 Helen E. Abud
Helen E. Abud Wing Hei Chan
Wing Hei Chan Thierry Jardé
Thierry Jardé- 1Department of Anatomy and Developmental Biology, Monash University, Clayton, VIC, Australia
- 2Development and Stem Cells Program, Monash Biomedicine Discovery Institute, Clayton, VIC, Australia
Epidermal Growth Factor (EGF) has long been known for its role in promoting proliferation of intestinal epithelial cells. EGF is produced by epithelial niche cells at the base of crypts in vivo and is routinely added to the culture medium to support the growth of intestinal organoids ex vivo. The recent identification of diverse stromal cell populations that reside underneath intestinal crypts has enabled the characterization of key growth factor cues supplied by these cells. The nature of these signals and how they are delivered to drive intestinal epithelial development, daily homeostasis and tissue regeneration following injury are being investigated. It is clear that aside from EGF, other ligands of the family, including Neuregulin 1 (NRG1), have distinct roles in supporting the function of intestinal stem cells through the ErbB pathway.
Introduction
A balance between cell proliferation and differentiation is exquisitely controlled in the intestinal epithelium throughout life (Beumer and Clevers, 2021). This is coordinated by a system of cues from surrounding niche cells that includes Paneth cells in the epithelium (Sato et al., 2011), diverse populations of stromal cells (Hageman et al., 2020; McCarthy et al., 2020a; Sphyris et al., 2021), enteric neural cells (Van Landeghem et al., 2011; Talbot et al., 2020) and macrophages (De Schepper et al., 2018; Sehgal et al., 2018). These signals act on the epithelium to modulate stem cell function and cell fate acquisition in progenitor cells (Tetteh et al., 2016). This complex array of cellular inputs has the ability to support the enormous expansion of the intestinal tract during development (Chin et al., 2017) and the strong proliferative response occurring during tissue repair following damage to ensure integrity of the epithelium (Hageman et al., 2020). Maintenance of the barrier and adequate tissue function is vital to prevent systemic infection from enteric pathogens and adequate digestion and nutrient absorption. Stem cells, which reside in the base of intestinal crypts, either self-renew or generate transit-amplifying progenitor cells that ultimately differentiate and generate the diversity of secretory and absorptive differentiated cell types required for a functional epithelium (Beumer and Clevers, 2021). Interplay of key signals from the WNT, Notch, Epidermal Growth Factor (EGF), and Bone Morphogenetic Protein (BMP) signaling pathways regulate the survival, self-renewal and differentiation of these cells to ensure a balance of cell types (Holik et al., 2013; Horvay and Abud, 2013; Tian et al., 2015; McCarthy et al., 2020b; Beumer and Clevers, 2021). The exact mechanisms of how signals are produced, what cell types secrete and receive signals and how tissues respond to promote the process of regeneration are being investigated. Some of the cell types that secrete molecules that either augment or inhibit WNT and BMP signaling have been described. EGF is present in Paneth cells and has primarily been shown to influence proliferation (Abud et al., 2005; Basak et al., 2017), but it is becoming increasingly clear that other ligands from this family can also influence the diversity of cells within the epithelium (Jardé et al., 2020; Holloway et al., 2021). Whether these ligands have distinct or functionally redundant activities and how signals influence the epithelium in different contexts is still being investigated. In this review, we discuss current evidence on the cellular source and role of ligands from the EGF family and how they interact with receptors in the epithelium to influence cellular proliferation, stem cell identity and lineage differentiation.
The EGF Family of Receptors and Ligands
The EGF family of ligands includes eleven structurally related proteins, namely EGF, transforming growth factor α (TGF-α), amphiregulin (AREG), epigen (EPGN), heparin-binding EGF-like growth factor (HB-EGF), epiregulin (EREG), betacellulin (BTC), and the neuregulins (NRG1-4) (Figure 1). These molecules have in common similar EGF-like motifs, and, due to their membrane-anchored nature, can act in a juxtacrine manner between two neighboring cells, or, in an autocrine/paracrine mode via proteolytic cleavage of the external EGF-like domain, which results in its release in the extracellular compartment (Singh and Harris, 2005; Rayego-Mateos et al., 2018). The EGF-like protein drives cellular signal transduction through the ErbB subclass of the Receptor Tyrosine Kinase superfamily, which consists of four members EGFR (also known as ErbB1), ErbB2, ErbB3, and ErbB4 (Downward et al., 1984; Schechter et al., 1984; Semba et al., 1985; Kraus et al., 1989; Plowman et al., 1993). The EGF family of ligands can be classified into four sub-groups based on distinct receptor binding specificities: (1) the ligands which recognize ErbB1 only (EGF, TGFα, AREG, and EPGN); (2) the ligands binding to both ErbB1 and ErbB4 (HB-EGF, EREG, and BTC); (3) the ligands which are specific for both ErbB3 and ErbB4 (NRG1 and NRG2); and (4) the ligands activating ErbB4 only (NRG3 and NRG4) (Figure 1). It should be noted that no ligands have been identified for ErbB2 to date. Nonetheless, all ErbB receptors contain an extracellular ligand binding site, a single membrane spanning region and a cytoplasmic tyrosine-kinase-containing domain (Lemmon et al., 2014). Upon ligand-induced conformational change, ErbB receptors form homodimers or heterodimers (Figure 1), which activates the intrinsic kinase domain, resulting in the phosphorylation of specific tyrosine residues within the cytoplasmic tails (Lemmon et al., 2014). Phosphorylated residues serve as docking sites for a range of molecules and regulatory proteins involved in various cascades of intracellular signaling, including MAPK, PI3K-AKT, and JAK-STAT (Singh and Harris, 2005; Iwakura and Nawa, 2013). These complex signaling routes regulate many key cellular functions, including cell proliferation, cell death and stem cell maintenance, which are essential for numerous body systems.
