 Lydia García-Gómez
Lydia García-Gómez Iker Castillo-Fernández
Iker Castillo-Fernández Ana Perez-Villalba
Ana Perez-Villalba- Laboratory of Animal Behavior Phenotype (L.A.B.P.), Department of Neuropsychology, Faculty of Psychology, Catholic University of Valencia, Valencia, Spain
Social behaviors have become more relevant to our understanding of the human nervous system because relationships with our peers may require and modulate adult neurogenesis. Here, we review the pieces of evidence we have to date for the divergence of social behaviors in mice by modulation of adult neurogenesis or if social behaviors and the social environment can drive a change in neurogenic processes. Social recognition and memory are deeply affected by antimitotic drugs and irradiation, while NSC transgenic mice may run with lower levels of social discrimination. Interestingly, social living conditions can create a big impact on neurogenesis. Social isolation and social defeat reduce the number of new neurons, while social dominance and enrichment of the social environment increase their number. These new “social neurons” trigger functional modifications with amazing transgenerational effects. All of these suggest that we are facing two bidirectional intertwined variables, and the great challenge now is to understand the cellular and genetic mechanisms that allow this relationship to be used therapeutically.
1 Introduction
Mice and humans are social animals, and both share a clear preference for social contact, group living, and a natural curiosity for novel social stimuli. This social behavior is associated not only with costs, like resource competition, but also with benefits, like cooperative breeding (Lee, 1994). Additionally, both mice and humans may also share the generation of new neurons in the adult brain (Kempermann et al., 2018). Adult neurogenesis is a complex biological process that occurs in two neurogenic niches of the adult mouse brain: the dentate gyrus (DG) of the hippocampus (Kempermann et al., 2004) and the subependymal zone (SEZ) of the lateral ventricles (Lois and Alvarez-Buylla, 1994). Sophisticated stages of proliferation, specification, and maturation are necessary for neuroblasts to be integrated in neural circuits. How frequent and feasible is this process in the adult human brain is still under interesting debate (Lucassen et al., 2020; Alvarez-Buylla et al., 2022).
In rodents, cognitive and emotional variables have been repeatedly correlated with variations in adult neurogenesis (Lazarini et al., 2009; Aimone et al., 2011). Extrinsic factors that regulate neurogenesis include enriched environment, voluntary exercise, and diet, skipping one of the most important variables: social behaviors. However, in recent years, an increasing number of articles include social behaviors in the list of parameters causing adult neurogenesis variations (Gobshtis et al., 2017; Pereira-Caixeta et al., 2017; Pereira-Caixeta et al., 2018; Lunardi et al., 2021), even though there is no clear agreement of their mutual influence.
Social behavior includes an extensive mosaic of different and complex behaviors, usually underrepresented or simplified but increasingly correlated with neurogenesis. Here, we review and analyze possible bidirectional effects: whether alterations in adult neurogenesis modulate social behavior and if social behaviors change neurogenesis.
2 Materials and methods
We performed a systematic literature search in PubMed and WoS (open access) in accordance with the “Preferred Reporting Items for Systematic Reviews and Meta-Analyses” (PRISMA) guidelines (Page et al., 2021). The Boolean expression used for our search was as follows: “adult neurogenesis” AND “social behavior” AND “mice.” This search retrieved 112 articles. All the reviewed articles included randomization in the distribution of experimental subjects and were published in peer-reviewed journals without time boundaries. We defined inclusion/exclusion criteria. Inclusion criteria were as follows: 1) the article should report specific measures of neurogenesis, concerning stem cells, progenitor cells, or neuroblast; 2) the article should include experiments of social behavior (Supplementary Figure S1); and 3) social behavior could be a consequence or an inductor of neurogenesis changes. Exclusion criteria were as follows: a) reviews, b) book chapters, c) articles not using mice, and d) documents not written in English. Exclusion criteria affected six articles. For inclusion criteria accomplishment, 12 articles did not meet criteria (i) and 14 did not meet inclusion criteria (ii) or (iii) (PRISMA flow diagram shown in Figure 1). Therefore, we included a total of 80 articles in this systematic review (Supplementary Table S1). Additional references were used for contextualizing.
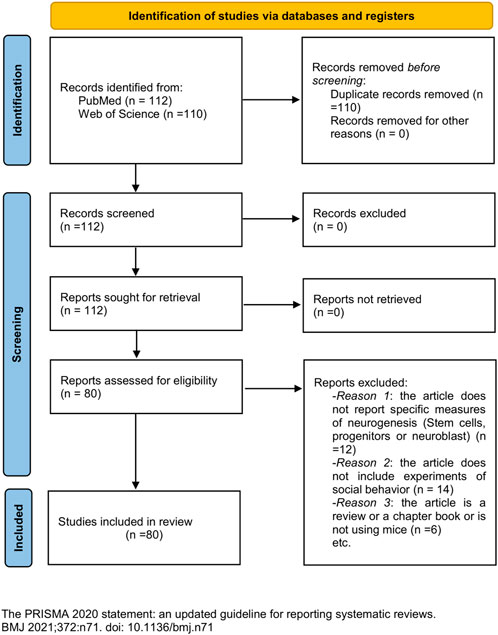
FIGURE 1. PRISMA flow diagram (2020 version).
3 Discussion
3.1 Irradiation, chemical damage, and chemical recovery of neurogenesis and their impact on social behavior
The most popular chemical methods to disrupt adult neurogenesis are brain delivery of cytosine-β-D-arabinofuranoside (AraC) and injection of temozolomide (TMZ). Both antimitotic drugs have been correlated with disrupted social recognition and memory, but the highest correlation was achieved by gamma irradiation (Pereira-Caixeta et al., 2018). Whole brain irradiation in adult mice drastically reduces subgranular zone (SGZ) cells by 93%–96% and immature neurons from 40% to 60% in a dose-dependent fashion (Mizumatsu et al., 2003), and it has been used as an acute and effective neurogenesis inhibitor. It also affects the dendritic complexity of new neurons, which is directly associated with social memory disruption even in young mice (Newton et al., 2020), especially when radiation affects CA2, a hippocampal region known to be important for social memory (Hitti and Siegelbaum, 2014) (Figure 2).
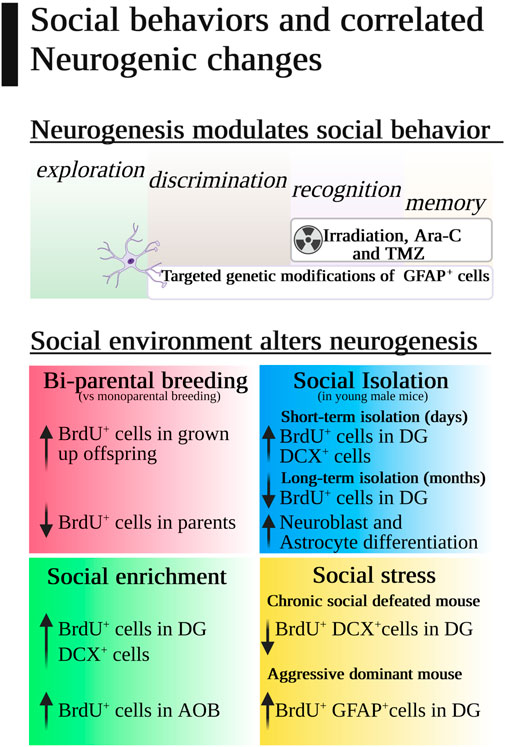
FIGURE 2. Social behaviors and correlated neurogenic changes.
