
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cardiovasc. Med., 25 July 2022
Sec. Atherosclerosis and Vascular Medicine
Volume 9 - 2022 | https://doi.org/10.3389/fcvm.2022.971491
This article is part of the Research TopicLipids and Inflammation in Health and Disease, Volume IIView all 11 articles
 Hong-Xuan Yang1
Hong-Xuan Yang1 Qiu-Ping Liu1
Qiu-Ping Liu1 Yan-Xi Zhou2,3Yu-Ying Chen1Pei An1Yi-Zhuo Xing1Lei Zhang4*Min Jia5*
Yan-Xi Zhou2,3Yu-Ying Chen1Pei An1Yi-Zhuo Xing1Lei Zhang4*Min Jia5* Hong Zhang1*
Hong Zhang1*Forsythiasides are a kind of phenylethanol glycosides existing in Forsythia suspensa (Thunb.) Vahl, which possesses extensive pharmacological activities. According to the different groups connected to the nucleus, forsythiasides can be divided into A-K. In recent years, numerous investigations have been carried out on forsythiasides A, B, C, D, E, and I, which have the effects of cardiovascular protection, anti-inflammation, anti-oxidation, neuroprotection, et al. Mechanistically, forsythiasides regulate toll-like receptor 4 (TLR4)/myeloid differentiation factor 88 (MyD88)/nuclear factor kappaB (NF-κB), nuclear factor-erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) and other signaling pathways, as well as the expression of related cytokines and kinases. Further exploration and development may unearth more treatment potential of forsythiasides and provide more evidence for their clinical applications. In summary, forsythiasides have high development and application value.
Forsythiae Fructus (Chinese name连翘) is the dried fruit of Forsythia suspensa (Thunb.) Vahl, a medicinal plant (Oleaceae) widely distributed in Shanxi, Henan, Shaanxi, and other provinces in China. It is commonly used not only in the clinical practice of traditional Chinese medicine (TCM), but also as an important raw material of many Chinese patent drugs due to its traditional efficacies of clearing heat and detoxification, detumescence and dispersing knot, dispelling wind, and clearing heat. Forsythiae Fructus possesses a variety of pharmacological activities, such as anti-inflammation, antioxidation, anti-virus, and anti-bacteria. Numerous compounds are isolated from F. suspensa, including phenylethanol glycosides, lignans, flavonoids, terpeniods, volatile oils, etc. (1–3). Forsythiasides are the main components of phenylethanol glycosides which have the highest content in F. suspensa.
As the important ingredients for Forsythiae Fructus to exert various pharmacological activities, forsythiasides A-K have been isolated from F. suspensa (4–6). Forsythiasides is a kind of glycosides formed by phenylethanol and sugar and have the stem nucleus as shown in Table 1. The glucose can connect with aglycone, rhamnose, xylose, apinose, and caffeoyl. The structures of forsythiasides A-K are displayed in Table 1, and the structures of forsythoside A and B are shown in Figure 1.

Table 1. The chemical structures of forsythiasides.
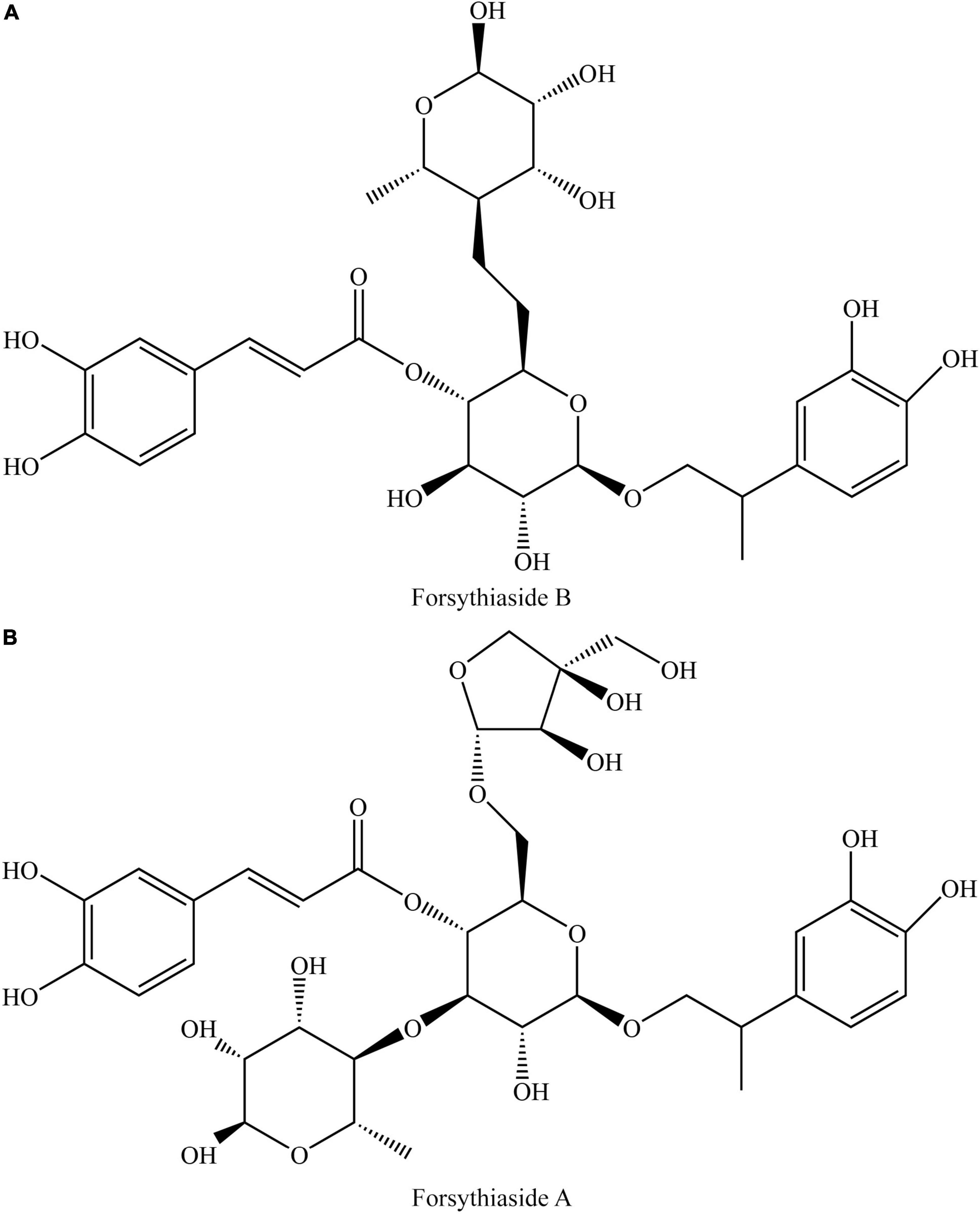
Figure 1. Structures of forsythiaside A (A) and forsythiaside B (B).
Forsythiaside A and forsythiaside B can reduce cardiovascular disease damage and exert cardiovascular protection due to their extensive anti-inflammatory and antioxidant effects. Intravenous injection of 5–20 mg/kg forsythoside B dose-dependently reduced polymorph nuclear leukocyte (PMN) infiltration and myeloperoxidase (MPO) activity in a rat model of myocardial ischemia-reperfusion injury (7). The former releases inflammatory factors to damage cardiomyocytes after being activated by ischemic injury, while the latter is considered to be related to the occurrence of cardiovascular disease (8, 9). High-mobility group box 1 (HMGB1) is an inflammatory mediator released by necrotic cells or activated innate immune cells and can activate the NF-κB signaling pathway (10). Forsythoside B could attenuate the expression of HMGB1 and NF-κB in myocardial tissue, decrease the levels of troponin-T, TNF-α, and IL-6 in serum, and reduce the severity of myocardial injury (7). After 40 mg/kg of forsythiaside A was injected intraperitoneally into heart failure mice, the protein expression of NF-κB was decreased, and the levels of inflammatory factors TNF-α, IL-6, and IL-1β in serum were reduced, displaying the cardioprotective effect of forsythiaside A (11).
Nuclear factor-erythroid 2-related factor 2 (Nrf2) is a cytoprotective factor that can regulate the expression of genes encoding antioxidant, anti-inflammatory, and other related proteins. It could bind to antioxidant response elements (AREs) to initiate the transcription of downstream antioxidant genes (12). Forsythiaside A alleviated oxidative stress in mice with heart failure by activating the Nrf2/heme oxygenase-1 (HO-1) signaling pathway (11). Forsythiaside B reduced the content of MDA in the serum of myocardial ischemia-reperfusion rats, and reverse the decrease of SOD and GPx activities caused by myocardial ischemia (7).
Besides, forsythoside A showed the vasoprotective effect through relaxing the isolated rat aorta. Specifically, forsythiaside A dose-dependently inhibited norepinephrine-induced vasoconstriction by reducing the influx of extracellular calcium ions caused by norepinephrine (13). When low-density lipoprotein is oxidized and accumulated on the arterial wall for a long time, it will cause arteriosclerosis. Forsythoside B could reverse the cytotoxicity elicited by oxidized low-density lipoprotein, inhibit the in vitro oxidation of low-density lipoprotein to play a cytoprotective role (14).
