 Rémy Char1*
Rémy Char1* Philippe Pierre1,2,3*
Philippe Pierre1,2,3*- 1Aix Marseille Université, Centre National de la Recherche Scientifique, Institut National de la Santé et de la Recherche Médicale, Centre d’Immunologie de Marseille-Luminy, Marseille, France
- 2Institute for Research in Biomedicine and Ilidio Pinho Foundation, Department of Medical Sciences, University of Aveiro, Aveiro, Portugal
- 3Shanghai Institute of Immunology, School of Medicine, Shanghai Jiao Tong University, Shanghai, China
Intracellular trafficking is essential for cell structure and function. In order to perform key tasks such as phagocytosis, secretion or migration, cells must coordinate their intracellular trafficking, and cytoskeleton dynamics. This relies on certain classes of proteins endowed with specialized and conserved domains that bridge membranes with effector proteins. Of particular interest are proteins capable of interacting with membrane subdomains enriched in specific phosphatidylinositol lipids, tightly regulated by various kinases and phosphatases. Here, we focus on the poorly studied RUFY family of adaptor proteins, characterized by a RUN domain, which interacts with small GTP-binding proteins, and a FYVE domain, involved in the recognition of phosphatidylinositol 3-phosphate. We report recent findings on this protein family that regulates endosomal trafficking, cell migration and upon dysfunction, can lead to severe pathology at the organismal level.
Introduction
The organization of cells into multiple membranous compartments with specific biochemical functions requires complex intracellular traffic and sorting of lipids and proteins, to transport them from their sites of synthesis to their functional destination. Intracellular transport involves lipid vesicles or tubules with the capacity to fuse with one another or to be secreted. They collectively participate in the dynamic exchanges necessary for cell homeostasis (Rothman, 2002; Søreng et al., 2018). Membrane traffic is tightly coordinated with protein synthesis, signal transduction of environmental stimuli and cytoskeleton organization, allowing the implementation of key cellular functions such as endocytosis, exocytosis, or migration (McMahon and Gallop, 2005; Habtezion et al., 2016; Vega-Cabrera and Pardo-López, 2017; MacGillavry and Hoogenraad, 2018; Margaria et al., 2019; Tapia et al., 2019; Buratta et al., 2020; Stalder and Gershlick, 2020).
Several families of molecular components required for orchestrating membrane vesicle exchange and transport during this process are conserved. They include adaptor and coat proteins, small GTP-binding proteins (GTPases), as well as Synaptosome Associated Protein (SNAP) Receptor (SNARE) proteins and SNARE binding proteins (Juliano, 2018). The vast superfamily of GTPases is involved in the establishment or regulation of virtually every step of intracellular membrane trafficking. They behave as molecular switches that can alternate between active and inactive states, through GTP binding and hydrolysis into GDP (Takai et al., 2001; Stenmark, 2009). The largest group of GTPases involved in intracellular membrane traffic is the Rab proteins family (Lamb et al., 2016). Rab GTPases specifically localize to different intracellular compartments, regulating vesicle formation and sorting, as well as transport along the cytoskeletal network. Each Rab protein can be recruited to specific membrane subdomains of a defined organelle and is associated to multiple effectors controlling membrane fusion and trafficking. Rab interaction with the membrane fusion complexes and cytoskeleton regulators is therefore crucial for cellular functions, including endocytosis and autophagy (Chen and Wandinger-Ness, 2001; Bruce et al., 2010; Geng et al., 2010; Thomas and Fromme, 2020; Yuan and Song, 2020).
Here, we review the literature concerning a less-well known family of proteins involved in the complex biochemical crosstalk established between the cytoskeleton and intracellular vesicles. This small group of proteins was named RUFY for “RUN and FYVE domain-containing.” RUFYs share a common structural domain organization, including an N-terminal RUN domain, one or several coiled-coil (CC) repeats and a C-terminal FYVE domain (Figure 1A). The molecular structures of the different RUFY proteins has been described (Dunkelberg and Gutierrez-Hartmann, 2001; Mari et al., 2001; Kukimoto-Niino et al., 2006; Kitagishi and Matsuda, 2013), but their function in endocytic regulation and their physiological relevance at the organismal level are still poorly characterized (Kitagishi and Matsuda, 2013; Terawaki et al., 2016). We revisit here how the rufy gene family was annotated, and propose the addition of a novel member, the fyco1 (FYVE and Coiled-coil containing domain 1) gene given its sequence and functional similarities with the other rufy genes (Pankiv et al., 2010; Terawaki et al., 2015). We also highlight recent findings on the implication of RUFY proteins in the regulation of cytoskeleton and endosome dynamics and their contribution to immunity, cancer and neurodegenerative diseases.
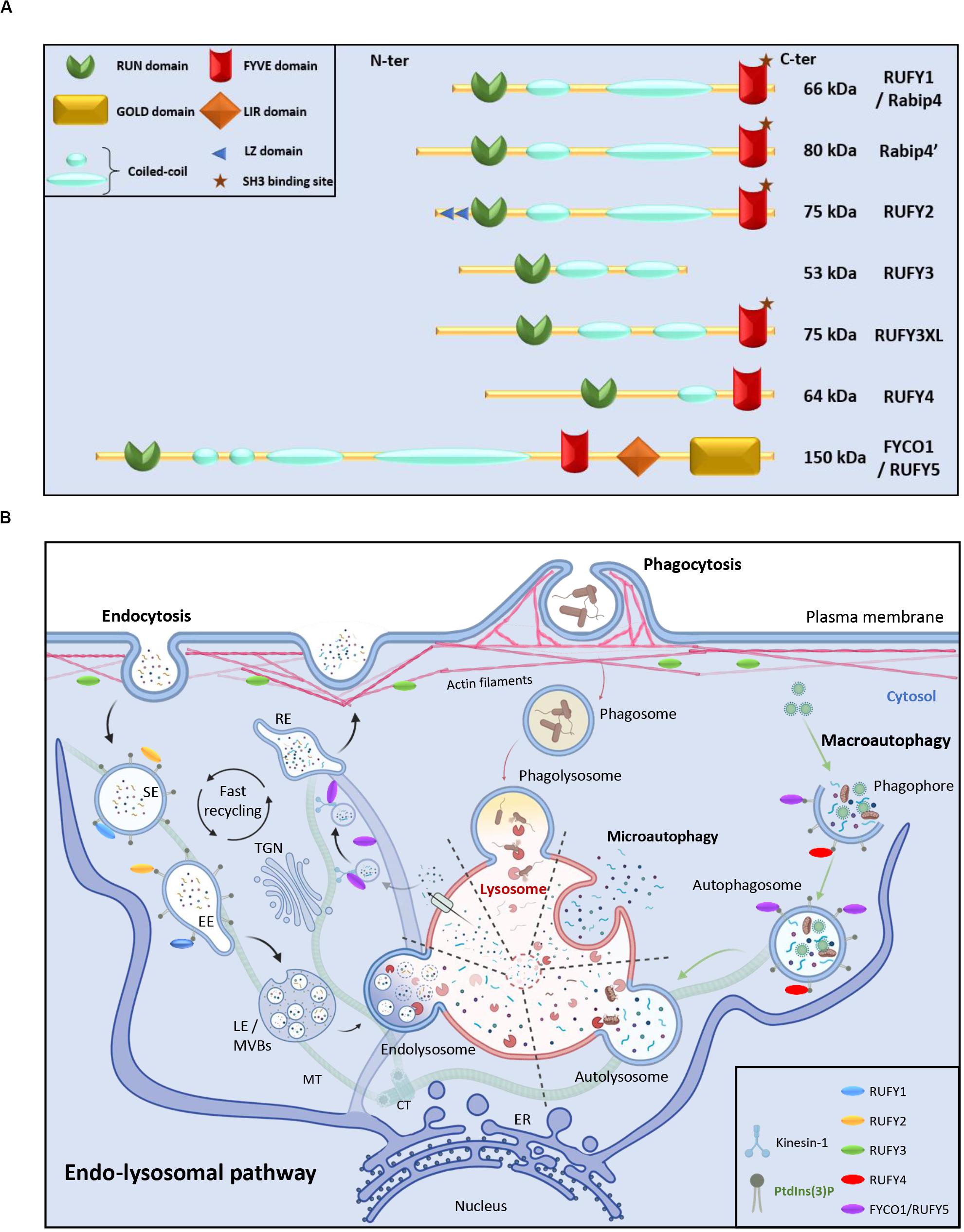
Figure 1. RUN and FYVE domain containing-proteins in the endo-lysosomal pathway. (A) Schematic representation of the RUFY proteins family. (B) Description of the endo-lysosomal and autophagy pathways and presumed functional locations of RUFY proteins. Extracellular material is ingested by endocytosis or phagocytosis. The action of different endosomes allows cargo to be sorted, recycled or degraded in a complex and regulated process involving fusion, maturation and transport along the cytoskeleton. Alternatively, during autophagy, obsolete components present in cytosol are captured in autophagosomes prior fusion with lysosomes and degradation (macroautophagy) or directly internalized through endosomal invagination (microautophagy). SE, sorting endosome; EE, early endosome; TGN, trans golgi network; LE, late endosome; MVBs, multi vesicular bodies; RE, recycling endosome, MT, microtubule; CT, centrioles; ER, endoplasmic reticulum. The location of PI3P and RUFY proteins known activity is shown. Created with BIoRender.com.
Endocytosis and Autophagy
Endocytosis and autophagy are membrane traffic pathways required for degradation and recycling of extracellular and intracellular components, respectively (Birgisdottir and Johansen, 2020). These pathways have a common endpoint at the lysosome, where their cargo is degraded. These both pathways intersect at several stages throughout vesicle formation, transport and fusion and share some of the components of their molecular machineries (Figure 1B).
There are numerous co-existing endocytic pathways, which initiate by the formation of nascent endocytic vesicles formed from plasma membrane invaginations and scissions. These endocytic vesicles undergo homotypic fusion and are rapidly targeted to sorting endosomes (SE). Sorting events initiated in SE determine the fate of internalized cargo molecules, such as recycling to plasma membrane, degradation in lysosomes, or other trafficking events (Naslavsky and Caplan, 2018; Figure 1B). On their way to degradation, sorted cargo accumulate in early endosomes (EE), that further mature into late endosomes (LE) through multiple events of cargo and lipid sorting. Late endosomes adopt a membrane organization termed multivesicular bodies, that are enriched in lysobisphosphatidic acid and contain intraluminal vesicles (Gruenberg, 2020). Next, LE potentiate their hydrolytic competence by fusing with lysosomes (Pillay et al., 2002) resulting in the degradation of their contents, providing nutrients and key factors to the cell (Doherty and McMahon, 2009; Kaksonen and Roux, 2018). Notably, endosomes play a role in signal transduction by serving as signaling platforms either for surface activated receptors like Toll-like receptors and epidermal growth factor receptor or metabolic sensors such as mechanistic target of rapamycin complex 1 (mTORC1; Argüello et al., 2016). Often they promote the degradation of their targets, leading to signal termination (Chung et al., 2010). The endocytic pathway has also specialized functions in differentiated cells such as neurotransmitter release and recycling in neurons, or antigen processing and presentation in professional antigen presenting cells, like B cells or dendritic cells (Argüello et al., 2016; Solé-Domènech et al., 2016; Hinze and Boucrot, 2018). Endocytosis events and endosomes positioning is highly dependent on the dynamic and spatial re-organization of the different cytoskeleton networks that include actin, intermediate filaments, or microtubules (Fletcher and Mullins, 2010; Pegoraro et al., 2017).
Complementary to endocytosis, autophagy is an intracellular process by which cells degrade and recycle their own cytoplasmic materials (Mizushima and Komatsu, 2011). Autophagy plays a central role in many physiological processes including stress management, development, immunity and aging (Puleston and Simon, 2014; Zhong et al., 2016; Fîlfan et al., 2017; Moretti et al., 2017; Doherty and Baehrecke, 2018). Autophagy is partially controlled though mTORC1 activity and is responsible for degradation and recycling of misfolded proteins, as well as obsolete organelles (Galluzzi et al., 2017). The endpoint of autophagy is to deliver cytoplasmic material to lysosomes, where like for endocytosed cargo, it is degraded. Several autophagy processes can be distinguished based on the entry mode of the cytosolic components destined for degradation (Figure 1B). Macroautophagy involves engulfment of cytoplasmic contents into a double membrane vesicle termed the autophagosome. The autophagosome fuses then with lysosomes, becoming an autolysosome, in which its cargo is degraded (Galluzzi et al., 2017). The presence of specific phosphoinositides lipids, together with Rab GTPases, at a given membrane compartment is often directly correlated with compartment function. One of the common mechanism regulating endocytosis and autophagy is an accumulation of phosphatidylinositol 3-phosphate (PtdIns(3)P) at surface of EE and on intraluminal vesicles of multivesicular endosomes and on autophagosomes (Nascimbeni et al., 2017; Figure 1B). PtdIns(3)P is also observed at sites of LC3−associated phagocytosis another pathway of internalization used by the cells to ingest large particulate material or microbes. PtdIns(3)P is therefore a beacon used by the cellular machinery to regulate endosomal sorting and autophagy (Birgisdottir and Johansen, 2020).
