
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cardiovasc. Med. , 14 March 2023
Sec. Coronary Artery Disease
Volume 10 - 2023 | https://doi.org/10.3389/fcvm.2023.1104223
This article is part of the Research Topic The Clinical Use of Coronary Functional Test in Coronary Artery Disease View all 7 articles
 PhongSon Dinh1,2
PhongSon Dinh1,2 JunHua Peng1,3
JunHua Peng1,3 ThanhLoan Tran1,4
ThanhLoan Tran1,4 DongFeng Wu5
DongFeng Wu5 ChauMyThanh Tran2
ChauMyThanh Tran2 ThiPhuongHoai Dinh6
ThiPhuongHoai Dinh6 ShangLing Pan1,3*
ShangLing Pan1,3*
Objects: To evaluate the hsa_circ_0001445 level in peripheral blood leukocytes of patients with coronary heart disease (CHD) and its related clinical factors, and predict its circRNA-miRNA-mRNA regulatory network in CHD pathogenesis via bioinformatics analysis.
Methods: Peripheral blood leukocytes were isolated from the whole blood samples of 94 CHD patients (aged 65.96 ± 9.78 years old) and 126 healthy controls (aged 60.75 ± 8.81 years old). qRT-PCR was used to quantify the expression level of circRNA and subsequently analyze its association with CHD clinical parameters. Via bioinformatics algorithm and GEO datasets, differential miRNA expression was evaluated using the Limma package. A miRNA-mRNA regulatory network was predicted by cyTargetLinker. ClusterProfiler was employed to perform functional enrichment analysis of the circRNA network to investigate its role in CHD pathogenesis.
Results: The expression of hsa_circ_0001445 in peripheral blood leukocytes of CHD patients was downregulated compared with that of healthy controls. Positive correlations were evident between hsa_circ_0001445 expression level and the levels of hemoglobin, triglycerides, high- and low-density lipoprotein cholesterol. A significant negative correlation was also found between hsa_circ_0001445 expression level and age and the neutrophil level. Low expression of hsa_circ_0001445 exhibited a discriminatory ability between CHD patients and healthy controls with a sensitivity of 67.5% and a specificity of 76.6% (p < 0.05). By bioinformatics analysis, 405 gene ontology terms were identified. The Kyoto Encyclopedia of Genes and Genomes terms focused principally on the PI3K-Akt signaling pathway. hsa_circ_0001445 was associated with the expression of three miRNAs that may regulate 18 genes involved in KEGG processes: hsa-miR-507, hsa-miR-375–3p, and hsa-miR-942–5p.
Conclusion: The hsa_circ_0001445 level in peripheral blood leukocytes may serve as a biomarker for CHD diagnosis. Our work on circRNA-miRNA-mRNA networks suggests a potential role for hsa_circ_0001445 in CHD development.
Coronary heart disease (CHD) is the most common heart condition worldwide and the leading cause of death of elderly men and women (1). Despite recent declines in developed countries, both CHD morbidity and mortality continue to increase rapidly in developing countries. Various factors are involved in CHD pathogenesis, including older age, dyslipidemia, obesity, psychological issues, hypercholesterolemia, diabetes, and family history (2–4). Currently, most efforts focus on biochemical tests, protein and gene based biomarkers to predict the incidence of CHD (5, 6). However, the ability of these factors in terms of early detection of CHD are still controversial (5). Recently, circular RNAs (circRNAs) have emerged as a novel potential non-invasive biomarker for diagnosis and prognostic of CHD patients.
CircRNAs are a kind of non-coding RNA that consists of continuous covalently closed loops without the 3′- and 5′ end like linear RNA, which enables it to resist degradation, and thus has relative conservation and stability (7). Recently, their functions and biological features have been extensively studied. They modify gene expression by serving as microRNA (miRNA) sponges that bind and inactivate certain miRNAs (8–10). Research on their roles in cardiovascular diseases has progressed rapidly. In particular, in-depth studies of the relationship between circRNA and CHD have provided effective tools to early diagnose CHD and by that reduced CHD mortality (5, 7, 11).
Recently, Vilades et al. (6) showed that the plasma levels of the circRNA hsa_circ_0001445 (hsa_circSMARCA5_013) were proportional to coronary atherosclerotic burden. hsa_circ_0001445 has been consistently detected in clinically relevant samples, including heart tissue (12), plasma (13), serum (14), and whole blood (15). In this study, we first investigated the levels of hsa_circ_0001445 in peripheral blood leukocytes (PBLs) of CHD patients from Guangxi, China (6), and analyzed its correlation with clinical characteristics. Next, we performed bioinformatics analyses to define a novel circRNA-miRNA-mRNA network involved in CHD. Finally, we conducted functional and pathway enrichment analyses of potentially relevant genes. Our findings may provide potential candidate for further studies on the pathogenesis of CHD.
The experimental group included 94 CHD patients aged 65.96 ± 9.78 years (58 men and 36 women) admitted to the Department of Cardiology of the People's Autonomous Hospital of Guangxi Zhuang from January 1 2019 to December 31 2020. All patients underwent coronary angiography (CAG), and those with stenoses ≥50% in at least one of the three main coronary arteries or their major branches (diameter ≥2 mm) were diagnosed with CHD. The exclusion criteria were diabetes mellitus, any other clinically acute or chronic inflammatory systemic disease, uncontrolled hypertension, liver or kidney dysfunction, endocrine disease, autoimmune disease, a malignancy, prior percutaneous coronary intervention (PCI) or coronary artery bypass grafting (CABG), and a history of CHD. The control group included 126 healthy subjects aged 60.75 ± 8.82 years (61 men and 65 women) recruited in the same period from the Second Affiliated Hospital of Guangxi Medical University. They were confirmed healthy after physical check-ups; none had a history of coronary atherosclerosis or microvascular disease. This study was conducted in accordance with the Declaration of Helsinki (1975) and was approved by the ethics committee of Guangxi Medical University (approval no. 2019-SB-060). All patients and controls gave written informed consent.
Total RNAs were extracted from PBLs of CHD patients and healthy controls using the SanPrep column microRNA extraction kits (Sangon Biotech, China); all samples were stored at −80°C.
RNA reverse transcription into cDNA was performed using 5× HiScript III qRT SuperMix kits (Vazyme Biotech, China) according to the manufacturer's instructions. One microgram of RNA and 4x gDNA wiper mix were incubated at 42°C for 2 min, then 5× HiScript III qRT SuperMix was added, followed by incubation at 37°C for 15 min and 85°C for 5 s. The products served as qRT-PCR templates.
