 Gianluigi Pironti
Gianluigi Pironti Daniel C. Andersson2,3
Daniel C. Andersson2,3
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Cell Dev. Biol., 11 February 2021
Sec. Molecular and Cellular Pathology
Volume 9 - 2021 | https://doi.org/10.3389/fcell.2021.640723
This article is part of the Research TopicExtracellular Vesicles as Next Generation TherapeuticsView all 11 articles
Extracellular vesicles (EVs), which are cell released double layered membrane particles, have been found in every circulating body fluid, and provide a tool for conveying diverse information between cells, influencing both physiological and pathological conditions. Viruses can hijack the EVs secretory pathway to exit infected cells and use EVs endocytic routes to enter uninfected cells, suggesting that EVs and viruses can share common cell entry and biogenesis mechanisms. SARS-CoV-2 is responsible of the coronavirus disease 2019 (Covid-19), which may be accompanied by severe multi-organ manifestations. EVs may contribute to virus spreading via transfer of virus docking receptors such as CD9 and ACE2. Covid-19 is known to affect the renin angiotensin system (RAS), and could promote secretion of harmful EVs. In this scenario EVs might be linked to cardiovascular manifestations of the Covid-19 disease through unbalance in RAS. In contrast EVs derived from mesenchymal stem cells or cardiosphere derived cells, may promote cardiovascular function due to their beneficial effect on angiogenesis, fibrosis, contractility and immuno-modulation. In this article we assessed the potential impact of EVs in cardiovascular manifestations of Covid-19 and highlight potential strategies to control the extracellular signaling for future therapies.
Cells are continuously secreting extracellular vesicles (EVs) that include large apoptotic bodies (1–5 μm), microvesicles (100–1,000 nm) or small exosomes (30–100 nm), which vary in their abundance, size, and composition. Enclosed by a lipid bilayer, EVs are thus very stable and convey cytoplasmic molecules including proteins, RNAs, lipids, and metabolites (Robbins and Morelli, 2014).
Exosomes originate from processing of early endosomes by endosomal sorting complex to form intraluminal vesicles within larger multivesicular bodies (MVB) that traffic in the cytoplasm (Thery et al., 2002). The fusion of MVB to the plasma membrane allows the release of their intraluminal vesicles (now called exosomes) into the extracellular microenvironment (Thery et al., 2002). Initially these vesicles were thought to be secreted by cells merely to eliminate obsolete molecules, but the importance of exosomes in conveying cellular information, with potent autocrine and paracrine biological activities, is now well known as they are released by many different cell types and can be found in most bodily fluids (van Niel et al., 2018).
Microvesicles have size overlap and share some surface markers, such as CD9 and CD63 (Crescitelli et al., 2013; Andreu and Yanez-Mo, 2014) making even more challenging the distinction from exosomes, thus they both are often referred as extracellular vesicles. However, microvesicles have a distinctive biogenesis originating from outward budding of plasma membrane (Kalra et al., 2016). The apoptotic bodies represent the largest EVs and are secreted by cells undergoing programmed cell death, by membrane blebbing and bulging, promoting the clearance of apoptotic material. The intrinsic capacity of EVs in conveying biologic material between proximal and distant cells have been used for therapeutic applications in order to facilitate drug delivery, but it is also used by pathogens to facilitate infection spreading and cause multi-organ diseases. This aspect should be taken in consideration in the perspective of engineering EVs with implemented delivery capacity. In this review we use the term EVs to refer to the two categories microvesicles and exosomes.
Many viruses, including coronaviruses, are known to enter the EVs avenue during synthesis and intra-host spreading, in a caveolin-1-dynamin-dependent mechanism (Owczarek et al., 2018; Badierah et al., 2020). Whether the SARS-CoV-2 virus, which causes the coronavirus disease 2019 (Covid-19) is involved in EVs trafficking is still unknown. However, in vitro data show that alveolar cells infected with SARS-CoV-1 present whole virions in the secretory vesicles near the plasma membrane of infected cells (Qian et al., 2013; Mason, 2020). Whether SARS-CoV-2 virions also can accumulate in secreting vesicles has not been validated yet, although electron microscopy images of lung biopsy in autopsy specimen from deceased Covid-19 patient intriguingly show SARS-CoV-2 virion within double membrane vesicles (Dittmayer et al., 2020). Moreover, histopathological analysis of renal specimen from deceased patients with Covid-19 showed that SARS-CoV-2 virions are mostly found in the cytoplasm and some vacuoles containing assembling virions similar to what has been seen with SARS-CoV-1 (Farkash et al., 2020; Su et al., 2020). Due to the size of coronavirus (60–140 nm) it is possible that bigger EVs such as microparticles might mediate whole virion transport, while exosomes might carry viral fragments. During an infection, big apoptotic bodies released from infected/injured cells might be an easy way to carry virions due to the larger size, however apoptotic bodies present surface bridging molecules that triggers phagocytosis, which could contribute to contain rather than spreading SARS-CoV-2 in the body (Arandjelovic and Ravichandran, 2015). However, exosomes and microparticles contain virus receptors that make EVs recipient cells susceptible to virus entry (Earnest et al., 2015). Indeed tetraspanin protein CD9 expressed by EVs formed cell membrane complex of the co-receptor dipeptidyl peptidase 4 (DPP4) and the type II transmembrane serine protease member TMPRSS2, a CoV-activating protease, facilitating proteolytic priming events with cells for MERS-CoV pseudovirus, which is necessary for virus cell entry and can be blocked by CD9 inhibition (Earnest et al., 2015). Similarly, SARS-CoV-2 uses DPP4 (Li Y. et al., 2020; Solerte et al., 2020) and TMPRSS2 (Hoffmann et al., 2020) to facilitate cell entry. Although it might be reasonable to think that also SARS-CoV-2 virulence relies on CD9 activity by clustering and scaffolding receptors and proteases for efficient cell entry, this hypothesis has not been validated yet in current literature.