RUN Domains
The presence of a single copy of a RUN and a FYVE domain at their extremities is the key characteristic defining the RUFY family members. RUN domains were named after three proteins bearing similar peptide motifs, RPIP8, UNC-14 and NESCA (new molecule containing SH3 at the carboxy−terminus) (Ogura et al., 1997; Matsuda et al., 2000). RUN domains are present in multiple proteins (RUN proteins) in a large panel of organisms (Figure 2) and principally allow direct interactions with small GTPases of the Rap and Rab families (Callebaut et al., 2001; Yoshida et al., 2011). RUN domains adopt a hydrophobic globular structure bearing six conserved blocks named A to F (Figure 3A). These blocks correspond to eight α-helices and some 310-helices. The first helix is crucial to limit hydrophobic exposure and maintain protein solubility of RUN-containing proteins (Callebaut et al., 2001; Kukimoto-Niino et al., 2006). In spite of strong conservation among the domains present in RUN-containing proteins, the proteins they interact with, their effectors, are highly variable (Mari et al., 2001) and the structural features of the RUN domain alone are not sufficient to define binding specificity for one or several members of the GTPase superfamily (Fukuda et al., 2011). Most RUN domain-bearing proteins bind small GTPases, but interactions with other molecules like kinesin 1 have also been described (Boucrot et al., 2005). A direct physical link between RUN proteins with actin filaments and microtubules has been also demonstrated (Torti et al., 1999), reinforcing the idea that these molecules are also critical for cellular functions requiring actin remodeling, such as migration or phagocytosis (Price and Bos, 2004; Bos, 2005; Miertzschke et al., 2007; Xu et al., 2007; Figure 4A). Additional functions for RUN domains have been described, for example for the RUN domain present in NESCA, which blocks TRAF6-mediated polyubiquitination of the NF-kappa-B essential modulator and consequently induces NF-kB activation. This is just one of the ways in which RUN proteins can act in signal transduction and the coordination of membrane traffic with actin dynamics upon external stimulation (Yoshida et al., 2011). As well as promoting endosomal fusion through their binding to Rab or Rap GTPases (Callebaut et al., 2001; Yoshida et al., 2011), their interaction with motor proteins, like kinesin or myosin, suggests a role for RUN domains in regulating vesicular and organelle transport (Callebaut et al., 2001; Yoshida et al., 2011). Via these different mechanisms, RUN proteins have been implicated in neuronal development (Honda et al., 2017b), signaling (Sun et al., 2012), migration (Yoshida et al., 2011), and regulation of various cellular function like endocytosis or exocytosis (Kitagishi and Matsuda, 2013).
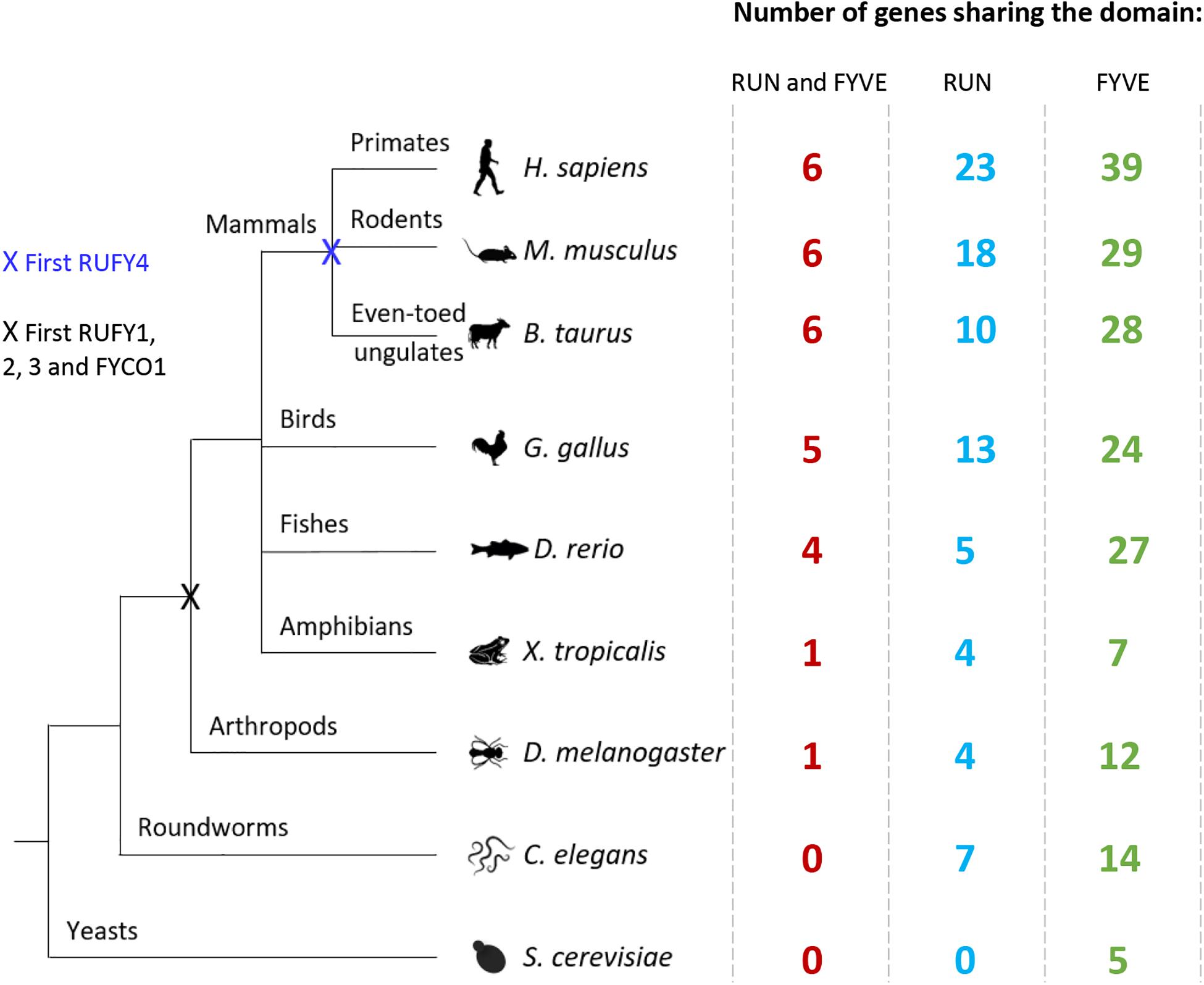
Figure 2. Evolution of RUN and FYVE domain or rufy genes among living organisms. Diagram illustrating the evolution of the rufy genes. Species representative of various taxonomic groups are listed, data were extracted from the Differential Expression Atlas Genes database (EMBL-EBI). Next to each species studied, the number corresponds to the number of genes having in its sequence a FYVE (green), RUN (blue) or both (red) domain. The “X” corresponds to the appearance of a common rufy ancestor gene.
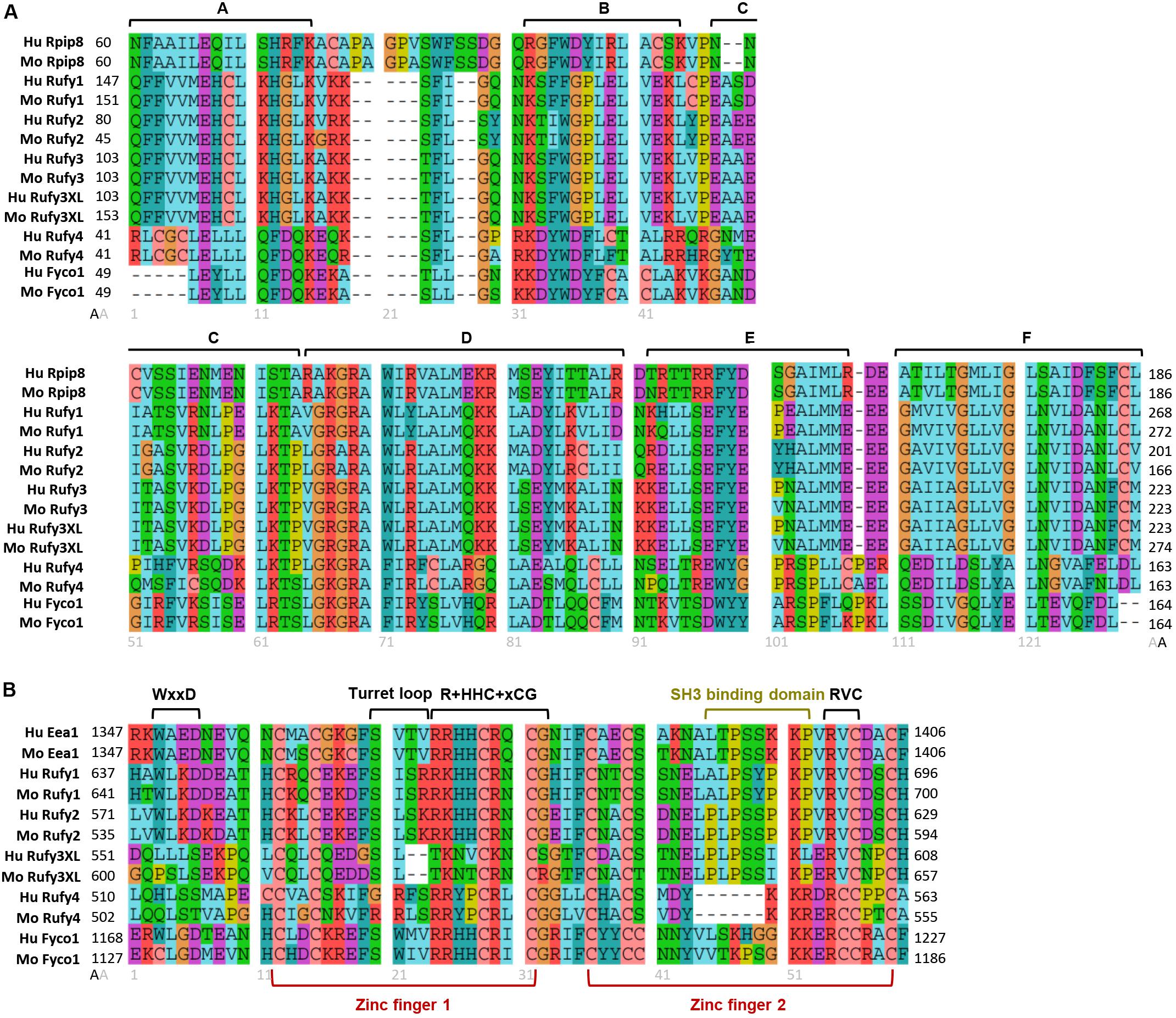
Figure 3. Molecular organization of RUN and FYVE domains from the RUFY proteins family. Alignment of the protein sequences of the RUN (A) and FYVE (B) domains of the RUFY proteins family in human and mouse. (A) RUN consensus blocks are represented by segments (A–F). Rpip8 sequence is used as RUN domain reference, (B) FYVE conserved motives and zinc fingers are represented by segments. In the alignment, “x” is any amino acid and “+” represents positively charged amino acid. Eea1 sequence is used as FYVE domain reference. For all alignment, amino acids are colored according to their properties: Cyan for hydrophobic positions (A,V,I,L,M), turquoise for aromatic positions (F,Y,W,H), red for basic residues (K,R), purple for acidic residues (D,E), green for polar uncharged (N,Q,S,T), salmon for cysteine (C), orange for glycine (G) and yellow for proline (P). Gray numbers below alignment means the amino acids position after alignment. Black numbers surrounding the alignments represent the start (left) and end (right) positions of the domains in the peptide sequence of each protein. Alignment were realized with Seaviewer analyzer software (Gouy et al., 2010). Accession numbers for protein are following: human Rpip8 (NP_001138297.1), mouse Rpip8 (NP_058039.1), human Eea1 (NP_003557.3), mouse Eea1 (NP_001001932.1), human RUFY1 (NP_079434.3), mouse RUFY1 (NP_766145.1), human RUFY2 (NP_060457.4), mouse RUFY2 (NP_081701.2), human RUFY3 (NP_055776.1), mouse RUFY3 (NP_081806.1) human RUFY3XL (NP_001032519.1), mouse RUFY3XL (NP_001276703.1), human RUFY4 (NP_940885.2), mouse RUFY4 (NP_001164112.1), human FYCO1 (NP_078789.2), mouse FYCO1 (NP_001103723.2).
