 Shihui Fu1,2†
Shihui Fu1,2† Jianqiu Hu3†Xiaoping Chen4†Bo Li1†Hongjuan Shun5Juelin Deng1*Yujie Zhang6*
Jianqiu Hu3†Xiaoping Chen4†Bo Li1†Hongjuan Shun5Juelin Deng1*Yujie Zhang6* Yao Yao7,8*
Yao Yao7,8* Yali Zhao4*
Yali Zhao4*- 1Department of Cardiology, Hainan Hospital of Chinese People's Liberation Army General Hospital, Sanya, China
- 2Department of Geriatric Cardiology, Chinese People's Liberation Army General Hospital, Beijing, China
- 3Department of Ultrasound, Hainan Hospital of Chinese People's Liberation Army General Hospital, Sanya, China
- 4Central Laboratory, Hainan Hospital of Chinese People's Liberation Army General Hospital, Sanya, China
- 5Department of Health Medicine, Hainan Hospital of Chinese People's Liberation Army General Hospital, Sanya, China
- 6Department of Epidemiology, School of Public Health, Southern Medical University, Guangzhou, China
- 7Center for the Study of Aging and Human Development and Geriatrics Division, Medical School of Duke University, Durham, NC, United States
- 8Center for Healthy Aging and Development Studies, National School of Development, Peking University, Beijing, China
In the current study, we aimed to determine the association of single nucleotide polymorphism rs189037 in ataxia-telangiectasia mutated (ATM) gene with cardiac structure and human longevity. Based on the China Hainan Centenarian Cohort Study performed in 18 cities and counties of Hainan Province, China, the current study enrolled 547 centenarians, 250 young participants aged 20–45 years, and 250 middle-aged and elderly participants aged 46–90 years. The frequency of TT genotype was significantly higher and that of CC genotype was significantly lower in middle-aged and elderly participants than in young (P = 0.012) and centenarian (P = 0.041) participants. There were no significant differences in the genotype and allele frequencies of SNP rs189037 between young and centenarian participants. Compared with CT genotype, TT genotype was positively and significantly associated with interventricular septum thickness (IVST) and left ventricular posterior wall thickness (LVPWT) in centenarian (IVST: P = 0.049; LVPWT: P = 0.047) and middle-aged and elderly (IVST: P = 0.008; LVPWT: P = 0.004) participants. Compared with CC genotype, TT genotype was positively and significantly associated with LVPWT in centenarian (P = 0.030) and middle-aged and elderly (P = 0.013) participants. Compared with CC genotype, CT genotype was negatively and significantly associated with left ventricular end-diastolic diameter (LVEDD) in centenarian (P = 0.011) and middle-aged and elderly (P = 0.040) participants. The current study demonstrated that mutant rs189037 in the ATM gene was more commonly identified in middle-aged and elderly participants than in young and centenarian participants, was significantly associated with increased left ventricular wall thickness and volume, and could induce left ventricular eccentric hypertrophy and shorten human lifespan. Therefore, rs189037 without mutation might be an indicator of youth health and successful aging, whereas mutant rs189037 might hinder human longevity.