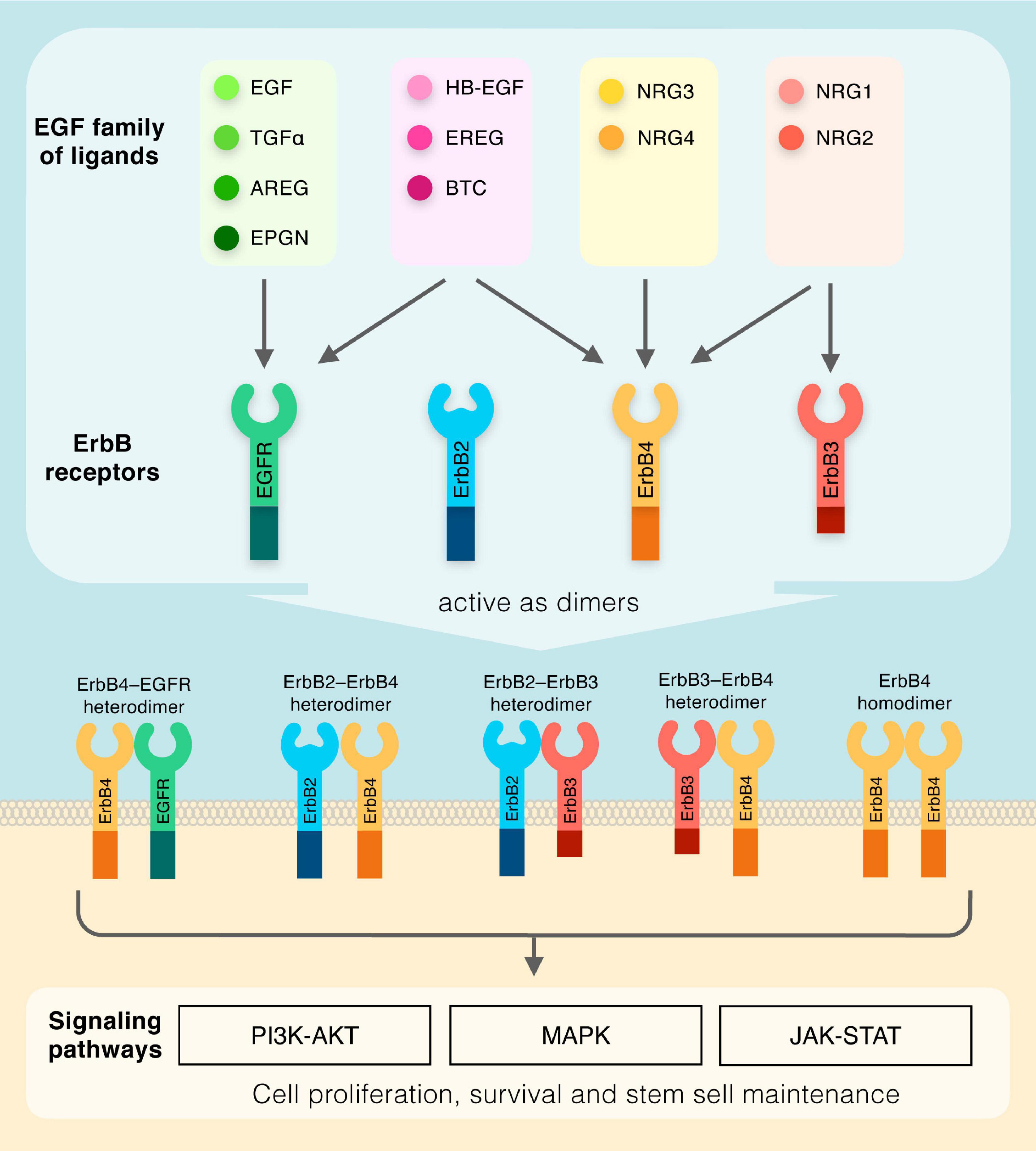
Figure 1. Diagram outlining the members of the EGF family of ligands and their respective receptors. Dimeric receptors activate downstream PI3K-AKT, MAPK, and JAK-STAT signaling pathways.
Location and Function of Signals and Receptors During Mammalian Homeostasis
In mice and humans, Paneth cells are localized within the epithelium adjacent to intestinal stem cells in the base of crypts. These specialized cells secrete WNT3, EGF and NRG1 during intestinal homeostasis (Sato et al., 2011; Jardé et al., 2020; Figure 2). Although initially identified as a key constituent of the niche, it is clear that when Paneth cells are depleted in vivo, other cellular sources can compensate for this loss, resulting in the maintenance of an intact epithelium (Durand et al., 2012; Farin et al., 2012). The stromal compartment of the adult intestinal tract contains sub-populations of cells including FOXL1+ telocytes that secrete WNT2B and the WNT signaling potentiator RSPO3 (Aoki et al., 2016; Shoshkes-Carmel et al., 2018) and PDGFRα+ cells (Greicius et al., 2018; McCarthy et al., 2020b) that also secrete WNT2B and RSPO3. Mesenchymal cells marked by GLI1 also secrete WNT in the colon (Degirmenci et al., 2018). Single cell sequencing has revealed distinct sub-populations that express different levels of PDGFRα, with those expressing higher levels being localized to the villi, and CD34+ PDGFRαlow cells residing at the base of crypts, where they express Grem1 and antagonize BMP signaling. These cells described as trophocytes are capable of fully supporting the growth of organoids ex vivo (McCarthy et al., 2020b). A similar sub-population of fibroblasts associated with crypts was also recently identified in the colon (Brugger et al., 2020). MAP3K2 expressing cells also reside at the base of crypts where they secrete RSPO1 (Wu et al., 2021). Macrophages identified by expression of CD11b and CSF1R are also closely associated with crypts and when depleted, loss of intestinal stem cells are observed (De Schepper et al., 2018; Sehgal et al., 2018). A unifying global expression analysis of the EGF family of ligands in both epithelial and mesenchymal intestinal niche sub-populations during intestinal homeostasis is currently missing. Re-analysis of recently published single cell RNA sequencing datasets would help understanding the complex ligand dynamics in the intestinal tract (Kinchen et al., 2018; Kim et al., 2020; Busslinger et al., 2021; Yu et al., 2021). Studies, which focused on individual ligands, have shown extremely low levels of EGF can be detected in mesenchymal cell populations, while NRG1 is expressed at relatively high levels in stromal cells, including PDGFRα+ cells, and is observed in macrophages (Jardé et al., 2020). Both EGF and NRG1-expressing cells are also found in the developing human intestinal tract, with EGF localized in the epithelial villus domain and NRG1 detected in PDGFRα+ cells within the subepithelial mesenchyme underlying crypts (Holloway et al., 2021; Yu et al., 2021). Other members of the NRG family of ligands display exclusive low expression in the mesenchyme (NRG2) or in the epithelium (NRG4) while NRG3 is absent in both compartments in adult intestinal tissues. AREG shares a similar expression pattern with NRG1, with an enrichment in subepithelial myofibroblasts and in rare F4/80+ macrophage-enriched cells but with limited expression in the epithelium (Inatomi et al., 2006; Shao and Sheng, 2010; Yang et al., 2017). EREG is also localized in an uncharacterized subpopulation of mesenchymal cells and is weakly expressed in the epithelium (Xian et al., 1999; Kallincos et al., 2000; Xia et al., 2003; Lee et al., 2004). In contrast, BTC, HB-EGF, and TGFα are enriched in the intestinal epithelium. The expression pattern of EPGN in both mesenchymal and epithelial compartments is currently unknown. Interestingly, EGFR/ERBB receptors are present within the epithelial cells of the epithelium (Figure 2). EGFR is enriched in stem and progenitor cells while ERBB2 and ERBB3 are detected throughout the crypt – villus axis (Suzuki et al., 2010; Jardé et al., 2020). Taken together, these data suggest a model where mesenchymal-secreted molecules, which includes NRG1, NRG2, AREG, and EREG, act on epithelial cells via a paracrine mechanism while epithelial-produced ligands such as EGF, BTC, HB-EGF, and TGFα regulate cellular function in an autocrine manner. It is important to note that the processes and signaling pathways regulating ligand production during intestinal homeostasis in these particular sub-types is not clearly defined. In addition, a comparative analysis of ligand-mediated downstream signaling pathways in normal intestinal cells is not available.