But irradiation studies also demonstrated that adult neurogenesis is not a requirement for performing some social behaviors, such as maternal behavior, and for female mice to distinguish among pups. In contrast, adult olfactory neurogenesis is essential for normal social interaction, especially in male mice (Feierstein et al., 2010).
Additionally, chemical recovery of adult hippocampal neurogenesis (AHN) with psychoactive drugs has given rise to not fully understood results on social behavior. For example, memantine, the glutamate receptor antagonist, was associated with increased AHN (Maekawa et al., 2009) but not necessarily with an improvement in long-term social memory after previous 3 min of social contact, but it shows an association after reducing that time (Ishikawa et al., 2014).
3.2 Neurogenic changes induced by genetic mutations and the effect on social behavior
Unlike the local action of irradiation, many gene mutations are associated with a broad spectrum of abnormalities, like changes in development, neurotransmitters, neurogenesis, and behavior. We reproduce some examples here. The BTBR mouse model of autism deals with multisystem physiological and behavioral alterations, including social behavior deficits and a reduced number of bromodeoxyuridine (BrdU)+ cells, doublecortin (DCX)+ cells, nestin+ cells, and glial fibrillary acidic protein (GFAP)+ Sox2+ cells in the hippocampus, but researchers can normalize their neurogenesis and social behavior by transplanting human amniotic epithelial cells (Zhang R. et al., 2019). In contrast, another model of autism, PTEN (phosphatase and tensin homolog on chromosome 10) mutant mice, showed deficits in social exploration despite having increased Ki67+ cells in the DG (Amiri et al., 2012) in addition to developmental and electrophysiological alterations. In the same line of indirect relationships, knock-out mice of the cell adhesion molecule neuroplastin-65 showed anxious-like and depressive-like behavior as well as more BrdU+ cells in the DG and SEZ (Li et al., 2019). Here, more NSCs are associated with not only increased social interaction but also downregulation of serotonin affecting neurogenic niches and the limbic system. The same lack of specificity was described with poly (ADP-ribose) polymerase-1 (PARP-1, a multi-functional nuclear enzyme that regulates DNA repair). PARP-1 knock-out mice showed not only a high impact on NSC survival, DCX+ cells, and low neuronal differentiation that is associated with diminished social interaction (Hong et al., 2019) but also with symptoms described as “schizophrenia-like behavioral deficits” that include hyperactivity and anxiety. This is a similar profile to other “schizophrenia-like” DISC1 and BRINP1 mouse models. The mutant expression of DISC1 produced more anxious and depression-like behavior and reduced social exploration in the social chamber test as well as less proliferation of neural progenitors and dendritic maturation in the DG (Terrillion et al., 2017). Reduced neurogenesis is presented with reduced sociability (among other alterations). In BRINP1 KO mice, the lack of this cell cycle suppressor resulted in increased cell division and neurogenesis (Kobayashi et al., 2014). They also presented hyperactivity and slight impulsive response. Then, they showed a reduced duration of contacts in the social interaction test but a radical increase in total distance travelled suggesting that hyperactivity may affect sociability by reducing the length of contact despite preserving social curiosity (with an equal number of contacts). Once again, this is a scenario of multi-functional alterations.
These broad-spectrum variables are presented in articles that related social exploration and neurogenesis triggered by genetic mutations that affected numerous systems (Lehmann et al., 2013; Guha et al., 2014; Garrett et al., 2018; Alen et al., 2019; Zhang et al., 2019; Amar et al., 2021; Velloso et al., 2022), types of cells and proteins (Jenniches et al., 2016; Zhang et al., 2016; Salminen et al., 2017; Alén et al., 2019; Zhang et al., 2020; Nacer et al., 2021) plus multisystem drugs (Poletaeva et al., 2012; Osborn et al., 2013; Subbanna and Basavarajappa 2014; Chiu et al., 2015; Di Nuzzo et al., 2015; Castilla-Ortega et al., 2016; Fu et al., 2018; Jiang et al., 2020; Kim et al., 2020), or in addition to electrophysiological changes in neurons (Alachkar et al., 2013). Therefore, the extensive effects of genetic mutations make it difficult to deduce a direct effect on social behavior (Bagnall-Moreau et al., 2020).
Instead, when genetic modifications were specifically guided with inducible and reversible genetic models to reduce neurogenesis, social discrimination between a familiar and a new mouse could be correlated with the number of DCX in the DG (Garrett et al., 2015) without altering other behaviors. Pharmacological reduction of AHN with transgenic mice expressing herpes-simplex virus-TK (thymidine kinase) under the GFAP promoter affected social memory (Cope et al., 2020).
Transgenic GFAP-TK-GFP mice injected with ganciclovir in the lateral ventricle showed a massive elimination of DCX+ cells in the MOB (main olfactory bulb) and DG without affecting astrocyte lineage or microglia (Wei et al., 2011). Curiously, when the investigators eliminated neurogenesis (DCX+ cells) in adolescence, there were no changes in memory and anxiety tests, but social exploration was severely affected. However, when the same experiment was performed on adult mice, there was no effect on social behavior. Normal socialization requires neurogenesis during adolescence, but once socialization is established, neurogenesis was no longer required. It would be interesting to study if this effect can be reversed over time.
Finally, social recognition and hence social memory in mice depend on the olfactory system, which receive neuroblasts through the rostral migratory stream from the SEZ (Lledó and Valley, 2016). Accordingly, the reduction of DCX in the MOB has a functional correlation with social memory (Garrett et al., 2018) even with high levels of proliferating cell nuclear antigen (PCNA) in the SEZ, suggesting that social memory formation needs intact olfactory and hippocampal neurogenesis.
3.3 Voluntary exercise as a neurogenic booster with social behavior modulation
Voluntary exercise can increase AHN (van Praag et al., 1999), and now we know several social variables that could modulate this classic effect. When young male mice are isolated and stressed, exercise increases short-term (Kannangara et al., 2009) and long-term (Hueston et al., 2017) BrdU+-retaining cells but only if they are left isolated and the effect is not reproduced if they are grouped, supporting the idea that social housing can buffer the effects of stress. Actually, the more severe and stressful the conditions in which neurogenesis is reduced, the greater the effect of anxiolytic variables, such as physical exercise and social contact. Moreover, social isolation during adolescence prevented exercise-induced neurogenesis in the ventral hippocampus with similar numbers in long-term BrdU+ and NeuN+ cells with or without voluntary wheel running (Kozareva et al., 2018). Another study found that environmental and social complexity did not directly regulate running-induced neurogenesis (Grégoire et al., 2014). Here, isolated mice with voluntary running had more Ki67+ and long-term BrdU+-retaining cells in the DG than isolated mice living with a locked running wheel, but not statistically different than socially enhanced conditions. This would suggest that exercise per se is not compensating neurogenesis by substituting the effect of isolation and poor environments.
3.4 Social behavior as a modulator of neurogenic changes
3.4.1 How social living conditions affect neurogenesis
Usually, inbred mice are housed to resemble social co-habitant conditions in nature, with some relevant exceptions with consequences in adult neurogenesis and social behavior (Figure 2). For instance, mice pups are weaned 21 days after birth, a period of time in which pups are very sensitive to the social environment. Single-parental conditions are the standard condition for breeding mice pups because pregnant mothers are separated from males and take care of the pups alone. However, bi-parental housing (mother and father or mother and another female) increases BrdU+ cells in the DG of male mice pups when they become adults in comparison with single-parental housing, and this effect improves context fear conditioning and is transmitted to the next generation (Mak et al., 2013). Curiously, offspring females with bi-parental housing did not change DG neurogenesis, but they increased oligodendrogenesis in the corpus callosum and their social exploration in the social chamber test, showing that female mice AHN is more resilient to early life social stressors than male mice AHN. Unfortunately, the neurogenic effect for progenitors was the opposite as parenting decreases BrdU+ cells in the DG in male and female parents compared to controls (Glasper et al., 2011).