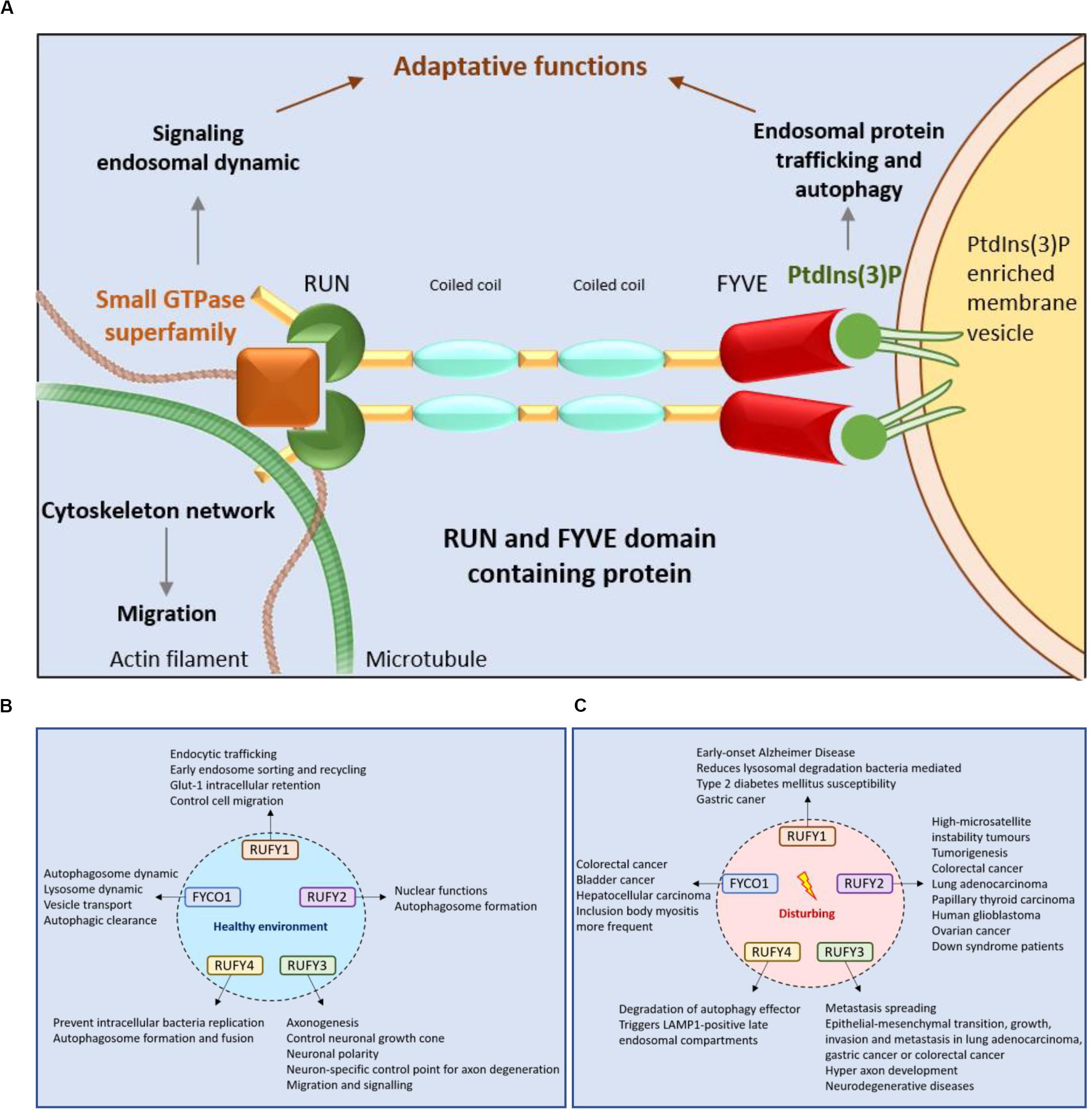
Figure 4. RUFY proteins are important for intracellular trafficking, signaling and cytoskeleton dynamics. (A) Schematic representation of the RUN and FYVE domains activity of RUFY proteins. RUN domains act on signaling, endosomal protein trafficking and cytoskeletal network dynamics via small GTPase proteins. FYVE domains bind PtdIns(3)P and regulates autophagy and endosome trafficking. (B) Function of RUFY proteins in homeostatic conditions. (C) Consequences of alterations in RUFY proteins functions at the cellular and organismal level.
FYVE Domains
FYVE-domain-bearing proteins (for Fab1, YOTB/ZK632.12, Vac1, and EEA1) are specifically found in association with membranous organelles enriched in PtdIns(3)P and highly conserved among eukaryotes, including yeast (Hayakawa et al., 2007; Figure 2). FYVE domains adopt a zinc finger conformation (Misra and Hurley, 1999; Kutateladze and Overduin, 2001). In addition to FYVE, ten types of zinc finger folds have been characterized, including conventional, Gal4, GATA-1, TFIIS, MetRS, LIM, RING domain, PKC CRD, and PHD domains. Zinc fingers are structural conformations adopted by peptide chains upon coordination of two Zn2+ cations within a cysteine rich region (Schwabe and Klug, 1994; Stenmark et al., 1996). Unlike most molecules bearing zinc fingers, FYVE proteins display only one copy of the domain located at any position along the peptide chain, highlighting its autonomy as a structural unit. FYVE zinc fingers can stabilize protein-protein or protein-DNA/RNA interactions (Dunkelberg and Gutierrez-Hartmann, 2001). A “classical” FYVE domain has eight potential zinc coordinating tandem cysteine positions and is characterized by having basic amino acids around the cysteines. Many members of this family also include two histidine residues in a sequence motif including WxxD, CxxC, R+HHC+xCG and RVC where “x” means any amino acid and “+” a positively charged amino acid (Figure 3B). Most deviations from this sequence can reduce the domain affinity for zinc and destabilize it (Stenmark et al., 1996; Misra and Hurley, 1999; Stenmark and Aasland, 1999; Kutateladze and Overduin, 2001). Within this structural framework, specific modifications in the non-conserved residues of the domain can radically affect FYVE protein subcellular localization and function, by forming a “turret loop” and a dimerization interface (Hayakawa et al., 2004).
With regard to their affinity for PtdIns(3)P, FYVE domain-containing proteins are mostly found associated to EE or phagosomes (Stenmark et al., 1996; Gaullier et al., 1998; Stenmark and Aasland, 1999; Figure 4A). The presence of FYVE domains is therefore correlated to the regulation of membrane traffic, through specific recognition of PtdIns(3)P domains by “R+HHC+xCG” motifs (Gaullier et al., 1998), and modulation by associated phosphatidylinositol kinases. PtdIns(3)P is generated from phosphatidylinositol by Class III PtdIns 3-kinases (PI3K), like Vps34, on target membranes such as nascent autophagosome (omegasomes) (Melia et al., 2020), or EE (Di Paolo and De Camilli, 2006; Raiborg et al., 2013; Scott et al., 2014; Figure 1B). In turn, accumulation of PtdIns(3)P recruits and activates effector proteins containing FYVE domains, favoring transport or fusion of target organelles (Stenmark and Gillooly, 2001; Axe et al., 2008; Burman and Ktistakis, 2010; Schink et al., 2013). Affinity for PtdIns(3)P is determined by the pair of histidine residues present in the “R+HHC+XCG” motif of the FYVE domain (Stahelin et al., 2002; Diraviyam et al., 2003; Lee et al., 2005; He et al., 2009). This affinity can also be harnessed by FYVE proteins to link endosomes with mRNA, ribonucleoprotein particles (mRNP) and associated ribosomes, playing a role in their long-distance transport in the cell (Pohlmann et al., 2015). Importantly, many FYVE proteins homodimerize. Dimerization multiplies the conserved residues displayed by the different signature motifs present in the FYVE domain and contributes to a network of hydrogen bonding and electrostatic interactions that provides positive selection for binding several PtdIns(3)P head groups. PH-dependent insertion of FYVE domain into cell membranes (He et al., 2009; Pankiv et al., 2010) is reinforced by additional hydrophobic membrane interactions with the turret loop and/or tandem lysine residues. These non-specific interactions promote FYVE domain access to phosphate head groups, that are hindered by the close packing of lipid molecules. This bivalent mechanism increases therefore greatly FYVE domains specificity for PtdIns(3)P-enriched domains and discrimination against other mono- or polyphosphorylated PtdIns species (Misra and Hurley, 1999; Stenmark and Aasland, 1999; Dumas et al., 2001; Kutateladze and Overduin, 2001).
FYVE proteins are therefore key players in endocytosis and autophagy and mutations in FYVE domains can alter profoundly these functions, as well as cellular homeostasis (Kamentseva et al., 2020). For example, EEA1 protein (early endosome antigen 1) is known to be crucial for endosome dynamics and any mutation in its conserved residues or the oligomerization site can drastically reduce the affinity between its FYVE domain and PtdIns(3)P (Stenmark et al., 1996; Gaullier et al., 2000). In this context, RUFYs proteins, by bearing a N-terminal RUN domain, one or several copies of a coiled-coil domain next to a C-terminal FYVE domain (Figure 1A) have all the features required to carry-out specific adaptor functions to regulate endocytosis or autophagy by impacting on organelle fusion and mobility along the cytoskeleton.
The RUFY Proteins Family
The RUFY family encompass four genes named rufy1 to 4, sharing homologies and displaying specific tissue expression and alternative splicing. Rufy genes are relatively conserved genes, absent from prokaryotes and fungi. Upon evolution, the emergence of the common ancestor appeared in vertebrates and arthropods, which possess one ortholog (CG31064) (Figure 2). No RUFY protein could be detected In Caenorhabditis elegans and only a FYVE-bearing protein (T10G3.5) considered as an ortholog of human EEA1 shows some sequence similarities with the RUFY family. T10G3.5 exhibits PtdIns(3)P binding activity and is involved in endocytosis, being mostly expressed in epidermis and intestine of C. elegans (Hayakawa et al., 2007). In chordates, Rubicon (RUN domain and cysteine-rich domain containing, Beclin 1-interacting protein) and FYVE And Coiled-Coil Domain Autophagy Adaptor 1 (FYCO1), display structural and functional features, potentially categorizing them as RUFY proteins. Rubicon was identified as a component of the Class III PI3K complex and a negative regulator of autophagy and endosomal trafficking (Matsunaga et al., 2009; Zhong et al., 2009). Like RUFYs, Rubicon contains multiple functional domains that interact with other proteins, including a RUN, a CC and a FYVE-like domains (Wong et al., 2018). However, despite these similarities, the poor degree of sequence homology and the lack of conservation of its FYVE-like domain, which was found not to bind to PI(3)P (Burman and Ktistakis, 2010), prevented Rubicon’s integration within the RUFY proteins family, conversely to FYCO1, which we propose here to name RUFY5 and detail the characteristics below.
RUFY1
RUFY1, previously named Rabip4 is an 80 kDa protein, mainly expressed in the brain, kidney, lung, placenta and testis. There are two RUFY1 isoforms Rabip4, and Rabip4’ that has an additional 108 amino acid upstream of the N-terminal RUN domain (Figure 1A). They were both shown to interact with the small endosomal GTPases Rab4, Rab5, and Rab14 (Fouraux et al., 2004; Vukmirica et al., 2006; Table 1). RUFY1 inactivation inhibits efficient recycling of endocytosed transferrin, implicating RUFY1 in the regulation of EE functions through cooperative interactions with Rab4 and Rab14 (Cormont et al., 2001; Yamamoto et al., 2010; Nag et al., 2018). This was further demonstrated by the alteration of epidermal growth factor receptor endocytic trafficking kinetics in cells depleted of RUFY1 (Gosney et al., 2018) and the hijacking of RUFY1 by the bacteria P. gingivalis to escape lysosomal degradation (Takeuchi et al., 2016). In melanocytes, RUFY1 was found to form a complex with rabenosyn-5, KIF3A-B, Rab4A and adaptor protein-3 (AP-3) to differentially regulate tyrosinase-related protein-1 and tyrosinase sorting in endosomes, contributing to melanosome maturation (Nag et al., 2018; Table 1). Moreover, silencing the Rabip4’ isoform of RUFY1 was shown to promote outgrowth of plasma membrane protrusions, and to regulate the spatial distribution of lysosomes at their tips, through an interaction with AP-3 (Ivan et al., 2012; Figures 1B, 4B). RUFY1 is also capable of controlling cell migration by regulating integrin trafficking (Vukmirica et al., 2006), presumably via endocytosis. In full agreement with a role of RUFY1 in regulating endosomal dynamics, a single nucleotide polymorphism (S705A) in the rufy1 gene was associated with high blood glucose levels and type 2 diabetes mellitus susceptibility in an exome-wide association study (EWAS; Yamada et al., 2017). This result is consistent with the early finding that Rabip4 expression leads to Glucose transporter-1 (Glut-1) intracellular retention (Cormont et al., 2001). Interestingly RUFY1 display a SH3-binding motif “PxxPxP” embedded in the FYVE domain and is able to interacting with the epithelial and endothelial tyrosine kinase (ETK), and possibly regulates endocytosis through this interaction (Yang et al., 2002). Another EWAS, aiming to find early-onset Alzheimer’s Disease (AD) susceptibility genes, identified RUFY1 among genes involved in endo-lysosomal transport and known to be important for the development of AD (Kunkle et al., 2017; Figure 4C).