Thus, coronavirus such as SARS-CoV-1 and MERS-CoV may be directed into the exosomal pathway or released from microvesicles, upon intracellular entry, and its components, such as mRNAs and proteins, including spike protein (Kuate et al., 2007), are packaged into EVs for secretion (Hassanpour et al., 2020).
Although conventional biochemistry techniques allow the detection of viral components such as viral RNA or spike proteins, there is currently no empirical evidence that EVs secreted from infected cells contain specific fragments of SARS-CoV-2. EM images potentially would be a method to investigate this, but purification techniques available at the moment cannot separate virions from EVs due to their overlapping size. Therefore, the isolation procedures need to be implemented in order to identify specific viral components (RNA or proteins) carried by EVs. Whether EVs released from infected cells have harmful or protective roles is controversial: vesicles containing only viral fragments might stimulate the immune system for their antigen priming activity, while the whole virion hidden in bigger vesicles could potentially elicit viral dissemination by escaping the immune recognition and further spread the virus within the body.
In addition to CD9, it has been shown that exosomes can transfer the angiotensin converting enzyme 2 (ACE2) receptor (Wang et al., 2020), which SARS-CoV-2 uses for cell entry. It is reasonable to believe that increased levels of ACE2 might promote more virus entry similarly to what has been observed in cellular models transfected with human ACE2 (Blanco-Melo et al., 2020; Ou et al., 2020). Alternatively EVs containing ACE2 might be used to decoy the SARS-CoV-2 and limit virus spreading (Inal, 2020). The presence of docking station for virus priming such as ACE2 is necessary but not sufficient for virus entry as the proteases activity of TMPRSS2 is critical for fusion and internalization of ACE-2/SARS-CoV2 spike complex (Hoffmann et al., 2020), and co-receptors such as neuropilin-1 (NRP-1) can significantly potentiate SARS-CoV-2 infectivity (Cantuti-Castelvetri et al., 2020). Both NRP-1 and TMPRSS2 have been found in EVs (Munshi et al., 2019; Chi et al., 2020). As the inter-cellular trafficking pathways of viruses and EVs are similar or shared, pharmacological inhibition of EVs trafficking has been proposed as an important antiviral approach in order to limit the systemic spreading of SARS-CoV-2 (Urciuoli and Peruzzi, 2020).
It has been reported that during SARS-CoV-1 and MERS-CoV infections there were increased levels of circulating exosomes enriched in lung-associated self-antigens as well as viral antigens (Gunasekaran et al., 2020). Thus, it might be possible that SARS-CoV-2 infected cells increase the production of exosomes enriched in viral components. In this way, exosomes containing pathogenic proteins and RNA released from infected cells may induce host humoral and cellular immune response (Gunasekaran et al., 2017) and could potentially play anti-infective roles (Zhang et al., 2018). In contrast, it is conceivable that viral material might also be hidden within EVs vesicles using a sort of “trojan horse” strategy for immune system evasion, which might potentially be linked to the variable detection of viral RNA in testing patients with ongoing, or recovered, Covid-19 (Elrashdy et al., 2020). Although different reasons (including methodological problems) may be underlying false negative PCR tests for viral detection, it is intriguing to speculate that EVs could be a hiding place for viral material.
In the progression of chronic heart failure, compensatory but maladaptive renin-angiotensin system (RAS) activation promotes cardiac remodeling (Zhou et al., 2016). Current literature describes a modulatory role of EVs in RAS through intercellular trafficking by exosomes. It has been shown that increased angiotensin type II (AngII) levels in heart failure, elicit secretion of exosomes by cardiac fibroblasts, which induces neighboring cardiomyocytes to release more AngII and express AngII receptors (AT1R and AT2R) through a feedback process that promotes myocardial hypertrophy (Lyu et al., 2015). Moreover, increased AngII in plasma and cardiac pressure overload leads to release of exosomes enriched in AT1R, which target resistance vessels and cardiomyocytes, aggravating cardiac remodeling (Pironti et al., 2015). Post-mortem autopsy analysis showed that almost 35% of patients who succumbed to SARS-CoV-1 (in 2003) presented detectable viral SARS-CoV-1 genome in heart samples, and this was associated with myocardial fibrosis, inflammation, and reduced myocardial ACE2 expression (Oudit et al., 2009). SARS-CoV-2 viral particles have been found in cardiac biopsies of Covid-19 patients (Dolhnikoff et al., 2020; Tavazzi et al., 2020), and it has been reported that this virus can infect human cardiomyocytes and induce cardiotoxicity (Bojkova et al., 2020).
Both SARS-CoV-1 and SARS-CoV-2 uses ACE2 as docking station to infect target cells and the internalization of virus/enzyme complex leads to loss of enzymatically active ACE2 at the cell surface (Gheblawi et al., 2020; Hoffmann et al., 2020).
As RAS-inhibitors can upregulate expression of ACE2, there were initial concerns that use of RAS-antagonists may increase risk or severity of Covid-19 as more docking stations would theoretically be present. However, the use of RAS inhibitor therapy was not associated with increased infectivity or severity of disease (Li J. et al., 2020; Zhang et al., 2020).