One of the most emotional and cognitive impacting housing conditions for mice is social isolation (Figure 2), especially for male mice (Guarnieri et al., 2020) although no clear effect on hippocampus cell proliferation has been demonstrated per se (Monteiro et al., 2014; Guarnieri et al., 2020). Young males isolated for a short term could have a greater number of BrdU+ cells in the DG after only 4 days of isolation, but surprisingly this difference was not found in females, and it disappeared 20 days later (Ruscio et al., 2015). Long-term isolation studies found a moderate influence on neurogenesis through an increase in neuroblast and astrocyte differentiation (Du Preez et al., 2020; Du Preez et al., 2021) that can be reversed with physical exercise (Hueston et al., 2017). This could mean that exercise, in comparison with social isolation, did not change the number of BrdU+ cells in isolated adult mice but could buffer the neuronal and astrocyte differentiation effect. Prolonged social isolation resulted in more undifferentiated NSCs and fewer DCX+ cells in the DG than in standard grouped housing (Dranovsky et al., 2011), proposing a time-dependent influence of social isolation on the fate and lineage of NSCs.
Furthermore, when social isolation was applied in mice pups, more important changes occur in NSCs. Some authors report an increase in proliferation in the SEZ with more BrdU+ cells, more neurospheres in the primary culture, and more BrdU+ GFAP+ cells in the SEZ of maternal and socially deprived mice (Daun et al., 2020), which could raise not only neurogenesis but also NSC exhaustion. However, others found lower levels of BrdU in the ventral DG of maternal deprived mice (O’Leary et al., 2014) or fewer NeuN+ cells in CA3 (Reshetnikov et al., 2020), showing that both neurogenic niches respond differently to social isolation. Nevertheless, this is not necessarily a contradictory result as new neurons accomplish different functions in the two neurogenic niches. Functions like olfactory exploration and memory consolidation could be stimulated or deactivated simultaneously by social isolation.
There are myriad molecular mechanisms that affect NSCs and their transition from quiescent, primed-for-activation, and activated cells. For example, inflammatory signals like interleukin 17A serum levels in a murine model of posttraumatic stress disorder modulate DCX and Ki67+cells in the DG (Willinger and Turgeman 2022) or extracellular and adhesion molecules (reviewed in Morante-Redolat and Porlan, 2019) can promote alertness and activation of NSCs in response to indicators that affect the entire organism. Tumor necrosis factor-α (TNF-α), a pro-inflammatory cytokine, induces multiple effects in the nervous system through two main receptors: TNF-R1 and TNF-R2, which play very different functions in neurogenic niches (Belenguer et al., 2021). Removing the TNF receptor results in different social behaviors. While TNFR1−/− mice seemed perfectly normal in social exploration, recognition, and memory, TNFR2−/− mice showed lower social exploration and social memory (Camara et al., 2013).
Threats to the immune system are linked with transgenerational effects on social behavior. Social novelty exploration is diminished in adult mice when the mother had viral immune activation during pregnancy. Consequently, a recreation of this condition in mice is counterbalanced with vaccination (Wu et al., 2018). Other authors found that social isolation primed the immune system of isolated mice for infection allowing them to clear bacterial threats more effectively than grouped mice (Hamilton et al., 2022). Thus, housing conditions could be a key variable to understand fluctuations in NSC activity through immune system modification.
Similarly, stimulating social contact has been presented as a neurogenic trigger in APP/PS1 mice, an established model for Alzheimer’s disease with progressive spatial memory failure. Notably, these mice showed improved performance when they are co-housed with healthy young mice (Hsiao et al., 2014). This benefit in cognitive performance through the BDNF-TrkB signaling pathway was activated in aged APP/PS1 mice but we do not know if this partnership is beneficial for younger mice or if this effect is seemingly happening in physiological aging.
3.4.2 How social enrichment affects neurogenesis
Adult neurogenesis in rodents can be enhanced by transferring mice from standard laboratory housing to a more complex and stimulating environment (Kempermann et al., 1997; Kempermann et al., 1998). One main component of this environmental enrichment was social stimuli (Figure 2). Social enrichment can trigger hippocampal DCX division by itself in the same way as that in contact with objects and running wheels (Moreno-Jiménez et al., 2019). After tamoxifen neurogenesis depletion in the hippocampus, social environmental enrichment was able to generate the highest level of new neurons compared to standard or isolated housing (Dranovsky et al., 2011). Moreover, female mice in an enriched environment with male pheromones would need adult neurogenesis to develop mate preferences and equivalently, environmental pheromone cues induce olfactory and DG neurogenesis (Mak et al., 2007), indicating that socially rich environments promote neurogenesis (Sakalem et al., 2017). In the olfactory system, social behaviors relying on the activation of the VNS regulate adult neurogenesis in the mouse AOB. For example, male mice presented more BrdU+ cells when they were intruders in a resident cage and female mice had higher numbers of BrdU+ cells when they were in contact with male urine, although both sexes decreased new neurons in the AOB with aging (Nunez-Parra et al., 2011). Finally, social enrichment could be associated with sexual contact that activates neurogenesis and counterbalances the effect of chronic stress (Kim et al., 2013).
Likewise, social and environmental enrichment in young mice promotes increased social exploration and interaction with a novel mouse or object, despite this being not necessarily associated with an increase in AHN (Silva et al., 2011; Buschert et al., 2016). Curiously, when environmental enrichment is used to get a recovery from previous social stress conditions, the stimulation of adult neurogenesis is required (Schloesser et al., 2010). Therefore, social enrichment could be a therapeutic resource as long as adult neurogenesis can be stimulated.
Excessive social contact in crowded housing did not induce BrdU changes in the hippocampus at short or long intervals (Ago et al., 2014), and there are no immediate consequences on anxious behavior, highlighting high sociality in mice.
3.4.3 How social stress affects neurogenesis
The social role of a mouse in a group is based on a relatively stable hierarchy with acute and mild social pressure and aggression to keep the individuals in the social rank (Jennings et al., 1998). Consequently, any alteration in this dynamic system induces an extra dose of social stress. This stressor can be recreated with repeated social defeat or with an unstable social hierarchy. In repeated social defeat, the experimental mouse is exposed to recurrent social aggressions from a dominant mouse. This procedure induces avoidance to the aggressor, anxious behavior (Mouri et al., 2018), long-term disruption in AHN (Mitra et al., 2006; Chen et al., 2015; McKim et al., 2016), inflammatory local response (Ito et al., 2017; Shen et al., 2022), and decreased social interaction that can be recovered with antidepressants (Wu et al., 2017) and glucocorticoids (Lehmann et al., 2013).
