2016). Suppressive effect of microRNA-143 in retinoblastoma. Int. J. Ophthalmol. 9 (11), 1584–1590. doi:10.18240/ijo.2016.11.08
Wang, M., Yu, F., Wu, W., Zhang, Y., Chang, W., Ponnusamy, M., et al. (2017). Circular RNAs: A novel type of non-coding RNA and their potential implications in antiviral immunity. Int. J. Biol. Sci. 13 (12), 1497–1506. doi:10.7150/ijbs.22531
Wang, M., Zhao, X., Zhu, D., Liu, T., Liang, X., Liu, F., et al. (2017). HIF-1α promoted vasculogenic mimicry formation in hepatocellular carcinoma through LOXL2 up-regulation in hypoxic tumor microenvironment. J. Exp. Clin. Cancer Res. 36 (1), 60–14. doi:10.1186/s13046-017-0533-1
Wang, P-S., Wang, Z., and Yang, C. (Editors) (2021). “Dysregulations of long non-coding RNAs− the emerging “lnc” in environmental carcinogenesis,” Seminars in cancer biology (Elsevier).
Wang, Q., Zhu, Y., Zuo, G., Chen, X., Cheng, J., and Zhang, S. (2020). LINC00858 promotes retinoblastoma cell proliferation, migration and invasion by inhibiting miR-3182. Exp. Ther. Med. 19 (2), 999–1005. doi:10.3892/etm.2019.8294
Wang, R., Geng, N., Zhou, Y., Zhang, D., Li, L., Li, J., et al. (2015). Aberrant Wnt-1/beta-catenin signaling and WIF-1 deficiency are important events which promote tumor cell invasion and metastasis in salivary gland adenoid cystic carcinoma. Biomed. Mat. Eng. 26 (1), S2145–S2153. doi:10.3233/BME-151520
Wang, S., Liu, J., Yang, Y., Hao, F., and Zhang, L. (2018). PlncRNA-1 is overexpressed in retinoblastoma and regulates retinoblastoma cell proliferation and motility through modulating CBR3. IUBMB life 70 (10), 969–975. doi:10.1002/iub.1886
Wang, X., Li, X., Lin, F., Sun, H., Lin, Y., Wang, Z., et al. (2021). The lnc-CTSLP8 upregulates CTSL1 as a competitive endogenous RNA and promotes ovarian cancer metastasis. J. Exp. Clin. Cancer Res. 40 (1), 151–217. doi:10.1186/s13046-021-01957-z
Wang, X., Su, Y., and Yin, C. (2021). Long non-coding RNA (lncRNA) five prime to Xist (FTX) promotes retinoblastoma progression by regulating the microRNA-320a/with-no-lysine kinases 1 (WNK1) axis. Bioengineered 12 (2), 11622–11633. doi:10.1080/21655979.2021.1994718
Wang, Y., Lian, Y., and Ge, C. (2019). MiR-145 changes sensitivity of non-small cell lung cancer to gefitinib through targeting ADAM19. Eur. Rev. Med. Pharmacol. Sci. 23 (13), 5831–5839. doi:10.26355/eurrev_201907_18323
Wang, Y., Mo, Y., Gong, Z., Yang, X., Yang, M., Zhang, S., et al. (2017). Circular RNAs in human cancer. Mol. Cancer 16 (1), 25–28. doi:10.1186/s12943-017-0598-7
Wang, Y., Sun, D., Sheng, Y., Guo, H., Meng, F., and Song, T. (2020). XIST promotes cell proliferation and invasion by regulating miR-140-5p and SOX4 in retinoblastoma. World J. Surg. Oncol. 18 (1), 49. doi:10.1186/s12957-020-01825-8
Wang, Y., Wang, J., Hao, H., and Luo, X. (2020). lncRNA KCNQ1OT1 promotes the proliferation, migration and invasion of retinoblastoma cells by upregulating HIF-1α via sponging miR-153-3p. J. Investig. Med. 68 (8), 1349–1356. doi:10.1136/jim-2020-001431
Wang, Y., Xin, D., and Zhou, L. (2020). LncRNA LINC00152 increases the aggressiveness of human retinoblastoma and enhances carboplatin and adriamycin resistance by regulating MiR-613/yes-associated protein 1 (YAP1) Axis. Med. Sci. Monit. 26, e920886. doi:10.12659/MSM.920886
Wang, Z., Yao, Y. J., Zheng, F., Guan, Z., Zhang, L., Dong, N., et al. (2017). Mir-138-5p acts as a tumor suppressor by targeting pyruvate dehydrogenase kinase 1 in human retinoblastoma. Eur. Rev. Med. Pharmacol. Sci. 21 (24), 5624–5629. doi:10.26355/eurrev_201712_14005
Watson, A. P., Evans, R. L., and Egland, K. A. (2013). Multiple functions of sushi domain containing 2 (SUSD2) in breast tumorigenesis. Mol. Cancer Res. 11 (1), 74–85. doi:10.1158/1541-7786.MCR-12-0501-T
Wei, D., Yang, L., Lv, B., and Chen, L. (2017). Genistein suppresses retinoblastoma cell viability and growth and induces apoptosis by upregulating miR-145 and inhibiting its target ABCE1. Mol. Vis. 23, 385–394.