Inflammation is a defense mechanism that occurs when tissues are stimulated by some damages such as trauma, infection by pathogens including bacteria, viruses, parasites, and other inflammatory agents. Inflammatory mediators released from activated mast cells, neutrophils, and macrophages serve as modulator to promote or inhibit the inflammation process (15–17). TLR4/NF-κB is a vital anti-inflammatory pathway, which has attracted much attention in numerous studies on the anti-inflammatory activity of forsythiasides.
Toll-like receptors are a family of pattern-recognition receptors mostly expressed on the surface of cells involved in innate immunity. As a Type I integral membrane protein, TLRs consist of two major parts: an ectodomain of leucine-rich repeats and a cytoplasmic domain of Toll/IL-1R homology domain (18). Among the ten different TLRs discovered in humans, TLR4 is well-known for its characteristic to detect lipopolysaccharide (LPS). LPS non-covalently associates with TLR4 to form an activated heterodimer (LPS/TLR4/myeloid differentiation-2) complex, assisted by membrane cluster differentiation-14 coreceptor (19). Then the complex dimerization recruits MyD88-adaptor-like protein and MyD88, which allows activation of several IL-1 receptor-associated kinases (IRAKs). These IRAKs lead to the ubiquitination of TNF-α receptor-associated factor 6 (TRAF-6), an adaptor molecule activating the transforming growth factor beta-activated kinase 1 (TAK1). Then TAK1 phosphorylates several kinases, resulting in NF-κB being released with other elements (20). However, unlike the above-mentioned TLR4/MyD88/NF-κB pathway, TLR4 can stimulate the production of type I interferons (IFN) as a result of acting on related proteins and phosphorylation of interferon regulatory factor 3 (IRF3) (20).
In dozens of studies on the anti-inflammatory effects of forsythosides A and B, LPS has been widely used to establish inflammation models. Forsythiaside A could significantly reverse the increase of TLR4 and NF-κB protein expression induced by LPS (21), which was also observed in the inflammation of PC12 cells induced by hypoxia/reoxygenation (22). Not only that, forsythosides A and B also reduced the expression of NF-κB through other pathways such as JNK-interacting protein (JIP)/c-Jun N-terminal kinase (JNK)/NF-κB and Nrf2/HO-1/NF-κB, thereby affecting the process of inflammation (23, 24). It is worth mentioning that although Nrf2 and NF-κB are the two key transcription factors regulating cellular response to oxidative stress and inflammation, respectively, the absence of Nrf2 can exacerbate NF-κB activity, leading to the increase of cytokine production (25). Both forsythiasides A and B could inhibit the expression of NF-κB by activating the Nrf2/HO-1 pathway, thereby preventing LPS-induced inflammation in BV2 microglia cells and RAW 264.7 macrophages as well as ovalbumin (OVA)-induced allergic airway inflammation in mice (24, 26–28). As a result, inflammatory factors such as TNF-α and ILs were also regulated in the inflammation model treated by forsythiasides. Forsythiasides could protect against inflammation via the NF-κB pathway by regulating TLR4 and Nrf2. Figure 2 shows how forsythiasides act on the TLR4/MyD88/NF-κB pathway to exert the anti-inflammatory effects. The anti-inflammation properties of forsythiasides A and B are presented in Table 2.
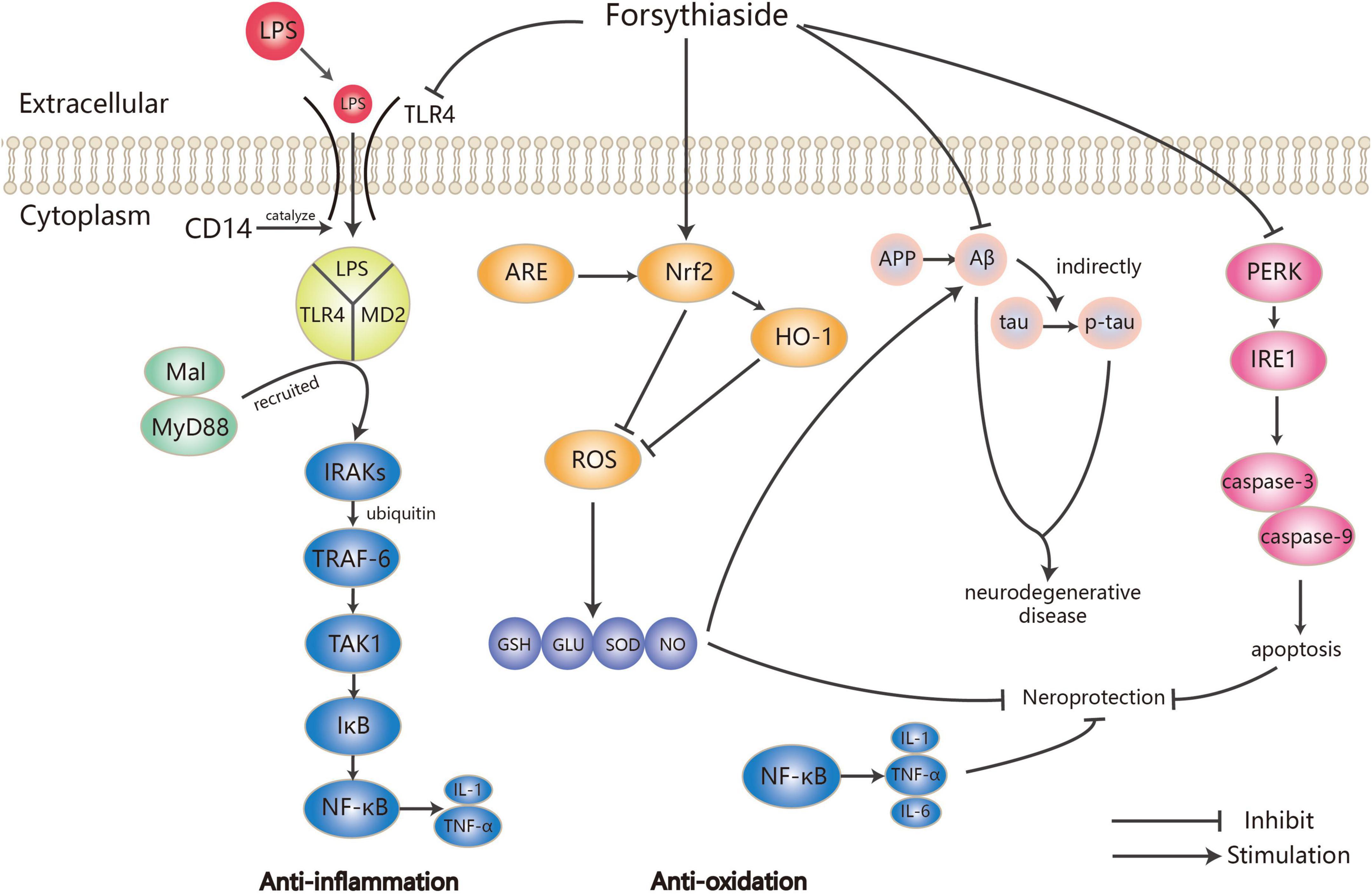
Figure 2. Mechanism of anti-inflammation and anti-oxidation by forsythiasides.