One month after social defeat, adult male mice reduced the numbers of BrdU+ DCX+ cells (McKim et al., 2016) or their BrdU+ Neu-N+ cells (Chen et al., 2015; Jiang et al., 2017), suggesting that social stress affects neural progenitor cell differentiation (Figure 2). Again, higher differences can be observed when juvenile mice were used in which chronic social defeat was associated not only with lower neurogenesis but also with diminished Ki67+cells (Mouri et al., 2018). In contrast, some reliance to social stress induced by defeated experience has been described in adult and adolescent GFAP-TK heterozygous mice with reduced neurogenesis (Lehmann et al., 2013; Kirshenbaum et al., 2014). Curiously, DCX knock-out mice reduced their aggressiveness in the resident-intruder paradigm (Germain et al., 2013), and hypertrophic PtenLoxPNestin-CreERT2 mice have more Ki67 in the DG without affecting spatial hippocampal functioning but with deficits in social interaction (Amiri et al., 2012). Different results appeared when we consider mouse strain and total BrdU+ cells in the DG. C57BL/6 × 129/Sv mice showed lower social interaction after repeated social stress together with higher number of BrdU+ cells (McAllister et al., 2020). C57BL/6N mice showed lower social interaction after repeated social stress but without changes in BrdU+ cell numbers (Ishikawa et al., 2019), and OF1 mice showed more aggressive behavior after repeated social stress with fewer BrdU+ cells (Ferragud et al., 2010). Therefore, the mouse strain could affect social stress output, neurogenesis, or both.
An alternative protocol for social stress in the setting of social hierarchy is called social confrontation stress. This is a repeated aggressive experience to establish social dominance, and it classifies mice between “winners” (more offensive behavior) and “losers” (more defensive behavior). Interestingly, winners had a higher number of BrdU+ cells, BrdU+GFAP+ cells, and DCX+ cells and amplifying neural progenitor cells (Smagin et al., 2015) but only when this social dominance was established through explicit aggressive experience. Non-aggressive social dominance in the social tube test was not associated with changes in AHN (Pallé et al., 2019). Thus, developing explicit social dominance and not just social status enhances AHN.
Unstable housing conditions are another chronic social stress paradigm that increases BrdU+ cells in the DG (Buschert et al., 2013), but this is not associated with increased neurogenesis (Yohn et al., 2019). Actually, mice with extra copies of S100β are more sensitive to this housing, which has been linked to human major depression (Buschert et al., 2013). This implies that unstable housing could be a mild stressor in comparison to chronic social defeat that gets a bigger impact in neurogenic niches.
Finally, adult neurogenesis can be modulated by the social environment, and also new “social neurons” can transform our social behavior (Figure 2). Now, we would need to understand how we can use this reciprocal relationship for therapeutic purposes.
Author contributions
LG-G and IC-F conducted the pre-screening and classification of the articles. LG-G prepared the list of references. AP-V conducted conceptualization, screening of the articles, writing of the manuscript, article discussion, creation of the figures, search of funding, and correspondence.
Funding
This work was supported by CITSAM (2021-283-001) and the Catholic University of Valencia (UCV). Figures were created at BioRender.com.
Acknowledgments
The authors are grateful to Isabel Fariñas and her laboratory members and Cristina Gil-Sanz and her laboratory members for training, support, and inspiration. The authors thank Perez-Bermejo for methodological advice. Finally, the authors are indebted to all the cited authors for their work, and this review is a tribute to all of them.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2022.1011657/full#supplementary-material
SUPPLEMENTARY FIGURE S1 | Social behavior tests used in the articles of this review. (A) Three-chamber test (or social chamber test) in which mice are placed in three open compartment spaces and find the possibility to explore objects and/or other mice (Rein et al., 2020 for protocol) or (B) social interaction test, in an open-field arena mice, interact for few minutes. Social hierarchy and dominance are evaluated using the (C) social tube test (Fan et al., 2019 for protocol). Social stress can be observed with the resident-intruder paradigm and repeated social defeat (D). The resident acts as an aggressor, and the intruder shows defensive and avoiding behavior.
SUPPLEMENTARY TABLE S1 | References reviewed in this systematic review according to the PRISMA flow diagram. Title, authors, year of publication, and DOI are reported for every article.
Abbreviations
AHN, adult hippocampal neurogenesis; AOB, accessory olfactory bulb; BrdU, 5-bromo-2-deoxyuridine; DCX, doublecortin; DG, dentate gyrus; GFAP, glial fibrillary acidic protein; MOB, main olfactory bulb; NSCs, neural stem cells; SEZ, subependymal zone; SGZ, subgranular zone; VNS, vomeronasal system.
References
Ago, Y., Tanaka, T., Ota, Y., Kitamoto, M., Imoto, E., Takuma, K., et al. (2014). Social crowding in the night-time reduces an anxiety-like behavior and increases social interaction in adolescent mice. Behav. Brain Res. 270, 37–46. doi:10.1016/j.bbr.2014.04.047
Aimone, J. B., Deng, W., and Gage, F. H. (2011). Resolving new memories: A critical look at the dentate gyrus, adult neurogenesis, and pattern separation. Neuron 70 (4), 589–596. doi:10.1016/j.neuron.2011.05.010
Alachkar, A., Jiang, D., Harrison, M., Zhou, Y., Chen, G., and Mao, Y. (2013). An EJC factor RBM8a regulates anxiety behaviors. Curr. Mol. Med. 13 (6), 887–899. doi:10.2174/15665240113139990019
Alén, F., Gómez-Redondo, I., Rivera, P., Suárez, J., Ramos-Ibeas, P., Pericuesta, E., et al. (2019). Sex-Dimorphic behavioral alterations and altered neurogenesis in U12 intron splicing-defective Zrsr1 mutant mice. Int. J. Mol. Sci. 20 (14), E3543. doi:10.3390/ijms20143543
Alvarez-Buylla, A., Cebrian-Silla, A., Sorrells, S. F., Nascimento, M. A., Paredes, M. F., Garcia-Verdugo, J. M., et al. (2022). Comment on “Impact of neurodegenerative diseases on human adult hippocampal neurogenesis”. Science 376 (6590), eabn7083. doi:10.1126/science.abn7083
Amar, M., Pramod, A. B., Yu, N. K., Herrera, V. M., Qiu, L. R., Moran-Losada, P., et al. (2021). Autism-linked Cullin3 germline haploinsufficiency impacts cytoskeletal dynamics and cortical neurogenesis through RhoA signaling. Mol. Psychiatry 26 (7), 3586–3613. doi:10.1038/s41380-021-01052-x