Wildeboer, D., Naus, S., Amy Sang, Q. X., Bartsch, J. W., and Pagenstecher, A. (2006). Metalloproteinase disintegrins ADAM8 and ADAM19 are highly regulated in human primary brain tumors and their expression levels and activities are associated with invasiveness. J. Neuropathol. Exp. Neurol. 65 (5), 516–527. doi:10.1097/01.jnen.0000229240.51490.d3
Wu, A., Zhou, X., Mi, L., and Shen, J. (2020). LINC00202 promotes retinoblastoma progression by regulating cell proliferation, apoptosis, and aerobic glycolysis through miR-204-5p/HMGCR axis. Open Life Sci. 15 (1), 437–448. doi:10.1515/biol-2020-0047
Wu, J., Fang, J., Yang, Z., Chen, F., Liu, J., and Wang, Y. (2012). Wnt inhibitory factor-1 regulates glioblastoma cell cycle and proliferation. J. Clin. Neurosci. 19 (10), 1428–1432. doi:10.1016/j.jocn.2011.12.023
Wu, L., Chen, Z., and Xing, Y. (2018). MiR-506-3p inhibits cell proliferation, induces cell cycle arrest and apoptosis in retinoblastoma by directly targeting NEK6. Cell Biol. Int. doi:10.1002/cbin.11041
Wu, L., Chen, Z., Zhang, J., and Xing, Y. (2012). Effect of miR-513a-5p on etoposide-stimulating B7-H1 expression in retinoblastoma cells. J. Huazhong Univ. Sci. Technol. Med. Sci. = Hua 32 (4), 601–606. doi:10.1007/s11596-012-1004-8
Wu, S., Ai, N., Liu, Q., and Zhang, J. (2018). MicroRNA-448 inhibits the progression of retinoblastoma by directly targeting ROCK1 and regulating PI3K/AKT signalling pathway. Oncol. Rep. 39 (5), 2402–2412. doi:10.3892/or.2018.6302
Wu, S., Han, M., and Zhang, C. (2019). Overexpression of microRNA-186 inhibits angiogenesis in retinoblastoma via the Hedgehog signaling pathway by targeting ATAD2. J. Cell. Physiol. 234 (10), 19059–19072. doi:10.1002/jcp.28545
Wu, X., Zeng, Y., Wu, S., Zhong, J., Wang, Y., and Xu, J. (2015). MiR-204, down-regulated in retinoblastoma, regulates proliferation and invasion of human retinoblastoma cells by targeting CyclinD2 and MMP-9. FEBS Lett. 589 (5), 645–650. doi:10.1016/j.febslet.2015.01.030
Wu, X. Z., Cui, H. P., Lv, H. J., and Feng, L. (2019). Knockdown of lncRNA PVT1 inhibits retinoblastoma progression by sponging miR-488-3p. Biomed. Pharmacother. 112, 108627. doi:10.1016/j.biopha.2019.108627
Wu, Z., Chen, L., Zhang, C., and Liu, Y. (2021). miR-4319 inhibited retinoblastoma cells proliferation, migration, invasion and EMT progress via suppressing CD147 mediated MMPs expression. J. Mol. Histol. 52 (2), 269–277. doi:10.1007/s10735-020-09946-w
Xia, G., Wang, H., Song, Z., Meng, Q., Huang, X., and Huang, X. (2017). Gambogic acid sensitizes gemcitabine efficacy in pancreatic cancer by reducing the expression of ribonucleotide reductase subunit-M2 (RRM2). J. Exp. Clin. Cancer Res. 36 (1), 107–114. doi:10.1186/s13046-017-0579-0
Xia, W., Wang, L., Yu, D., Mu, X., and Zhou, X. (2019). Lidocaine inhibits the progression of retinoblastoma in vitro and in vivo by modulating the miR-520a-3p/EGFR axis. Mol. Med. Rep. 20 (2), 1333–1342. doi:10.3892/mmr.2019.10363