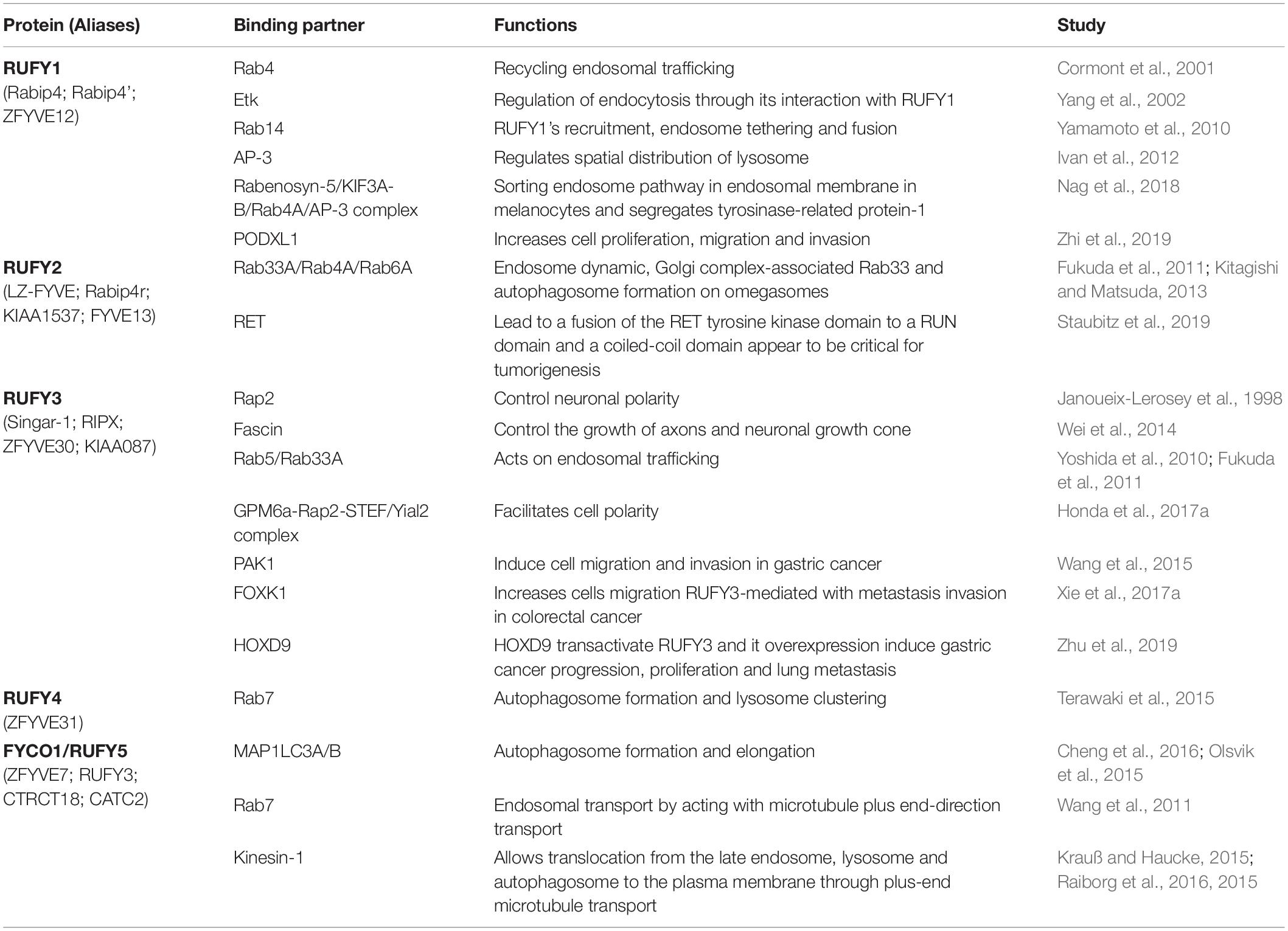
Table 1. Summary of RUFY proteins functional interactions.
RUFY2
RUFY2 (or Leucine zipper FYVE-finger protein, LZ-FYVE) is a 75 kDa protein originally identified as an activating transcription factor-2 interactor embryogenesis (Dunkelberg and Gutierrez-Hartmann, 2001), preferentially located in the nucleus and expressed during. After development, RUFY2 expression remains high in the brain, lung, liver and the gastrointestinal tract (Yang et al., 2002). RUFY2 displays two N-terminal leucine zipper domains as well as a C-terminal FYVE-finger domain. Although it is likely to have a nuclear function at early stages of embryonic development, the presence of a FYVE domain suggests a cytoplasmic role for RUFY2 in regulating membrane traffic in fully differentiated cells. Importantly, the RUN domain of RUFY2 was shown to associate specifically with the Golgi complex-associated Rab33A (Fukuda et al., 2011; Table 1). Given the reported interaction of Rab33A and Rab33B with Atg16L and its putative role in regulating autophagy (Fukuda and Itoh, 2008), RUFY2 could contribute to autophagosome formation through a dual interaction with Rab33A and PtdIns(3)P on omegasomes (Figures 1B, 4B). Irrespective of its function, rufy2 expression is subject to modulation by the micro RNA miR-155 (Bofill-De Ros et al., 2015), which is an important regulator of immune cells development and inflammatory responses (Ceppi et al., 2009). The rufy2 gene is also frequently found mutated in cancer cells, with the most frequent mutations converting it into a strong target for nonsense mediated mRNA decay, thereby decreasing considerably its expression (Shin et al., 2011; Figure 4C).
RUFY3
RUFY3, also known as Rap2-interacting protein X (RIPX) (Kukimoto-Niino et al., 2006) or Single Axon-Related 1 (Singar1) (Mori et al., 2007), is the best characterized member of the RUFY family. RUFY3, the smallest of the RUFY proteins with a molecular weight of 53 kDa (Figure 1A), is mostly expressed in neurons (Kitagishi and Matsuda, 2013). Neuronal RUFY3 is atypical, since it lacks a FYVE domain and is considered as part of the RUFY family based on strong sequence similarities with the other members, notably in the RUN and coiled-coil domains (Figure 2A). RUFY3 is distributed between the cytosol and at the plasma membrane, but not in intracellular vesicles, presumably because it lacks a FYVE domain. In artificial conditions, like following expression of the dominant gain of function mutant form of Rab5 (Q79L) in U937 cells, RUFY3 was found associated in large vesicle structures and to co-immunoprecipitate with Rab5, via an interaction with its carboxyl terminal domain and surprisingly not its RUN domain (Yoshida et al., 2010). Like RUFY2, RUFY3 was also shown in a 2-hybrid screen and by co-immunoprecipitation to bind Rab33, through its coiled-coil domain 1 (CC1; Fukuda et al., 2011). In 293T and 3Y1 cell lines however, RUFY3 was shown not to interact with several small GTPases, including Rab2, Rab5, Rab7, Rho, and Ras. This suggests that either RUFY3 requires cell specific partner proteins or post-translation modifications to be able to bind to small GTPases. RUFY3 was first described as interacting with Rap2, a small Ras-like GTPase, via a 173 residue fragment (83–255) located in the RUN domain (Janoueix-Lerosey et al., 1998; Kukimoto-Niino et al., 2006; Table 1). Together with Rap1, Rap2 interacts with Ras effectors, such as Raf, PI3K, and Ral guanine nucleotide dissociation stimulator, inhibiting activation of their downstream targets, and thus suppressing Ras oncogenic activity (Kukimoto-Niino et al., 2006; Nussinov et al., 2020). In the adult nervous system, Rap1 and Rap2 also regulate the maturation and plasticity of dendritic spine and synapses. By forming a complex together with Rap2 and Fascin, RUFY3 interacts with the filamentous actin network and controls the growth of axons and neuronal growth cone (Wei et al., 2014; Table 1). Recent mechanistic studies indicate that RUFY3 accumulates in lipid rafts by forming a Glycoprotein M6A (GPM6a)-RUFY3-Rap2-STEF/Yial2 complex (Honda et al., 2017a; Table 1). This complex activates the Rac guanine nucleotide exchange factor (Honda et al., 2017b), impacting actin organization and promoting neuronal polarity and growth (Figure 4B). RUFY3 seems therefore to have different axogenic functions in brain (Mori et al., 2007; Honda et al., 2017b) and not surprisingly, roles for RUFY3 in amyotrophic lateral sclerosis (Arosio et al., 2016), major depressive disorder (Aberg et al., 2018) and AD (Zelaya et al., 2015) have been reported. Olfactory dysfunction occurs in 90% of AD cases and is correlated with elevated rufy3 expression in glomerular and mitral layers of the olfactory bulb (Zelaya et al., 2015). RUFY3 is cleaved by caspase 3 and critically required for caspase-mediated degeneration of tropomyosin receptor kinase A positive sensory axons in vitro and in vivo (Hertz et al., 2019; Figure 4C). Removal of neuronally enriched RUFY3 is able to block caspase 3-dependent apoptosis, while dephosphorylation of RUFY3 at residue S34 appears required for its degradation (Hertz et al., 2019). Analysis of rufy3-deficient mice supports a second distinct function for RUFY3 in neuronal growth and polarity, since mutant embryos show defects in axonal projection patterns. These occur in addition to the prevention of CASP3-dependent apoptosis in dorsal root ganglions. RUFY3 appears therefore to be key for nervous system development, remodeling and function, explaining the embryonic lethality displayed upon rufy3 genetic inactivation in mouse (Hertz et al., 2019).
With the current advance in genomics and single cell RNA sequencing, specific gene expression patterns can be revised and more accurately defined. Analysis of several genomic databases (BioGPS, NCBI, Human Atlas Protein, ImmGen, Ensembl) reveal that, in addition to neurons, RUFY3 expression can be detected in other tissues and cell types. Moreover, the rufy3 gene appears to have many transcriptional variants, leading to the expression of different protein isoforms. Two of these isoforms display a C-terminal region extended by 150 amino acids, compared to the previously identified neuronal isoform of RUFY3. Importantly, these previously uncharacterized longer isoforms (RUFY3XL) possess the same RUN domain and a putative FYVE domain in their C-terminus (Figure 1A), indicating that RUFY3 is a legitimate member of the RUFY family. In contrast to classic FYVE zinc fingers, genomic databases reveal this putative FYVE domain appears to lack the tandem histidine residue cluster that defines affinity for PtdIns(3)P (Figure 3B). Interestingly, the SH3 binding site embedded in the RUFY1 and RUFY2 FYVE domains is also present in RUFY3XL, suggesting a potential signal transduction activity for this uncharacterized isoform. The translation of rufy3xl mRNA into a functional protein and its capacity to bind PtdIns(3)P remain to be demonstrated. If true, a role for RUFY3 in the coordination of endosome dynamics or organelle transport could be hypothesized. This idea is supported by the observation that RUFY3 is present in Staufen2-containing messenger ribonucleoprotein particles, that are used to transport mRNAs along neuronal dendrites to their site of translation (Maher-Laporte et al., 2010). FYVE proteins have already been implicated in endosome-mediated transport of mRNP (Pohlmann et al., 2015) and RUFY3XL could therefore also perform this function. The existence of FYVE domain bearing isoforms, might extend and diversify its function in other specialized cells.
RUFY4
RUFY4 is a 64 kDa that is atypical among the RUFY family members, since it bears several non-conserved residues in its RUN domain and it lacks the tandem histidine cluster and the SH3 binding domain normally present in the FYVE domain (Figures 1A, 3A,B). RUFY4 was shown to interact with PtdIns(3)P enriched membranes (Terawaki et al., 2015, 2016). Interestingly, EMBL-EBI and SMART genomic databases show that rufy4 is present only in mammals, suggesting that rufy4 is the most recently evolved gene in the RUFY family (Figure 2). RUFY4 levels are extremely low in most cells and tissues with the exception of lungs and lymphoid organs. RUFY4 was found to be strongly induced in vitro in dendritic cells differentiated from bone marrow progenitors in presence of GM-CSF and IL-4. In vivo, its expression was confirmed in alveolar macrophages and in lung dendritic cells isolated from asthmatic mice (Terawaki et al., 2015). RUFY4 interacts with Rab7 through its RUN domain and promotes the generation of large autophagosomes (Terawaki et al., 2015; Figure 4B and Table 1). RUFY4 over-expression induces the degradation of the autophagy effector LC3/ATG8 and triggers clustering of LAMP1-positive late endosomal compartments. These compartments are distinct from large abnormal autophagosome-like structures positive for RUFY4 and Syntaxin-17, a Qa SNARE involved in autophagosome formation and fusion (Figure 4C). RUFY4 was also proposed to interact with PLEKHM1 and the HOPS complex, which are implicated in LE and lysosome dynamics and positioning (Terawaki et al., 2016). RUFY4 seems therefore able to harness the classical autophagy machinery to facilitate autophagosome formation and increase autophagy flux by acting at different biochemical steps (Terawaki et al., 2015). By optimizing effector protein activity and organelle distribution, RUFY4 expression facilitates the elimination of both damaged mitochondria and intracellular bacteria in phagocytes. RUFY4 expression in HeLa cells can prevent replication of Brucella abortus (Terawaki et al., 2015) and Salmonella typhimurium (Lassen et al., 2016) suggesting that RUFY4 has a key role in anti-bacterial responses in the lung. It also potentially acts to drive immunity though the regulation of endocytosis and autophagy, necessary for the presentation at the cell surface of antigens from intracellular pathogens (Terawaki et al., 2015).