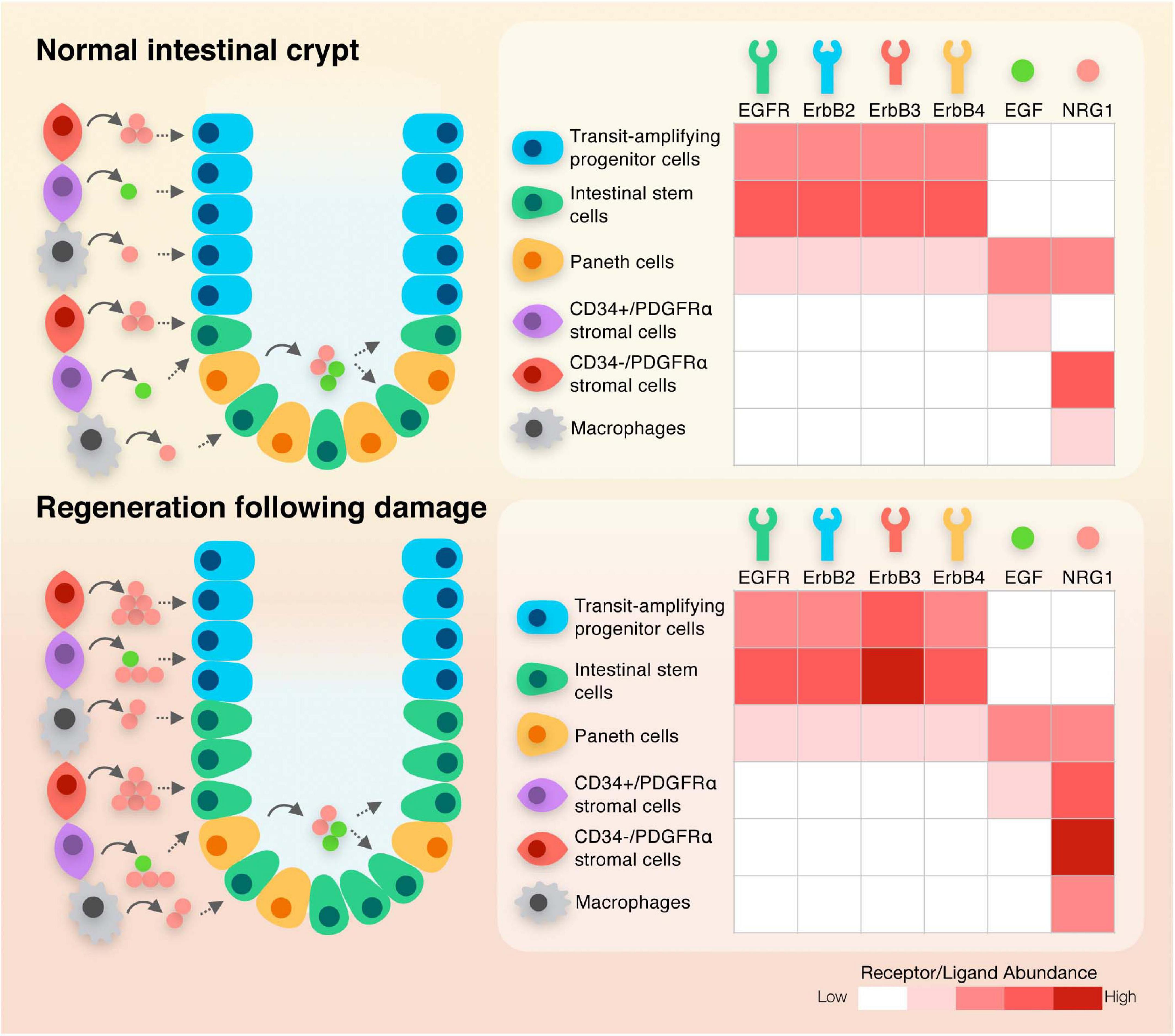
Figure 2. Schematic diagram depicting the expression of ligands and receptors within the different cellular compartments of the intestinal crypt during normal homeostasis and following regeneration.
Numerous mouse knockout studies have investigated the functional roles of individual ligands (summarized in Table 1). There is likely to be some functional redundancy given triple knockout of three of the ligands EGF, AREG, and TGFα has little effect on the intestinal epithelium with animals being viable and fertile (Luetteke et al., 1999). Similarly, single knockout of the other EGF ligands does not affect embryonic development, with the exception of Nrg1, which results in aberrant heart and neural development leading to embryonic lethality (Meyer and Birchmeier, 1995; Erickson et al., 1997). Although knockout of EGF in mice produces little effect on the epithelium, the loss of NRG1 under homeostatic conditions produces a significant reduction in proliferative stem and progenitor cells (Jardé et al., 2020; Table 1). Loss of each individual ERBB receptor is lethal and produces severe defects with significant intestinal abnormalities observed (Gassmann et al., 1995; Lee et al., 1995; Miettinen et al., 1995; Threadgill et al., 1995; Erickson et al., 1997; Riethmacher et al., 1997). Genetic background has a considerable effect on the penetrance of phenotypes (see Table 1) with knockout of EGFR exhibiting variable phenotypes from peri-implantation lethality to viable animals with multi-organ defects (Sibilia and Wagner, 1995; Threadgill et al., 1995). Individual intestinal-specific knockout of ERBB receptors has also been reported to have minimal phenotypic effects during tissue homeostasis, suggesting some functional redundancy (Lee et al., 2009; Zhang et al., 2012; Almohazey et al., 2017; Srivatsa et al., 2017). However, double and/or triple knockout combinations that are intestinal-specific will be required to confirm this.
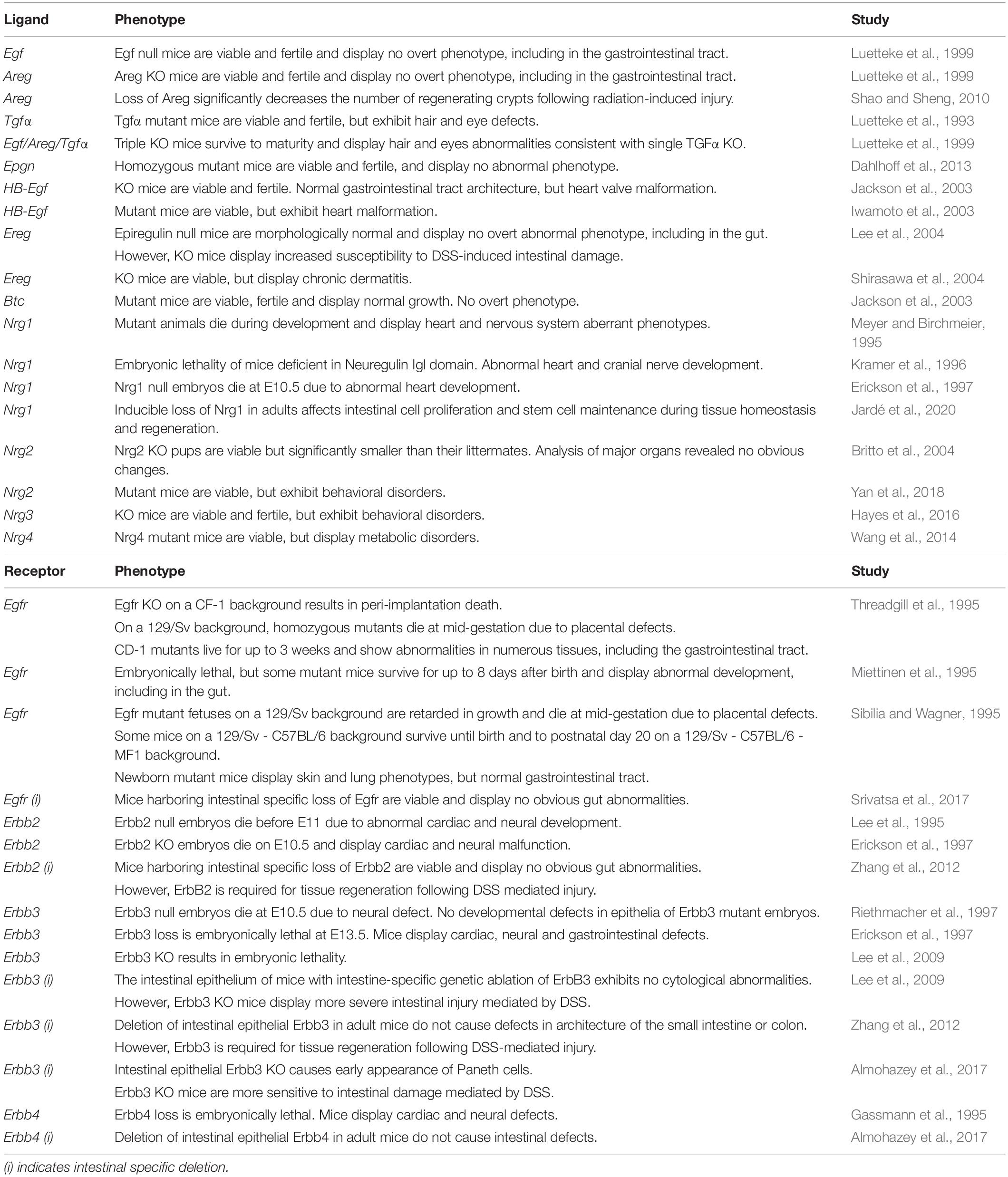
Table 1. Knockout mouse models of EGF family of ligands and receptors and their associated phenotypes.
