 Maxim V. C. Greenberg
Maxim V. C. Greenberg
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Cell Dev. Biol. , 07 January 2021
Sec. Stem Cell Research
Volume 8 - 2020 | https://doi.org/10.3389/fcell.2020.629068
This article is part of the Research Topic Chromatin Regulation in Cell Fate Decisions View all 15 articles
Vertebrate genomes are marked by notably high levels of 5-cytosine DNA methylation (5meC). The clearest function of DNA methylation among members of the subphylum is repression of potentially deleterious transposable elements (TEs). However, enrichment in the bodies of protein coding genes and pericentromeric heterochromatin indicate an important role for 5meC in those genomic compartments as well. Moreover, DNA methylation plays an important role in silencing of germline-specific genes. Impaired function of major components of DNA methylation machinery results in lethality in fish, amphibians and mammals. Despite such apparent importance, mammals exhibit a dramatic loss and regain of DNA methylation in early embryogenesis prior to implantation, and then again in the cells specified for the germline. In this minireview we will highlight recent studies that shine light on two major aspects of embryonic DNA methylation reprogramming: (1) The mechanism of DNA methylation loss after fertilization and (2) the protection of discrete loci from ectopic DNA methylation deposition during reestablishment. Finally, we will conclude with some extrapolations for the evolutionary underpinnings of such extraordinary events that seemingly put the genome under unnecessary risk during a particularly vulnerable window of development.
5-cytosine DNA methylation (5meC) is a modification conserved across all kingdoms of eukaryotes. Most generally it is found in the CpG dinucleotide context, and there are factors that ensure faithful daughter strand methylation after each round of DNA synthesis (Bostick et al., 2007; Sharif et al., 2007; Woo et al., 2008). Given the tight link with DNA replication, 5meC has high potential to exhibit epigenetic stability. In flowering plants, so-called “epialleles”—alleles that differ in 5meC content, not their DNA sequence—can persist for an indefinite number of generations (Weigel and Colot, 2012). Recently it was demonstrated in the yeast Cryptococcus neoformans that DNA methylation patterns have endured for millions of years exclusively through a maintenance mechanism, as there is an absence of de novo DNA methylation enzymes in the genome (Catania et al., 2020).
Standing apart from other eukaryotic lineages, it has been known for decades that mammals exhibit two rounds of dramatic DNA methylation reprogramming during embryonic development: first immediately after fertilizations, and a second time in the germline (Monk et al., 1987). The most facile explanation for the initial 5meC erasure event is that the embryo must level the high DNA methylation asymmetry exhibited by the paternal and maternal gametic genomes that arrive in the zygote (Wang et al., 2014), thus mitigating dosage discrepancies between alleles. By the blastocyst stage, which coincides with naive pluripotency, residual DNA methylation (~20% of CpGs) is largely restricted to genomic imprints and TEs (Wang et al., 2014, 2018; Zhu et al., 2018). As the embryo implants in the uterus, the de novo DNA methyltransferases, DNMT3A and DNMT3B, rapidly remethylate the genome to 70–80% CpG methylation, establishing a pattern that is globally maintained in somatic tissue-types (Borgel et al., 2010; Seisenberger et al., 2012; Smith et al., 2017; Zhang et al., 2018). This whole process repeats itself in primordial germ cells (PGCs), with a key difference being that in the germline, genomic imprints are erased and then reset in a sex-specific manner (Hajkova et al., 2002; Lee et al., 2002).
The last decade has seen advances in sequencing protocols and technology that have allowed for stunning temporal resolution of allele-specific DNA methylation patterns in early mammalian development (Wang et al., 2014, 2018; Gkountela et al., 2015; Zhu et al., 2018; Grosswendt et al., 2020). Nevertheless, much of the mechanistic underpinning of these processes remains lacking. To this end, embryonic stem cells (ESCs), which are derived from the inner cell mass (ICM) of the blastocyst, remain a powerful model for exploring the bases of the phenomenology of DNA methylation reprogramming. In this mini-review, we will highlight recent findings made in mouse ESCs (mESCs) that may help explain (1) how rapid global demethylation occurs, and (2) how promoters remain protected from the onslaught of DNA methylation establishment during implantation. Finally, given that DNA methylation reprogramming is a peculiarity—one that does not even appear to occur in our non-mammalian vertebrate cousins—we will discuss a possible clue that might explain the evolutionary rise of counterintuitive events.
DNA demethylation can occur via either passive or active means. Passive demethylation simply requires the impairment of maintenance DNA methylation machinery, which results in 2-fold dilution of methyl-CpGs during each round of DNA synthesis. In mammals, active DNA demethylation occurs through action of Ten-eleven translocase (TET) family of dioxygenases, although the mechanism is far from intuitive: iterative oxidation of 5meC to hydroxymethylcytosine (5hmC), formylcytosine (5fC), and finally carboxylcytosine (5caC) will trigger the thymine DNA glycosylase-dependent base excision repair (TDG BER) pathway to replace modified cytosines with unmodified versions (Kriaucionis and Heintz, 2009; Tahiliani et al., 2009; He et al., 2011; Maiti and Drohat, 2011; Weber et al., 2016). Oxidized forms of 5meC also impede DNA methylation maintenance (Hashimoto et al., 2012; Otani et al., 2013; Ji et al., 2014), thus in that sense, TET proteins contribute to passive DNA demethylation, as well.
The extent to which the TET enzymes contribute to the global demethylation exhibited during embryonic 5meC reprogramming remains an active, and somewhat controversial, area of research. Soon after fertilization, the DNA within the paternal pronucleus becomes strongly enriched for TET3-dependent 5hmC, relative to its maternal counterpart (Gu et al., 2011; Iqbal et al., 2011; Wossidlo et al., 2011). Moreover, paternal DNA methylation is rapidly erased in the zygote, before passive dilution can even occur (Mayer et al., 2000; Oswald et al., 2000). Therefore, it seems quite logical to posit that TET3 is responsible for demethylating the paternal genome. However, careful genetic dissection indicates that this is not entirely the case (Amouroux et al., 2016). Instead, TET3 activity may help protect the paternal genome from DNMT3A-dependent de novo DNA methylation (Amouroux et al., 2016; Albert et al., 2020). Of course, this would then imply there is an undiscovered mechanism of active DNA demethylation that must be occurring in the zygote. It should also be noted that in the early embryo, TET3 activity can lead to a passive loss of DNA methylation, independently of base-excision repair (Guo et al., 2014; Shen et al., 2014).
In fact, impaired DNA methylation maintenance plays an undeniably important role in reprogramming during the first cell divisions of preimplantation development. The maintenance pathway primarily consists of two proteins: the methyltransferase DNMT1 and its co-factor Ubiquitin-like, containing PHD and RING finger domains, 1 (UHRF1, also known as NP95). While there are many intricate layers to UHRF1/DNMT1 regulation, at its most fundamental level the mechanism is elegantly simple: UHRF1 recognizes hemimethylated CpG sites after DNA replication, and recruits DNMT1 to methylated the cytosine on the daughter strand (Bostick et al., 2007; Sharif et al., 2007) (Figure 1).