FYCO1
FYCO1 is a 150 kDa protein bearing a RUN and a FYVE domains. In several databases, fyco1 was misidentified as rufy3, although these two genes are present on completely distinct chromosomes, in human chromosome 3 and 4, respectively. At the sequence level, although it is larger, FYCO1 appears to be a RUFY4 ortholog gene (Figures 3A,B), suggesting that FYCO1 belongs to the RUFY family. We therefore propose that it could be annotated as RUFY5 to fit the family nomenclature. Separating its N-terminal RUN domain from the FYVE zinc finger, FYCO1 has several CC domains, as well as a LC3/ATG8 Interacting Region (LIR) and a Golgi Dynamic (GOLD) domain in its C-terminus (Figure 1A). FYCO1 preferentially interacts with MAP1LC3A/B of the Atg8-familly proteins through its LIR (Olsvik et al., 2015; Cheng et al., 2016). Coiled-coil domains promote FYCO1 dimerization and have been shown to mediate the formation of a complex with Rab7, via a part of the CC located upstream of the FYVE domain (Pankiv et al., 2010; Wang et al., 2011; Table 1). Overexpression of FYCO1 was shown to redistribute LC3- and Rab7-positive structures to the cell periphery in a microtubule-dependent manner (Pankiv et al., 2010). This effect is mediated by the central part of the CC region and suggests a role for FYCO1 in the transport of autophagic vesicles (Figure 4B). The capacity of FYCO1 to interact with Rab7 and LC3A/B on the external surface of autophagosomes, and PtdIns3P enriched membranes through its FYVE domain, is likely to be key to its function as an adaptor protein. Indeed, these interactions allow microtubule plus end-directed transport and protrusion of endocytic organelles, including autophagosomes (Pankiv and Johansen, 2010), LE (Raiborg et al., 2015, 2016), lysosomes (Mrakovic et al., 2012; Hong et al., 2017; Lie and Nixon, 2019), and phagosomes (Ma et al., 2014). Endoplasmic reticulum (ER) and endosomes are connected through contact sites, the numbers of which increase as endosomes mature. The functions of such sites include to control the association of endosomes with the minus-end-directed microtubule motor dynein and to mediate endosome fission. Repeated LE–ER contacts promote microtubule-dependent translocation of LEs to the cell periphery and subsequent fusion with the plasma membrane (Raiborg et al., 2016). Such fusion induces outgrowth of protrusions and neurites in the neuroendocrine cell line PC12, which require the ER-associated protein protrudin on the ER and FYCO1 to interact with LEs and kinesin 1 (Krauß and Haucke, 2015; Table 1). FYCO1 has been described as a novel mediators of invalopodia formation and function of Protrudin-mediated ER–endosome contact sites (Pedersen et al., 2020). Multiple studies highlight the critical function of FYCO1 in autophagy and autophagosome/endosome trafficking (Dionne et al., 2017) with pathological consequences arising when FYCO1 function is altered (Figures 4B,C). Mutations in the fyco1 gene affect autophagy and cause autosomal-recessive congenital cataracts by altering lens development and transparency in patients (Chen et al., 2011, 2017; Brennan et al., 2012; Costello et al., 2013; Chauss et al., 2014; Frost et al., 2014; Khan et al., 2015; Gunda et al., 2018; Li et al., 2018). Sequencing studies of candidate genes potentially involved in several neuromuscular or neurodegenerative diseases have identified rare variants of autophagy related proteins like VCP and SQSTM1. Among these genes, a missense fyco1 variant was identified to cause sporadic inclusion body myositis (Güttsches et al., 2017; Rothwell et al., 2017; Britson et al., 2018; Figure 4C). Finally FYCO1 has been implicated in the autophagic clearance of specialized particles or aggregates, like male germ cell-specific RNP ribonucleoprotein granules (Da Ros et al., 2017), post-mitotic bodies (Dionne et al., 2017) or α-synuclein aggregates (Saridaki et al., 2018).
RUFY Proteins and Cancer
As describe above, RUFY proteins play a central role in cellular functions by regulating vesicular trafficking and its interactions with the cytoskeleton. Neuronal deficit and neurodegeneration are the most obvious manifestations of RUFY proteins alteration. Not surprisingly, however, given their relatively broad adaptors functions, RUFY proteins have taken center stage in the oncology field.
The ETK tyrosine kinase has been shown to play a pivotal role in a variety of cellular processes including proliferation, differentiation, motility, and apoptosis (Yang et al., 2002; Kung, 2011; Zhuang et al., 2014; Wang et al., 2018). Tyrosine phosphorylation of RUFY1 by ETK appears to be important for its endosomal localization and could play an important role promoting tumoral transformation by affecting downstream effectors of PI3-kinase. RUFY1 was also shown to interact with podocalyxin-like protein (PODXL), a transmembrane glycoprotein with anti-adhesive properties associated with poor prognosis of several cancers (Taniuchi et al., 2018; He et al., 2020; Table 1). Gastric cancer progression is significantly increased upon PODXL expression, a phenotype reduced by concomitant RUFY1 silencing. Depletion of RUFY1 inactivates the PI3K/AKT, NF-κB and MAPK/ERK signaling pathways and reduces drastically migration and invasion of cancer cells in vitro (Zhi et al., 2019). Given the positive correlation between podxl and rufy1 expression in tissues and serum, rufy1 was proposed as a potential biomarker for gastric cancers stratification (Zhi et al., 2019; Figure 3C). Like RUFY1, a role for RUFY2 in various cancer has been reported (Shin et al., 2011; Zheng et al., 2014; Staubitz et al., 2019). Rufy2 is one of the most frequently mutated genes in high-microsatellite instability tumors and colorectal cancer (Shin et al., 2011). Gene rearrangement of the proto-oncogene ret with rufy2 have been shown to drive tumorigenesis in lung adenocarcinoma (Zheng et al., 2014) and papillary thyroid carcinoma (Staubitz et al., 2019). The gene rearrangement leads to a fusion of the RET tyrosine kinase domain with RUFY2 RUN domain and coiled-coil domain; this appears to be critical for tumorigenesis (Staubitz et al., 2019; Table 1). Rufy2 mRNA is the target of several microRNAs, including miR-146a, miR-196a-5p and miR-155 (Bofill-De Ros et al., 2015). Dysregulated microRNA targeting of RUFY2 expression was found important for the development of human glioblastoma and ovarian cancer, suggesting a tumor suppression role for RUFY2 (Lukács et al., 2019; Zheng et al., 2020; Figure 4C). Given the key role of RUFY3 in cell migration, membrane transport, and cellular signaling, through its interaction with rap2, it is not surprising that RUFY3 dysregulation has been implicated in several cancer processes and metastatic tumor spread. The abnormal expression of RUFY3 is linked to poor prognosis. It can promote growth, invasion and metastasis in lung adenocarcinoma, gastric cancer cells or colorectal cancer (Xie et al., 2017a, b; Men et al., 2019; Zhu et al., 2019). RUFY3 overexpression and its interaction with P21-activated kinase-1 (PAK1) leads to the formation of F-actin-enriched protrusive structures, increased epithelial-mesenchymal transition and gastric cancer cell migration (Kumar and Vadlamudi, 2002; Vadlamudi and Kumar, 2003). Several transcription factors, including Forkhead box k1 (FOXK1) and Homebox D9 (HOXD9) involved in cancer progression (Moens and Selleri, 2006; Tabuse et al., 2011; Lv et al., 2015; Peng et al., 2016; Wu et al., 2016; Liu et al., 2018; Zhu et al., 2019), have been shown to regulate RUFY3 expression and activity (Xie et al., 2017a; Zhu et al., 2019; Table 1). So far, no correlation has been found between RUFY4 and any type of cancer. FYCO1 has also been implicated in colorectal cancer progression (Sillars-Hardebol et al., 2010) and recent studies have concluded that FYCO1 may serve as a biomarker in bladder cancer (Eissa et al., 2017) or hepatocellular carcinoma (Vongchan and Linhardt, 2017; Figure 4C). Plus, FYCO1 can indirectly associated with cell invasion (Pedersen et al., 2020).
Conclusion
Although they have been poorly characterized to date, RUFY proteins play a central role in cellular homeostasis by regulating endocytosis, autophagy and coordinating organelle transport with signal transduction cascades. It is important to note that RUFY proteins also provide a regulatory link between cytoskeletal dynamics and membrane trafficking. Consequently, these proteins have adaptive functions by acting on localized actions (through PtdIns(3)P) and signaling (through small GTPases), which can affect key biological functions in specialized cells, such as migration, tissue repair or targeted secretion. The dysregulated expression of RUFY proteins has therefore severe consequences on cell differentiation and polarization, causing cancers or neurodegenerative diseases. However, further molecular and physiological analyses will be required to understand how these proteins exert their functions in specialized cell types like immune cells or neurons. Immunocytes require endocytosis and migration to perform their functions within primary and secondary lymphoid organs or at sites of infection. The restricted expression of RUFY4, as well as the existence of splicing variants of RUFY3 in alveolar macrophages and dendritic cells, suggest a role for these molecules in phagocytes. Of importance will be the characterization of the different molecules interacting either with their RUN or FYVE domains in a cell specific manner. Identification of these RUFY’s interactors will be crucial to establish the functionality of the domains and their importance for signaling on one end and subcellular targeting at the other end. The coiled-coil structural domains found in the central part of the RUFY proteins should also be scrutinized. CC domains, in addition to support homodimerization and increase affinity for PtdIns(3)P, could also be determinant in promoting RUFY proteins interactions with effector molecules, like Rab7, as observed for FYCO1. The nature and pattern of expression of these effector molecules will allow to sort the different activities displayed by the RUFYs in individual cell types and thereby shed light on their physiological importance in health and diseases.
Author Contributions
Both authors contributed equally to the design and implementation of the research and writing of the manuscript.
Funding
The PP laboratory is “Equipe de la Fondation de la Recherche Médicale” (FRM) sponsored by the grant DEQ20180339212. The project was supported by grants from l’Agence Nationale de la Recherche (ANR) «DCBIOL Labex ANR-11-LABEX-0043», «INFORM Labex ANR-11-LABEX-0054» funded by the “Investissements d’Avenir” French government program. The research was also supported by the Ilídio Pinho foundation and FCT – Fundaçăo para a Ciência e a Tecnologia – and Programa Operacional Competitividade e Internacionalizaçăo – Compete2020 (FEDER) – references POCI-01-0145-FEDER-016768 and POCI-01-0145-FEDER-030882.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge Dr. Jonathan Ewbank for his constructive comments.
References
Aberg, K. A., Dean, B., Shabalin, A. A., Chan, R. F., Han, L. K. M., Zhao, M., et al. (2018). Methylome-wide association findings for major depressive disorder overlap in blood and brain and replicate in independent brain samples. Mol. Psychiatry 25, 1344–1354. doi: 10.1038/s41380-018-0247-6
Argüello, R. J., Reverendo, M., Gatti, E., and Pierre, P. (2016). Regulation of protein synthesis and autophagy in activated dendritic cells: implications for antigen processing and presentation. Immunol. Rev. 272, 28–38. doi: 10.1111/imr.12427
Arosio, A., Sala, G., Rodriguez-Menendez, V., Grana, D., Gerardi, F., Lunetta, C., et al. (2016). MEF2D and MEF2C pathways disruption in sporadic and familial ALS patients. Mol. Cell. Neurosci. 74, 10–17. doi: 10.1016/j.mcn.2016.02.002
Axe, E. L., Walker, S. A., Manifava, M., Chandra, P., Roderick, H. L., Habermann, A., et al. (2008). Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum. J. Cell Biol. 182, 685–701. doi: 10.1083/jcb.200803137
Birgisdottir, ÅB., and Johansen, T. (2020). Autophagy and endocytosis – interconnections and interdependencies. J. Cell Sci. 133:jcs228114. doi: 10.1242/jcs.228114
Bofill-De Ros, X., Santos, M., Vila-Casadesús, M., Villanueva, E., Andreu, N., Dierssen, M., et al. (2015). Genome-wide miR-155 and miR-802 target gene identification in the hippocampus of Ts65Dn Down syndrome mouse model by miRNA sponges. BMC Genomics 16:907. doi: 10.1186/s12864-015-2160-6
Bos, J. L. (2005). Linking rap to cell adhesion. Curr. Opin. Cell Biol. 17, 123–128. doi: 10.1016/j.ceb.2005.02.009
Boucrot, E., Henry, T., Borg, J.-P., Gorvel, J.-P., and Méresse, S. (2005). The intracellular fate of Salmonella depends on the recruitment of kinesin. Science 308, 1174–1178. doi: 10.1126/science.1110225
Brennan, L. A., Kantorow, W. L., Chauss, D., McGreal, R., He, S., Mattucci, L., et al. (2012). Spatial expression patterns of autophagy genes in the eye lens and induction of autophagy in lens cells. Mol. Vis. 18, 1773–1786.