Introduction
The average lifespan of a human ranges from ~65 to 85 years, and the frequency of centenarians is 1 in 10,000 worldwide (1). In studies conducted on twins, human longevity has been indicated to be moderately heritable, and different genetic mutations account for ~25% of human longevity (2). Although studies have been performed to evaluate the role of genetic mutations in human longevity, only the apolipoprotein E (APOE) gene has been analyzed in several populations (3, 4). Following the discovery of the association between APOE gene and human longevity, lipid metabolism has become the most studied pathway; however, there is some evidence that human longevity is associated with an increased resistance to oxidative stress (5). Chen and colleagues have reported the relationship of centenarian longevity with single nucleotide polymorphism (SNP) rs189037 of the ataxia-telangiectasia mutated (ATM) gene (6). The protein encoded by ATM gene is resistant to oxidative stress owing to its role in the detection of reactive oxygen species (ROS)-induced lesions and repair of DNA defects (7, 8). ROS is now considered to contribute to senescence primarily by inducing DNA double-strand breaks (DSBs) and causing telomere dysfunction (9). ATM gene detects DNA damage and phosphorylates and activates downstream kinases, such as checkpoint kinases ChK1 and ChK2 and P53 (10). Moreover, an antioxidant system exists, which can reduce ROS levels. ATM gene plays a crucial role in balancing redox state, and it is widely expressed in many tissues, including the brain, skin, and endothelial cells (11). ATM expression and function are markedly decreased in aged mice. Patients with mutations in ATM gene present with insulin resistance, growth retardation, immune deficiency, and increased susceptibility to cancer. These observations suggest that ATM is critical to human lifespan (12). Studies have investigated the association between mutations in ATM gene and exceptional human longevity, but to the best of our knowledge, no studies have been conducted to assess the influence of the gene across human lifespan (6). The genetic basis for influencing human lifespan could improve the understanding of aging mechanisms, thus providing new insights and targets to avoid structural abnormality and promote successful aging. An abnormal cardiac structure is responsible for increased mortality rates and a significant obstacle in improving human longevity. At present, studies on SNPs in ATM gene have primarily focused on cancer and lifespan, and limited information regarding the specific effects of mutant rs189037 in ATM gene on cardiac structure and human longevity is available. In the current study, we aimed to determine the association of SNP rs189037 in ATM gene with cardiac structure and human longevity.
Materials and Methods
Population
As a population-based study, the China Hainan Centenarian Cohort Study (CHCCS) was performed in 18 cities and counties of Hainan Province, China, from July 2014 to December 2016 (13). It enrolled 547 centenarians identified by the National Civil Registry from the Hainan Civil Affairs Bureau. Age was ascertained from national identification cards. Among 547 centenarians, the median age was 102 years (101, 104; range: 100–116 years). The current study enrolled 250 young participants aged 20–45 years [median age: 32 years (28, 37) years; range: 22–45 years] and 250 middle-aged and elderly participants aged 46–90 years [median age: 64 years (55, 73); range: 46–89 years] in the same geographical area. These groups were carefully interviewed to exclude the presence of relatives with exceptional longevity in the past generations. The current study was approved by the Ethics Committee of Hainan Hospital of Chinese People's Liberation Army General Hospital (Sanya, Hainan; Number: 301hn11201601) and conducted in accordance with the provisions of the Helsinki Declaration. Written informed consents were obtained from all participants prior to the start of this study.
Procedures
Based on a standardized protocol, in-person interviews, physical examinations, and blood analyses were conducted by a well-trained research team of the Chinese People's Liberation Army General Hospital by home visits. This interdisciplinary research team included internists, geriatricians, cardiologists, endocrinologists, nephrologists, and nurses. Blood pressure was measured on the right arm of participants using a calibrated desktop sphygmomanometer (Yuwell Medical Equipment & Supply Co., Ltd., Jiangsu, China). Participants sat on a chair for 5 min with feet on the floor and arm supported at heart level. Systolic blood pressure (SBP) and diastolic blood pressure (DBP) were recorded at the first and fifth Korotkoff sounds, respectively. Participants with SBP ≥140 mmHg or DBP ≥90 mmHg or those receiving medications for treatment of hypertension were defined as patients with hypertension (14). Ultrasound (Philips CX50, Philips Medical Systems, Andover, MA, USA) was used to measure the parameters of cardiac structure and function by experienced radiologists who were unaware of clinical and laboratory data (15). All participants were asked to lie down in a supine position, and chest was scanned using a 1–5-MHz cardiac probe (S5–1).
Samples of venous blood were routinely collected by venipuncture and stored at 4°C and delivered to the central laboratory at the Department of Biochemistry, Hainan Hospital of Chinese People's Liberation Army General Hospital, within 4 h. Serum levels of triglyceride (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), and fasting blood glucose (FBG) were measured by enzymatic assays (Roche Products Ltd, Basel, Switzerland) using a fully automatic biochemical autoanalyzer (Cobas c702; Roche Products Ltd, Basel, Switzerland), and low-density lipoprotein cholesterol (LDL-C) level was calculated. Qualified technicians who performed laboratory analyses were blinded to clinical data.