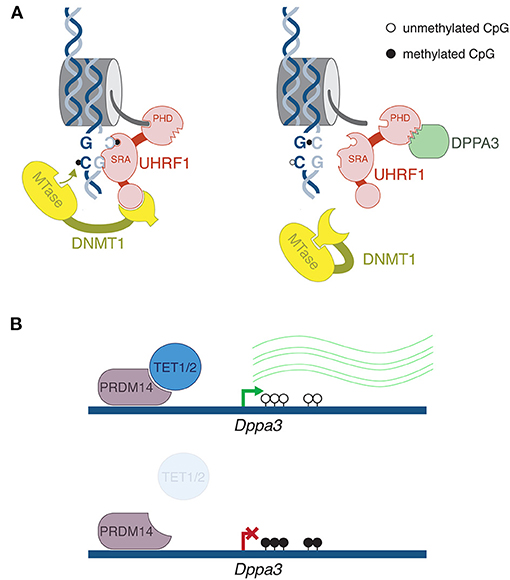
Figure 1. Mechanism of DPPA3-mediated DNA demethylation. (A) After DNA replication, the SRA domain of UHRF1 binds to hemimethylated CpG, flipping out the methylated cytosine. Recruitment of DNMT1 directs methylation of the symmetrical unmethylated cytosine on the daughter strand. Crucially, UHRF1 must bind to histone H3 through its PHD domain. DPPA3 binds to the PHD domain, disrupting UHRF1's interaction with chromatin leading to impaired DNA maintenance methylation. (B) Top panel: In mouse ESCs, PRDM14 recruits TET1 and TET2 roughly 2 kb upstream of the Dppa3 promoter, leading to its demethylation and activation. Bottom panel: in Tet1/2 double mutants, increased DNA methylation at the Dppa3 gene corresponds with repression. Thus, active demethylation of this one gene is required for global passive demethylation. This mechanism likely does not occur during embryonic DNA methylation reprogramming, but may help explain the wave of DNA demethylation that occurs in the male and female germlines.
In the past few years, studies have emerged that show UHRF1 disruption may be an important means of DNA demethylation. On the spectrum of differentiated cell types, oocytes are quite DNA hypomethylated: roughly 50% of CpGs are methylated in mouse and human oocytes, compared to 70–80% in somatic tissues (Kobayashi et al., 2012; Shirane et al., 2013; Okae et al., 2014). Recently it was demonstrated that the protein Developmental Pluripotency-Associated 3 (DPPA3, also known as STELLA and PGC7) shuttles UHRF1 to the cytoplasm of mouse oocytes (Li et al., 2018a). In the Dppa3 KO oocytes, UHRF1 returns to the nucleus, and oocyte methylation is increased (Li et al., 2018a; Han et al., 2019). It has been known for decades in mouse preimplantation embryos, an oocyte-specific isoform form of DNMT1 (oDNMT1) is enriched in the cytoplasm, hence its access to DNA is restricted (Carlson et al., 1992; Howell et al., 2001). While immunofluorescence indicates that oDNMT1 still remains largely cytoplasmic in Dppa3 mutants, a proportion does enter the nucleus as well—that is, DPPA3 activity is the best explanation for the lowly methylated state of the oocyte genome.
Our understanding of the DPPA3 regulation of UHRF1 has been greatly elucidated in a mESC system. When cultured in serum-free conditions and in the presence of MAPK and GSK3β inhibitors (2i), mESCs exhibit globally low levels of DNA methylation—more or less on par with the ICM cells from which ESCs are derived (Ficz et al., 2013; Habibi et al., 2013; Leitch et al., 2013). It was demonstrated that the underlying reason for such low methylation was impaired UHRF1 activity (von Meyenn et al., 2016). While initially UHRF1 was shown to be unstable at the post-translational level in 2i ESCs (von Meyenn et al., 2016), subsequently it was demonstrated that DPPA3 is the factor that is impeding UHRF1 function (Mulholland et al., 2020). Through a rigorous set of biochemical and microscopy-based experiments, Mulholland et al. showed that DPPA3 binds to UHRF1, thus impairing the latter's ability to bind to chromatin (Figure 1). In the mESC system, Dppa3 mutants are marked by reduction of UHRF1 localization to the cytoplasm, reminiscent of the phenotype in oocytes. However, this aspect of DPPA3-mediated UHRF1 regulation is much less drastic in the cell culture system, thus it is not completely clear if the mechanism of action is exactly the same between oocytes and mESCs. Moreover, DPPA3 can lead to demethylation without its nuclear export function (Du et al., 2019). Regardless, like in oocytes, absence of DPPA3 leads to an increased nuclear fraction of UHRF1 and a gain of DNA methylation. Consistently, when DPPA3 is overexpressed in other cell culture systems, there is a decrease in DNA methylation (Funaki et al., 2014). It should be noted that in addition to DPPA3 action, a number of features of 2i-cultured mESCs likely contribute to the globally DNA hypomethylated state, such as TET protein activity, de novo DNA methyltransferase repression (Leitch et al., 2013), and a chromatin state refractory to DNA methylation (van Mierlo et al., 2019), to name three pertinent examples. However, DPPA3 has clearly emerged as a dominant player for this particular feature.
Does the role of DPPA3 in mESCs reflect a role in the DNA demethylation occurring in preimplantation development? While formal demonstration awaits, there are some intriguing indications that indeed DPPA3 performs an analogous function in vivo: during normal development, expression of certain classes of TEs are important for activation of the proper embryonic transcription program (Macfarlan et al., 2012; Ishiuchi et al., 2015; Jachowicz et al., 2017); Dppa3 mutant mice exhibit repression of at least a proportion of these elements at the 2-cell stage (Huang et al., 2017). Given the important role of 5meC in transposon silencing, it is not outlandish to suggest that maintenance of DNA methylation on TEs at the heart of the Dppa3 transposon expression phenotype, with the caveat that it is difficult to tease apart the maternal from the embryonic effect, given these embryos were generated from Dppa3 mutant oocytes. Moreover, it should be noted that DPPA3 has been reported to protect the maternal genome from TET3-mediated demethylation in zygotes, i.e., the inverse phenomenon (Nakamura et al., 2007, 2012; Han et al., 2019). While not trivial given the severe phenotype of Dppa3 maternal/zygotic mutants (Payer et al., 2003), hopefully future work will help resolve these apparently contradictory functions.