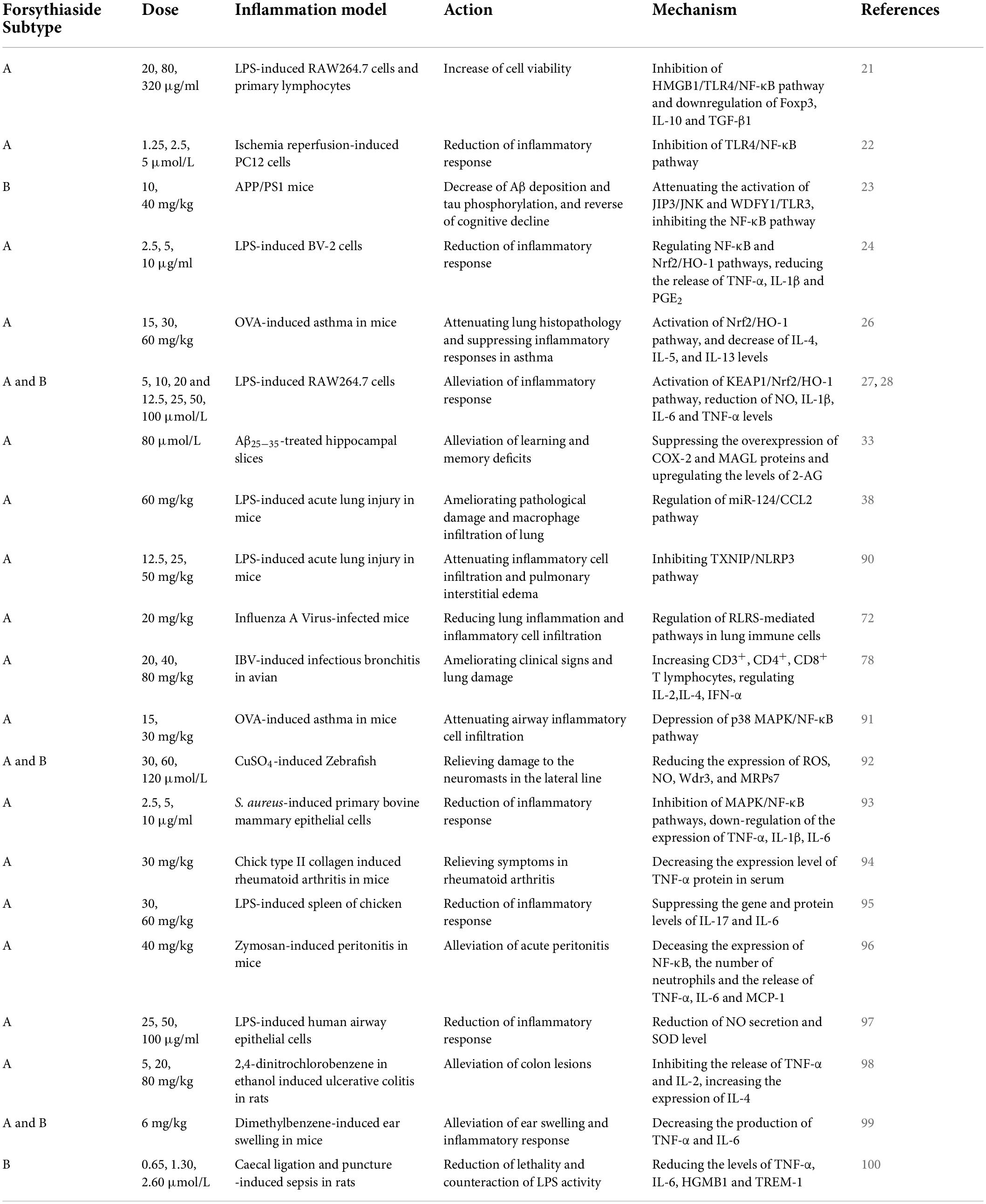
Table 2. Anti-inflammatory effects of forsythiasides.
Alzheimer’s disease (AD) is an age-related neurodegenerative disease, whose pathological features are the neurofibrillary tangles formed by the deposition of amyloid-β (Aβ), hyperphosphorylated tau protein, and neuron loss (29). Among them, Aβ is a 38-43 amino acid polypeptide processed from amyloid precursor protein (APP), which plays an important role in causing the imbalance of synaptic homeostasis and clearance dysfunction of lysosomal (30, 31). Recently, deposition was reported to be promoted by interferon-induced transmembrane protein 3, a γ-secretase modulatory protein induced by inflammatory cytokines.
The neuroprotective effect of forsythiasides is related to their anti-neuroinflammatory activity. Specifically, they could significantly down-regulate inflammatory factors such as interleukin and tumor necrosis factor. The levels of inflammatory factors TNF-α, IL-1β, and IL-6 in the brain tissue of APP/PS1 mice and SAMP8 mice were higher than those of wild-type mice. Interestingly, forsythiasides could reverse these enhanced inflammatory factors. Furthermore, forsythiaside B inhibited NF-κB to exert anti-neuroinflammatory effect by insulting the activation of JIP-3/JNK and reducing the expression of WDFY1/TLR3 (32). Besides, forsythiaside A inhibited the cannabinoid receptor 1 (CB1R)-dependent NF-κB signaling pathway to reduce the secretion of TNF-α and PEG2 in organotypic hippocampal slices of mice (33).
In addition, forsythiasides increased the content of acetylcholine in Aβ-induced AD mouse, which is one of the important neurotransmitters in the central cholinergic system, indicating that forsythiasides have the potential to improve the function of acetylcholine (34).
Acute lung injury (ALI) is one of the earliest and highest morbidity complications after severe trauma or infection. Up-regulation of inflammatory factors serves as a sign of direct response and the development of sustained cell damage (35). A large number of studies have shown that forsythiasides A and B have protective effects on LPS-induced ALI. After the ALI mouse model was treated with forsythiasides, the pathological damage of the lung tissue was relieved in varying degrees (36, 37). At the same time, the gene and protein expression of TLR4, MyD88, and NF-κB were suppressed (36), and the levels of inflammatory factors such as TNF-α, IL-1β, and IL-6 were down-regulated (37). Furthermore, forsythiaside A could reduce the production of chemotactic protein CCL-2, inhibit the adhesion and migration of monocytes to type II lung epithelial cells, thereby decreasing the pathological symptoms of acute lung injury (38). In addition, forsythiaside A could reduce the inflammatory injury of H9N2 avian influenza virus-induced ALI mice by inhibiting MyD88 and NF-κB signaling pathways to down-regulate inflammatory factors such as TNF-α, IL-6, IL-1β (39, 40). Moreover, in mice with cigarette smoke-induced chronic obstructive pulmonary disease, forsythoside A not only decreased inflammatory cytokines and NO production by inhibiting NF-κB signaling pathway, but also increased glutathione/glutathione ratio of peptide disulfides that prevent lung damage (41).
Nephrotic syndrome (NS) is a common type of glomerular disease in children and adults and has many complications resulting in difficult prognosis. Forsythiaside A significantly reduced the levels of urine protein, blood creatinine and urea nitrogen in a nephropathy rat model induced by adriamycin, and decreased the number of apoptotic cells in the kidney tissue (42). Forsythiaside A also dose-dependently depressed the expression of NF-κB p65/Macrophage inflammatory protein-2 in the renal tissue and the levels of inflammatory factors such as IL-6, IL-1β, and TNF-α in serum, improving survival in rats with doxorubicin-induced nephropathy (42).
Diabetic nephropathy (DN) is a common cause of mortality in diabetic patients, and is generally caused by persistent high glucose (HG). Podocytes play an important role in maintaining glomerular structure and filtration. Due to the limited division ability, the damage and reduction of podocytes are the key factors to accelerate the progression of DN. It was found that forsythiaside A inhibited matrix metalloproteinase-12, down-regulated extracellular regulated protein kinases (ERK)/p38 mitogen-activated protein kinase (MAPK) signaling pathway, reduced the expression levels of p-ERK, p-p38, and p-JNP in cells, and inactivated MAPK signaling to alleviate the oxidative stress and inflammation caused by HG (43).
Galactosamine (GalN) is a hepatotoxic substance that can inhibit the synthesis of RNA and protein in liver cells, causing diffuse liver necrosis and inflammation. In a mouse model of acute liver injury induced by LPS/GalN, forsythiaside A inhibited NF-κB activation and reduced TNF-α levels in serum, protecting against liver damage (44).
Osteoclasts, one of the components of bone tissue, have the function of bone resorption. They cooperate with osteoblasts to play a key role in the development and formation of bone (45). Forsythiaside A suppressed LPS-induced skull osteolysis in mice by decreasing the differentiation and formation of osteoclasts to restrain bone resorption. Besides, forsythiaside A also inhibited osteoclast differentiation and reduce ovariectomy-elicited bone loss in mice (46).
Nuclear factor-erythroid 2-related factor 2 plays a vital role in transcriptional activation of genes driven by the antioxidant responsive element (ARE), which mediates transcriptional regulation of phase II detoxification enzyme and antioxidant proteins to clear reactive oxygen species (ROS) (12). HO-1, one of Nrf2-target genes, exerts protection against oxidative injury and modulation of inflammation as well as contribution to angiogenesis (47). In the LPS-induced RAW 264.7 macrophage model, oxidative stress was observed (28). Forsythiaside A inhibited the activation of phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) pathway. At the same time, it activated the Nrf2/HO-1 pathway and reduced the level of ROS. Forsythoside A also depressed the production of inflammatory mediators NO and PGE2, and decreased the expression of pro-inflammatory cytokines TNF-α and IL-1β (28). Meanwhile, the regulatory effect of forsythiaside A on the Nrf2/HO-1 pathway was observed in the OVA-induced mouse asthma model (26). Figure 1 displays how forsythiasides exerts the anti-oxidation effects by regulation of the Nrf2/HO-1 pathway.
Free radicals are the product of the body’s oxidation process, and the appropriate number of free radicals helps maintain the body’s normal physiological functions. The body can maintain the free radical level in a stable range by scavenging the excessive free radicals. Too high concentration of free radicals in the body acts on lipids to undergo peroxidation. The resulting peroxide product MDA denatures cross-link proteins, damages DNA, inactivates enzymes and hormones to damage cells, and finally accelerates aging (48). Forsythiaside A exerted neuroprotective and hepatoprotective effects by clearing MDA (44, 49). Moreover, it was found that forsythiaside A had a dose-dependent anti-oxidant effect on scavenging DPPH free radicals, hydroxyl free radicals, and superoxide anion free radicals, etc. (50–52).