Genotyping
Genomic DNA was isolated from peripheral blood samples using a QIAamp DNA Mini kit and eluted using 60 μl of buffer AE (Qiagen, Hilden, Germany). The concentration and purity of DNA were assessed using the Gen5 CHS 2.01 Software on the Eon microplate spectrophotometer (BioTeK, Winooski, Vermont, USA). SNP rs189037 in ATM gene was genotyped in all participants using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) and confirmed in 180 participants (>15%) by sequencing using a 3730XL automatic DNA sequencer (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA). The rate of concurrence of both methods was 100%, indicating PCR-RFLP as a reliable method. Primer design and DNA sequencing were performed by BGI (Shenzhen, China; Figure 1A). The forward and reverse primers were 5′ -GCTGCTTGGCGTTGCTTC-3′and 5′-CATGAGATTGGCGGTCTGG3′ (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA), respectively. PCR was performed in 12.5 μl total volume containing 1 μl of genomic DNA. The following PCR procedures were performed: an initial denaturation step at 94°C for 4 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 56°C for 30 s, elongation for 30 s at 72°C, and a final extension at 72°C for 5 min. ATM promoter region contains two Sac II restriction enzyme sites, and one of them is present in SNP rs189037 (C/T) only when T → C substitution at nucleotide 116 occurs (Figure 1B). Amplification products (287 bp) of ATM gene were visualized on a 1% agarose gel by staining with gelgreen (Tiangen, Beijing, China; Figure 1C). PCR products were digested with SacII (New England Biolabs, Ipswich, MA, USA) at 37°C overnight and stained with silver nitrate after resolution by electrophoresis on a 20% polyacrylamide gel (Figure 1D). All genotypes were determined as follows: three fragments of 46, 116, and 125 bp, respectively, for CC genotype; two fragments of 125 and 162 bp, respectively, for TT genotype; and four fragments of 46, 116, 125, and 162 bp, respectively, for CT genotype.
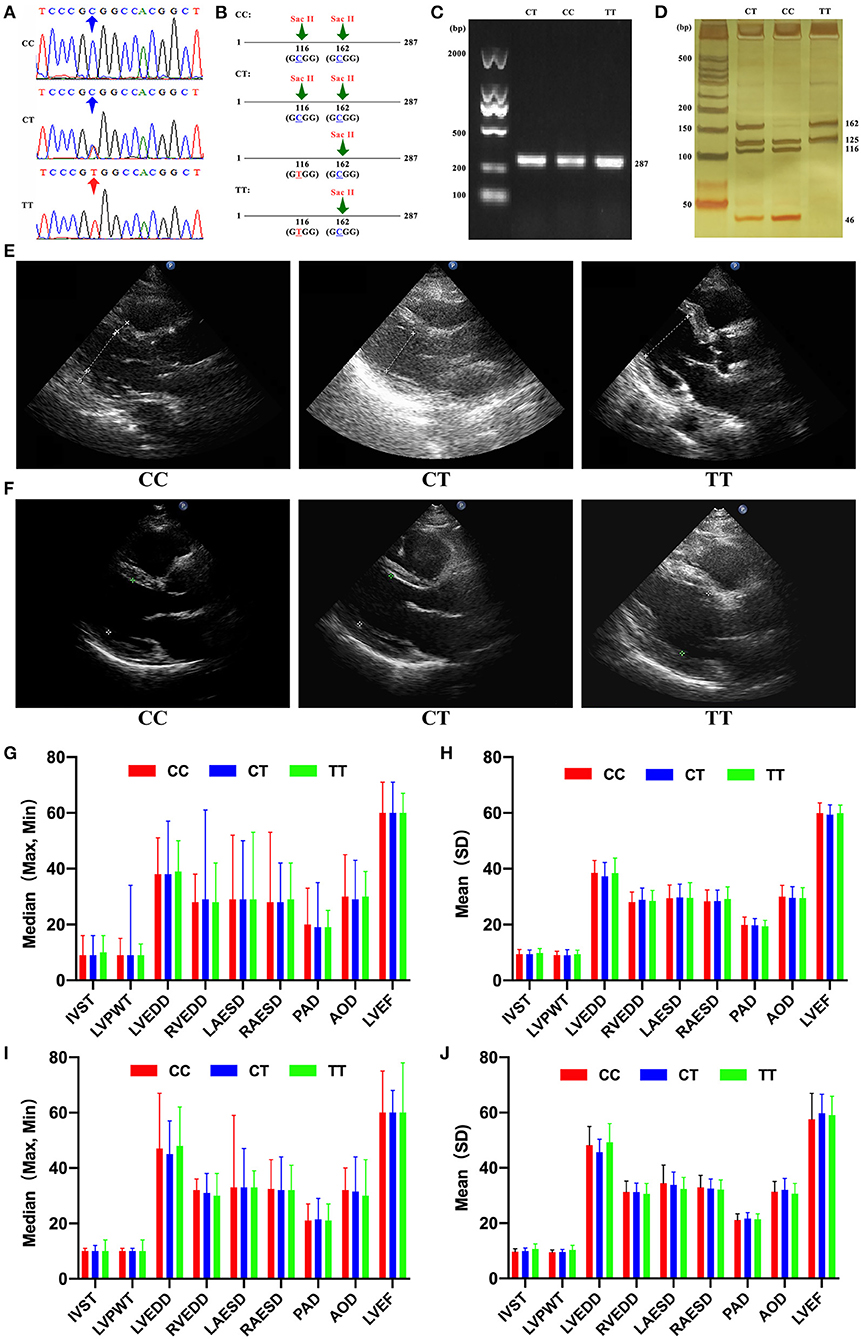