Finally, Mulholland et al. showed that TET1 and TET2 are required for demethylation of Dppa3 regulatory regions, thus its activation (Figure 1) (Mulholland et al., 2020). In other words, targeted demethylation of one gene supports global passive demethylation. While this is a compelling finding, it is likely not the mechanism occurring after fertilization. Firstly, DPPA3 is already present in the maternal store of protein inherited from the oocyte (Li et al., 2018a); secondly, TET3 is the active TET enzyme after fertilization, not TET1 or 2; and thirdly, the Dppa3 gene arrives unmethylated from the oocyte, thus does not require further demethylation (Wang et al., 2014). However, this indeed might be the mechanism to activate Dppa3 prior to the germline DNA demethylation program. Dppa3 is expressed in the early stages of germline specification, and importantly, in vivo genetic analyses have revealed a role for DPPA3-mediated demethylation in PGCs, although the link with UHRF1 was not made (Nakashima et al., 2013). Incidentally, it has been reported that UHRF1 is downregulated during PGC progression at both the RNA and protein level (Kagiwada et al., 2013; Ohno et al., 2013). It will be necessary to eventually demonstrate if the DPPA3 phenomenology observed in mESCs occur at the relevant stages of in vivo development.
Shortly after reaching its lowest global levels of DNA methylation, the embryo implants into the uterine wall, which coincides with upregulation of the de novo DNA methylation program. In a few short days, the genome becomes highly DNA methylated. DNMT3A and DNMT3B show preference for histone 3 lysine 36 di- and trimethylation (H3K36me2/3)-marked regions, respectively, via binding by their PWWP domains (Dhayalan et al., 2010). H3K36me2/3 are broadly deposited in the genome, and may serve to enhance DNMT3A/B activity (Baubec et al., 2015; Weinberg et al., 2019). In general though, the de novo methyltransferases exhibit very little discrimination for genomic compartments, with one key exception: CpG island (CGI) promoters, which are distinguished by their markedly high CpG content. Roughly two-thirds of promoters are CGIs, and comprise most housekeeping and developmental genes (Gardiner-Garden and Frommer, 1987; Larsen et al., 1992; Ku et al., 2008). There is nothing about the sequence, per se, that should repel de novo DNA methylation; in fact, DNMT3A/B show a preference for CpG-rich sequences (Baubec et al., 2015). However, keeping promoters free of methylation is absolutely crucial for proper cellular function; DNA methylation is a very stable epigenetic mark, and ectopic promoter methylation can lead to long-term silencing of important genes. The ADD domain, also harbored by both enzymes, is repelled by H3K4 methylation (Ooi et al., 2007; Otani et al., 2009; Zhang et al., 2010; Guo et al., 2015). Given that H3K4me3 is strongly linked with active promoters, therein lies a simple mechanism to protect promoter sequences from DNA methylation deposition.
During the dramatic flux of DNA methylation in embryogenesis, there must be a means by which genes that are not expressed during reprogramming do not become unwilling targets of DNMT3A/B. Several years ago the discovery was made in mESCs that a large proportion of developmental gene promoters are “bivalently” marked—that is marked by both H3K4me3 and the polycomb repressive complex 2 (PRC2)-deposited H3K27me3 (Azuara et al., 2006; Bernstein et al., 2006). Also known as poised genes, these genes are silent, thanks to PRC2-mediated repression, but at the same time protected from DNA methylation and ready to activate upon the proper developmental cue. Moreover, there is some evidence that H3K27me3 marked regions may be refractory to de novo methylation independently of the H3K4 mark (Greenberg et al., 2017; Li et al., 2018b). Finally, TET1 is enriched at bivalent gene promoters (Manzo et al., 2017; Gu et al., 2018). Incidentally, TET1 contains a CxxC domain, which binds specifically to unmethylated CpG-rich sequences, however this domain does not appear to determine TET1 CGI localization (Zhang et al., 2016). Both TET proteins and KDM2B—a CxxC-domain containing complex associated with PRC1—have been demonstrated to protect CGIs from de novo DNA methylation (Boulard et al., 2015; Verma et al., 2018). Thus, there are several layers of regulation to prevent DNA methylation-based silencing.
In addition to TET and polycomb action, there must be a sequence-based recruitment of H3K4 methyltransferase complexes, i.e., through transcription factors. CGIs serve as important platforms of transcription factor binding, which is associated with alterations in DNA methylation level (Lienert et al., 2011; Krebs et al., 2014). Indeed, integration of a number of genome-wide datasets indicated the main determinant of sequences that do not exhibit DNA methylation is transcription factor binding (Kremsky and Corces, 2020). Recently, two studies converged on two factors that play a role in protecting bivalent genes: DPPA2 and DPPA4 (Eckersley-Maslin et al., 2020; Gretarsson and Hackett, 2020). While both studies utilized an mESC system, the discoveries were through different means. DPPA2/4 are heterodimeric transcription factors that are known to play a role in zygotic genome activation (De Iaco et al., 2019; Eckersley-Maslin et al., 2019; Yan et al., 2019), but they also are bound to bivalent genes (Engelen et al., 2015; Hernandez et al., 2018; Klein et al., 2018). Intriguingly, Dppa2/4 mutant mice exhibit developmental defects far after the embryonic stages where developmental genes are enriched for bivalent marks (Madan et al., 2009; Nakamura et al., 2011). Following up on this curiosity, Eckersley-Maslin et al. showed that in Dppa2/4 mutant mESCs, a subset of DPPA2/4 targets lose both H3K4me3 and H3K27me3 enrichment, indicating a role for these transcription factors in recruiting both silencing and activating complexes (Eckersley-Maslin et al., 2020) (Figure 2). Moreover, during differentiation to embryoid bodies, this subset of genes acquire DNA methylation and fail to activate, likely because they no longer are protected from de novo DNA methyltransferases (Figure 2).
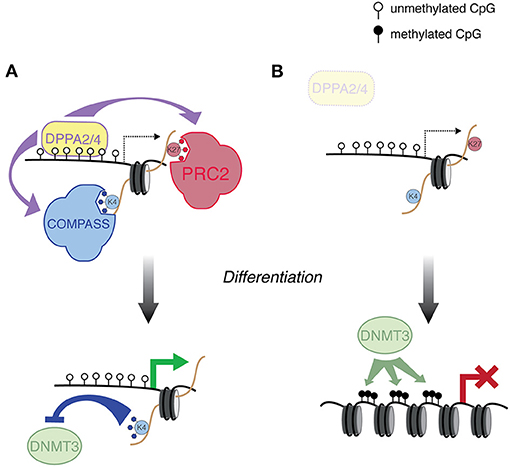
Figure 2. DPPA2/4-mediated regulation of developmental genes. (A) DPPA2/4 bind a subset of developmental genes, and recruit the COMPASS and PRC2 complexes, which deposit H3K4me3 and H3K27me3, respectively. These genes are then poised to activate in the proper developmental context. The H3K4me3 protects this class of promoter from de novo DNA methylation. (B) In Dppa2/4 mutant ESCs, genes that are normally targeted by DPPA2/4 are susceptible to aberrant de novo DNA methyltransferase activity, preventing the ability of this class of genes to activate during differentiation.
