 Elia Bosch-Rué1†
Elia Bosch-Rué1† Leire Diez-Tercero1†
Leire Diez-Tercero1† Barbara Giordano-Kelhoffer1
Barbara Giordano-Kelhoffer1 Luis M. Delgado1
Luis M. Delgado1 Begoña M. Bosch1Mireia Hoyos-Nogués1
Begoña M. Bosch1Mireia Hoyos-Nogués1 Miguel Angel Mateos-Timoneda1
Miguel Angel Mateos-Timoneda1 Phong A. Tran2,3Francisco Javier Gil1
Phong A. Tran2,3Francisco Javier Gil1 Roman A. Perez1*
Roman A. Perez1*- 1Bioengineering Institute of Technology, Universitat Internacional de Catalunya, Barcelona, Spain
- 2Centre for Biomedical Technologies, Queensland University of Technology (QUT), Brisbane, QLD, Australia
- 3Interface Science and Materials Engineering Group, School of Mechanical, Medical and Process Engineering, Queensland University of Technology, Brisbane, QLD, Australia
Bone is the most studied tissue in the field of tissue regeneration. Even though it has intrinsic capability to regenerate upon injury, several pathologies and injuries could hamper the highly orchestrated bone formation and resorption process. Bone tissue engineering seeks to mimic the extracellular matrix of the tissue and the different biochemical pathways that lead to successful regeneration. For many years, the use of extrinsic factors (i.e., growth factors and drugs) to modulate these biological processes have been the preferred choice in the field. Even though it has been successful in some instances, this approach presents several drawbacks, such as safety-concerns, short release profile and half-time life of the compounds. On the other hand, the use of inorganic ions has attracted significant attention due to their therapeutic effects, stability and lower biological risks. Biomaterials play a key role in such strategies where they serve as a substrate for the incorporation and release of the ions. In this review, the methodologies used to incorporate ions in biomaterials is presented, highlighting the osteogenic properties of such ions and the roles of biomaterials in controlling their release.
Introduction
Bone is a complex and hierarchical organ that is in constant remodeling depending on the specific macro- and microenvironments. The main function of bone tissue is to provide mechanical stability to the body and protect the main organs (Florencio-Silva et al., 2015). Moreover, it has a high turnover which helps maintaining an ion balance in the body. Diseases or traumatic injuries can compromise bone function. For this purpose, it is of great importance to restore the lost functionality in a short and efficient manner. The gold standard has been the use of natural grafts (such as auto- and allografts), although their limited availability and possible transmission of diseases have limited their applications (Younger and Chapman, 1989; Betz, 2002). With this in mind, there is a great need to develop novel synthetic grafts that may provide similar functions to those of natural grafts while avoiding the possible related issues (Moore et al., 2001).
For successful bone regeneration, it is important that the bone substitute mimics to the highest extent the highly orchestrated bone regeneration steps (Perez et al., 2015b). Novel strategies involve the continuous adaptation of the inserted matrices to trigger a series of biological processes that may ultimately lead to bone regeneration (Pérez et al., 2013). These processes will mainly stimulate cells, through specific intrinsic features provided by the biomaterial and/or the release of extrinsic factors allocated within the substrate, such as molecules with biological activity (Perez et al., 2016). Among the different steps, the most critical are mitigation of the possible bacterial infection, the control of the initial inflammatory response, the attraction of cells to the site of defect through the formation of blood vessels, and the final maturation of the bone tissue (Perez et al., 2015b).
The different processes have been generally modulated by a combinatorial approach of intrinsic and extrinsic features of the defined biomaterials. Intrinsic properties, mainly the mechanical properties (both at a local and bulk-level), the surface roughness or the surface charge, are common parameters that can be tuned by a proper design (Navarrete et al., 2017). On the contrary, extrinsic factors are based on the use of biologically active molecules (such as growth factors) and peptides that can exert a positive response on tissue regeneration (Wang Z. et al., 2017). These extrinsic biologically active molecules have shown great potential in vitro. However, controlling their adequate delivery as well as their low half-life once implanted in the defect area are major hurdles for their clinical/industrial translation. To this purpose, the use of ions (mainly metal ions) is an attractive option because their therapeutic effect is well-known, they have an increased stability and, in terms of safety, they have lower risk than the use of biomolecules (Mouriño et al., 2012; Perez et al., 2015b). These therapeutic ions released from biomaterials can modulate the tissue regeneration steps in a similar way to biologically active molecules.
Hence, the scope of this review is to describe the vast possibilities of ion incorporation within biomaterials. The review will cover the different possible methodologies of ion incorporation within ceramic, metallic, and polymeric materials and the effect that the different methodologies will have on their final release. The attention will be given to their osteogenic properties, while the antibacterial and angiogenic properties have been reviewed elsewhere (Hoppe et al., 2011; Kaya et al., 2018).
Bone Regeneration Steps
Once a biomaterial is implanted, a dynamic interaction between the biomaterial surface and the plasma proteins takes place and a temporary matrix around the biomaterial or implanted device is formed; this phenomenon is known as the Vroman effect (Vroman et al., 1980). The initial protein adsorption depends on the surface properties of the biomaterial, which relate to wettability, surface charge, topography or stiffness, among others (Hallab et al., 2001; MacDonald et al., 2002; Xu and Siedlecki, 2007; Shiu et al., 2014), which modulate the initial cell response and subsequent bone healing phases. Bone healing is a complex biological process that is controlled by several growth factors, cytokines, cell types, and mechanical stimuli that direct the different overlapping phases, mainly haemostasis, acute inflammation, progenitor and stem cell homing, angiogenesis, osteogenic differentiation, and mineralisation (Perez et al., 2015b). The last step can be divided in mainly an osteogenic differentiation of stem cells followed by the mineralisation of the differentiated cells as described below. The role of ions has been proved in many of the bone regeneration processes, primarily acting as enzyme co-factors and, therefore, they influence the different signaling pathways (Zhi Lin Sun and Hanks, 1997). The potential role of ions is summarized in Figure 1. Understanding the different phases that take place during bone regeneration might be useful for the design of biomaterials that can deliver cues at specific time points.
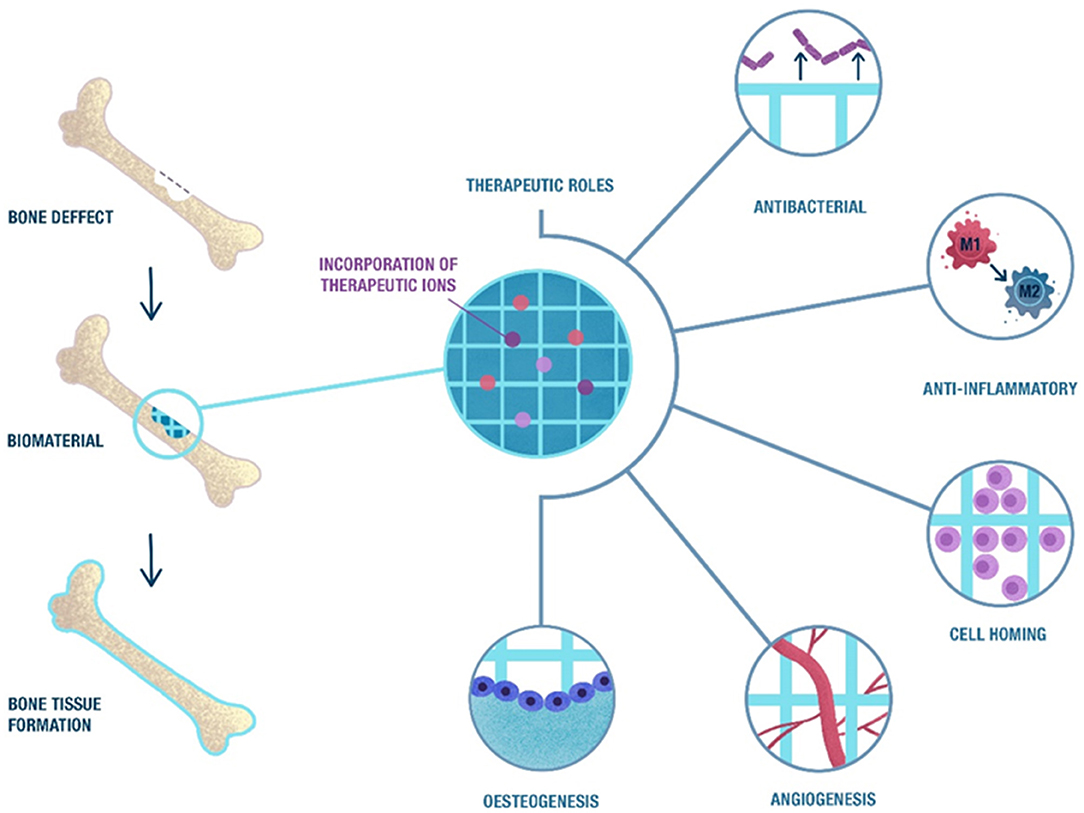
Figure 1. Schematic representation of the biological phases that take place during bone healing in which ions may have a therapeutic role. The natural bone healing starts with a pro-inflammatory response, followed by stem cell homing and the formation of new blood vessels. This process ends with the differentiation of stem cells into osteocytes and the formation of new bone. The release of ions from scaffolds may induce a therapeutic effect in these processes, combined with an anti-bacterial effect that may reduce the risk of infection after implantation.
Osteogenic Differentiation
Following stem cell homing, mesenchymal stromal cells (MSCs) proliferate and sequentially differentiate into chondroblasts and osteoblasts. Osteogenic differentiation only happens under specific microenvironments and stimuli, because MSCs have the ability to differentiate into other tissues, such as cartilage and fat (Dominici et al., 2006). Several different factors are implicated in this osteogenic differentiation. For example, physical cues such as stiffness or topographies have been shown to modulate MSCs differentiation (Engler et al., 2006; Dobbenga et al., 2016). On the other hand, growth factors such as fibroblast growth factors (FGFs), bone morphogenetic proteins (BMPs), and platelet derived growth factors (PDGFs) have also been demonstrated to be osteogenic stimulators (Behr et al., 2010; Kempen et al., 2010; Jeon et al., 2012). Therefore, scaffolds can be designed to provide biophysical and biochemical cues for stimulating osteogenic differentiation of stem cells and other osteoprogenitor cells. Moreover, this osteogenic differentiation is marked by an up-regulation of transcription factor Runx2, alkaline phosphatase (ALP), osteocalcin (OCN), osteopontin (OPN), bone sialoprotein (BSP), and ECM phosphoglycoproteins, among others, which stimulate mineralisation (Fakhry et al., 2013). Interestingly, it is known that many ions are involved in this process, such as Si4+, Sr2+, B3+, Ca2+, and Zn2+ and others which will be described more in detail in section Osteogenic Actions (Reffitt et al., 2003; Fukada et al., 2013; Hashimoto, 2020).
Mineralisation
Mineralisation of the extracellular matrix (ECM) is the last phase of bone healing and consists in a gradual reabsorption of the primary cartilaginous callus and replacement by a hard callus (Dimitriou et al., 2005). The arrival of MSCs to the site of injury activates their differentiation into chondroblasts and the molecular cascade of type I and type II collagen secretion. At this point, the transforming growth factor beta (TGF-β) and BMPs play an important role as they are involved in the chondrogenesis and the initiation of the bone healing cascade (Cho et al., 2002; Tsuji et al., 2006; Marsell and Einhorn, 2009). Consequently, this mineralisation step is the result of multiple interactions between cells, ECM proteins and environmental factors such as pH, ionic strength, and mechanical stimulus (Marsell and Einhorn, 2011).
During mineralisation, osteoblasts secrete matrix vesicles containing small calcium phosphate crystal precursors and lipids that are capable of attracting calcium (Golub, 2009). Moreover, collagen type I (COL1) acts as a template for the precipitation of the inorganic matter (Fedde et al., 1999). Together with COL1, other proteins from the ECM of bone, such as OCN, OPN, BSP, and phosphoglycoproteins, have a key role in the mineralisation process. Some of these proteins have calcium binding domains, which in turn bind to phosphate ions, allowing the formation of apatite crystals (Chen et al., 1992).
Incorporating Ions Within Biomaterials
Osteogenic ions have been incorporated into biomaterials to enhance bone regeneration. Depending on the type and morphology of the biomaterials, different incorporation techniques can be used that result in different ion release characteristics. In this section, a brief description of the incorporation of ions in different materials and the overall morphology of the composite (or the potential bone substitute), which, in fact, will modulate the ion release, will be described.
Types of Materials and Incorporation Methods
An overview of the possible designs of biomaterials incorporating ions is shown in Figure 2, where different types of morphologies are established as well as different types of compositions. The different possibilities will be briefly described.
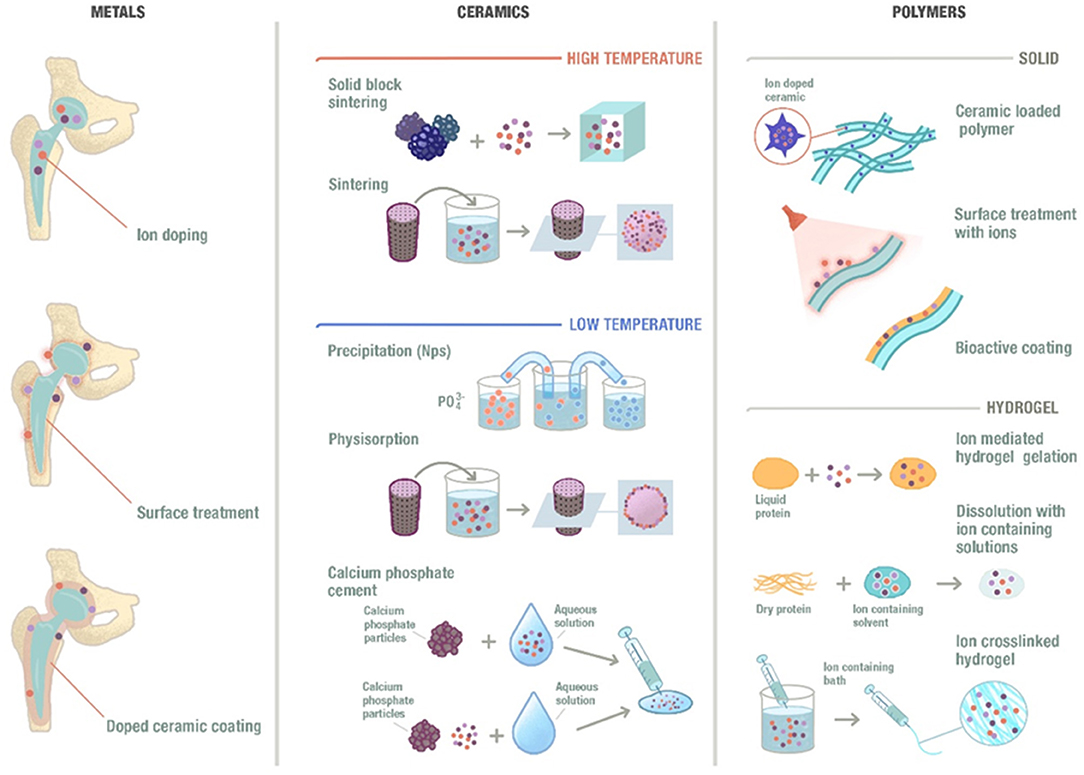
Figure 2. Schematic representation of the ion incorporation strategies depending on the different types of material and morphologies. Ions could be incorporated to metallic biomaterials by integrating them into their composition, performing a substrate treatment or including them into a ceramic coating that will be applied to the surface of the metal. Moreover, ions can also be incorporated into ceramic materials at high temperatures or low temperatures and into polymeric materials following different methods.