Amiri, A., Cho, W., Zhou, J., Birnbaum, S. G., Sinton, C. M., McKay, R. M., et al. (2012). Pten deletion in adult hippocampal neural stem/progenitor cells causes cellular abnormalities and alters neurogenesis. J. Neurosci. 32 (17), 5880–5890. doi:10.1523/JNEUROSCI.5462-11.2012
Bagnall-Moreau, C., Huerta, P. T., Comoletti, D., La-Bella, A., Berlin, R., Zhao, C., et al. (2020). In utero exposure to endogenous maternal polyclonal anti-Caspr2 antibody leads to behavioral abnormalities resembling autism spectrum disorder in male mice. Sci. Rep. 10 (1), 14446. doi:10.1038/s41598-020-71201-9
Belenguer, G., Duart-Abadia, P., Jordán-Pla, A., Domingo-Muelas, A., Blasco-Chamarro, L., Ferrón, S. R., et al. (2021). Adult neural stem cells are alerted by systemic inflammation through TNF-α receptor signaling. Cell Stem Cell 28 (2), 285–299.e9. doi:10.1016/j.stem.2020.10.016
Buschert, J., Hohoff, C., Touma, C., Palme, R., Rothermundt, M., Arolt, V., et al. (2013). S100B overexpression increases behavioral and neural plasticity in response to the social environment during adolescence. J. Psychiatr. Res. 47 (11), 1791–1799. doi:10.1016/j.jpsychires.2013.08.001
Buschert, J., Sakalem, M. E., Saffari, R., Hohoff, C., Rothermundt, M., Arolt, V., et al. (2016). Prenatal immune activation in mice blocks the effects of environmental enrichment on exploratory behavior and microglia density. Prog. Neuropsychopharmacol. Biol. Psychiatry 67, 10–20. doi:10.1016/j.pnpbp.2016.01.005
Camara, M. L., Corrigan, F., Jaehne, E. J., Jawahar, M. C., Anscomb, H., Koerner, H., et al. (2013). TNF-α and its receptors modulate complex behaviours and neurotrophins in transgenic mice. Psychoneuroendocrinology 38 (12), 3102–3114. doi:10.1016/j.psyneuen.2013.09.010
Castilla-Ortega, E., Blanco, E., Serrano, A., Ladrón de Guevara-Miranda, D., Pedraz, M., Estivill-Torrús, G., et al. (2016). Pharmacological reduction of adult hippocampal neurogenesis modifies functional brain circuits in mice exposed to a cocaine conditioned place preference paradigm. Addict. Biol. 21 (3), 575–588. doi:10.1111/adb.12248
Chen, C. C., Huang, C. C., and Hsu, K. S. (2015). Chronic social stress affects synaptic maturation of newly generated neurons in the adult mouse dentate gyrus. Int. J. Neuropsychopharmacol. 19 (3), pyv097. doi:10.1093/ijnp/pyv097
Chiu, W. H., Depboylu, C., Hermanns, G., Maurer, L., Windolph, A., Oertel, W. H., et al. (2015). Long-term treatment with L-DOPA or pramipexole affects adult neurogenesis and corresponding non-motor behavior in a mouse model of Parkinson's disease. Neuropharmacology 95, 367–376. doi:10.1016/j.neuropharm.2015.03.020
Cope, E. C., Waters, R. C., Diethorn, E. J., Pagliai, K. A., Dias, C. G., Tsuda, M., et al. (2020). Adult-Born neurons in the Hippocampus are essential for social memory maintenance. eNeuro 7 (6), ENEURO.0182–20.2020. doi:10.1523/ENEURO.0182-20.2020
Daun, K. A., Fuchigami, T., Koyama, N., Maruta, N., Ikenaka, K., and Hitoshi, S. (2020). Early maternal and social deprivation expands neural stem cell population size and reduces Hippocampus/amygdala- dependent fear memory. Front. Neurosci. 14, 22. doi:10.3389/fnins.2020.00022
di Nuzzo, L., Orlando, R., Tognoli, C., Di Pietro, P., Bertini, G., Miele, J., et al. (2015). Antidepressant activity of fingolimod in mice. Pharmacol. Res. Perspect. 3 (3), e00135. doi:10.1002/prp2.135
Dranovsky, A., Picchini, A. M., Moadel, T., Sisti, A. C., Yamada, A., Kimura, S., et al. (2011). Experience dictates stem cell fate in the adult hippocampus. Neuron 70 (5), 908–923. doi:10.1016/j.neuron.2011.05.022
Du Preez, A., Law, T., Onorato, D., Lim, Y. M., Eiben, P., Musaelyan, K., et al. (2020). The type of stress matters: Repeated injection and permanent social isolation stress in male mice have a differential effect on anxiety- and depressive-like behaviours, and associated biological alterations. Transl. Psychiatry 10 (1), 325. doi:10.1038/s41398-020-01000-3
Du Preez, A., Onorato, D., Eiben, I., Musaelyan, K., Egeland, M., Zunszain, P. A., et al. (2021). Chronic stress followed by social isolation promotes depressive-like behaviour, alters microglial and astrocyte biology and reduces hippocampal neurogenesis in male mice. Brain Behav. Immun. 91, 24–47. doi:10.1016/j.bbi.2020.07.015
Fan, Z., Zhu, H., Zhou, T., Wang, S., Wu, Y., and Hu, H. (2019). Author Correction: Using the tube test to measure social hierarchy in mice. Nat. Protoc. 14 (8), 2595. doi:10.1038/s41596-019-0158-2
Feierstein, C. E., Lazarini, F., Wagner, S., Gabellec, M. M., de Chaumont, F., Olivo-Marin, J. C., et al. (2010). Disruption of adult neurogenesis in the olfactory bulb affects social interaction but not maternal behavior. Front. Behav. Neurosci. 4, 176. doi:10.3389/fnbeh.2010.00176
Ferragud, A., Haro, A., Sylvain, A., Velázquez-Sánchez, C., Hernández-Rabaza, V., and Canales, J. J. (2010). Enhanced habit-based learning and decreased neurogenesis in the adult hippocampus in a murine model of chronic social stress. Behav. Brain Res. 210 (1), 134–139. doi:10.1016/j.bbr.2010.02.013
Fu, J., Gao, J., Gong, L., Ma, Y., Xu, H., Gu, Z., et al. (2018). Silica nanoparticle exposure during the neonatal period impairs hippocampal precursor proliferation and social behavior later in life. Int. J. Nanomedicine 13, 3593–3608. doi:10.2147/IJN.S160828
Garrett, L., Becker, L., Rozman, J., Puk, O., Stoeger, T., Yildirim, A., et al. (2018). Fgf9 (Y162C) mutation alters information processing and social memory in mice. Mol. Neurobiol. 55 (6), 4580–4595. doi:10.1007/s12035-017-0659-3
Garrett, L., Zhang, J., Zimprich, A., Niedermeier, K. M., Fuchs, H., Gailus-Durner, V., et al. (2015). Conditional reduction of adult born doublecortin-positive neurons reversibly impairs selective behaviors. Front. Behav. Neurosci. 9, 302. doi:10.3389/fnbeh.2015.00302
Germain, J., Bruel-Jungerman, E., Grannec, G., Denis, C., Lepousez, G., Giros, B., et al. (2013). Doublecortin knockout mice show normal hippocampal- dependent memory despite CA3 lamination defects. PLoS One 8 (9), 74992. doi:10.1371/journal.pone.0074992
Glasper, E. R., Kozorovitskiy, Y., Pavlic, A., and Gould, E. (2011). Paternal experience suppresses adult neurogenesis without altering hippocampal function in Peromyscus californicus. J. Comp. Neurol. 519 (11), 2271–2281. doi:10.1002/cne.22628
Gobshtis, N., Tfilin, M., Wolfson, M., Fraifeld, V. E., and Turgeman, G. (2017). Transplantation of mesenchymal stem cells reverses behavioural deficits and impaired neurogenesis caused by prenatal exposure to valproic acid. Oncotarget 8 (11), 17443–17452. doi:10.18632/oncotarget.15245