The expression level of hsa_circ_0001445 was detected by qRT-PCR with Light Cycler 96 platform (Roche, USA), and Glyceraldehyde-3-phosphate dehydrogenase (hGAPDH) served as the internal standard for normalization. The specific primers were listed as follows: (hsa_circ_0001445) forward primer: 5′-TGGGCGAAAGTTCACTTAGAA-3′, reverse primer: 5′- CACATGTGTTGCTCCATGTCT-3′; (hGAPDH) forward primer: 5′- TGTTGCCATCAATGACCCCTT-3′, reverse primer: 5′-CTCCACGACGTACTCAGCG-3′ (6, 16). Each sample was performed in triplicate. The reaction conditions included of 40 cycles at 95°C for 3 min, 95°C for 10 s, and 60°C for 60s; the dissolution curves were obtained via one cycle at 95°C for 5 s, 60°C for 1 min, and 97°C for 1 s. The expression level of hsa_circ_0001445 was calculated using the 2−ΔCt method relative to hGAPDH.
Data were statistically analyzed using SPSS 22.0 (SPSS Inc. Chicago, IL, USA) and GraphPad Prism 8 (GraphPad Software, San Diego, California, USA). Continuous data were presented as mean ± standard deviation (means ± SD) if normally distributed, and otherwise as median (interquartile range). The circRNA expression levels between CHD and controls were compared using the Student's t-test (calculated by SPSS) to determine statistically significant difference between the means of two groups. Spearman's rho coefficient was used to assess the correlation between continuous variables. Logistic regression was used to assess relationships between various factors and the PBL levels of hsa_circ_0001445. A p-value < 0.05 was considered significant. Analysis of receiver operating characteristic (ROC) curves was performed to calculate the optimal area under the ROC curve (AUC) for evaluating the CHD diagnostic ability of hsa_circ_0001445.
miRNA expression levels of CHD patients were collected from the public Gene Expression Omnibus database (GEO) (https://www.ncbi.nlm.nih.gov/geo/) using the following criteria: peripheral blood cells from humans, ≥3 samples of patients and normal controls, and miRNAs expression in CHD patients. The relevant datasets were GSE105449 (GPL22949) (17) and GSE61741 (GPL9040) (18). There were 236 samples in total (136 control and 100 CHD) that met the above criteria. GSE IDs, the high-throughput data, and the annotated subject characteristics of the control and CHD groups were collected in a series matrix file and were analyzed using R software ver. 3.6.2.
R software (Version 3.6.2; R Foundation for Statistical Computing, Vienna, Austria) (https://www.r-project.org/), the Bioconductor software package (https://bioconductor.org/packages) (19), and the limma package (20) were used to analyze differential miRNA expression based on miRNA expression data. All p-values were corrected using the false discovery rate (FDR) correction toolkit. A p-value <0.05 and with an FDR < 0.05 for all GSE files (fold change >1) were considered significant. Then, Venn diagrams were framed to identify overlapping miRNAs among the predicted datas in order to determine which potential miRNAs were associated with hsa_circ_0001445 in CHD. miRNAs identified as significant were entered into Cytoscape ver. 3.6.1 (http://cytoscape.org/) and were used to establish a network. A regulatory network was predicted by cyTargetLinker (https://cytargetlinker.github.io/) using the targetscan-hsa-7 and miRTarBase-hsa-7 databases (21). Finally, a miRNA-mRNA network was constructed.
ClusterProfiler ver. 3.14.3 package in R software was used to analyze the gene ontology and signaling pathways. Genome-wide annotation for humans was based on mapping employing the Entrez gene identifiers; we used several methods to visualize and interpret the functional enrichment results. A p-value <0.05 served as a cut-off when determining significant enrichments of GO terms and KEGG pathways. That is, the more likely that the gene associated with the listed entry/pathway influences cellular life activities and warrants further research (22). Venny 2.1 diagram was used to improve predictive accuracies (via intersection) (https://bioinfogp.cnb.csic.es/tools/venny/). Finally, the interaction network of miRNAs with genes of the KEGG pathway was established.
The expression level of hsa_circ_0001445 in PBLs of CHD patients was significantly lower than that of healthy controls (0.640 ± 0.254 and 1.079 ± 0.453, respectively) (p < 0.001) (Figure 1).
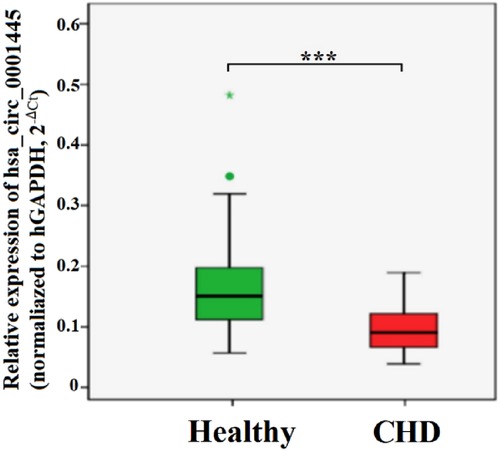
Figure 1. Expression levels of hsa_circ_0001445 in the healthy and CHD groups. p < 0.001.
We compared the age, sex, blood parameters, fasting blood glucose levels (GLU), three renal function items (UREA, CREA, and UA), three liver enzymes (AST, ALT, GGT) and cholesterol levels of the two groups. As shown in Table 1, age, white blood cell count (WBC), and levels of hematocrit (HCT), neutrophils (NEU), red blood cells (RBCs), hemoglobin (HGB), platelets (PLT), alanine aminotransferase (ALT), aspartate transaminase (AST), glutamyl transpeptidase (GGT), glucose (GLU), total cholesterol (T-Cho), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C) significantly differed between the two groups (all p < 0.05).
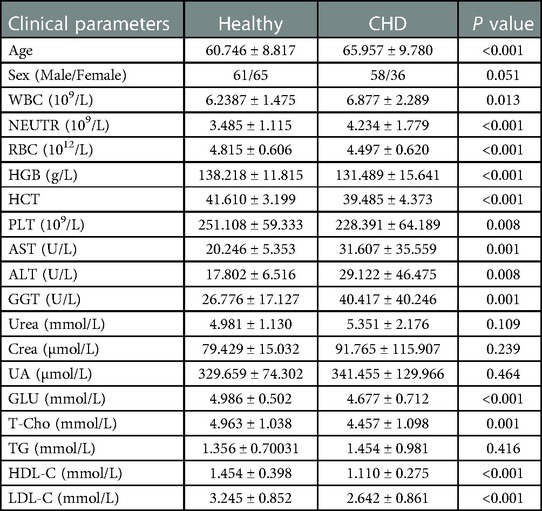
Table 1. Clinical parameters of the CHD patients and healthy controls.
Clinical parameters associated with hsa_circ_0001445 expression level in CHD patients were shown in Table 2. Its expression was negatively correlated with age and NEU but positively correlated with HGB, T-Cho, HDL-C, and LDL-C (p < 0.05).