Xia, Z., Yang, X., Wu, S., Feng, Z., Qu, L., Chen, X., et al. (2019). LncRNA TP73-AS1 down-regulates miR-139-3p to promote retinoblastoma cell proliferation. Biosci. Rep. 39 (5), BSR20190475. doi:10.1042/BSR20190475
Xiang, J., Wu, Y., Li, D-S., Wang, Z-Y., Shen, Q., Sun, T-Q., et al. (2015). miR-584 suppresses invasion and cell migration of thyroid carcinoma by regulating the target oncogene ROCK1. Oncol. Res. Treat. 38 (9), 436–440. doi:10.1159/000438967
Xie, H., Kang, Y., Wang, S., Zheng, P., Chen, Z., Roy, S., et al. (2020). E2f5 is a versatile transcriptional activator required for spermatogenesis and multiciliated cell differentiation in zebrafish. PLoS Genet. 16 (3), e1008655. doi:10.1371/journal.pgen.1008655
Xu, C., and Tian, L. H. (2020). LncRNA XIST promotes proliferation and epithelial-mesenchymal transition of retinoblastoma cells through sponge action of miR-142-5p. Eur. Rev. Med. Pharmacol. Sci. 24 (18), 9256–9264. doi:10.26355/eurrev_202009_23007
Xu, F., Liu, G., Wang, L., Wang, X., Jin, X., and Bo, W. (2020). miR-494 promotes progression of retinoblastoma via PTEN through PI3K/AKT signaling pathway. Oncol. Lett. 20 (2), 1952–1960. doi:10.3892/ol.2020.11749
Xu, H. K., Wang, X. D., Wang, D. G., Wei, D. D., Liang, L., and Liu, C. H. (2021). miR-340 exerts suppressive effect on retinoblastoma progression by targeting KIF14. Curr. Eye Res. 46 (2), 232–238. doi:10.1080/02713683.2020.1795202
Xu, L., Li, W., Shi, Q., Wang, M., Li, H., Yang, X., et al. (2020). MicroRNA-936 inhibits the malignant phenotype of retinoblastoma by directly targeting HDAC9 and deactivating the PI3K/AKT pathway. Oncol. Rep. 43 (2), 635–645. doi:10.3892/or.2020.7456
Xu, L., Long, H., Zhou, B., Jiang, H., and Cai, M. (2021). CircMKLN1 suppresses the progression of human retinoblastoma by modulation of miR-425-5p/PDCD4 Axis. Curr. Eye Res. 46 (11), 1751–1761. doi:10.1080/02713683.2021.1927110
Xu, L., Zhu, S., Tang, A., and Liu, W. (2021). LncRNA MBLN1-AS1 inhibits the progression of retinoblastoma through targeting miR-338-5p-Wnt/β-catenin signaling pathway. Inflamm. Res. 70 (2), 217–227. doi:10.1007/s00011-020-01432-z
Xu, X., Jia, R., Zhou, Y., Song, X., Wang, J., Qian, G., et al. (2011). Microarray-based analysis: Identification of hypoxia-regulated microRNAs in retinoblastoma cells. Int. J. Oncol. 38 (5), 1385–1393. doi:10.3892/ijo.2011.961
Xu, X., Zhao, Y., Duan, G., and Du, B. (2021). Downregulation of MIAT reduces the proliferation and migratory and invasive abilities of retinoblastoma cells by sponging miR-665 and regulating LASP1. Exp. Ther. Med. 22 (5), 1342. doi:10.3892/etm.2021.10777
Xu, Y., Fu, Z., Gao, X., Wang, R., and Li, Q. (2021). Long non-coding RNA XIST promotes retinoblastoma cell proliferation, migration, and invasion by modulating microRNA-191-5p/brain derived neurotrophic factor. Bioengineered 12 (1), 1587–1598. doi:10.1080/21655979.2021.1918991