As cells are infected by SARS-CoV-2, the ACE2 expression is down-regulated due to virus/enzyme complex internalization. The reduction of ACE2 enzymatic activity results in an imbalance within the RAS, which stimulates neutrophil infiltration, unopposed AngII accumulation and may promote acute lung injury. Thus, RAS-inhibitors may also be protective. Less attention has been given to the fact that increased circulating AngII levels could elicit the endocytosis of ACE-2 bound to SARS-CoV-2 via a AT1R-dependent mechanism (Offringa et al., 2020), which may promote ACE2 degradation (Deshotels et al., 2014). Furthermore, reduced expression levels of ACE2 in the heart, following SARS-CoV-2 infection, may impair the conversion of AngII to the Ang1-7 heptapeptide, which is anti-inflammatory, anti-fibrotic, and cardioprotective (Patel et al., 2014) and represents the main mediators of the protective RAS signaling activating via AT2R and MasR (Santos et al., 2003). Thus, in addition to downregulating ACE2 and Ang1-7, unbalanced RAS activation through exosomes during Covid-19 may represent a novel detrimental pathway that could be responsible for some of the cardiovascular disease manifestations in Covid-19 (Figure 1). Indeed in a yet ongoing promising clinical trial, a non-peptide AT2R agonist is used as therapy to counterbalance the potentially harmful RAS effects during Covid-19 (Angiotensin II Type Two Receptor Agonist in COVID-19 Trial ATTRACT study, NCT04452435) (Steckelings and Sumners, 2020).
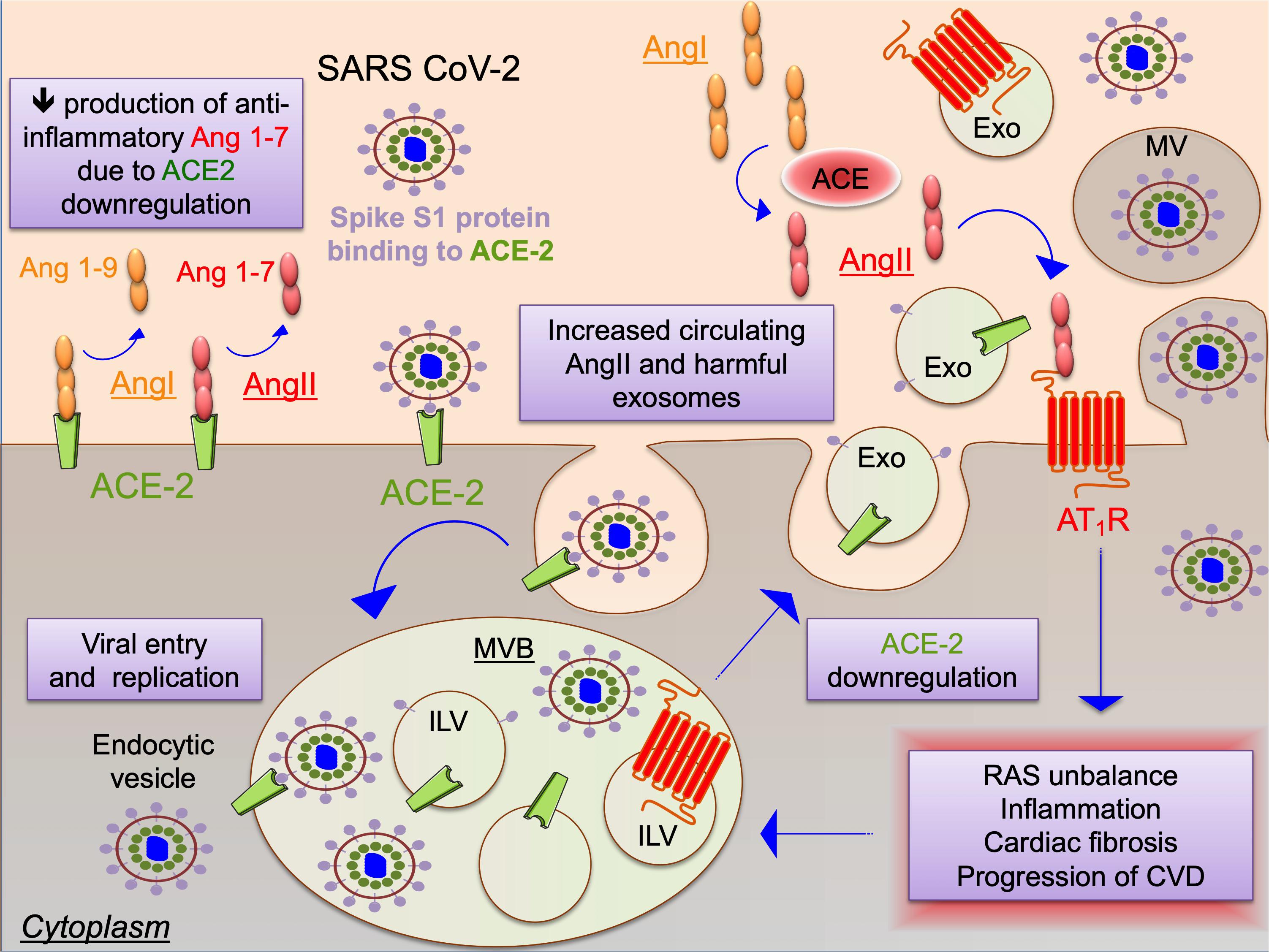
Figure 1. Schematic representation of the potential interplay between the renin-angiotensin system (RAS) and SARS-CoV-2 as mediated by extracellular vesicles. SARS coronaviruses use spike protein S1 to bind ACE-2 as docking station leading to cell entry and also to ACE2 down-regulation. Increased levels of angiotensin type II (AngII) aggravate RAS, promoting inflammation, cardiac fibrosis and progression of cardiovascular diseases (CVD). SARS-CoV-2 infection and RAS trigger the endocytic machinery, leading to internalization of ACE2, virion components and angiotentin II type 1 receptor (AT1R) into intraluminal vesicles (ILV) within multivesicular bodies (MVB), which are released as exosomes (Exo) upon fusion of the MVB with the cell plasma membrane. SARS-CoV-2 spreading may be enhanced also through release of virions in larger microvesicles (MV) upon plasma membrane budding of infected cells. Exosomes or other EVs might be responsible for manifestations and progression of cardiovascular disease in Covid-19 through maladaptive RAS signaling.