Ceramics
Ceramics are commonly used materials in the field of bone tissue regeneration. A well-known ceramic is hydroxyapatite (HA), which presents multiple synthesis routes at low or high temperature (Habraken et al., 2016). One of the main disadvantages of HA is the low degradation and resorption rates, mainly related with its high degree of crystallinity. In order to reduce its crystallinity, and hence to trigger reabsorption, its structure is usually disrupted by incorporating different ions, such as Sr2+, Zn2+, or Mg2+, among others (Reger et al., 2019). For the high temperature ceramics, these ions are generally incorporated by doping the apatites prior to synthesis. Another strategy to incorporate ions is to sinter the apatites in the absence of the ions, which can then be soaked in different solutions containing the desired ions to allow their physisorption (Garley et al., 2019). For the low temperature ceramics, these ions can be introduced by simply incorporating the desired ion into the calcium and phosphate solutions, forcing its reaction and, consequently, leading to the precipitation of the doped apatite (Evis and Webster, 2011; Ofudje et al., 2019). A slightly different approach for the low temperature fabrication is the use of calcium phosphate cements (CPC), which is based on the mixing of a powder and a liquid to form solid apatite at room temperature (Perez et al., 2012, 2015c). In this case, the ion can be incorporated either as a salt within the powder, or as an ionic solution within the liquid phase (Schamel et al., 2017).
Ion-doped glass-ceramics and glasses have also been used in bone tissue engineering due to their bioactivity, resorbability and appropriate mechanical properties. The incorporation of trace elements modifies their biological activity, allowing the release of therapeutic ions from their structure (Hoppe et al., 2011). The release kinetics of these ions can be controlled depending on the incorporation method used as well as on the physicochemical properties of the material (Castaño et al., 2017).
Polymers
The ability of polymeric materials to incorporate ions differs depending on the type of polymer used. Synthetic polymers are less susceptible to easily incorporate metallic ions, since tedious and complicated chemical routes are needed. Other option is the blending of such polymers with ionic-releasing particles, such as bioactive glass (Olmos Buitrago et al., 2015; Alizadeh-Osgouei et al., 2019). Among the different natural polymers that can be used, polysaccharides (such as alginate) and natural proteins (such as collagen) are the most commonly used. Amongst the polysaccharides, alginate is an interesting example of a versatile polymer that allows the incorporation of ions by simple crosslinking of the polysaccharide chains with divalent ions (Perez and Kim, 2013; Perez et al., 2015a). This allows the gelation of the polymer, which maintains the ion within the structure. Once the hydrogel is placed within a monovalent ion solution, the ions are exchanged, releasing the divalent ions within the alginate.
Metals
Different ions have also been successfully incorporated to metallic biomaterials. Since ions are generally of metallic origin, these can be integrated within the overall composition of the metals, allowing a slow and sustained release (Alrabeah et al., 2017). While this may take long time to allow the release of the therapeutic ions, the continuous exchange of ions and the liquid environment may lead to certain corrosion issues that need to be controlled (Noumbissi et al., 2019). Furthermore, the ions can be incorporated on the surface of the metallic substrates by surface treatments, such as etching, plasma treatment or coatings (Lu et al., 2012). For example, a ceramic coating can be applied to the surface of the metallic device to incorporate the therapeutic ions (Kose et al., 2013). These ions will mainly be found on the surface and their release may be more easily tuned without jeopardizing the bulk properties of the implant.
Overall Morphology
Besides the biomaterial composition, different morphologies of the final device might determine how ions are released (concentration and time) and sensed by the cells, (Figure 3).
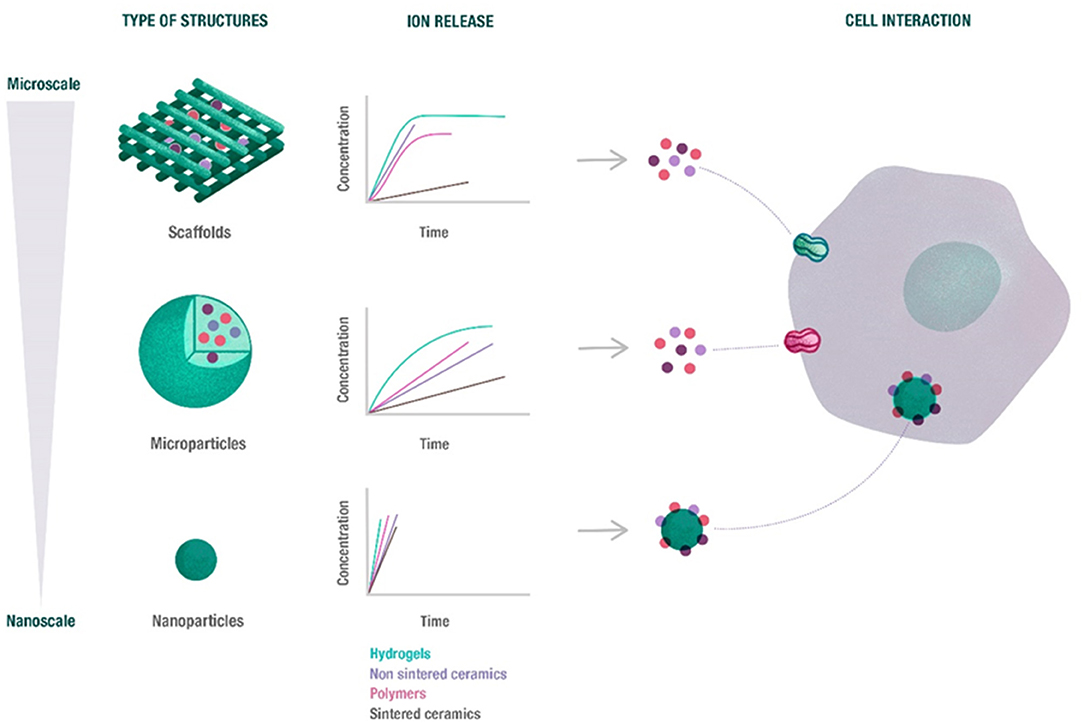
Figure 3. Overview of the potential release of ions depending on the biomaterial morphology. The release profile of ions from scaffolds, microparticles and nanoparticles depends on biomaterial size and morphology, as well as its composition. Nanoparticles rapidly released the ions, while microparticles exhibited a sustained release. Moreover, the ions released from scaffolds and microparticles are able to enter cell through ion channels, while nanoparticles can be directly internalized.
Scaffolds (Porous and Fibrous)
Three dimensional scaffolds are the most commonly used structures for bone tissue regeneration. There are several requirements for an ideal scaffold for bone tissue engineering, mainly to provide similar mechanical properties to those of the native bone (especially in load-bearing areas), to degrade while the new bone is being formed, to present sufficient porosity allowing cell infiltration, as well as to give osteoconductive and osteoinductive cues for bone maturation (Perez and Mestres, 2016). These scaffolds can be fabricated by several methods, including freeze-drying, solvent casting, solid freeform fabrication or phase separation, among others (Chocholata et al., 2019). A special type of preparation is electrospinning, which allows forming thin layers of nanofibrous materials, mimicking the fibrillar ECM structure (Xue et al., 2019). Its main disadvantage is the limited space for cell migration (i.e., small pore size and interconnectivity) and low mechanical stiffness. To overcome these limitations, 3D printing is receiving significant attraction, due to the ability to fabricate custom-made structures with greater control over the porosity (size, morphology, and interconnectivity) (Murphy and Atala, 2014; Kyle et al., 2017; Jammalamadaka and Tappa, 2018). These scaffolds, however, generally require an invasive surgery since the devices need to be implanted into the site of the defect.
Hydrogels
Hydrogels are a special kind of polymer scaffold. They entrap high amounts of water within the polymeric network. Many hydrogels are fully injectable into the site of defect (Drury and Mooney, 2003). This property offers a great advantage since it reduces the risk of infection in surgery, due to their less invasive characteristics. Furthermore, their low temperature processing (mainly at room or body temperature) allows the delivery of biologically active molecules as well as cells (Bai et al., 2018). The most commonly used hydrogels are based on polysaccharides and natural proteins. Their degradation rates are generally fast but can be strategically cross-linked to tune their degradation rates to match with the formation of new bone. Their main disadvantage is the poor mechanical properties which make them more suitable for non-load bearing applications.
Microparticles (MPs)
Based on the gaps provided by the packing of spheres, this allows to use microspheres or microcarriers as a matrix that provides sufficient porosity for cell and bone ingrowth (Park et al., 2013; Salerno et al., 2013). Their main advantage is that each microsphere, in the range between 100 and 400 μm can be used to individually grow cells and allow placing an interconnected macro-scaffold with individually seeded microscaffolds into the site of defect. Overall, this would allow for a uniform distribution of cells throughout the site of defect. Furthermore, their homogenous size allows a constant release of the incorporated molecules or ions (Perez et al., 2014; Timin et al., 2018). Moreover, these can be made injectable/printable by simply mixing the microspheres with a proper hydrogel (Levato et al., 2014).
Nanoparticles (NPs)
In a similar way to MPs, NPs present unique properties based on their nano sizes. Their range of sizes down to the nano-level confers them high specific surface areas with electrostatic charge that allows their penetration into cells (Vieira et al., 2017). These are generally synthesized by chemical routes and their size ranges between 20 and 200 nm. These particles can be made of polymeric, ceramic or metallic materials. It is worth highlighting that NPs will offer a significant differential effect compared to MPs based on the fact that a number of NPs are able to enter into the cells and release ions within the cell cytoplasm, whereas MPs are only able to release ions outside the cytoplasm (Palombella et al., 2017) (Figure 3).
Osteogenic Actions
Some ions are trace elements of bone, being some of them essential for the normal development of the skeleton. Besides, their deficiency can cause diseases such as osteoporosis, in which there is a reduction of the bone mineral mass and change in the microarchitecture (Aaseth et al., 2012). The addition of ions in biomaterials, such as Ca2+, Cu2+, Sr2+, Mg2+, Zn2+, and B3+ amongst others, have shown to enhance osteogenesis and, lead to better overall bone regeneration (Glenske et al., 2018; Lu et al., 2019). For this reason, it is critical to understand how ions interact with cells in vitro and how these ions could be incorporated in the appropriate doses within biomaterials to be beneficial both in vitro as well as in vivo. The main mechanisms of actions of the key ions are schematically represented in Figure 4 and their incorporation into and delivery from biomaterials described below. In order to summarize the different actions promoted by ions, three different summarizing tables are presented. Table 1 shows the effects of the different ions incorporated and released from the structure of metals. Table 2 shows the effects of different ions incorporated and released from ceramics while Table 3 shows the effects of incorporated ions within polymeric materials.
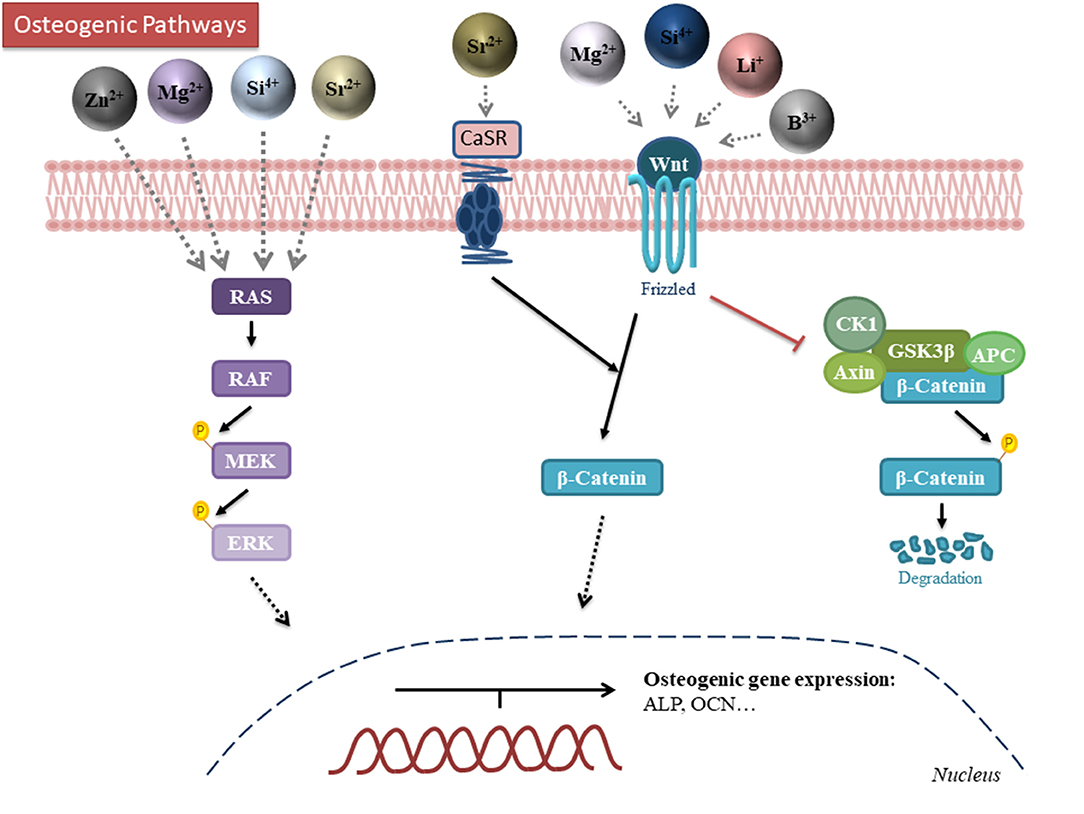
Figure 4. Schematic representation of the mechanism of action for the different osteogenic ions. Zinc, magnesium, silicon and strontium modulate the MAPK-ERK pathway. Strontium, magnesium, silicon, lithium and boron activate the canonical Wnt/β-catenin pathway. Darker color intensity on the ions indicates a more established pathway. B, boron; Li, Lithium; Mg, magnesium; Si, silicon; Sr, Strontium; Zn, Zinc; RAS, Rat sarcoma; RAF, Rapidly activated Fibrosarcoma; MEK, mitogen-activated protein/extracellular signal-regulated kinase kinase (MAPK/Erk kinase); CK1, casein kinase 1; APC, Adenomatous Polyposis Coli.