Grégoire, C. A., Bonenfant, D., Le Nguyen, A., Aumont, A., and Fernandes, K. J. (2014). Untangling the influences of voluntary running, environmental complexity, social housing and stress on adult hippocampal neurogenesis. PLoS One 9 (1), 86237. doi:10.1371/journal.pone.0086237
Guarnieri, L. O., Pereira-Caixeta, A. R., Medeiros, D. C., Aquino, N. S. S., Szawka, R. E., Mendes, E., et al. (2020). Pro-neurogenic effect of fluoxetine in the olfactory bulb is concomitant to improvements in social memory and depressive-like behavior of socially isolated mice. Transl. Psychiatry 10 (1), 33. doi:10.1038/s41398-020-0701-5
Guha, S. K., Tillu, R., Sood, A., Patgaonkar, M., Nanavaty, I. N., Sengupta, A., et al. (2014). Single episode of mild murine malaria induces neuroinflammation, alters microglial profile, impairs adult neurogenesis, and causes deficits in social and anxiety-like behavior. Brain Behav. Immun. 42, 123–137. doi:10.1016/j.bbi.2014.06.009
Hamilton, A., Rizzo, R., Brod, S., Ono, M., Perretti, M., Cooper, D., et al. (2022). The immunomodulatory effects of social isolation in mice are linked to temperature control. Brain Behav. Immun. 102, 179–194. doi:10.1016/j.bbi.2022.02.022
Hitti, F. L., and Siegelbaum, S. A. (2014). The hippocampal CA2 region is essential for social memory. Nature 508 (7494), 88–92. doi:10.1038/nature13028
Hong, S., Yi, J. H., Lee, S., Park, C. H., Ryu, J. H., Shin, K. S., et al. (2019). Defective neurogenesis and schizophrenia-like behavior in PARP-1-deficient mice. Cell Death Dis. 10 (12), 943. doi:10.1038/s41419-019-2174-0
Hsiao, Y. H., Hung, H. C., Chen, S. H., and Gean, P. W. (2014). Social interaction rescues memory deficit in an animal model of Alzheimer's disease by increasing BDNF-dependent hippocampal neurogenesis. J. Neurosci. 34 (49), 16207–16219. doi:10.1523/JNEUROSCI.0747-14.2014
Hueston, C. M., Cryan, J. F., and Nolan, Y. M. (2017). Adolescent social isolation stress unmasks the combined effects of adolescent exercise and adult inflammation on hippocampal neurogenesis and behavior. Neuroscience 365, 226–236. doi:10.1016/j.neuroscience.2017.09.020
Ishikawa, R., Kim, R., Namba, T., Kohsaka, S., Uchino, S., and Kida, S. (2014). Time-dependent enhancement of hippocampus-dependent memory after treatment with memantine: Implications for enhanced hippocampal adult neurogenesis. Hippocampus 24 (7), 784–793. doi:10.1002/hipo.22270
Ishikawa, R., Uchida, C., Kitaoka, S., Furuyashiki, T., and Kida, S. (2019). Improvement of PTSD-like behavior by the forgetting effect of hippocampal neurogenesis enhancer memantine in a social defeat stress paradigm. Mol. Brain 12 (1), 68. doi:10.1186/s13041-019-0488-6
Ito, N., Hirose, E., Ishida, T., Hori, A., Nagai, T., Kobayashi, Y., et al. (2017). Kososan, a Kampo medicine, prevents a social avoidance behavior and attenuates neuroinflammation in socially defeated mice. J. Neuroinflammation 14 (1), 98. doi:10.1186/s12974-017-0876-8
Jenniches, I., Ternes, S., Albayram, O., Otte, D. M., Bach, K., Bindila, L., et al. (2016). Anxiety, stress, and fear response in mice with reduced endocannabinoid levels. Biol. Psychiatry 79 (10), 858–868. doi:10.1016/j.biopsych.2015.03.033
Jennings, M., Batchelor, G. R., Brain, P. F., Dick, A., Elliott, H., Francis, R. J., et al. (1998). Refining rodent husbandry: The mouse. Report of the rodent refinement working party. Lab. Anim. 32 (3), 233–259. doi:10.1258/002367798780559301
Jiang, B., Wang, Y. J., Wang, H., Song, L., Huang, C., Zhu, Q., et al. (2017). Antidepressant-like effects of fenofibrate in mice via the hippocampal brain-derived neurotrophic factor signalling pathway. Br. J. Pharmacol. 174 (2), 177–194. doi:10.1111/bph.13668
Jiang, N., Lv, J., Wang, H., Huang, H., Wang, Q., Lu, C., et al. (2020). Ginsenoside Rg1 ameliorates chronic social defeat stress-induced depressive-like behaviors and hippocampal neuroinflammation. Life Sci. 252, 117669. doi:10.1016/j.lfs.2020.117669
Kannangara, T. S., Webber, A., Gil-Mohapel, J., and Christie, B. R. (2009). Stress differentially regulates the effects of voluntary exercise on cell proliferation in the dentate gyrus of mice. Hippocampus 19 (10), 889–897. doi:10.1002/hipo.20514
Kempermann, G., Brandon, E. P., and Gage, F. H. (1998). Environmental stimulation of 129/SvJ mice causes increased cell proliferation and neurogenesis in the adult dentate gyrus. Curr. Biol. 8 (16), 939–942. doi:10.1016/s0960-9822(07)00377-6
Kempermann, G., Gage, F. H., Aigner, L., Song, H., Curtis, M. A., Thuret, S., et al. (2018). Human adult neurogenesis: Evidence and remaining questions. Cell Stem Cell 23 (1), 25–30. doi:10.1016/j.stem.2018.04.004
Kempermann, G., Jessberger, S., Steiner, B., and Kronenberg, G. (2004). Milestones of neuronal development in the adult hippocampus. Trends Neurosci. 27 (8), 447–452. doi:10.1016/j.tins.2004.05.013
Kempermann, G., Kuhn, H. G., and Gage, F. H. (1997). More hippocampal neurons in adult mice living in an enriched environment. Nature 386 (6624), 493–495. doi:10.1038/386493a0
Kim, J. I., Lee, J. W., Lee, Y. A., Lee, D. H., Han, N. S., Choi, Y. K., et al. (2013). Sexual activity counteracts the suppressive effects of chronic stress on adult hippocampal neurogenesis and recognition memory. Brain Res. 1538, 26–40. doi:10.1016/j.brainres.2013.09.007
Kim, J., Lim, J., Moon, S. H., Liu, K. H., and Choi, H. J. (2020). Toluene inhalation causes early anxiety and delayed depression with regulation of dopamine turnover, 5-HT(1A) receptor, and adult neurogenesis in mice. Biomol. Ther. 28 (3), 282–291. doi:10.4062/biomolther.2020.055
Kirshenbaum, G. S., Lieberman, S. R., Briner, T. J., Leonardo, E. D., and Dranovsky, A. (2014). Adolescent but not adult-born neurons are critical for susceptibility to chronic social defeat. Front. Behav. Neurosci. 8, 289. doi:10.3389/fnbeh.2014.00289
Kobayashi, M., Nakatani, T., Koda, T., Matsumoto, K., Ozaki, R., Mochida, N., et al. (2014). Absence of BRINP1 in mice causes increase of hippocampal neurogenesis and behavioral alterations relevant to human psychiatric disorders. Mol. Brain 7, 12. doi:10.1186/1756-6606-7-12
Kozareva, D. A., O'Leary, O. F., Cryan, J. F., and Nolan, Y. M. (2018). Deletion of TLX and social isolation impairs exercise-induced neurogenesis in the adolescent hippocampus. Hippocampus 28 (1), 3–11. doi:10.1002/hipo.22805
Lazarini, F., Mouthon, M. A., Gheusi, G., de Chaumont, F., Olivo-Marin, J. C., Lamarque, S., et al. (2009). Cellular and behavioral effects of cranial irradiation of the subventricular zone in adult mice. PLoS One 4 (9), e7017. doi:10.1371/journal.pone.0007017
Lee, P. C. (1994). Social structure and evolution', Behaviour and evolution. New York, NY, US: Cambridge University Press, 266–303.