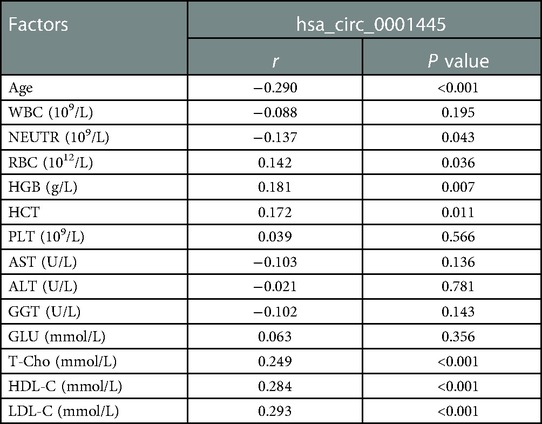
Table 2. Correlations between hsa_circ_0001445 expression level and clinical parameters of CHD patients.
We used multivariate logistic regression to explore whether hsa_circ_0001445 independently predicted CHD. Model 1 included NEU and HGB; model 2 included T-Cho, HDL-C, and LDL-C; and model 3 included NEU, HGB, T-Cho, HDL-C, and LDL-C. Based on the median 2−ΔCt values for hsa_circ_0001445, the CHD group was subdivided into those with low and high circRNA expression. In addition, based on the normal ranges and median values of NEU, HGB, T-Cho, and LDL-C, patients were subdivided into high and normal subgroups. The HDL-C values were used to define normal- and low-expression subgroups.
Low hsa_circ_0001445 expression level was an independent risk factor for CHD in all three models (Table 3).
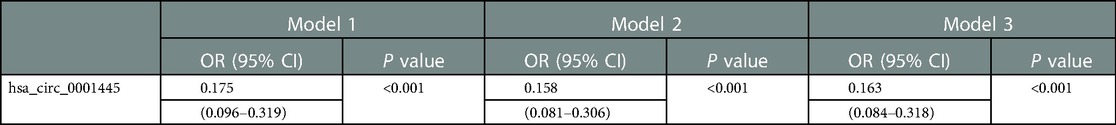
Table 3. Logistic regression results.
ROC curve analyses of the healthy and CHD groups showed that the AUC for hsa_circ_0001445 was 0.816 ± 0.028 (95% CI 0.761–0.871; p < 0.001; Figure 2). It had the highest J index [(Se + Sp − 1) = 0.441]; a 2−ΔCt value of 0.814 was thus chosen as the cut-off. The sensitivity was 67.5%, the specificity was 76.6%, and the likelihood ratio [Se/(1-Sp)] was 2.88.
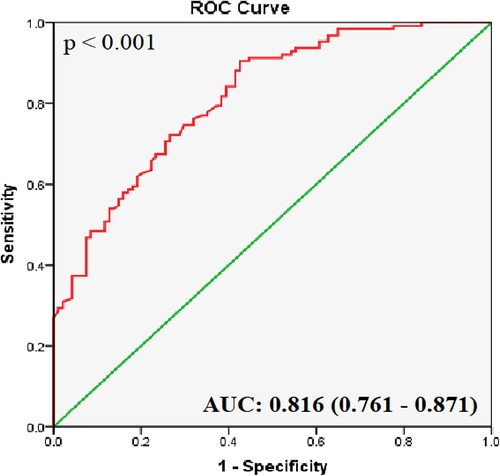
Figure 2. ROC curve analysis of hsa_circ_0001445. The AUC shows the diagnostic ability of hsa_circ_0001445 in terms of CHD (p < 0.001).
We found 132 differentially expressed miRNAs in CHD, including 79 that were upregulated and 53 that were downregulated in GSE105449 and GSE61741 dataset; all met the cutoff criteria of abs (log2FC) >1 and p < 0.05 (Supplementary Table S1). The top 10 up- and downregulated miRNAs are listed in Tables 4, 5.

Table 4. Top 10 DmiREs from GSE105449.
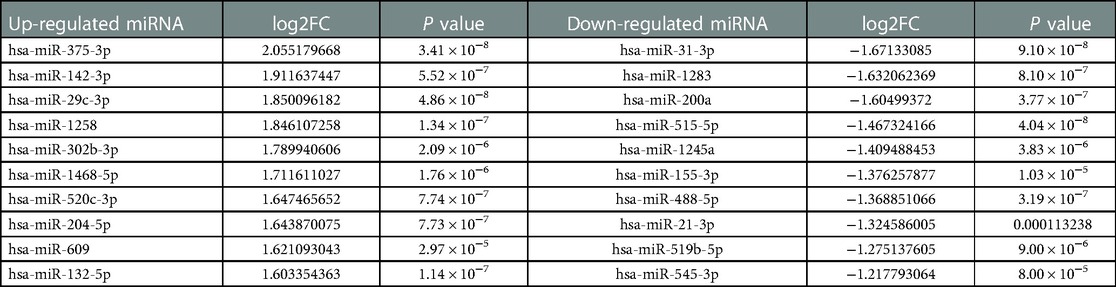
Table 5. Top 10 DmiREs from GSE61741.
The Circbank and Circinteractome databases were used to predict interactions between hsa_circ_0001445 and miRNAs. We identified 26 interacting pairs.
We used Venny ver. 2.1 to visualize the Circbank and Circinteractome data, the upregulated miRNAs of GSE105449 and GSE61741 dataset; and their intersections (Figure 3). The predicted target miRNAs of hsa_circ_0001445 were hsa-miR-507, hsa-miR-375–3p, hsa-miR-576–5p, and hsa-miR-942–5p. These four miRNAs were entered into Cytoscape ver. 3.6.1. The CyTargetLinker app simply links Cytoscape networks to miRNA-mRNA interactions. A total of 1,175 mRNAs were obtained and the Gene IDs were converted into Entrez IDs for GO and KEGG analyses using R software and the Perl tool.
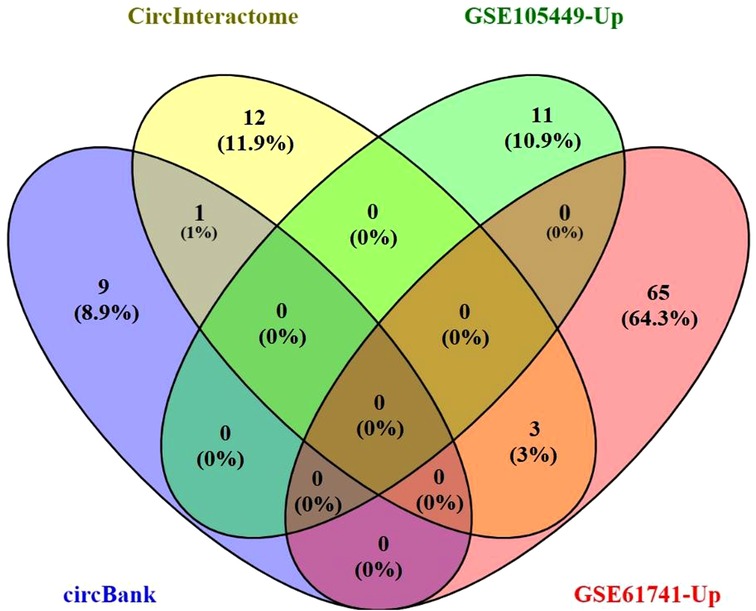
Figure 3. The Venn diagram for the overlap of numbers of the predicted miRNAs.