Superoxide dismutase (SOD) is a free radical scavenger widely present in aerobic metabolizing cells, which plays a key role in the body’s oxidation and antioxidant balance. Studies have shown that when inflammation occurs, the level of ROS in cells increases significantly (28). Forsythiaside A could strengthen the body’s antioxidant capacity, and relieve oxidative stress caused by inflammation or ischemic damage through enhancing the activity of cell SOD and increasing the content of glutathione (GSH), glutathione peroxidase (GSH-Px) and catalase (53, 54). The mechanism of action was related to regulation of the PI3K/AKT/Nrf2/HO-1 signaling pathway (28, 55), indicating that forsythiaside A has a certain potential in alleviating inflammation and oxidative damage.
There is certain evidence that excessive oxidative stress factors can produce toxicity to nerve cells, which is related to degenerative memory impairment in AD patients. SOD and GSH-Px are endogenous antioxidant enzymes, which reflect the body’s ability to scavenge free radicals and play a key role in the antioxidant capacity of brain cells. Forsythoside A exerted neuroprotective effects by activating Nrf2 and endoplasmic reticulum stress pathway to reduce cerebral ischemic damage. Forsythiaside A increased the activities of SOD and GSH-Px in the Aβ-induced aging mouse model, and reduced the levels of peroxidation product MDA and important oxygen free radicals NO in vivo and in vitro, alleviating learning and memory deficits in aging mice (49). Forsythiaside A also had a certain effect on delaying the body’s aging by increase of the antioxidant capacity.
Cell protective enzymes such as SOD and GPX can prevent excessive levels of active oxygen to maintain the body’s stability. Forsythiaside B had strong antioxidant and free radical scavenging activities. It activated the expression of Nrf2, regulated the level of nuclear transcription factors of cell protective enzymes, and induced the protective ability of phase II cells to exert skin protection (56). Transient receptor potential vanilloid 3 (TRPV3) channel plays an important role in skin physiology, which is associated with genetic Olmsted syndrome characterized by palmoplantar keratoderma and severe pruritus. Forsythiaside B had the therapeutic potential for the treatment of chronic pruritus, skin allergies or inflammation-related skin diseases due to its specific inhibition of overactive TRPV3. It suppressed channel currents activated by TRPV3 agonists in a dose-dependent manner and significantly attenuated acute and chronic pruritus in a dry skin mouse model (57). Another report showed that forsythiaside B had a protective effect on free radical-induced functional endothelial injury (58).
In the acute liver injury mouse model induced by LPS/GalN, forsythiaside A significantly reduced the content of MDA and attenuated the pathological injury of the mouse liver by activation of the Nrf2/HO-1 signaling pathway, indicating its liver protection effect related to anti-oxidation (44).
Forsythiasides A, B, and I show a widely anti-bacterial activity in many investigations. For instance, enrofloxacin, one of the most used fluoroquinolones, has good therapeutic effect on many pathogens such as Escherichia coli, Pseudomonas and Aeromonas hydrophila. Owing to the widespread use of quinolones, the bacterial resistance has become more and more prominent (59). Studies have found that forsythiaside A could down-regulate the expression of key genes in the differentiation family of drug-resistant nodular cells, and significantly inhibit enrofloxacin resistance to A. hydrophila (60, 61). Forsythia suspensa affected the growth of drug-resistant bacteria of A. hydrophila, presumably mainly through amino acid metabolism, glycolysis, carbon, and nitrogen metabolism, as well as stress-related ABC transport and chemotaxis pathways. Bacterial drug resistance is closely related to its efflux pump system, which can be reduced through inhibiting the activity or reducing the expression for efflux pump.
In addition, forsythiaside A has an obvious antibacterial activity against E. coli, Staphylococcus aureus, and Streptococcus pneumoniae. The K-B paper disk method to measure the diameter of the inhibition zone showed its successive decrease effect on the three bacteria (62). In vivo and in vitro experiments suggested that forsythiaside A had certain inhibitory effect on S. aureus, Streptococcus lactis, and Streptococcus agalactiae (63). Similarly, forsythiaside B was also found to have antibacterial activity against Proteus mirabilis and S. aureus (64). There are also many reports that forsythiaside A had strong inhibitory effect on E. coli, Pseudomonas aeruginosa, and Bacillus subtilis [32](65, 66). Moreover, forsythoside B displayed strong antibacterial activity against multi-drug resistant S. aureus (67). Although forsythiasides have extensive antibacterial effects, the related mechanism needs to be further studied. The antibacterial properties of forsythiasides are summarized in Table 3.

Table 3. Antibacterial effects of forsythiasides.
As a commonly used Chinese medicine, F. suspensa has strong antiviral activity. Studies have exhibited that forsythiaside has inhibitory effect on a variety of viruses. Forsythiaside A could directly kill chicken infectious bronchitis viruses in vitro and inhibit the infection of infectious bronchitis viruses in a dose-dependent manner, but no obvious effect was observed in the infected cells. When a large dose of forsythiaside A was used to pretreat chicken embryo kidney cells, the infectivity of infectious bronchitis virus was significantly inhibited (68). The mechanism was probably related to the induction of IFN-α by forsythiaside A, which up-regulated the relevant factors of the janus kinase–signal transducer and activator of transcription signaling pathway (68). Further research found that forsythiaside A significantly inhibited the replication of the viruses in the cells, and enhance the expression of mRNA related to receptors such as melanoma differentiation-associated gene-5 laboratory of genetics and physiology 2, nod-like receptor family caspase activation and recruitment domain containing 5 and antiviral proteins such as IRF7, IFN-α, and IFN-β (69). The above evidence shows that forsythiaside A has the effect of anti-infectious bronchitis.
Matrix protein 1 (M1 protein), the most abundant one in viruses, plays an important role in maintaining the virus structure and the process of virus replication assembly, and germination. Forsythiaside A could cause slow or abnormal germination of influenza A viruses and the mechanism may relate to decrease in the expression of M1 protein and interfere with the germination process of newly formed viruses (70). Forsythiaside A alleviated the symptoms of weight loss and lung tissue damage caused by influenza virus infection in mice by attenuating the mRNA expression of TLR7, MyD88, IRAK4, TRAF6 in TLR7 signaling pathway and NF-κB p65, reducing the gene and protein expression of retinoicacidinduciblegene-1, mitochondrial antiviral signaling protein and NF-κB, and inhibiting the replication of influenza A viruses to control infection (71, 72). In addition, studies have found that forsythosides C and D were two main components against influenza viruses in the anti-influenza capsules of Forsythiae Fructus (73). It can be seen that forsythiasides have great potential in anti-virus.
Regulatory T cells (Tregs), a subgroup of T cells, are important immune cells with independent functions in the body. Tregs not only participate in cellular immunity, but also maintain the body’s immune balance by suppressing effector T cells to avoid excessive immunity. Therefore, Tregs play a huge role in preventing autoimmune diseases, anti-graft rejection, and tumor immunity (74). The cell-specific nuclear transcription factor Foxp3 can characteristically mark Tregs and play a key role in the development and functional maintenance of Tregs (75).
Forsythiaside A was found to possess immunomodulatory effect by regulating Tregs (76). When the body is infected by endotoxin, endotoxemia will result. Bacterial endotoxin can increase the survival and proliferation ability of Tregs, and enhance their immunosuppressive function to inhibit the body’s immune response. Forsythiaside A exerted the immunomodulatory effect by reducing the level of peripheral blood Tregs and the expression of foxp3 transcription factors in endotoxin mice (21). In addition, forsythiaside A inhibited the replication of bovine viral diarrhea viruses in peripheral blood mononuclear cells cultured in vitro. It plays an important role in immune regulation by promoting the transcription of OX40, 4-1BB, and 4-1BBL which increase the regulation of the activation and proliferation of T cells (77). In addition, forsythiaside A significantly enhanced the number of CD3+, CD4+, CD8+ T lymphocytes in the blood of chickens infected with infectious bronchitis viruses (IBV), showing regulatory effect on the immune function of chickens (78).
The hypothalamus is the advanced center of body temperature regulation, and some nerve nuclei play an important role in this process, such as the paraventricular nucleus (PVN) and supraoptic nucleus. Furthermore, there are abundant temperature-sensing neurons in the periphery such as the dorsal root ganglia (DRG), which also exert a key role in thermoregulation. The temperature-sensitive protein TRPA1 is abundantly expressed in the body temperature regulation center (79). Forsythiaside A effectively reduced the body temperature of fever mice induced by subcutaneous injection of yeast suspension through increasing the expression of TRPA1 in the PVN, supraoptic nucleus and DRG (80). Not only that, forsythiaside A also inhibited the secretion of inflammatory mediators such as PGE2 and IL-8, and mitigated symptoms of fever in mice injected with yeast suspension subcutaneously. The mechanism of action may be related to inhibiting the expression of TRPV1 and reducing calcium influx and MAPK phosphorylation (81). Forsythiaside I, the main component of Qingqiao, was also found to possess the function of clearing heat and removing toxicity (82).