Lehmann, M. L., Brachman, R. A., Martinowich, K., Schloesser, R. J., and Herkenham, M. (2013). Glucocorticoids orchestrate divergent effects on mood through adult neurogenesis. J. Neurosci. 33 (7), 2961–2972. doi:10.1523/JNEUROSCI.3878-12.2013
Li, H., Liu, Y., Gao, X., Liu, L., Amuti, S., Wu, D., et al. (2019). Neuroplastin 65 modulates anxiety- and depression-like behavior likely through adult hippocampal neurogenesis and central 5-HT activity. Febs J. 286 (17), 3401–3415. doi:10.1111/febs.14865
Lledó, P. M., and Valley, M. (2016). Adult olfactory bulb neurogenesis. Cold Spring Harb. Perspect. Biol. 8 (8), a018945. doi:10.1101/cshperspect.a018945
Lois, C., and Alvarez-Buylla, A. (1994). Long-distance neuronal migration in the adult mammalian brain. Science 264 (5162), 1145–1148. doi:10.1126/science.8178174
Lucassen, P. J., Fitzsimons, C. P., Salta, E., and Maletic-Savatic, M. (2020). Adult neurogenesis, human after all (again): Classic, optimized, and future approaches. Behav. Brain Res. 381, 112458. doi:10.1016/j.bbr.2019.112458
Lunardi, P., Mansk, L. M. Z., Jaimes, L. F., and Pereira, G. S. (2021). On the novel mechanisms for social memory and the emerging role of neurogenesis. Brain Res. Bull. 171, 56–66. doi:10.1016/j.brainresbull.2021.03.006
Maekawa, M., Namba, T., Suzuki, E., Yuasa, S., Kohsaka, S., and Uchino, S. (2009). NMDA receptor antagonist memantine promotes cell proliferation and production of mature granule neurons in the adult hippocampus. Neurosci. Res. 63 (4), 259–266. doi:10.1016/j.neures.2008.12.006
Mak, G. K., Antle, M. C., Dyck, R. H., and Weiss, S. (2013). Bi-parental care contributes to sexually dimorphic neural cell Genesis in the adult mammalian brain. PLoS One 8 (5), 62701. doi:10.1371/journal.pone.0062701
Mak, G. K., Enwere, E. K., Gregg, C., Pakarainen, T., Poutanen, M., Huhtaniemi, I., et al. (2007). Male pheromone-stimulated neurogenesis in the adult female brain: Possible role in mating behavior. Nat. Neurosci. 10 (8), 1003–1011. doi:10.1038/nn1928
McAllister, B. B., Pochakom, A., Fu, S., and Dyck, R. H. (2020). Effects of social defeat stress and fluoxetine treatment on neurogenesis and behavior in mice that lack zinc transporter 3 (ZnT3) and vesicular zinc. Hippocampus 30 (6), 623–637. doi:10.1002/hipo.23185
McKim, D. B., Niraula, A., Tarr, A. J., Wohleb, E. S., Sheridan, J. F., and Godbout, J. P. (2016). Neuroinflammatory dynamics underlie memory impairments after repeated social defeat. J. Neurosci. 36 (9), 2590–2604. doi:10.1523/JNEUROSCI.2394-15.2016
Mitra, R., Sundlass, K., Parker, K. J., Schatzberg, A. F., and Lyons, D. M. (2006). Social stress- related behavior affects hippocampal cell proliferation in mice. Physiol. Behav. 89 (2), 123–127. doi:10.1016/j.physbeh.2006.05.047
Mizumatsu, S., Monje, M. L., Morhardt, D. R., Rola, R., Palmer, T. D., and Fike, J. R. (2003). Extreme sensitivity of adult neurogenesis to low doses of X-irradiation. Cancer Res. 63 (14), 4021–4027.
Monteiro, B. M., Moreira, F. A., Massensini, A. R., Moraes, M. F., and Pereira, G. S. (2014). Enriched environment increases neurogenesis and improves social memory persistence in socially isolated adult mice. Hippocampus 24 (2), 239–248. doi:10.1002/hipo.22218
Morante-Redolat, J. M., and Porlan, E. (2019). Neural stem cell regulation by adhesion molecules within the subependymal niche. Front. Cell Dev. Biol. 7, 102. doi:10.3389/fcell.2019.00102
Moreno-Jiménez, E. P., Jurado-Arjona, J., Ávila, J., and Llorens-Martín, M. (2019). The social component of environmental enrichment is a pro-neurogenic stimulus in adult c57BL6 female mice. Front. Cell Dev. Biol. 7, 62. doi:10.3389/fcell.2019.00062
Mouri, A., Ukai, M., Uchida, M., Hasegawa, S., Taniguchi, M., Ito, T., et al. (2018). Juvenile social defeat stress exposure persistently impairs social behaviors and neurogenesis. Neuropharmacology 133, 23–37. doi:10.1016/j.neuropharm.2018.01.016
Nacer, S. A., Letsinger, A. C., Otto, S., DeFilipp, J. S., Nikolova, V. D., Riddick, N. V., et al. (2021). Loss of α7 nicotinic acetylcholine receptors in GABAergic neurons causes sex-dependent decreases in radial glia-like cell quantity and impairments in cognitive and social behavior. Brain Struct. Funct. 226 (2), 365–379. doi:10.1007/s00429-020-02179-3
Newton, J., Brown, T., Corley, C., Alexander, T., Trujillo, M., McElroy, T., et al. (2020). Cranial irradiation impairs juvenile social memory and modulates hippocampal physiology. Brain Res. 1748, 147095. doi:10.1016/j.brainres.2020.147095
Nunez-Parra, A., Pugh, V., and Araneda, R. C. (2011). Regulation of adult neurogenesis by behavior and age in the accessory olfactory bulb. Mol. Cell. Neurosci. 47 (4), 274–285. doi:10.1016/j.mcn.2011.05.003
O'Leary, O. F., Felice, D., Galimberti, S., Savignac, H. M., Bravo, J. A., Crowley, T., et al. (2014). GABAB (1) receptor subunit isoforms differentially regulate stress resilience. Proc. Natl. Acad. Sci. U. S. A. 111 (42), 15232–15237. doi:10.1073/pnas.1404090111
Osborn, M., Rustom, N., Clarke, M., Litteljohn, D., Rudyk, C., Anisman, H., et al. (2013). Antidepressant-like effects of erythropoietin: A focus on behavioural and hippocampal processes. PLoS One 8 (9), e72813. doi:10.1371/journal.pone.0072813
Page, M. J., McKenzie, J. E., Bossuyt, P. M., Boutron, I., Hoffmann, T. C., Mulrow, C. D., et al. (2021). The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Rev. Esp. Cardiol. 74 (9), 790–799. doi:10.1016/j.rec.2021.07.010
Pallé, A., Zorzo, C., Luskey, V. E., McGreevy, K. R., Fernández, S., and Trejo, J. L. (2019). Social dominance differentially alters gene expression in the medial prefrontal cortex without affecting adult hippocampal neurogenesis or stress and anxiety-like behavior. Faseb J. 33 (6), 6995–7008. doi:10.1096/fj.201801600R
Pereira-Caixeta, A. R., Guarnieri, L. O., Medeiros, D. C., Mendes, E., Ladeira, L. C. D., Pereira, M. T., et al. (2018). Inhibiting constitutive neurogenesis compromises long-term social recognition memory. Neurobiol. Learn. Mem. 155, 92–103. doi:10.1016/j.nlm.2018.06.014