GO terms were analyzed using R ver. 3.6.2 and clusterProfiler ver. 3.14.3 followed by pathway enrichment. A total of 405 GO terms were found (Supplementary Table S2). These covered molecular function pathways, as shown in the table. The GO terms were ranked by their adjusted p-values (p-value <0.05; p-adjusted <0.05) (Figure 4).
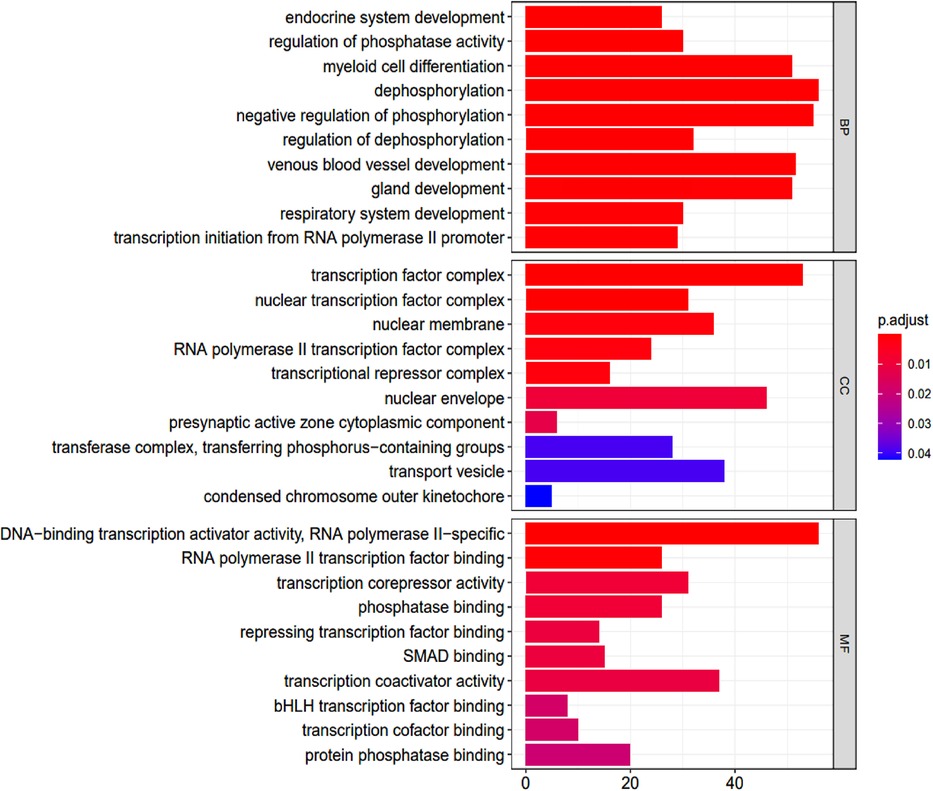
Figure 4. Go enrichment analysis. BP, biological process; CC, cellular component; MF, molecular function.
The GO terms indicated biological process pathways such as dephosphorylation, negative regulation of phosphorylation, and venous blood vessel development (Figure 5).
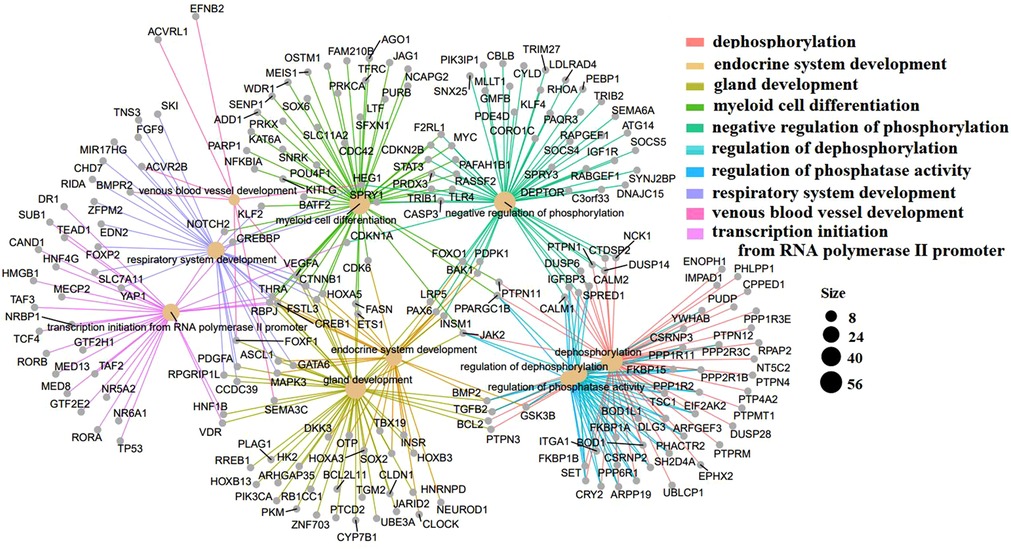
Figure 5. Top 10 GO terms with the lowest p-values. Biological pathways and associated genes were identified. Each node indicates a biological process in the path. The lines are the interactions between the genes and the terms.
KEGG enrichment analysis identified 70 pathways (p-values <0.05) (Supplementary Table S3). The top 30 are summarized in Figure 6.