As active ingredients from a multifunctional medicinal plant, forsythiasides display an anticancer activity to a certain degree. Forsythia Fructus could prolong the survival time of melanoma mice, whose components forsythiasides A, E, and I significantly inhibited the cell viability of B16-F10 cells (83). In addition, forsythoside B suppressed the proliferative activity of cervical cancer cells by blocking the expression of NF-κB and up-regulating p21 binding to the cyclin E/CDK2 complex (84).
Besides, forsythiaside A significantly improved the survival rate of rats with cerebral ischemia and reduced neurological deficits by inhibiting neuronal apoptosis and attenuating the expression of caspase-3 and caspase-9 (85). Forsythiaside A obviously increased hair density and thickness in mice with androgenetic alopecia by 50 and 30%, respectively, and decreased the expression of caspase-9 and caspase-3 in the skin by 40 and 53%, respectively. It also inhibited DHT-induced apoptosis of human hair dermal papilla cells and human keratinocytes. Forsythiaside A is a natural product with the potential to treat androgenetic alopecia due to its protective effect on hair loss by inhibiting apoptosis and delaying hair cells entering the degenerative phase (86). Forsythiaside A attenuated APAP-induced hepatocyte degeneration and necrosis by inhibiting the PI3K/AKT pathway associated with apoptosis and reversing the abnormal expression of caspase-3, caspase-8, caspase-9, bax, and bcl-2 (87).
As one of commonly used Chinese medicines, Forsythiae Fructus has extensive pharmacological effects, such as anti-inflammatory, clearing heat, removing toxicity, antibacterial, and antioxidant. Forsythiasides are abundant in F. suspensa. This article summarized the pharmacological activities of forsythiasides A-K (Table 4), which have a similar chemical structure. However, most of the current investigations focus on forsythiasides A and B. NF-κB is a DNA-binding transcription factor existing in eukaryotic cells, which participates in normal physiological processes such as immune and inflammatory response, and is seen as the convergence point of multiple signal pathways. The NF-κB immune signal pathway is closely related to the biological processes of development, proliferation, differentiation, and apoptosis in immune cells, and plays an important role in the regulation of inflammatory cytokine gene expression. In addition, studies have shown that the NF-κB signaling pathway is closely related to the occurrence of cancer (88). Forsythiasides exert anti-inflammatory, anti-tumor, neuroprotective and lung damage protective effects by inhibiting the activation of NF-κB signaling pathway. Furthermore, forsythiasides can also perform the pharmacological activity through activating the Nrf2 signaling pathway and down-regulated the expression of HMGB1 and TLR4. Many documents reported that the strong neuroprotective effect of forsythiasides is related to reduction of Aβ deposition, alleviation of inflammation and oxidative stress of nerve cells, and decrease of caspase-3 activation to inhibit cell apoptosis. Forsythiasides have potential to treat AD by reversing nerve damage and memory dysfunction.
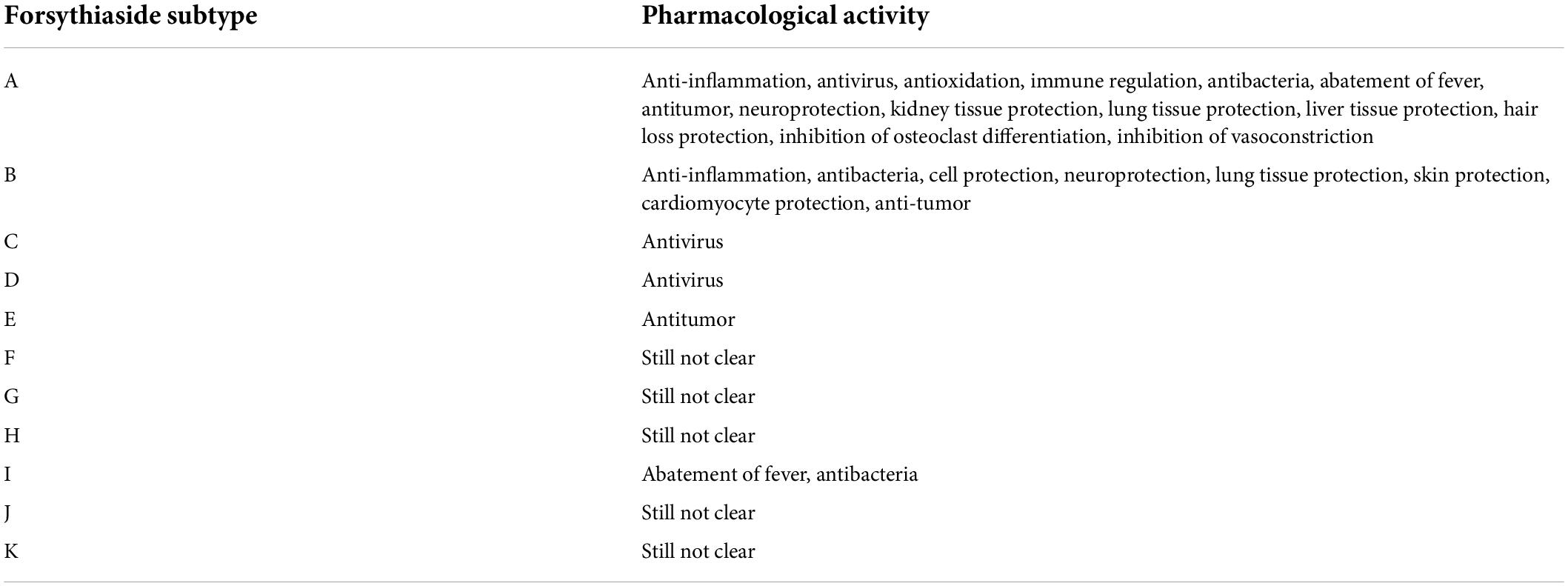
Table 4. Pharmacological activities of forsythiasides.
Although forsythiasides possess many pharmacological activities, there exist some limitations. Forsythiasides A and B show strong antibacterial activity and the ability to scavenge free radicals in vitro. In addition to normal bacteria, forsythiasides also have inhibitory effect on drug-resistant bacteria. However, the current investigations on antibacterial and free radical scavenging effects of forsythiasides are mostly concentrated on in vitro experiments, failing to further validate these in vivo. Forsythiasides interfere with the replication process of the virus, exhibiting the evident antiviral activity. However, the current literature only reported their inhibitory effects on chicken infectious bronchitis viruses and influenza A viruses. In addition, forsythiasides A and B were considered to be closely related to the pseudo-allergic reaction caused by Shuanghuanglian injection (89). However, there are few toxicology investigations reported.
In summary, subsequent studies on forsythiasides should be carried out in view of the above limitations. The NF-κB signaling pathway is closely relevant to a variety of human diseases such as inflammation, tumor, and tissue damage. The other pharmacological effects should be explored further based on NF-κB signaling pathway. Forsythiasides have potential to be an adjuvant drug for the treatment of Alzheimer’s disease owing to the significant neuroprotective effect, so their anti-AD mechanism should be explored further. The antibacterial and antioxidant activities of forsythiasides in vivo should also be investigated and relevant toxicological studies need to be carried out. Finally, it is hoped that the above-mentioned problems will be solved further and more evidence is provided for the clinical applications of forsythiasides.
HZ, MJ, and LZ: conceptualization. H-XY and Q-PL: writing-original. H-XY and HZ: writing-review and editing. HZ, MJ, LZ, Y-XZ, Y-YC, PA, and Y-ZX: verification and recommendation. All authors contributed to the article and approved the submitted version.
This work was supported by funds from the National Natural Science Foundation of China (No. 82174023), National Key R&D Program for Key Research Project of Modernization of Traditional Chinese Medicine (2019YFC1711602 and 2019YFC1711604), Three-year Action Plan for Shanghai TCM Development and Inheritance Program [ZY(2021-2023)- 0401], Clinical Innovation Project of Shenkang Hospital Development Center in Shanghai (SHDC12019X33), and Research on Green Space Life Support System for Extreme Environments Based on Renewable Energy (BX221C010).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Hu J, Ma L, Zhang J, Xu C. Research progress of Forsythia suspensa. Cent South Pharm. (2012) 10:760–4.
2. Xia W, Dong C, Yang C, Chen H. Research progress on chemical constituents and pharmacology of Forsythia suspensa. Mod Chin Med. (2016) 18:1670–4. doi: 10.13313/j.issn.1673-4890.2016.12.031
3. Zhang T, Shi L, Liu W, Zhang M, Yang J, Li F. research progress on chemical constituents and pharmacological effects of Forsythia. J Liaoning U Tradit Chinese Med. (2016) 18:222–4. doi: 10.13194/j.issn.1673-842x.2016.12.067
4. Wang Z, Xia Q, Liu X, Liu W, Huang W, Mei X, et al. Phytochemistry, pharmacology, quality control and future research of Forsythia suspensa (Thunb.) Vahl: a review. J Ethnopharmacol. (2018) 210:318–39. doi: 10.1016/j.jep.2017.08.040
5. Wei Q, Li P, Wu T, Li C, Zhang R. Progress in phenylethanoid glycosides from Forsythia suspensa. Chin J Clin Pharmacol. (2018) 34:2481–5. doi: 10.13699/j.cnki.1001-6821.2018.20.024
6. Yan X, Xiang Z, Wen J, Zhang W, Yang B, Qu Z, et al. Phenylethanoid glycosides from fruits of Forsythia Suspensa. Chin Tradit Herb Drugs. (2016) 47:3362–5.