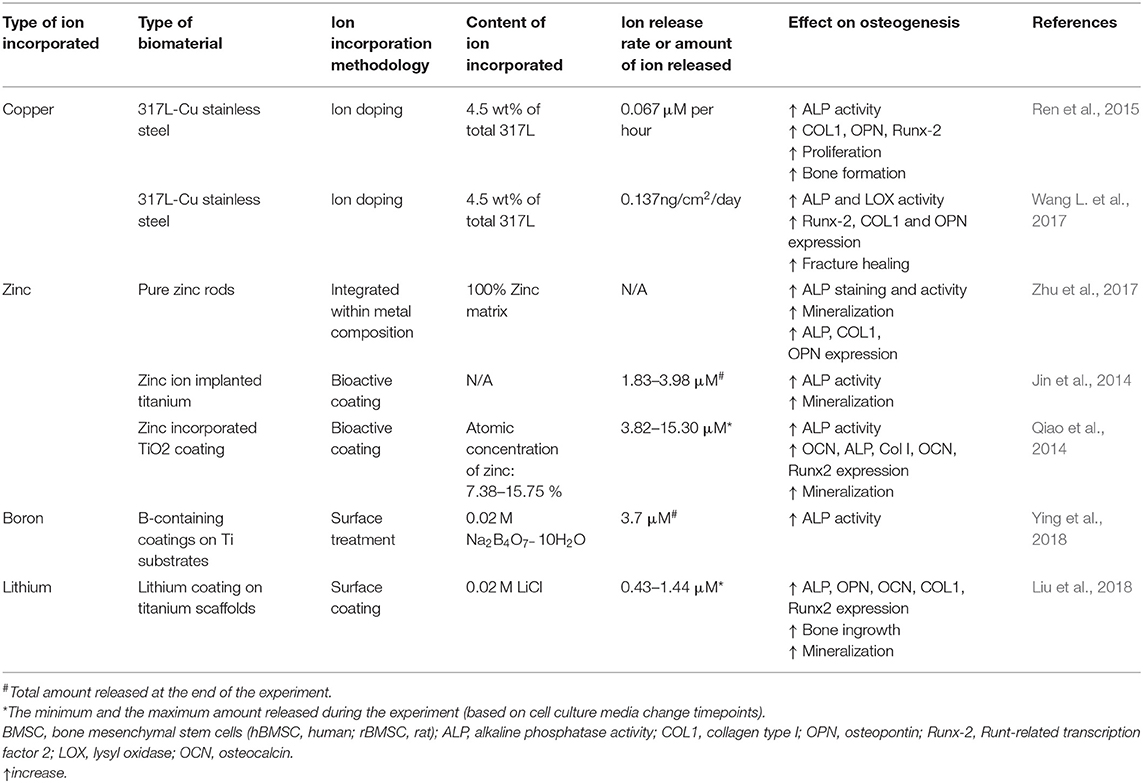
Table 1. Summary of studies incorporating different ions within metal biomaterials.
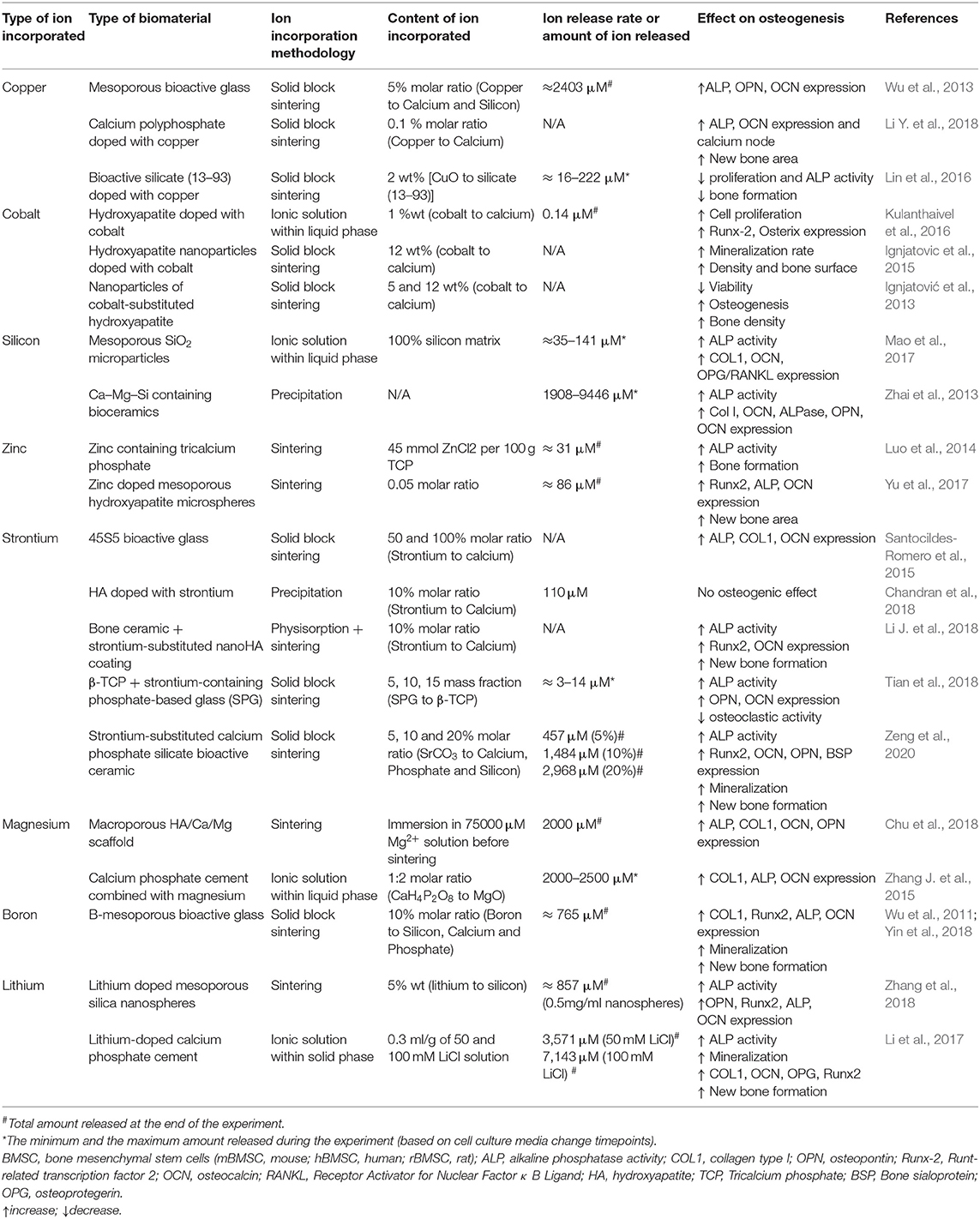
Table 2. Summary of studies incorporating different ions within ceramic biomaterials.
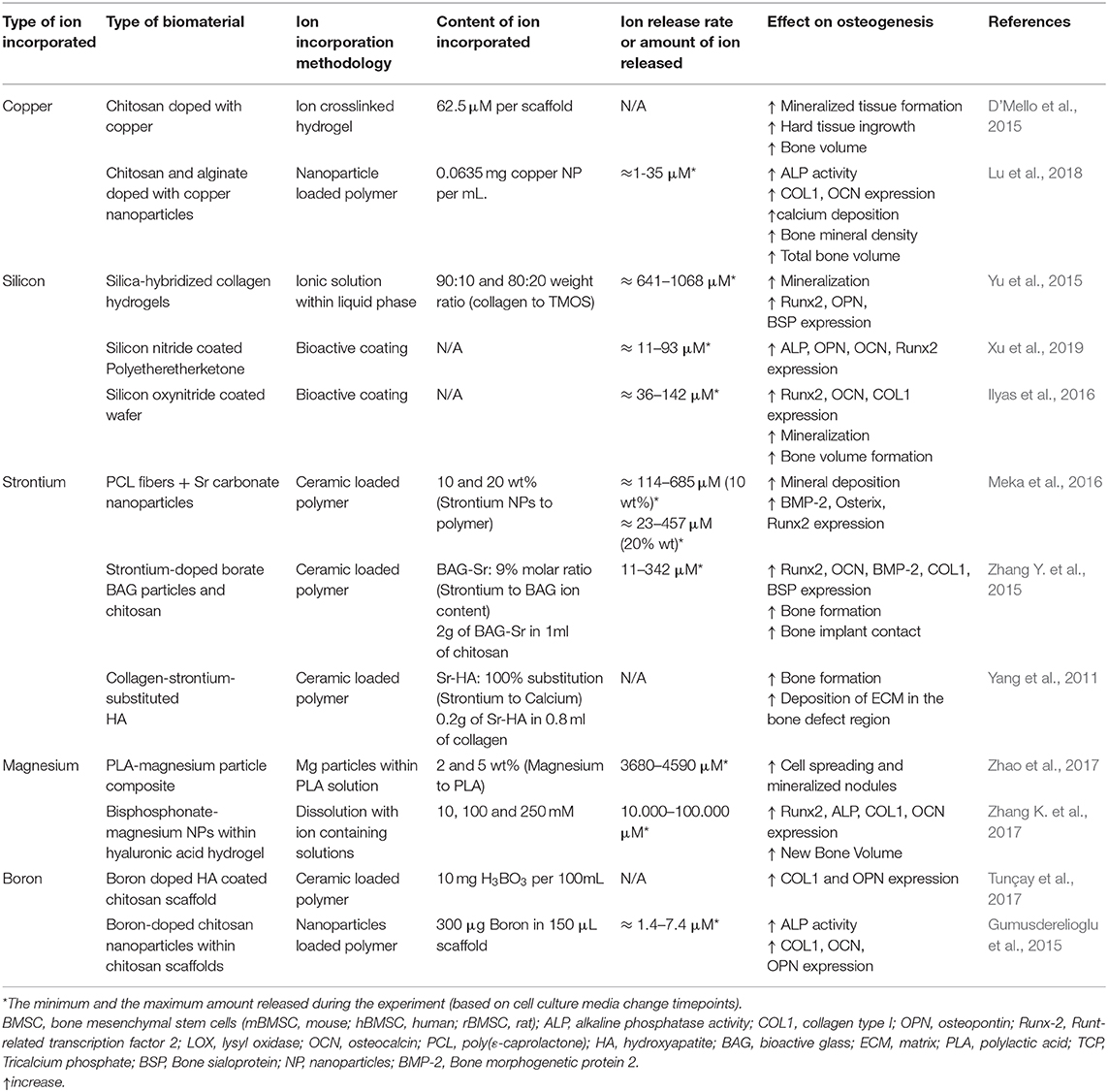
Table 3. Summary of studies incorporating different ions within polymer biomaterials.
Copper
Copper has been described to enhance the osteogenic differentiation in in vitro studies. The incorporation of copper in cell culture medium reduced the proliferation of MSCs although it increased the capacity of osteogenic differentiation, inducing an early expression of ALP and mineralisation (Pablo Rodrguez et al., 2002). Little is known on the exact mechanism of how copper affects the osteogenic differentiation. Nevertheless, a possible hypothesis is that it takes place through ECM processing, since several enzymes involved in the crosslinking of collagen and elastin, such as lysyl oxidase (LOX), are copper-dependent, and it is well-known that the ECM composition and structure influence stem cell differentiation (Smith et al., 2018). Some works have also reported a negative effect of copper. A study reported that the supplementation of culture media with copper showed a lower expression of osteogenic genes in rat bone marrow stromal cells (rBMSC), suggesting that copper suppressed the Runx2 signaling pathway which in turn suppressed the collagen deposition (Li et al., 2014). Hence, further studies are needed to elucidate a more realistic and accurate mechanism.
Copper has often been incorporated in metallic materials, as well as in other biomaterials, in order to stimulate bone regeneration. Regarding the metallic materials, several prostheses as well as dental implants present copper in their structure. Interestingly, the presence of copper in dental implants showed increased ALP activity and increased COL1, osteoprotegerin and OPN expression, which together enhanced the mineralisation potential of MSCs (Burghardt et al., 2015). The copper containing implants were then placed in an in vivo animal study, showing an increase in bone formation and bone-implant integration (Ren et al., 2015). Similar results were shown for nano-copper-bearing stainless steel, which were shown to increase osteogenic cell expression in vitro and to enhance bone formation in vivo (Wang L. et al., 2017).
Similar to metals, ceramics have also been doped with copper. Several ceramics, such as bioactive glasses and phosphate-based ceramics, have shown enhanced bone regeneration capabilities. An in vitro study showed that copper-containing microporous bioactive glass (Cu-MBG) or its ionic products cultured with human bone marrow stromal cells (hBMSCs) induced a higher expression of ALP, OPN, and OCN (Wu et al., 2013). Similar results were observed in vivo, showing enhanced bone regeneration (Li Y. et al., 2018). Furthermore, copper is able to enhance the apatite-forming ability when incorporated into bioactive glass NPs (Zheng et al., 2017). In some cases, copper has been shown to negatively impact osteogenic induction. For instance, pre-osteoblastic MC3T3-E1 cells cultured with bioactive silicate (13–93) doped with different concentrations of copper (0–2 wt%) showed no significant differences in proliferation or ALP activity. Moreover, the highest dose (2 wt% CuO) presented a negative effect in osteogenic response, showing lower ALP activity than the pristine samples. These results were also confirmed with an in vivo study in a rat calvaria defect model (Lin et al., 2016).
Unlike metals and ceramics, polymeric materials have rarely been combined with copper ions. Nevertheless, a recent study mixed 2% w/v chitosan solution with 0.625 mM copper solution to fabricate freeze-dried scaffolds containing a total of 62.5 μM of copper within chitosan hydrogels. The resulting doped-scaffolds significantly increased bone volume formation in a rat calvaria defect model, although authors suggest a possible angiogenic role of copper which might benefit bone formation (D'Mello et al., 2015). The main limitation of the study was the fast release of the ions, limiting the presence of the copper ions in the site of defect during the osteogenic induction period. In order to overcome the limitations and the fast release from polymeric materials, copper containing NPs rather than ions have been used. For instance, copper NPs combined with alginate and chitosan significantly increased COL1 and OCN gene expression, as well as the genes involved in cell adhesion, together with an increase in the extracellular calcium deposition. The results were then extrapolated into in vivo studies, showing increased bone mineral density and greater total bone volume compared to control (Lu et al., 2018).