Although EVs release as results of RAS disturbance during SARS-CoV-2 infection has not yet been demonstrated, we here provide a provocative hypothesis of a link between EVs, Covid-19 and RAS based on two demonstrated evidences: (i) SARS-CoV-2 infection causes disturbance of RAS; (ii) RAS-dependent release of exosomes in e.g., hypertension are known to aggravate cardiovascular diseases and myocardial remodeling.
Severe Covid-19 display a systemic pro-inflammatory state often referred to as a cytokine storm with critical illness and manifestations such as acute respiratory distress syndrome (ARDS), pulmonary edemea, thromboembolism, and acute cardiac injury, which come with high mortality (Huang et al., 2020; Leisman et al., 2020).
Mesenchymal stem cells (MSCs) have powerful immuno-modulatory effects and are considered potentially effective therapeutic assets to combat a wide spectrum of inflammatory-driven diseases but also clinically useful in preventing inflammatory activation from acute graft vs. host disease following allogenic hematopoietic stem-cell transplantation (Hashmi et al., 2016; Bazzoni et al., 2020). Thus, MSCs might be used to prevent or reduce the cytokine storm caused by SARS-CoV-2. Interestingly, EVs released from MSCs is one of the mechanisms whereby MSCs may promote anti-inflammatory effects (Hashmi et al., 2016; Bazzoni et al., 2020). Indeed, several studies have described beneficial effect of MSCs-derived EVs following myocardial injury by increasing angiogenesis (Liang et al., 2016; Gong et al., 2017) improving cardiac contractility (Kang et al., 2015; Ma et al., 2017; Mayourian et al., 2017). Moreover stem cells derived from cardiac tissues, i.e., cardiosphere derived cells (CDC), produce EVs able to reduce inflammation and fibrosis signaling (Gray et al., 2015; Gallet et al., 2017; Rogers et al., 2020). The intercellular communication between macrophages, fibroblasts and endothelial cells orchestrates the progression of chronic heart failure and elicit myocardial fibrosis. Thus, in the context of anti-fibrogenesis, EVs may have a beneficial effect by promoting the trans-differentiation of macrophages from pro-inflammatory M1 into M2-like cells with immunosuppressive phenotype (Silva et al., 2017). This process is important for the resolution of inflammation and might be particularly relevant in the context of therapy meant to combat the systemic pro-inflammatory state in Covid-19.
Interferon gamma (INF-γ) is a major proinflammatory cytokine secreted by activated T cells and natural killer cells, which promotes macrophage activation and mediate host defense against pathogen infection. Decreased plasma levels of INF-γ in the early stage of the disease (before the onset of cytokine storm) were associated with increased risk of pulmonary fibrosis in Covid-19 patients (Hu et al., 2020). Indeed INF-γ has antiviral action, through enhancing the viral antigen processing and presentation to cytotoxic T lymphocytes for virus clearance, and has also anti-fibrotic effect. Thus early intervention of anti-viral infection using IFN-γ could be significant in the inhibition of fibrosis for better functional recovery (Hu et al., 2020). Stem cells exposed to proinflammatory cytokine signaling released EVs enriched in mRNA coding for INF-γ signaling pathway, such as STAT1, JAK 1-2 (Cossetti et al., 2014). Interestingly EVs can recycle and deliver free INF-γ and by that, promoting the continuation of pro-inflammatory signaling response in target cells (Cossetti et al., 2014).
Therapeutic approaches for Covid-19 based on MSCs-derived EVs aimed to enhancing the healing process following lung injury, have been taken in consideration (Akbari and Rezaie, 2020; Al-Khawaga and Abdelalim, 2020; Pinky et al., 2020; Tsuchiya et al., 2020). However, experimental evidences of their mechanism of action, and whether MSCs-derived EVs therapy can ameliorate cardiovascular manifestations have not been elucidated. Interestingly, the first clinical trial using MSC derived exosomes that were intravenously injected in Covid-19 patients with moderate-to-severe symptoms (Sengupta et al., 2020) (NCT04276987) showed positive results in terms of safety profile, capacity to restore oxygenation, downregulation of cytokines, and reconstitute immunity. In another clinical trial (NCT04276987), inhalation of aerosol enriched in allogenic EVs derived from adipose MSC have been tested for safety and efficacy in Covid-19 patients with severe symptoms (Pocsfalvi et al., 2020).
The timing of an intervention seems to be a critical aspect in Covid-19 disease. The acute phase of Covid-19 illness is characterized by three pathological phases: (i) the early infection phase when SARS-CoV-2 infiltrates and replicate; (ii) the pulmonary phase characterized by respiratory dysfunction; (iii) finally the hyperinflammation phase driven by the host immunity with an exaggerate inflammatory response (Akhmerov and Marban, 2020). The specific stage of disease must be taken into consideration when commencing different therapeutic interventions e.g., with antiviral, immunopotentiating or anti-inflammatory properties (de Simone and Mancusi, 2020). Although each phase can be defined based on clinical and laboratory findings the preciseness of this diagnosing is not complete thus providing a major challenge for timing of intervention. A too early anti-inflammatory intervention might hinder the viral elimination by host immunity, while a too late anti-inflammatory therapy might have a slight efficacy.
Although EVs containing viral components might elicit the adaptive immune response by presenting the antigen in a process similar to a vaccination, EVs secreted from infected/injured cells could activate macrophage secreting cytokines and contribute to the cytokine storm formation during Covid-19 (Perez et al., 2019). Indeed, EVs can carry pathogen-associated molecular patterns (PAMPs) released from stressed or injured cells and therefore triggering inflammation upon interaction with innate immune cells contributing to inflammation induction and persistence (Bhatnagar et al., 2007). Additionally, EVs containing cytokines (Pizzirani et al., 2007), enzymes involved in the biosynthesis of lipid mediators (Cossetti et al., 2014) and other chemotactic signals (Esser et al., 2010; Kriebel et al., 2018) can contribute to the propagation of inflammation. Interestingly, ACE2 enriched EVs derived from engineered MSC has been considered as a possible Covid-19 therapy in order to decoy the SARS-CoV-2 and limit virus spreading (Inal, 2020).