7. Jiang W, Fu F, Xu B, Tian J, Zhu H, Hou J. Cardioprotection with forsythoside B in rat myocardial ischemia-reperfusion injury: relation to inflammation response. Phytomedicine. (2010) 17:635–9. doi: 10.1016/j.phymed.2009.10.017
8. Piccolo EB, Thorp EB, Sumagin R. Functional implications of neutrophil metabolism during ischemic tissue repair. Curr Opin Pharmacol. (2022) 63:102191. doi: 10.1016/j.coph.2022.102191
9. Maiocchi SL, Ku J, Thai T, Chan E, Rees MD, Thomas SR. Myeloperoxidase: a versatile mediator of endothelial dysfunction and therapeutic target during cardiovascular disease. Pharmacol Ther. (2021) 221:107711. doi: 10.1016/j.pharmthera.2020.107711
10. Andrassy M, Volz HC, Igwe JC, Funke B, Eichberger SN, Kaya Z, et al. High-mobility group box-1 in ischemia-reperfusion injury of the heart. Circulation. (2008) 117:3216–26. doi: 10.1161/circulationaha.108.769331
11. Fu Q, Ma D, Tong Y, Li H, Shen C. Protective effect and possible mechanism of forsythiaside A on pressure overload heart failure in mice. Prog Anat Sci. (2021) 27:65–7. doi: 10.16695/j.cnki.1006-2947.2021.01.017
12. Lee J, Calkins M, Chan K, Kan Y, Johnson J. Identification of the NF-E2-related factor-2-dependent genes conferring protection against oxidative stress in primary cortical astrocytes using oligonucleotide microarray analysis. J Biol Chem. (2003) 278:12029–38. doi: 10.1074/jbc.M211558200
13. Iizuka T, Nagai M. Vasorelaxant effects of forsythiaside from the fruits of Forsythia suspensa. Yakugaku Zasshi. (2005) 125:219–24. doi: 10.1248/yakushi.125.219
14. Martin-Nizard F, Sahpaz S, Furman C, Fruchart J, Duriez P, Bailleul F. Natural phenylpropanoids protect endothelial cells against oxidized LDL-induced cytotoxicity. Planta Med. (2003) 69:207–11. doi: 10.1055/s-2003-38474
15. Metz M, Grimbaldeston M, Nakae S, Piliponsky A, Tsai M, Galli S. Mast cells in the promotion and limitation of chronic inflammation. Immunol Rev. (2007) 217:304–28. doi: 10.1111/j.1600-065X.2007.00520.x
16. Soehnlein O, Steffens S, Hidalgo A, Weber C. Neutrophils as protagonists and targets in chronic inflammation. Nat Rev Immunol. (2017) 17:248–61. doi: 10.1038/nri.2017.10
17. Hamidzadeh K, Christensen S, Dalby E, Chandrasekaran P, Mosser D. Macrophages and the recovery from acute and chronic inflammation. Annu Rev Physiol. (2017) 79:567–92. doi: 10.1146/annurev-physiol-022516-034348
18. Kashani B, Zandi Z, Pourbagheri-Sigaroodi A, Bashash D, Ghaffari S. The role of toll-like receptor 4 (TLR4) in cancer progression: a possible therapeutic target? J Cell Physiol. (2021) 236:4121–37. doi: 10.1002/jcp.30166
19. Jagtap P, Prasad P, Pateria A, Deshmukh S, Gupta SA. Single step in vitro bioassay mimicking TLR4-LPS pathway and the role of MD2 and CD14 coreceptors. Front Immunol. (2020) 11:5. doi: 10.3389/fimmu.2020.00005
20. Brooks D, Barr L, Wiscombe S, McAuley D, Simpson A, Rostron A. Human lipopolysaccharide models provide mechanistic and therapeutic insights into systemic and pulmonary inflammation. Eur Respir J. (2020) 56:1901298. doi: 10.1183/13993003.01298-2019
21. Zeng X, Yuan W, Zhou L, Wang S, Xie Y, Fu Y. Forsythoside A exerts an anti-endotoxin effect by blocking the LPS/TLR4 signaling pathway and inhibiting Tregs in vitro. Int J Mol Med. (2017) 40:243–50. doi: 10.3892/ijmm.2017.2990
22. Ma Z, Zhang D, Sun J, Niu J, Ren Q, Wu Q, et al. Forsythiaside A inhibits inflammation induced by cerebral ischemia through TLR4/NF-κB. Acta U Med Anhui. (2021) 56:730–4. doi: 10.19405/j.cnki.issn1000-1492.2021.05.011
23. Kong F, Jiang X, Wang R, Zhai S, Zhang Y, Wang D. Forsythoside B attenuates memory impairment and neuroinflammation via inhibition on NF-κB signaling in Alzheimer’s disease. J Neuroinflamm. (2020) 17:305. doi: 10.1186/s12974-020-01967-2
24. Wang Y, Zhao H, Lin C, Ren J, Zhang S. Forsythiaside A exhibits anti-inflammatory effects in LPS-stimulated BV2 microglia cells through activation of NRF2/HO-1 signaling pathway. Neurochem Res. (2016) 41:659–65. doi: 10.1007/s11064-015-1731-x
25. Wardyn J, Ponsford A, Sanderson C. Dissecting molecular cross-talk between NRF2 and NF-κB response pathways. Biochem Soc Trans. (2015) 43:621–6. doi: 10.1042/bst20150014
26. Qian J, Ma X, Xun Y, Pan L. Protective effect of forsythiaside A on OVA-induced asthma in mice. Eur J Pharmacol. (2017) 812:250–5. doi: 10.1016/j.ejphar.2017.07.033
27. Wu A, Yang Z, Huang Y, Yuan H, Lin C, Wang T, et al. Natural phenylethanoid glycosides isolated from Callicarpa kwangtungensis suppressed lipopolysaccharide-mediated inflammatory response via activating Keap1/NRF2/HO-1 pathway in RAW 264.7 macrophages cell. J Ethnopharmacol. (2020) 258:112857. doi: 10.1016/j.jep.2020.112857
28. Tang H, Che N, Liu H, Ma X, Li J, Li L. Forsythiaside A suppresses LPS-induced inflammation and oxidative stress by inhibiting PI3K/Akt pathway and activating Nrf2/HO-1 pathway. Immunol J. (2021) 37:390–6. doi: 10.13431/j.cnki.immunol.j.20210054
29. Hyman BT, Phelps CH, Beach TG, Bigio EH, Cairns NJ, Carrillo MC, et al. National institute on aging-Alzheimer’s association guidelines for the neuropathologic assessment of Alzheimer’s disease. Alzheimers Dement. (2012) 8:1–13. doi: 10.1016/j.jalz.2011.10.007
30. Thinakaran G, Koo EH. Amyloid precursor protein trafficking. Proc Funct J Biologic Chem. (2008) 283:29615–9. doi: 10.1074/jbc.R800019200
31. Knopman D, Amieva H, Petersen R, Chételat G, Holtzman D, Hyman B, et al. Alzheimer disease. Nat Rev Dis Primers. (2021) 7:33. doi: 10.1038/s41572-021-00269-y
32. Kong F. Study on the Protective effect of Forsythiaside B on Alzheimer’s Disease and its undering mechanism Master’s thesis. Changchun: Jilin University (2020).
33. Chen L, Yan Y, Chen T, Zhang L, Gao X, Du C, et al. Forsythiaside prevents β-amyloid-induced hippocampal slice injury by upregulating 2-arachidonoylglycerol via cannabinoid receptor 1-dependent NF-κB pathway. Neurochem Int. (2019) 125:57–66. doi: 10.1016/j.neuint.2019.02.008
34. Yan X, Chen T, Zhang L, Du H. Protective effects of Forsythoside A on amyloid beta-induced apoptosis in PC12 cells by downregulating acetylcholinesterase. Eur J Pharmacol. (2017) 810:141–8. doi: 10.1016/j.ejphar.2017.07.009
35. Johnson E, Matthay M. Acute lung injury: epidemiology, pathogenesis, and treatment. J Aerosol Med Pulm Drug Deliv. (2010) 23:243–52. doi: 10.1089/jamp.2009.0775
36. Zhou L, Yang H, Ai Y, Xie Y, Fu Y. Protective effect of forsythiaside A on acute lung injure induced by lipopolysaccharide in mice. Chin J Cell Mol Immunol. (2014) 30:151–4. doi: 10.13423/j.cnki.cjcmi.007031
37. Liu J, Li X, Yan F, Pan Q, Yang C, Wu M, et al. Protective effect of forsythoside B against lipopolysaccharide-induced acute lung injury by attenuating the TLR4/NF-κB pathway. Int Immunopharmacol. (2019) 66:336–46. doi: 10.1016/j.intimp.2018.11.033
38. Lu Z, Liu S, Ou J, Cao H, Shi L, Liu D, et al. Forsythoside A inhibits adhesion and migration of monocytes to type II alveolar epithelial cells in lipopolysaccharide-induced acute lung injury through upregulating miR-124. Toxicol Appl Pharmacol. (2020) 407:115252. doi: 10.1016/j.taap.2020.115252
39. Sheng N, Lu G, Fu Y, Hu G. Effect of forsythoside A on the expression of inflammatory genes in H9N2-AIV Mice. China Anim Health. (2020) 22:49–50.