Role of the EGF Family of Ligands and Receptors During Regeneration Following Injury
The intestinal epithelium is a selective permeable barrier that permits uptake of nutrients from the luminal contents while forming a barrier against the toxic by-products of digestion and pathogenic bacteria (Beumer and Clevers, 2021). As the epithelial monolayer is exposed to an extremely harsh chemical and mechanical environment, it is highly vulnerable to damage. This is partially compensated for by the daily dynamic renewal of the epithelial layer, with differentiated cells being replaced every few days via the activity and neutral competition of a small population of stem cells (Snippert et al., 2010). Damage induced by pathogenic bacteria that primarily impacts the villi is rapidly repaired, but severe infection can also compromise the function of stem cells deep within crypts and result in more extensive damage (Mileto et al., 2020). Inflammation and treatments such as chemotherapy and radiotherapy also impact the integrity of cells within crypts. Recovery from these insults involves a repair and regeneration process that involves extensive remodeling of both cells within the epithelium and the surrounding niche cells (Hageman et al., 2020). Following injury, macrophages are recruited and secrete WNT ligands and IL6 (Taniguchi et al., 2015), innate lymphoid cells secrete IL22 (Aparicio-Domingo et al., 2015), stromal cells are remodeled (Kinchen et al., 2018) and there are distinct changes in the extracellular matrix that generate the mechanical cues to activate YAP/TAZ signaling in the epithelium (Yui et al., 2018). Strikingly, despite the widely reported observation of EGF stimulating proliferation of intestinal cells, there is little change in the expression of EGF during the regenerative response following injury in the intestinal epithelium (Jardé et al., 2020; Figure 2). In contrast, NRG1 is robustly up-regulated following injury in macrophages, endothelial cells and in PDGFRα+ stromal cells (Figure 2). The effect of NRG1 on the epithelium induces both a strong proliferative response and induction of stem cell characteristics in regenerating crypts (Jardé et al., 2020). AREG and EREG have also been observed to be induced in epithelial cells following injury, with knockout animals displaying a significant decrease in the number of regenerating crypt domains and a more significant weight loss following injury, respectively (Lee et al., 2004; Shao and Sheng, 2010; Table 1). The expression of ErbB3 is up-regulated during regeneration (Jardé et al., 2020) and the requirement for ErbB receptor function during tissue regeneration has been clearly demonstrated using knockout models. Indeed, loss of epithelial ErbB2 or ErbB3 decreases the ability of the intestine to efficiently regenerate following DSS-mediated injury (Lee et al., 2009; Zhang et al., 2012; Almohazey et al., 2017; Table 1).
Utilizing Organoid Cultures to Interrogate Contributions of Ligands and Differential Activation of Signaling Pathways
It is clear that coordination of stem cell maintenance, progenitor proliferation and differentiation of mature cell types in the intestinal epithelium is orchestrated by gradients of active growth factors, agonists and antagonists in vivo (Tian et al., 2015; Basak et al., 2017; McCarthy et al., 2020b; Beumer and Clevers, 2021). The behavior of cells can change depending on the threshold and length of active signals and crosstalk between signaling pathways. Intestinal organoid culture was developed based on substitution of key in vivo niche signals (Sato et al., 2009, 2011) with Matrigel providing the appropriate stiffness to mimic the extracellular matrix. Under these conditions, the epithelium has the capacity to self organize, with many different epithelial cell types forming from single stem cells (Sato et al., 2009). Although this system has clear limitations, it provides an opportunity to study the intestinal epithelial population in isolation. Complex cellular interactions can be replicated by performing co-cultures with fibroblasts, nerve cells and immune cells (Kabiri et al., 2014; Rogoz et al., 2015; Pastula et al., 2016), in which genetic deletion of specific ligands can be performed and the impact on epithelial cells characterized. The effect of the microbiome and its by-products can also be evaluated (Mileto et al., 2020). However, and as opposed to applying the ligand-mediated stimulation in an unspecific manner in the current organoid technology, new systems that fully replicate the growth factor gradients observed in vivo, including WNT and EGF enrichment at the bottom of intestinal crypts, are required. Indeed, recent technological advances have allowed preservation of such complex tissue systems and gradients in culture using intestine- and organoid-on-a-chip models (Wang et al., 2018; Nikolaev et al., 2020). These systems will expand our understanding of complexity of the cellular microenvironment in which the cellular gradients of EGF family of ligands can be tested and assessed.
The relative proportion of different epithelial cell types can be manipulated in organoids by altering the culture conditions. For example, addition of CHIR99021 to elevate WNT signaling can enrich for Lgr5+ stem cells and inhibition of Notch induces secretory cells (Yin et al., 2014). Therefore, the organoid culture system permits the function of different environmental signals and signaling components to be interrogated by addition of proteins, toxins and chemical inhibitors (Clevers and Tuveson, 2019; Hageman et al., 2020). The function of the EGF family of ligands and receptors have been investigated using this methodology. EGF was included in the medium utilized for the first intestinal organoid cultures based on observations that EGF could promote proliferation of intestinal epithelial cells (McKenna et al., 1994; Dignass and Sturm, 2001; Abud et al., 2005; Sato et al., 2009). Along with R-spondin 1, EGF is required to maintain organoid crypt growth. Inhibition of signaling through the EGFR by addition of gefitinib and/or withdrawal of EGF from mouse small intestinal organoid cultures dramatically reduces proliferation within organoids and induces quiescence and an enteroendocrine molecular signature in Lgr5+ cells (Basak et al., 2017). However, one of the limitations in studying EGF signaling in intestinal organoids is the production by niche epithelial cells of EGF itself, which makes the analysis of exogenous supplementation vs. endogenous challenging.
It is important to keep in mind that the composition of the extracellular matrix surrounding epithelial cells in adult tissues, which is not fully replicated in organoid culture, might also affect the downstream molecular response to EGFR/ErbB activation (Yarwood and Woodgett, 2001). Other ligands from the family have also been tested in organoids. NRG1 can substitutes for EGF and robustly induces proliferation and budding of mouse small intestinal organoids through prolonged activation of MAPK and AKT signaling that is dramatically more effective than EGF (Jardé et al., 2020). In addition, HB-EGF supports the growth of human adult intestinal organoids in a similar fashion to EGF, which contrasts with the decreased ability of EREG to sustain organoid growth (Fujii et al., 2018). Human fetal enteroid cultures established in either NRG1 or EGF also display different phenotypes. EGF promotes proliferation and intestinal lineage identity while NRG1 supports cellular diversity and intestinal epithelial stem cell maturation (Holloway et al., 2021; Yu et al., 2021).