Gretarsson and Hackett ultimately arrived at similar conclusions, although they discovered a role for DPPA2/4 through different means. Using a clever mESC-based CRISPR screen, they searched for mutants that failed to activate a methylated reporter during an in vitro model of global DNA demethylation (Gretarsson and Hackett, 2020). Dppa2/4 were two of the top hits from the screen, but mutations did not lead to global DNA methylation perturbations. Consistent with the study from Eckersley-Maslin, DNA methylation abnormalities were restricted to a proportion of bivalently marked gene promoters. Curiously, DPPA2/4 also target LINE1 retrotransposons, although their role in transposon regulation is less clear. It should be noted that DNA methylation defects do not occur at all DPPA2/4 targets in the double mutant background; moreover, there are bivalent promoters that are not DPPA2/4 targets. That is all to say, DPPA2/4 are only two of the transcription factors important for DNA methylation protection during epigenetic reprogramming and there is a high probability others exist. Nevertheless, these are two important studies that undergird a model wherein sequence-specific transcription factors not only prime genes for proper expression during development, but also play a protective role against ectopic silencing.
DNA methylation, if misregulated, can have dire consequences. As discussed above, the improper de novo DNA methylation of promoters can lead to stable silencing of key developmental regulators. Perhaps more worrisome, the absence of DNA methylation can result not only in the ectopic expression of protein coding genes, but massive transposon derepression, which can have far ranging and deleterious effects (Walsh et al., 1998). Mouse embryos lacking either de novo or maintenance DNA methylation machinery die shortly after gastrulation (Li et al., 1992; Okano et al., 1999). Therefore, mammals must have evolved compensatory mechanisms to control the genome during not one, but two waves of DNA methylation reprogramming (Walter et al., 2016; Hill et al., 2018). What is more, the lowest levels of DNA methylation occur in pluripotent stem cells and primordial germ cells—the cell types that give rise to all somatic tissues and the germline, respectively. It is difficult to imagine cells that are more important for proper organismal development and transmission of genetic material to the next generation. Why then do mammals take such extraordinary apparent risks with their genome? Even among vertebrates this phenomenon is odd: zebrafish, by comparison, undergo nothing so drastic with their methylome during embryonic development (Skvortsova et al., 2019).
In investigating the evolutionary conservation of the Dppa3 gene, Mulholland et al. revealed it is not found outside mammals. Amazingly, when the mouse DPPA3 protein was incubated with the egg extracts of the amphibian Xenopus laevis, the mouse protein inhibited the frog UHRF1. Furthermore, when the fertilized eggs from the model fish medaka were injected with Dppa3 mRNA, the embryos exhibited dramatic hypomethylation (Mulholland et al., 2020). Therefore, DPPA3 has evolved as a potent DNA demethylation factor that can disrupt UHRF1 function in distant species. The authors suggest that perhaps Dppa3 arose in the mammalian lineage in concert with the role of transposon expression regulating the early transcription program. This certainly is possible, however it does not explain why global demethylation occurs exclusively in mammals, and also why it happens twice in development.
It is notable that Dppa3 is only present in placental mammals; marsupials and egg-laying monotremes lack the gene. Classic experiments in the 1980s demonstrated that mouse paternal and maternal genomes are not equivalent—androgenetic diploids exhibit robust placenta, whereas conversely gynogenetic diploids harbor severely undersized placenta (Surani and Barton, 1983; Barton et al., 1984; McGrath and Solter, 1984; Surani et al., 1984). This result is consistent with genetic conflict theory, which states that there is a conflict between the parental genes with regards to offspring development (Moore and Haig, 1991). In this case, the paternally expressed genes promote larger placenta leading to greater resource allocation to the developing embryo and fetus at the mother's expense, and for the maternally expressed genes it is the inverse. This theory was developed to explain the existence of genomic imprinting, which is parent-specific gene expression controlled by DNA methylation patterns inherited from the gametes. Pertinently, neither androgenetic nor gynogenetic embryos are viable. Given the stark DNA methylation asymmetry between gametes, perhaps Dppa3 evolved in order to prevent either the paternal or maternal genome exerting too much control with regards to inherent conflict. Imprint control regions, in turn, evolved mechanisms to escape DNA methylation erasure during embryogenesis. In the case of marsupials, there is evidence of germline reprogramming and genomic imprinting (Renfree et al., 2008; Suzuki et al., 2013; Ishihara et al., 2019), however evidence for embryonic reprogramming in both marsupials and monotremes is limited. Hopefully future studies will interrogate dynamic DNA methylation after conception in non-placental mammals, which will not only provide insights into DPPA3 function, but into evolutionary theory.
MVCG wrote the manuscript and prepared the figures.
MVCG was supported by a Laboratoire d'excellence Who Am I? (Labex 11-LABX-0071) Emerging Teams Grant and the European Research Council (ERC-StG-2019 DyNAmecs).