Cobalt
Similar to copper, cobalt is also known to initially stimulate the angiogenic response, which may also contribute to the osteogenic process. Nevertheless, its mechanism is not clearly elucidated. Actually, cobalt-supplemented culture media has not shown a positive osteogenic induction but rather an impairment in the osteogenic differentiation (Patntirapong et al., 2009; Birgani et al., 2016; Drynda et al., 2018). Despite the previous results, cobalt has been incorporated in several biomaterials, mainly in ceramics. For instance, HA powders doped with different concentrations of cobalt (0.5, 1, 5, and 10 w/w%) showed that in samples with 1% cobalt, there was an increase in proliferation of MG-63 cells, as well as an increased expression of Runx2 and Osterix. The proposed mechanism of action was that cobalt acted as a calcium agonist, activating Wingless-related integration site 5 (Wnt5) signaling pathway cascade and hence leading to the expression of Runx2 and subsequently, the expression of Osterix (Kulanthaivel et al., 2016). Positive results were also found in vivo, showing that HA NPs doped with cobalt produced dense collagen fibers with a mineralisation rate higher than HA, showing as well a higher bone surface and density in a rat bone mandibular defect (Ignjatovic et al., 2015). However, different results have been found in in vitro and in vivo studies. For instance, cobalt-doped NPs produced a decrease in viability and cytoskeletal deformation in osteoblastic cells in vitro. Interestingly, when implanted in vivo, an increased rate of osteogenesis with higher alveolar bone density with a well formed vasculature was found (Ignjatović et al., 2013). The differences between the in vitro and in vivo results are probably ascribed to the possible negative effects caused at the cellular level by NPs, since it is known that size, shape and morphology may have profound negative effects on cell morphology, which are not necessarily found in vivo (Kwon et al., 2013; Perez et al., 2017). With the controversial results found in the literature, it is not clear whether cobalt can be beneficial or useful for osteogenic differentiation. It has been recently hypothesized that the main role of cobalt in bone regeneration is to stimulate angiogenesis with new blood vessel formation, which is an important step to successfully achieve a proper osteogenic differentiation and a proper overall bone regeneration (Perez et al., 2015a; O'Neill et al., 2018).
Silicon
Another well-known ion that has shown significant effects on osteogenic differentiation and mineralization is silicon. Silicon is found in the active calcification sites in bones and is clearly implicated in the mineralisation process of bone growth (Jugdaohsingh et al., 2002; Han et al., 2013). It was previously suggested that silicon is a key element in the initiation of the pre-osseous tissue mineralisation (Shi et al., 2015). Supplementation with silanol in vivo showed that silicon partially stopped the trabecular bone loss in mature ovariectomized rats by minimizing bone resorption and enhancing bone formation, which was hypothesized to be ascribed to the mineralisation of the organic bone matrix stimulation (Hott et al., 1993). The main known mechanism by which silicon interacts with cells is based on the activation of the Mitogen-Activated Protein Kinase-Extracellular signal-regulated kinase (MAPK-ERK) pathway in osteoblast-like cells (Shie et al., 2011). It has also been demonstrated that silicon ions may stimulate cell proliferation and osteogenic differentiation of hBMSCs through the activation of Wnt and sonic hedgehog (SHH)-related gene expression (Han et al., 2013). It has also been demonstrated that silicon promotes in vitro osteogenesis in osteoblasts through the activation of mTOR in the PI3K-Akt-mTOR pathway (Zhou et al., 2019). However, another study revealed that the inhibition of mTOR promoted osteogenic differentiation in MC3T3 cells, although this was using strontium ions (Cheng et al., 2019). A possible explanation to this contradiction could be that there are two types of mTOR complexes so they could be playing different roles in osteogenesis (Cheng et al., 2019).
Since silicon is generally present in combination with oxygen, forming silica, the most commonly used strategy for delivering silicon ions is incorporating them in ceramics, generally in the form of scaffolds, either as the bulk materials or as a coating, as well as in the form of NPs. A common strategy to prepare silica based materials is based on the sol-gel reaction of the silica precursors, such as tetraethyl orthosilicate or tetramethyl orthosilicate. Silica based microcarriers are prepared using the silica-based precursors that allowed the hardening of the sol in a water in oil emulsion, producing hardened microspheres after the complete condensation reaction. The microcarriers were shown to allow cell attachment and to induce osteogenic differentiation of MSCs, which was even more enhanced in the presence of other ions, such as calcium (Perez et al., 2014). Similar silica based structures have as well-been incorporated within calcium based ceramics. For instance, calcium-magnesium-silicon-containing bioceramics were compared with β-tricalcium phosphate (β-TCP) ceramics to analyze the osteogenic potential of hBMSCs cultured on the different scaffolds. The proliferation, ALP activity and the protein expression of COL1, ALP, OPN, BSP, and OCN was increased in the silicon based ceramics compared to the β -TCP due to the release of the different ions from the scaffolds. Interestingly, among the three different silicate based scaffolds, which were bredigite Ca7MgSi4O16, akermanite Ca2MgSi2O7 and diopside CaMgSi2O6, bredigite showed the highest osteogenic potential. The results showed a direct correlation between the scaffolds with the highest osteogenic potential with the amount of silicon ions present (Zhai et al., 2013). Although silicon has been shown to be one of the ions with the highest potential to induce the osteogenic differentiation compared to other ions, its combinations with other ions has been shown to demonstrate even higher synergistic effects. For instance, a bioceramic material containing silicon and strontium components was investigated in vitro. The results demonstrated that silicon ions alone had a higher induction effect on ALP activity than strontium ions, and that the combination of both ions resulted on the highest result of osteogenesis. Furthermore, mandibular defects of ovariectomized rat models were tested in vivo and demonstrated that bioceramics containing silicon and strontium promoted osteogenesis and mineralisation when compared with β-TCP bioceramics (Mao et al., 2017). This synergistic effect is probably explained by the different activation mechanisms of each ion, since silicon induces the hypoxia inducible factors (HIF) production whereas strontium interacts directly with other membrane receptors (Caverzasio and Thouverey, 2011).
Besides pristine ceramics, composite scaffolds have also been prepared incorporating different silicon-based ceramics within polymer structures. As an example, a siloxane-doped polylactic acid (PLA) and vaterite composite coated with hydroxycarbonate apatite (SPV-H) was used to study the osteogenic activity on MSCs for inducing mineralisation and differentiation. The experimental group was compared with the composite in the absence of silicon using PLA and vaterite as a control (PV-H). In general, MSCs cultured on SPV-H formed bone nodules after 21 days of culture with higher osteogenic differentiation compared to PV-H (Obata and Kasuga, 2009). Another example was observed when silicon ions were incorporated into collagen hydrogels via sol-gel reaction. These collagen/silica hybrid gels were able to increase apatite formation, as well as to induce the expression of Runx2, OPN and BSP osteogenic markers compared to collagen hydrogels (Yu et al., 2015). Moreover, silicon ions could also be incorporated as a coating on several types of surfaces by plasma enhanced chemical deposition. The release of silicon ions from these coatings resulted in an increase of Runx2, OCN, ALP and Col I gene expression in vitro. In vivo studies further confirmed the osteogenic effect of the coatings, showing the promotion of bone formation and mineralization (Ilyas et al., 2016; Xu et al., 2019).
Apart from the described 3D scaffolds, mesoporous silica NPs have attained great interest due to their ability to encapsulate molecules and hence stimulate different biological processes at the cellular level (Castillo et al., 2019). Silica NPs have been widely synthesized and have shown great potential as delivery cargo carriers for osteogenic induction (Kwon et al., 2013; Perez et al., 2017). These silica NPs are generally in the range of tens of nanometers and have been functionalized by coatings, tethering or co-precipitation of other ions. Readers are referred to previous reviews for a deep understanding of their synthesis and cellular effects (Kwon et al., 2013; Perez et al., 2017).
Silicon-containing glasses have been long used in bone tissue engineering, since their discovery by Larry Hench fifty years ago (Hench, 2006). It has been established that their biological activity comes from the release of biologically-active ions (Baino et al., 2018). Moreover, many new compositions and other types of bioglasses have been proposed for optimizing the body's response according to the specific clinical applications, incorporating other ions such as silver and strontium (Jones et al., 2016).
Zinc
Zinc is an essential trace metal that promotes osteoblastic proliferation and differentiation, stimulates mineralisation and inhibits osteoclastic cells through promoting bone cell proliferation, ALP activity, and collagen and protein synthesis (Jin et al., 2014; He et al., 2016; Paul et al., 2017). It is known to enhance bone metabolism through 1, 25-dihydroxyvitamin D3, a hormone which regulates calcium action (Yamaguchi et al., 1988; Park et al., 2018). Furthermore, zinc may promote ALP activity and collagen synthesis during osteogenesis. An in vitro study analyzed the effect of different zinc containing culture media on osteoblast differentiation in hBMSCs. The results showed that zinc promoted the expression of Runx2 via the cAMP-PKA-CREB pathway, which is an important regulator involved in the osteogenesis of MSCs (Zhu et al., 2017; Park et al., 2018). As previously described, this pathway is known to stimulate the expression of genes such as BMP2, which is well-known to be involved in osteogenesis and bone formation (Kim et al., 2013). Low concentrations of zinc (2 and 5 μg/mL) have also been described to stimulate osteogenesis in rBMSCs through the activation of MAPK-ERK pathway (Yu et al., 2020). However, higher zinc concentrations (15 μg/mL) have been shown to induce cell apoptosis by ROS generation (Yu et al., 2020).
Zinc has been generally added into biomaterials for orthopedic applications as an alternative to other biodegradable metals such as magnesium and iron as well as in ceramic based biomaterials (Zhu et al., 2017; Glenske et al., 2018). Regarding the metallic materials, these have been doped with zinc ions in the field of dental implantology. Titanium implants can be coated with zinc using a plasma immersion technique, which was previously shown to lead to an increased osteogenic activity (Jin et al., 2014). Interestingly, a recent study compared the osteogenic effect of incorporating zinc in titanium with two different coating techniques both in vitro and in vivo. One method incorporated zinc into the sub-surface of TiO2 coatings by plasma immersion ion implantation followed by deposition, whereas the other method consisted of a bulk-doped TiO2 by plasma electrolytic oxidation (Qiao et al., 2014). Both the in vitro and the in vivo results showed enhanced osteogenic behavior on the coating performed by plasma immersion ion implantation. The authors concluded that the samples that incorporated zinc on the surface of TiO2 implants, although they had a lower total zinc content, they had the best osteogenic potential and better bone regeneration activity both in vitro and in vivo, compared to the bulk doped samples. Hence, the results suggest that cells prefer the presence of zinc ions on the surface of the biomaterial rather than zinc ions in the surrounding medium (Qiao et al., 2014).
Nowadays, in a similar trend to other ions, it is becoming more common to incorporate zinc in ceramic based biomaterials. For instance, β-TCP ceramics were doped with three different concentrations of zinc (3, 6, and 9% w/w) with same silicon ion concentration (15% w/w silicon). Biological studies performed with mouse pre-osteoblasts showed adequate cell proliferation and osteogenic differentiation when cells were cultured on the low and intermediate content of zinc (3 and 6% w/w), whereas it was shown that higher zinc content (9% w/w) induced cell death (Paul et al., 2017). The zinc containing ceramics were then implanted into critical sized rabbit femoral condyle defects, showing lower bone formation with the higher zinc concentration (Paul et al., 2017). In a similar way, zinc containing TCP with different concentrations (0, 5, 15, and 45 mmol ZnCl2/100 mg TCP) were tested. The results showed that the highest zinc amount promoted the ALP activity of hBMSCs compared to the pristine ceramic. Furthermore, the implantation of the ceramics in the paraspinal muscle of canines for 12 weeks resulted in ectopic bone formation in a dose dependent manner, showing as well no bone formation in the pristine ceramic (Luo et al., 2014). Similarly, the calcium phosphate-based ceramics can be shaped into microsphere with the help of polymeric materials showing as well positive cell osteogenic stimulation in the presence of zinc. Interestingly, the ability to simulate bone regeneration in vivo was shown to be highest in the presence of zinc regardless of the presence or absence of collagen and calcium phosphate (Yu et al., 2017).
Strontium
Strontium is known to increase bone formation and inhibit osteoclast activity and has been proposed as a promising therapy for osteoporosis treatment and prevention (Peng et al., 2009; Aimaiti et al., 2017; Zeng et al., 2020). It has been commonly described that strontium promoted osteogenesis through the activation of the Wnt/β-catenin pathway, increasing Glycogen synthase kinase 3 beta (GSK3β) phosphorylation. This allows two mechanisms: the degradation of phosphorylated β-catenin and the translocation of β-catenin into the nucleus increasing the expression of Runx2 (Yang et al., 2011; Zeng et al., 2020). Another study evaluated that the activation of this pathway was performed through the activation of the calcium-sensing receptor (CaSR), which increases the release of calcium and, subsequently, activates the Wnt signal pathway (Saidak and Marie, 2012). It has also been suggested that strontium activates the Ras/MAPK signaling pathway, subsequently leading to Runx2 transcription downstream (Peng et al., 2009).
Strontium has been mainly incorporated in ceramics or in composite biomaterials. In the case of ceramics, when strontium was added into bioactive glasses, it enhanced the osteogenic gene expression in rat MSCs (Santocildes-Romero et al., 2015). Not only has strontium been able to promote osteogenesis, but also has been shown to reduce the osteoclastic activity (Tian et al., 2018). Their results have not only been shown in vitro, but also in vivo (Chandran et al., 2018; Li J. et al., 2018). While ceramics are an interesting and an appealing source for bone regeneration, composite materials are as well an interesting type of materials to dope with strontium. A nanocomposite fibrous scaffold made of poly(ε-caprolactone) (PCL) doped with strontium carbonate NPs increased the proliferation of human MSCs, as well as the expression of BMP2, Runx2 and Osterix in the mRNA and protein level, and also increased the deposition of calcium minerals even with the absence of osteogenic factors (Meka et al., 2016). Similarly, strontium doped composites have shown an enhanced bone regeneration (Zhang Y. et al., 2015; Henriques Lourenço et al., 2017). For instance, strontium-doped HA microspheres embedded in strontium-crosslinked RGD-alginate hydrogel were implanted in vivo in a rat critical-sized bone defect model and showed increased bone formation, stimulating the cell migration and collagen deposition (Henriques Lourenço et al., 2017).
Magnesium
Magnesium and its alloys have been used for biomedical applications for over 2 centuries and it has been observed that its degradation products (magnesium particles and ions) enhance new bone formation. Magnesium ions have been shown to increase ALP activity, matrix mineralisation and the expression of differentiation markers such as Runx2, BMP2, OPN, and COL1 in osteoblasts and MSCs (Yoshizawa et al., 2014; Díaz-Tocados et al., 2017; Wang J. et al., 2017; Zhang X. et al., 2017). Magnesium ions have been described to enhance bone formation mainly through the activation of MAPK-ERK pathway and regulate osteogenic gene expression (Wang et al., 2018, 2020; Yan et al., 2019; Qi et al., 2020). Other molecular pathway activated by magnesium ions is Wnt/β-catenin pathway, which phosphorylates GSK3β as described above and results in an increased osteogenic gene expression (Wang et al., 2018; Hung et al., 2019; Park et al., 2019). It has also been determined that magnesium ions are able to stimulate the activation of Notch pathway, which acts as a regulator of osteogenesis although its exact role in this process remains controversial (Engin and Lee, 2010; Long, 2012; Díaz-Tocados et al., 2017). PI3K pathway is also activated by magnesium ions that enter the cell through TRPM7 ion channel, but the exact molecular mechanisms are still unknown (Zhang X. et al., 2017).