Conclusion
A striking feature of the intestinal epithelium is the high turnover of cells which occurs on a daily basis, the dramatic expansion of tissue during embryonic development and the rapid remodeling observed in response to injury. Members of the EGF family of ligands and their receptors contribute substantially to these processes where the current evidence suggests there are distinct functions for particular ligands but also some functional redundancy (Gregorieff et al., 2015; Jardé et al., 2020; Holloway et al., 2021). The downstream signaling pathways that mediate these processes and the crosstalk that may occur with other pathways are less defined. Lineage tracing studies have revealed the high level of plasticity present within intestinal crypts with progeny of Lgr5+ cells having the capacity to de-differentiate following injury to replace the stem cell pool (Tian et al., 2011; Metcalfe et al., 2014; Schmitt et al., 2018; Yu et al., 2018; Ayyaz et al., 2019; Jones et al., 2019; Murata et al., 2020). Although several of the intrinsic signals required for this process have been identified, the influence of signals in the microenvironment that control this are less clear. It is likely that ligands such as NRG1, which is significantly upregulated in stromal cells during regeneration, play a role (Jardé et al., 2020). Organoid cultures offer a resource to further define these activities, especially for human tissues, and future studies incorporating co-cultures of specific niche cell types will further clarify cellular mechanisms. Defining these signals could ultimately inform strategies to improve epithelial repair in conditions such as inflammatory bowel disease, necrotizing enterocolitis and short gut syndrome.
Author Contributions
HA and TJ wrote the manuscript. WC prepared the figures. HA, TJ, and WC edited and approved the final manuscript. All authors contributed to the article and approved the submitted version.
Funding
HA and TJ are supported by NHMRC Australia grants 1129600 and 2003693 (TJ), 1011187 and 1100531 (HA), and 1188689 (HA and TJ).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abud, H. E., Watson, N., and Heath, J. K. (2005). Growth of intestinal epithelium in organ culture is dependent on EGF signalling. Exp. Cell Res. 303, 252–262. doi: 10.1016/j.yexcr.2004.10.006
Almohazey, D., Lo, Y. H., Vossler, C. V., Simmons, A. J., Hsieh, J. J., Bucar, E. B., et al. (2017). The ErbB3 receptor tyrosine kinase negatively regulates Paneth cells by PI3K-dependent suppression of Atoh1. Cell Death Differ. 24, 855–865. doi: 10.1038/cdd.2017.27
Aoki, R., Shoshkes-Carmel, M., Gao, N., Shin, S., May, C. L., Golson, M. L., et al. (2016). Foxl1-expressing mesenchymal cells constitute the intestinal stem cell niche. Cell. Mol. Gastroenterol. Hepatol. 2, 175–188. doi: 10.1016/j.jcmgh.2015.12.004
Aparicio-Domingo, P., Romera-Hernandez, M., Karrich, J. J., Cornelissen, F., Papazian, N., Lindenbergh-Kortleve, D. J., et al. (2015). Type 3 innate lymphoid cells maintain intestinal epithelial stem cells after tissue damage. J. Exp. Med. 212, 1783–1791. doi: 10.1084/jem.20150318
Ayyaz, A., Kumar, S., Sangiorgi, B., Ghoshal, B., Gosio, J., Ouladan, S., et al. (2019). Single-cell transcriptomes of the regenerating intestine reveal a revival stem cell. Nature 569, 121–125. doi: 10.1038/s41586-019-1154-y
Basak, O., Beumer, J., Wiebrands, K., Seno, H., van Oudenaarden, A., and Clevers, H. (2017). Induced Quiescence of Lgr5+ Stem Cells in Intestinal Organoids Enables Differentiation of Hormone-Producing Enteroendocrine Cells. Cell Stem Cell 20, 177–190.e4.
Beumer, J., and Clevers, H. (2021). Cell fate specification and differentiation in the adult mammalian intestine. Nat. Rev. Mol. Cell Biol. 22, 39–53. doi: 10.1038/s41580-020-0278-0
Britto, J. M., Lukehurst, S., Weller, R., Fraser, C., Qiu, Y., Hertzog, P., et al. (2004). Generation and characterization of neuregulin-2-deficient mice. Mol. Cell Biol. 24, 8221–8226. doi: 10.1128/MCB.24.18.8221-8226.2004
Brugger, M. D., Valenta, T., Fazilaty, H., Hausmann, G., and Basler, K. (2020). Distinct populations of crypt-associated fibroblasts act as signaling hubs to control colon homeostasis. PLoS Biol 18:e3001032. doi: 10.1371/journal.pbio.3001032
Busslinger, G. A., Weusten, B. L. A., Bogte, A., Begthel, H., Brosens, L. A. A., and Clevers, H. (2021). Human gastrointestinal epithelia of the esophagus, stomach, and duodenum resolved at single-cell resolution. Cell Rep. 34:108819. doi: 10.1016/j.celrep.2021.108819
Chin, A. M., Hill, D. R., Aurora, M., and Spence, J. R. (2017). Morphogenesis and maturation of the embryonic and postnatal intestine. Semin. Cell Dev. Biol. 66, 81–93. doi: 10.1016/j.semcdb.2017.01.011
Clevers, H., and Tuveson, D. A. (2019). Organoid Models for Cancer Research. Annu. Rev. Cancer Biol. 3, 223–234. doi: 10.1146/annurev-cancerbio-030518-055702
Dahlhoff, M., Schäfer, M., Wolf, E., and Schneider, M. R. (2013). Genetic deletion of the EGFR ligand epigen does not affect mouse embryonic development and tissue homeostasis. Exp. Cell Res. 319, 529–535. doi: 10.1016/j.yexcr.2012.11.001
De Schepper, S., Verheijden, S., Aguilera-Lizarraga, J., Viola, M. F., Boesmans, W., Stakenborg, N., et al. (2018). Self-Maintaining Gut Macrophages Are Essential for Intestinal Homeostasis. Cell 175, 400–415.e13.
Degirmenci, B., Valenta, T., Dimitrieva, S., Hausmann, G., and Basler, K. (2018). GLI1-expressing mesenchymal cells form the essential Wnt-secreting niche for colon stem cells. Nature 558, 449–453. doi: 10.1038/s41586-018-0190-3
Dignass, A. U., and Sturm, A. (2001). Peptide growth factors in the intestine. Eur. J. Gastroenterol. Hepatol. 13, 763–770. doi: 10.1097/00042737-200107000-00002
Downward, J., Yarden, Y., Mayes, E., Scrace, G., Totty, N., Stockwell, P., et al. (1984). Close similarity of epidermal growth factor receptor and v-erb-B oncogene protein sequences. Nature 307, 521–527. doi: 10.1038/307521a0
Durand, A., Donahue, B., Peignon, G., Letourneur, F., Cagnard, N., Slomianny, C., et al. (2012). Functional intestinal stem cells after Paneth cell ablation induced by the loss of transcription factor Math1 (Atoh1). Proc. Natl. Acad. Sci. U. S. A. 109, 8965–8970. doi: 10.1073/pnas.1201652109
Erickson, S. L., O’Shea, K. S., Ghaboosi, N., Loverro, L., Frantz, G., Bauer, M., et al. (1997). ErbB3 is required for normal cerebellar and cardiac development: a comparison with ErbB2-and heregulin-deficient mice. Development 124, 4999–5011. doi: 10.1242/dev.124.24.4999
Farin, H. F., Van Es, J. H., and Clevers, H. (2012). Redundant sources of Wnt regulate intestinal stem cells and promote formation of Paneth cells. Gastroenterology 143, 1518–1529.e7.
Fujii, M., Matano, M., Toshimitsu, K., Takano, A., Mikami, Y., Nishikori, S., et al. (2018). Human Intestinal Organoids Maintain Self-Renewal Capacity and Cellular Diversity in Niche-Inspired Culture Condition. Cell Stem Cell 23, 787–793.e6.