Thus, a therapeutic application of EVs for Covid-19 must include donor cells with anti-inflammatory activity such as mesenchymal stem cells or cardiosphere derived cells. The anti-inflammatory activity of EVs indicate that this platform can work well in xenogenic applications (de Couto et al., 2017; Gallet et al., 2017), which could simplify the large scale production of EVs for therapeutic applications by using MSCs or CDC derived from large animals rather than humans.
We have discussed the possible function of EVs in Covid-19 and highlight potential therapeutic application for EVs in modulating the virus infection, the pro-inflammatory state and cardiovascular disease in Covid-19. EVs, such as exosomes could be involved in aggravated cardiovascular manifestations of Covid-19 either by promoting viral docking in cardiac cells or shifting RAS to promote inflammation, coagulation, fibrosis and endothelial dysfunction. Furthermore, EVs derived from engineered MSCs or CDC used as drug delivery system to modulate the cytokine storm of virus spreading represent a promising strategy to combat Covid-19. In summary, understanding the molecular mechanisms behind the entry, replication, and spreading of SARS-CoV-2, in conjunction to EVs trafficking, may provide tools to limit the manifestation of Covid-19.
GP substantially contributed to the conception and design of the perspective article, literature search, drafting the article and revising the article critically for important intellectual content. DA and LL contributed in literature search, drafting the article and revising the article critically for important intellectual content. All the authors approved the final version of the article to be published.
LL and GP were funded by grants from the Swedish Heart-Lung Foundation (20150557) and the Swedish Research Council (523-2014-2336). DA was funded by the Swedish Heart-Lung Foundation (20180637 and 2016074), the Swedish Society for Medical research (SSMF and S16-0159).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Akbari, A., and Rezaie, J. (2020). Potential therapeutic application of mesenchymal stem cell-derived exosomes in SARS-CoV-2 pneumonia. Stem Cell Res. Ther. 11:356.
Al-Khawaga, S., and Abdelalim, E. M. (2020). Potential application of mesenchymal stem cells and their exosomes in lung injury: an emerging therapeutic option for COVID-19 patients. Stem Cell Res. Ther. 11:437.
Andreu, Z., and Yanez-Mo, M. (2014). Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 5:442.
Arandjelovic, S., and Ravichandran, K. S. (2015). Phagocytosis of apoptotic cells in homeostasis. Nat. Immunol. 16, 907–917. doi: 10.1038/ni.3253
Badierah, R. A., Uversky, V. N., and Redwan, E. M. (2020). Dancing with Trojan horses: an interplay between the extracellular vesicles and viruses. J. Biomol. Struct. Dyn. doi: 10.1080/07391102.2020.1756409 Online ahead of print.
Bazzoni, R., Takam Kamga, P., Tanasi, I., and Krampera, M. (2020). Extracellular vesicle-dependent communication between mesenchymal stromal cells and immune effector cells. Front. Cell Dev. Biol. 8:596079.
Bhatnagar, S., Shinagawa, K., Castellino, F. J., and Schorey, J. S. (2007). Exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and in vivo. Blood 110, 3234–3244. doi: 10.1182/blood-2007-03-079152
Blanco-Melo, D., Nilsson-Payant, B. E., Liu, W. C., Uhl, S., Hoagland, D., Moller, R., et al. (2020). Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 181, 1036–1045.e1039.
Bojkova, D., Wagner, J. U. G., Shumliakivska, M., Aslan, G. S., Saleem, U., Hansen, A., et al. (2020). SARS-CoV-2 infects and induces cytotoxic effects in human cardiomyocytes. Cardiovasc. Res. 116, 2207–2215. doi: 10.1093/cvr/cvaa267
Cantuti-Castelvetri, L., Ojha, R., Pedro, L. D., Djannatian, M., Franz, J., Kuivanen, S., et al. (2020). Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 370, 856–860.
Chi, M., Shi, X., Huo, X., Wu, X., Zhang, P., and Wang, G. (2020). Dexmedetomidine promotes breast cancer cell migration through Rab11-mediated secretion of exosomal TMPRSS2. Ann. Transl. Med. 8:531. doi: 10.21037/atm.2020.04.28
Cossetti, C., Iraci, N., Mercer, T. R., Leonardi, T., Alpi, E., Drago, D., et al. (2014). Extracellular vesicles from neural stem cells transfer IFN-gamma via Ifngr1 to activate Stat1 signaling in target cells. Mol. Cell. 56, 193–204. doi: 10.1016/j.molcel.2014.08.020
Crescitelli, R., Lasser, C., Szabo, T. G., Kittel, A., Eldh, M., Dianzani, I., et al. (2013). Distinct RNA profiles in subpopulations of extracellular vesicles: apoptotic bodies, microvesicles and exosomes. J. Extracell. Vesicles. 2:20677.
de Couto, G., Gallet, R., Cambier, L., Jaghatspanyan, E., Makkar, N., Dawkins, J. F., et al. (2017). Exosomal MicroRNA transfer into macrophages mediates cellular postconditioning. Circulation 136, 200–214. doi: 10.1161/circulationaha.116.024590
de Simone, G., and Mancusi, C. (2020). Finding the right time for anti-inflammatory therapy in COVID-19. Int. J. Infect. Dis. 101, 247–248. doi: 10.1016/j.ijid.2020.09.1454
Deshotels, M. R., Xia, H., Sriramula, S., Lazartigues, E., and Filipeanu, C. M. (2014). Angiotensin II mediates angiotensin converting enzyme type 2 internalization and degradation through an angiotensin II type I receptor-dependent mechanism. Hypertension 64, 1368–1375. doi: 10.1161/hypertensionaha.114.03743
Dittmayer, C., Meinhardt, J., Radbruch, H., Radke, J., Heppner, B. I., Heppner, F. L., et al. (2020). Why misinterpretation of electron micrographs in SARS-CoV-2-infected tissue goes viral. Lancet 396, e64–e65.