When incorporated and released from biomaterials, magnesium ions have been shown to have different effects. For example, magnesium was incorporated into macroporous scaffolds by hydrothermal calcination. The released magnesium ions were able to promote the osteogenic differentiation of MC3T3-E1 pre-osteoblasts. When calcium was incorporated in combination with magnesium, the surface of these scaffolds presented nano-crystal microstructures with topographical and morphological features that could regulate cell adhesion and could increase even more the expression of the osteogenic markers. In the absence of calcium, although these structures were not observed, the surface roughness was increased compared to the unloaded scaffold (Pan et al., 2015; Chu et al., 2018). Magnesium has also been combined with CPC with different ratios of magnesium to calcium. Interestingly, the greater surface roughness and proper wettability of magnesium/CPC combined with a moderate calcium/magnesium ratio allowed the adhesion of fibronectin in a conformation that enhanced cell attachment and promoted the expression of integrin α5β1 in rBMSCs, which is also involved in cell adhesion. Moreover, the presence of magnesium increased the osteogenic differentiation of these cells compared to the pristine based CPC (Zhang J. et al., 2015). Despite its positive results, further experiments are required to determine whether the increased osteogenic differentiation was due to the effect of the released ions or due to the nanotopograhical structure. PLA/Magnesium composites with evenly distributed magnesium particles embedded in the PLA matrix have been developed by mixing pure magnesium powder with particle sizes about 100 μm with previously dissolved PLA. MC3T3-E1 cells were able to adhere to the surface of the composite, showing that the release of magnesium ions from the material promoted matrix mineralisation (Zhao et al., 2017).
Despite the fact that magnesium is generally incorporated in the presence of other ions, its superior activity in terms of cell adhesion and osteogenic differentiation has been widely studied. For instance, magnesium or strontium were incorporated within the structure of layered titanates. This material was developed using hydrothermal alkaline treatment with NaOH to form titanate nanofibers on commercially available titanium discs. The ions were incorporated by ionic exchange with Na+ ions. The exchange efficiency of strontium was higher than the possible doses of magnesium, leading to a lower release rate of magnesium ions. Nevertheless, the cell-material interaction results showed that despite its lower concentrations, magnesium promoted to a higher extent the expression of osteogenic markers Runx2, OPN and OCN in MC3T3-E1 pre-osteoblast cell line in a direct cell culture (Song et al., 2018).
Nanoparticle based systems have also shown to be an effective source of magnesium for its sustained release. In this sense, self-assembled bisphosphonate-magnesium NPs were incorporated within a hyaluronic acid based hydrogel. The formation of bisphosphonate-magnesium NPs was done by simple mixing of methacrylated hyaluronic acid, acrylated bisphosphonate and MgCl2 at 10, 100, and 250 mM concentrations. Subsequently, the free acrylated groups of the NPs and methacrylated groups in the hyaluronic acid were crosslinked using UV light. In order to verify the osteogenic capacity of the composite hydrogel, MSCs were loaded within the 3D hydrogels, showing that the material promoted in vitro mineralisation and an upregulation of osteogenic genes, which was then confirmed in vivo by implanting the hydrogel in craniotomy defects created in Sprague Dawley rats (Zhang K. et al., 2017).
Boron
Boron is a trace element that is essential for several biological processes such as embryogenesis, immune function, cognitive functions and bone growth and maintenance (Nielsen, 2000). Several studies have shown that boron supplementation could have a beneficial effect in bone health, improving bone strength and microstructure (Devirian and Volpe, 2003; Dessordi et al., 2017). Boron ions have been demonstrated to enhance osteogenesis through the activation of Wnt/β-catenin pathway; in particular, osteogenesis stimulated by boron is promoted by a transcription factor (TCF7L2), which is up-regulated by the activation of the Wnt pathway (Yin et al., 2020). Hence, the delivery of this ion from biomaterials could be a promising therapeutic approach for bone regeneration.
For example, boron ions released from boron nitride nanotubes increased protein adsorption, and MSCs attachment, ALP activity and OCN protein content, indicating that the nanotubes were able to induce osteogenesis (Li et al., 2016). Boron containing coatings have also shown promising results regarding their osteogenic potential. One study showed that a boron doped HA coated chitosan scaffold promoted cell adhesion, ECM synthesis and Runx2, OCN and OPN differentiation marker expression in MC3T3-E1 cells, suggesting that they are able to support osteoblastic differentiation and mineralisation (Tunçay et al., 2017). Another study prepared a boron-containing bioceramic coating generated by micro-arc oxidation onto titanium discs and was able to release low concentrations of boron ions that induced an increase in ALP activity (Ying et al., 2018).
Boron has not only been incorporated into coatings, but also has been integrated in scaffolds. For instance, boron containing chitosan NPs blended within a chitosan scaffold were able to increase osteogenic expression of osteoblastic cells cultured on the scaffolds (Gumusderelioglu et al., 2015). Boron has as well-been directly incorporated into bioactive glass scaffolds (Wu et al., 2011). In vitro studies using hBMSCs showed that the scaffolds highly activated Wnt/β-catenin pathway, increasing the presence of β-catenin, p-GSK3β and Setd7 at the protein level. To determine the role of Setd7, a knock down of this protein was performed, observing a reduction in the mineralisation activity and in the expression of osteogenic markers ALP and Runx2 when cells were stimulated with B-MBG, suggesting that promoting osteoblast differentiation is a Setd7 dependent process when cells were stimulated with boron (Yin et al., 2018). However, more research is needed to evaluate if other factors are involved during boron-induced osteoblast differentiation.
Lithium
Lithium is a microelement present in the human body that has been evaluated to enhance osteoblast proliferation and bone formation (Ma et al., 2019). It has been reported that the molecular mechanism stimulated by lithium is through the activation of the Wnt/β-catenin pathway, which promotes osteogenic differentiation enhancing OCN production and increasing ALP activity (Li D. et al., 2018; Huang et al., 2019).
Lithium doped mesoporous silica nanospheres have been created using sol-gel method showing the ability to rapidly release lithium ions within the first 3 days followed by a sustained release until 14 days. Controlled release of lithium increased cell proliferation, differentiation and the expression of Runx2, OCN, OPN, and ALP in rBMSCs compared to bare mesoporous silica nanospheres (Zhang et al., 2018). A lithium containing coating has been used on a scaffold made of entangled titanium wire in order to enhance bone regeneration. The coatings were developed by micro-arc oxidation of the titanium wire using concentrations of lithium chloride of 0.01 M (L1-MAO) or 0.02 M (L2-MAO). The release of lithium ions from L2-MAO coating was higher compared to L1-MAO coating and it was observed that only L2-MAO was able to enhance MG-63 osteoblast adhesion and spreading, ALP activity, and it up-regulated the expression of Runx2, as well as early (COL1A1 and OPN), middle (ALP) and late (OCN) markers of osteogenesis (Liu et al., 2018).
Lithium chloride dissolved in citric acid at different concentrations (0, 50, 100, and 200 mM) has also been combined with a 1:1 mixture of tetracalcium phosphate and dicalcium phosphate to form a CPC (Li/CPC). A lithium release of up to 60 mg/L from Li/CPC increased the proliferation, ALP activity, mineralisation and expression of COL1A1 and Runx2 in MC3T3-E1 cells through the activation of Wnt/β-catenin pathway. In vivo, this material increased the formation of new bone compared to CPC and accelerated bone formation in an ovariectomized rat model (Li et al., 2017). Lithium ions have been recently incorporated into bioactive glass NPs, but their effect on osteogenesis still remains unknown and more studies should be performed (El-Kady et al., 2016).
Conclusions and Future Perspectives
Overall, diverse ions have been shown to have different effects on the different osteogenic actions. Figure 5 summarizes the differential effect for each of the steps considered in bone regeneration, mainly covering the ability of ions to induce cell proliferation, to trigger initial, middle or late osteogenesis, as well as the capacity for the different ions to inhibit osteoclastogenesis.

Figure 5. Schematic illustration of the effect of ions during osteoblastogenesis and osteoclastogenesis. First, cell proliferation is stimulated by boron and silicon. Then, zinc, silicon, strontium, boron and magnesium enhance the expression of early genes such as Runx2 and Osx. Later, zinc, boron, magnesium, silicon and lithium stimulate the gene expression of intermediate markers like ALP and COL1. Late gene expression markers OCN and OPN are stimulated by zinc, lithium and silicon. Osteoclastogenesis is inhibited by zinc, silicon, magnesium and strontium. B, boron; Li, Lithium; Mg, magnesium; Si, silicon; Sr, Strontium; Zn, Zinc.
The recent development of biomaterials that incorporate and release different ions has shown great potential to trigger bone tissue regeneration. While the effective doses vary significantly among these ions, these are still far below the concentrations of growth factors and other molecules needed to produce similar tissue regeneration process. Furthermore, their inorganic nature allows a more stable and prolonged effect compared to other biological molecules. Nevertheless, few similar developments have been established for the sustained and controlled release of ions. For this purpose, further methodologies should be established on how to control on demand ion delivery as well as how to obtain sustained on delivery systems. Currently, significant controversial issues arise from the optimum doses and the release time points, which limits the proper incorporation within biomaterials. Moreover, in order to obtain greater insights of their potential, deeper understating of the signaling pathways involved is needed for the different ions in order to clearly identify the doses required as well as the time points in which these are needed. Overall, the incorporation of ions in biomaterials and their release are promising in providing signaling cues for bone tissue regeneration.
Author Contributions
EB-R, LD-T, and BG-K contributed to literature search. EB-R, LD-T, BG-K, LD, BB, and MH-N contributed to writing. MM-T, PT, FG, and RP contributed to reviewing and editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Government of Catalonia (2017 SGR 708), the Spanish Ministry (Ramón y Cajal fellowship (RYC2018-025977-I) and project RTI2018-096088-J-100 (MINECO/FEDER), and pre-doctoral fellowship from Universitat Internacional de Catalunya (UIC).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Alex Mas for his support in figure preparation.
Abbreviations
ALP, Alkaline phosphatase; BMPs, Bone morphogenetic proteins; BSP, Bone sialoprotein; CaSR, Calcium sensing receptor; COL1, Collagen type I; CPC, Calcium Phosphate Cements; ECM, Extracellular matrix; ERK, Extracellular signal-regulated kinase; FGFs, Fibroblast growth factors; GSK3β, Glycogen synthase kinase 3 beta; HA, Hydroxyapatite; hBMSCs, Human bone marrow stromal cells; HIF, Hypoxia inducible factors; MAPK, Mitogen-Activated Protein Kinase; MPs, Microparticles; MSCs, Mesenchymal stromal cells; NPs, Nanoparticles; OCN, Osteocalcin; OPN, Osteopontin; PCL, Poly(ε-caprolactone); PDGFs, Platelet derived growth factors; PLA, Polylactic acid; rBMSCs, Rat bone marrow stromal cells; SHH, Sonic hedgehog; TCP, Tricalcium phosphate; TGF-β, Transforming growth factor β; Wnt, Wingless-related integration site.
References
Aaseth, J., Boivin, G., and Andersen, O. (2012). Osteoporosis and trace elements - an overview. J. Trace Elem. Med. Biol. 26, 149–152. doi: 10.1016/j.jtemb.2012.03.017
Aimaiti, A., Maimaitiyiming, A., Boyong, X., Aji, K., Li, C., and Cui, L. (2017). Low-dose strontium stimulates osteogenesis but high-dose doses cause apoptosis in human adipose-derived stem cells via regulation of the ERK1/2 signaling pathway. Stem Cell Res. Ther. 8, 1–12. doi: 10.1186/s13287-017-0726-8
Alizadeh-Osgouei, M., Li, Y., and Wen, C. (2019). A comprehensive review of biodegradable synthetic polymer-ceramic composites and their manufacture for biomedical applications. Bioact. Mater. 4, 22–36. doi: 10.1016/j.bioactmat.2018.11.003
Alrabeah, G. O., Brett, P., Knowles, J. C., and Petridis, H. (2017). The effect of metal ions released from different dental implant-abutment couples on osteoblast function and secretion of bone resorbing mediators. J. Dent. 66, 91–101. doi: 10.1016/j.jdent.2017.08.002
Bai, X., Gao, M., Syed, S., Zhuang, J., Xu, X., and Zhang, X. Q. (2018). Bioactive hydrogels for bone regeneration. Bioact. Mater. 3, 401–417. doi: 10.1016/j.bioactmat.2018.05.006
Baino, F., Hamzehlou, S., and Kargozar, S. (2018). Bioactive glasses: where are we and where are we going? J. Funct. Biomater. 9:25. doi: 10.3390/jfb9010025
Behr, B., Leucht, P., Longaker, M. T., and Quarto, N. (2010). Fgf-9 is required for angiogenesis and osteogenesis in long bone repair. Proc. Natl. Acad. Sci. U. S. A. 107, 11853–11858. doi: 10.1073/pnas.1003317107
Betz, R. R. (2002). Limitations of autograft and allograft: new synthetic solutions. Orthopedics 25, 561–570. doi: 10.3928/0147-7447-20020502-04
Birgani, Z. T., Gharraee, N., Malhotra, A., Van Blitterswijk, C. A., and Habibovic, P. (2016). Combinatorial incorporation of fluoride and cobalt ions into calcium phosphates to stimulate osteogenesis and angiogenesis. Biomed. Mater. 11:015020. doi: 10.1088/1748-6041/11/1/015020
Burghardt, I., Lüthen, F., Prinz, C., Kreikemeyer, B., Zietz, C., Neumann, H. G., et al. (2015). A dual function of copper in designing regenerative implants. Biomaterials 44, 36–44. doi: 10.1016/j.biomaterials.2014.12.022
Castaño, O., Pérez, S., Mateos-Timoneda, M. A., and Engel, E. (2017). “Chapter 13: cell interactions with calcium phosphate glasses,” in RSC Smart Materials, eds H.-J. Schneider and M. Shahinpoor (London: Royal Society of Chemistry), 303–315. doi: 10.1039/9781782622017-00303
Castillo, R. R., Lozano, D., González, B., Manzano, M., Izquierdo-Barba, I., and Vallet-Regí, M. (2019). Advances in mesoporous silica nanoparticles for targeted stimuli-responsive drug delivery: an update. Expert Opin. Drug Deliv. 16, 415–439. doi: 10.1080/17425247.2019.1598375
Caverzasio, J., and Thouverey, C. (2011). Activation of FGF receptors is a new mechanism by which strontium ranelate induces osteoblastic cell growth. Cell. Physiol. Biochem. 27, 243–250. doi: 10.1159/000327950
Chandran, S., Shenoy, S. J., Babu, S. S. P., Nair, R., et al. (2018). Strontium hydroxyapatite scaffolds engineered with stem cells aid osteointegration and osteogenesis in osteoporotic sheep model. Coll. Surf. B Biointerf. 163, 346–354. doi: 10.1016/j.colsurfb.2017.12.048
Chen, Y., Bal, B. S., and Gorski, J. P. (1992). Calcium and collagen binding properties of osteopontin, bone sialoprotein, and bone acidic glycoprotein-75 from bone. J. Biol. Chem. 267, 24871–24878.