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Albert, J. R., Au Yeung, W. K., Toriyama, K., Kobayashi, H., Hirasawa, R., Brind'Amour, J., et al. (2020). Maternal DNMT3A-dependent de novo methylation of the paternal genome inhibits gene expression in the early embryo. Nat. Commun. 11:5417. doi: 10.1038/s41467-020-19279-7
Amouroux, R., Nashun, B., Shirane, K., Nakagawa, S., Hill, P. W., D'Souza, Z., et al. (2016). De novo DNA methylation drives 5hmC accumulation in mouse zygotes. Nat. Cell Biol. 18, 225–233. doi: 10.1038/ncb3296
Azuara, V., Perry, P., Sauer, S., Spivakov, M., Jørgensen, H. F., John, R. M., et al. (2006). Chromatin signatures of pluripotent cell lines. Nat. Cell Biol. 8, 532–538. doi: 10.1038/ncb1403
Barton, S. C., Surani, M. A., and Norris, M. L. (1984). Role of paternal and maternal genomes in mouse development. Nature 311, 374–376. doi: 10.1038/311374a0
Baubec, T., Colombo, D. F., Wirbelauer, C., Schmidt, J., Burger, L., Krebs, A. R., et al. (2015). Genomic profiling of DNA methyltransferases reveals a role for DNMT3B in genic methylation. Nature 520, 243–247. doi: 10.1038/nature14176
Bernstein, B. E., Mikkelsen, T. S., Xie, X., Kamal, M., Huebert, D. J., Cuff, J., et al. (2006). A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 125, 315–326. doi: 10.1016/j.cell.2006.02.041
Borgel, J., Guibert, S., Li, Y., Chiba, H., Schübeler, D., Sasaki, H., et al. (2010). Targets and dynamics of promoter DNA methylation during early mouse development. Nat. Genet. 42, 1093–1100. doi: 10.1038/ng.708
Bostick, M., Kim, J. K., Esteve, P. O., Clark, A., Pradhan, S., and Jacobsen, S. E. (2007). UHRF1 plays a role in maintaining DNA methylation in mammalian cells. Science 317, 1760–1764. doi: 10.1126/science.1147939
Boulard, M., Edwards, J. R., and Bestor, T. H. (2015). FBXL10 protects Polycomb-bound genes from hypermethylation. Nat. Genet. 47, 479–485. doi: 10.1038/ng.3272
Carlson, L. L., Page, A. W., and Bestor, T. H. (1992). Subcellular localization and properties of DNA methyltransferase in preimplantation mouse embryos: implications for genomic imprinting. Genes Dev. 6, 2536–2541. doi: 10.1101/gad.6.12b.2536
Catania, S., Dumesic, P. A., Pimentel, H., Nasif, A., Stoddard, C. I., Burke, J. E., et al. (2020). Evolutionary persistence of DNA methylation for millions of years after ancient loss of a de novo methyltransferase. Cell 180, 263–277.e20. doi: 10.1016/j.cell.2020.02.006
De Iaco, A., Coudray, A., Duc, J., and Trono, D. (2019). DPPA2 and DPPA4 are necessary to establish a 2C-like state in mouse embryonic stem cells. EMBO Rep. 20:e47382. doi: 10.15252/embr.201847382
Dhayalan, A., Rajavelu, A., Rathert, P., Tamas, R., Jurkowska, R. Z., Ragozin, S., et al. (2010). The Dnmt3a PWWP domain reads histone 3 lysine 36 trimethylation and guides DNA methylation. J. Biol. Chem. 285, 26114–26120. doi: 10.1074/jbc.M109.089433
Du, W., Dong, Q., Zhang, Z., Liu, B., Zhou, T., Xu, R.-M., et al. (2019). Stella protein facilitates DNA demethylation by disrupting the chromatin association of the RING finger–type E3 ubiquitin ligase UHRF1. J. Biol. Chem. 294, 8907–8917. doi: 10.1074/jbc.RA119.008008
Eckersley-Maslin, M., Alda-Catalinas, C., Blotenburg, M., Kreibich, E., Krueger, C., and Reik, W. (2019). Dppa2 and Dppa4 directly regulate the Dux-driven zygotic transcriptional program. Genes Dev. 33, 194–208. doi: 10.1101/gad.321174.118
Eckersley-Maslin, M. A., Parry, A., Blotenburg, M., Krueger, C., Ito, Y., Franklin, V. N. R., et al. (2020). Epigenetic priming by Dppa2 and 4 in pluripotency facilitates multi-lineage commitment. Nat. Struct. Mol. Biol. 27, 696–705. doi: 10.1038/s41594-020-0443-3
Engelen, E., Brandsma, J. H., Moen, M. J., Signorile, L., Dekkers, D. H. W., Demmers, J., et al. (2015). Proteins that bind regulatory regions identified by histone modification chromatin immunoprecipitations and mass spectrometry. Nat. Commun. 6:7155. doi: 10.1038/ncomms8155
Ficz, G., Hore, T. A., Santos, F., Lee, H. J., Dean, W., Arand, J., et al. (2013). FGF signaling inhibition in ESCs drives rapid genome-wide demethylation to the epigenetic ground state of pluripotency. Cell Stem Cell 13, 351–359. doi: 10.1016/j.stem.2013.06.004
Funaki, S., Nakamura, T., Nakatani, T., Umehara, H., Nakashima, H., and Nakano, T. (2014). Inhibition of maintenance DNA methylation by stella. Biochem. Biophys. Res. Commun. 453, 455–460. doi: 10.1016/j.bbrc.2014.09.101
Gardiner-Garden, M., and Frommer, M. (1987). CpG islands in vertebrate genomes. J. Mol. Biol. 196, 261–282. doi: 10.1016/0022-2836(87)90689-9
Gkountela, S., Zhang, K. X., Shafiq, T. A., Liao, W.-W., Hargan-Calvopiña, J., Chen, P.-Y., et al. (2015). DNA demethylation dynamics in the human prenatal germline. Cell 161, 1425–1436. doi: 10.1016/j.cell.2015.05.012
Greenberg, M. V. C., Glaser, J., Borsos, M., Marjou, F. E., Walter, M., Teissandier, A., et al. (2017). Transient transcription in the early embryo sets an epigenetic state that programs postnatal growth. Nat. Genet. 49, 110–118. doi: 10.1038/ng.3718
Gretarsson, K. H., and Hackett, J. A. (2020). Dppa2 and Dppa4 counteract de novo methylation to establish a permissive epigenome for development. Nat. Struct. Mol. Biol. 27, 706–716. doi: 10.1038/s41594-020-0445-1
Grosswendt, S., Kretzmer, H., Smith, Z. D., Kumar, A. S., Hetzel, S., Wittler, L., et al. (2020). Epigenetic regulator function through mouse gastrulation. Nature 584, 102–108. doi: 10.1038/s41586-020-2552-x
Gu, T., Lin, X., Cullen, S. M., Luo, M., Jeong, M., Estecio, M., et al. (2018). DNMT3A and TET1 cooperate to regulate promoter epigenetic landscapes in mouse embryonic stem cells. Genome Biol. 19:88. doi: 10.1186/s13059-018-1464-7
Gu, T. P., Guo, F., Yang, H., Wu, H. P., Xu, G. F., Liu, W., et al. (2011). The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 477, 606–610. doi: 10.1038/nature10443
