 Marc Herb
Marc Herb Alexander Gluschko
Alexander Gluschko Michael Schramm
Michael Schramm- Institute for Medical Microbiology, Immunology and Hygiene, Cologne, Germany
Reactive oxygen species (ROS), such as the superoxide anion or hydrogen peroxide, have been established over decades of research as, on the one hand, important and versatile molecules involved in a plethora of homeostatic processes and, on the other hand, as inducers of damage, pathologies and diseases. Which effects ROS induce, strongly depends on the cell type and the source, amount, duration and location of ROS production. Similar to cellular pH and calcium levels, which are both strictly regulated and only altered by the cell when necessary, the redox balance of the cell is also tightly regulated, not only on the level of the whole cell but in every cellular compartment. However, a still widespread view present in the scientific community is that the location of ROS production is of no major importance and that ROS randomly diffuse from their cellular source of production throughout the whole cell and hit their redox-sensitive targets when passing by. Yet, evidence is growing that cells regulate ROS production and therefore their redox balance by strictly controlling ROS source activation as well as localization, amount and duration of ROS production. Hopefully, future studies in the field of redox biology will consider these factors and analyze cellular ROS more specifically in order to revise the view of ROS as freely flowing through the cell.
Reactive Oxygen Species: Handle With Care!
Reactive oxygen species (ROS) are molecules with higher reactivity than molecular oxygen (O2) (Fenton, 1894; Haber et al., 1997; D’Autreaux and Toledano, 2007; Hatz et al., 2007; Prousek, 2007; Das and Roychoudhury, 2014; Odobasic et al., 2016; Aratani, 2018; Juan et al., 2021) and include highly reactive free radicals, such as the superoxide anion (O2–) (Hayyan et al., 2016), and non-radical species, such as hydrogen peroxide (H2O2) (Chavez et al., 2009; Odobasic et al., 2016; Aratani, 2018).
A plethora of different ROS sources, such as NADPH oxidases (Nox) (Bedard and Krause, 2007; Nakano et al., 2008; Lambeth and Neish, 2014; Gluschko et al., 2018; Wolf et al., 2020), mitochondria (Afanas’ev et al., 1999; Han et al., 2003; Muller et al., 2004; Murphy, 2009; Brand, 2010; Shoshan-Barmatz et al., 2010; West et al., 2011a,b; Bezawork-Geleta et al., 2017; Zhao et al., 2019; Hadrava Vanova et al., 2020), xanthine oxidase (Nomura et al., 2014; Al-Shehri et al., 2020), peroxisomes (Lismont et al., 2015; Fransen et al., 2017; Shai et al., 2018) and cytochrome P450 oxidases (Omura and Sato, 1962; Zhang et al., 2020) can be responsible for cellular ROS production and it highly depends on the stimulus and the cell type whether a single or multiple ROS sources are activated, for how long this occurs and for what purpose (Banfi et al., 2004; Gavazzi et al., 2006; Bedard and Krause, 2007; Aguirre and Lambeth, 2010; Brand, 2010; Dikalova et al., 2010; Donko et al., 2010; Carnesecchi et al., 2011; Al-Mehdi et al., 2012; Lanciano et al., 2013; Kim et al., 2014; Lambeth and Neish, 2014; Ives et al., 2015; Xu et al., 2017; Herb et al., 2019b; Hernansanz-Agustin et al., 2020; Herb and Schramm, 2021).
O2– is the precursor of all cellular ROS (Chavez et al., 2009; Niethammer et al., 2009) and, under physiological pH, cannot diffuse over cell membranes due to its negative charge (Takahashi and Asada, 1983; Shoshan-Barmatz et al., 2010; Cordeiro, 2014). It has a half-life of a couple of seconds or even less (Marklund, 1976; Rapoport et al., 1994; D’Autreaux and Toledano, 2007; Taverne et al., 2013). Despite being not a strong oxidizing substance by itself, O2– readily oxidizes iron-sulfur structures of proteins (Liochev and Fridovich, 1999; Imlay, 2003), which can lead to protein malfunction and iron release from proteins. The released iron reacts with H2O2 to form the highly reactive and toxic OH radical (Liochev and Fridovich, 1999). Therefore, under healthy conditions cells keep O2– levels low (∼ 10–11-10–12 M) by compartmentalization and quick removal of O2–by superoxide dismutates (D’Autreaux and Toledano, 2007; Kehrer et al., 2010; Wang et al., 2018), which are expressed in all cellular compartments (Melov et al., 1999; Lambeth and Neish, 2014; Wang et al., 2018). A sustained increase in cellular O2– levels is associated with damage to cellular structures (Davies, 2016; Pizzino et al., 2017; Sies et al., 2017; Gutteridge and Halliwell, 2018; Su et al., 2019; Juan et al., 2021). However, O2– can also contribute to cellular signaling (Chen et al., 2009; Xu et al., 2017; Ren et al., 2021).
O2– quickly dismutates to H2O2, which is more, although not freely, diffusible for cellular membranes (Bienert et al., 2006; Wang et al., 2020; Chauvigne et al., 2021), which questions saturation of the cell with H2O2 to fulfill signaling functions in compartments, which are not in direct proximity to the ROS source (Beretta et al., 2020; Sies, 2021). Communication between cellular compartments can be achieved by aquaporins, which facilitate a controlled passage of H2O2 over membranes (Bienert and Chaumont, 2014; Wang et al., 2020). H2O2 has a longer cellular half-life (∼1 ms) with concentrations of ∼10–7 M under cellular homoeostatic conditions (D’Autreaux and Toledano, 2007). Because of these properties, it functions as an important signaling molecule involved in many different cellular processes (Kamata et al., 2005; Tonks, 2005; Rhee, 2006; Marinho et al., 2013; Holmstrom and Finkel, 2014; Romero et al., 2014; Jones et al., 2016; Short et al., 2016; Zhang et al., 2016; Herb et al., 2019b; Sies and Jones, 2020; Chauvigne et al., 2021). H2O2-mediated signaling is mainly based on the oxidation of cysteine residues of proteins (Chiarugi et al., 2001; Rhee, 2006; Herscovitch et al., 2008; Romero et al., 2014; Jones et al., 2016; Short et al., 2016; Herb et al., 2019b). These cysteine residues have a low pKa, are exposed to the cytosol and deprotonated to thiolate groups (Finkel, 2011; Poole, 2015). An increase to nanomolar concentrations (∼100 nM) of H2O2 is sufficient to induce reversible oxidation. This can lead to allosteric protein changes that alter the enzymatic function of the target proteins in many ways (Lee et al., 1998; Meng et al., 2002; Kamata et al., 2005; Tonks, 2005). ROS-mediated oxidation can also lead to covalent linkage of cysteine residues by disulfide bonds (Herscovitch et al., 2008; Zhou et al., 2014; Herb et al., 2019b). Since these H2O2-mediated protein oxidations can be reversed by the antioxidant defense system, they represent important redox switches involved in various cellular processes (Barford, 2004; Holmstrom and Finkel, 2014). Excessive H2O2 production, however, leads to further oxidation of the oxidized cysteines, which is an irreversible process and results in protein malfunction (Winterbourn and Hampton, 2008).
Growing evidence indicates that the redox status in different cellular compartments varies greatly (Fransen et al., 2017; Beretta et al., 2020; Chauvigne et al., 2021; Sies, 2021; Wang et al., 2021b), is tightly regulated (Nakamura, 2005; Kirkman and Gaetani, 2007; Kelley et al., 2010; Brigelius-Flohe and Maiorino, 2013; Poljsak et al., 2013; Couto et al., 2016; Jones et al., 2016; Chauvigne et al., 2021) and every elevation of ROS levels is controlled by the cell in various ways (Babior, 2002; Bulua et al., 2011; Hoeven et al., 2011; West et al., 2011a; Gluschko et al., 2018; Herb et al., 2019b; Chauvigne et al., 2021). The condition of ROS levels exceeding the capacity of cellular antioxidant defense systems is termed oxidative stress (Niki, 2016; Sies and Jones, 2020). Oxidative stress can be further divided into two subforms: (1) Oxidative distress, which represents excessive and prolonged oxidative stress, causes damage to cellular components and results in a number of different pathologies (Cross et al., 1987; Rangasamy et al., 2005; Thimmulappa et al., 2006). Notably, excessive oxidative distress is not always detrimental, if produced at the right place. Exceeded generation of oxidative stress in pathogen-containing phagosomes of phagocytes, for example, is an important factor of antimicrobial immunity (West et al., 2011a; Winterbourn and Kettle, 2013; Gluschko et al., 2018; Herb and Schramm, 2021). (2) Oxidative eustress represents a tightly controlled increase in cellular ROS levels (Niki, 2016; Sies and Jones, 2020), which are sufficient to fulfill important cellular processes, but do not induce critical damage to cellular structures (Tai et al., 2009; Finkel, 2011; Nathan and Cunningham-Bussel, 2013; Reczek and Chandel, 2014; Herb et al., 2019b).
Unfortunately, a lot of studies, which show the important role of ROS in various cellular processes, often suggest that ROS are produced in excess, saturate the cell and react randomly with redox-sensitive targets. This is mainly due to experimental setups that might lead to misinterpretation of the location of ROS production in cells.
Many studies use only one type of ROS probe, but do not provide an explanation for the choice, such as specificity for a cellular compartment or a defined type of ROS subspecies. Often probes are used that show neither a specificity for a ROS subspecies nor a defined cellular compartment, which leads to the frequently used terms “intracellular ROS” or “total cellular ROS,” which implicate that ROS once produced are equally distributed in the cell. Common examples for diffusible ROS probes are luminol (Caldefie-Chezet et al., 2002; Pavelkova and Kubala, 2004), 2′,7′-dichlordihydrofluorescein-diacetat (H2DCF-DA) (Ushijima et al., 1997; Hempel et al., 1999; Kim and Xue, 2020; Kim et al., 2021; Wang et al., 2021a) or dihydroethidium (DHE) (Gatliff et al., 2017; Wang and Zou, 2018; Zeller et al., 2021), which are regarded as compartment-specific but in fact they are not (Lundqvist and Dahlgren, 1996; Ushijima et al., 1997; Hempel et al., 1999; Wang and Zou, 2018). There are compartment-specific derivates available for these ROS probes, namely Isoluminol (Lundqvist and Dahlgren, 1996; Dahlgren and Karlsson, 1999; Caldefie-Chezet et al., 2002; Gluschko et al., 2018; Herb et al., 2019b; Wolf et al., 2020), 5-(and −6)-carboxy-2′,7′-dihydrochlorofluorescein-diacetat (5/6-Carboxy-DCF) (Hempel et al., 1999; Mak et al., 2017; Herb et al., 2019b; Wolf et al., 2020) and MitoSOX Red (MitoSOX) (Robinson et al., 2006; Mukhopadhyay et al., 2007) as alternatives, whose combined usage gives a much more confluent picture of the cellular ROS production. The preferable option for most precise ROS measurements concerning compatibility and specificity for ROS subspecies is represented by genetically modified cells, which express the ROS probe of choice in the cellular compartment of choice, like the HyPer family reporters and roGFP2-Orp1 (Belousov et al., 2006; Gutscher et al., 2009; Markvicheva et al., 2011; Bilan et al., 2013; Hernández-Barrera et al., 2013; Wang et al., 2021b). Another precise approach for compartment-specific ROS measurements is the coupling of ROS probes to cargo/particles, which can be engulfed by cells. This technique is especially useful in phagocytes like macrophages, to determine ROS levels in the phagosome (Geng et al., 2015; Ligeon et al., 2021a,b). For further reading on topics regarding ROS detection methods we want to point out to other reviews (Ermakova et al., 2014; Herb and Schramm, 2021).
Also the combined use of only globally working ROS scavengers in combination with ROS probes that detect total cellular ROS can lead to results, which suggest that ROS are present in the whole cell after diffusion from the location of their production. With NAC as most prominent globally working ROS scavenger (Patriarca et al., 2005; Aldini et al., 2018; Ezerina et al., 2018) only the general involvement of ROS in the cellular process of interest can be investigated, but no compartment-specific ROS production can be analyzed. More examples of globally working ROS scavengers are Tempol (4-Hydroxy-Tempo) (dismutation of O2– into H2O2) (Bernardy et al., 2017; Herb et al., 2019b), Tiron (a global O2– scavenger) (Krishna et al., 1992; Hein and Kuo, 1998; Manzano et al., 2000), Trolox (globally scavenges OOH and OOR) (Davies et al., 1988; Dugas et al., 2000) and ebselen (effectively removes H2O2 and ONOO–) (Nakamura et al., 2002; Matsushita et al., 2004; Mugesh, 2013). All of the scavengers mentioned above are diffusible (Davies et al., 1988; Krishna et al., 1992; Hein and Kuo, 1998; Haj-Yehia et al., 1999; Dugas et al., 2000; Manzano et al., 2000; Rak et al., 2000; Nakamura et al., 2002; Matsushita et al., 2004; Mugesh, 2013; Herb and Schramm, 2021). Assessment of specific removal of ROS subspecies and therefore their involvement in cellular processes is possible with these substances, but they cannot be used to identify the specific compartment in which the ROS exert their function.