Cheng, Y., Huang, L., Wang, Y., Huo, Q., Shao, Y., Bao, H., et al. (2019). Strontium promotes osteogenic differentiation by activating autophagy via the the AMPK/mTOR signaling pathway in MC3T3-E1 cells. Int. J. Mol. Med. 44, 652–660. doi: 10.3892/ijmm.2019.4216
Cho, T.-J., Gerstenfeld, L. C., and Einhorn, T. A. (2002). Differential temporal expression of members of the transforming growth factor β superfamily during murine fracture healing. J. Bone Miner. Res. 17, 513–520. doi: 10.1359/jbmr.2002.17.3.513
Chocholata, P., Kulda, V., and Babuska, V. (2019). Fabrication of scaffolds for bone-tissue regeneration. Materials 12:568. doi: 10.3390/ma12040568
Chu, L., Jiang, G., Hu, X., Le, J. T. D., He, X. P., Li, Y., et al. (2018). Biodegradable macroporous scaffold with nano-crystal surface microstructure for highly effective osteogenesis and vascularization. J. Mater. Chem. B 6, 1658–1667. doi: 10.1039/C7TB03353B
Dessordi, R., Spirlandeli, A. L., Zamarioli, A., Volpon, J. B., and Navarro, A. M. (2017). Boron supplementation improves bone health of non-obese diabetic mice. J. Trace Elem. Med. Biol. 39, 169–175. doi: 10.1016/j.jtemb.2016.09.011
Devirian, T. A., and Volpe, S. L. (2003). The physiological effects of dietary boron. Crit. Rev. Food Sci. Nutr. 43, 219–231. doi: 10.1080/10408690390826491
Díaz-Tocados, J. M., Herencia, C., Martínez-Moreno, J. M., Montes de Oca, A., Rodríguez-Ortiz, M. E., Vergara, N., et al. (2017). Magnesium chloride promotes osteogenesis through notch signaling activation and expansion of mesenchymal stem cells. Sci. Rep. 7, 1–12. doi: 10.1038/s41598-017-08379-y
Dimitriou, R., Tsiridis, E., and Giannoudis, P. V. (2005). Current concepts of molecular aspects of bone healing. Injury 36, 1392–1404. doi: 10.1016/j.injury.2005.07.019
D'Mello, S., Elangovan, S., Hong, L., Ross, R. D., Sumner, D. R., and Salem, A. K. (2015). Incorporation of copper into chitosan scaffolds promotes bone regeneration in rat calvarial defects. J. Biomed. Mater. Res. Part B Appl. Biomater. 103, 1044–1049. doi: 10.1002/jbm.b.33290
Dobbenga, S., Fratila-Apachitei, L. E., and Zadpoor, A. A. (2016). Nanopattern-induced osteogenic differentiation of stem cells – a systematic review. Acta Biomater. 46, 3–14. doi: 10.1016/j.actbio.2016.09.031
Dominici, M., Le Blanc, K., Mueller, I., Slaper-Cortenbach, I., Marini, F., Krause, D. S., et al. (2006). Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8, 315–317. doi: 10.1080/14653240600855905
Drury, J. L., and Mooney, D. J. (2003). Hydrogels for tissue engineering: scaffold design variables and applications. Biomaterials 24, 4337–4351. doi: 10.1016/S0142-9612(03)00340-5
Drynda, A., Drynda, S., Kekow, J., Lohmann, C. H., and Bertrand, J. (2018). Differential effect of cobalt and chromium ions as well as cocr particles on the expression of osteogenic markers and osteoblast function. Int. J. Mol. Sci. 19:3034. doi: 10.3390/ijms19103034
El-Kady, A. M., Farag, M. M., and El-Rashedi, A. M. I. (2016). Bioactive glass nanoparticles designed for multiple deliveries of lithium ions and drugs: curative and restorative bone treatment. Eur. J. Pharm. Sci. 91, 243–250. doi: 10.1016/j.ejps.2016.05.004
Engin, F., and Lee, B. (2010). NOTCHing the bone: insights into multi-functionality. Bone 46, 274–280. doi: 10.1016/j.bone.2009.05.027
Engler, A. J., Sen, S., Sweeney, H. L., and Discher, D. E. (2006). Matrix elasticity directs stem cell lineage specification. Cell 126, 677–689. doi: 10.1016/j.cell.2006.06.044
Evis, Z., and Webster, T. J. (2011). Nanosize hydroxyapatite: doping with various ions. Adv. Appl. Ceram. 110, 311–320. doi: 10.1179/1743676110Y.0000000005
Fakhry, M., Hamade, E., Badran, B., Buchet, R., and Magne, D. (2013). Molecular mechanisms of mesenchymal stem cell differentiation towards osteoblasts. World J. Stem Cells 5, 136–148. doi: 10.4252/wjsc.v5.i4.136
Fedde, K. N., Blair, L., Silverstein, J., Coburn, S. P., Ryan, L. M., Weinstein, R. S., et al. (1999). Alkaline phosphatase knock-out mice recapitulate the metabolic and skeletal defects of infantile hypophosphatasia. J. Bone Miner. Res. 14, 2015–2026. doi: 10.1359/jbmr.1999.14.12.2015
Florencio-Silva, R., Rodrigues Da, G., Sasso, S., Sasso-Cerri, E., Simões, M. J., and Cerri, P. S. (2015). Biology of bone tissue: structure, function, and factors that influence bone cells. Biomed. Res. Int. 2015:421746. doi: 10.1155/2015/421746
Fukada, T., Hojyo, S., and Furuichi, T. (2013). Zinc signal: a new player in osteobiology. J. Bone Miner. Metab. 31, 129–135. doi: 10.1007/s00774-012-0409-6
Garley, A., Hoff, S. E., Saikia, N., Jamadagni, S., Baig, A., and Heinz, H. (2019). Adsorption and substitution of metal ions on hydroxyapatite as a function of crystal facet and electrolyte pH. J. Phys. Chem. C 123, 16982–16993. doi: 10.1021/acs.jpcc.9b02808
Glenske, K., Donkiewicz, P., Köwitsch, A., Milosevic-Oljaca, N., Rider, P., Rofall, S., et al. (2018). Applications of metals for bone regeneration. Int. J. Mol. Sci. 19:826. doi: 10.20944/preprints201802.0051.v1
Golub, E. E. (2009). Role of matrix vesicles in biomineralization. Biochim. Biophys. Acta - Gen. Subj. 1790, 1592–1598. doi: 10.1016/j.bbagen.2009.09.006
Gumusderelioglu, M., Tunçay, E. T., Kaynak, G., Demirtaş, T. T., Aydin, S. T., and Hakki, S. S. (2015). Encapsulated boron as an osteoinductive agent for bone scaffolds. J. Trace Elem. Med. Biol. 31, 120–128. doi: 10.1016/j.jtemb.2015.03.008
Habraken, W., Habibovic, P., Epple, M., and Bohner, M. (2016). Calcium phosphates in biomedical applications: materials for the future? Mater. Today 19, 69–87. doi: 10.1016/j.mattod.2015.10.008
Hallab, N. J., Bundy, K. J., O'Connor, K., Moses, R. L., and Jacobs, J. J. (2001). Evaluation of metallic and polymeric biomaterial surface energy and surface roughness characteristics for directed cell adhesion. Tissue Eng. 7, 55–71. doi: 10.1089/107632700300003297
Han, P., Wu, C., and Xiao, Y. (2013). The effect of silicate ions on proliferation, osteogenic differentiation and cell signalling pathways (WNT and SHH) of bone marrow stromal cells. Biomater. Sci. 1, 379–392. doi: 10.1039/C2BM00108J
Hashimoto, R. (2020). “Extracellular Ca2+ in bone marrow,” in Advances in Experimental Medicine and Biology, eds W. E. Crusio, H. Dong, H. H. Radeke, N. Rezaei, and J. Xiao (New York, NY: Springer New York LLC), 1065–1078. doi: 10.1007/978-3-030-12457-1_42
He, M., Chen, X., Cheng, K., Weng, W., and Wang, H. (2016). Enhanced osteogenic activity of TiO2 nanorod films with microscaled distribution of Zn-CaP. ACS Appl. Mater. Interfaces 8, 6944–6952. doi: 10.1021/acsami.6b01284
Hench, L. L. (2006). The story of Bioglass®. J. Mater. Sci. Mater. Med. 17, 967–978. doi: 10.1007/s10856-006-0432-z
Henriques Lourenço, A., Neves, N., Ribeiro-Machado, C., Sousa, S. R., Lamghari, M., Barrias, C. C., et al. (2017). Injectable hybrid system for strontium local delivery promotes bone regeneration in a rat critical-sized defect model. Sci. Rep. 7, 1–15. doi: 10.1038/s41598-017-04866-4
Hoppe, A., Güldal, N. S., and Boccaccini, A. R. (2011). A review of the biological response to ionic dissolution products from bioactive glasses and glass-ceramics. Biomaterials 32, 2757–2774. doi: 10.1016/j.biomaterials.2011.01.004
Hott, M., de Pollak, C., Modrowski, D., and Marie, P. J. (1993). Short-term effects of organic silicon on trabecular bone in mature ovariectomized rats. Calcif. Tissue Int. 53, 174–179. doi: 10.1007/BF01321834
Huang, T., Ben, L. Y. Z., Yu, K., Yu, Z., Wang, Y., Jiang, Z. W., et al. (2019). Effect of the Wnt signal-RANKL/OPG axis on the enhanced osteogenic integration of a lithium incorporated surface. Biomater. Sci. 7, 1101–1116. doi: 10.1039/C8BM01411F
Hung, C. C., Chaya, A., Liu, K., Verdelis, K., and Sfeir, C. (2019). The role of magnesium ions in bone regeneration involves the canonical Wnt signaling pathway. Acta Biomater. 98, 246–255. doi: 10.1016/j.actbio.2019.06.001
Ignjatovic, N., Ajdukovic, Z., Rajkovic, J., Najman, S., Mihailovic, D., and Uskokovic, D. (2015). Enhanced osteogenesis of nanosized cobalt-substituted hydroxyapatite. J. Bionic Eng. 12, 604–612. doi: 10.1016/S1672-6529(14)60150-5
Ignjatović, N., Ajduković, Z., Savić, V., Najman, S., Mihailović, D., Vasiljević, P., et al. (2013). Nanoparticles of cobalt-substituted hydroxyapatite in regeneration of mandibular osteoporotic bones. J. Mater. Sci. Mater. Med. 24, 343–354. doi: 10.1007/s10856-012-4793-1
Ilyas, A., Odatsu, T., Shah, A., Monte, F., Kim, H. K. W., Kramer, P., et al. (2016). Amorphous silica: a new antioxidant role for rapid critical-sized bone defect healing. Adv. Healthc. Mater. 5, 2199–2213. doi: 10.1002/adhm.201600203
Jammalamadaka, U., and Tappa, K. (2018). Recent advances in biomaterials for 3D printing and tissue engineering. J. Funct. Biomater. 9:4. doi: 10.3390/jfb9010022
Jeon, E., Yun, Y.-R., Kang, W., Lee, S., Koh, Y.-H., Kim, H.-W., et al. (2012). Investigating the role of FGF18 in the cultivation and osteogenic differentiation of mesenchymal stem cells. PLoS ONE 7:e43982. doi: 10.1371/annotation/77eec9d5-379f-4895-8b85-308ee29facf4
Jin, G., Cao, H., Qiao, Y., Meng, F., Zhu, H., and Liu, X. (2014). Osteogenic activity and antibacterial effect of zinc ion implanted titanium. Coll. Surf. B Biointerf. 117, 158–165. doi: 10.1016/j.colsurfb.2014.02.025
Jones, J. R., Brauer, D. S., Hupa, L., and Greenspan, D. C. (2016). Bioglass and bioactive glasses and their impact on healthcare. Int. J. Appl. Glas. Sci. 7, 423–434. doi: 10.1111/ijag.12252
Jugdaohsingh, R., Anderson, S. H. C., Tucker, K. L., Elliott, H., Kiel, D. P., Thompson, R. P. H., et al. (2002). Dietary silicon intake and absorption. Am. J. Clin. Nutr. 75, 887–893. doi: 10.1093/ajcn/75.5.887
Kaya, S., Cresswell, M., and Boccaccini, A. R. (2018). Mesoporous silica-based bioactive glasses for antibiotic-free antibacterial applications. Mater. Sci. Eng. C 83, 99–107. doi: 10.1016/j.msec.2017.11.003
Kempen, D. H. R., Creemers, L. B., Alblas, J., Lu, L., Verbout, A. J., Yaszemski, M. J., et al. (2010). Growth factor interactions in bone regeneration. Tissue Eng. Part B Rev. 16, 551–566. doi: 10.1089/ten.teb.2010.0176
Kim, J. M., Choi, J. S., Kim, Y. H., Jin, S. H., Lim, S., Jang, H. J., et al. (2013). An activator of the cAMP/PKA/CREB pathway promotes osteogenesis from human mesenchymal stem cells. J. Cell. Physiol. 228, 617–626. doi: 10.1002/jcp.24171
Kose, N., Otuzbir, A., Pekşen, C., Kiremitçi, A., and Dogan, A. (2013). “A silver ion-doped calcium phosphate-based ceramic nanopowder-coated prosthesis increased infection resistance basic research,” in Clinical Orthopaedics and Related Research, ed S. S. Leopold (New York, NY: Springer New York LLC), 2532–2539. doi: 10.1007/s11999-013-2894-x
Kulanthaivel, S., Roy, B., Agarwal, T., Giri, S., Pramanik, K., Pal, K., et al. (2016). Cobalt doped proangiogenic hydroxyapatite for bone tissue engineering application. Mater. Sci. Eng. C 58, 648–658. doi: 10.1016/j.msec.2015.08.052
Kwon, S., Singh, R. K., Perez, R. A., Neel, E. A. A., Kim, H.-W., and Chrzanowski, W. (2013). Silica-based mesoporous nanoparticles for controlled drug delivery. J. Tissue Eng. 4:2041731413503357. doi: 10.1177/2041731413503357
Kyle, S., Jessop, Z. M., Al-Sabah, A., and Whitaker, I. S. (2017). “Printability” of candidate biomaterials for extrusion based 3D printing: state-of-the-art. Adv. Healthc. Mater. 6:1700264. doi: 10.1002/adhm.201700264
Levato, R., Visser, J., Planell, J. A., Engel, E., Malda, J., and Mateos-Timoneda, M. A. (2014). Biofabrication of tissue constructs by 3D bioprinting of cell-laden microcarriers. Biofabrication 6:035020. doi: 10.1088/1758-5082/6/3/035020
Li, D., Huifang, L., Zhao, J., Yang, Z., Xie, X., Wei, Z., et al. (2018). Porous lithium-doped hydroxyapatite scaffold seeded with hypoxia-preconditioned bone-marrow mesenchymal stem cells for bone-tissue regeneration. Biomed. Mater. 13:055002. doi: 10.1088/1748-605X/aac627
Li, J., Yang, L., Guo, X., Cui, W., Yang, S., Wang, J., et al. (2018). Osteogenesis effects of strontium-substituted hydroxyapatite coatings on true bone ceramics surface in vitro and in vivo. Biomed. Mater. 13:015018. doi: 10.1088/1748-605X/aa89af