Guo, F., Li, X., Liang, D., Li, T., Zhu, P., Guo, H., et al. (2014). Active and passive demethylation of male and female pronuclear DNA in the mammalian zygote. Cell Stem Cell 15, 447–459. doi: 10.1016/j.stem.2014.08.003
Guo, X., Wang, L., Li, J., Ding, Z., Xiao, J., Yin, X., et al. (2015). Structural insight into autoinhibition and histone H3-induced activation of DNMT3A. Nature 517, 640–644. doi: 10.1038/nature13899
Habibi, E., Brinkman, A. B., Arand, J., Kroeze, L. I., Kerstens, H. H. D., Matarese, F., et al. (2013). Whole-genome bisulfite sequencing of two distinct interconvertible DNA methylomes of mouse embryonic stem cells. Cell Stem Cell 13, 360–369. doi: 10.1016/j.stem.2013.06.002
Hajkova, P., Erhardt, S., Lane, N., Haaf, T., El-Maarri, O., Reik, W., et al. (2002). Epigenetic reprogramming in mouse primordial germ cells. Mech. Dev. 117, 15–23. doi: 10.1016/S0925-4773(02)00181-8
Han, L., Ren, C., Zhang, J., Shu, W., and Wang, Q. (2019). Differential roles of Stella in the modulation of DNA methylation during oocyte and zygotic development. Cell Discov 5:9. doi: 10.1038/s41421-019-0081-2
Hashimoto, H., Liu, Y., Upadhyay, A. K., Chang, Y., Howerton, S. B., Vertino, P. M., et al. (2012). Recognition and potential mechanisms for replication and erasure of cytosine hydroxymethylation. Nucl. Acids Res. 40, 4841–4849. doi: 10.1093/nar/gks155
He, Y. F., Li, B. Z., Li, Z., Liu, P., Wang, Y., Tang, Q., et al. (2011). Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science 333, 1303–1307. doi: 10.1126/science.1210944
Hernandez, C., Wang, Z., Ramazanov, B., Tang, Y., Mehta, S., Dambrot, C., et al. (2018). Dppa2/4 facilitate epigenetic remodeling during reprogramming to pluripotency. Cell Stem Cell 23, 396–411.e8. doi: 10.1016/j.stem.2018.08.001
Hill, S., Leitch, H. G., Requena, C. E., Sun, Z., Amouroux, R., Roman-trufero, M., et al. (2018). Epigenetic reprogramming enables the transition from primordial germ cell to gonocyte. Nature 555, 392–396. doi: 10.1038/nature25964
Howell, C. Y., Bestor, T. H., Ding, F., Latham, K. E., Mertineit, C., Trasler, J. M., et al. (2001). Genomic imprinting disrupted by a maternal effect mutation in the Dnmt1 gene. Cell 104, 829–838. doi: 10.1016/S0092-8674(01)00280-X
Huang, Y., Kim, J. K., Do, D. V., Lee, C., Penfold, C. A., Zylicz, J. J., et al. (2017). Stella modulates transcriptional and endogenous retrovirus programs during maternal-to-zygotic transition. Elife 6:e22345. doi: 10.7554/eLife.22345
Iqbal, K., Jin, S.-G., Pfeifer, G. P., and Szabó, P. E. (2011). Reprogramming of the paternal genome upon fertilization involves genome-wide oxidation of 5-methylcytosine. Proc. Natl. Acad. Sci. U.S.A. 108, 3642–3647. doi: 10.1073/pnas.1014033108
Ishihara, T., Hickford, D., Shaw, G., Pask, A. J., and Renfree, M. B. (2019). DNA methylation dynamics in the germline of the marsupial tammar wallaby, Macropus eugenii. DNA Res. 26, 85–94. doi: 10.1093/dnares/dsy040
Ishiuchi, T., Enriquez-Gasca, R., Mizutani, E., Boškoviä, A., Ziegler-Birling, C., Rodriguez-Terrones, D., et al. (2015). Early embryonic-like cells are induced by downregulating replication-dependent chromatin assembly. Nat. Struct. Mol. Biol. 22, 662–671. doi: 10.1038/nsmb.3066
Jachowicz, J. W., Bing, X., Pontabry, J., Bošković, A., Rando, O. J., and Torres-Padilla, M. E. (2017). LINE-1 activation after fertilization regulates global chromatin accessibility in the early mouse embryo. Nat. Genet. 49, 1502–1510. doi: 10.1038/ng.3945
Ji, D., Lin, K., Song, J., and Wang, Y. (2014). Effects of Tet-induced oxidation products of 5-methylcytosine on Dnmt1- and DNMT3a-mediated cytosine methylation. Mol. BioSyst. 10, 1749–1750. doi: 10.1039/c4mb00150h
Kagiwada, S., Kurimoto, K., Hirota, T., Yamaji, M., and Saitou, M. (2013). Replication-coupled passive DNA demethylation for the erasure of genome imprints in mice. EMBO J. 32, 340–353. doi: 10.1038/emboj.2012.331
Klein, R. H., Tung, P.-Y., Somanath, P., Fehling, H. J., and Knoepfler, P. S. (2018). Genomic functions of developmental pluripotency associated factor 4 (Dppa4) in pluripotent stem cells and cancer. Stem Cell Res. 31, 83–94. doi: 10.1016/j.scr.2018.07.009
Kobayashi, H., Sakurai, T., Imai, M., Takahashi, N., Fukuda, A., Yayoi, O., et al. (2012). Contribution of intragenic DNA methylation in mouse gametic DNA methylomes to establish Oocyte-specific heritable marks. PLoS Genet. 8:e1002440. doi: 10.1371/journal.pgen.1002440
Krebs, A. R., Dessus-Babus, S., Burger, L., and Schübeler, D. (2014). High-throughput engineering of a mammalian genome reveals building principles of methylation states at CG rich regions. Elife 3:e04094. doi: 10.7554/eLife.04094.019
Kremsky, I., and Corces, V. G. (2020). Protection from DNA re-methylation by transcription factors in primordial germ cells and pre-implantation embryos can explain trans-generational epigenetic inheritance. Genome Biol. 21:118. doi: 10.1186/s13059-020-02036-w
Kriaucionis, S., and Heintz, N. (2009). The nuclear DNA base 5-hydroxymethylcytosine is present in purkinje neurons and the brain. Science 324, 929–930. doi: 10.1126/science.1169786
Ku, M., Koche, R. P., Rheinbay, E., Mendenhall, E. M., Endoh, M., Mikkelsen, T. S., et al. (2008). Genomewide analysis of PRC1 and PRC2 occupancy identifies two classes of bivalent domains. PLoS Genet. 4:e1000242. doi: 10.1371/journal.pgen.1000242
Larsen, F., Gundersen, G., Lopez, R., and Prydz, H. (1992). CpG islands as gene markers in the human genome. Genomics 13, 1095–1107. doi: 10.1016/0888-7543(92)90024-M
Lee, J., Inoue, K., Ono, R., Ogonuki, N., Kohda, T., Kaneko-Ishino, T., et al. (2002). Erasing genomic imprinting memory in mouse clone embryos produced from day 11.5 primordial germ cells. Development 129, 1807–1817.Available online at: https://dev.biologists.org/content/129/8/1807.