Not only the location of ROS production, but also their various sources and their activation, regulation and termination is of major importance for the understanding of the complex redox maintenance in cells. For the identification of ROS sources it is not always possible to provide genetic evidence with a knock-out system or by siRNA usage. ROS source inhibitors are in these cases an option to block ROS production and analyze possible ROS sources. There are a lot of specific ROS source inhibitors commercially available and the choice is continuously expanded (Murphy, 2009; Wind et al., 2010; Altenhofer et al., 2015; Herb and Schramm, 2021).
For Nox enzymes, as one of the most prominent ROS sources in many cell types, the well-validated general Nox inhibitors VAS2870 (Leusen et al., 1995; ten Freyhaus et al., 2006; Wind et al., 2010; Altenhofer et al., 2012, 2015) or GKT 137831 (Laleu et al., 2010; Sedeek et al., 2010; Gaggini et al., 2011; Aoyama et al., 2012; Strengert et al., 2014; Kim et al., 2021) can be used. Both inhibitors show no intrinsic antioxidant activity and do not inhibit other flavoproteins (Wind et al., 2010; Altenhofer et al., 2012; Teixeira et al., 2017). Both also inhibit Nox-derived ROS production in vitro and in vivo (Carnesecchi et al., 2009, 2011; Aoyama et al., 2012; Green et al., 2012; Bettaieb et al., 2015; Gorin et al., 2015). In sharp contrast to VAS2870 and GKT 137831, the substances apocynin and DPI are still used and falsely addressed as specific Nox inhibitors in many otherwise convincing and excellent studies (Barbieri et al., 2003; Kiritoshi et al., 2003; Dostert et al., 2008; Choi et al., 2011; Abuaita et al., 2015; Gatliff et al., 2017; Alonso et al., 2019; Fan et al., 2019; Damiano et al., 2020; Geng et al., 2020; Inomata et al., 2020; Prestes et al., 2020; Ahmad et al., 2021; Ligeon et al., 2021b; Martinez et al., 2021 #1039; Troia et al., 2021). Several studies have shown that apocynin directly scavenges ROS due to its antioxidant capacities (Aldieri et al., 2008; Heumuller et al., 2008; Mora-Pale et al., 2009; Wingler et al., 2011; Trevelin et al., 2016), while DPI inhibits flavoproteins in general (O’Donnell et al., 1993; Wind et al., 2010; Altenhofer et al., 2015) including Nox2 (Reis et al., 2020), but also various other targets, such as complex I of the mitochondrial electron transport chain (Bloxham, 1979; Lambeth et al., 2008; Bulua et al., 2011), iNOS (Stuehr et al., 1991; Geyer et al., 1997) or xanthine oxidase (O’Donnell et al., 1993; Wind et al., 2010) as well as calcium transporters (Tazzeo et al., 2009). Since genetic knock-out models with the CRISPR-Cas9 technology (Ledford, 2015; Anzalone et al., 2020; Carlson-Stevermer et al., 2020), either for cell lines, ex vivo cells or mice, as well as knock-down via siRNA (Han, 2018) are readily available tools for analyzing possible roles of Nox enzymes in cellular processes, the use of apocynin or DPI, especially in combination with diffusible ROS probes, should not be recommended, since it may lead to false interpretations of results regarding Nox enzyme involvement and the location of ROS production.
In mitochondria, the complexes of the ETC not only are essential for energy generation of the cell, but are also ROS production sites (Nohl et al., 2003; Lambeth and Neish, 2014). Inhibition of the complexes for analysis of ROS production might also result in energy deprivation and the energy status of the cell has to be checked every time these inhibitors are used. Typically used inhibitors are rotenone (Stowe and Camara, 2009; Heinz et al., 2017; Scialo et al., 2017), which inhibits complex I and increases ROS production inside the mitochondrial matrix (St-Pierre et al., 2002; Lambert and Brand, 2004; Panov et al., 2005; Stowe and Camara, 2009; Sena et al., 2013) and antimycin A (Murphy, 2009; Bleier and Drose, 2013), which inhibits complex III and increases ROS production into the intermembrane space (IMS) (Chen et al., 2003; Han et al., 2003; Al-Mehdi et al., 2012; Quinlan et al., 2012; Herb et al., 2019a). The most commonly used ROS probe for detection of mitochondrial ROS is MitoSOX, which measures O2– exclusively inside the mitochondrial matrix (Robinson et al., 2006; Mukhopadhyay et al., 2007; Ernst et al., 2021). However, since the ETC complexes show compartment-specific differences concerning ROS production (Fridovich, 1997; Murphy, 2009; Brand, 2010; West et al., 2011b; Herb and Schramm, 2021), this probe can only be used to measure ROS production inside mitochondria and, therefore, other cellular compartments should always be analyzed in addition. In healthy, undamaged mitochondria, ROS cannot escape the mitochondrial matrix because of the very effective antioxidative defense system (Roca and Ramakrishnan, 2013; Briston et al., 2017; Hos et al., 2017; Hernansanz-Agustin et al., 2020; Lin et al., 2020; Wang et al., 2020; Zhao et al., 2020). Only after prolonged overproduction or when the structure of the mitochondrial membranes is ruptured, either by opening of the mitochondrial permeability transition pore or direct damage, e.g., by pathogenic toxins, ROS can escape from the matrix into the cytosol (Koterski et al., 2005; Stavru et al., 2011; Roca and Ramakrishnan, 2013; Briston et al., 2017; Hos et al., 2017; Zhang Y. et al., 2019; Zhao et al., 2020). Nevertheless, the general term “mitochondrial ROS” is used in many studies, which often is synonymous for matrix-located mitochondrial ROS production measured by MitoSOX. ROS measurements in other cellular compartments as well as an explanation if and how the mitochondrial ROS escape from the matrix and fulfill their role in the cell, with a few exceptions (Koterski et al., 2005; Zhou et al., 2011; Roca and Ramakrishnan, 2013; Briston et al., 2017; Hos et al., 2017; Herb et al., 2019b; Zhao et al., 2020), are often not provided. Additionally, the usage of inhibitors of the ETC, like rotenone or antimycin A, which have compartment-specific effects on ROS production in combination with diffusible ROS probes can also lead to misinterpretations of the performed ROS measurements. For further reading concerning ROS scavengers and inhibitors, we like to point to other reviews (Wind et al., 2010; Altenhofer et al., 2015; Herb and Schramm, 2021).
ROS Production: The Dose Makes the Poison
A model that involves an uncontrolled increase in total cellular ROS levels implies that cells take into account the collateral damage that ROS can inflict while enroute to their redox-sensitive target, that can be at a completely different cellular location (Bulua et al., 2011; Nazarewicz et al., 2013; Kelly et al., 2015; Garaude et al., 2016; Kim et al., 2017; Figure 1). But oxidative distress (Buczynski et al., 2013; Li et al., 2013; Bhattacharyya et al., 2014; Niki, 2016; Sweeney and McAuley, 2016; Yang et al., 2016; Sies and Jones, 2020) is a situation for the healthy cell that has to be avoided. Tightly controlled production of ROS in direct vicinity of a redox-sensitive target (Tai et al., 2009; Finkel, 2011; Nathan and Cunningham-Bussel, 2013; Reczek and Chandel, 2014; Herb et al., 2019b) requires much less ROS production and hence results in much less collateral damage, while fulfilling important cellular functions, i.e., oxidative eustress (Niki, 2016; Sies and Jones, 2020; Figure 2).
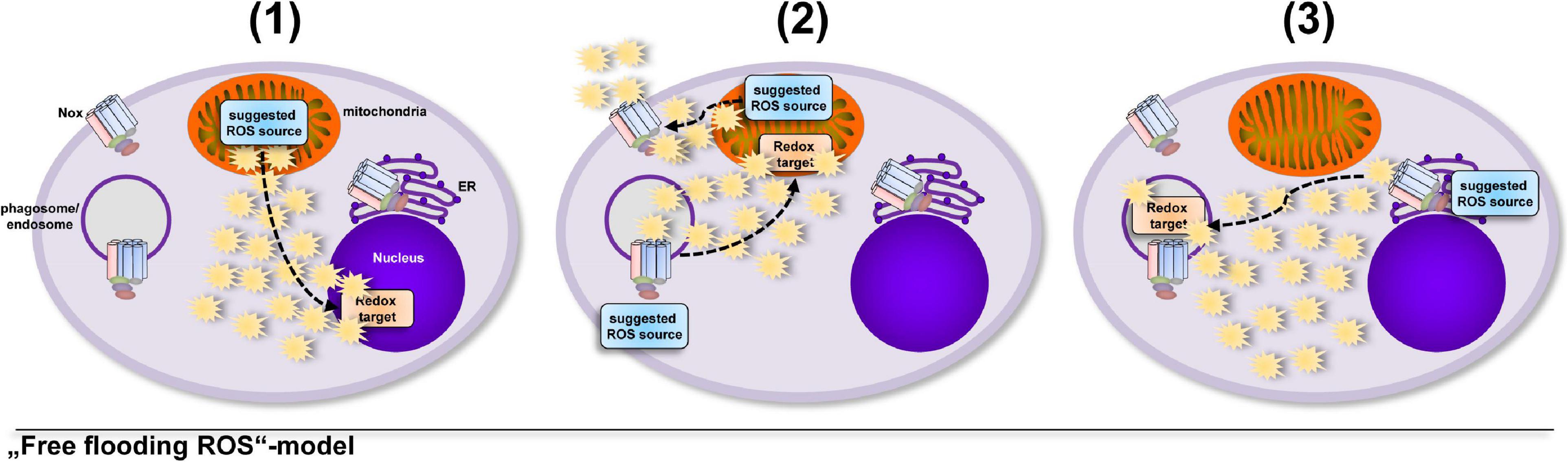
Figure 1. Several studies suggested that ROS are produced in excess, saturate the cell and find their redox-sensitive targets at random. Usage of diffusible ROS probes, globally working ROS scavengers and unspecific inhibitors often place the suggested ROS source at a completely different location than the redox-sensitive target, which might lead to the interpretation that cells “take into account” the damage that ROS can inflict on their way to the target molecule. The importance of ROS in general for various cellular processes was shown by many excellent studies (Bulua et al., 2011; Nazarewicz et al., 2013; Kelly et al., 2015; Garaude et al., 2016; Kim et al., 2017), however, diffusible ROS probes or only one ROS probe are often used to determine ROS production in cells, which might lead to the suggestions, e.g., that (1) ROS escape from the mitochondrial matrix and regulate expression and secretion of cytokines (Bulua et al., 2011; Kelly et al., 2015), (2) extracellular Nox2-derived ROS modify enzyme activity in the mitochondrial matrix or matrix-derived ROS modulate Nox2 activity (Nazarewicz et al., 2013; Garaude et al., 2016) or (3) ROS produced by ER-located Nox4 reach the phagosome for inactivation of phagocytosed parasites (Kim et al., 2017).
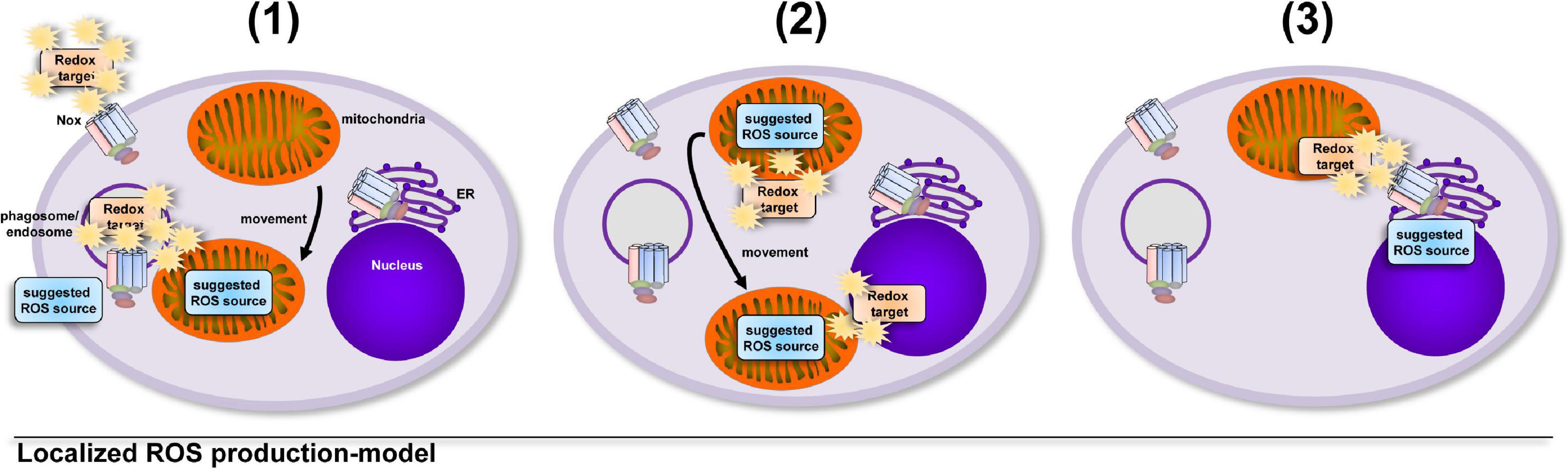
Figure 2. Growing evidence supports the hypothesis that cellular compartments show big differences and tight regulation of their redox status. The induction of ROS production is controlled by the cell in terms of location, source, duration and amount. The localized and timely controlled ROS production in the direct vicinity of the redox-sensitive target reduces the induced damage to cellular components and results in beneficial consequences for the cell, a condition termed as oxidative eustress (Sies, 2021). Examples of localized ROS production are (1) the production of antimicrobial ROS by Nox2 (Craig and Slauch, 2009; Gluschko et al., 2018) or mitochondria, which are recruited to pathogen-containing phagosomes (West et al., 2011a; Geng et al., 2015), (2) the recruitment of the redox-regulated target to ROS-producing mitochondria for NLRP3 inflammasome activation (Zhou et al., 2011) or the relocation of mitochondria to the nucleus for ROS-mediated nuclear signaling (Al-Mehdi et al., 2012) and (3) ROS production by ER-localized Nox4 during formation of mitochondria-associated membranes for regulation of calcium signaling (Beretta et al., 2020).