Yan, G., Su, Y., Ma, Z., Yu, L., and Chen, N. (2019). Long noncoding RNA LINC00202 promotes tumor progression by sponging miR-3619-5p in retinoblastoma. Cell Struct. Funct. 44 (1), 51–60. doi:10.1247/csf.18033
Yan, J., Deng, Y. X., Cai, Y. L., and Cong, W. D. (2022). LncRNA MIR17HG promotes the proliferation, migration, and invasion of retinoblastoma cells by up-regulating HIF-1α expression via sponging miR-155-5p. Kaohsiung J. Med. Sci. 38 (6), 554–564. doi:10.1002/kjm2.12523
Yan, P., Liu, J., Zhou, R., Lin, C., Wu, K., Yang, S., et al. (2020). LASP1 interacts with N-WASP to activate the Arp2/3 complex and facilitate colorectal cancer metastasis by increasing tumour budding and worsening the pattern of invasion. Oncogene 39 (35), 5743–5755. doi:10.1038/s41388-020-01397-7
Yan, T-f., Wu, M-j., Xiao, B., Hu, Q., Fan, Y-H., and Zhu, X-G. (2018). Knockdown of HOXC6 inhibits glioma cell proliferation and induces cell cycle arrest by targeting WIF-1 in vitro and vivo. Pathol. Res. Pract. 214 (11), 1818–1824. doi:10.1016/j.prp.2018.09.001
Yanai, H., Ban, T., Wang, Z., Choi, M. K., Kawamura, T., Negishi, H., et al. (2009). HMGB proteins function as universal sentinels for nucleic-acid-mediated innate immune responses. Nature 462 (7269), 99–103. doi:10.1038/nature08512
Yang, F., Zhou, X., Du, S., Zhao, Y., Ren, W., Deng, Q., et al. (2014). LIM and SH3 domain protein 1 (LASP-1) overexpression was associated with aggressive phenotype and poor prognosis in clear cell renal cell cancer. PloS one 9 (6), e100557. doi:10.1371/journal.pone.0100557
Yang, G., Fu, Y., Lu, X., Wang, M., Dong, H., and Li, Q. (2019). miR-34a regulates the chemosensitivity of retinoblastoma cells via modulation of MAGE-A/p53 signaling. Int. J. Oncol. 54 (1), 177–187. doi:10.3892/ijo.2018.4613
Yang, G., Fu, Y., Zhang, L., Lu, X., and Li, Q. (2017). miR106b regulates retinoblastoma Y79 cells through Runx3. Oncol. Rep. 38 (5), 3039–3043. doi:10.3892/or.2017.5931
Yang, H., Huang, Y., He, J., Chai, G., Di, Y., Wang, A., et al. (2020). MiR-486-3p inhibits the proliferation, migration and invasion of retinoblastoma cells by targeting ECM1. Biosci. Rep. 40 (6), BSR20200392. doi:10.1042/BSR20200392
Yang, L., Wei, N., Wang, L., Wang, X., and Liu, Q. H. (2018). miR-498 promotes cell proliferation and inhibits cell apoptosis in retinoblastoma by directly targeting CCPG1. Childs Nerv. Syst. 34 (3), 417–422. doi:10.1007/s00381-017-3622-8
Yang, L., Zhang, L., Lu, L., and Wang, Y. (2020). lncRNA UCA1 increases proliferation and multidrug resistance of retinoblastoma cells through downregulating miR-513a-5p. DNA Cell Biol. 39 (1), 69–77. doi:10.1089/dna.2019.5063
Yang, L., Zhang, L., Lu, L., and Wang, Y. (2019). Long noncoding RNA SNHG16 sponges miR-182-5p and miR-128-3p to promote retinoblastoma cell migration and invasion by targeting LASP1. Onco. Targets. Ther. 12, 8653–8662. doi:10.2147/OTT.S212352