Gassmann, M., Casagranda, F., Orioli, D., Simon, H., Lai, C., Klein, R., et al. (1995). Aberrant neural and cardiac development in mice lacking the ErbB4 neuregulin receptor. Nature 378, 390–394. doi: 10.1038/378390a0
Gregorieff, A., Liu, Y., Inanlou, M. R., Khomchuk, Y., and Wrana, J. L. (2015). Yap-dependent reprogramming of Lgr5(+) stem cells drives intestinal regeneration and cancer. Nature 526, 715–718. doi: 10.1038/nature15382
Greicius, G., Kabiri, Z., Sigmundsson, K., Liang, C., Bunte, R., Singh, M. K., et al. (2018). PDGFRalpha(+) pericryptal stromal cells are the critical source of Wnts and RSPO3 for murine intestinal stem cells in vivo. Proc. Natl. Acad. Sci. U. S. A. 115, E3173–E3181.
Hageman, J. H., Heinz, M. C., Kretzschmar, K., van der Vaart, J., Clevers, H., and Snippert, H. J. G. (2020). Intestinal regeneration: regulation by the microenvironment. Dev. Cell 54, 435–446. doi: 10.1016/j.devcel.2020.07.009
Hayes, L. N., Shevelkin, A., Zeledon, M., Steel, G., Chen, P. L., Obie, C., et al. (2016). Neuregulin 3 knockout mice exhibit behaviors consistent with psychotic disorders. Mol. Neuropsychiatry 2, 79–87. doi: 10.1159/000445836
Holik, A. Z., Krzystyniak, J., Young, M., Richardson, K., Jarde, T., Chambon, P., et al. (2013). Brg1 is required for stem cell maintenance in the murine intestinal epithelium in a tissue-specific manner. Stem Cells 31, 2457–2466.
Holloway, E. M., Czerwinski, M., Tsai, Y. H., Wu, J. H., Wu, A., Childs, C. J., et al. (2021). Mapping Development of the Human Intestinal Niche at Single-Cell Resolution. Cell Stem Cell 28, 568–580.e4.
Horvay, K., and Abud, H. E. (2013). Regulation of intestinal stem cells by Wnt and Notch signalling. Adv. Exp. Med. Biol. 786, 175–186. doi: 10.1007/978-94-007-6621-1_10
Inatomi, O., Andoh, A., Yagi, Y., Bamba, S., Tsujikawa, T., and Fujiyama, Y. (2006). Regulation of amphiregulin and epiregulin expression in human colonic subepithelial myofibroblasts. Int. J. Mol. Med. 18, 497–503.
Iwakura, Y., and Nawa, H. (2013). ErbB1-4-dependent EGF/neuregulin signals and their cross talk in the central nervous system: pathological implications in schizophrenia and Parkinson’s disease. Front. Cell Neurosci. 7:4. doi: 10.3389/fncel.2013.00004
Iwamoto, R., Yamazaki, S., Asakura, M., Takashima, S., Hasuwa, H., Miyado, K., et al. (2003). Heparin-binding EGF-like growth factor and ErbB signaling is essential for heart function. Proc. Natl. Acad. Sci. U.S.A. 100, 3221–3226. doi: 10.1073/pnas.0537588100
Jackson, L. F., Qiu, T. H., Sunnarborg, S. W., Chang, A., Zhang, C., Patterson, C., et al. (2003). Defective valvulogenesis in HB-EGF and TACE-null mice is associated with aberrant BMP signaling. EMBO J. 22, 2704–2716. doi: 10.1093/emboj/cdg264
Jardé, T., Chan, W. H., Rossello, F. J., Kaur Kahlon, T., Theocharous, M., Kurian Arackal, T., et al. (2020). Mesenchymal Niche-Derived Neuregulin-1 Drives Intestinal Stem Cell Proliferation and Regeneration of Damaged Epithelium. Cell Stem Cell 27, 646–662.e7.
Jones, J. C., Brindley, C. D., Elder, N. H., Myers, M. G. Jr., Rajala, M. W., Dekaney, C. M., et al. (2019). Cellular Plasticity of Defa4(Cre)-Expressing Paneth Cells in Response to Notch Activation and Intestinal Injury. Cell. Mol. Gastroenterol. Hepatol. 7, 533–554. doi: 10.1016/j.jcmgh.2018.11.004
Kabiri, Z., Greicius, G., Madan, B., Biechele, S., Zhong, Z., Zaribafzadeh, H., et al. (2014). Stroma provides an intestinal stem cell niche in the absence of epithelial Wnts. Development 141, 2206–2215. doi: 10.1242/dev.104976
Kallincos, N. C., Xian, C. J., Dunbar, A. J., Couper, R. T., and Read, L. C. (2000). Cloning of rat betacellulin and characterization of its expression in the gastrointestinal tract. Growth Factors 18, 203–213. doi: 10.3109/08977190009003245
Kim, J. E., Fei, L., Yin, W. C., Coquenlorge, S., Rao-Bhatia, A., Zhang, X., et al. (2020). Single cell and genetic analyses reveal conserved populations and signaling mechanisms of gastrointestinal stromal niches. Nat. Commun. 11:334.
Kinchen, J., Chen, H. H., Parikh, K., Antanaviciute, A., Jagielowicz, M., Fawkner-Corbett, D., et al. (2018). Structural Remodeling of the Human Colonic Mesenchyme in Inflammatory Bowel Disease. Cell 175, 372–386.e17.
Kramer, R., Bucay, N., Kane, D. J., Martin, L. E., Tarpley, J. E., and Theill, L. E. (1996). Neuregulins with an Ig-like domain are essential for mouse myocardial and neuronal development. Proc. Natl. Acad. Sci. U.S.A. 93, 4833–4838. doi: 10.1073/pnas.93.10.4833
Kraus, M. H., Issing, W., Miki, T., Popescu, N. C., and Aaronson, S. A. (1989). Isolation and characterization of ERBB3, a third member of the ERBB/epidermal growth factor receptor family: evidence for overexpression in a subset of human mammary tumors. Proc. Natl. Acad. Sci. U. S. A. 86, 9193–9197. doi: 10.1073/pnas.86.23.9193
Lee, D., Pearsall, R. S., Das, S., Dey, S. K., Godfrey, V. L., and Threadgill, D. W. (2004). Epiregulin is not essential for development of intestinal tumors but is required for protection from intestinal damage. Mol. Cell Biol. 24, 8907–8916. doi: 10.1128/mcb.24.20.8907-8916.2004
Lee, D., Yu, M., Lee, E., Kim, H., Yang, Y., Kim, K., et al. (2009). Tumor-specific apoptosis caused by deletion of the ERBB3 pseudo-kinase in mouse intestinal epithelium. J. Clin. Invest. 119, 2702–2713. doi: 10.1172/jci36435
Lee, K. F., Simon, H., Chen, H., Bates, B., Hung, M. C., and Hauser, C. (1995). Requirement for neuregulin receptor erbB2 in neural and cardiac development. Nature 378, 394–398. doi: 10.1038/378394a0
Lemmon, M. A., Schlessinger, J., and Ferguson, K. M. (2014). The EGFR family: not so prototypical receptor tyrosine kinases. Cold Spring Harb. Perspect. Biol. 6:a020768. doi: 10.1101/cshperspect.a020768
Luetteke, N. C., Qiu, T. H., Fenton, S. E., Troyer, K. L., Riedel, R. F., Chang, A., et al. (1999). Targeted inactivation of the EGF and amphiregulin genes reveals distinct roles for EGF receptor ligands in mouse mammary gland development. Development 126, 2739–2750. doi: 10.1242/dev.126.12.2739
Luetteke, N. C., Qiu, T. H., Peiffer, R. L., Oliver, P., Smithies, O., and Lee, D. C. (1993). TGF alpha deficiency results in hair follicle and eye abnormalities in targeted and waved-1 mice. Cell 73, 263–278. doi: 10.1016/0092-8674(93)90228-i
McCarthy, N., Kraiczy, J., and Shivdasani, R. A. (2020a). Cellular and molecular architecture of the intestinal stem cell niche. Nat. Cell Biol. 22, 1033–1041. doi: 10.1038/s41556-020-0567-z
McCarthy, N., Manieri, E., Storm, E. E., Saadatpour, A., Luoma, A. M., Kapoor, V. N., et al. (2020b). Distinct Mesenchymal Cell Populations Generate the Essential Intestinal BMP Signaling Gradient. Cell Stem Cell 26, 391–402.e5.