Of note, to reach levels at which the ROS can fulfill their important cellular roles, ROS production has to overcome the highly effective antioxidative defense systems of one or more cellular compartments (Nakamura, 2005; Kirkman and Gaetani, 2007; Kelley et al., 2010; Brigelius-Flohe and Maiorino, 2013; Poljsak et al., 2013; Lismont et al., 2015; Couto et al., 2016; Jones et al., 2016; Shai et al., 2018; Chauvigne et al., 2021; Herb and Schramm, 2021). Also H2O2-detoxifying enzymes, such as glutathione peroxidases or catalase, which can quickly decrease cellular H2O2 concentrations (Winterbourn and Hampton, 2008) as well as the inactivation of these scavenger enzymes by H2O2 itself broaden the regulatory potential of cells for selective and localized ROS signaling (Wood et al., 2003; Woo et al., 2010). This further places the model of ROS molecules, which “flood” the whole cell from a single location of ROS production in rather unrealistic light. Nevertheless, the model of ROS as “flooding” the cell is still popular in the scientific community, often referred to as “total cellular ROS levels” (Wang et al., 2013, 2019; Dinakar et al., 2016; Kim and Xue, 2020; Loth et al., 2020; Thorne et al., 2021), “intracellular ROS levels” (Tepel et al., 2000; Bensaad et al., 2009; Choi et al., 2011; Lee et al., 2017; Wang et al., 2019, 2021a; Wei et al., 2019; Zaidieh et al., 2019; Zhang W. et al., 2019; Mendiola et al., 2020; Winitchaikul et al., 2021; Zhong et al., 2021) or simply “ROS levels” (Chen et al., 2012, 2021; Wei et al., 2019; Agarwal and Ganesh, 2020; Kim et al., 2021; Knight et al., 2021; Zeller et al., 2021) in many studies, mainly because of the usage of diffusible ROS probes, which suggest free diffusion of ROS through the cell without regard of the location of ROS production. Mutations, e.g., in cancer cells (Schumacker, 2006; Liou and Storz, 2010; Reczek and Chandel, 2017; Zaidieh et al., 2019; Perillo et al., 2020), pathogenic invasion (West et al., 2011a; Abuaita et al., 2018; Gluschko et al., 2018; Roca et al., 2019) or metabolic disbalance (Li et al., 2016; Mak et al., 2017; Peng et al., 2021) are prominent examples, in which the ROS production of the cell can enter an uncontrolled stage and quickly overcome the antioxidative defense system leading to rapidly increased ROS levels in nearly every compartment of the cell with often detrimental consequences. In this context, cells can be regarded as “overflowing with ROS,” however, in healthy cells a redox balance between all producing and eliminating ROS sources, mediated by the antioxidant defense system is crucial for cellular functioning (Nathan and Cunningham-Bussel, 2013; Deshmukh et al., 2017; Suzuki et al., 2019; Wei et al., 2019; Baird and Yamamoto, 2020; Saito and Kimura, 2021). Importantly, in the extracellular milieu H2O2 can travel much further than inside the cell and fulfills important signaling (Levine et al., 1994; Sharma et al., 2012; Das and Roychoudhury, 2014; Hervera et al., 2018; Huang et al., 2019; Janku et al., 2019; Deng et al., 2020) and chemotactic functions (Niethammer et al., 2009; Rieger and Sagasti, 2011).
Concluding Remarks
In recent years, more and more studies supported the model–and highlighted the importance–of localized cellular ROS production in direct vicinity of the redox target (Meinhard and Grill, 2001; Veal et al., 2007; Go and Jones, 2008; Craig and Slauch, 2009; West et al., 2011a; Wink et al., 2011; Zhou et al., 2011; Al-Mehdi et al., 2012; Naviaux, 2012; Allan et al., 2014; Romero et al., 2014; Geng et al., 2015; To et al., 2017; Gluschko et al., 2018; Herb et al., 2019b; Acin-Perez et al., 2020; Beretta et al., 2020; Chanin et al., 2020; Miller et al., 2020; Sies and Jones, 2020; Herb and Schramm, 2021; Ligeon et al., 2021b; Sies, 2021; Wong et al., 2021). Therefore, the model of ROS molecules as “omnipresent and freely diffusing throughout the cell” should always be interpreted carefully in the context of research and highly depends on the proper use of ROS probes, scavengers and inhibitors. In healthy cells, ROS should be considered as molecules, whose production is tightly controlled in terms of stimulus, source, location, duration and amount.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
MH and MS: conceptualization. MH: writing–original draft preparation and visualization. MH, AG, and MS: writing–review and editing and funding acquisition. MS: supervision and project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Deutsche Forschungsgemeins- chaft (DFG) grant SCHR 1627/2-1 to MS, by the Köln Fortune grant 278/2019 to AG, and by the Köln Fortune grant 302/2020 to MH.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abuaita, B. H., Burkholder, K. M., Boles, B. R., and O’Riordan, M. X. (2015). The endoplasmic reticulum stress sensor inositol-requiring enzyme 1alpha augments bacterial killing through sustained oxidant production. mBio 6:e00705.
Abuaita, B. H., Schultz, T. L., and O’Riordan, M. X. (2018). Mitochondria-derived vesicles deliver antimicrobial reactive oxygen species to control phagosome-localized Staphylococcus aureus. Cell Host Microbe 24, 625–636.e5.
Acin-Perez, R., Iborra, S., Marti-Mateos, Y., Cook, E. C. L., Conde-Garrosa, R., Petcherski, A., et al. (2020). Fgr kinase is required for proinflammatory macrophage activation during diet-induced obesity. Nat. Metab. 2, 974–988. doi: 10.1038/s42255-020-00273-8
Afanas’ev, I. B., Ostrachovitch, E. A., and Korkina, L. G. (1999). Lucigenin is a mediator of cytochrome C reduction but not of superoxide production. Arch. Biochem. Biophys. 366, 267–274. doi: 10.1006/abbi.1999.1215
Agarwal, S., and Ganesh, S. (2020). Perinuclear mitochondrial clustering, increased ROS levels, and HIF1 are required for the activation of HSF1 by heat stress. J. Cell Sci. 133:jcs245589.
Aguirre, J., and Lambeth, J. D. (2010). Nox enzymes from fungus to fly to fish and what they tell us about Nox function in mammals. Free Radic. Biol. Med. 49, 1342–1353. doi: 10.1016/j.freeradbiomed.2010.07.027
Ahmad, A., Nawaz, M. I., Siddiquei, M. M., and Abu El-Asrar, A. M. (2021). Apocynin ameliorates NADPH oxidase 4 (NOX4) induced oxidative damage in the hypoxic human retinal Muller cells and diabetic rat retina. Mol. Cell. Biochem. 476, 2099–2109. doi: 10.1007/s11010-021-04071-y
Aldieri, E., Riganti, C., Polimeni, M., Gazzano, E., Lussiana, C., Campia, I., et al. (2008). Classical inhibitors of NOX NAD(P)H oxidases are not specific. Curr. Drug Metab. 9, 686–696. doi: 10.2174/138920008786049285
Aldini, G., Altomare, A., Baron, G., Vistoli, G., Carini, M., Borsani, L., et al. (2018). N-Acetylcysteine as an antioxidant and disulphide breaking agent: the reasons why. Free Radic. Res. 52, 751–762. doi: 10.1080/10715762.2018.1468564
Allan, E. R., Tailor, P., Balce, D. R., Pirzadeh, P., Mckenna, N. T., Renaux, B., et al. (2014). NADPH oxidase modifies patterns of MHC class II-restricted epitopic repertoires through redox control of antigen processing. J. Immunol. 192, 4989–5001. doi: 10.4049/jimmunol.1302896
Al-Mehdi, A. B., Pastukh, V. M., Swiger, B. M., Reed, D. J., Patel, M. R., Bardwell, G. C., et al. (2012). Perinuclear mitochondrial clustering creates an oxidant-rich nuclear domain required for hypoxia-induced transcription. Sci. Signal. 5:ra47. doi: 10.1126/scisignal.2002712
Alonso, D., Serrano, E., Bermejo, F. J., and Corral, R. S. (2019). HIF-1alpha-regulated MIF activation and Nox2-dependent ROS generation promote Leishmania amazonensis killing by macrophages under hypoxia. Cell. Immunol. 335, 15–21. doi: 10.1016/j.cellimm.2018.10.007
Al-Shehri, S. S., Duley, J. A., and Bansal, N. (2020). Xanthine oxidase-lactoperoxidase system and innate immunity: biochemical actions and physiological roles. Redox Biol. 34:101524. doi: 10.1016/j.redox.2020.101524
Altenhofer, S., Kleikers, P. W., Radermacher, K. A., Scheurer, P., Rob Hermans, J. J., Schiffers, P., et al. (2012). The NOX toolbox: validating the role of NADPH oxidases in physiology and disease. Cell. Mol. Life Sci. 69, 2327–2343. doi: 10.1007/s00018-012-1010-9
Altenhofer, S., Radermacher, K. A., Kleikers, P. W., Wingler, K., and Schmidt, H. H. (2015). Evolution of NADPH oxidase inhibitors: selectivity and mechanisms for target engagement. Antioxid. Redox Signal. 23, 406–427. doi: 10.1089/ars.2013.5814
Anzalone, A. V., Koblan, L. W., and Liu, D. R. (2020). Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 38, 824–844. doi: 10.1038/s41587-020-0561-9
Aoyama, T., Paik, Y. H., Watanabe, S., Laleu, B., Gaggini, F., Fioraso-Cartier, L., et al. (2012). Nicotinamide adenine dinucleotide phosphate oxidase in experimental liver fibrosis: GKT137831 as a novel potential therapeutic agent. Hepatology 56, 2316–2327. doi: 10.1002/hep.25938
Aratani, Y. (2018). Myeloperoxidase: its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 640, 47–52. doi: 10.1016/j.abb.2018.01.004
Babior, B. M. (2002). The activity of leukocyte NADPH oxidase: regulation by p47PHOX cysteine and serine residues. Antioxid. Redox Signal. 4, 35–38. doi: 10.1089/152308602753625834
Baird, L., and Yamamoto, M. (2020). The molecular mechanisms regulating the KEAP1-NRF2 Pathway. Mol. Cell. Biol. 40:e00099-20.
Banfi, B., Malgrange, B., Knisz, J., Steger, K., Dubois-Dauphin, M., and Krause, K. H. (2004). NOX3, a superoxide-generating NADPH oxidase of the inner ear. J. Biol. Chem. 279, 46065–46072. doi: 10.1074/jbc.m403046200
Barbieri, S. S., Eligini, S., Brambilla, M., Tremoli, E., and Colli, S. (2003). Reactive oxygen species mediate cyclooxygenase-2 induction during monocyte to macrophage differentiation: critical role of NADPH oxidase. Cardiovasc. Res. 60, 187–197. doi: 10.1016/s0008-6363(03)00365-1
Barford, D. (2004). The role of cysteine residues as redox-sensitive regulatory switches. Curr. Opin. Struct. Biol. 14, 679–686. doi: 10.1016/j.sbi.2004.09.012
Bedard, K., and Krause, K. H. (2007). The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87, 245–313. doi: 10.1152/physrev.00044.2005
Belousov, V. V., Fradkov, A. F., Lukyanov, K. A., Staroverov, D. B., Shakhbazov, K. S., Terskikh, A. V., et al. (2006). Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nat. Methods 3, 281–286. doi: 10.1038/nmeth866
Bensaad, K., Cheung, E. C., and Vousden, K. H. (2009). Modulation of intracellular ROS levels by TIGAR controls autophagy. EMBO J. 28, 3015–3026. doi: 10.1038/emboj.2009.242
Beretta, M., Santos, C. X., Molenaar, C., Hafstad, A. D., Miller, C. C., Revazian, A., et al. (2020). Nox4 regulates InsP3 receptor-dependent Ca(2+) release into mitochondria to promote cell survival. EMBO J. 39:e103530.