Figure 1. (A) DNA sequencing of ataxia-telangiectasia-mutated (ATM) gene with different genotypes of single nucleotide polymorphism (SNP) rs189037. (B) ATM promoter region contains two SacII restriction enzyme sites, and one of them is present in SNP rs189037 (C/T) only when T → C institution at nucleotide 116 occurs. (C) Amplification products (287 bp) of ATM gene stained with gelgreen on a 1% agarose gel. (D) Polymerase chain reaction products digested with SacII and stained with silver nitrate on a 20% polyacrylamide gel. (E) Cardiac structure shown by ultrasound in centenarians with different genotypes of SNP rs189037. (F) Cardiac structure shown by ultrasound in middle-aged and elderly participants with different genotypes of SNP rs189037. (G) Median [maximum (Max), minimum (Min)] of cardiac structure and function in centenarians with different genotypes of SNP rs189037. (H) Mean [standard deviation (SD)] of cardiac structure and function in centenarians with different genotypes of SNP rs189037. (I) Median [maximum (Max), minimum (Min)] of cardiac structure and function in middle-aged and elderly participants with different genotypes of SNP rs189037. (J) Mean [standard deviation (SD)] of cardiac structure and function in middle-aged and elderly participants with different genotypes of SNP rs189037.
Statistical Analyses
Statistical analyses were performed using SPSS software (version 17.0, SPSS Inc., Chicago, IL, USA). Data are presented as means and standard deviations (continuous variables with normal distributions), medians and interquartile ranges (continuous variables with skewed distributions), and numbers and percentages (categorical variables). Continuous data were compared using analysis of variance (ANOVA; for variables with normal distribution) or Kruskal-Wallis test (for variables with skewed distribution). Categorical data, including genotype and allele frequencies, were compared using Chi-square test. Hardy-Weinberg equilibrium was tested by Chi-square test. Multivariate logistic regression was applied to analyze the associations between the genotypes of SNP rs189037 and age groups with adjustments for sex; nationality; hypertension; and FBG, TC, TG, HDL-C, and LDL-C levels. Multivariate logistic regression was applied to analyze the associations between the genotypes of SNP rs189037 and cardiac structure and function with adjustments for age; sex; nationality; hypertension; and FBG, TC, TG, HDL-C, and LDL-C levels. Statistical significance was set at a two-sided P < 0.05.