Yang, L., Zhang, L., Lu, L., and Wang, Y. (2020). miR-214-3p regulates multi-drug resistance and apoptosis in retinoblastoma cells by targeting ABCB1 and XIAP. Onco. Targets. Ther. 13, 803–811. doi:10.2147/OTT.S235862
Yang, L. L., Li, Q., Zhang, X., and Cao, T. (2020). Long non-coding RNA XIST confers aggressive progression via miR-361-3p/STX17 in retinoblastoma cells. Eur. Rev. Med. Pharmacol. Sci. 24 (20), 10433–10444. doi:10.26355/eurrev_202010_23395
Yao, L., Yang, L., Song, H., Liu, T. G., and Yan, H. (2020). Silencing of lncRNA XIST suppresses proliferation and autophagy and enhances vincristine sensitivity in retinoblastoma cells by sponging miR-204-5p. Eur. Rev. Med. Pharmacol. Sci. 24 (7), 3526–3537. doi:10.26355/eurrev_202004_20812
Yao, S., and Yan, W. (2018). Overexpression of Mst1 reduces gastric cancer cell viability by repressing the AMPK-Sirt3 pathway and activating mitochondrial fission. Onco. Targets. Ther. 11, 8465–8479. doi:10.2147/OTT.S180851
Yao, X., Shen, H., Peng, Q., and Yu, J. (2021). TP53/miR-129/MDM2/4/TP53 feedback loop modulates cell proliferation and apoptosis in retinoblastoma. Cell cycleGeorget. Tex) 20 (5-6), 603–615. doi:10.1080/15384101.2021.1892321
Yin, D. F., Zhou, X. J., Li, N., Liu, H. J., and Yuan, H. (2021). Long non-coding RNA SND1-IT1 accelerates cell proliferation, invasion and migration via regulating miR-132-3p/SMAD2 axis in retinoblastoma. Bioengineered 12 (1), 1189–1201. doi:10.1080/21655979.2021.1909962
Yin, X., Liao, Y., Xiong, W., Zhang, Y., Zhou, Y., and Yang, Y. (2020). Hypoxia-induced lncRNA ANRIL promotes cisplatin resistance in retinoblastoma cells through regulating ABCG2 expression. Clin. Exp. Pharmacol. Physiol. 47 (6), 1049–1057. doi:10.1111/1440-1681.13279
Yu, B., Zhao, J., and Dong, Y. (2021). Circ_0000527 promotes retinoblastoma progression through modulating miR-98-5p/XIAP pathway. Curr. Eye Res. 46 (9), 1414–1423. doi:10.1080/02713683.2021.1891255
Yuan, X., Sun, Z., and Cui, C. (2021). Knockdown of lncRNA HOTTIP inhibits retinoblastoma progression by modulating the miR-101-3p/STC1 Axis. Technol. Cancer Res. Treat. 20, 1533033821997831. doi:10.1177/1533033821997831
Yun, M-S., Kim, S-E., Jeon, S. H., Lee, J-S., and Choi, K-Y. (2005). Both ERK and Wnt/beta-catenin pathways are involved in Wnt3a-induced proliferation. J. Cell Sci. 118 (2), 313–322. doi:10.1242/jcs.01601
Zehavi, L., Schayek, H., Jacob-Hirsch, J., Sidi, Y., Leibowitz-Amit, R., and Avni, D. (2015). MiR-377 targets E2F3 and alters the NF-kB signaling pathway through MAP3K7 in malignant melanoma. Mol. Cancer 14, 68. doi:10.1186/s12943-015-0338-9
Zeng, Z., Li, Y., Pan, Y., Lan, X., Song, F., Sun, J., et al. (2018). Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat. Commun. 9 (1), 5395–5414. doi:10.1038/s41467-018-07810-w
Zhang, A., Shang, W., Nie, Q., Li, T., and Li, S. (2018). Long non-coding RNA H19 suppresses retinoblastoma progression via counteracting miR-17-92 cluster. J. Cell. Biochem. 119 (4), 3497–3509. doi:10.1002/jcb.26521