Bernardy, C. C. F., Zarpelon, A. C., Pinho-Ribeiro, F. A., Calixto-Campos, C., Carvalho, T. T., Fattori, V., et al. (2017). Tempol, a superoxide dismutase mimetic agent, inhibits superoxide anion-induced inflammatory pain in mice. Biomed Res. Int. 2017:9584819.
Bettaieb, A., Jiang, J. X., Sasaki, Y., Chao, T. I., Kiss, Z., Chen, X., et al. (2015). Hepatocyte Nicotinamide Adenine Dinucleotide Phosphate Reduced Oxidase 4 regulates stress signaling, fibrosis, and insulin sensitivity during development of steatohepatitis in mice. Gastroenterology 149, 468–480.e10.
Bezawork-Geleta, A., Rohlena, J., Dong, L., Pacak, K., and Neuzil, J. (2017). Mitochondrial complex II: at the crossroads. Trends Biochem. Sci. 42, 312–325. doi: 10.1016/j.tibs.2017.01.003
Bhattacharyya, S., Sinha, K., and Sil, P. C. (2014). Cytochrome P450s: mechanisms and biological implications in drug metabolism and its interaction with oxidative stress. Curr. Drug 15, 719–742. doi: 10.2174/1389200215666141125121659
Bienert, G. P., and Chaumont, F. (2014). Aquaporin-facilitated transmembrane diffusion of hydrogen peroxide. Biochim. Biophys. Acta 1840, 1596–1604. doi: 10.1016/j.bbagen.2013.09.017
Bienert, G. P., Schjoerring, J. K., and Jahn, T. P. (2006). Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta 1758, 994–1003.
Bilan, D. S., Pase, L., Joosen, L., Gorokhovatsky, A. Y., Ermakova, Y. G., Gadella, T. W., et al. (2013). HyPer-3: a genetically encoded H(2)O(2) probe with improved performance for ratiometric and fluorescence lifetime imaging. ACS Chem. Biol. 8, 535–542. doi: 10.1021/cb300625g
Bleier, L., and Drose, S. (2013). Superoxide generation by complex III: from mechanistic rationales to functional consequences. Biochim. Biophys. Acta 1827, 1320–1331. doi: 10.1016/j.bbabio.2012.12.002
Bloxham, D. P. (1979). The relationship of diphenyleneiodonium-induced hypoglycaemia to the specific covalent modification of NADH-ubiquinone oxidoreductase. Biochem. Soc. Trans. 7, 103–106. doi: 10.1042/bst0070103
Brand, M. D. (2010). The sites and topology of mitochondrial superoxide production. Exp. Gerontol. 45, 466–472. doi: 10.1016/j.exger.2010.01.003
Brigelius-Flohe, R., and Maiorino, M. (2013). Glutathione peroxidases. Biochim. Biophys. Acta 1830, 3289–3303.
Briston, T., Roberts, M., Lewis, S., Powney, B., Staddon, J. M., Szabadkai, G., et al. (2017). Mitochondrial permeability transition pore: sensitivity to opening and mechanistic dependence on substrate availability. Sci. Rep. 7:10492.
Buczynski, B. W., Maduekwe, E. T., and O’reilly, M. A. (2013). The role of hyperoxia in the pathogenesis of experimental BPD. Semin. Perinatol. 37, 69–78. doi: 10.1053/j.semperi.2013.01.002
Bulua, A. C., Simon, A., Maddipati, R., Pelletier, M., Park, H., Kim, K. Y., et al. (2011). Mitochondrial reactive oxygen species promote production of proinflammatory cytokines and are elevated in TNFR1-associated periodic syndrome (TRAPS). J. Exp. Med. 208, 519–533. doi: 10.1084/jem.20102049
Caldefie-Chezet, F., Walrand, S., Moinard, C., Tridon, A., Chassagne, J., and Vasson, M. P. (2002). Is the neutrophil reactive oxygen species production measured by luminol and lucigenin chemiluminescence intra or extracellular? Comparison with DCFH-DA flow cytometry and cytochrome c reduction. Clin. Chim. Acta 319, 9–17. doi: 10.1016/s0009-8981(02)00015-3
Carlson-Stevermer, J., Kelso, R., Kadina, A., Joshi, S., Rossi, N., Walker, J., et al. (2020). CRISPRoff enables spatio-temporal control of CRISPR editing. Nat. Commun. 11:5041.
Carnesecchi, S., Deffert, C., Donati, Y., Basset, O., Hinz, B., Preynat-Seauve, O., et al. (2011). A key role for NOX4 in epithelial cell death during development of lung fibrosis. Antioxid. Redox Signal. 15, 607–619. doi: 10.1089/ars.2010.3829
Carnesecchi, S., Deffert, C., Pagano, A., Garrido-Urbani, S., Metrailler-Ruchonnet, I., Schappi, M., et al. (2009). NADPH oxidase-1 plays a crucial role in hyperoxia-induced acute lung injury in mice. Am. J. Respir. Crit. Care Med. 180, 972–981. doi: 10.1164/rccm.200902-0296oc
Chanin, R. B., Winter, M. G., Spiga, L., Hughes, E. R., Zhu, W., Taylor, S. J., et al. (2020). Epithelial-derived reactive Oxygen Species enable AppBCX-mediated aerobic respiration of Escherichia coli during intestinal inflammation. Cell Host Microbe 28, 780–788.e5.
Chauvigne, F., Ducat, C., Ferre, A., Hansen, T., Carrascal, M., Abian, J., et al. (2021). A multiplier peroxiporin signal transduction pathway powers piscine spermatozoa. Proc. Natl. Acad. Sci. U.S.A. 118:e2019346118. doi: 10.1073/pnas.2019346118
Chavez, V., Mohri-Shiomi, A., and Garsin, D. A. (2009). Ce-Duox1/BLI-3 generates reactive oxygen species as a protective innate immune mechanism in Caenorhabditis elegans. Infect. Immun. 77, 4983–4989. doi: 10.1128/iai.00627-09
Chen, C., Mahar, R., Merritt, M. E., Denlinger, D. L., and Hahn, D. A. (2021). ROS and hypoxia signaling regulate periodic metabolic arousal during insect dormancy to coordinate glucose, amino acid, and lipid metabolism. Proc. Natl. Acad. Sci. U.S.A. 118:e2017603118. doi: 10.1073/pnas.2017603118
Chen, H. C., Su, L. T., Gonzalez-Pagan, O., Overton, J. D., and Runnels, L. W. (2012). A key role for Mg(2+) in TRPM7’s control of ROS levels during cell stress. Biochem. J. 445, 441–448. doi: 10.1042/bj20120248
Chen, Q., Vazquez, E. J., Moghaddas, S., Hoppel, C. L., and Lesnefsky, E. J. (2003). Production of reactive oxygen species by mitochondria: central role of complex III. J. Biol. Chem. 278, 36027–36031.
Chen, Y., Azad, M. B., and Gibson, S. B. (2009). Superoxide is the major reactive oxygen species regulating autophagy. Cell Death Differ. 16, 1040–1052. doi: 10.1038/cdd.2009.49
Chiarugi, P., Fiaschi, T., Taddei, M. L., Talini, D., Giannoni, E., Raugei, G., et al. (2001). Two vicinal cysteines confer a peculiar redox regulation to low molecular weight protein tyrosine phosphatase in response to platelet-derived growth factor receptor stimulation. J. Biol. Chem. 276, 33478–33487. doi: 10.1074/jbc.m102302200
Choi, J., Ifuku, M., Noda, M., and Guilarte, T. R. (2011). Translocator protein (18 kDa)/peripheral benzodiazepine receptor specific ligands induce microglia functions consistent with an activated state. Glia 59, 219–230. doi: 10.1002/glia.21091
Cordeiro, R. M. (2014). Reactive oxygen species at phospholipid bilayers: distribution, mobility and permeation. Biochim. Biophys. Acta 1838, 438–444. doi: 10.1016/j.bbamem.2013.09.016
Couto, N., Wood, J., and Barber, J. (2016). The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 95, 27–42. doi: 10.1016/j.freeradbiomed.2016.02.028
Craig, M., and Slauch, J. M. (2009). Phagocytic superoxide specifically damages an extracytoplasmic target to inhibit or kill Salmonella. PLoS One 4:e4975. doi: 10.1371/journal.pone.0004975
Cross, C. E., Halliwell, B., Borish, E. T., Pryor, W. A., Ames, B. N., Saul, R. L., et al. (1987). Oxygen radicals and human disease. Ann. Intern. Med. 107, 526–545. doi: 10.7326/0003-4819-107-4-526
Dahlgren, C., and Karlsson, A. (1999). Respiratory burst in human neutrophils. J. Immunol. Methods 232, 3–14. doi: 10.1016/s0022-1759(99)00146-5
Damiano, S., Sozio, C., La Rosa, G., and Santillo, M. (2020). NOX-dependent signaling dysregulation in severe COVID-19: clues to effective treatments. Front. Cell. Infect. Microbiol. 10:608435. doi: 10.3389/fcimb.2020.608435
Das, K., and Roychoudhury, A. (2014). Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2:53. doi: 10.3389/fenvs.2014.00053
D’Autreaux, B., and Toledano, M. B. (2007). ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 8, 813–824. doi: 10.1038/nrm2256
Davies, M. J., Forni, L. G., and Willson, R. L. (1988). Vitamin E analogue Trolox C. E.s.r. and pulse-radiolysis studies of free-radical reactions. Biochem. J. 255, 513–522.
Deng, H., Yang, W., Zhou, Z., Tian, R., Lin, L., Ma, Y., et al. (2020). Targeted scavenging of extracellular ROS relieves suppressive immunogenic cell death. Nat. Commun. 11:4951.
Deshmukh, P., Unni, S., Krishnappa, G., and Padmanabhan, B. (2017). The Keap1-Nrf2 pathway: promising therapeutic target to counteract ROS-mediated damage in cancers and neurodegenerative diseases. Biophys. Rev. 9, 41–56. doi: 10.1007/s12551-016-0244-4
Dikalova, A. E., Gongora, M. C., Harrison, D. G., Lambeth, J. D., Dikalov, S., and Griendling, K. K. (2010). Upregulation of Nox1 in vascular smooth muscle leads to impaired endothelium-dependent relaxation via eNOS uncoupling. Am. J. Physiol. Heart Circ. Physiol. 299, H673–H679.
Dinakar, C., Vishwakarma, A., Raghavendra, A. S., and Padmasree, K. (2016). Alternative oxidase pathway optimizes photosynthesis during osmotic and temperature stress by regulating cellular ROS, malate valve and antioxidative systems. Front. Plant Sci. 7:68. doi: 10.3389/fpls.2016.00068
Donko, A., Ruisanchez, E., Orient, A., Enyedi, B., Kapui, R., Peterfi, Z., et al. (2010). Urothelial cells produce hydrogen peroxide through the activation of Duox1. Free Radic. Biol. Med. 49, 2040–2048. doi: 10.1016/j.freeradbiomed.2010.09.027
Dostert, C., Petrilli, V., Van Bruggen, R., Steele, C., Mossman, B. T., and Tschopp, J. (2008). Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 320, 674–677. doi: 10.1126/science.1156995
Dugas, A. J. Jr., Castaneda-Acosta, J., Bonin, G. C., Price, K. L., Fischer, N. H., and Winston, G. W. (2000). Evaluation of the total peroxyl radical-scavenging capacity of flavonoids: structure-activity relationships. J. Nat. Prod. 63, 327–331. doi: 10.1021/np990352n
Ermakova, Y. G., Bilan, D. S., Matlashov, M. E., Mishina, N. M., Markvicheva, K. N., Subach, O. M., et al. (2014). Red fluorescent genetically encoded indicator for intracellular hydrogen peroxide. Nat. Commun. 5:5222.
Ernst, O., Sun, J., Lin, B., Banoth, B., Dorrington, M. G., Liang, J., et al. (2021). A genome-wide screen uncovers multiple roles for mitochondrial nucleoside diphosphate kinase D in inflammasome activation. Sci. Signal. 14:eabe0387. doi: 10.1126/scisignal.abe0387
Ezerina, D., Takano, Y., Hanaoka, K., Urano, Y., and Dick, T. P. (2018). N-Acetyl cysteine functions as a fast-acting antioxidant by triggering intracellular H2S and Sulfane Sulfur Production. Cell Chem. Biol. 25, 447–459.e4.