Leitch, H. G., McEwen, K. R., Turp, A., Encheva, V., Carroll, T., Grabole, N., et al. (2013). Naive pluripotency is associated with global DNA hypomethylation. Nat. Struct. Mol. Biol. 20, 311–316. doi: 10.1038/nsmb.2510
Li, E., Bestor, T. H., and Jaenisch, R. (1992). Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell 69, 915–926. doi: 10.1016/0092-8674(92)90611-F
Li, Y., Zhang, Z., Chen, J., Liu, W., Lai, W., Liu, B., et al. (2018a). Stella safeguards the oocyte methylome by preventing de novo methylation mediated by DNMT1. Nature 564, 136–140. doi: 10.1038/s41586-018-0751-5
Li, Y., Zheng, H., Wang, Q., Zhou, C., Wei, L., Liu, X., et al. (2018b). Genome-wide analyses reveal a role of Polycomb in promoting hypomethylation of DNA methylation valleys. Genome Biol. 19:18. doi: 10.1186/s13059-018-1390-8
Lienert, F., Wirbelauer, C., Som, I., Dean, A., Mohn, F., and Schübeler, D. (2011). Identification of genetic elements that autonomously determine DNA methylation states. Nat. Genet. 43, 1091–1097. doi: 10.1038/ng.946
Macfarlan, T. S., Gifford, W. D., Driscoll, S., Lettieri, K., Rowe, H. M., Bonanomi, D., et al. (2012). Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature 487, 57–63. doi: 10.1038/nature11244
Madan, B., Madan, V., Weber, O., Tropel, P., Blum, C., Kieffer, E., et al. (2009). The pluripotency-associated gene Dppa4 is dispensable for embryonic stem cell identity and germ cell development but essential for embryogenesis. Mol. Cell Biol. 29, 3186–3203. doi: 10.1128/MCB.01970-08
Maiti, A., and Drohat, A. C. (2011). Thymine DNA glycosylase can rapidly excise 5-formylcytosine and 5-carboxylcytosine: potential implications for active demethylation of CpG sites. J. Biol. Chem. 286, 35334–35338. doi: 10.1074/jbc.C111.284620
Manzo, M., Wirz, J., Ambrosi, C., Villaseñor, R., Roschitzki, B., and Baubec, T. (2017). Isoform-specific localization of DNMT3A regulates DNA methylation fidelity at bivalent CpG islands. EMBO J. 36, 3421–3434. doi: 10.15252/embj.201797038
Mayer, W., Niveleau, A., Walter, J., Fundele, R., and Haaf, T. (2000). Demethylation of the zygotic paternal genome. Nature 403, 501–502. doi: 10.1038/35000656
McGrath, J., and Solter, D. (1984). Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell 37, 179–183. doi: 10.1016/0092-8674(84)90313-1
Monk, M., Boubelik, M., and Lehnert, S. (1987). Temporal and regional changes in DNA methylation in the embryonic, extraembryonic and germ cell lineages during mouse embryo development. Development 99, 371–382.
Moore, T., and Haig, D. (1991). Genomic imprinting in mammalian development: a parental tug-of-war. Trends Genet. 7, 45–49. doi: 10.1016/0168-9525(91)90040-W
Mulholland, C. B., Nishiyama, A., Ryan, J., Nakamura, R., Yigit, M., Glück, I. M., et al. (2020). Recent evolution of a TET-controlled and DPPA3/STELLA-driven pathway of passive DNA demethylation in mammals. Nat. Commun. 11:5972. doi: 10.1038/s41467-020-19603-1
Nakamura, T., Arai, Y., Umehara, H., Masuhara, M., Kimura, T., Taniguchi, H., et al. (2007). PGC7/Stella protects against DNA demethylation in early embryogenesis. Nat. Cell Biol. 9, 64–71. doi: 10.1038/ncb1519
Nakamura, T., Liu, Y.-J., Nakashima, H., Umehara, H., Inoue, K., Matoba, S., et al. (2012). PGC7 binds histone H3K9me2 to protect against conversion of 5mC to 5hmC in early embryos. Nature 486, 415–419. doi: 10.1038/nature11093
Nakamura, T., Nakagawa, M., Ichisaka, T., Shiota, A., and Yamanaka, S. (2011). Essential roles of ECAT15-2/Dppa2 in functional lung development. Mol. Cell Biol. 31, 4366–4378. doi: 10.1128/MCB.05701-11
Nakashima, H., Kimura, T., Kaga, Y., Nakatani, T., Seki, Y., Nakamura, T., et al. (2013). Effects of dppa3 on DNA methylation dynamics during primordial germ cell development in mice. Biol. Reprod. 88:125. doi: 10.1095/biolreprod.112.105932
Ohno, R., Nakayama, M., Naruse, C., Okashita, N., Takano, O., Tachibana, M., et al. (2013). A replication-dependent passive mechanism modulates DNA demethylation in mouse primordial germ cells. Development 140, 2892–2903. doi: 10.1242/dev.093229
Okae, H., Chiba, H., Hiura, H., Hamada, H., Sato, A., Utsunomiya, T., et al. (2014). Genome-wide analysis of DNA methylation dynamics during early human development. PLoS Genet. 10:e1004868. doi: 10.1371/journal.pgen.1004868
Okano, M., Bell, D. W., Haber, D. A., and Li, E. (1999). DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 99, 247–257. doi: 10.1016/S0092-8674(00)81656-6
Ooi, S. K., Qiu, C., Bernstein, E., Li, K., Jia, D., Yang, Z., et al. (2007). DNMT3L connects unmethylated lysine 4 of histone H3 to de novo methylation of DNA. Nature 448, 714–717. doi: 10.1038/nature05987
Oswald, J., Engemann, S., Lane, N., Mayer, W., Olek, A., Fundele, R., et al. (2000). Active demethylation of the paternal genome in the mouse zygote. Curr. Biol. 10, 475–478. doi: 10.1016/S0960-9822(00)00448-6
Otani, J., Kimura, H., Sharif, J., Endo, T. A., Mishima, Y., Kawakami, T., et al. (2013). Cell cycle-dependent turnover of 5-hydroxymethyl cytosine in mouse embryonic stem cells. PLoS ONE 8:e82961. doi: 10.1371/journal.pone.0082961
Otani, J., Nankumo, T., Arita, K., Inamoto, S., Ariyoshi, M., and Shirakawa, M. (2009). Structural basis for recognition of H3K4 methylation status by the DNA methyltransferase 3A ATRX–DNMT3–DNMT3L domain. EMBO Rep. 10, 1235–1241. doi: 10.1038/embor.2009.218
Payer, B., Saitou, M., Barton, S. C., Thresher, R., Dixon, J. P. C., Zahn, D., et al. (2003). Stella is a maternal effect gene required for normal early development in mice. Curr. Biol. 13, 2110–2117. doi: 10.1016/j.cub.2003.11.026
Renfree, M. B., Ager, E. I., Shaw, G., and Pask, A. J. (2008). Genomic imprinting in marsupial placentation. Reproduction 136, 523–531. doi: 10.1530/REP-08-0264
Seisenberger, S., Andrews, S., Krueger, F., Arand, J., Walter, J., Santos, F., et al. (2012). The dynamics of genome-wide DNA methylation reprogramming in mouse primordial germ cells. Mol. Cell 48, 849–862. doi: 10.1016/j.molcel.2012.11.001
Sharif, J., Muto, M., Takebayashi, S., Suetake, I., Iwamatsu, A., Endo, T. A., et al. (2007). The SRA protein Np95 mediates epigenetic inheritance by recruiting Dnmt1 to methylated DNA. Nature 450, 908–912. doi: 10.1038/nature06397