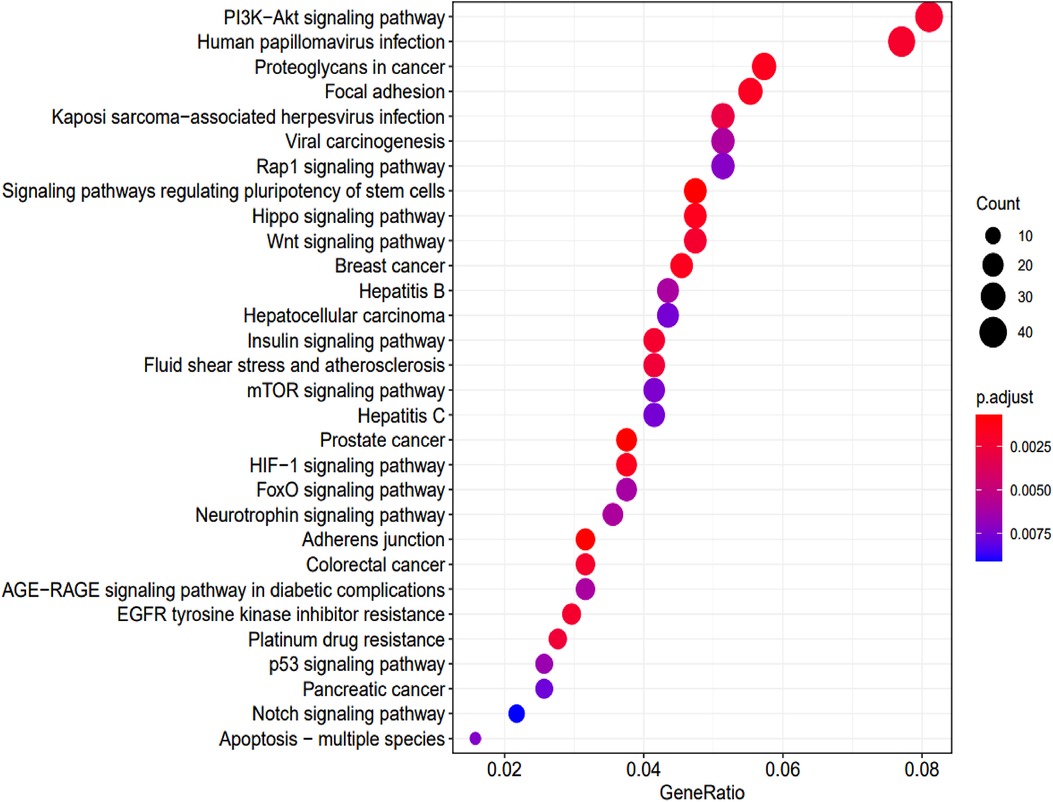
Figure 6. KEGG pathway analysis and the numbers of associated genes. The pathways and the associated genes were ranked based on the p-values. The sizes of the dots represent the gene counts of each row (KEGG categories).
To enhance accuracy, the results afforded by the mirtarbase and targetscan databases were intersected with those described above (Figure 7). This yielded 63 genes that were compared to KEGG pathway genes. We found 18 potential KEGG genes. Finally, we built a subnetwork of the interactions between miRNAs and these genes. This included the signaling pathways mentioned.
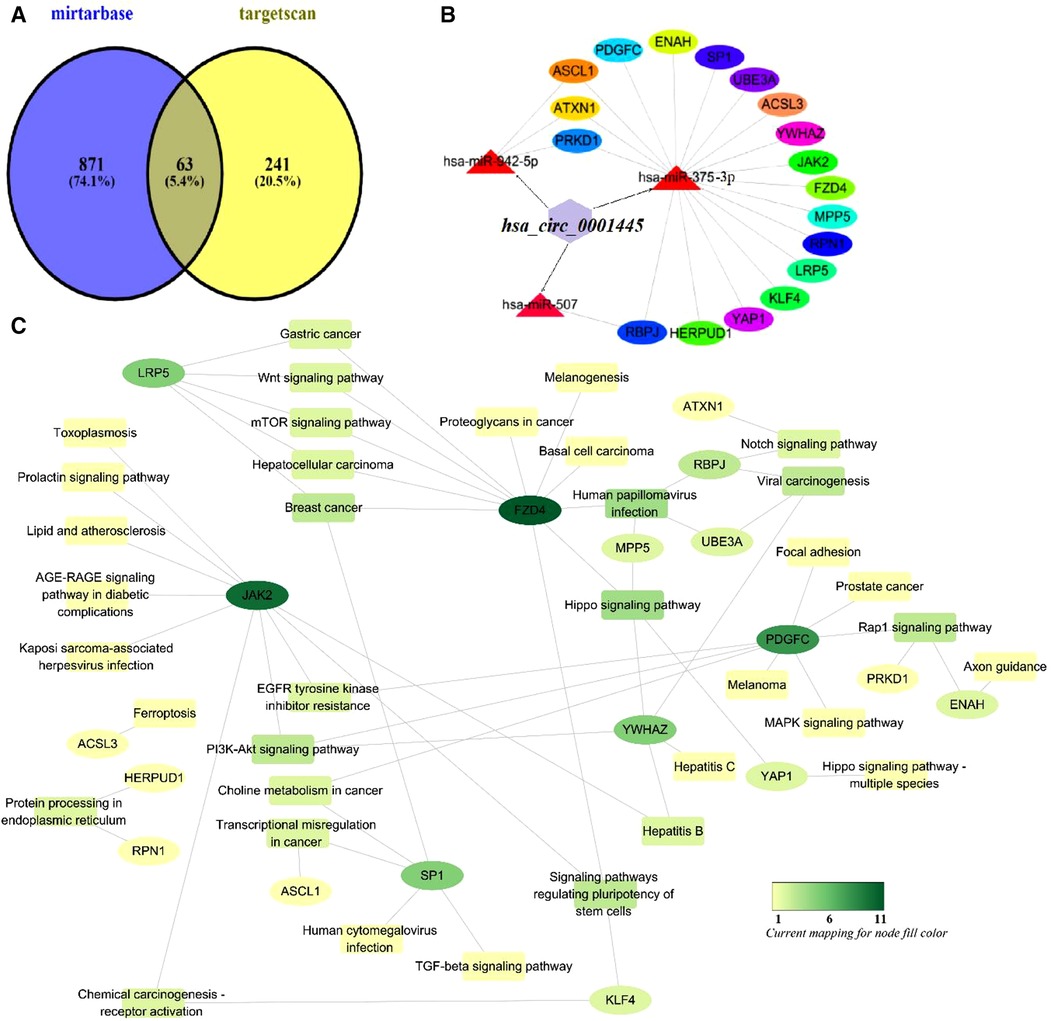
Figure 7. Mechanism of action of hsa_circ_0001445. (A) Venn diagram of genes predicted by mirtarbase and targetscan. (B) The subnetwork of the interactions between miRNAs and these genes. (C) Functional analysis of potential genes of the KEGG pathway using Cytoscape ver. 3.6.1.