Britson, K. A., Yang, S. Y., and Lloyd, T. E. (2018). New developments in the genetics of inclusion body myositis. Curr. Rheumatol. Rep. 20:26. doi: 10.1007/s11926-018-0738-0
Bruce, E. A., Digard, P., and Stuart, A. D. (2010). The Rab11 pathway is required for influenza A virus budding and filament formation. J. Virol. 84, 5848–5859. doi: 10.1128/JVI.00307-10
Buratta, S., Tancini, B., Sagini, K., Delo, F., Chiaradia, E., Urbanelli, L., et al. (2020). Lysosomal exocytosis, exosome release and secretory autophagy: the autophagic- and endo-lysosomal systems go extracellular. Int. J. Mol. Sci. 21:2576. doi: 10.3390/ijms21072576
Burman, C., and Ktistakis, N. T. (2010). Regulation of autophagy by phosphatidylinositol 3-phosphate. FEBS Lett. 584, 1302–1312. doi: 10.1016/j.febslet.2010.01.011
Callebaut, I., de Gunzburg, J., Goud, B., and Mornon, J.-P. (2001). RUN domains: a new family of domains involved in Ras-like GTPase signaling. Trends Biochem. Sci. 26, 79–83. doi: 10.1016/S0968-0004(00)01730-8
Ceppi, M., Schmidt, E., and Pierre, P. (2009). Genetic modification of murine dendritic cells by RNA transfection. Methods Mol. Biol. 531, 145–156. doi: 10.1007/978-1-59745-396-7_10
Chauss, D., Basu, S., Rajakaruna, S., Ma, Z., Gau, V., Anastas, S., et al. (2014). Differentiation state-specific mitochondrial dynamic regulatory networks are revealed by global transcriptional analysis of the developing chicken lens. G3 4, 1515–1527. doi: 10.1534/g3.114.012120
Chen, J., Ma, Z., Jiao, X., Fariss, R., Kantorow, W. L., Kantorow, M., et al. (2011). Mutations in FYCO1 cause autosomal-recessive congenital cataracts. Am. J. Hum. Genet. 88, 827–838. doi: 10.1016/j.ajhg.2011.05.008
Chen, J., Wang, Q., Cabrera, P. E., Zhong, Z., Sun, W., Jiao, X., et al. (2017). Molecular genetic analysis of Pakistani families with autosomal recessive congenital cataracts by homozygosity screening. Invest. Ophthalmol. Vis. Sci. 58, 2207–2217. doi: 10.1167/iovs.17-21469
Chen, W., and Wandinger-Ness, A. (2001). Expression and functional analyses of Rab8 and Rab11a in exocytic transport from trans-Golgi network. Methods Enzymol. 329, 165–175. doi: 10.1016/s0076-6879(01)29077-6
Cheng, X., Wang, Y., Gong, Y., Li, F., Guo, Y., Hu, S., et al. (2016). Structural basis of FYCO1 and MAP1LC3A interaction reveals a novel binding mode for Atg8-family proteins. Autophagy 12, 1330–1339. doi: 10.1080/15548627.2016.1185590
Chung, I., Akita, R., Vandlen, R., Toomre, D., Schlessinger, J., and Mellman, I. (2010). Spatial control of EGF receptor activation by reversible dimerization on living cells. Nature 464, 783–787. doi: 10.1038/nature08827
Cormont, M., Mari, M., Galmiche, A., Hofman, P., and Le Marchand-Brustel, Y. (2001). A FYVE-finger-containing protein, Rabip4, is a Rab4 effector involved in early endosomal traffic. Proc. Natl. Acad. Sci. U.S.A. 98, 1637–1642. doi: 10.1073/pnas.031586998
Costello, M. J., Brennan, L. A., Basu, S., Chauss, D., Mohamed, A., Gilliland, K. O., et al. (2013). Autophagy and mitophagy participate in ocular lens organelle degradation. Exp. Eye Res. 116:141–150. doi: 10.1016/j.exer.2013.08.017
Da Ros, M., Lehtiniemi, T., Olotu, O., Fischer, D., Zhang, F.-P., Vihinen, H., et al. (2017). FYCO1 and autophagy control the integrity of the haploid male germ cell-specific RNP granules. Autophagy 13, 302–321. doi: 10.1080/15548627.2016.1261319
Di Paolo, G., and De Camilli, P. (2006). Phosphoinositides in cell regulation and membrane dynamics. Nature 443, 651–657. doi: 10.1038/nature05185
Dionne, L. K., Peterman, E., Schiel, J., Gibieža, P., Skeberdis, V. A., Jimeno, A., et al. (2017). FYCO1 regulates accumulation of post-mitotic midbodies by mediating LC3-dependent midbody degradation. J. Cell Sci. 130, 4051–4062. doi: 10.1242/jcs.208983
Diraviyam, K., Stahelin, R. V., Cho, W., and Murray, D. (2003). Computer modeling of the membrane interaction of FYVE domains. J. Mol. Biol. 328, 721–736. doi: 10.1016/S0022-2836(03)00325-5
Doherty, G. J., and McMahon, H. T. (2009). Mechanisms of endocytosis. Annu. Rev. Biochem. 78, 857–902. doi: 10.1146/annurev.biochem.78.081307.110540
Doherty, J., and Baehrecke, E. H. (2018). Life, death and autophagy. Nat. Cell Biol. 20, 1110–1117. doi: 10.1038/s41556-018-0201-5
Dumas, J. J., Merithew, E., Sudharshan, E., Rajamani, D., Hayes, S., Lawe, D., et al. (2001). Multivalent endosome targeting by homodimeric EEA1. Mol. Cell 8, 947–958. doi: 10.1016/S1097-2765(01)00385-9
Dunkelberg, J. C., and Gutierrez-Hartmann, A. (2001). LZ-FYVE: a novel developmental stage-specific leucine zipper, FYVE-finger protein. DNA Cell Biol. 20, 403–412. doi: 10.1089/104454901750361460
Eissa, S., Matboli, M., Awad, N., and Kotb, Y. (2017). Identification and validation of a novel autophagy gene expression signature for human bladder cancer patients. Tumour Biol. 39:1010428317698360. doi: 10.1177/1010428317698360
Fîlfan, M., Sandu, R. E., Zăvăleanu, A. D., GreşiŢă, A., Glăvan, D. G., Olaru, D. G., et al. (2017). Autophagy in aging and disease. Rom. J. Morphol. Embryol. 58, 27–31.
Fletcher, D. A., and Mullins, R. D. (2010). Cell mechanics and the cytoskeleton. Nature 463, 485–492. doi: 10.1038/nature08908
Fouraux, M. A., Deneka, M., Ivan, V., van der Heijden, A., Raymackers, J., van Suylekom, D., et al. (2004). Rabip4’ is an effector of rab5 and rab4 and regulates transport through early endosomes. Mol. Biol. Cell 15, 611–624. doi: 10.1091/mbc.e03-05-0343
Frost, L. S., Mitchell, C. H., and Boesze-Battaglia, K. (2014). Autophagy in the eye: implications for ocular cell health. Exp. Eye Res. 124, 56–66. doi: 10.1016/j.exer.2014.04.010
Fukuda, M., and Itoh, T. (2008). Direct link between Atg protein and small GTPase Rab: Atg16L functions as a potential Rab33 effector in mammals. Autophagy 4, 824–826. doi: 10.4161/auto.6542
Fukuda, M., Kobayashi, H., Ishibashi, K., and Ohbayashi, N. (2011). Genome-wide investigation of the Rab binding activity of RUN domains: development of a novel tool that specifically traps GTP-Rab35. Cell Struct. Funct. 36, 155–170. doi: 10.1247/csf.11001
Galluzzi, L., Baehrecke, E. H., Ballabio, A., Boya, P., Bravo-San Pedro, J. M., Cecconi, F., et al. (2017). Molecular definitions of autophagy and related processes. EMBO J. 36, 1811–1836. doi: 10.15252/embj.201796697
Gaullier, J.-M., Rønning, E., Gillooly, D. J., and Stenmark, H. (2000). Interaction of the EEA1 FYVE finger with phosphatidylinositol 3-phosphate and early endosomes. Role of conserved residues. J. Biol. Chem. 275, 24595–24600. doi: 10.1074/jbc.M906554199
Gaullier, J.-M., Simonsen, A., D’Arrigo, A., Bremnes, B., Stenmark, H., and Aasland, R. (1998). FYVE fingers bind PtdIns(3)P. Nature 394, 432–433. doi: 10.1038/28767
Geng, J., Nair, U., Yasumura-Yorimitsu, K., and Klionsky, D. J. (2010). Post-Golgi Sec proteins are required for autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 21, 2257–2269. doi: 10.1091/mbc.e09-11-0969
Gosney, J. A., Wilkey, D. W., Merchant, M. L., and Ceresa, B. P. (2018). Proteomics reveals novel protein associations with early endosomes in an epidermal growth factor-dependent manner. J. Biol. Chem. 293, 5895–5908. doi: 10.1074/jbc.RA117.000632
Gouy, M., Guindon, S., and Gascuel, O. (2010). Seaview version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27, 221–224. doi: 10.1093/molbev/msp259
Gruenberg, J. (2020). Life in the lumen: the multivesicular endosome. Traffic 21, 76–93. doi: 10.1111/tra.12715
Gunda, P., Manne, M., Adeel, S. S., Kondareddy, R. K. R., and Tirunilai, P. (2018). Detection of c.139G>A (D47N) mutation in GJA8 gene in an extended family with inheritance of autosomal dominant zonular cataract without pulverulent opacities by exome sequencing. J. Genet. 97, 879–885. doi: 10.1007/s12041-018-0971-3
Güttsches, A.-K., Brady, S., Krause, K., Maerkens, A., Uszkoreit, J., Eisenacher, M., et al. (2017). Proteomics of rimmed vacuoles define new risk allele in inclusion body myositis. Ann. Neurol. 81, 227–239. doi: 10.1002/ana.24847
Habtezion, A., Nguyen, L. P., Hadeiba, H., and Butcher, E. C. (2016). Leukocyte trafficking to the small intestine and colon. Gastroenterology 150, 340–354. doi: 10.1053/j.gastro.2015.10.046
Hayakawa, A., Hayes, S., Leonard, D., Lambright, D., and Corvera, S. (2007). Evolutionarily conserved structural and functional roles of the FYVE domain. Biochem. Soc. Symb. 74, 95–105.
Hayakawa, A., Hayes, S. J., Lawe, D. C., Sudharshan, E., Tuft, R., Fogarty, K., et al. (2004). Structural basis for endosomal targeting by FYVE domains. J. Biol. Chem. 279, 5958–5966. doi: 10.1074/jbc.M310503200
He, J., Vora, M., Haney, R. M., Filonov, G. S., Musselman, C. A., Burd, C. G., et al. (2009). Membrane insertion of the FYVE domain is modulated by pH. Proteins 76, 852–860. doi: 10.1002/prot.22392
He, S., Du, W., Li, M., Yan, M., and Zheng, F. (2020). PODXL might be a new prognostic biomarker in various cancers: a meta-analysis and sequential verification with TCGA datasets. BMC Cancer 20:620. doi: 10.1186/s12885-020-07108-5
Hertz, N. T., Adams, E. L., Weber, R. A., Shen, R. J., O’Rourke, M. K., Simon, D. J., et al. (2019). Neuronally enriched RUFY3 is required for caspase-mediated axon degeneration. Neuron 103, 412–422.e4. doi: 10.1016/j.neuron.2019.05.030
Hinze, C., and Boucrot, E. (2018). Endocytosis in proliferating, quiescent and terminally differentiated cells. J. Cell Sci. 131:jcs216804. doi: 10.1242/jcs.216804
Honda, A., Ito, Y., Takahashi-Niki, K., Matsushita, N., Nozumi, M., Tabata, H., et al. (2017a). Extracellular signals induce glycoprotein M6a clustering of lipid rafts and associated signaling molecules. J. Neurosci. 37, 4046–4064. doi: 10.1523/JNEUROSCI.3319-16.2017
Honda, A., Usui, H., Sakimura, K., and Igarashi, M. (2017b). Rufy3 is an adapter protein for small GTPases that activates a Rac guanine nucleotide exchange factor to control neuronal polarity. J. Biol. Chem. 292, 20936–20946. doi: 10.1074/jbc.M117.809541
Hong, Z., Pedersen, N. M., Wang, L., Torgersen, M. L., Stenmark, H., and Raiborg, C. (2017). PtdIns3P controls mTORC1 signaling through lysosomal positioning. J. Cell Biol. 216, 4217–4233. doi: 10.1083/jcb.201611073
Ivan, V., Martinez-Sanchez, E., Sima, L. E., Oorschot, V., Klumperman, J., Petrescu, S. M., et al. (2012). AP-3 and Rabip4’ coordinately regulate spatial distribution of lysosomes. PLoS One 7:e48142. doi: 10.1371/journal.pone.0048142
Janoueix-Lerosey, I., Pasheva, E., de Tand, M.-F., Tavitian, A., and de Gunzburg, J. (1998). Identification of a specific effector of the small GTP-binding protein Rap2. Eur. J. Biochem. 252, 290–298. doi: 10.1046/j.1432-1327.1998.2520290.x
Juliano, R. L. (2018). Intracellular trafficking and endosomal release of oligonucleotides: what we know and what we don’t. Nucleic Acid Ther. 28, 166–177. doi: 10.1089/nat.2018.0727
Kaksonen, M., and Roux, A. (2018). Mechanisms of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 19, 313–326. doi: 10.1038/nrm.2017.132
Kamentseva, R., Kosheverova, V., Kharchenko, M., Zlobina, M., Salova, A., Belyaeva, T., et al. (2020). Functional cycle of EEA1-positive early endosome: direct evidence for pre-existing compartment of degradative pathway. PLoS One 15:e0232532. doi: 10.1371/journal.pone.0232532
Khan, A. O., Aldahmesh, M. A., and Alkuraya, F. S. (2015). Phenotypes of recessive pediatric cataract in a cohort of children with identified homozygous gene mutations (an American ophthalmological society thesis). Trans. Am. Ophthalmol. Soc. 113:T7.