Li, L., Peng, X., Qin, Y., Wang, R., Tang, J., Cui, X., et al. (2017). Acceleration of bone regeneration by activating Wnt/β-catenin signalling pathway via lithium released from lithium chloride/calcium phosphate cement in osteoporosis. Sci. Rep. 7, 1–12. doi: 10.1038/srep45204
Li, S., Wang, M., Chen, X., Li, S. F., Li-Ling, J., and Xie, H. Q. (2014). Inhibition of osteogenic differentiation of mesenchymal stem cells by copper supplementation. Cell Prolif. 47, 81–90. doi: 10.1111/cpr.12083
Li, X., Wang, X., Jiang, X., Yamaguchi, M., Ito, A., Bando, Y., et al. (2016). Boron nitride nanotube-enhanced osteogenic differentiation of mesenchymal stem cells. J. Biomed. Mater. Res. - Part B Appl. Biomater. 104, 323–329. doi: 10.1002/jbm.b.33391
Li, Y., Wang, J., Wang, Y., Du, W., and Wang, S. (2018). Transplantation of copper-doped calcium polyphosphate scaffolds combined with copper (II) preconditioned bone marrow mesenchymal stem cells for bone defect repair. J. Biomater. Appl. 32, 738–753. doi: 10.1177/0885328217739456
Lin, Y., Xiao, W., Bal, B. S., and Rahaman, M. N. (2016). Effect of copper-doped silicate 13-93 bioactive glass scaffolds on the response of MC3T3-E1 cells in vitro and on bone regeneration and angiogenesis in rat calvarial defects in vivo. Mater. Sci. Eng. C 67, 440–452. doi: 10.1016/j.msec.2016.05.073
Liu, W., Chen, D., Jiang, G., Li, Q., Wang, Q., Cheng, M., et al. (2018). A lithium-containing nanoporous coating on entangled titanium scaffold can enhance osseointegration through Wnt/β-catenin pathway. Nanomed. Nanotechnol. Biol. Med. 14, 153–164. doi: 10.1016/j.nano.2017.09.006
Long, F. (2012). Building strong bones: molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 13, 27–38. doi: 10.1038/nrm3254
Lu, T., Qiao, Y., and Liu, X. (2012). Surface modification of biomaterials using plasma immersion ion implantation and deposition. Interface Focus 2, 325–336. doi: 10.1098/rsfs.2012.0003
Lu, X., Li, K., Xie, Y., Qi, S., Shen, Q., Yu, J., et al. (2019). Improved osteogenesis of boron incorporated calcium silicate coatings via immunomodulatory effects. J. Biomed. Mater. Res. - Part A 107, 12–24. doi: 10.1002/jbm.a.36456
Lu, Y., Li, L., Zhu, Y., Wang, X., Li, M., Lin, Z., et al. (2018). Multifunctional copper-containing carboxymethyl chitosan/alginate scaffolds for eradicating clinical bacterial infection and promoting bone formation. ACS Appl. Mater. Interfaces 10, 127–138. doi: 10.1021/acsami.7b13750
Luo, X., Barbieri, D., Davison, N., Yan, Y., De Bruijn, J. D., and Yuan, H. (2014). Zinc in calcium phosphate mediates bone induction: in vitro and in vivo model. Acta Biomater. 10, 477–485. doi: 10.1016/j.actbio.2013.10.011
Ma, Y., Li, Y., Hao, J., Ma, B., Di, T., and Dong, H. (2019). Evaluation of the degradation, biocompatibility and osteogenesis behavior of lithium-doped calcium polyphosphate for bone tissue engineering. Biomed. Mater. Eng. 30, 23–36. doi: 10.3233/BME-181030
MacDonald, D. E., Deo, N., Markovic, B., Stranick, M., and Somasundaran, P. (2002). Adsorption and dissolution behavior of human plasma fibronectin on thermally and chemically modified titanium dioxide particles. Biomaterials 23, 1269–1279. doi: 10.1016/S0142-9612(01)00317-9
Mao, L., Xia, L., Chang, J., Liu, J., Jiang, L., Wu, C., et al. (2017). The synergistic effects of Sr and Si bioactive ions on osteogenesis, osteoclastogenesis and angiogenesis for osteoporotic bone regeneration. Acta Biomater. 61, 217–232. doi: 10.1016/j.actbio.2017.08.015
Marsell, R., and Einhorn, T. A. (2009). The role of endogenous bone morphogenetic proteins in normal skeletal repair. Injury 40, S4–7. doi: 10.1016/S0020-1383(09)70003-8
Marsell, R., and Einhorn, T. A. (2011). The biology of fracture healing. Injury 42, 551–555. doi: 10.1016/j.injury.2011.03.031
Meka, S. R. K., Jain, S., and Chatterjee, K. (2016). Strontium eluting nanofibers augment stem cell osteogenesis for bone tissue regeneration. Coll. Surf. B Biointerf. 146, 649–656. doi: 10.1016/j.colsurfb.2016.07.012
Moore, W. R., Graves, S. E., and Bain, G. I. (2001). Synthetic bone graft substitutes. ANZ J. Surg. 71, 354–361. doi: 10.1046/j.1440-1622.2001.02128.x
Mouriño, V., Cattalini, J. P., and Boccaccini, A. R. (2012). Metallic ions as therapeutic agents in tissue engineering scaffolds: an overview of their biological applications and strategies for new developments. J. R. Soc. Interface 9, 401–419. doi: 10.1098/rsif.2011.0611
Murphy, S. V., and Atala, A. (2014). 3D bioprinting of tissues and organs. Nat. Biotechnol. 32, 773–785. doi: 10.1038/nbt.2958
Navarrete, R. O., Lee, E. M., Smith, K., Hyzy, S. L., Doroudi, M., Williams, J. K., et al. (2017). Substrate stiffness controls osteoblastic and chondrocytic differentiation of mesenchymal stem cells without exogenous stimuli. PLoS ONE 12:e0170312. doi: 10.1371/journal.pone.0170312
Nielsen, F. H. (2000). The emergence of boron as nutritionally important throughout the life cycle. Nutrition 16, 512–514. doi: 10.1016/S0899-9007(00)00324-5
Noumbissi, S., Scarano, A., and Gupta, S. (2019). A literature review study on atomic ions dissolution of titanium and its alloys in implant dentistry. Materials 12:368. doi: 10.3390/ma12030368
Obata, A., and Kasuga, T. (2009). Stimulation of human mesenchymal stem cells and osteoblasts activities in vitro on silicon-releasable scaffolds. J. Biomed. Mater. Res. - Part A 91, 11–17. doi: 10.1002/jbm.a.32181
Ofudje, E. A., Adeogun, A. I., Idowu, M. A., and Kareem, S. O. (2019). Synthesis and characterization of Zn-Doped hydroxyapatite: scaffold application, antibacterial and bioactivity studies. Heliyon 5:e01716. doi: 10.1016/j.heliyon.2019.e01716
Olmos Buitrago, J., Perez, R., El-Fiqi, A., Singh, R. K., Kim, J. H., et al. (2015). Core-shell fibrous stem cell carriers incorporating osteogenic nanoparticulate cues for bone tissue engineering. Acta Biomater. 28, 183–192. doi: 10.1016/j.actbio.2015.09.021
O'Neill, E., Awale, G., Daneshmandi, L., Umerah, O., and Lo, K. W. H. (2018). The roles of ions on bone regeneration. Drug Discov. Today 23, 879–890. doi: 10.1016/j.drudis.2018.01.049
Pablo Rodrguez, J., Ros, S., and Gonzlez, M. (2002). Modulation of the proliferation and differentiation of human mesenchymal stem cells by copper. J. Cell. Biochem. 85, 92–100. doi: 10.1002/jcb.10111
Palombella, S., Pirrone, C., Rossi, F., Armenia, I., Cherubino, M., Valdatta, L., et al. (2017). Effects of metal micro and nano-particles on hASCs: an in vitro model. Nanomaterials 7:212. doi: 10.3390/nano7080212
Pan, H., Xie, Y., Li, K., Hu, D., Zhao, J., Zheng, X., et al. (2015). ROCK-regulated synergistic effect of macropore/nanowire topography on cytoskeletal distribution and cell differentiation. RSC Adv. 5, 101834–101842. doi: 10.1039/C5RA19691D
Park, J.-H., Pérez, R. A., Jin, G.-Z., Choi, S.-J., Kim, H.-W., and Wall, I. B. (2013). Microcarriers designed for cell culture and tissue engineering of bone. Tissue Eng. - Part B Rev. 19, 172–190. doi: 10.1089/ten.teb.2012.0432
Park, J. W., Hanawa, T., and Chung, J. H. (2019). The relative effects of Ca and Mg ions on MSC osteogenesis in the surface modification of microrough Ti implants. Int. J. Nanomedicine 14, 5697–5711. doi: 10.2147/IJN.S214363
Park, K. H., Choi, Y., Yoon, D. S., Lee, K.-M., Kim, D., and Lee, J. W. (2018). Zinc promotes osteoblast differentiation in human mesenchymal stem cells via activation of the cAMP-PKA-CREB signaling pathway. Stem Cells Dev. 27, 1125–1135. doi: 10.1089/scd.2018.0023
Patntirapong, S., Habibovic, P., and Hauschka, P. V. (2009). Effects of soluble cobalt and cobalt incorporated into calcium phosphate layers on osteoclast differentiation and activation. Biomaterials 30, 548–555. doi: 10.1016/j.biomaterials.2008.09.062
Paul, K., Lee, B. Y., Abueva, C., Kim, B., Choi, H. J., Bae, S. H., et al. (2017). In vivo evaluation of injectable calcium phosphate cement composed of Zn- and Si-incorporated β-tricalcium phosphate and monocalcium phosphate monohydrate for a critical sized defect of the rabbit femoral condyle. J. Biomed. Mater. Res. - Part B Appl. Biomater. 105, 260–271. doi: 10.1002/jbm.b.33537
Peng, S., Zhou, G., Luk, K. D. K., Cheung, K. M. C., Li, Z., Lam, W. M., et al. (2009). Strontium promotes osteogenic differentiation of mesenchymal stem cells through the Ras/MAPK signaling pathway. Cell. Physiol. Biochem. 23, 165–174. doi: 10.1159/000204105
Perez, R. A., Choi, S.-J., Han, C.-M., Kim, J.-J., Shim, H., Leong, K. W., et al. (2016). Biomaterials control of pluripotent stem cell fate for regenerative therapy. Prog. Mater. Sci. 82, 234–293. doi: 10.1016/j.pmatsci.2016.05.003
Perez, R. A., El-Fiqi, A., Park, J. H., Kim, T. H., Kim, J. H., and Kim, H. W. (2014). Therapeutic bioactive microcarriers: co-delivery of growth factors and stem cells for bone tissue engineering. Acta Biomater. 10, 520–530. doi: 10.1016/j.actbio.2013.09.042
Perez, R. A., and Kim, H.-W. (2013). Core-shell designed scaffolds of alginate/alpha-tricalcium phosphate for the loading and delivery of biological proteins. J. Biomed. Mater. Res. - Part A 101A, 1103–1112. doi: 10.1002/jbm.a.34406
Perez, R. A., Kim, H.-W., and Ginebra, M.-P. (2012). Polymeric additives to enhance the functional properties of calcium phosphate cements. J. Tissue Eng. 3:2041731412439555. doi: 10.1177/2041731412439555
Perez, R. A., Kim, J.-H., Buitrago, J. O., Wall, I. B., and Kim, H.-W. (2015a). Novel therapeutic core-shell hydrogel scaffolds with sequential delivery of cobalt and bone morphogenetic protein-2 for synergistic bone regeneration. Acta Biomater. 23, 295–308. doi: 10.1016/j.actbio.2015.06.002
Perez, R. A., and Mestres, G. (2016). Role of pore size and morphology in musculo-skeletal tissue regeneration. Mater. Sci. Eng. C 61, 922–939. doi: 10.1016/j.msec.2015.12.087
Perez, R. A., Seo, S.-J., Won, J.-E., Lee, E.-J., Jang, J.-H., Knowles, J. C., et al. (2015b). Therapeutically relevant aspects in bone repair and regeneration. Mater. Today 18, 573–589. doi: 10.1016/j.mattod.2015.06.011
Perez, R. A., Shin, S.-H., Han, C.-M., and Kim, H.-W. (2015c). Bioactive injectables based on calcium phosphates for hard tissues: a recent update. Tissue Eng. Regen. Med. 12, 143–153. doi: 10.1007/s13770-015-0096-1
Perez, R. A., Singh, R. K., Kim, T. H., and Kim, H. W. (2017). Silica-based multifunctional nanodelivery systems toward regenerative medicine. Mater. Horizons 4, 772–799. doi: 10.1039/C7MH00017K
Pérez, R. A., Won, J.-E., Knowles, J. C., and Kim, H.-W. (2013). Naturally and synthetic smart composite biomaterials for tissue regeneration. Adv. Drug Deliv. Rev. 65, 471–496. doi: 10.1016/j.addr.2012.03.009
Qi, H., Liu, Y., Wu, L., Ni, S., Sun, J., Xue, J., et al. (2020). MicroRNA-16, via FGF2 regulation of the ERK/MAPK pathway, is involved in the magnesium-promoted osteogenic differentiation of mesenchymal stem cells. Oxid. Med. Cell. Longev. 2020:3894926. doi: 10.1155/2020/3894926
Qiao, Y., Zhang, W., Tian, P., Meng, F., Zhu, H., Jiang, X., et al. (2014). Stimulation of bone growth following zinc incorporation into biomaterials. Biomaterials 35, 6882–6897. doi: 10.1016/j.biomaterials.2014.04.101
Reffitt, D. M., Ogston, N., Jugdaohsingh, R., Cheung, H. F. J., Evans, B. A. J., Thompson, R. P. H., et al. (2003). Orthosilicic acid stimulates collagen type 1 synthesis and osteoblastic differentiation in human osteoblast-like cells in vitro. Bone 32, 127–135. doi: 10.1016/S8756-3282(02)00950-X
Reger, N. C., Bhargava, A. K., Ratha, I., Kundu, B., and Balla, V. K. (2019). Structural and phase analysis of multi-ion doped hydroxyapatite for biomedical applications. Ceram. Int. 45, 252–263. doi: 10.1016/j.ceramint.2018.09.160
Ren, L., Wong, H. M., Yan, C. H., Yeung, K. W. K., and Yang, K. (2015). Osteogenic ability of Cu-bearing stainless steel. J. Biomed. Mater. Res. - Part B Appl. Biomater. 103, 1433–1444. doi: 10.1002/jbm.b.33318
Saidak, Z., and Marie, P. J. (2012). Strontium signaling: molecular mechanisms and therapeutic implications in osteoporosis. Pharmacol. Ther. 136, 216–226. doi: 10.1016/j.pharmthera.2012.07.009
Salerno, A., Levato, R., Mateos-Timoneda, M. A., Engel, E., Netti, P. A., and Planell, J. A. (2013). Modular polylactic acid microparticle-based scaffolds prepared via microfluidic emulsion/solvent displacement process: fabrication, characterization, and in vitro mesenchymal stem cells interaction study. J. Biomed. Mater. Res. - Part A 101A, 720–732. doi: 10.1002/jbm.a.34374