Dolhnikoff, M., Ferreira Ferranti, J., De Almeida Monteiro, R. A., Duarte-Neto, A. N., Soares Gomes-Gouvea, M., Viu Degaspare, N., et al. (2020). SARS-CoV-2 in cardiac tissue of a child with COVID-19-related multisystem inflammatory syndrome. Lancet Child Adolesc. Health 4, 790–794.
Earnest, J. T., Hantak, M. P., Park, J. E., and Gallagher, T. (2015). Coronavirus and influenza virus proteolytic priming takes place in tetraspanin-enriched membrane microdomains. J. Virol. 89, 6093–6104. doi: 10.1128/jvi.00543-15
Elrashdy, F., Aljaddawi, A. A., Redwan, E. M., and Uversky, V. N. (2020). On the potential role of exosomes in the COVID-19 reinfection/reactivation opportunity. J. Biomol. Struct. Dyn. doi: 10.1080/07391102.2020.1790426 Online ahead of print.
Esser, J., Gehrmann, U., D’alexandri, F. L., Hidalgo-Estevez, A. M., Wheelock, C. E., Scheynius, A., et al. (2010). Exosomes from human macrophages and dendritic cells contain enzymes for leukotriene biosynthesis and promote granulocyte migration. J. Allergy Clin. Immunol. 126, 1032–1040. doi: 10.1016/j.jaci.2010.06.039
Farkash, E. A., Wilson, A. M., and Jentzen, J. M. (2020). Ultrastructural evidence for direct renal infection with SARS-CoV-2. J. Am. Soc. Nephrol. 31, 1683–1687. doi: 10.1681/asn.2020040432
Gallet, R., Dawkins, J., Valle, J., Simsolo, E., De Couto, G., Middleton, R., et al. (2017). Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 38, 201–211.
Gheblawi, M., Wang, K., Viveiros, A., Nguyen, Q., Zhong, J. C., Turner, A. J., et al. (2020). Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: celebrating the 20th anniversary of the discovery of ACE2. Circ. Res. 126, 1456–1474. doi: 10.1161/circresaha.120.317015
Gong, M., Yu, B., Wang, J., Wang, Y., Liu, M., Paul, C., et al. (2017). Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget 8, 45200–45212. doi: 10.18632/oncotarget.16778
Gray, W. D., French, K. M., Ghosh-Choudhary, S., Maxwell, J. T., Brown, M. E., Platt, M. O., et al. (2015). Identification of therapeutic covariant microRNA clusters in hypoxia-treated cardiac progenitor cell exosomes using systems biology. Circ. Res. 116, 255–263. doi: 10.1161/circresaha.116.304360
Gunasekaran, M., Bansal, S., Ravichandran, R., Sharma, M., Perincheri, S., Rodriguez, F., et al. (2020). Respiratory viral infection in lung transplantation induces exosomes that trigger chronic rejection. J. Heart Lung Transplant. 39, 379–388. doi: 10.1016/j.healun.2019.12.009
Gunasekaran, M., Xu, Z., Nayak, D. K., Sharma, M., Hachem, R., Walia, R., et al. (2017). Donor-derived exosomes with lung self-antigens in human lung allograft rejection. Am. J. Transplant. 17, 474–484. doi: 10.1111/ajt.13915
Hashmi, S., Ahmed, M., Murad, M. H., Litzow, M. R., Adams, R. H., Ball, L. M., et al. (2016). Survival after mesenchymal stromal cell therapy in steroid-refractory acute graft-versus-host disease: systematic review and meta-analysis. Lancet Haematol. 3, e45–e52.
Hassanpour, M., Rezaie, J., Nouri, M., and Panahi, Y. (2020). The role of extracellular vesicles in COVID-19 virus infection. Infect. Genet. Evol. 85:104422. doi: 10.1016/j.meegid.2020.104422
Hoffmann, M., Kleine-Weber, H., Schroeder, S., Kruger, N., Herrler, T., Erichsen, S., et al. (2020). SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280.e278.
Hu, Z. J., Xu, J., Yin, J. M., Li, L., Hou, W., Zhang, L. L., et al. (2020). Lower circulating interferon-gamma is a risk factor for lung fibrosis in COVID-19 patients. Front. Immunol. 11:585647.
Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., et al. (2020). Clinical features of patients infected with 2019 novel coronavirus in Wuhan. China. Lancet 395, 497–506.
Inal, J. M. (2020). Decoy ACE2-expressing extracellular vesicles that competitively bind SARS-CoV-2 as a possible COVID-19 therapy. Clin. Sci. 134, 1301–1304. doi: 10.1042/cs20200623
Kalra, H., Drummen, G. P., and Mathivanan, S. (2016). Focus on extracellular vesicles: introducing the next small big thing. Int. J. Mol. Sci. 17:170. doi: 10.3390/ijms17020170
Kang, K., Ma, R., Cai, W., Huang, W., Paul, C., Liang, J., et al. (2015). Exosomes secreted from CXCR4 overexpressing mesenchymal stem cells promote cardioprotection via Akt signaling pathway following myocardial infarction. Stem Cells Int. 2015:659890.