Kitagishi, Y., and Matsuda, S. (2013). RUFY, Rab and Rap family proteins involved in a regulation of cell polarity and membrane trafficking. Int. J. Mol. Sci. 14, 6487–6498. doi: 10.3390/ijms14036487
Krauß, M., and Haucke, V. (2015). A grab to move on: ER-endosome contacts in membrane protrusion formation and neurite outgrowth. EMBO J. 34, 1442–1444. doi: 10.15252/embj.201591553
Kukimoto-Niino, M., Takagi, T., Akasaka, R., Murayama, K., Uchikubo-Kamo, T., Terada, T., et al. (2006). Crystal structure of the RUN domain of the RAP2-interacting protein X. J. Biol. Chem. 281, 31843–31853. doi: 10.1074/jbc.M604960200
Kumar, R., and Vadlamudi, R. K. (2002). Emerging functions of p21-activated kinases in human cancer cells. J. Cell. Physiol. 193, 133–144. doi: 10.1002/jcp.10167
Kung, H.-J. (2011). Targeting tyrosine kinases and autophagy in prostate cancer. Horm. Cancer 2, 38–46. doi: 10.1007/s12672-010-0053-3
Kunkle, B. W., Vardarajan, B. N., Naj, A. C., Whitehead, P. L., Rolati, S., Slifer, S., et al. (2017). Early-onset Alzheimer disease and candidate risk genes involved in endolysosomal transport. JAMA Neurol. 74, 1113–1122. doi: 10.1001/jamaneurol.2017.1518
Kutateladze, T., and Overduin, M. (2001). Structural mechanism of endosome docking by the FYVE domain. Science 291, 1793–1796. doi: 10.1126/science.291.5509.1793
Lamb, C. A., Longatti, A., and Tooze, S. A. (2016). Rabs and GAPs in starvation-induced autophagy. Small GTPases 7, 265–269. doi: 10.1080/21541248.2016.1220779
Lassen, K. G., McKenzie, C. I., Mari, M., Murano, T., Begun, J., Baxt, L. A., et al. (2016). Genetic coding variant in GPR65 alters lysosomal pH and links lysosomal dysfunction with colitis risk. Immunity 44, 1392–1405. doi: 10.1016/j.immuni.2016.05.007
Lee, S. A., Eyeson, R., Cheever, M. L., Geng, J., Verkhusha, V. V., Burd, C., et al. (2005). Targeting of the FYVE domain to endosomal membranes is regulated by a histidine switch. Proc. Natl. Acad. Sci. U.S.A. 102, 13052–13057. doi: 10.1073/pnas.0503900102
Li, J., Leng, Y., Han, S., Yan, L., Lu, C., Luo, Y., et al. (2018). Clinical and genetic characteristics of Chinese patients with familial or sporadic pediatric cataract. Orphanet J. Rare Dis. 13:94. doi: 10.1186/s13023-018-0828-0
Lie, P. P. Y., and Nixon, R. A. (2019). Lysosome trafficking and signaling in health and neurodegenerative diseases. Neurobiol. Dis. 122, 94–105. doi: 10.1016/j.nbd.2018.05.015
Liu, H., Zhang, M., Xu, S., Zhang, J., Zou, J., Yang, C., et al. (2018). HOXC8 promotes proliferation and migration through transcriptional up-regulation of TGFβ1 in non-small cell lung cancer. Oncogenesis 7:1. doi: 10.1038/s41389-017-0016-4
Lukács, J., Soltész, B., Penyige, A., Nagy, B., and Póka, R. (2019). Identification of miR-146a and miR-196a-2 single nucleotide polymorphisms at patients with high-grade serous ovarian cancer. J. Biotechnol. 297, 54–57. doi: 10.1016/j.jbiotec.2019.03.016
Lv, X., Li, L., Lv, L., Qu, X., Jin, S., Li, K., et al. (2015). HOXD9 promotes epithelial–mesenchymal transition and cancer metastasis by ZEB1 regulation in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 34:133. doi: 10.1186/s13046-015-0245-3
Ma, J., Becker, C., Reyes, C., and Underhill, D. M. (2014). Cutting edge: FYCO1 recruitment to Dectin-1 phagosomes is accelerated by light chain 3 protein and regulates phagosome maturation and reactive oxygen production. J. Immunol. 192, 1356–1360. doi: 10.4049/jimmunol.1302835
MacGillavry, H. D., and Hoogenraad, C. C. (2018). Membrane trafficking and cytoskeletal dynamics in neuronal function. Mol. Cell. Neurosci. 91, 1–208. doi: 10.1016/j.mcn.2018.08.003
Maher-Laporte, M., Berthiaume, F., Moreau, M., Julien, L.-A., Lapointe, G., Mourez, M., et al. (2010). Molecular composition of Staufen2-containing ribonucleoproteins in embryonic rat brain. PLoS One 5:e11350. doi: 10.1371/journal.pone.0011350
Margaria, J. P., Ratto, E., Gozzelino, L., Li, H., and Hirsch, E. (2019). Class II PI3Ks at the intersection between signal transduction and membrane trafficking. Biomolecules 9:104. doi: 10.3390/biom9030104
Mari, M., Macia, E., Le Marchand-Brustel, Y., and Cormont, M. (2001). Role of the FYVE finger and the RUN domain for the subcellular localization of Rabip4. J. Biol. Chem. 276, 42501–42508. doi: 10.1074/jbc.M104885200
Matsuda, S., Miyazaki, K., Ichigotani, Y., Kurata, H., Takenouchi, Y., Yamamoto, T., et al. (2000). Molecular cloning and characterization of a novel human gene (NESCA) which encodes a putative adapter protein containing SH3. Biochim. Biophys. Acta 1491, 321–326. doi: 10.1016/S0167-4781(00)00049-X
Matsunaga, K., Saitoh, T., Tabata, K., Omori, H., Satoh, T., Kurotori, N., et al. (2009). Two Beclin 1-binding proteins, Atg14L and Rubicon, reciprocally regulate autophagy at different stages. Nat. Cell Biol. 11, 385–396. doi: 10.1038/ncb1846
McMahon, H. T., and Gallop, J. L. (2005). Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 438, 590–596. doi: 10.1038/nature04396
Melia, T. J., Lystad, A. H., and Simonsen, A. (2020). Autophagosome biogenesis: from membrane growth to closure. J. Cell Biol. 219:e202002085. doi: 10.1083/jcb.202002085
Men, W., Li, W., Li, Y., Zhao, J., Qu, X., Li, P., et al. (2019). RUFY3 predicts poor prognosis and promotes metastasis through epithelial-mesenchymal transition in lung adenocarcinoma. J. Cancer 10, 6278–6285. doi: 10.7150/jca.35072
Miertzschke, M., Stanley, P., Bunney, T. D., Rodrigues-Lima, F., Hogg, N., and Katan, M. (2007). Characterization of interactions of adapter protein RAPL/Nore1B with RAP GTPases and their role in T cell migration. J. Biol. Chem. 282, 30629–30642. doi: 10.1074/jbc.M704361200
Misra, S., and Hurley, J. H. (1999). Crystal structure of a phosphatidylinositol 3-phosphate-specific membrane-targeting motif, the FYVE domain of Vps27p. Cell 97, 657–666. doi: 10.1016/S0092-8674(00)80776-X
Mizushima, N., and Komatsu, M. (2011). Autophagy: renovation of cells and tissues. Cell 147, 728–741. doi: 10.1016/j.cell.2011.10.026
Moens, C. B., and Selleri, L. (2006). Hox cofactors in vertebrate development. Dev. Biol. 291, 193–206. doi: 10.1016/j.ydbio.2005.10.032
Moretti, J., Roy, S., Bozec, D., Martinez, J., Chapman, J. R., Ueberheide, B., et al. (2017). STING senses microbial viability to orchestrate stress-mediated autophagy of the endoplasmic reticulum. Cell 171, 809–823.e13. doi: 10.1016/j.cell.2017.09.034
Mori, T., Wada, T., Suzuki, T., Kubota, Y., and Inagaki, N. (2007). Singar1, a novel RUN domain-containing protein, suppresses formation of surplus axons for neuronal polarity. J. Biol. Chem. 282, 19884–19893. doi: 10.1074/jbc.M700770200
Mrakovic, A., Kay, J. G., Furuya, W., Brumell, J. H., and Botelho, R. J. (2012). Rab7 and Arl8 GTPases are necessary for lysosome tubulation in macrophages. Traffic 13, 1667–1679. doi: 10.1111/tra.12003
Nag, S., Rani, S., Mahanty, S., Bissig, C., Arora, P., Azevedo, C., et al. (2018). Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. J. Cell. Sci. 131:jcs216226. doi: 10.1242/jcs.216226
Nascimbeni, A. C., Codogno, P., and Morel, E. (2017). Phosphatidylinositol-3-phosphate in the regulation of autophagy membrane dynamics. FEBS J. 284, 1267–1278. doi: 10.1111/febs.13987
Naslavsky, N., and Caplan, S. (2018). The enigmatic endosome - sorting the ins and outs of endocytic trafficking. J. Cell. Sci. 131:jcs216499. doi: 10.1242/jcs.216499
Nussinov, R., Jang, H., Zhang, M., Tsai, C.-J., and Sablina, A. A. (2020). The mystery of Rap1 suppression of oncogenic Ras. Trends Cancer 6, 369–379. doi: 10.1016/j.trecan.2020.02.002
Ogura, K., Shirakawa, M., Barnes, T. M., Hekimi, S., and Ohshima, Y. (1997). The UNC-14 protein required for axonal elongation and guidance in Caenorhabditis elegans interacts with the serine/threonine kinase UNC-51. Genes Dev. 11, 1801–1811. doi: 10.1101/gad.11.14.1801
Olsvik, H. L., Lamark, T., Takagi, K., Larsen, K. B., Evjen, G., Øvervatn, A., et al. (2015). FYCO1 contains a C-terminally extended, LC3A/B-preferring LC3-interacting region (LIR) motif required for efficient maturation of autophagosomes during basal autophagy. J. Biol. Chem. 290, 29361–29374. doi: 10.1074/jbc.M115.686915
Pankiv, S., Alemu, E. A., Brech, A., Bruun, J.-A., Lamark, T., Øvervatn, A., et al. (2010). FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end–directed vesicle transport. J. Cell Biol. 188, 253–269. doi: 10.1083/jcb.200907015
Pankiv, S., and Johansen, T. (2010). FYCO1: linking autophagosomes to microtubule plus end-directing molecular motors. Autophagy 6, 550–552. doi: 10.4161/auto.6.4.11670
Pedersen, N. M., Wenzel, E. M., Wang, L., Antoine, S., Chavrier, P., Stenmark, H., et al. (2020). Protrudin-mediated ER–endosome contact sites promote MT1-MMP exocytosis and cell invasion. J. Cell Biol. 219, 28. doi: 10.1083/jcb.202003063
Pegoraro, A. F., Janmey, P., and Weitz, D. A. (2017). Mechanical properties of the cytoskeleton and cells. Cold Spring Harb. Perspect. Biol. 9:a022038. doi: 10.1101/cshperspect.a022038
Peng, Y., Zhang, P., Huang, X., Yan, Q., Wu, M., Xie, R., et al. (2016). Direct regulation of FOXK1 by C-jun promotes proliferation, invasion and metastasis in gastric cancer cells. Cell Death Dis. 7:e2480. doi: 10.1038/cddis.2016.225
Pillay, C. S., Elliott, E., and Dennison, C. (2002). Endolysosomal proteolysis and its regulation. Biochem. J. 363, 417–429. doi: 10.1042/0264-6021:3630417
Pohlmann, T., Baumann, S., Haag, C., Albrecht, M., and Feldbrügge, M. (2015). A FYVE zinc finger domain protein specifically links mRNA transport to endosome trafficking. eLife 4:e06041. doi: 10.7554/eLife.06041
Price, L. S., and Bos, J. L. (2004). RAPL: taking the Rap in immunity. Nat. Immunol. 5, 1007–1008. doi: 10.1038/ni1004-1007
Puleston, D. J., and Simon, A. K. (2014). Autophagy in the immune system. Immunology 141, 1–8. doi: 10.1111/imm.12165
Raiborg, C., Schink, K. O., and Stenmark, H. (2013). Class III phosphatidylinositol 3–kinase and its catalytic product PtdIns3P in regulation of endocytic membrane traffic. FEBS J. 280, 2730–2742. doi: 10.1111/febs.12116
Raiborg, C., Wenzel, E. M., Pedersen, N. M., Olsvik, H., Schink, K. O., Schultz, S. W., et al. (2015). Repeated ER-endosome contacts promote endosome translocation and neurite outgrowth. Nature 520, 234–238. doi: 10.1038/nature14359
Raiborg, C., Wenzel, E. M., Pedersen, N. M., and Stenmark, H. (2016). ER–endosome contact sites in endosome positioning and protrusion outgrowth. Biochem. Soc. Trans. 44, 441–446. doi: 10.1042/BST20150246
Rothman, J. E. (2002). The machinery and principles of vesicle transport in the cell. Nat. Med. 8, 1059–1062. doi: 10.1038/nm770
Rothwell, S., Lilleker, J. B., and Lamb, J. A. (2017). Genetics in inclusion body myositis. Curr. Opin. Rheumatol. 29, 639–644. doi: 10.1097/BOR.0000000000000431
Saridaki, T., Nippold, M., Dinter, E., Roos, A., Diederichs, L., Fensky, L., et al. (2018). FYCO1 mediates clearance of α-synuclein aggregates through a Rab7-dependent mechanism. J. Neurochem. 146, 474–492. doi: 10.1111/jnc.14461
Schink, K. O., Raiborg, C., and Stenmark, H. (2013). Phosphatidylinositol 3-phosphate, a lipid that regulates membrane dynamics, protein sorting and cell signalling. Bioessays 35, 900–912. doi: 10.1002/bies.201300064
Schwabe, J. W. R., and Klug, A. (1994). Zinc mining for protein domains. Nat. Struct. Mol. Biol. 1, 345–349. doi: 10.1038/nsb0694-345
Scott, C. C., Vacca, F., and Gruenberg, J. (2014). Endosome maturation, transport and functions. Semin. Cell Dev. Biol. 31, 2–10. doi: 10.1016/j.semcdb.2014.03.034
Shin, N., You, K. T., Lee, H., Kim, W. K., Song, M., Choi, H.-J., et al. (2011). Identification of frequently mutated genes with relevance to nonsense mediated mRNA decay in the high microsatellite instability cancers. Int. J. Cancer 128, 2872–2880. doi: 10.1002/ijc.25641
Sillars-Hardebol, A. H., Carvalho, B., de Wit, M., Postma, C., Delis-van Diemen, P. M., Mongera, S., et al. (2010). Identification of key genes for carcinogenic pathways associated with colorectal adenoma-to-carcinoma progression. Tumour Biol. 31, 89–96. doi: 10.1007/s13277-009-0012-1
Solé-Domènech, S., Cruz, D. L., Capetillo-Zarate, E., and Maxfield, F. R. (2016). The endocytic pathway in microglia during health, aging and Alzheimer’s disease. Ageing Res. Rev. 32, 89–103. doi: 10.1016/j.arr.2016.07.002
Søreng, K., Neufeld, T. P., and Simonsen, A. (2018). “Chapter One - Membrane trafficking in autophagy,” in International Review of Cell and Molecular Biology, ed. L. Galluzzi (Cambridge, MA: Academic Press), 1–92. doi: 10.1016/bs.ircmb.2017.07.001
Stahelin, R. V., Long, F., Diraviyam, K., Bruzik, K. S., Murray, D., and Cho, W. (2002). Phosphatidylinositol 3-phosphate induces the membrane penetration of the FYVE domains of Vps27p and Hrs. J. Biol. Chem. 277, 26379–26388. doi: 10.1074/jbc.M201106200
Stalder, D., and Gershlick, D. C. (2020). Direct trafficking pathways from the Golgi apparatus to the plasma membrane. Semin. Cell Dev. Biol. doi: 10.1016/j.semcdb.2020.04.001 [Epub ahead of print].