McKenna, K. J., Ligato, S., Kauffman, G. L. Jr., Abt, A. B., Stryker, J. A., and Conter, R. L. (1994). Epidermal growth factor enhances intestinal mitotic activity and DNA content after acute abdominal radiation. Surgery 115, 626–632.
Metcalfe, C., Kljavin, N. M., Ybarra, R., and de Sauvage, F. J. (2014). Lgr5+ stem cells are indispensable for radiation-induced intestinal regeneration. Cell Stem Cell 14, 149–159. doi: 10.1016/j.stem.2013.11.008
Meyer, D., and Birchmeier, C. (1995). Multiple essential functions of neuregulin in development. Nature 378, 386–390. doi: 10.1038/378386a0
Miettinen, P. J., Berger, J. E., Meneses, J., Phung, Y., Pedersen, R. A., Werb, Z., et al. (1995). Epithelial immaturity and multiorgan failure in mice lacking epidermal growth factor receptor. Nature 376, 337–341. doi: 10.1038/376337a0
Mileto, S. J., Jarde, T., Childress, K. O., Jensen, J. L., Rogers, A. P., Kerr, G., et al. (2020). Clostridioides difficile infection damages colonic stem cells via TcdB, impairing epithelial repair and recovery from disease. Proc. Natl. Acad. Sci. U. S. A. 117, 8064–8073. doi: 10.1073/pnas.1915255117
Murata, K., Jadhav, U., Madha, S., van Es, J., Dean, J., Cavazza, A., et al. (2020). Ascl2-Dependent Cell Dedifferentiation Drives Regeneration of Ablated Intestinal Stem Cells. Cell Stem Cell 26, 377–390.e6.
Nikolaev, M., Mitrofanova, O., Broguiere, N., Geraldo, S., Dutta, D., Tabata, Y., et al. (2020). Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature 585, 574–578. doi: 10.1038/s41586-020-2724-8
Pastula, A., Middelhoff, M., Brandtner, A., Tobiasch, M., Hohl, B., Nuber, A. H., et al. (2016). Three-Dimensional gastrointestinal organoid culture in combination with nerves or fibroblasts: a method to characterize the gastrointestinal stem cell niche. Stem Cells Int. 2016:3710836.
Plowman, G. D., Culouscou, J. M., Whitney, G. S., Green, J. M., Carlton, G. W., Foy, L., et al. (1993). Ligand-specific activation of HER4/p180erbB4, a fourth member of the epidermal growth factor receptor family. Proc. Natl. Acad. Sci. U. S. A. 90, 1746–1750. doi: 10.1073/pnas.90.5.1746
Rayego-Mateos, S., Rodrigues-Diez, R., Morgado-Pascual, J. L., Valentijn, F., Valdivielso, J. M., Goldschmeding, R., et al. (2018). Role of Epidermal Growth Factor Receptor (EGFR) and Its Ligands in Kidney Inflammation and Damage. Mediators Inflamm. 2018:8739473.
Riethmacher, D., Sonnenberg-Riethmacher, E., Brinkmann, V., Yamaai, T., Lewin, G. R., and Birchmeier, C. (1997). Severe neuropathies in mice with targeted mutations in the ErbB3 receptor. Nature 389, 725–730. doi: 10.1038/39593
Rogoz, A., Reis, B. S., Karssemeijer, R. A., and Mucida, D. (2015). A 3-D enteroid-based model to study T-cell and epithelial cell interaction. J. Immunol. Methods 421, 89–95. doi: 10.1016/j.jim.2015.03.014
Sato, T., van Es, J. H., Snippert, H. J., Stange, D. E., Vries, R. G., van den Born, M., et al. (2011). Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 469, 415–418. doi: 10.1038/nature09637
Sato, T., Vries, R. G., Snippert, H. J., van de Wetering, M., Barker, N., Stange, D. E., et al. (2009). Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459, 262–265. doi: 10.1038/nature07935
Schechter, A. L., Stern, D. F., Vaidyanathan, L., Decker, S. J., Drebin, J. A., Greene, M. I., et al. (1984). The neu oncogene: an erb-B-related gene encoding a 185,000-Mr tumour antigen. Nature 312, 513–516. doi: 10.1038/312513a0
Schmitt, M., Schewe, M., Sacchetti, A., Feijtel, D., van de Geer, W. S., Teeuwssen, M., et al. (2018). Paneth Cells Respond to Inflammation and Contribute to Tissue Regeneration by Acquiring Stem-like Features through SCF/c-Kit Signaling. Cell Rep. 24, 2312–2328.e7.
Sehgal, A., Donaldson, D. S., Pridans, C., Sauter, K. A., Hume, D. A., and Mabbott, N. A. (2018). The role of CSF1R-dependent macrophages in control of the intestinal stem-cell niche. Nat. Commun. 9:1272.
Semba, K., Kamata, N., Toyoshima, K., and Yamamoto, T. (1985). A v-erbB-related protooncogene, c-erbB-2, is distinct from the c-erbB-1/epidermal growth factor-receptor gene and is amplified in a human salivary gland adenocarcinoma. Proc. Natl. Acad. Sci. U. S. A. 82, 6497–6501. doi: 10.1073/pnas.82.19.6497
Shao, J., and Sheng, H. (2010). Amphiregulin promotes intestinal epithelial regeneration: roles of intestinal subepithelial myofibroblasts. Endocrinology 151, 3728–3737. doi: 10.1210/en.2010-0319
Shirasawa, S., Sugiyama, S., Baba, I., Inokuchi, J., Sekine, S., Ogino, K., et al. (2004). Dermatitis due to epiregulin deficiency and a critical role of epiregulin in immune-related responses of keratinocyte and macrophage. Proc. Natl. Acad. Sci. U.S.A. 101, 13921–13926. doi: 10.1073/pnas.0404217101
Shoshkes-Carmel, M., Wang, Y. J., Wangensteen, K. J., Toth, B., Kondo, A., Massasa, E. E., et al. (2018). Subepithelial telocytes are an important source of Wnts that supports intestinal crypts. Nature 557, 242–246. doi: 10.1038/s41586-018-0084-4
Sibilia, M., and Wagner, E. F. (1995). Strain-dependent epithelial defects in mice lacking the EGF receptor. Science 269, 234–238. doi: 10.1126/science.7618085
Singh, A. B., and Harris, R. C. (2005). Autocrine, paracrine and juxtacrine signaling by EGFR ligands. Cell Signal. 17, 1183–1193. doi: 10.1016/j.cellsig.2005.03.026
Snippert, H. J., van der Flier, L. G., Sato, T., van Es, J. H., van den Born, M., Kroon-Veenboer, C., et al. (2010). Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell 143, 134–144. doi: 10.1016/j.cell.2010.09.016
Sphyris, N., Hodder, M. C., and Sansom, O. J. (2021). Subversion of niche-signalling pathways in colorectal cancer: what makes and breaks the intestinal stem cell. Cancers 13:1000. doi: 10.3390/cancers13051000
Srivatsa, S., Paul, M. C., Cardone, C., Holcmann, M., Amberg, N., Pathria, P., et al. (2017). EGFR in Tumor-Associated Myeloid Cells Promotes Development of Colorectal Cancer in Mice and Associates With Outcomes of Patients. Gastroenterology 153, 178–190.e10.