Fan, L. M., Geng, L., Cahill-Smith, S., Liu, F., Douglas, G., Mckenzie, C. A., et al. (2019). Nox2 contributes to age-related oxidative damage to neurons and the cerebral vasculature. J. Clin. Invest. 129, 3374–3386. doi: 10.1172/jci125173
Fenton, H. J. H. (1894). LXXIII.—Oxidation of tartaric acid in presence of iron. J. Chem. Soc. Trans. 65, 899–910. doi: 10.1039/ct8946500899
Fransen, M., Lismont, C., and Walton, P. (2017). The peroxisome-mitochondria connection: How and Why? Int. J. Mol. Sci. 18:1126. doi: 10.3390/ijms18061126
Fridovich, I. (1997). Superoxide anion radical (O2-.), superoxide dismutases, and related matters. J. Biol. Chem. 272, 18515–18517. doi: 10.1074/jbc.272.30.18515
Gaggini, F., Laleu, B., Orchard, M., Fioraso-Cartier, L., Cagnon, L., Houngninou-Molango, S., et al. (2011). Design, synthesis and biological activity of original pyrazolo-pyrido-diazepine, -pyrazine and -oxazine dione derivatives as novel dual Nox4/Nox1 inhibitors. Bioorg. Med. Chem. 19, 6989–6999. doi: 10.1016/j.bmc.2011.10.016
Garaude, J., Acin-Perez, R., Martinez-Cano, S., Enamorado, M., Ugolini, M., Nistal-Villan, E., et al. (2016). Mitochondrial respiratory-chain adaptations in macrophages contribute to antibacterial host defense. Nat. Immunol. 17, 1037–1045. doi: 10.1038/ni.3509
Gatliff, J., East, D. A., Singh, A., Alvarez, M. S., Frison, M., Matic, I., et al. (2017). A role for TSPO in mitochondrial Ca(2+) homeostasis and redox stress signaling. Cell Death Dis. 8:e2896. doi: 10.1038/cddis.2017.186
Gavazzi, G., Banfi, B., Deffert, C., Fiette, L., Schappi, M., Herrmann, F., et al. (2006). Decreased blood pressure in NOX1-deficient mice. FEBS Lett. 580, 497–504. doi: 10.1016/j.febslet.2005.12.049
Geng, J., Sun, X., Wang, P., Zhang, S., Wang, X., Wu, H., et al. (2015). Kinases Mst1 and Mst2 positively regulate phagocytic induction of reactive oxygen species and bactericidal activity. Nat. Immunol. 16, 1142–1152. doi: 10.1038/ni.3268
Geng, L., Fan, L. M., Liu, F., Smith, C., and Li, J. (2020). Nox2 dependent redox-regulation of microglial response to amyloid-beta stimulation and microgliosis in aging. Sci. Rep. 10:1582.
Geyer, O., Podos, S. M., and Mittag, T. (1997). Nitric oxide synthase activity in tissues of the bovine eye. Graefes Arch. Clin. Exp. Ophthalmol. 235, 786–793. doi: 10.1007/bf02332864
Gluschko, A., Herb, M., Wiegmann, K., Krut, O., Neiss, W. F., Utermohlen, O., et al. (2018). The beta2 Integrin Mac-1 induces protective LC3-associated phagocytosis of Listeria monocytogenes. Cell Host Microbe 23, 324–337.e5.
Go, Y. M., and Jones, D. P. (2008). Redox compartmentalization in eukaryotic cells. Biochim. Biophys. Acta 1780, 1273–1290. doi: 10.1016/j.bbagen.2008.01.011
Gorin, Y., Cavaglieri, R. C., Khazim, K., Lee, D. Y., Bruno, F., Thakur, S., et al. (2015). Targeting NADPH oxidase with a novel dual Nox1/Nox4 inhibitor attenuates renal pathology in type 1 diabetes. Am. J. Physiol. Renal Physiol. 308, F1276–F1287.
Green, D. E., Murphy, T. C., Kang, B. Y., Kleinhenz, J. M., Szyndralewiez, C., Page, P., et al. (2012). The Nox4 inhibitor GKT137831 attenuates hypoxia-induced pulmonary vascular cell proliferation. Am. J. Respir. Cell Mol. Biol. 47, 718–726. doi: 10.1165/rcmb.2011-0418oc
Gutscher, M., Sobotta, M. C., Wabnitz, G. H., Ballikaya, S., Meyer, A. J., Samstag, Y., et al. (2009). Proximity-based protein thiol oxidation by H2O2-scavenging peroxidases. J. Biol. Chem. 284, 31532–31540. doi: 10.1074/jbc.m109.059246
Gutteridge, J. M. C., and Halliwell, B. (2018). Mini-Review: Oxidative stress, redox stress or redox success? Biochem. Biophys. Res. Commun. 502, 183–186. doi: 10.1016/j.bbrc.2018.05.045
Haber, F., Weiss, J., and Pope, W. J. (1997). The catalytic decomposition of hydrogen peroxide by iron salts. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 147, 332–351. doi: 10.1098/rspa.1934.0221
Hadrava Vanova, K., Kraus, M., Neuzil, J., and Rohlena, J. (2020). Mitochondrial complex II and reactive oxygen species in disease and therapy. Redox Rep. 25, 26–32. doi: 10.1080/13510002.2020.1752002
Haj-Yehia, A. I., Nassar, T., Assaf, P., Nassar, H., and Anggard, E. E. (1999). Effects of the superoxide dismutase-mimic compound TEMPOL on oxidant stress-mediated endothelial dysfunction. Antioxid. Redox Signal. 1, 221–232. doi: 10.1089/ars.1999.1.2-221
Han, D., Antunes, F., Canali, R., Rettori, D., and Cadenas, E. (2003). Voltage-dependent anion channels control the release of the superoxide anion from mitochondria to cytosol. J. Biol. Chem. 278, 5557–5563. doi: 10.1074/jbc.m210269200
Han, H. (2018). RNA interference to knock down gene expression. Methods Mol. Biol. 1706, 293–302. doi: 10.1007/978-1-4939-7471-9_16
Hatz, S., Lambert, J. D., and Ogilby, P. R. (2007). Measuring the lifetime of singlet oxygen in a single cell: addressing the issue of cell viability. Photochem. Photobiol. Sci. 6, 1106–1116. doi: 10.1039/b707313e
Hayyan, M., Hashim, M. A., and Alnashef, I. M. (2016). Superoxide ion: generation and chemical implications. Chem. Rev. 116, 3029–3085. doi: 10.1021/acs.chemrev.5b00407
Hein, T. W., and Kuo, L. (1998). LDLs impair vasomotor function of the coronary microcirculation: role of superoxide anions. Circ. Res. 83, 404–414. doi: 10.1161/01.res.83.4.404
Heinz, S., Freyberger, A., Lawrenz, B., Schladt, L., Schmuck, G., and Ellinger-Ziegelbauer, H. (2017). Mechanistic investigations of the mitochondrial complex I inhibitor rotenone in the context of pharmacological and safety evaluation. Sci. Rep. 7:45465.
Hempel, S. L., Buettner, G. R., O’malley, Y. Q., Wessels, D. A., and Flaherty, D. M. (1999). Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: comparison with 2′,7′-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic. Biol. Med. 27, 146–159.
Herb, M., and Schramm, M. (2021). Functions of ROS in macrophages and antimicrobial immunity. Antioxidants10:313. doi: 10.3390/antiox10020313
Herb, M., Farid, A., Gluschko, A., Kronke, M., and Schramm, M. (2019a). Highly efficient transfection of primary macrophages with in vitro transcribed mRNA. J. Vis. Exp. 153.
Herb, M., Gluschko, A., Wiegmann, K., Farid, A., Wolf, A., Utermohlen, O., et al. (2019b). Mitochondrial reactive oxygen species enable proinflammatory signaling through disulfide linkage of NEMO. Sci. Signal. 12:eaar5926. doi: 10.1126/scisignal.aar5926
Hernández-Barrera, A., Quinto, C., Johnson, E. A., Wu, H.-M., Cheung, A. Y., and Cárdenas, L. (2013). Using hyper as a molecular probe to visualize hydrogen peroxide in living plant cells: a method with virtually unlimited potential in plant biology. Methods Enzymol. 527, 275–290. doi: 10.1016/b978-0-12-405882-8.00015-5
Hernansanz-Agustin, P., Choya-Foces, C., Carregal-Romero, S., Ramos, E., Oliva, T., Villa-Pina, T., et al. (2020). Na(+) controls hypoxic signalling by the mitochondrial respiratory chain. Nature 586, 287–291.
Herscovitch, M., Comb, W., Ennis, T., Coleman, K., Yong, S., Armstead, B., et al. (2008). Intermolecular disulfide bond formation in the NEMO dimer requires Cys54 and Cys347. Biochem. Biophys. Res. Commun. 367, 103–108. doi: 10.1016/j.bbrc.2007.12.123
Hervera, A., De Virgiliis, F., Palmisano, I., Zhou, L., Tantardini, E., Kong, G., et al. (2018). Reactive oxygen species regulate axonal regeneration through the release of exosomal NADPH oxidase 2 complexes into injured axons. Nat. Cell Biol. 20, 307–319. doi: 10.1038/s41556-018-0039-x
Heumuller, S., Wind, S., Barbosa-Sicard, E., Schmidt, H. H., Busse, R., Schroder, K., et al. (2008). Apocynin is not an inhibitor of vascular NADPH oxidases but an antioxidant. Hypertension 51, 211–217. doi: 10.1161/hypertensionaha.107.100214
Hoeven, R., Mccallum, K. C., Cruz, M. R., and Garsin, D. A. (2011). Ce-Duox1/BLI-3 generated reactive oxygen species trigger protective SKN-1 activity via p38 MAPK signaling during infection in C. elegans. PLoS Pathog. 7:e1002453. doi: 10.1371/journal.ppat.1002453
Holmstrom, K. M., and Finkel, T. (2014). Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 15, 411–421. doi: 10.1038/nrm3801
Hos, N. J., Ganesan, R., Gutierrez, S., Hos, D., Klimek, J., Abdullah, Z., et al. (2017). Type I interferon enhances necroptosis of Salmonella Typhimurium-infected macrophages by impairing antioxidative stress responses. J. Cell Biol. 216, 4107–4121.
Huang, H., Ullah, F., Zhou, D. X., Yi, M., and Zhao, Y. (2019). Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 10:800. doi: 10.3389/fpls.2019.00800
Imlay, J. A. (2003). Pathways of oxidative damage. Annu. Rev. Microbiol. 57, 395–418. doi: 10.1146/annurev.micro.57.030502.090938
Inomata, M., Xu, S., Chandra, P., Meydani, S. N., Takemura, G., Philips, J. A., et al. (2020). Macrophage LC3-associated phagocytosis is an immune defense against Streptococcus pneumoniae that diminishes with host aging. Proc. Natl. Acad. Sci. U.S.A. 117, 33561–33569. doi: 10.1073/pnas.2015368117
Ives, A., Nomura, J., Martinon, F., Roger, T., Leroy, D., Miner, J. N., et al. (2015). Xanthine oxidoreductase regulates macrophage IL1beta secretion upon NLRP3 inflammasome activation. Nat. Commun. 6:6555.
Janku, M., Luhova, L., and Petrivalsky, M. (2019). On the Origin and Fate of Reactive Oxygen Species in Plant Cell Compartments. Antioxidants 8:105. doi: 10.3390/antiox8040105
Jones, A. I., Meshulam, T., Oliveira, M. F., Burritt, N., and Corkey, B. E. (2016). Extracellular redox regulation of intracellular reactive oxygen generation, mitochondrial function and lipid turnover in cultured human Adipocytes. PLoS One 11:e0164011. doi: 10.1371/journal.pone.0164011
Juan, C. A., De La Lastra, J. M. P., Plou, F. J., and Perez-Lebena, E. (2021). The chemistry of reactive oxygen species (ROS) revisited: outlining their role in biological macromolecules (DNA, Lipids and Proteins) and induced pathologies. Int. J. Mol. Sci. 22, 4642.
Kamata, H., Honda, S., Maeda, S., Chang, L., Hirata, H., and Karin, M. (2005). Reactive oxygen species promote TNFalpha-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell 120, 649–661. doi: 10.1016/j.cell.2004.12.041
Kehrer, J. P., Robertson, J. D., and Smith, C. V. (2010). “1.14 - Free radicals and reactive oxygen species,” in Comprehensive Toxicology, 2nd Edn, ed. C. A. Mcqueen (Oxford: Elsevier), 277–307.
Kelley, E. E., Khoo, N. K., Hundley, N. J., Malik, U. Z., Freeman, B. A., and Tarpey, M. M. (2010). Hydrogen peroxide is the major oxidant product of xanthine oxidase. Free Radic. Biol. Med. 48, 493–498. doi: 10.1016/j.freeradbiomed.2009.11.012
Kelly, B., Tannahill, G. M., Murphy, M. P., and O’neill, L. A. (2015). Metformin Inhibits the Production of Reactive Oxygen Species from NADH:Ubiquinone Oxidoreductase to Limit Induction of Interleukin-1beta (IL-1beta) and Boosts Interleukin-10 (IL-10) in Lipopolysaccharide (LPS)-activated Macrophages. J. Biol. Chem. 290, 20348–20359. doi: 10.1074/jbc.m115.662114
Kim, H. K., Lee, H. Y., Riaz, T. A., Bhattarai, K. R., Chaudhary, M., Ahn, J. H., et al. (2021). Chalcone suppresses tumor growth through NOX4-IRE1alpha sulfonation-RIDD-miR-23b axis. Redox Biol. 40:101853. doi: 10.1016/j.redox.2021.101853
Kim, H., and Xue, X. (2020). Detection of total reactive Oxygen Species in Adherent Cells by 2′,7′-Dichlorodihydrofluorescein Diacetate Staining. J. Vis. Exp. 160:10.3791/60682. doi: 10.3791/60682
Kim, J. H., Lee, J., Bae, S. J., Kim, Y., Park, B. J., Choi, J. W., et al. (2017). NADPH oxidase 4 is required for the generation of macrophage migration inhibitory factor and host defense against Toxoplasma gondii infection. Sci. Rep. 7:6361.