Shen, L., Inoue, A., He, J., Liu, Y., Lu, F., and Zhang, Y. (2014). Tet3 and DNA replication mediate demethylation of both the maternal and paternal genomes in mouse zygotes. Cell Stem Cell 15, 459–471. doi: 10.1016/j.stem.2014.09.002
Shirane, K., Toh, H., Kobayashi, H., Miura, F., Chiba, H., Ito, T., et al. (2013). Mouse oocyte methylomes at base resolution reveal genome-wide accumulation of non-CpG methylation and role of DNA methyltransferases. PLoS Genet. 9:e1003439. doi: 10.1371/journal.pgen.1003439
Skvortsova, K., Tarbashevich, K., Stehling, M., Lister, R., Irimia, M., Raz, E., et al. (2019). Retention of paternal DNA methylome in the developing zebrafish germline. Nat. Commun. 10:3054. doi: 10.1038/s41467-019-10895-6
Smith, Z. D., Shi, J., Gu, H., Donaghey, J., Clement, K., Cacchiarelli, D., et al. (2017). Epigenetic restriction of extraembryonic lineages mirrors the somatic transition to cancer. Nature 549, 543–547. doi: 10.1038/nature23891
Surani, M. A., and Barton, S. C. (1983). Development of gynogenetic eggs in the mouse: implications for parthenogenetic embryos. Science 222, 1034–1036. doi: 10.1126/science.6648518
Surani, M. A., Barton, S. C., and Norris, M. L. (1984). Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis. Nature 308, 548–550. doi: 10.1038/308548a0
Suzuki, S., Shaw, G., and Renfree, M. B. (2013). Postnatal epigenetic reprogramming in the germline of a marsupial, the tammar wallaby. Epigenetics Chromatin 6:14. doi: 10.1186/1756-8935-6-14
Tahiliani, M., Koh, K. P., Shen, Y., Pastor, W. A., Bandukwala, H., Brudno, Y., et al. (2009). Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324, 930–935. doi: 10.1126/science.1170116
van Mierlo, G., Dirks, R. A. M., De Clerck, L., Brinkman, A. B., Huth, M., Kloet, S. L., et al. (2019). Integrative proteomic profiling reveals PRC2-dependent epigenetic crosstalk maintains ground-state pluripotency. Cell Stem Cell 24, 123–137.e8. doi: 10.1016/j.stem.2018.10.017
Verma, N., Pan, H., Doré, L. C., Shukla, A., Li, Q. V., Pelham-Webb, B., et al. (2018). TET proteins safeguard bivalent promoters from de novo methylation in human embryonic stem cells. Nat. Genet. 50, 83–95. doi: 10.1038/s41588-017-0002-y
von Meyenn, F., Iurlaro, M., Habibi, E., Liu, N. Q., Salehzadeh-Yazdi, A., Santos, F., et al. (2016). Impairment of DNA methylation maintenance is the main cause of global demethylation in naive embryonic stem cells. Mol. Cell 62, 848–861. doi: 10.1016/j.molcel.2016.04.025
Walsh, C. P., Chaillet, J. R., and Bestor, T. H. (1998). Transcription of IAP endogenous retroviruses is constrained by cytosine methylation. Nat. Genet. 20, 116–117. doi: 10.1038/2413
Walter, M., Teissandier, A., Pérez-Palacios, R., Bourc'his, D., Akalin, A., Kormaksson, M., et al. (2016). An epigenetic switch ensures transposon repression upon dynamic loss of DNA methylation in embryonic stem cells. Elife 5:e11418. doi: 10.7554/eLife.11418
Wang, C., Liu, X., Gao, Y., Yang, L., Li, C., Liu, W., et al. (2018). Reprogramming of H3K9me3-dependent heterochromatin during mammalian embryo development. Nat. Cell Biol. 20, 620–631. doi: 10.1038/s41556-018-0093-4
Wang, L., Zhang, J., Duan, J., Gao, X., Zhu, W., Lu, X., et al. (2014). Programming and inheritance of parental DNA methylomes in mammals. Cell 157, 979–991. doi: 10.1016/j.cell.2014.04.017
Weber, A. R., Krawczyk, C., Robertson, A. B., Kuśnierczyk, A., Vågbø, C. B., Schuermann, D., et al. (2016). Biochemical reconstitution of TET1–TDG–BER-dependent active DNA demethylation reveals a highly coordinated mechanism. Nat. Commun. 7:10806. doi: 10.1038/ncomms10806
Weigel, D., and Colot, V. (2012). Epialleles in plant evolution. Genome Biol. 13:249. doi: 10.1186/gb-2012-13-10-249
Weinberg, D. N., Papillon-Cavanagh, S., Chen, H., Yue, Y., Chen, X., Rajagopalan, K. N., et al. (2019). The histone mark H3K36me2 recruits DNMT3A and shapes the intergenic DNA methylation landscape. Nature 573, 281–286. doi: 10.1038/s41586-019-1534-3
Woo, H. R., Dittmer, T. A., and Richards, E. J. (2008). Three SRA-domain methylcytosine-binding proteins cooperate to maintain global CpG methylation and epigenetic silencing in Arabidopsis. PLoS Genet. 4:e1000156. doi: 10.1371/journal.pgen.1000156
Wossidlo, M., Nakamura, T., Lepikhov, K., Marques, C. J., Zakhartchenko, V., Boiani, M., et al. (2011). 5-Hydroxymethylcytosine in the mammalian zygote is linked with epigenetic reprogramming. Nat. Commun. 2:1240. doi: 10.1038/ncomms1240
Yan, Y.-L., Zhang, C., Hao, J., Wang, X.-L., Ming, J., Mi, L., et al. (2019). DPPA2/4 and SUMO E3 ligase PIAS4 opposingly regulate zygotic transcriptional program. PLoS Biol. 17:e3000324. doi: 10.1371/journal.pbio.3000324
Zhang, W., Xia, W., Wang, Q., Towers, A. J., Chen, J., Gao, R., et al. (2016). Isoform switch of TET1 regulates DNA demethylation and mouse development. Mol. Cell 64, 1062–1073. doi: 10.1016/j.molcel.2016.10.030
Zhang, Y., Jurkowska, R., Soeroes, S., Rajavelu, A., Dhayalan, A., Bock, I., et al. (2010). Chromatin methylation activity of Dnmt3a and Dnmt3a/3L is guided by interaction of the ADD domain with the histone H3 tail. Nucl. Acids Res. 38, 4246–4253. doi: 10.1093/nar/gkq147
Zhang, Y., Xiang, Y., Yin, Q., Du, Z., Peng, X., Wang, Q., et al. (2018). Dynamic epigenomic landscapes during early lineage specification in mouse embryos. Nat. Genet. 50, 96–105. doi: 10.1038/s41588-017-0003-x
Keywords: DNA methylation, epigenetics, reprogramming, mammalian development, embryonic stem cells (ESC)
Citation: Greenberg MVC (2021) Get Out and Stay Out: New Insights Into DNA Methylation Reprogramming in Mammals. Front. Cell Dev. Biol. 8:629068. doi: 10.3389/fcell.2020.629068
Received: 13 November 2020; Accepted: 09 December 2020;
Published: 07 January 2021.
Edited by:
Bruno Di Stefano, Baylor College of Medicine, United StatesReviewed by:
Zachary Smith, Yale University, United StatesCopyright © 2021 Greenberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maxim V. C. Greenberg, bWF4aW0uZ3JlZW5iZXJnQGlqbS5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.