Suzuki, A., Sekiya, S., Gunshima, E., Fujii, S., and Taniguchi, H. (2010). EGF signaling activates proliferation and blocks apoptosis of mouse and human intestinal stem/progenitor cells in long-term monolayer cell culture. Lab. Invest. 90, 1425–1436. doi: 10.1038/labinvest.2010.150
Talbot, J., Hahn, P., Kroehling, L., Nguyen, H., Li, D., and Littman, D. R. (2020). Feeding-dependent VIP neuron-ILC3 circuit regulates the intestinal barrier. Nature 579, 575–580. doi: 10.1038/s41586-020-2039-9
Taniguchi, K., Wu, L. W., Grivennikov, S. I., de Jong, P. R., Lian, I., Yu, F. X., et al. (2015). A gp130-Src-YAP module links inflammation to epithelial regeneration. Nature 519, 57–62. doi: 10.1038/nature14228
Tetteh, P. W., Basak, O., Farin, H. F., Wiebrands, K., Kretzschmar, K., Begthel, H., et al. (2016). Replacement of Lost Lgr5-Positive Stem Cells through Plasticity of Their Enterocyte-Lineage Daughters. Cell Stem Cell 18, 203–213. doi: 10.1016/j.stem.2016.01.001
Threadgill, D. W., Dlugosz, A. A., Hansen, L. A., Tennenbaum, T., Lichti, U., Yee, D., et al. (1995). Targeted disruption of mouse EGF receptor: effect of genetic background on mutant phenotype. Science 269, 230–234. doi: 10.1126/science.7618084
Tian, H., Biehs, B., Chiu, C., Siebel, C. W., Wu, Y., Costa, M., et al. (2015). Opposing activities of Notch and Wnt signaling regulate intestinal stem cells and gut homeostasis. Cell Rep. 11, 33–42. doi: 10.1016/j.celrep.2015.03.007
Tian, H., Biehs, B., Warming, S., Leong, K. G., Rangell, L., Klein, O. D., et al. (2011). A reserve stem cell population in small intestine renders Lgr5-positive cells dispensable. Nature 478, 255–259. doi: 10.1038/nature10408
Van Landeghem, L., Chevalier, J., Mahe, M. M., Wedel, T., Urvil, P., Derkinderen, P., et al. (2011). Enteric glia promote intestinal mucosal healing via activation of focal adhesion kinase and release of proEGF. Am. J. Physiol. Gastrointest. Liver Physiol. 300, G976–G987. doi: 10.1016/j.jss.2005.11.269
Wang, G. X., Zhao, X. Y., Meng, Z. X., Kern, M., Dietrich, A., Chen, Z., et al. (2014). The brown fat-enriched secreted factor Nrg4 preserves metabolic homeostasis through attenuation of hepatic lipogenesis. Nat. Med. 20, 1436–1443. doi: 10.1038/nm.3713
Wang, Y., Kim, R., Gunasekara, D. B., Reed, M. I., DiSalvo, M., Nguyen, D. L., et al. (2018). Formation of Human Colonic Crypt Array by Application of Chemical Gradients Across a Shaped Epithelial Monolayer. Cell. Mol. Gastroenterol. Hepatol. 5, 113–130. doi: 10.1016/j.jcmgh.2017.10.007
Wu, N., Sun, H., Zhao, X., Zhang, Y., Tan, J., Qi, Y., et al. (2021). MAP3K2-regulated intestinal stromal cells define a distinct stem cell niche. Nature 592, 606–610. doi: 10.1038/s41586-021-03283-y
Xia, G., Rachfal, A. W., Martin, A. E., and Besner, G. E. (2003). Upregulation of endogenous heparin-binding EGF-like growth factor (HB-EGF) expression after intestinal ischemia/reperfusion injury. J. Invest. Surg. 16, 57–63. doi: 10.1080/713829588
Xian, C. J., Mardell, C. E., and Read, L. C. (1999). Specificity of the localization of transforming growth factor-alpha immunoreactivity in colon mucosa. J. Histochem. Cytochem. 47, 949–958. doi: 10.1177/002215549904700712
Yan, L., Shamir, A., Skirzewski, M., Leiva-Salcedo, E., Kwon, O. B., Karavanova, I., et al. (2018). Neuregulin-2 ablation results in dopamine dysregulation and severe behavioral phenotypes relevant to psychiatric disorders. Mol. Psychiatry 23, 1233–1243. doi: 10.1038/mp.2017.22
Yang, Y. P., Ma, H., Starchenko, A., Huh, W. J., Li, W., Hickman, F. E., et al. (2017). A Chimeric Egfr Protein Reporter Mouse Reveals Egfr Localization and Trafficking In Vivo. Cell Rep. 19, 1257–1267. doi: 10.1016/j.celrep.2017.04.048
Yarwood, S. J., and Woodgett, J. R. (2001). Extracellular matrix composition determines the transcriptional response to epidermal growth factor receptor activation. Proc. Natl. Acad. Sci. U. S. A. 98, 4472–4477. doi: 10.1073/pnas.081069098
Yin, X., Farin, H. F., van Es, J. H., Clevers, H., Langer, R., and Karp, J. M. (2014). Niche-independent high-purity cultures of Lgr5+ intestinal stem cells and their progeny. Nat. Methods 11, 106–112. doi: 10.1038/nmeth.2737
Yu, Q., Kilik, U., Holloway, E. M., Tsai, Y. H., Harmel, C., Wu, A., et al. (2021). Charting human development using a multi-endodermal organ atlas and organoid models. Cell 184, 3281–3298.e22.
Yu, S., Tong, K., Zhao, Y., Balasubramanian, I., Yap, G. S., Ferraris, R. P., et al. (2018). Paneth Cell Multipotency Induced by Notch Activation following Injury. Cell Stem Cell 23, 46–59.e5.
Yui, S., Azzolin, L., Maimets, M., Pedersen, M. T., Fordham, R. P., Hansen, S. L., et al. (2018). YAP/TAZ-Dependent Reprogramming of Colonic Epithelium Links ECM Remodeling to Tissue Regeneration. Cell Stem Cell 22, 35–49.e7.
Keywords: Epidermal Growth Factor, neuregulin 1, intestinal stem cells, signaling, niche, ErbB, tissue regeneration, organoids
Citation: Abud HE, Chan WH and Jardé T (2021) Source and Impact of the EGF Family of Ligands on Intestinal Stem Cells. Front. Cell Dev. Biol. 9:685665. doi: 10.3389/fcell.2021.685665
Received: 25 March 2021; Accepted: 27 June 2021;
Published: 19 July 2021.
Edited by:
Kai Kretzschmar, University Hospital Würzburg, GermanyReviewed by:
Hugo Snippert, University Medical Center Utrecht, NetherlandsDelilah Hendriks, Hubrecht Institute (KNAW), Netherlands
Copyright © 2021 Abud, Chan and Jardé. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Helen E. Abud, aGVsZW4uYWJ1ZEBtb25hc2guZWR1; Thierry Jardé, dGhpZXJyeS5qYXJkZUBtb25hc2guZWR1