40. Sheng N, Lu G, Fu Y, Tian X, Hu G. Effect of forsythoside A on Toll-like receptor signal pathway in mice infected with H9N2 Avian influenza. J Beijing U Agric. (2020) 35:94–100. doi: 10.13473/j.cnki.issn.1002-3186.2020.0317
41. Cheng L, Li F, Ma R, Hu X. Forsythiaside inhibits cigarette smoke-induced lung inflammation by activation of NRF2 and inhibition of NF-κB. Int Immunopharmacol. (2015) 28:494–9. doi: 10.1016/j.intimp.2015.07.011
42. Lu C, Zheng S, Liu J. Forsythiaside A alleviates renal damage in adriamycin-induced nephropathy. Front Biosci. (2020) 25:526–35. doi: 10.2741/4818
43. Quan X, Liu H, Ye D, Ding X, Su X. Forsythoside A alleviates high glucose-induced oxidative stress and inflammation in podocytes by inactivating MAPK signaling via MMP12 inhibition. Diabetes Metab Syndr. (2021) 14:1885–95. doi: 10.2147/dmso.s305092
44. Pan C, Zhou G, Chen W, Zhuge L, Jin L, Zheng Y, et al. Protective effect of forsythiaside A on lipopolysaccharide/d-galactosamine-induced liver injury. Int Immunopharmacol. (2015) 26:80–5. doi: 10.1016/j.intimp.2015.03.009
45. Chen H, Wang Y. Function and regulation of the osteoclast in the pathological changes of bone destruction in rheumatoid arthritis. Chin J Osteoporos. (2016) 22:1168–73.
46. Sun X. The Mechanism of Forsythiaside A Inhibiting Osteoclast Differentiation and Its Therapeutic Application Master Thesis. Hangzhou: Zhejiang University (2018).
47. Loboda A, Damulewicz M, Pyza E, Jozkowicz A, Dulak J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: an evolutionarily conserved mechanism. Cell Mol Life Sci. (2016) 73:3221–47. doi: 10.1007/s00018-016-2223-0
48. Liu J, Zhang L. Functions of forsythiaside on the damage of DNA induced by hydroxyl radical. J Shanxi Coll Tradit Chin Med. (2006) 7:23–4.
49. Wang H, Wang L, Liu X, Li C, Xu S, Farooq A. Neuroprotective effects of forsythiaside on learning and memory deficits in senescence-accelerated mouse prone (SAMP8) mice. Pharmacol Biochem Be. (2013) 105:134–41. doi: 10.1016/j.pbb.2012.12.016
50. Delazar A, Sabzevari A, Mojarrab M, Nazemiyeh H, Esnaashari S, Nahar L, et al. Free-radical-scavenging principles from Phlomis caucasica. J Nat Med. (2008) 62:464–6. doi: 10.1007/s11418-008-0255-y
51. Georgiev M, Alipieva K, Orhan I, Abrashev R, Denev P, Angelova M. Antioxidant and cholinesterases inhibitory activities of Verbascum xanthophoeniceum GRISEB. and its phenylethanoid glycosides. Food Chem. (2011) 128:100–5. doi: 10.1016/j.foodchem.2011.02.083
52. Piao X, Wu Q, Tian Y. Antioxidants from Forsythia suspensa. Lishizhen Med Materia Med Res. (2010) 21:1307–9.
53. Zhao Y. Effect of Forsythaside on the Cytokine and Antionxidative Function in Bursa of Fabricius of Chickens. Master Thesis Harbin: Northeast Agricultural University (2013).
54. Li H, Chen J, Zhao Y, Wu Y, Li X, Li Y. Effect of forsythiaside on antioxidation induced by LPS in liver of chickens. China Poultry. (2013) 35:20–2.
55. Ma Z, Zhang D, Sun J, Niu J, Ren Q, Wu Q, et al. Forsythiaside A inhibited cerebral ischemic induced oxidative damage through AKT/NRF2 signaling pathway. Prog Mod Biomed. (2021) 21:214–8. doi: 10.13241/j.cnki.pmb.2021.02.004
56. Sgarbossa A, Dal Bosco M, Pressi G, Cuzzocrea S, Dal Toso R, Menegazzi M. Phenylpropanoid glycosides from plant cell cultures induce heme oxygenase 1 gene expression in a human keratinocyte cell line by affecting the balance of NRF2 and BACH1 transcription factors. Chem Biol Interact. (2012) 199:87–95. doi: 10.1016/j.cbi.2012.06.006
57. Zhang H, Sun X, Qi H, Ma Q, Zhou Q, Wang W, et al. Pharmacological inhibition of the temperature-sensitive and ca-permeable transient receptor potential vanilloid TRPV3 Channel by natural forsythoside b attenuates pruritus and cytotoxicity of keratinocytes. J Pharmacol Exp Ther. (2019) 368:21–31. doi: 10.1124/jpet.118.254045
58. Ismailoglu U, Saracoglu I, Harput U, Sahin-Erdemli I. Effects of phenylpropanoid and iridoid glycosides on free radical-induced impairment of endothelium-dependent relaxation in rat aortic rings. J Ethnopharmacol. (2002) 79:193–7. doi: 10.1016/s0378-8741(01)00377-4
59. Nelson J, Chiller T, Powers J, Angulo F. Fluoroquinolone-resistant Campylobacter species and the withdrawal of fluoroquinolones from use in poultry: a public health success story. Clin Infect Dis. (2007) 44:977–80. doi: 10.1086/512369
60. Ying X, Yang Y. New veterinary antibacterial drug−enrofloxacin. Chin J Vet Drug. (1995) 29:53–6.
61. Dong Y, Feng D, Sun J, Zhang X, Hu K, Yang X. Delaying effect of forsythiaside A Aeromonas hydrophila resistance to enrofloxacin and its exocytosis. J Southern Agric. (2019) 50:187–93.
62. Zhang Y, Han Y, Liang L, Shang C, Xue H. Preparation and antibacterial antioxidant activity of phillyrin, forsythiaside A on n-butanol part extract from qingqiao of Forsythia suspensa. Chin J Exp Tradit Med. (2014) 20:192–6. doi: 10.13422/j.cnki.syfjx.2014210192
63. Wang H, Jiang H, Wu G. Research on the antibacterial effect of forsythiaside in vitro and in vivo. China Feed. (2005) 26–7.
64. Didry N, Seidel V, Dubreuil L, Tillequin F, Bailleul F. Isolation and antibacterial activity of phenylpropanoid derivatives from Ballota nigra. J Ethnopharmacol. (1999) 67:197–202. doi: 10.1016/s0378-8741(99)00019-7
65. Qin Z, Xu J, Zhang L. Studies on the anti-bacteria effect of fructus forsythia and folium forsythia in vitro. Food Eng. (2013) 49–52.
66. Qu H, Zhang Y, Wang Y, Li B, Sun W. Antioxidant and antibacterial activity of two compounds (forsythiaside and forsythin) isolated from Forsythia suspensa. J Pharma Pharmacol. (2008) 60:261–6. doi: 10.1211/jpp.60.2.0016
67. Nazemiyeh H, Rahman M, Gibbons S, Nahar L, Delazar A, Ghahramani M, et al. Assessment of the antibacterial activity of phenylethanoid glycosides from Phlomis lanceolata against multiple-drug-resistant strains of Staphylococcus aureus. J Nat Med. (2008) 62:91–5. doi: 10.1007/s11418-007-0194-z
68. Li H, Wu J, Zhang Z, Ma Y, Liao F, Zhang Y, et al. Forsythoside a inhibits the avian infectious bronchitis virus in cell culture. Phytother Res. (2011) 25:338–42. doi: 10.1002/ptr.3260
69. Zhang T, Liu B, Yang X, Lv A, Zhang Z, Gong P, et al. Effect of forsythoside A on expression of intracellular receptors and antiviral gene in IBV-infected cells. J Beijing U Agric. (2017) 32:37–42. doi: 10.13473/j.cnki.issn.1002-3186.2017.0118
70. Law A, Yang C, Lau A, Chan G. Antiviral effect of forsythoside A from Forsythia suspensa (Thunb.) Vahl fruit against influenza A virus through reduction of viral M1 protein. J Ethnopharmacol. (2017) 24:236–47. doi: 10.1016/j.jep.2017.07.015
71. Deng L, Pang P, Zheng K, Nie J, Xu H, Wu S, et al. Forsythoside A controls influenza A virus infection and improves the prognosis by inhibiting virus replication in mice. Molecules. (2016) 21:524. doi: 10.3390/molecules21050524
72. Zheng X, Fu Y, Shi S, Wu S, Yan Y, Xu L, et al. Effect of forsythiaside A on the RLRs signaling pathway in the lungs of mice infected with the influenza A virus FM1 strain. Molecules. (2019) 24:4219. doi: 10.3390/molecules24234219
73. Zhao W, Shi R, Liu B, Zhang J. Isolation and elucidation of antiviral substances from lianqiao bingduqing capsule on influenza virus. Chin Tradit Pat Med. (2005) 27:81–5.