Zhang, B., Wang, Q., and Pan, X. (2007). MicroRNAs and their regulatory roles in animals and plants. J. Cell. Physiol. 210 (2), 279–289. doi:10.1002/jcp.20869
Zhang, C., and Wu, S. (2020). microRNA -378a-3p restrains the proliferation of retinoblastoma cells but promotes apoptosis of retinoblastoma cells via inhibition of FOXG1. Invest. Ophthalmol. Vis. Sci. 61 (5), 31. doi:10.1167/iovs.61.5.31
Zhang, G., Yang, W., Li, D., Li, X., Huang, J., Huang, R., et al. (2020). lncRNA FEZF1-AS1 promotes migration, invasion and epithelial-mesenchymal transition of retinoblastoma cells by targeting miR-1236-3p. Mol. Med. Rep. 22 (5), 3635–3644. doi:10.3892/mmr.2020.11478
Zhang, H., Qiu, X., Song, Z., Lan, L., Ren, X., and Ye, B. (2022). CircCUL2 suppresses retinoblastoma cells by regulating miR-214-5p/E2F2 Axis. Anticancer. Drugs 33 (1), e218–e227. doi:10.1097/CAD.0000000000001190
Zhang, H., Yang, X., Xu, Y., and Li, H. (2021). KCNQ1OT1 regulates the retinoblastoma cell proliferation, migration and SIRT1/JNK signaling pathway by targeting miR-124/SP1 axis. Biosci. Rep. 41 (1), BSR20201626. doi:10.1042/BSR20201626
Zhang, H., Zhong, J., Bian, Z., Fang, X., Peng, Y., and Hu, Y. (2017). Long non-coding RNA CCAT1 promotes human retinoblastoma SO-RB50 and Y79 cells through negative regulation of miR-218-5p. Biomed. Pharmacother. 87, 683–691. doi:10.1016/j.biopha.2017.01.004
Zhang, J., He, J., and Zhang, L. (2018). The down-regulation of microRNA-137 contributes to the up-regulation of retinoblastoma cell proliferation and invasion by regulating COX-2/PGE2 signaling. Biomed. Pharmacother. 106, 35–42. doi:10.1016/j.biopha.2018.06.099
Zhang, J., Knight, R., Wang, Y., Sawyer, T. W., Martyniuk, C. J., and Langlois, V. S. (2019). Hair follicle miRNAs: A novel biomarker for primary blast induced-mild traumatic brain injury. Biomarkers 24 (2), 166–179. doi:10.1080/1354750X.2018.1531929
Zhang, L. J., Wang, F., Qi, P. Y., Zhou, W. Y., and Wang, B. (2021). miR-513b-5p inhibits the proliferation and promotes apoptosis of retinoblastoma cells by targeting TRIB1. Open Med. 16 (1), 1364–1371. doi:10.1515/med-2021-0343
Zhang, M., Li, Q., Pan, Y., Wang, H., Liu, G., and Yin, H. (2018). MicroRNA-655 attenuates the malignant biological behaviours of retinoblastoma cells by directly targeting PAX6 and suppressing the ERK and p38 MAPK signalling pathways. Oncol. Rep. 39 (4), 2040–2050. doi:10.3892/or.2018.6264
Zhang, P., Lu, Y., Liu, X. Y., and Zhou, Y. H. (2015). Knockdown of Rho-associated protein kinase 1 suppresses proliferation and invasion of glioma cells. Tumour Biol. 36 (1), 421–428. doi:10.1007/s13277-014-2673-7
Zhang, Q., Yu, L., Qin, D., Huang, R., Jiang, X., Zou, C., et al. (2015). Role of microRNA-30c targeting ADAM19 in colorectal cancer. PLoS One 10 (3), e0120698. doi:10.1371/journal.pone.0120698