Identifying factors involved in CHD pathogenesis can not only improves our understanding of its development but also suggests new approaches to the diagnosis, prognosis, and management of CHD. Numerous biomarkers associated with CHD have been applied, however, to identify its primary stage by regular examinations, such as cardiac ultrasound and electrocardiography, remains challenging. CHD diagnosis in its early stage with a sensitive biomarker are crucial for treatment and prognosis. Recent large-scale studies have suggested that circRNAs play an essential role in the pathogenesis and progression of CHD (5, 6, 12, 23). In this study, we identified for the first time the circRNA hsa_circ_0001445 as a potential diagnostic biomarker for CHD. By evaluating the expression level of hsa_circ_0001445 in CHD patients and control group, our results showed that the expression of hsa_circ_0001445 in PBLs of CHD patients was significantly lower than that of healthy controls. ROC curve analysis indicated that its expression level well-distinguished CHD patients from healthy subjects with AUC valued 0.816 ± 0.028 (95% CI 0.761–0.871; p < 0.001). In addition, logistic regression revealed that hsa_circ_0001445 was an independent predictor of CHD. Further studies on hsa_circ_0001445 as well as other circRNAs involved in CHD is required to investigate their potential in CHD diagnosis.
The RNA binding protein Quaking (QKI) promotes circRNA formation (24). QKI was proven to be involved in cell differentiation, apoptosis, proliferation, and migration (25). Interestingly, recent works have shown that QKI affects cardiovascular development and function (26–29), suggesting that QKI may affect the expression of hsa_circ_0001445 in CHD patients and consequently act on the disease progression. Several studies have also indicated that hsa_circ_0001445 may be involved in CHD development. Vilades et al. (6) found that plasma levels of hsa_circ_0001445 are lower in patients with higher coronary atherosclerotic burdens. Moreover, hsa_circ_0001445 is expressed by human coronary smooth muscle cells in vitro; its secretion is reduced under atherosclerotic conditions (6). Cai et al. (30) found that hsa_circ_0001445 is downregulated during low lipoprotein-induced oxidation of human umbilical vein endothelial cells. Overexpression of hsa_circ_0001445 promotes cell proliferation and inhibits the inflammatory response and apoptosis. These findings and our work suggest that hsa_circ_0001445 may involve in CHD development. Since the molecular mechanism of CHD is complicated, as is the effect of hsa_circ_0001445 on CHD, further research is required to elucidate its role in CHD pathogenesis.
The circRNA-miRNA-mRNA axis has been recently researched in terms of how circRNAs regulate CHD development. Lin et al. (31) constructed a network graph of the correlations between hsa-miR-101–5 and each of hsa_circ_0030769, hsa_circ_15486–161, hsa_circ_0122274, and hsa_circ_0079828. Miao et al. (5) hypothesized that the miRNAs regulated by hsa_circ_0016274 are miRNA-361–5p, miR-21–3p, miRNA-296–3p, and miRNA-375. The target genes of the circ-YOD1-miR-21–3p/miR-296–3p axis are BCL6, FBXL18, MMP9, and FCGR3B, as confirmed in other studies describing associations of MMP9, BCL6 (32, 33), hsa-miR-21–3p (32), and hsa-miR-296–3p (34) with CHD. In another study, the network of circ_ZNF609-related miRNAs included hsa-miR-615–5p, hsa-miR-145–5p, hsa-miR-138–5p, hsa-miR-150–5p (35), and AKT1 (a downstream target of miR-138–5p) (36). In this study, we predicted that certain genes affect CHD through bioinformatics analysis. This finding is in agreement with previous reports as JAK2 (37, 38), FZD4 (39), PDGFC (40), YWHAZ (41), SP1 (42), and LRP5 (43–45) which are all targets of hsa-miR-375–3p (46, 47). These genes participate in many signaling pathways, of which the PI3K-Akt pathway may play a major role in CHD (48–52). Phosphoinositide 3-kinase (PI3K) lies downstream of many receptor tyrosine kinases. PI3Ks play crucial roles in many aspects of biological response, such as membrane trafficking, cytoskeletal organization, cell growth and apoptosis. A serine/threonine kinase Akt, also known as protein kinase B, is the most well characterized target of PI3K. Akt is known to mediate cell survival signal by regulating several effectors, increases the rate of initiation of translation of mRNA by ribosomes such as Bad or procaspase-9, p70S6K. Stimulating and activating the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling pathway can regulate the expression of vascular endothelial cytokines, the polarization and survival of macrophages, the expression of inflammatory factors and platelet function, thus affecting the occurrence and development of CHD. This reinforces the authenticity of our interactive network prediction. The integrated biological information predicts the potential molecular mechanism of hsa_circ_0001445.
This study had certain limitations. First, it was a cross-sectional study with a modest sample size. Our findings require confirmation in larger studies to obtain higher reliability. Second, we did not perform luciferase assay, WB assay; but only database-derived links. Those binding assays would reinforce our suggestion that hsa_circ_0001445 is a good candidate biomarker of CHD. The mechanism by which the circRNA-miRNA-mRNA axis regulates CHD pathogenesis requires further in vivo and in vitro research.
We identified hsa_circ_0001445 as a potential biomarker for CHD diagnosis. The predicted genes involved in CHD participate in many signaling pathways, of which PI3K-Akt signaling may be particularly relevant to CHD. Our results provide a basis for further research on the molecular mechanism of hsa_circ_0001445 in CHD pathogenesis.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
The studies involving human participants were reviewed and approved by No. 2019-SB-060. The patients/participants provided their written informed consent to participate in this study.
PSD interpreted data, performed the statistical analyses and drafted the manuscript. TPHD and CMTT participated in the design and analyzed the bioinformatics results. TLT and DFW carried out the epidemiological survey and collected the samples. JHP, SLP designed the study, revised the manuscript and supervised the whole work. All authors contributed to the article and approved the submitted version.
The study was supported by the National Natural Science Foundation of China (grant no. 81660241, 81960266, 81860205).
We would like to thank patients and all the staff members in the Departments of Cardiology of The People's Hospital of Guangxi Zhuang Autonomous Region and The Second Affiliated Hospital of Guangxi Medical University for their support and cooperation in sample collection.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcvm.2023.1104223/full#supplementary-material.
Supplementary Table 1
The analysis for differentiallyexpressed miRNAs.
Supplementary Table 2
The analysis for Gene Ontology.
Supplementary Table 3
The analysis for KEGG pathway.