Results
Table 1 describes the basic characteristics and genotype distribution of SNP rs189037 in young, middle-aged and elderly, and centenarian participants. The frequency of TT genotype was significantly higher and that of CC genotype was significantly lower in middle-aged and elderly participants than in young (P = 0.012) and centenarian (P = 0.041) participants. C allele in middle-aged and elderly participants was significantly less frequent compared with young (P = 0.005) and centenarian (P = 0.019) participants. There were no significant differences in the genotype and allele frequencies of SNP rs189037 between young and centenarian participants. Distributions of genotype frequencies in young (P = 0.232), middle-aged and elderly (P = 0.471), and centenarian (P = 0.982) participants were compatible with the Hardy-Weinberg equilibrium (P > 0.05).
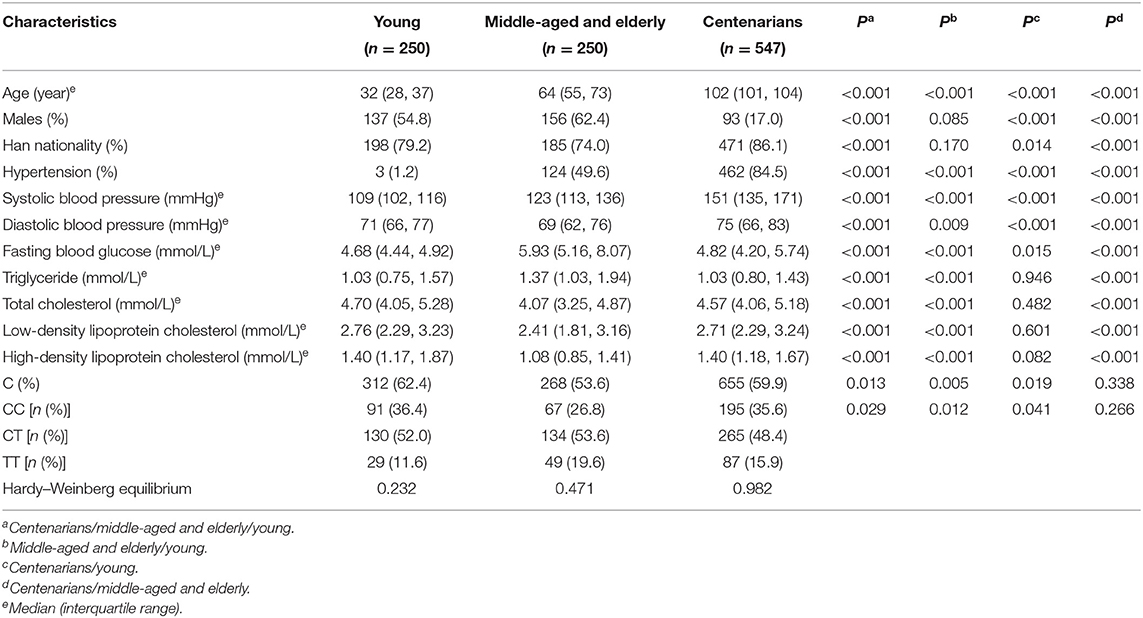
Table 1. Basic characteristics of participants with different genotypes of rs189037.
Table 2 shows the multiple associations of age groups with different genotypes of SNP rs189037. Compared with CC genotype, TT genotype was positively and significantly associated with middle-aged and elderly/young participants [Exp(B) (95% CI): 3.303 (1.107–9.859); P = 0.032]. The genotypes of SNP rs189037 were not significantly associated with centenarian/young participants.