Staubitz, J. I., Schad, A., Springer, E., Rajalingam, K., Lang, H., Roth, W., et al. (2019). Novel rearrangements involving the RET gene in papillary thyroid carcinoma. Cancer Genet. 230, 13–20. doi: 10.1016/j.cancergen.2018.11.002
Stenmark, H. (2009). Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 10, 513–525. doi: 10.1038/nrm2728
Stenmark, H., and Aasland, R. (1999). FYVE-finger proteins–effectors of an inositol lipid. J. Cell Sci. 112, 4175–4183.
Stenmark, H., Aasland, R., Toh, B. H., and D’Arrigo, A. (1996). Endosomal localization of the autoantigen EEA1 is mediated by a zinc-binding FYVE finger. J. Biol. Chem. 271, 24048–24054.
Stenmark, H., and Gillooly, D. J. (2001). Intracellular trafficking and turnover of phosphatidylinositol 3-phosphate. Semin. Cell Dev. Biol. 12, 193–199. doi: 10.1006/scdb.2000.0236
Sun, Q., Han, C., Liu, L., Wang, Y., Deng, H., Bai, L., et al. (2012). Crystal structure and functional implication of the RUN domain of human NESCA. Protein Cell 3, 609–617. doi: 10.1007/s13238-012-2052-3
Tabuse, M., Ohta, S., Ohashi, Y., Fukaya, R., Misawa, A., Yoshida, K., et al. (2011). Functional analysis of HOXD9 in human gliomas and glioma cancer stem cells. Mol. Cancer 10:60. doi: 10.1186/1476-4598-10-60
Takai, Y., Sasaki, T., and Matozaki, T. (2001). Small GTP-binding proteins. Physiol. Rev. 81, 153–208. doi: 10.1152/physrev.2001.81.1.153
Takeuchi, H., Takada, A., Kuboniwa, M., and Amano, A. (2016). Intracellular periodontal pathogen exploits recycling pathway to exit from infected cells. Cell. Microbiol. 18, 928–948. doi: 10.1111/cmi.12551
Taniuchi, K., Tsuboi, M., Sakaguchi, M., and Saibara, T. (2018). Measurement of serum PODXL concentration for detection of pancreatic cancer. Onco Targets Ther. 11, 1433–1445. doi: 10.2147/OTT.S155367
Tapia, D., Jiménez, T., Zamora, C., Espinoza, J., Rizzo, R., González-Cárdenas, A., et al. (2019). KDEL receptor regulates secretion by lysosome relocation- and autophagy-dependent modulation of lipid-droplet turnover. Nat. Commun. 10:735. doi: 10.1038/s41467-019-08501-w
Terawaki, S., Camosseto, V., Pierre, P., and Gatti, E. (2016). RUFY4: Immunity piggybacking on autophagy? Autophagy 12, 598–600. doi: 10.1080/15548627.2015.1136772
Terawaki, S., Camosseto, V., Prete, F., Wenger, T., Papadopoulos, A., Rondeau, C., et al. (2015). RUN and FYVE domain–containing protein 4 enhances autophagy and lysosome tethering in response to Interleukin-4. J. Cell Biol. 210, 1133–1152. doi: 10.1083/jcb.201501059
Thomas, L. L., and Fromme, J. C. (2020). Extensive GTPase crosstalk regulates Golgi trafficking and maturation. Curr. Opin. Cell Biol. 65, 1–7. doi: 10.1016/j.ceb.2020.01.014
Torti, M., Bertoni, A., Canobbio, I., Sinigaglia, F., Lapetina, E. G., and Balduini, C. (1999). Interaction of the low-molecular-weight GTP-binding protein rap2 with the platelet cytoskeleton is mediated by direct binding to the actin filaments. J. Cell. Biochem. 75, 675–685.
Vadlamudi, R. K., and Kumar, R. (2003). P21-activated kinases in human cancer. Cancer Metastasis Rev. 22, 385–393. doi: 10.1023/a:1023729130497
Vega-Cabrera, L. A., and Pardo-López, L. (2017). Membrane remodeling and organization: elements common to prokaryotes and eukaryotes. IUBMB Life 69, 55–62. doi: 10.1002/iub.1604
Vongchan, P., and Linhardt, R. J. (2017). Characterization of a new monoclonal anti-glypican-3 antibody specific to the hepatocellular carcinoma cell line, HepG2. World J. Hepatol. 9, 368–384. doi: 10.4254/wjh.v9.i7.368
Vukmirica, J., Monzo, P., Le Marchand-Brustel, Y., and Cormont, M. (2006). The Rab4A effector protein Rabip4 is involved in migration of NIH 3T3 fibroblasts. J. Biol. Chem. 281, 36360–36368. doi: 10.1074/jbc.M602920200
Wang, G., Zhang, Q., Song, Y., Wang, X., Guo, Q., Zhang, J., et al. (2015). PAK1 regulates RUFY3-mediated gastric cancer cell migration and invasion. Cell Death Dis. 6:e1682. doi: 10.1038/cddis.2015.50
Wang, Q., Zeng, F., Sun, Y., Qiu, Q., Zhang, J., Huang, W., et al. (2018). Etk Interaction with PFKFB4 modulates chemoresistance of small-cell lung cancer by regulating autophagy. Clin. Cancer Res. 24, 950–962. doi: 10.1158/1078-0432.CCR-17-1475
Wang, T., Ming, Z., Xiaochun, W., and Hong, W. (2011). Rab7: role of its protein interaction cascades in endo-lysosomal traffic. Cell. Signal. 23, 516–521. doi: 10.1016/j.cellsig.2010.09.012
Wei, Z., Sun, M., Liu, X., Zhang, J., and Jin, Y. (2014). Rufy3, a protein specifically expressed in neurons, interacts with actin-bundling protein Fascin to control the growth of axons. J. Neurochem. 130, 678–692. doi: 10.1111/jnc.12740
Wong, S.-W., Sil, P., and Martinez, J. (2018). Rubicon: LC3-associated phagocytosis and beyond. FEBS J. 285, 1379–1388. doi: 10.1111/febs.14354
Wu, Y., Peng, Y., Wu, M., Zhang, W., Zhang, M., Xie, R., et al. (2016). Oncogene FOXK1 enhances invasion of colorectal carcinoma by inducing epithelial-mesenchymal transition. Oncotarget 7, 51150–51162. doi: 10.18632/oncotarget.9457
Xie, R., Wang, J., Liu, X., Wu, L., Zhang, H., Tang, W., et al. (2017a). RUFY3 interaction with FOXK1 promotes invasion and metastasis in colorectal cancer. Sci. Rep. 7:3709. doi: 10.1038/s41598-017-04011-1
Xie, R., Wang, J., Tang, W., Li, Y., Peng, Y., Zhang, H., et al. (2017b). Rufy3 promotes metastasis through epithelial–mesenchymal transition in colorectal cancer. Cancer Lett. 390, 30–38. doi: 10.1016/j.canlet.2017.01.001
Xu, J., Shi, S., Matsumoto, N., Noda, M., and Kitayama, H. (2007). Identification of Rgl3 as a potential binding partner for Rap-family small G-proteins and profilin II. Cell. Signal. 19, 1575–1582. doi: 10.1016/j.cellsig.2007.02.004
Yamada, Y., Sakuma, J., Takeuchi, I., Yasukochi, Y., Kato, K., Oguri, M., et al. (2017). Identification of five genetic variants as novel determinants of type 2 diabetes mellitus in Japanese by exome-wide association studies. Oncotarget 8, 80492–80505. doi: 10.18632/oncotarget.19287
Yamamoto, H., Koga, H., Katoh, Y., Takahashi, S., Nakayama, K., and Shin, H.-W. (2010). Functional cross-talk between Rab14 and Rab4 through a dual effector, RUFY1/Rabip4. Mol. Biol. Cell 21, 2746–2755. doi: 10.1091/mbc.e10-01-0074
Yang, J., Kim, O., Wu, J., and Qiu, Y. (2002). Interaction between tyrosine kinase Etk and a RUN domain- and FYVE domain-containing protein RUFY1. A possible role of ETK in regulation of vesicle trafficking. J. Biol. Chem. 277, 30219–30226. doi: 10.1074/jbc.M111933200
Yoshida, H., Kitagishi, Y., Okumura, N., Murakami, M., Nishimura, Y., and Matsuda, S. (2011). How do you RUN on? FEBS Lett. 585, 1707–1710. doi: 10.1016/j.febslet.2011.05.011
Yoshida, H., Okumura, N., Kitagishi, Y., Shirafuji, N., and Matsuda, S. (2010). Rab5(Q79L) interacts with the carboxyl terminus of RUFY3. Int. J. Biol. Sci. 6, 187–189. doi: 10.7150/ijbs.6.187
Yuan, W., and Song, C. (2020). The emerging role of Rab5 in membrane receptor trafficking and signaling pathways. Biochem. Res. Int. 2020:4186308. doi: 10.1155/2020/4186308
Zelaya, M. V., Pérez-Valderrama, E., de Morentin, X. M., Tuñon, T., Ferrer, I., Luquin, M. R., et al. (2015). Olfactory bulb proteome dynamics during the progression of sporadic Alzheimer’s disease: identification of common and distinct olfactory targets across Alzheimer-related co-pathologies. Oncotarget 6, 39437–39456. doi: 10.18632/oncotarget.6254
Zheng, J., Su, Z., Kong, Y., Lin, Q., Liu, H., Wang, Y., et al. (2020). LncRNAs predicted to interfere with the gene regulation activity of miR-637 and miR-196a-5p in GBM. Front. Oncol. 10:303. doi: 10.3389/fonc.2020.00303
Zheng, Z., Liebers, M., Zhelyazkova, B., Cao, Y., Panditi, D., Lynch, K. D., et al. (2014). Anchored multiplex PCR for targeted next-generation sequencing. Nat. Med. 20, 1479–1484. doi: 10.1038/nm.3729
Zhi, Q., Chen, H., Liu, F., Han, Y., Wan, D., Xu, Z., et al. (2019). Podocalyxin-like protein promotes gastric cancer progression through interacting with RUN and FYVE domain containing 1 protein. Cancer Sci. 110, 118–134. doi: 10.1111/cas.13864
Zhong, Y., Wang, Q. J., Li, X., Yan, Y., Backer, J. M., Chait, B. T., et al. (2009). Distinct regulation of autophagic activity by Atg14L and Rubicon associated with Beclin 1-phosphatidylinositol-3-kinase complex. Nat. Cell Biol. 11, 468–476. doi: 10.1038/ncb1854
Zhong, Z., Sanchez-Lopez, E., and Karin, M. (2016). Autophagy, inflammation, and immunity: a troika governing cancer and its treatment. Cell 166, 288–298. doi: 10.1016/j.cell.2016.05.051
Zhu, H., Dai, W., Li, J., Xiang, L., Wu, X., Tang, W., et al. (2019). HOXD9 promotes the growth, invasion and metastasis of gastric cancer cells by transcriptional activation of RUFY3. J. Exp. Clin. Cancer Res. 38:412. doi: 10.1186/s13046-019-1399-1
Keywords: RUFY, cancer, neurodegenerative diseases, immunity, RUN, FYVE, phosphatidylinositol 3-phosphate, cytoskeleton
Citation: Char R and Pierre P (2020) The RUFYs, a Family of Effector Proteins Involved in Intracellular Trafficking and Cytoskeleton Dynamics. Front. Cell Dev. Biol. 8:779. doi: 10.3389/fcell.2020.00779
Received: 11 June 2020; Accepted: 24 July 2020;
Published: 11 August 2020.
Edited by:
Roberto Botelho, Ryerson University, CanadaReviewed by:
Daniel G. S. Capelluto, Virginia Tech, United StatesBrian Paul Ceresa, University of Louisville, United States
Copyright © 2020 Char and Pierre. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rémy Char, Y2hhckBjaW1sLnVuaXYtbXJzLmZy; Philippe Pierre, cGllcnJlQGNpbWwudW5pdi1tcnMuZnI=