1. Virani S, Alonso A, Aparicio H, Benjamin E, Bittencourt M, Callaway C, et al. Heart disease and stroke statistics-2021 update A report from the American heart association. Circulation. (2021) 143:E254–743. doi: 10.1161/CIR.0000000000000950
2. Medina-Leyte DJ, Zepeda-García O, Domínguez-Pérez M, González-Garrido A, Villarreal-Molina T, Jacobo-Albavera L. Endothelial dysfunction, inflammation and coronary artery disease: potential biomarkers and promising therapeutical approaches. Int J Mol Sci. (2021) 22:3850. doi: 10.3390/ijms22083850
3. Sun HJ, Wu ZY, Nie XW, Bian JS. Role of endothelial dysfunction in cardiovascular diseases: the link between inflammation and hydrogen sulfide. Front Pharmacol. (2020) 10:1568. doi: 10.3389/fphar.2019.01568
4. Medina-Leyte DJ, Domínguez-Pérez M, Mercado I, Villarreal-Molina MT, Jacobo-Albavera L. Use of human umbilical vein endothelial cells (HUVEC) as a model to study cardiovascular disease: a review. Appl. Sci. (2020) 10:938. doi: 10.3390/app10030938
5. Miao L, Yin RX, Zhang QH, Liao PJ, Wang Y, Nie RJ, et al. A novel circRNA-miRNA-mRNA network identifies circ-YOD1 as a biomarker for coronary artery disease. Sci Rep. (2019) 9:18314. doi: 10.1038/s41598-019-54603-2
6. Vilades D, Martínez-Camblor P, Ferrero-Gregori A, Bär C, Lu DC, Xiao K, et al. Plasma circular RNA hsa_circ_0001445 and coronary artery disease: performance as a biomarker. FASEB J. (2020) 34:4403–14. doi: 10.1096/fj.201902507R
7. Fan X, Weng X, Zhao Y, Chen W, Gan T, Xu D. Circular RNAs in cardiovascular disease: an overview. Biomed Res Int. (2017) 2017:5135781. doi: 10.1155/2017/5135781
8. Barrett SP, Salzman J. Circular RNAs: analysis, expression and potential functions. Development. (2016) 143:1838–47. doi: 10.1242/dev.128074
9. Starke S, Jost I, Rossbach O, Schneider T, Schreiner S, Hung LH, et al. Exon circularization requires canonical splice signals. Cell Rep. (2015) 10:103–11. doi: 10.1016/j.celrep.2014.12.002
10. Lu M. Circular RNA: functions, applications and prospects. ExRNA. (2020) 2:1–7. doi: 10.1186/s41544-019-0046-5
11. Song CL, Wang JP, Xue X, Liu N, Zhang XH, Zhao Z, et al. Effect of circular ANRIL on the inflammatory response of vascular endothelial cells in a rat model of coronary atherosclerosis. Cell Physiol Biochem. (2017) 42:1202–12. doi: 10.1159/000478918
12. Schulte C, Barwari T, Joshi A, Theofilatos K, Zampetaki A, Barallobre-Barreiro J, et al. Comparative analysis of circulating noncoding rnas versus protein biomarkers in the detection of myocardial injury. Circ Res. (2019) 125:328–40. doi: 10.1161/CIRCRESAHA.119.314937
13. Maass PG, Glažar P, Memczak S, Dittmar G, Hollfinger I, Schreyer L, et al. A map of human circular RNAs in clinically relevant tissues. J Mol Med. (2017) 95:1179–89. doi: 10.1007/s00109-017-1582-9
14. Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J, et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. (2015) 25:981–4. doi: 10.1038/cr.2015.82
15. Sebastian M, Panagiotis P, Oliver P, Nikolaus R. Identification and characterization of circular RNAs as a new class of putative biomarkers in human blood. PLoS One. (2015) 10:e0141214. doi: 10.1371/journal.pone.0141214
16. Zhao Z, Li X, Gao C, Jian D, Hao P, Rao L, et al. Peripheral blood circular RNA hsa-circ-0124644 can be used as a diagnostic biomarker of coronary artery disease. Sci Rep. (2017) 7:39918. doi: 10.1038/srep39918
17. Kok MGM, de Ronde MWJ, Moerland PD, Ruijter JM, Creemers EE, Pinto-Sietsma SJ. Small sample sizes in high-throughput miRNA screens: a common pitfall for the identification of miRNA biomarkers. Biomol Detect Quantif. (2018) 15:1–5. doi: 10.1016/j.bdq.2017.11.002
18. Keller A, Leidinger P, Vogel B, Backes C, ElSharawy A, Galata V, et al. miRNAs can be generally associated with human pathologies as exemplified for miR-144. BMC Med. (2014) 12:224. doi: 10.1186/s12916-014-0224-0
19. Gentleman R, Carey V, Huber W, Irizarry R, Dudoit S. Bioinformatics and computational biology solutions using R and bioconductor. J Am Stat Assoc. (2007) 102:388–9. doi: 10.1198/jasa.2007.s179
20. Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. (2015) 43:e47. doi: 10.1093/nar/gkv007
21. Kohl M, Wiese S, Warscheid B. Cytoscape: software for visualization and analysis of biological networks. Methods Mol Biol. (2011) 696:291–303. doi: 10.1007/978-1-60761-987-1_18
22. Yu G, Wang G, Han Y, He Y. Clusterprofiler: an R package for comparing biological themes among gene clusters. OMICS. (2012) 16:284–7. doi: 10.1089/omi.2011.0118
23. Zhang S, Wang W, Wu X, Zhou X. Regulatory roles of circular RNAs in coronary artery disease. Mol Ther Nucleic Acids. (2020) 21:172–9. doi: 10.1016/j.omtn.2020.05.024
24. Conn SJ, Pillman KA, Toubia J, Conn VM, Salmanidis M, Phillips CA, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. (2015) 160:1125–34. doi: 10.1016/j.cell.2015.02.014
25. Chénard CA, Richard S. New implications for the QUAKING RNA binding protein in human disease. J Neurosci Res. (2008) 86:233–42. doi: 10.1002/jnr.21485
26. Chen X, Yin J, Cao D, et al. The emerging roles of the RNA binding protein QKI in cardiovascular development and function. Front Cell Dev Biol. (2021) 9:668659. doi: 10.3389/fcell.2021.668659
27. Chen X, Liu Y, Xu C, Ba L, Liu Z, Li XY, et al. QKI Is a critical pre-mRNA alternative splicing regulator of cardiac myofibrillogenesis and contractile function. Nat Commun. (2021) 12:89. doi: 10.1038/s41467-020-20327-5
28. Dehghan A, Bis JC, White CC, Smith AV, Morrison AC, Cupples LA, et al. Genome-Wide association study for incident myocardial infarction and coronary heart disease in prospective cohort studies: the CHARGE consortium. PLoS ONE. (2016) 11:e0144997. doi: 10.1371/journal.pone.0144997