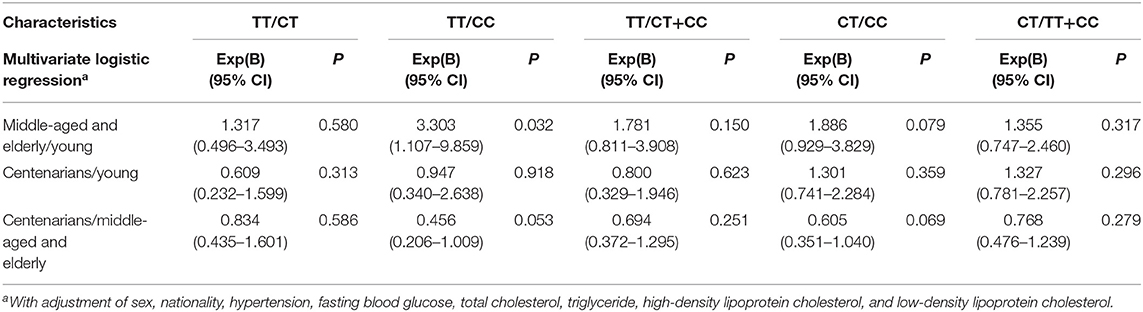
Table 2. Associations of age group with different genotypes of rs189037.
The basic characteristics of centenarians with different genotypes of SNP rs189037 are presented in Table 3. Centenarians with TT and CT genotypes showed significant differences in left ventricular posterior wall thickness (LVPWT; P = 0.004). Centenarians with TT and CC genotypes showed significant differences in LVPWT (P = 0.047; Figures 1E,G,H). Centenarians with CT and CC genotypes showed significant differences in LVEDD (P = 0.008).
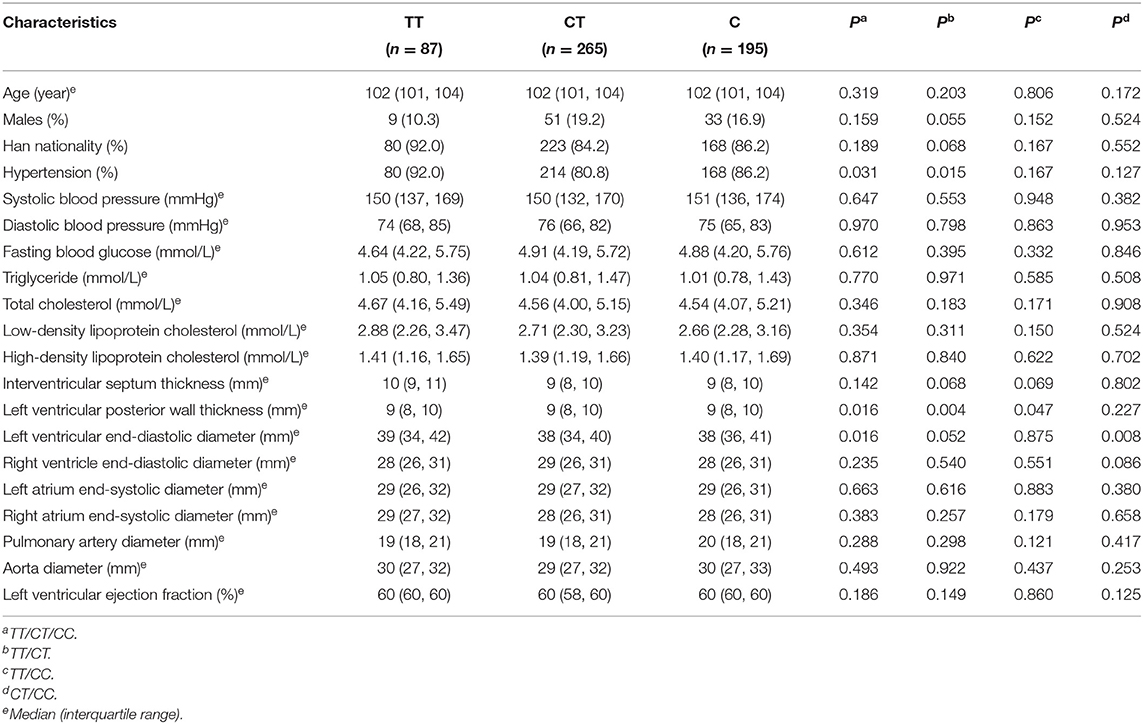
Table 3. Basic characteristics of centenarians with different genotypes of rs189037.