Zhang, S., Long, J., and Hu, Y. (2020). Long noncoding RNA LINC00205 enhances the malignant characteristics of retinoblastoma by acting as a molecular sponge of microRNA-665 and consequently increasing HMGB1 expression. Biochem. Biophys. Res. Commun. 526 (2), 396–403. doi:10.1016/j.bbrc.2020.03.083
Zhang, T., Yang, J., Gong, F., Li, L., and Li, A. (2020). Long non-coding RNA CASC9 promotes the progression of retinoblastoma via interacting with miR-145-5p. Cell cycleGeorget. Tex) 19 (18), 2270–2280. doi:10.1080/15384101.2020.1802813
Zhang, W., Levi, L., Banerjee, P., Jain, M., and Noy, N. (2015). Kruppel-like factor 2 suppresses mammary carcinoma growth by regulating retinoic acid signaling. Oncotarget 6 (34), 35830–35842. doi:10.18632/oncotarget.5767
Zhang, X., Song, L., Huang, Y., Han, S., Hou, M., and Li, H. (2020). Downregulation of MST4 underlies a novel inhibitory role of MicroRNA let-7a in the progression of retinoblastoma. Invest. Ophthalmol. Vis. Sci. 61 (6), 28. doi:10.1167/iovs.61.6.28
Zhang, Y., Dou, X., Kong, Q., Li, Y., and Zhou, X. (2022). Circ_0075804 promotes the malignant behaviors of retinoblastoma cells by binding to miR-138-5p to induce PEG10 expression. Int. Ophthalmol. 42 (2), 509–523. doi:10.1007/s10792-021-02067-7
Zhang, Y., Wang, X., and Zhao, Y. (2018). MicroRNA-874 prohibits the proliferation and invasion of retinoblastoma cells by directly targeting metadherin. Mol. Med. Rep. 18 (3), 3099–3105. doi:10.3892/mmr.2018.9295
Zhang, Y., Wu, D., Xia, F., Xian, H., Zhu, X., Cui, H., et al. (2016). Downregulation of HDAC9 inhibits cell proliferation and tumor formation by inducing cell cycle arrest in retinoblastoma. Biochem. Biophys. Res. Commun. 473 (2), 600–606. doi:10.1016/j.bbrc.2016.03.129
Zhang, Y., Wu, J. H., Han, F., Huang, J. M., Shi, S. Y., Gu, R. D., et al. (2013). Arsenic trioxide induced apoptosis in retinoblastoma cells by abnormal expression of microRNA-376a. Neoplasma 60 (3), 247–253. doi:10.4149/neo_2013_033
Zhang, Y., Xue, C., Zhu, X., Zhu, X., Xian, H., and Huang, Z. (2016). Suppression of microRNA-125a-5p upregulates the TAZ-EGFR signaling pathway and promotes retinoblastoma proliferation. Cell. Signal. 28 (8), 850–860. doi:10.1016/j.cellsig.2016.04.002
Zhang, Y., and Zheng, A. (2021). MiR-142-5p promotes retinoblastoma cell proliferation, migration and invasion by targeting PTEN. J. Biochem. 170 (2), 195–202. doi:10.1093/jb/mvaa121
Zhang, Y., Zhu, X., Zhu, X., Wu, Y., Liu, Y., Yao, B., et al. (2017). MiR-613 suppresses retinoblastoma cell proliferation, invasion, and tumor formation by targeting E2F5. Tumour Biol. 39 (3), 1010428317691674. doi:10.1177/1010428317691674
Zhao, H., Wan, J., and Zhu, Y. (2020). Carboplatin inhibits the progression of retinoblastoma through IncRNA XIST/miR-200a-3p/NRP1 Axis. Drug Des. devel. Ther. 14, 3417–3427. doi:10.2147/DDDT.S256813
Zhao, W., An, Y., Liang, Y., and Xie, X. (2014). Role of HOTAIR long noncoding RNA in metastatic progression of lung cancer. Eur. Rev. Med. Pharmacol. Sci. 18 (13), 1930–1936.