Kriebel, P. W., Majumdar, R., Jenkins, L. M., Senoo, H., Wang, W., Ammu, S., et al. (2018). Extracellular vesicles direct migration by synthesizing and releasing chemotactic signals. J. Cell Biol. 217, 2891–2910. doi: 10.1083/jcb.201710170
Kuate, S., Cinatl, J., Doerr, H. W., and Uberla, K. (2007). Exosomal vaccines containing the S protein of the SARS coronavirus induce high levels of neutralizing antibodies. Virology 362, 26–37. doi: 10.1016/j.virol.2006.12.011
Leisman, D. E., Ronner, L., Pinotti, R., Taylor, M. D., Sinha, P., Calfee, C. S., et al. (2020). Cytokine elevation in severe and critical COVID-19: a rapid systematic review, meta-analysis, and comparison with other inflammatory syndromes. Lancet Respir. Med. 8, 1233–1244. doi: 10.1016/s2213-2600(20)30404-5
Li, J., Wang, X., Chen, J., Zhang, H., and Deng, A. (2020). Association of renin-angiotensin system inhibitors with severity or risk of death in patients with hypertension hospitalized for coronavirus disease 2019 (COVID-19) infection in Wuhan. China. JAMA Cardiol. 5, 825–830. doi: 10.1001/jamacardio.2020.1624
Li, Y., Zhang, Z., Yang, L., Lian, X., Xie, Y., Li, S., et al. (2020). The MERS-CoV receptor DPP4 as a candidate binding target of the SARS-CoV-2 Spike. iScience 23:101160. doi: 10.1016/j.isci.2020.101160
Liang, X., Zhang, L., Wang, S., Han, Q., and Zhao, R. C. (2016). Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 129, 2182–2189. doi: 10.1242/jcs.170373
Lyu, L., Wang, H., Li, B., Qin, Q., Qi, L., Nagarkatti, M., et al. (2015). A critical role of cardiac fibroblast-derived exosomes in activating renin angiotensin system in cardiomyocytes. J. Mol. Cell Cardiol. 89, 268–279. doi: 10.1016/j.yjmcc.2015.10.022
Ma, J., Zhao, Y., Sun, L., Sun, X., Zhao, X., Sun, X., et al. (2017). Exosomes derived from akt-modified human umbilical cord mesenchymal stem cells improve cardiac regeneration and promote angiogenesis via activating platelet-derived growth factor D. Stem Cells Transl. Med. 6, 51–59. doi: 10.5966/sctm.2016-0038
Mason, R. J. (2020). Pathogenesis of COVID-19 from a cell biology perspective. Eur. Respir. J. 55:2000607. doi: 10.1183/13993003.00607-2020
Mayourian, J., Cashman, T. J., Ceholski, D. K., Johnson, B. V., Sachs, D., Kaji, D. A., et al. (2017). Experimental and computational insight into human mesenchymal stem cell paracrine signaling and heterocellular coupling effects on cardiac contractility and arrhythmogenicity. Circ. Res. 121, 411–423. doi: 10.1161/circresaha.117.310796
Munshi, A., Mehic, J., Creskey, M., Gobin, J., Gao, J., Rigg, E., et al. (2019). A comprehensive proteomics profiling identifies NRP1 as a novel identity marker of human bone marrow mesenchymal stromal cell-derived small extracellular vesicles. Stem Cell Res. Ther. 10:401.
Offringa, A., Montijn, R., Singh, S., Paul, M., Pinto, Y. M., and Pinto-Sietsma, S. J. (2020). The mechanistic overview of SARS-CoV-2 using angiotensin-converting enzyme 2 to enter the cell for replication: possible treatment options related to the renin-angiotensin system. Eur. Heart J. Cardiovasc. Pharmacother. 6, 317–325. doi: 10.1093/ehjcvp/pvaa053
Ou, X., Liu, Y., Lei, X., Li, P., Mi, D., Ren, L., et al. (2020). Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 11:1620.
Oudit, G. Y., Kassiri, Z., Jiang, C., Liu, P. P., Poutanen, S. M., Penninger, J. M., et al. (2009). SARS-coronavirus modulation of myocardial ACE2 expression and inflammation in patients with SARS. Eur. J. Clin. Invest. 39, 618–625. doi: 10.1111/j.1365-2362.2009.02153.x
Owczarek, K., Szczepanski, A., Milewska, A., Baster, Z., Rajfur, Z., Sarna, M., et al. (2018). Early events during human coronavirus OC43 entry to the cell. Sci. Rep. 8:7124.
Patel, V. B., Clarke, N., Wang, Z., Fan, D., Parajuli, N., Basu, R., et al. (2014). Angiotensin II induced proteolytic cleavage of myocardial ACE2 is mediated by TACE/ADAM-17: a positive feedback mechanism in the RAS. J. Mol. Cell Cardiol. 66, 167–176. doi: 10.1016/j.yjmcc.2013.11.017
Perez, P. S., Romaniuk, M. A., Duette, G. A., Zhao, Z., Huang, Y., Martin-Jaular, L., et al. (2019). Extracellular vesicles and chronic inflammation during HIV infection. J. Extracell. Vesicles 8:1687275. doi: 10.1080/20013078.2019.1687275
Pinky, G. S., Krishnakumar, V., Sharma, Y., Dinda, A. K., and Mohanty, S. (2020). Mesenchymal stem cell derived exosomes: a nano platform for therapeutics and drug delivery in combating COVID-19. Stem Cell Rev. Rep. doi: 10.1007/s12015-020-10002-z Online ahead of print.