74. Hall BT. Cells: soldiers and spies–the surveillance and control of effector T cells by regulatory T cells. Clin J Am Soc Nephrol. (2015) 10:2050–64. doi: 10.2215/cjn.06620714
75. Yuan W, Yang H, Fu Y. Research progress of traditional chinese medicine on the immune regulatory function of regulatory T cells. Chin Tradit Pat Med. (2014) 36:1041–4.
76. Zhang X. Effect of forsythiaside A on immune regulation in endotoxemia mice and mechanism of action. Guid J Tradit Chin Med Pharm. (2016) 22:57–60. doi: 10.13862/j.cnki.cn43-1446/r.2016.21.018
77. Song Q, Weng X, Cai D, Zhang W, Wang J. Forsythoside A inhibits BVDV Replication via TRAF2-Dependent CD28-4-1BB Signaling in Bovine PBMCs. PLoS One. (2016) 11:e0162791. doi: 10.1371/journal.pone.0162791
78. Wang X, Li X, Wang X, Chen L, Ning E, Fan Y, et al. Experimental study of forsythoside A on prevention and treatment of avian infectious bronchitis. Res Vet Sci. (2021) 135:523–31. doi: 10.1016/j.rvsc.2020.11.009
79. Ding Z, Gomez T, Werkheiser J, Cowan A, Rawls S. Icilin induces a hyperthermia in rats that is dependent on nitric oxide production and NMDA receptor activation. Eur J Pharmacol. (2008) 578:201–8. doi: 10.1016/j.ejphar.2007.09.030
80. Su H, Wan H, Liu C, Kong X, Lin N. Effect of forsythiaseide A on temperature and expression of TRPA1 in mice with yeast induced pyrexia. Chin J Exp Tradit Med Formulae. (2016) 22:134–8. doi: 10.13422/j.cnki.syfjx.2016010134
81. Liu C, Su H, Wan H, Qin Q, Wu X, Kong X, et al. Forsythoside A exerts antipyretic effect on yeast-induced pyrexia mice via inhibiting transient receptor potential vanilloid 1 function. Int J Biol Sci. (2017) 13:65–75. doi: 10.7150/ijbs.18045
82. Ma L, Gao J, Zhang L. Screening for active components of heat-clearing and detoxifying from green fructus forsythiae. Chem Res Appl. (2019) 31:271–7.
83. Bao J, Ding R, Liang Y, Liu F, Wang K, Jia X, et al. Differences in chemical component and anticancer activity of green and ripe forsythiae fructus. Am J Chin Med. (2017) 45:1513–36. doi: 10.1142/s0192415x17500823
84. Zhang Z, Lv Y, Huang P. Forsythiaside B inhibits the proliferation activity of HeLa cells though transcription factor NF-κB. Chin J Clin Pharmacol Ther. (2020) 25:387–92.
85. Ma T, Shi Y, Wang Y. Forsythiaside A protects against focal cerebral ischemic injury by mediating the activation of the NRF2 and endoplasmic reticulum stress pathways. Mol Med Rep. (2019) 20:1313–20. doi: 10.3892/mmr.2019.10312
86. Shin HS, Park SY, Song HG, Hwang E, Lee DG, Yi TH. The androgenic alopecia protective effects of forsythiaside-A and the molecular regulation in a mouse model. Phytother Res. (2015) 29:870–6. doi: 10.1002/ptr.5324
87. Gong L, Zhou H, Wang C, He L, Guo C, Peng C, et al. Hepatoprotective effect of forsythiaside a against acetaminophen-induced liver injury in zebrafish: coupling network pharmacology with biochemical pharmacology. J Ethnopharmacol. (2021) 271:113890. doi: 10.1016/j.jep.2021.113890
88. Taniguchi K, Karin M. NF-κB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol. (2018) 18:309–24. doi: 10.1038/nri.2017.142
89. Han J, Zhang Y, Pan C, Xian Z, Pan C, Zhao Y, et al. Forsythoside A and Forsythoside B contribute to shuanghuanglian injection-induced pseudoallergic reactions through the RHOA/ROCK Signaling Pathway. Int J Mol Sci. (2019) 20:6266. doi: 10.3390/ijms20246266
90. Liang H, Liu K, Zhuang Z, Sun H, Sun B, Jiao M, et al. Potential of Forsythoside I as a therapeutic approach for acute lung injury: involvement of TXNIP/NLRP3 inflammasome. Mol Immunol. (2021) 134:192–201. doi: 10.1016/j.molimm.2021.03.004
91. Lin X, Li J, Che N, Li L, Li L. Forsythiaside A suppresses airway inflammation of asthma through inhibition of p38 MAPK/NF-kappa B signaling pathway. Chin J Immunol. (2019) 35:2971–4.
92. Gong L, Yu L, Gong X, Wang C, Hu N, Dai X, et al. Exploration of anti-inflammatory mechanism of forsythiaside A and forsythiaside B in CuSO4-induced inflammation in zebrafish by metabolomic and proteomic analyses. J Neuroinflamma. (2020) 17:173. doi: 10.1186/s12974-020-01855-9
93. Zhang J, Zhang Y, Huang H, Zhang H, Lu W, Fu G, et al. Forsythoside A inhibited S. Aureus stimulated inflammatory response in primary bovine mammary epithelial cells. Microb Pathog. (2018) 116:158–63. doi: 10.1016/j.micpath.2018.01.002
94. Cheng G, Lv D, Liu L, Wang B, Wang W, Zhang W, et al. Study on the Inhibitory mechanism of forsythiaside A against rheumatoid arthritis of rats. Animal Husbandry Feed Sci. (2014) 35:1–3. doi: 10.16003/j.cnki.issn1672-5190.2014.05.062
95. Cheng G, Zhang Q, Yue L, Xu L, Zhu D, Liu H, et al. Effect of forsythiaside A on the Levels of IL-17 and IL-6 in spleen of chickens inflamed by endotoxin. Heilongjiang Animal Sci Vet Med. (2017) 149–52. doi: 10.13881/j.cnki.hljxmsy.2017.2096
96. Zhang X, Ding Y, Kang P, Zhang X, Zhang T. Forsythoside A modulates zymosan-induced peritonitis in mice. Molecules. (2018) 23:593. doi: 10.3390/molecules23030593
97. Liu J, Zhang L. Anti-inflammatory activity of Forsythia suspensa extract on human airway epithelial cells inflammation model. Nat Product Res Dev. (2015) 27:1248–53. doi: 10.16333/j.1001-6880.2015.07.023
98. Yu L, Jiang H, Liu H. Effect of forsythiaside A on immune function in rats with ulcerative colitis. Chin J Pathophysiol. (2020) 36:1128–32.
99. Quan Y, Yuan A, Gong X, Peng C, Li Y. Investigation on anti-inflammatory components of Forsythia suspensa. Nat Product Res Dev. (2017) 29:435–8. doi: 10.16333/j.1001-6880.2017.3.013
100. Jiang W, Xu Y, Zhang S, Zhu H, Hou J. Forsythoside B protects against experimental sepsis by modulating inflammatory factors. Phytother Res. (2012) 26:981–7. doi: 10.1002/ptr.3668
Keywords: forsythiasides, Forsythia suspensa, cardiovascular protection, anti-inflammation, anti-oxidation
Citation: Yang H-X, Liu Q-P, Zhou Y-X, Chen Y-Y, An P, Xing Y-Z, Zhang L, Jia M and Zhang H (2022) Forsythiasides: A review of the pharmacological effects. Front. Cardiovasc. Med. 9:971491. doi: 10.3389/fcvm.2022.971491
Received: 17 June 2022; Accepted: 04 July 2022;
Published: 25 July 2022.
Edited by:
Alexander Nikolaevich Orekhov, Institute for Aterosclerosis Research, RussiaCopyright © 2022 Yang, Liu, Zhou, Chen, An, Xing, Zhang, Jia and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lei Zhang, c2VlLWV5ZUAxNjMuY29t; Min Jia, am03LjFAMTYzLmNvbQ==; Hong Zhang, aHF6aGFuZzUxQDEyNi5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.