Table 4 shows multiple associations of cardiac structure and function with different genotypes of SNP rs189037 in centenarians. Compared with CT genotype, TT genotype was positively and significantly associated with IVS [Exp(B) (95% CI): 1.181 (1.001–1.394); P = 0.049] and LVPWT [Exp(B) (95% CI): 1.152 (1.002–1.324); P = 0.047] in centenarians. Compared with CC genotype, TT genotype was positively and significantly associated with LVPWT [Exp(B) (95% CI): 1.244 (1.021–1.517); P = 0.030] in centenarians. Compared with CC genotype, CT genotype was negatively and significantly associated with LVEDD [Exp(B) (95% CI): 0.947 (0.908–0.988); P = 0.011] and positively and significantly associated with RVEDD [Exp(B) (95% CI): 1.064 (1.009–1.123); P = 0.022] in centenarians.
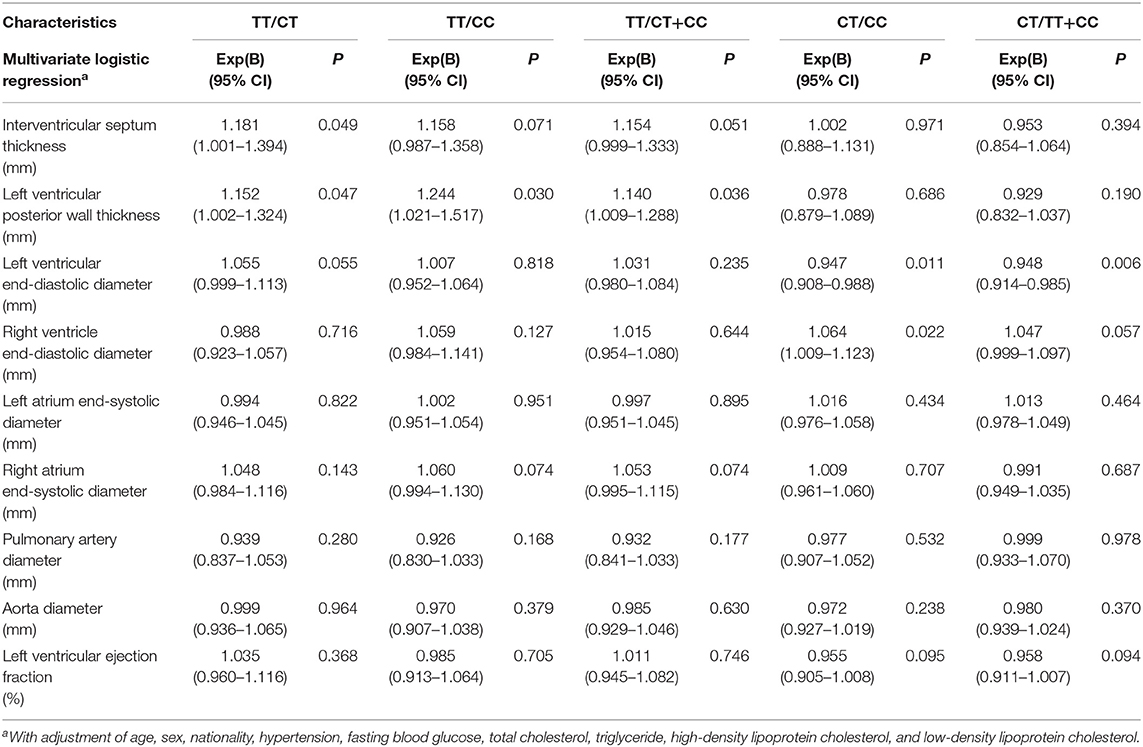
Table 4. Associations of cardiac structure and function with different genotypes of rs189037 in the centenarians.