Pironti, G., Strachan, R. T., Abraham, D., Mon-Wei, Yu, S., Chen, M., et al. (2015). Circulating exosomes induced by cardiac pressure overload contain functional angiotensin II Type 1 receptors. Circulation 131, 2120–2130. doi: 10.1161/circulationaha.115.015687
Pizzirani, C., Ferrari, D., Chiozzi, P., Adinolfi, E., Sandona, D., Savaglio, E., et al. (2007). Stimulation of P2 receptors causes release of IL-1beta-loaded microvesicles from human dendritic cells. Blood 109, 3856–3864. doi: 10.1182/blood-2005-06-031377
Pocsfalvi, G., Mammadova, R., Ramos Juarez, A. P., Bokka, R., Trepiccione, F., and Capasso, G. (2020). COVID-19 and extracellular vesicles: an intriguing interplay. Kidney Blood Press. Res. 45, 661–670. doi: 10.1159/000511402
Qian, Z., Travanty, E. A., Oko, L., Edeen, K., Berglund, A., Wang, J., et al. (2013). Innate immune response of human alveolar type II cells infected with severe acute respiratory syndrome-coronavirus. Am. J. Respir. Cell Mol. Biol. 48, 742–748. doi: 10.1165/rcmb.2012-0339oc
Robbins, P. D., and Morelli, A. E. (2014). Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 14, 195–208. doi: 10.1038/nri3622
Rogers, R. G., Ciullo, A., Marban, E., and Ibrahim, A. G. (2020). Extracellular vesicles as therapeutic agents for cardiac fibrosis. Front. Physiol. 11:479.
Santos, R. A., Simoes, E., Silva, A. C., Maric, C., Silva, D. M., Machado, R. P., et al. (2003). Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc. Natl. Acad. Sci. U.S.A. 100, 8258–8263.
Sengupta, V., Sengupta, S., Lazo, A., Woods, P., Nolan, A., and Bremer, N. (2020). Exosomes derived from bone marrow mesenchymal stem cells as treatment for severe COVID-19. Stem Cells Dev. 29, 747–754. doi: 10.1089/scd.2020.0080
Silva, A. M., Teixeira, J. H., Almeida, M. I., Goncalves, R. M., Barbosa, M. A., and Santos, S. G. (2017). Extracellular vesicles: immunomodulatory messengers in the context of tissue repair/regeneration. Eur. J. Pharm. Sci. 98, 86–95. doi: 10.1016/j.ejps.2016.09.017
Solerte, S. B., Di Sabatino, A., Galli, M., and Fiorina, P. (2020). Dipeptidyl peptidase-4 (DPP4) inhibition in COVID-19. Acta Diabetol. 57, 779–783. doi: 10.1007/s00592-020-01539-z
Steckelings, U. M., and Sumners, C. (2020). Correcting the imbalanced protective RAS in COVID-19 with angiotensin AT2-receptor agonists. Clin. Sci. 134, 2987–3006. doi: 10.1042/cs20200922
Su, H., Yang, M., Wan, C., Yi, L. X., Tang, F., Zhu, H. Y., et al. (2020). Renal histopathological analysis of 26 postmortem findings of patients with COVID-19 in China. Kidney Int. 98, 219–227. doi: 10.1016/j.kint.2020.04.003
Tavazzi, G., Pellegrini, C., Maurelli, M., Belliato, M., Sciutti, F., Bottazzi, A., et al. (2020). Myocardial localization of coronavirus in COVID-19 cardiogenic shock. Eur. J. Heart Fail. 22, 911–915. doi: 10.1002/ejhf.1828
Thery, C., Zitvogel, L., and Amigorena, S. (2002). Exosomes: composition, biogenesis and function. Nat. Rev. Immunol. 2, 569–579. doi: 10.1038/nri855
Tsuchiya, A., Takeuchi, S., Iwasawa, T., Kumagai, M., Sato, T., Motegi, S., et al. (2020). Therapeutic potential of mesenchymal stem cells and their exosomes in severe novel coronavirus disease 2019 (COVID-19) cases. Inflamm. Regen. 40:14.
Urciuoli, E., and Peruzzi, B. (2020). Inhibiting extracellular vesicle trafficking as antiviral approach to Corona virus disease 2019 infection. Front. Pharmacol. 11:580505.
van Niel, G., D’angelo, G., and Raposo, G. (2018). Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 19, 213–228. doi: 10.1038/nrm.2017.125
Wang, J., Chen, S., and Bihl, J. (2020). Exosome-mediated transfer of ACE2 (angiotensin-converting enzyme 2) from endothelial progenitor cells promotes survival and function of endothelial cell. Oxid. Med. Cell Longev. 2020: 4213541.
Zhang, P., Zhu, L., Cai, J., Lei, F., Qin, J. J., Xie, J., et al. (2020). Association of inpatient use of angiotensin-converting enzyme inhibitors and angiotensin II receptor blockers with mortality among patients with hypertension hospitalized with COVID-19. Circ. Res. 126, 1671–1681.
Zhang, W., Jiang, X., Bao, J., Wang, Y., Liu, H., and Tang, L. (2018). Exosomes in pathogen infections: a bridge to deliver molecules and link functions. Front. Immunol. 9:90.
Keywords: extracellular vescicles, COVID–19, renin—angiotensin—aldosterone system, cardiovascular disease, MSC derived exosomes, cardiosphere-derived cell, exosomes, cardiomyopathy
Citation: Pironti G, Andersson DC and Lund LH (2021) Mechanistic and Therapeutic Implications of Extracellular Vesicles as a Potential Link Between Covid-19 and Cardiovascular Disease Manifestations. Front. Cell Dev. Biol. 9:640723. doi: 10.3389/fcell.2021.640723
Received: 11 December 2020; Accepted: 21 January 2021;
Published: 11 February 2021.
Edited by:
Eduardo Marbán, Independent Researcher, Los Angeles, United StatesReviewed by:
Ahmed Gamal Ibrahim, Cedars-Sinai Medical Center, United StatesCopyright © 2021 Pironti, Andersson and Lund. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gianluigi Pironti, Z2lhbmx1aWdpLnBpcm9udGlAa2kuc2U=; orcid.org/0000-0001-9401-3029
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.