Zhao, W., Wang, S., Qin, T., and Wang, W. (2020). Circular RNA (circ-0075804) promotes the proliferation of retinoblastoma via combining heterogeneous nuclear ribonucleoprotein K (HNRNPK) to improve the stability of E2F transcription factor 3 E2F3. J. Cell. Biochem. 121 (7), 3516–3525. doi:10.1002/jcb.29631
Zhao, Y., Zhang, S., and Zhang, Y. (2017). MicroRNA-320 inhibits cell proliferation, migration and invasion in retinoblastoma by targeting specificity protein 1. Mol. Med. Rep. 16 (2), 2191–2198. doi:10.3892/mmr.2017.6767
Zhao, Z., Liu, W., Liu, J., Wang, J., and Luo, B. (2017). The effect of EBV on WIF1, NLK, and APC gene methylation and expression in gastric carcinoma and nasopharyngeal cancer. J. Med. Virol. 89 (10), 1844–1851. doi:10.1002/jmv.24863
Zheng, Q., Zhu, Q., Li, C., Hao, S., Li, J., Yu, X., et al. (2020). microRNA-144 functions as a diagnostic and prognostic marker for retinoblastoma. Clinics 75, e1804. doi:10.6061/clinics/2020/e1804
Zheng, T., Chen, W., Wang, X., Cai, W., Wu, F., and Lin, C. (2021). Circular RNA circ-FAM158A promotes retinoblastoma progression by regulating miR-138-5p/SLC7A5 axis. Exp. Eye Res. 211, 108650. doi:10.1016/j.exer.2021.108650
Zhong, W., Yang, J., Li, M., Li, L., and Li, A. (2019). Long noncoding RNA NEAT1 promotes the growth of human retinoblastoma cells via regulation of miR-204/CXCR4 axis. J. Cell. Physiol. 234 (7), 11567–11576. doi:10.1002/jcp.27812
Zhou, Y., Li, Z., Ding, Y., Zhang, P., Wang, J., Zhang, J., et al. (2018). Promoter methylation of WNT inhibitory factor-1 may be associated with the pathogenesis of multiple human tumors. J. Cancer Res. Ther. 14 (9), S381. doi:10.4103/0973-1482.235357
Zhu, Y., and Hao, F. (2021). Knockdown of long non-coding RNA HCP5 suppresses the malignant behavior of retinoblastoma by sponging miR-3619-5p to target HDAC9. Int. J. Mol. Med. 47 (5), 74. doi:10.3892/ijmm.2021.4907
Zou, J. Y., and Crews, F. T. (2014). Release of neuronal HMGB1 by ethanol through decreased HDAC activity activates brain neuroimmune signaling. PloS one 9 (2), e87915. doi:10.1371/journal.pone.0087915
Keywords: Retinoblastma, non-coding RNAs, exosome, microRNA, circular RNA
Citation: Ahangar Davoodi N, Najafi S, Naderi Ghale-Noie Z, Piranviseh A, Mollazadeh S, Ahmadi Asouri S, Asemi Z, Morshedi M, Tamehri Zadeh SS, Hamblin MR, Sheida A and Mirzaei H (2022) Role of non-coding RNAs and exosomal non-coding RNAs in retinoblastoma progression. Front. Cell Dev. Biol. 10:1065837. doi: 10.3389/fcell.2022.1065837
Received: 10 October 2022; Accepted: 05 December 2022;
Published: 23 December 2022.
Edited by:
Viviana C. Valadez-Graham, Institute of Biotechnology, National Autonomous University of Mexico, MexicoReviewed by:
Vanesa Olivares-Illana, Autonomous University of San Luis Potosi, MexicoCopyright © 2022 Ahangar Davoodi, Najafi, Naderi Ghale-Noie, Piranviseh, Mollazadeh, Ahmadi Asouri, Asemi, Morshedi, Tamehri Zadeh, Hamblin, Sheida and Mirzaei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amirhossein Sheida, c2hlaWRhYW1pcmhvc3NlaW5AZ21haWwuY29t; Hamed Mirzaei, bWlyemFlaS1oQGthdW1zLmFjLmly, aC5taXJ6YWVpMjAwMkBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.