Pereira-Caixeta, A. R., Guarnieri, L. O., Pena, R. R., Dias, T. L., and Pereira, G. S. (2017). Neurogenesis inhibition prevents enriched environment to prolong and strengthen social recognition memory, but not to increase BDNF expression. Mol. Neurobiol. 54 (5), 3309–3316. doi:10.1007/s12035-016-9922-2
Poletaeva, , Perepelkina, O. V., Boiarshinova, O. S., Lil'p, I. G., Markina, N. V., Timoshenko, T. V., et al. (2012). Neonatal injections of pharmacological agents and their remote genotype- dependent effects in mice and rats. Russ. J. Dev. Biol. 43 (6), 319–331. doi:10.1134/s1062360412060045
Rein, B., Ma, K., and Yan, Z. (2020). A standardized social preference protocol for measuring social deficits in mouse models of autism. Nat. Protoc. 15 (10), 3464–3477. doi:10.1038/s41596-020-0382-9
Reshetnikov, V. V., Kovner, A. V., Lepeshko, A. A., Pavlov, K. S., Grinkevich, L. N., and Bondar, N. P. (2020). Stress early in life leads to cognitive impairments, reduced numbers of CA3 neurons and altered maternal behavior in adult female mice. Genes Brain Behav. 19 (3), e12541. doi:10.1111/gbb.12541
Ruscio, M. G., Bradley King, S., and Haun, H. L. (2015). Social isolation increases cell proliferation in male and cell survival in female California mice (Peromyscus californicus). Physiol. Behav. 151, 570–576. doi:10.1016/j.physbeh.2015.08.039
Sakalem, M. E., Seidenbecher, T., Zhang, M., Saffari, R., Kravchenko, M., Wördemann, S., et al. (2017). Environmental enrichment and physical exercise revert behavioral and electrophysiological impairments caused by reduced adult neurogenesis. Hippocampus 27 (1), 36–51. doi:10.1002/hipo.22669
Salminen, A. V., Garrett, L., Schormair, B., Rozman, J., Giesert, F., Niedermeier, K. M., et al. (2017). Meis1: Effects on motor phenotypes and the sensorimotor system in mice. Dis. Model. Mech. 10 (8), 981–991. doi:10.1242/dmm.030080
Schloesser, R. J., Lehmann, M., Martinowich, K., Manji, H. K., and Herkenham, M. (2010). Environmental enrichment requires adult neurogenesis to facilitate the recovery from psychosocial stress. Mol. Psychiatry 15 (12), 1152–1163. doi:10.1038/mp.2010.34
Shen, S. Y., Yu, R., Li, W., Liang, L. F., Han, Q. Q., Huang, H. J., et al. (2022). The neuroprotective effects of GPR55 against hippocampal neuroinflammation and impaired adult neurogenesis in CSDS mice. Neurobiol. Dis. 169, 620 105743. doi:10.1016/j.nbd.2022.105743
Silva, C. F., Duarte, F. S., Lima, T. C., and de Oliveira, C. L. (2011). Effects of social isolation and enriched environment on behavior of adult Swiss mice do not require hippocampal neurogenesis. Behav. Brain Res. 225 (1), 85–90. doi:10.1016/j.bbr.2011.07.007
Smagin, D. A., Park, J. H., Michurina, T. V., Peunova, N., Glass, Z., Sayed, K., et al. (2015). Altered hippocampal neurogenesis and amygdalar neuronal activity in adult mice with repeated experience of aggression. Front. Neurosci. 9, 443. doi:10.3389/fnins.2015.00443
Subbanna, S., and Basavarajappa, B. S. (2014). Pre-administration of G9a/GLP inhibitor during synaptogenesis prevents postnatal ethanol-induced LTP deficits and neurobehavioral abnormalities in adult mice. Exp. Neurol. 261, 34–43. doi:10.1016/j.expneurol.2014.07.003
Terrillion, C. E., Abazyan, B., Yang, Z., Crawford, J., Shevelkin, A. V., Jouroukhin, Y., et al. (2017). DISC1 in astrocytes influences adult neurogenesis and hippocampus-dependent behaviors in mice. Neuropsychopharmacology 42 (11), 2242–2251. doi:10.1038/npp.2017.129
van Praag, H., Kempermann, G., and Gage, F. H. (1999). Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 2 (3), 266–270. doi:10.1038/6368
Velloso, F. J., Kumari, E., Buono, K. D., Frondelli, M. J., and Levison, S. W. (2022). Analyzing mouse neural stem cell and progenitor cell proliferation using EdU incorporation and multicolor flow cytometry. Star. Protoc. 3 (1), 101065. doi:10.1016/j.xpro.2021.101065
Wei, L., Meaney, M. J., Duman, R. S., and Kaffman, A. (2011). Affiliative behavior requires juvenile, but not adult neurogenesis. J. Neurosci. 31 (40), 14335–14345. doi:10.1523/JNEUROSCI.1333-11.2011
Willinger, Y., and Turgeman, G. (2022). Interleukine-17 modulates neurogenesis and behavior following exposure to trauma in mice. Cells 11 (3), 343. doi:10.3390/cells11030343
Wu, Q., Cai, H., Song, J., and Chang, Q. (2017). The effects of sEH inhibitor on depression-like behavior and neurogenesis in male mice. J. Neurosci. Res. 95 (12), 2483–2492. doi:10.1002/jnr.24080
Wu, Y., Qi, F., Song, D., He, Z., Zuo, Z., Yang, Y., et al. (2018). Prenatal influenza vaccination rescues impairments of social behavior and lamination in a mouse model of autism. J. Neuroinflammation 15 (1), 228. doi:10.1186/s12974-018-1252-z
Yohn, C. N., Ashamalla, S. A., Bokka, L., Gergues, M. M., Garino, A., and Samuels, B. A. (2019). Social instability is an effective chronic stress paradigm for both male and female mice. Neuropharmacology 160, 107780. doi:10.1016/j.neuropharm.2019.107780
Zhang, H., Kang, E., Wang, Y., Yang, C., Yu, H., Wang, Q., et al. (2016). Brain-specific Crmp2 deletion leads to neuronal development deficits and behavioural impairments in mice. Nat. Commun. 7, 11773. doi:10.1038/ncomms11773
Zhang, R., Cai, Y., Xiao, R., Zhong, H., Li, X., Guo, L., et al. (2019). Human amniotic epithelial cell transplantation promotes neurogenesis and ameliorates social deficits in BTBR mice. Stem Cell Res. Ther. 10 (1), 153. doi:10.1186/s13287-019-1267-0
Keywords: adult neurogenesis, hippocampus, social behavior, social memory, social stress, housing conditions, neural stem cells, parental behavior
Citation: García-Gómez L, Castillo-Fernández I and Perez-Villalba A (2022) In the pursuit of new social neurons. Neurogenesis and social behavior in mice: A systematic review. Front. Cell Dev. Biol. 10:1011657. doi: 10.3389/fcell.2022.1011657
Received: 04 August 2022; Accepted: 17 October 2022;
Published: 04 November 2022.
Edited by:
Carmen Castro, Universidad de Cádiz, SpainReviewed by:
Chaojuan Yang, Peking University, ChinaMiriam Corraliza-Gomez, University of Cádiz, Spain
Copyright © 2022 García-Gómez, Castillo-Fernández and Perez-Villalba. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Perez-Villalba, YW5hcGVyZXpAdWN2LmVz