29. Cochrane A, Kelaini S, Tsifaki M, Bojdo J, Vilà-González M, Drehmer D, et al. Quaking is a key regulator of endothelial cell differentiation, neovascularization, and angiogenesis. Stem Cells. (2017) 35:952–66. doi: 10.1002/stem.2594
30. Cai Y, Xu L, Xu C, Wang Y, Fan C. Hsa_circ_0001445 inhibits ox-LDL-induced HUVECs inflammation, oxidative stress and apoptosis by regulating miRNA-640. Perfusion. (2020) 37:86–94. doi: 10.1177/0267659120979472
31. Lin F, Zhao G, Chen Z, Wang X, Lv F, Zhang Y, et al. CircRNA-miRNA association for coronary heart disease. Mol Med Rep. (2019) 19:2527–36. doi: 10.3892/mmr.2019.9905
32. Dai J, Chen W, Lin Y, Wang S, Guo X, Zhang Q. Exposure to concentrated ambient fine particulate matter induces vascular endothelial dysfunction via miR-21. Int J Biol Sci. (2017) 13:868–77. doi: 10.7150/ijbs.19868
33. Mahajan A, Go MJ, Zhang W, Below JE, Gaulton KJ, Ferreira T, et al. Genome-wide trans-ancestry meta-analysis provides insight into the genetic architecture of type 2 diabetes susceptibility. Nat Genet. (2014) 46:234–44. doi: 10.1038/ng.2897
34. Li H, Ouyang XP, Jiang T, Zheng XL, He PP, Zhao GJ. MicroRNA-296: a promising target in the pathogenesis of atherosclerosis? Mol Med. (2018) 24:12. doi: 10.1186/s10020-018-0012-y
35. Liang B, Li M, Deng Q, Wang C, Rong J, He S, et al. CircRNA ZNF609 in peripheral blood leukocytes acts as a protective factor and a potential biomarker for coronary artery disease. Ann Transl Med. (2020) 8:741–741. doi: 10.21037/atm-19-4728
36. Tang B, Li X, Ren Y, Wang J, Xu D, Hang Y, et al. MicroRNA-29a regulates lipopolysaccharide (LPS)-induced inflammatory responses in murine macrophages through the Akt1/ NF-κB pathway. Exp Cell Res. (2017) 360:74–80. doi: 10.1016/j.yexcr.2017.08.013
37. Bakshi C, Vijayvergiya R, Dhawan V. Aberrant DNA methylation of M1-macrophage genes in coronary artery disease. Sci Rep. (2019) 9:1429. doi: 10.1038/s41598-018-38040-1
38. Dou H, Kotini A, Liu W, Fidler T, Endo-Umeda K, Sun X, et al. Oxidized phospholipids promote NETosis and arterial thrombosis in LNK(SH2B3) deficiency. Circulation. (2021) 144:1940–54. doi: 10.1161/CIRCULATIONAHA.121.056414
39. Liu Y, Neogi A, Mani A. The role of Wnt signalling in development of coronary artery disease and its risk factors. Open Biol. (2020) 10:200128. doi: 10.1098/rsob.200128
40. Lupu IE, De Val S, Smart N. Coronary vessel formation in development and disease: mechanisms and insights for therapy. Nat Rev Cardiol. (2020) 17:790–806. doi: 10.1038/s41569-020-0400-1
41. Molina C, Jacquet E, Ponien P. Identification of optimal reference genes for transcriptomic analyses in normal and diseased human heart. Cardiovasc Res. (2018) 114:247–58. doi: 10.1093/cvr/cvx182
42. Todur SP, Ashavaid TF. Association of Sp1 tandem repeat polymorphism of ALOX5 with coronary artery disease in Indian subjects. Clin Transl Sci. (2012) 5:408–11. doi: 10.1111/j.1752-8062.2011.00396.x
43. Joiner DM, Ke J, Zhong Z, Xu HE, Williams BO. LRP5 And LRP6 in development and disease. Trends Endocrinol Metab. (2013) 24:31–9. doi: 10.1016/j.tem.2012.10.003
44. Foer D, Zhu M, Cardone RL, Simpson C, Sullivan R, Nemiroff S, et al. Impact of gain-of-function mutations in the low-density lipoprotein receptor-related protein 5 (LRP5) on glucose and lipid homeostasis. Osteoporos Int. (2017) 28:2011–7. doi: 10.1007/s00198-017-3977-4
45. Albanese I, Khan K, Barratt B, Al-Kindi H, Schwertani A. Atherosclerotic calcification: Wnt is the hint. J Am Heart Assoc. (2018) 7:e007356. doi: 10.1161/JAHA.117.007356
46. Schober A, Nazari-Jahantigh M, Wei Y, Bidzhekov K, Gremse F, Grommes J, et al. MicroRNA-126-5p promotes endothelial proliferation and limits atherosclerosis by suppressing Dlk1. Nat Med. (2014) 20:368–76. doi: 10.1038/nm.3487
47. Karacorlu OF, Cetin M, Yumrutas O, Bozgeyik I, Dumlupinar E, Bagis H. Circulating miR-196a-5p miR-373-3p and miR-375: novel candidate biomarkers for diagnosis of acute coronary syndrome. Meta Gene. (2018) 17:1–8. doi: 10.1016/j.mgene.2018.03.013
48. Ghigo A, Li M. Phosphoinositide 3-kinase: friend and foe in cardiovascular disease. Front Pharmacol. (2015) 6:169. doi: 10.3389/fphar.2015.00169
49. Bian WK, Jiang XX, Wang ZC, Zhu YR, Zhang HS, Li XB, et al. Comprehensive analysis of the ceRNA network in coronary artery disease. Sci Rep. (2021) 11:24279. doi: 10.1038/s41598-021-03688-9
50. Liu J, Xu P, Liu D, Wang R, Cui S, Zhang Q, et al. TCM Regulates PI3K/Akt signal pathway to intervene atherosclerotic cardiovascular disease. Evid Based Complement Alternat Med. (2021) 2021:4854755. doi: 10.1155/2021/4854755
51. Ghigo A, Laffargue M, Li M, Hirsch E. PI3K and calcium signaling in cardiovascular disease. Circ Res. (2017) 121:282–92. doi: 10.1161/CIRCRESAHA.117.310183
Keywords: coronary heart disease, hsa_circ_0001445, circRNA-miRNA-mRNA network, diagnostic markers, bioinformatics analysis
Citation: Dinh P, Peng J, Tran T, Wu D, Tran C, Dinh T and Pan S (2023) Identification of hsa_circ_0001445 of a novel circRNA-miRNA-mRNA regulatory network as potential biomarker for coronary heart disease. Front. Cardiovasc. Med. 10:1104223. doi: 10.3389/fcvm.2023.1104223
Received: 21 November 2022; Accepted: 22 February 2023;
Published: 14 March 2023.
Edited by:
Qingchun Zeng, Southern Medical University, ChinaReviewed by:
Jingyi Ren, China-Japan Friendship Hospital, China© 2023 Dinh, Peng, Tran, Feng, Tran, Dinh and Pan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: ShangLing Pan c2xwYW5AZ3htdS5lZHUuY24=
Specialty Section: This article was submitted to Coronary Artery Disease, a section of the journal Frontiers in Cardiovascular Medicine
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.