Kim, N. H., Choi, S., Han, E. J., Hong, B. K., Choi, S. Y., Kwon, H. M., et al. (2014). The xanthine oxidase-NFAT5 pathway regulates macrophage activation and TLR-induced inflammatory arthritis. Eur. J. Immunol. 44, 2721–2736. doi: 10.1002/eji.201343669
Kiritoshi, S., Nishikawa, T., Sonoda, K., Kukidome, D., Senokuchi, T., Matsuo, T., et al. (2003). Reactive oxygen species from mitochondria induce cyclooxygenase-2 gene expression in human mesangial cells: potential role in diabetic nephropathy. Diabetes Metab. Res. Rev. 52, 2570–2577. doi: 10.2337/diabetes.52.10.2570
Kirkman, H. N., and Gaetani, G. F. (2007). Mammalian catalase: a venerable enzyme with new mysteries. Trends Biochem. Sci. 32, 44–50. doi: 10.1016/j.tibs.2006.11.003
Knight, L. J., Martis, R. M., Acosta, M. L., Donaldson, P. J., and Lim, J. C. (2021). Detection of reduced mitochondrial ROS production but increased ROS levels and oxidative damage in the young xCT knockout mouse retina. Invest. Ophthalmol. Vis. Sci. 62, 2230–2230.
Koterski, J. F., Nahvi, M., Venkatesan, M. M., and Haimovich, B. (2005). Virulent Shigella flexneri causes damage to mitochondria and triggers necrosis in infected human monocyte-derived macrophages. Infect. Immun. 73, 504–513. doi: 10.1128/iai.73.1.504-513.2005
Krishna, C. M., Liebmann, J. E., Kaufman, D., Degraff, W., Hahn, S. M., Mcmurry, T., et al. (1992). The catecholic metal sequestering agent 1,2-dihydroxybenzene-3,5-disulfonate confers protection against oxidative cell damage. Arch. Biochem. Biophys. 294, 98–106. doi: 10.1016/0003-9861(92)90142-j
Laleu, B., Gaggini, F., Orchard, M., Fioraso-Cartier, L., Cagnon, L., Houngninou-Molango, S., et al. (2010). First in class, potent, and orally bioavailable NADPH oxidase isoform 4 (Nox4) inhibitors for the treatment of idiopathic pulmonary fibrosis. J. Med. Chem. 53, 7715–7730. doi: 10.1021/jm100773e
Lambert, A. J., and Brand, M. D. (2004). Inhibitors of the quinone-binding site allow rapid superoxide production from mitochondrial NADH:ubiquinone oxidoreductase (complex I). J. Biol. Chem. 279, 39414–39420. doi: 10.1074/jbc.m406576200
Lambeth, J. D., and Neish, A. S. (2014). Nox enzymes and new thinking on reactive oxygen: a double-edged sword revisited. Annu. Rev. Pathol. 9, 119–145. doi: 10.1146/annurev-pathol-012513-104651
Lambeth, J. D., Krause, K. H., and Clark, R. A. (2008). NOX enzymes as novel targets for drug development. Semin. Immunopathol. 30, 339–363. doi: 10.1007/s00281-008-0123-6
Lanciano, P., Khalfaoui-Hassani, B., Selamoglu, N., Ghelli, A., Rugolo, M., and Daldal, F. (2013). Molecular mechanisms of superoxide production by complex III: a bacterial versus human mitochondrial comparative case study. Biochim. Biophys. Acta 1827, 1332–1339. doi: 10.1016/j.bbabio.2013.03.009
Lee, E. Y., Bae, H. C., Lee, H., Jang, Y., Park, Y. H., Kim, J. H., et al. (2017). Intracellular ROS levels determine the apoptotic potential of keratinocyte by Quantum Dot via blockade of AKT Phosphorylation. Exp. Dermatol. 26, 1046–1052. doi: 10.1111/exd.13365
Lee, S. R., Kwon, K. S., Kim, S. R., and Rhee, S. G. (1998). Reversible inactivation of protein-tyrosine phosphatase 1B in A431 cells stimulated with epidermal growth factor. J. Biol. Chem. 273, 15366–15372. doi: 10.1074/jbc.273.25.15366
Leusen, J. H., Fluiter, K., Hilarius, P. M., Roos, D., Verhoeven, A. J., and Bolscher, B. G. (1995). Interactions between the cytosolic components p47phox and p67phox of the human neutrophil NADPH oxidase that are not required for activation in the cell-free system. J. Biol. Chem. 270, 11216–11221.
Levine, A., Tenhaken, R., Dixon, R., and Lamb, C. (1994). H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 79, 583–593. doi: 10.1016/0092-8674(94)90544-4
Li, T. Y., Sun, Y., Liang, Y., Liu, Q., Shi, Y., Zhang, C. S., et al. (2016). ULK1/2 constitute a bifurcate node controlling glucose metabolic fluxes in addition to autophagy. Mol. Cell 62, 359–370. doi: 10.1016/j.molcel.2016.04.009
Li, X., Fang, P., Mai, J., Choi, E. T., Wang, H., and Yang, X. F. (2013). Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers. J. Hematol. Oncol. 6:19. doi: 10.1186/1756-8722-6-19
Ligeon, L. A., Pena-Francesch, M., and Munz, C. (2021a). Measuring oxidation within LC3-associated phagosomes that optimizes MHC class II restricted antigen presentation. Methods Cell Biol. 164, 187–200. doi: 10.1016/bs.mcb.2021.02.003
Ligeon, L. A., Pena-Francesch, M., Vanoaica, L. D., Nunez, N. G., Talwar, D., Dick, T. P., et al. (2021b). Oxidation inhibits autophagy protein deconjugation from phagosomes to sustain MHC class II restricted antigen presentation. Nat. Commun. 12:1508.
Lin, T. K., Lin, K. J., Lin, K. L., Liou, C. W., Chen, S. D., Chuang, Y. C., et al. (2020). When friendship turns sour: effective communication between mitochondria and intracellular organelles in Parkinson’s disease. Front. Cell. Dev. Biol. 8:607392. doi: 10.3389/fcell.2020.607392
Liochev, S. I., and Fridovich, I. (1999). Superoxide and iron: partners in crime. IUBMB Life 48, 157–161. doi: 10.1080/713803492
Lismont, C., Nordgren, M., Van Veldhoven, P. P., and Fransen, M. (2015). Redox interplay between mitochondria and peroxisomes. Front. Cell Dev. Biol. 3:35. doi: 10.3389/fcell.2015.00035
Loth, M. K., Guariglia, S. R., Re, D. B., Perez, J., De Paiva, V. N., Dziedzic, J. L., et al. (2020). A novel interaction of translocator protein 18 kDa (TSPO) with NADPH Oxidase in Microglia. Mol. Neurobiol. 57, 4467–4487. doi: 10.1007/s12035-020-02042-w
Lundqvist, H., and Dahlgren, C. (1996). Isoluminol-enhanced chemiluminescence: a sensitive method to study the release of superoxide anion from human neutrophils. Free Radic. Biol. Med. 20, 785–792. doi: 10.1016/0891-5849(95)02189-2
Mak, T. W., Grusdat, M., Duncan, G. S., Dostert, C., Nonnenmacher, Y., Cox, M., et al. (2017). Glutathione primes T cell metabolism for inflammation. Immunity 46, 675–689. doi: 10.1016/j.immuni.2017.03.019
Manzano, V. M., Munoz, J. C., Jimenez, J. R., Puyol, M. R., Puyol, D. R., Kitamura, M., et al. (2000). Human renal mesangial cells are a target for the anti-inflammatory action of 9-cis retinoic acid. Br. J. Pharmacol. 131, 1673–1683. doi: 10.1038/sj.bjp.0703728
Marinho, H. S., Cyrne, L., Cadenas, E., and Antunes, F. (2013). The cellular steady-state of H2O2: latency concepts and gradients. Methods Enzymol. 527, 3–19. doi: 10.1016/b978-0-12-405882-8.00001-5
Marklund, S. (1976). Spectrophotometric study of spontaneous disproportionation of superoxide anion radical and sensitive direct assay for superoxide dismutase. J. Biol. Chem. 251, 7504–7507. doi: 10.1016/s0021-9258(17)32878-8
Markvicheva, K. N., Bilan, D. S., Mishina, N. M., Gorokhovatsky, A. Y., Vinokurov, L. M., Lukyanov, S., et al. (2011). A genetically encoded sensor for H2O2 with expanded dynamic range. Bioorg. Med. Chem. 19, 1079–1084. doi: 10.1016/j.bmc.2010.07.014
Martinez, M. A., Ubeda, A., and Trillo, M. A. (2021). Role of NADPH oxidase in MAPK signaling activation by a 50 Hz magnetic field in human neuroblastoma cells. Electromagn. Biol. Med. 40, 103–116. doi: 10.1080/15368378.2020.1851250
Matsushita, T., Fukuda, K., Yamamoto, H., Yamazaki, K., Tomiyama, T., Oh, M., et al. (2004). Effect of ebselen, a scavenger of reactive oxygen species, on chondrocyte metabolism. Mod. Rheumatol. 14, 25–30. doi: 10.3109/s10165-003-0261-6
Meinhard, M., and Grill, E. (2001). Hydrogen peroxide is a regulator of ABI1, a protein phosphatase 2C from Arabidopsis. FEBS Lett. 508, 443–446. doi: 10.1016/s0014-5793(01)03106-4
Melov, S., Coskun, P., Patel, M., Tuinstra, R., Cottrell, B., Jun, A. S., et al. (1999). Mitochondrial disease in superoxide dismutase 2 mutant mice. Proc. Natl. Acad. Sci. U.S.A. 96, 846–851.
Mendiola, A. S., Ryu, J. K., Bardehle, S., Meyer-Franke, A., Ang, K. K., Wilson, C., et al. (2020). Transcriptional profiling and therapeutic targeting of oxidative stress in neuroinflammation. Nat. Immunol. 21, 513–524.
Meng, T. C., Fukada, T., and Tonks, N. K. (2002). Reversible oxidation and inactivation of protein tyrosine phosphatases in vivo. Mol. Cell 9, 387–399. doi: 10.1016/s1097-2765(02)00445-8
Miller, B. M., Liou, M. J., Zhang, L. F., Nguyen, H., Litvak, Y., Schorr, E. M., et al. (2020). Anaerobic respiration of NOX1-derived hydrogen peroxide licenses bacterial growth at the colonic surface. Cell Host Microbe 28, 789–797.e5.
Mora-Pale, M., Weiwer, M., Yu, J., Linhardt, R. J., and Dordick, J. S. (2009). Inhibition of human vascular NADPH oxidase by apocynin derived oligophenols. Bioorg. Med. Chem. 17, 5146–5152. doi: 10.1016/j.bmc.2009.05.061
Mugesh, G. (2013). Glutathione peroxidase activity of ebselen and its analogues: some insights into the complex chemical mechanisms underlying the antioxidant activity. Curr. Chem. Biol. 7, 47–56. doi: 10.2174/2212796811307010005
Mukhopadhyay, P., Rajesh, M., Hasko, G., Hawkins, B. J., Madesh, M., and Pacher, P. (2007). Simultaneous detection of apoptosis and mitochondrial superoxide production in live cells by flow cytometry and confocal microscopy. Nat. Protoc. 2, 2295–2301. doi: 10.1038/nprot.2007.327
Muller, F. L., Liu, Y., and Van Remmen, H. (2004). Complex III releases superoxide to both sides of the inner mitochondrial membrane. J. Biol. Chem. 279, 49064–49073. doi: 10.1074/jbc.m407715200
Murphy, M. P. (2009). How mitochondria produce reactive oxygen species. Biochem. J. 417, 1–13. doi: 10.1042/bj20081386
Nakamura, H. (2005). Thioredoxin and its related molecules: update 2005. Antioxid. Redox Signal. 7, 823–828. doi: 10.1089/ars.2005.7.823
Nakamura, Y., Feng, Q., Kumagai, T., Torikai, K., Ohigashi, H., Osawa, T., et al. (2002). Ebselen, a glutathione peroxidase mimetic seleno-organic compound, as a multifunctional antioxidant. Implication for inflammation-associated carcinogenesis. J. Biol. Chem. 277, 2687–2694. doi: 10.1074/jbc.m109641200
Nakano, Y., Longo-Guess, C. M., Bergstrom, D. E., Nauseef, W. M., Jones, S. M., and Banfi, B. (2008). Mutation of the Cyba gene encoding p22phox causes vestibular and immune defects in mice. J. Clin. Invest. 118, 1176–1185.