Santocildes-Romero, M. E., Crawford, A., Hatton, P. V., Goodchild, R. L., Reaney, I. M., and Miller, C. A. (2015). The osteogenic response of mesenchymal stromal cells to strontium-substituted bioactive glasses. J. Tissue Eng. Regen. Med. 9, 619–631. doi: 10.1002/term.2003
Schamel, M., Bernhardt, A., Quade, M., Würkner, C., Gbureck, U., Moseke, C., et al. (2017). Cu2 +, Co2 + and Cr3 + doping of a calcium phosphate cement influences materials properties and response of human mesenchymal stromal cells. Mater. Sci. Eng. C 73, 99–110. doi: 10.1016/j.msec.2016.12.052
Shi, M., Zhou, Y., Shao, J., Chen, Z., Song, B., Chang, J., et al. (2015). Stimulation of osteogenesis and angiogenesis of hBMSCs by delivering Si ions and functional drug from mesoporous silica nanospheres. Acta Biomater. 21, 178–189. doi: 10.1016/j.actbio.2015.04.019
Shie, M. Y., Ding, S. J., and Chang, H. C. (2011). The role of silicon in osteoblast-like cell proliferation and apoptosis. Acta Biomater. 7, 2604–2614. doi: 10.1016/j.actbio.2011.02.023
Shiu, H. T., Goss, B., Lutton, C., Crawford, R., and Xiao, Y. (2014). Formation of blood clot on biomaterial implants influences bone healing. Tissue Eng. Part B Rev. 20, 697–712. doi: 10.1089/ten.teb.2013.0709
Smith, L. R., Cho, S., and Discher, D. E. (2018). Stem cell differentiation is regulated by extracellular matrix mechanics. Physiology 33, 16–25. doi: 10.1152/physiol.00026.2017
Song, X., Tang, W., Gregurec, D., Yate, L., Moya, S. E., and Wang, G. (2018). Layered titanates with fibrous nanotopographic features as reservoir for bioactive ions to enhance osteogenesis. Appl. Surf. Sci. 436, 653–661. doi: 10.1016/j.apsusc.2017.12.027
Tian, Y., Lu, T., He, F., Xu, Y., Shi, H., Shi, X., et al. (2018). β-tricalcium phosphate composite ceramics with high compressive strength, enhanced osteogenesis and inhibited osteoclastic activities. Coll. Surf. B Biointerfaces 167, 318–327. doi: 10.1016/j.colsurfb.2018.04.028
Timin, A. S., Litvak, M. M., Gorin, D. A., Atochina-Vasserman, E. N., Atochin, D. N., and Sukhorukov, G. B. (2018). Cell-based drug delivery and use of nano-and microcarriers for cell functionalization. Adv. Healthc. Mater. 7:1700818. doi: 10.1002/adhm.201700818
Tsuji, K., Bandyopadhyay, A., Harfe, B. D., Cox, K., Kakar, S., Gerstenfeld, L., et al. (2006). BMP2 activity, although dispensable for bone formation, is required for the initiation of fracture healing. Nat. Genet. 38, 1424–1429. doi: 10.1038/ng1916
Tunçay, E., Demirtaş, T. T., and Gümüşderelioglu, M. (2017). Microwave-induced production of boron-doped HAp (B-HAp) and B-HAp coated composite scaffolds. J. Trace Elem. Med. Biol. 40, 72–81. doi: 10.1016/j.jtemb.2016.12.005
Vieira, S., Vial, S., Reis, R. L., and Oliveira, J. M. (2017). Nanoparticles for bone tissue engineering. Biotechnol. Prog. 33, 590–611. doi: 10.1002/btpr.2469
Vroman, L., Adams, A. L., Fischer, G. C., and Munoz, P. C. (1980). Interaction of high molecular weight kininogen, factor XII, and fibrinogen in plasma at interfaces. Blood 55, 156–159. doi: 10.1182/blood.V55.1.156.156
Wang, J., Ma, X. Y., Feng, Y. F., Ma, Z. S., Ma, T. C., Zhang, Y., et al. (2017). Magnesium ions promote the biological behaviour of rat calvarial osteoblasts by activating the PI3K/Akt signalling pathway. Biol. Trace Elem. Res. 179, 284–293. doi: 10.1007/s12011-017-0948-8
Wang, L., Li, G., Ren, L., Kong, X., Wang, Y., Han, X., et al. (2017). Nano-copper-bearing stainless steel promotes fracture healing by accelerating the callus evolution process. Int. J. Nanomedicine 12, 8443–8457. doi: 10.2147/IJN.S146866
Wang, Y., Geng, Z., Huang, Y., Jia, Z., Cui, Z., Li, Z., et al. (2018). Unraveling the osteogenesis of magnesium by the activity of osteoblasts: in vitro. J. Mater. Chem. B 6, 6615–6621. doi: 10.1039/C8TB01746H
Wang, Z., Liu, Q., Liu, C., Tan, W., Tang, M., Zhou, X., et al. (2020). Mg2+ in β-TCP/Mg–Zn composite enhances the differentiation of human bone marrow stromal cells into osteoblasts through MAPK-regulated Runx2/Osx. J. Cell. Physiol. 235, 5182–5191. doi: 10.1002/jcp.29395
Wang, Z., Wang, Z., Lu, W. W., Zhen, W., Yang, D., and Peng, S. (2017). Novel biomaterial strategies for controlled growth factor delivery for biomedical applications. NPG Asia Mater. 9:e435. doi: 10.1038/am.2017.171
Wu, C., Miron, R., Sculean, A., Kaskel, S., Doert, T., Schulze, R., et al. (2011). Proliferation, differentiation and gene expression of osteoblasts in boron-containing associated with dexamethasone deliver from mesoporous bioactive glass scaffolds. Biomaterials 32, 7068–7078. doi: 10.1016/j.biomaterials.2011.06.009
Wu, C., Zhou, Y., Xu, M., Han, P., Chen, L., Chang, J., et al. (2013). Copper-containing mesoporous bioactive glass scaffolds with multifunctional properties of angiogenesis capacity, osteostimulation and antibacterial activity. Biomaterials 34, 422–433. doi: 10.1016/j.biomaterials.2012.09.066
Xu, L.-C., and Siedlecki, C. A. (2007). Effects of surface wettability and contact time on protein adhesion to biomaterial surfaces. Biomaterials 28, 3273–3283. doi: 10.1016/j.biomaterials.2007.03.032
Xu, Z., Wu, H., Wang, F., Kaewmanee, R., Pan, Y., Wang, D., et al. (2019). A hierarchical nanostructural coating of amorphous silicon nitride on polyetheretherketone with antibacterial activity and promoting responses of rBMSCs for orthopedic applications. J. Mater. Chem. B 7, 6035–6047. doi: 10.1039/C9TB01565E
Xue, J., Wu, T., Dai, Y., and Xia, Y. (2019). Electrospinning and electrospun nanofibers: methods, materials, and applications. Chem. Rev. 119, 5298–5415. doi: 10.1021/acs.chemrev.8b00593
Yamaguchi, M., Oishi, H., and Suketa, Y. (1988). Zinc stimulation of bone protein synthesis in tissue culture. Activation of aminoacyl-tRNA synthetase. Biochem. Pharmacol. 37, 4075–4080. doi: 10.1016/0006-2952(88)90098-6
Yan, Y., Wei, Y., Yang, R., Xia, L., Zhao, C., Gao, B., et al. (2019). Enhanced osteogenic differentiation of bone mesenchymal stem cells on magnesium-incorporated titania nanotube arrays. Coll. Surf. B Biointerfaces 179, 309–316. doi: 10.1016/j.colsurfb.2019.04.013
Yang, F., Yang, D., Tu, J., Zheng, Q., Cai, L., and Wang, L. (2011). Strontium enhances osteogenic differentiation of mesenchymal stem cells and in vivo bone formation by activating Wnt/catenin signaling. Stem Cells 29, 981–991. doi: 10.1002/stem.646
Yin, C., Jia, X., Miron, R. J., Long, Q., Xu, H., Wei, Y., et al. (2018). Setd7 and its contribution to Boron-induced bone regeneration in Boron-mesoporous bioactive glass scaffolds. Acta Biomater 73, 522–530. doi: 10.1016/j.actbio.2018.04.033
Yin, C., Jia, X., Zhao, Q., Zhao, Z., Wang, J., Zhang, Y., et al. (2020). Transcription factor 7-like 2 promotes osteogenic differentiation and boron-induced bone repair via lipocalin 2. Mater. Sci. Eng. C 110:110671. doi: 10.1016/j.msec.2020.110671
Ying, D., Ouyang, Z., Liu, T., Liu, X., and Huang, Q. (2018). Boron-containing micro/nano-structured TiO 2 /bioceramics coatings with modulatory effects on SaOS-2 cell response. Mater. Lett. 228, 29–32. doi: 10.1016/j.matlet.2018.05.122
Yoshizawa, S., Brown, A., Barchowsky, A., and Sfeir, C. (2014). Magnesium ion stimulation of bone marrow stromal cells enhances osteogenic activity, simulating the effect of magnesium alloy degradation. Acta Biomater 10, 2834–2842. doi: 10.1016/j.actbio.2014.02.002
Younger, E. M., and Chapman, M. W. (1989). Morbidity at bone graft donor sites. J. Orthop. Trauma 3, 192–195. doi: 10.1097/00005131-198909000-00002
Yu, H. S., Lee, E. J., Seo, S. J., Knowles, J. C., and Kim, H. W. (2015). Feasibility of silica-hybridized collagen hydrogels as three-dimensional cell matrices for hard tissue engineering. J. Biomater. Appl. 30, 338–350. doi: 10.1177/0885328215590108
Yu, W., Sun, T. W., Qi, C., Ding, Z., Zhao, H., Zhao, S., et al. (2017). Evaluation of zinc-doped mesoporous hydroxyapatite microspheres for the construction of a novel biomimetic scaffold optimized for bone augmentation. Int. J. Nanomedicine 12, 2293–2306. doi: 10.2147/IJN.S126505
Yu, Y., Liu, K., Wen, Z., Liu, W., Zhang, L., and Su, J. (2020). Double-edged effects and mechanisms of Zn2+ microenvironments on osteogenic activity of BMSCs: osteogenic differentiation or apoptosis. RSC Adv. 10, 14915–14927. doi: 10.1039/D0RA01465F
Zeng, J., Guo, J., Sun, Z., Deng, F., Ning, C., and Xie, Y. (2020). Osteoblastic and anti-osteoclastic activities of strontium-substituted silicocarnotite ceramics: in vitro and in vivo studies. Bioact. Mater. 5, 435–446. doi: 10.1016/j.bioactmat.2020.03.008
Zhai, W., Lu, H., Wu, C., Chen, L., Lin, X., Naoki, K., et al. (2013). Stimulatory effects of the ionic products from Ca-Mg-Si bioceramics on both osteogenesis and angiogenesis in vitro. Acta Biomater. 9, 8004–8014. doi: 10.1016/j.actbio.2013.04.024
Zhang, J., Cai, L., Tang, L., Zhang, X., Yang, L., Zheng, K., et al. (2018). Highly dispersed lithium doped mesoporous silica nanospheres regulating adhesion, proliferation, morphology, ALP activity and osteogenesis related gene expressions of BMSCs. Coll. Surf. B Biointerfaces 170, 563–571. doi: 10.1016/j.colsurfb.2018.06.038
Zhang, J., Ma, X., Lin, D., Shi, H., Yuan, Y., Tang, W., et al. (2015). Magnesium modification of a calcium phosphate cement alters bone marrow stromal cell behavior via an integrin-mediated mechanism. Biomaterials 53, 251–264. doi: 10.1016/j.biomaterials.2015.02.097
Zhang, K., Lin, S., Feng, Q., Dong, C., Yang, Y., Li, G., et al. (2017). Nanocomposite hydrogels stabilized by self-assembled multivalent bisphosphonate-magnesium nanoparticles mediate sustained release of magnesium ion and promote in-situ bone regeneration. Acta Biomater. 64, 389–400. doi: 10.1016/j.actbio.2017.09.039
Zhang, X., Zu, H., Zhao, D., Yang, K., Tian, S., Yu, X., et al. (2017). Ion channel functional protein kinase TRPM7 regulates Mg ions to promote the osteoinduction of human osteoblast via PI3K pathway: in vitro simulation of the bone-repairing effect of Mg-based alloy implant. Acta Biomater. 63, 369–382. doi: 10.1016/j.actbio.2017.08.051
Zhang, Y., Cui, X., Zhao, S., Wang, H., Rahaman, M. N., Liu, Z., et al. (2015). Evaluation of injectable strontium-containing borate bioactive glass cement with enhanced osteogenic capacity in a critical-sized rabbit femoral condyle defect model. ACS Appl. Mater. Interfaces 7, 2393–2403. doi: 10.1021/am507008z
Zhao, C., Wu, H., Ni, J., Zhang, S., and Zhang, X. (2017). Development of PLA/Mg composite for orthopedic implant: tunable degradation and enhanced mineralization. Compos. Sci. Technol. 147, 8–15. doi: 10.1016/j.compscitech.2017.04.037
Zheng, K., Dai, X., Lu, M., Hüser, N., Taccardi, N., and Boccaccini, A. R. (2017). Synthesis of copper-containing bioactive glass nanoparticles using a modified Stöber method for biomedical applications. Coll. Surf. B Biointerfaces 150, 159–167. doi: 10.1016/j.colsurfb.2016.11.016
Zhi Lin Sun Wataha, J. C., and Hanks, C. T. (1997). Effects of metal ions on osteoblast-like cell metabolism and differentiation. J. Biomed. Mater. Res. 34, 29–37. doi: 10.1002/(SICI)1097-4636(199701)34:1<29::AID-JBM5>3.0.CO;2-P
Zhou, H., Jiao, G., Dong, M., Chi, H., Wang, H., Wu, W., et al. (2019). Orthosilicic acid accelerates bone formation in human osteoblast-like cells through the PI3K–Akt–mTOR pathway. Biol. Trace Elem. Res. 190, 327–335. doi: 10.1007/s12011-018-1574-9
Keywords: bone, scaffolds, tissue engineering, biomaterials, tissue regeneration, therapeutic ions
Citation: Bosch-Rué E, Diez-Tercero L, Giordano-Kelhoffer B, Delgado LM, Bosch BM, Hoyos-Nogués M, Mateos-Timoneda MA, Tran PA, Gil FJ and Perez RA (2021) Biological Roles and Delivery Strategies for Ions to Promote Osteogenic Induction. Front. Cell Dev. Biol. 8:614545. doi: 10.3389/fcell.2020.614545
Received: 06 October 2020; Accepted: 17 December 2020;
Published: 14 January 2021.
Edited by:
Naresh Kumar Rajendran, University of Rochester, United StatesReviewed by:
Ahmed El-Fiqi, Dankook University, South KoreaRajprasad Loganathan, Johns Hopkins University, United States
Copyright © 2021 Bosch-Rué, Diez-Tercero, Giordano-Kelhoffer, Delgado, Bosch, Hoyos-Nogués, Mateos-Timoneda, Tran, Gil and Perez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roman A. Perez, cnBlcmV6YW5AdWljLmVz
†These authors share first authorship