Nathan, C., and Cunningham-Bussel, A. (2013). Beyond oxidative stress: an immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 13, 349–361. doi: 10.1038/nri3423
Naviaux, R. K. (2012). Oxidative shielding or oxidative stress? J. Pharmacol. Exp. Ther. 342, 608–618. doi: 10.1124/jpet.112.192120
Nazarewicz, R. R., Dikalova, A. E., Bikineyeva, A., and Dikalov, S. I. (2013). Nox2 as a potential target of mitochondrial superoxide and its role in endothelial oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 305, H1131– H1140.
Niethammer, P., Grabher, C., Look, A. T., and Mitchison, T. J. (2009). A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in Zebrafish. Nature 459, 996–999. doi: 10.1038/nature08119
Niki, E. (2016). Oxidative stress and antioxidants: Distress or eustress? Arch. Biochem. Biophys. 595, 19–24. doi: 10.1016/j.abb.2015.11.017
Nohl, H., Kozlov, A. V., Gille, L., and Staniek, K. (2003). Cell respiration and formation of reactive oxygen species: facts and artefacts. Biochem. Soc. Trans. 31, 1308–1311. doi: 10.1042/bst0311308
Nomura, J., Busso, N., Ives, A., Matsui, C., Tsujimoto, S., Shirakura, T., et al. (2014). Xanthine oxidase inhibition by febuxostat attenuates experimental atherosclerosis in mice. Sci. Rep. 4:4554.
Odobasic, D., Kitching, A. R., and Holdsworth, S. R. (2016). Neutrophil-mediated regulation of innate and adaptive immunity: the role of Myeloperoxidase. J. Immunol. Res. 2016:2349817.
O’Donnell, B. V., Tew, D. G., Jones, O. T., and England, P. J. (1993). Studies on the inhibitory mechanism of iodonium compounds with special reference to neutrophil NADPH oxidase. Biochem. J. 290(Pt 1), 41–49. doi: 10.1042/bj2900041
Omura, T., and Sato, R. (1962). A new cytochrome in liver microsomes. J. Biol. Chem. 237, 1375–1376.
Panov, A., Dikalov, S., Shalbuyeva, N., Taylor, G., Sherer, T., and Greenamyre, J. T. (2005). Rotenone model of Parkinson disease: multiple brain mitochondria dysfunctions after short term systemic rotenone intoxication. J. Biol. Chem. 280, 42026–42035.
Patriarca, S., Furfaro, A. L., Domenicotti, C., Odetti, P., Cottalasso, D., Marinari, U. M., et al. (2005). Supplementation with N-acetylcysteine and taurine failed to restore glutathione content in liver of streptozotocin-induced diabetics rats but protected from oxidative stress. Biochim. Biophys. Acta 1741, 48–54. doi: 10.1016/j.bbadis.2005.04.003
Pavelkova, M., and Kubala, L. (2004). Luminol-, isoluminol- and lucigenin-enhanced chemiluminescence of rat blood phagocytes stimulated with different activators. Luminescence 19, 37–42. doi: 10.1002/bio.754
Peng, H. Y., Lucavs, J., Ballard, D., Das, J. K., Kumar, A., Wang, L., et al. (2021). Metabolic reprogramming and reactive Oxygen Species in T cell immunity. Front. Immunol. 12:652687. doi: 10.3389/fimmu.2021.652687
Perillo, B., Di Donato, M., Pezone, A., Di Zazzo, E., Giovannelli, P., Galasso, G., et al. (2020). ROS in cancer therapy: the bright side of the moon. Exp. Mol. Med. 52, 192–203. doi: 10.1038/s12276-020-0384-2
Pizzino, G., Irrera, N., Cucinotta, M., Pallio, G., Mannino, F., Arcoraci, V., et al. (2017). Oxidative stress: harms and benefits for human health. Oxid. Med. Cell. Longev. 2017:8416763.
Poljsak, B., Suput, D., and Milisav, I. (2013). Achieving the balance between ROS and antioxidants: when to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013:956792.
Poole, L. B. (2015). The basics of thiols and cysteines in redox biology and chemistry. Free Radic. Biol. Med. 80, 148–157. doi: 10.1016/j.freeradbiomed.2014.11.013
Prestes, E. B., Alves, L. S., Rodrigues, D. A. S., Dutra, F. F., Fernandez, P. L., Paiva, C. N., et al. (2020). Mitochondrial reactive oxygen species participate in signaling triggered by Heme in macrophages and upon Hemolysis. J. Immunol. 205, 2795–2805. doi: 10.4049/jimmunol.1900886
Prousek, J. (2007). Fenton chemistry in biology and medicine. Pure Appl. Chem. 79, 2325–2338. doi: 10.1351/pac200779122325
Quinlan, C. L., Orr, A. L., Perevoshchikova, I. V., Treberg, J. R., Ackrell, B. A., and Brand, M. D. (2012). Mitochondrial complex II can generate reactive oxygen species at high rates in both the forward and reverse reactions. J. Biol. Chem. 287, 27255–27264. doi: 10.1074/jbc.m112.374629
Rak, R., Chao, D. L., Pluta, R. M., Mitchell, J. B., Oldfield, E. H., and Watson, J. C. (2000). Neuroprotection by the stable nitroxide Tempol during reperfusion in a rat model of transient focal ischemia. J. Neurosurg. 92, 646–651. doi: 10.3171/jns.2000.92.4.0646
Rangasamy, T., Guo, J., Mitzner, W. A., Roman, J., Singh, A., Fryer, A. D., et al. (2005). Disruption of Nrf2 enhances susceptibility to severe airway inflammation and asthma in mice. J. Exp. Med. 202, 47–59. doi: 10.1084/jem.20050538
Rapoport, R., Hanukoglu, I., and Sklan, D. (1994). A fluorimetric assay for hydrogen peroxide, suitable for NAD(P)H-dependent superoxide generating redox systems. Anal. Biochem. 218, 309–313. doi: 10.1006/abio.1994.1183
Reczek, C. R., and Chandel, N. S. (2014). ROS-dependent signal transduction. Curr. Opin. Cell Biol. 33C, 8–13. doi: 10.1016/j.ceb.2014.09.010
Reczek, C. R., and Chandel, N. S. (2017). The two faces of reactive oxygen species in cancer. Annu. Rev. Cancer Biol. 1, 79–98. doi: 10.1146/annurev-cancerbio-041916-065808
Reis, J., Massari, M., Marchese, S., Ceccon, M., Aalbers, F. S., Corana, F., et al. (2020). A closer look into NADPH oxidase inhibitors: validation and insight into their mechanism of action. Redox Biol. 32:101466. doi: 10.1016/j.redox.2020.101466
Ren, X., Wang, M., Wang, Y., and Huang, A. (2021). Superoxide anion generation response to wound in Arabidopsis hypocotyl cutting. Plant Signal. Behav. 16:1848086. doi: 10.1080/15592324.2020.1848086
Rhee, S. G. (2006). Cell signaling. H2O2, a necessary evil for cell signaling. Science 312, 1882–1883. doi: 10.1126/science.1130481
Rieger, S., and Sagasti, A. (2011). Hydrogen peroxide promotes injury-induced peripheral sensory axon regeneration in the Zebrafish skin. PLoS Biol. 9:e1000621. doi: 10.1371/journal.pbio.1000621
Robinson, K. M., Janes, M. S., Pehar, M., Monette, J. S., Ross, M. F., Hagen, T. M., et al. (2006). Selective fluorescent imaging of superoxide in vivo using ethidium-based probes. Proc. Natl. Acad. Sci. U.S.A. 103, 15038–15043. doi: 10.1073/pnas.0601945103
Roca, F. J., and Ramakrishnan, L. (2013). TNF dually mediates resistance and susceptibility to mycobacteria via mitochondrial reactive oxygen species. Cell 153, 521–534. doi: 10.1016/j.cell.2013.03.022
Roca, F. J., Whitworth, L. J., Redmond, S., Jones, A. A., and Ramakrishnan, L. (2019). TNF induces pathogenic programmed macrophage necrosis in tuberculosis through a mitochondrial-lysosomal-endoplasmic reticulum circuit. Cell 178, 1344–1361.e11.
Romero, L. C., Aroca, M. A., Laureano-Marin, A. M., Moreno, I., Garcia, I., and Gotor, C. (2014). Cysteine and cysteine-related signaling pathways in Arabidopsis thaliana. Mol. Plant 7, 264–276. doi: 10.1093/mp/sst168
Saito, Y., and Kimura, W. (2021). Roles of phase separation for cellular redox maintenance. Front. Genet. 12:691946. doi: 10.3389/fgene.2021.691946
Schumacker, P. T. (2006). Reactive oxygen species in cancer cells: live by the sword, die by the sword. Cancer Cell 10, 175–176. doi: 10.1016/j.ccr.2006.08.015
Scialo, F., Fernandez-Ayala, D. J., and Sanz, A. (2017). Role of mitochondrial reverse electron transport in ROS signaling: potential roles in health and disease. Front. Physiol. 8:428. doi: 10.3389/fphys.2017.00428
Sedeek, M., Callera, G., Montezano, A., Gutsol, A., Heitz, F., Szyndralewiez, C., et al. (2010). Critical role of Nox4-based NADPH oxidase in glucose-induced oxidative stress in the kidney: implications in type 2 diabetic nephropathy. Am. J. Physiol. Renal Physiol. 299, F1348–F1358.
Sena, L. A., Li, S., Jairaman, A., Prakriya, M., Ezponda, T., Hildeman, D. A., et al. (2013). Mitochondria are required for antigen-specific T cell activation through reactive oxygen species signaling. Immunity 38, 225–236. doi: 10.1016/j.immuni.2012.10.020
Shai, N., Yifrach, E., Van Roermund, C. W. T., Cohen, N., Bibi, C., IJlst, L., et al. (2018). Systematic mapping of contact sites reveals tethers and a function for the peroxisome-mitochondria contact. Nat. Commun. 9:1761.
Sharma, P., Jha, A. B., Dubey, R. S., and Pessarakli, M. (2012). Reactive oxygen Species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012:217037.
Short, J. D., Downs, K., Tavakoli, S., and Asmis, R. (2016). Protein Thiol Redox signaling in monocytes and macrophages. Antioxid. Redox Signal. 25, 816–835. doi: 10.1089/ars.2016.6697
Shoshan-Barmatz, V., De Pinto, V., Zweckstetter, M., Raviv, Z., Keinan, N., and Arbel, N. (2010). VDAC, a multi-functional mitochondrial protein regulating cell life and death. Mol. Aspects Med. 31, 227–285. doi: 10.1016/j.mam.2010.03.002
Sies, H. (2021). Oxidative eustress: on constant alert for redox homeostasis. Redox Biol. 41:101867. doi: 10.1016/j.redox.2021.101867
Sies, H., and Jones, D. P. (2020). Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 21, 363–383. doi: 10.1038/s41580-020-0230-3
Stavru, F., Bouillaud, F., Sartori, A., Ricquier, D., and Cossart, P. (2011). Listeria monocytogenes transiently alters mitochondrial dynamics during infection. Proc. Natl. Acad. Sci. U.S.A. 108, 3612–3617. doi: 10.1073/pnas.1100126108
Stowe, D. F., and Camara, A. K. (2009). Mitochondrial reactive oxygen species production in excitable cells: modulators of mitochondrial and cell function. Antioxid. Redox Signal. 11, 1373–1414. doi: 10.1089/ars.2008.2331
St-Pierre, J., Buckingham, J. A., Roebuck, S. J., and Brand, M. D. (2002). Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 277, 44784–44790. doi: 10.1074/jbc.m207217200
Strengert, M., Jennings, R., Davanture, S., Hayes, P., Gabriel, G., and Knaus, U. G. (2014). Mucosal reactive oxygen species are required for antiviral response: role of Duox in influenza a virus infection. Antioxid. Redox Signal. 20, 2695–2709. doi: 10.1089/ars.2013.5353
Stuehr, D. J., Fasehun, O. A., Kwon, N. S., Gross, S. S., Gonzalez, J. A., Levi, R., et al. (1991). Inhibition of macrophage and endothelial cell nitric oxide synthase by diphenyleneiodonium and its analogs. FASEB J. 5, 98–103. doi: 10.1096/fasebj.5.1.1703974
Su, L. J., Zhang, J. H., Gomez, H., Murugan, R., Hong, X., Xu, D., et al. (2019). Reactive Oxygen Species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid. Med. Cell. Longev. 2019:5080843.
Suzuki, T., Muramatsu, A., Saito, R., Iso, T., Shibata, T., Kuwata, K., et al. (2019). Molecular mechanism of cellular oxidative stress sensing by Keap1. Cell Rep. 28, 746–758.e4.
Sweeney, R. M., and McAuley, D. F. (2016). Acute respiratory distress syndrome. Lancet 388, 2416–2430.
Tai, T. C., Wong-Faull, D. C., Claycomb, R., and Wong, D. L. (2009). Hypoxic stress-induced changes in adrenergic function: role of HIF1 alpha. J. Neurochem. 109, 513–524. doi: 10.1111/j.1471-4159.2009.05978.x
Takahashi, M. A., and Asada, K. (1983). Superoxide anion permeability of phospholipid membranes and chloroplast thylakoids. Arch. Biochem. Biophys. 226, 558–566. doi: 10.1016/0003-9861(83)90325-9
Taverne, Y. J., Bogers, A. J., Duncker, D. J., and Merkus, D. (2013). Reactive oxygen species and the cardiovascular system. Oxid. Med. Cell. Longev. 2013: 862423.
Tazzeo, T., Worek, F., and Janssen, L. (2009). The NADPH oxidase inhibitor diphenyleneiodonium is also a potent inhibitor of cholinesterases and the internal Ca(2+) pump. Br. J. Pharmacol. 158, 790–796. doi: 10.1111/j.1476-5381.2009.00394.x
Teixeira, G., Szyndralewiez, C., Molango, S., Carnesecchi, S., Heitz, F., Wiesel, P., et al. (2017). Therapeutic potential of NADPH oxidase 1/4 inhibitors. Br. J. Pharmacol. 174, 1647–1669.
ten Freyhaus, H., Huntgeburth, M., Wingler, K., Schnitker, J., Baumer, A. T., Vantler, M., et al. (2006). Novel Nox inhibitor VAS2870 attenuates PDGF-dependent smooth muscle cell chemotaxis, but not proliferation. Cardiovasc. Res. 71, 331–341. doi: 10.1016/j.cardiores.2006.01.022
Tepel, M., Echelmeyer, M., Orie, N. N., and Zidek, W. (2000). Increased intracellular reactive oxygen species in patients with end-stage renal failure: effect of hemodialysis. Kidney Int. 58, 867–872. doi: 10.1046/j.1523-1755.2000.00236.x
Thimmulappa, R. K., Lee, H., Rangasamy, T., Reddy, S. P., Yamamoto, M., Kensler, T. W., et al. (2006). Nrf2 is a critical regulator of the innate immune response and survival during experimental sepsis. J. Clin. Invest. 116, 984–995. doi: 10.1172/jci25790
Thorne, L. S., Rochford, G., Williams, T. D., Southam, A. D., Rodriguez-Blanco, G., Dunn, W. B., et al. (2021). Cytoglobin protects cancer cells from apoptosis by regulation of mitochondrial cardiolipin. Sci. Rep. 11:985.
To, E. E., Vlahos, R., Luong, R., Halls, M. L., Reading, P. C., King, P. T., et al. (2017). Endosomal NOX2 oxidase exacerbates virus pathogenicity and is a target for antiviral therapy. Nat. Commun. 8:69.
Tonks, N. K. (2005). Redox redux: revisiting PTPs and the control of cell signaling. Cell 121, 667–670. doi: 10.1016/j.cell.2005.05.016
Trevelin, S. C., Dos Santos, C. X., Ferreira, R. G., De Sa Lima, L., Silva, R. L., Scavone, C., et al. (2016). Apocynin and Nox2 regulate NF-kappaB by modifying thioredoxin-1 redox-state. Sci. Rep. 6:34581.
Troia, A., Knutsen, R. H., Halabi, C. M., Malide, D., Yu, Z. X., Wardlaw-Pickett, A., et al. (2021). Inhibition of NOX1 Mitigates Blood Pressure Increases in Elastin Insufficiency. Function (Oxf) 2:zqab015.
Ushijima, Y., Totsune, H., Nishida, A., and Nakano, M. (1997). Chemiluminescence from human polymorphonuclear leukocytes activated with opsonized zymosan. Free Radic. Biol. Med. 22, 401–409. doi: 10.1016/s0891-5849(96)00329-2
Veal, E. A., Day, A. M., and Morgan, B. A. (2007). Hydrogen peroxide sensing and signaling. Mol. Cell 26, 1–14. doi: 10.1016/j.molcel.2007.03.016
Wang, H., Schoebel, S., Schmitz, F., Dong, H., and Hedfalk, K. (2020). Characterization of aquaporin-driven hydrogen peroxide transport. Biochim. Biophys. Acta Biomembr. 1862:183065. doi: 10.1016/j.bbamem.2019.183065
Wang, J., Zhang, W., Wang, S., Wang, Y., Chu, X., Ji, H., et al. (2021a). Lactobacillus plantarum exhibits antioxidant and cytoprotective activities in porcine intestinal epithelial cells exposed to hydrogen peroxide. Oxid. Med. Cell. Longev. 2021:8936907.
Wang, X., Jiang, Z., Yue, N., Jin, X., Zhang, X., Li, Z., et al. (2021b). Barley stripe mosaic virus gammab protein disrupts chloroplast antioxidant defenses to optimize viral replication. EMBO J 40:e107660.
Wang, P., Geng, J., Gao, J., Zhao, H., Li, J., Shi, Y., et al. (2019). Macrophage achieves self-protection against oxidative stress-induced ageing through the Mst-Nrf2 axis. Nat. Commun. 10:755.
Wang, Q., and Zou, M. H. (2018). Measurement of reactive oxygen Species (ROS) and Mitochondrial ROS in AMPK Knockout Mice Blood Vessels. Methods Mol. Biol. 1732, 507–517. doi: 10.1007/978-1-4939-7598-3_32
Wang, Y., Branicky, R., Noe, A., and Hekimi, S. (2018). Superoxide dismutases: dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 217, 1915–1928. doi: 10.1083/jcb.201708007
Wang, Z., Cai, F., Chen, X., Luo, M., Hu, L., and Lu, Y. (2013). The role of mitochondria-derived reactive oxygen species in hyperthermia-induced platelet apoptosis. PLoS One 8:e75044. doi: 10.1371/journal.pone.0075044
Wei, R., Enaka, M., and Muragaki, Y. (2019). Activation of KEAP1/NRF2/P62 signaling alleviates high phosphate-induced calcification of vascular smooth muscle cells by suppressing reactive oxygen species production. Sci. Rep. 9:10366.
West, A. P., Brodsky, I. E., Rahner, C., Woo, D. K., Erdjument-Bromage, H., Tempst, P., et al. (2011a). TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 472, 476–480. doi: 10.1038/nature09973
West, A. P., Shadel, G. S., and Ghosh, S. (2011b). Mitochondria in innate immune responses. Nat. Rev. Immunol. 11, 389–402.
Wind, S., Beuerlein, K., Eucker, T., Muller, H., Scheurer, P., Armitage, M. E., et al. (2010). Comparative pharmacology of chemically distinct NADPH oxidase inhibitors. Br. J. Pharmacol. 161, 885–898. doi: 10.1111/j.1476-5381.2010.00920.x
Wingler, K., Hermans, J. J., Schiffers, P., Moens, A., Paul, M., and Schmidt, H. H. (2011). NOX1, 2, 4, 5: counting out oxidative stress. Br. J. Pharmacol. 164, 866–883. doi: 10.1111/j.1476-5381.2011.01249.x
Winitchaikul, T., Sawong, S., Surangkul, D., Srikummool, M., Somran, J., Pekthong, D., et al. (2021). Calotropis gigantea stem bark extract induced apoptosis related to ROS and ATP production in colon cancer cells. PLoS One 16:e0254392. doi: 10.1371/journal.pone.0254392
Wink, D. A., Hines, H. B., Cheng, R. Y., Switzer, C. H., Flores-Santana, W., Vitek, M. P., et al. (2011). Nitric oxide and redox mechanisms in the immune response. J. Leukoc. Biol. 89, 873–891.
Winterbourn, C. C., and Hampton, M. B. (2008). Thiol chemistry and specificity in redox signaling. Free Radic. Biol. Med. 45, 549–561. doi: 10.1016/j.freeradbiomed.2008.05.004
Winterbourn, C. C., and Kettle, A. J. (2013). Redox reactions and microbial killing in the neutrophil phagosome. Antioxid. Redox Signal. 18, 642–660. doi: 10.1089/ars.2012.4827
Wolf, A., Herb, M., Schramm, M., and Langmann, T. (2020). The TSPO-NOX1 axis controls phagocyte-triggered pathological angiogenesis in the eye. Nat. Commun. 11:2709.
Wong, H. S., Mezera, V., Dighe, P., Melov, S., Gerencser, A. A., Sweis, R. F., et al. (2021). Superoxide produced by mitochondrial site IQ inactivates cardiac succinate dehydrogenase and induces hepatic steatosis in Sod2 knockout mice. Free Radic. Biol. Med. 164, 223–232. doi: 10.1016/j.freeradbiomed.2020.12.447
Woo, H. A., Yim, S. H., Shin, D. H., Kang, D., Yu, D. Y., and Rhee, S. G. (2010). Inactivation of peroxiredoxin I by phosphorylation allows localized H(2)O(2) accumulation for cell signaling. Cell 140, 517–528. doi: 10.1016/j.cell.2010.01.009
Wood, Z. A., Poole, L. B., and Karplus, P. A. (2003). Peroxiredoxin evolution and the regulation of hydrogen peroxide signaling. Science 300, 650–653. doi: 10.1126/science.1080405
Xu, J., Tran, T., Padilla Marcia, C. S., Braun, D. M., and Goggin, F. L. (2017). Superoxide-responsive gene expression in Arabidopsis thaliana and Zea mays. Plant Physiol. Biochem. 117, 51–60. doi: 10.1016/j.plaphy.2017.05.018
Yang, Y., Karakhanova, S., Hartwig, W., D’haese, J. G., Philippov, P. P., Werner, J., et al. (2016). Mitochondria and mitochondrial ROS in cancer: novel targets for anticancer therapy. J. Cell. Physiol. 231, 2570–2581. doi: 10.1002/jcp.25349
Zaidieh, T., Smith, J. R., Ball, K. E., and An, Q. (2019). ROS as a novel indicator to predict anticancer drug efficacy. BMC Cancer 19:1224. doi: 10.1186/s12885-019-6438-y
Zeller, J., Bogner, B., Kiefer, J., Braig, D., Winninger, O., Fricke, M., et al. (2021). CRP Enhances the Innate Killing Mechanisms Phagocytosis and ROS formation in a conformation and complement-dependent manner. Front. Immunol. 12:754. doi: 10.3389/fimmu.2018.00754
Zhang, J., Wang, X., Vikash, V., Ye, Q., Wu, D., Liu, Y., et al. (2016). ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016:4350965.
Zhang, W., Hu, X., Shen, Q., and Xing, D. (2019). Mitochondria-specific drug release and reactive oxygen species burst induced by polyprodrug nanoreactors can enhance chemotherapy. Nat. Commun. 10:1704.
Zhang, X. D., Peng, Y. Q., Zhao, J., Li, Q., Yu, X. J., Acevedo-Rocha, C. G., et al. (2020). Bacterial cytochrome P450-catalyzed regio- and stereoselective steroid hydroxylation enabled by directed evolution and rational design. Bioresour. Bioprocess. 7:2.
Zhang, Y., Yao, Y., Qiu, X., Wang, G., Hu, Z., Chen, S., et al. (2019). Listeria hijacks host mitophagy through a novel mitophagy receptor to evade killing. Nat. Immunol. 20, 433–446. doi: 10.1038/s41590-019-0324-2
Zhao, Q., Liu, J., Deng, H., Ma, R., Liao, J. Y., Liang, H., et al. (2020). Targeting mitochondria-located circRNA SCAR Alleviates NASH via Reducing mROS Output. Cell 183, 76–93.e22.
Zhao, R. Z., Jiang, S., Zhang, L., and Yu, Z. B. (2019). Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 44, 3–15.
Zhong, J., Li, Q., Luo, H., and Holmdahl, R. (2021). Neutrophil-derived reactive oxygen species promote tumor colonization. Commun. Biol. 4:865.
Zhou, L., Yeo, A. T., Ballarano, C., Weber, U., Allen, K. N., Gilmore, T. D., et al. (2014). Disulfide-mediated stabilization of the IkappaB kinase binding domain of NF-kappaB essential modulator (NEMO). Biochemistry 53, 7929–7944. doi: 10.1021/bi500920n
Keywords: mitochondria, NADPH oxidases, ROS sources, reactive oxygen species, redox balance, oxidative stress, ROS probes, ROS inhibitors
Citation: Herb M, Gluschko A and Schramm M (2021) Reactive Oxygen Species: Not Omnipresent but Important in Many Locations. Front. Cell Dev. Biol. 9:716406. doi: 10.3389/fcell.2021.716406
Received: 28 May 2021; Accepted: 19 August 2021;
Published: 07 September 2021.
Edited by:
Bechan Sharma, Allahabad University, IndiaReviewed by:
Takuya Sakaguchi, Cleveland Clinic, United StatesAndrea N. Beltrán González, Molecular Biology and Neurosciences (IFIBYNE), Argentina
Roberto Giovannoni, University of Pisa, Italy
Copyright © 2021 Herb, Gluschko and Schramm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marc Herb, bWFyYy5oZXJiQHVrLWtvZWxuLmRl