The basic characteristics of middle-aged and elderly participants with different genotypes of SNP rs189037 are presented in Table 5. Middle-aged and elderly participants with TT and CT genotypes showed significant differences in LVPWT (P = 0.037), LVEDD (P = 0.015), and left ventricular ejection fraction (LVEF, P = 0.032). Middle-aged and elderly participants with TT and CC genotypes showed significant differences in LVPWT (P = 0.043; Figures 1F,I,J).
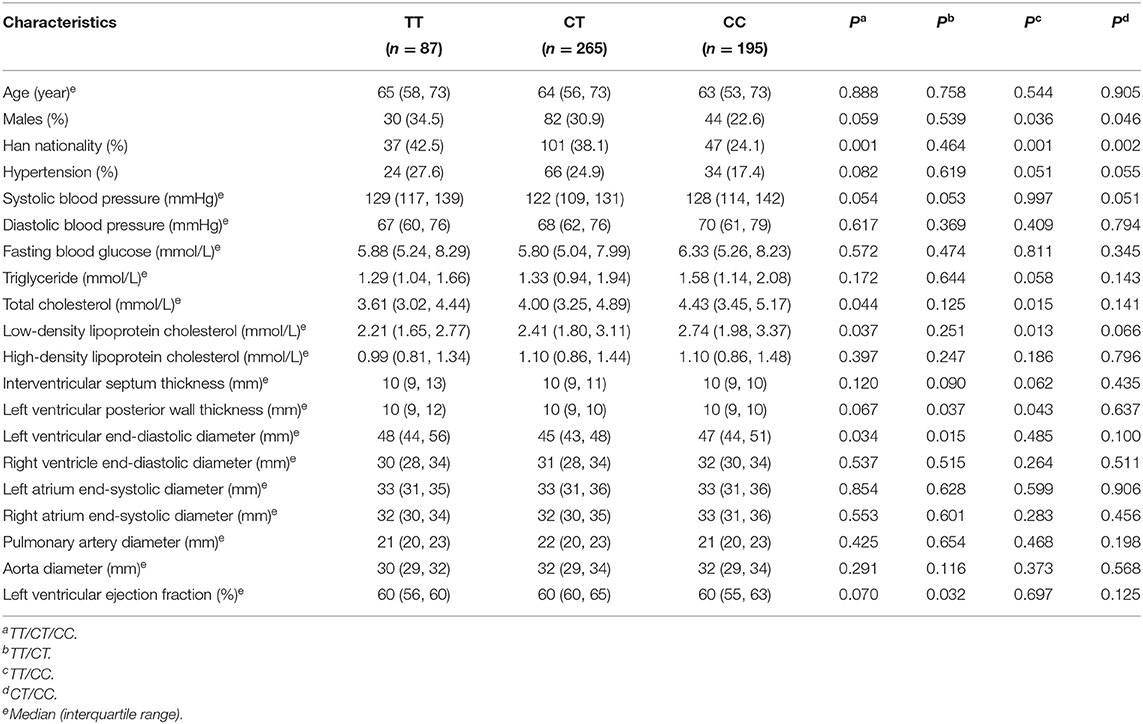
Table 5. Basic characteristics of middle-aged and elderly with different genotypes of rs189037.
Table 6 shows multiple associations of cardiac structure and function with different genotypes of SNP rs189037 in middle-aged and elderly participants. Compared with CT genotype, TT genotype was positively and significantly associated with IVST [Exp(B) (95% CI): 1.739 (1.156–2.615); P = 0.008], LVPWT [Exp(B) (95% CI): 2.016 (1.244–3.267); P = 0.004], and LVEDD [Exp(B) (95% CI): 1.207 (1.070–1.361); P = 0.002] in middle-aged and elderly participants. Compared with CC genotype, TT genotype was positively and significantly associated with IVST [Exp(B) (95% CI): 1.801 (1.141–2.843); P = 0.012] and LVPWT [Exp(B) (95% CI): 1.925 (1.145–3.236); P = 0.013] in middle-aged and elderly participants. Compared with CC genotype, CT genotype was negatively and significantly associated with LVEDD [Exp(B) (95% CI): 0.909 (0.829–0.996); P = 0.040] in middle-aged and elderly participants.
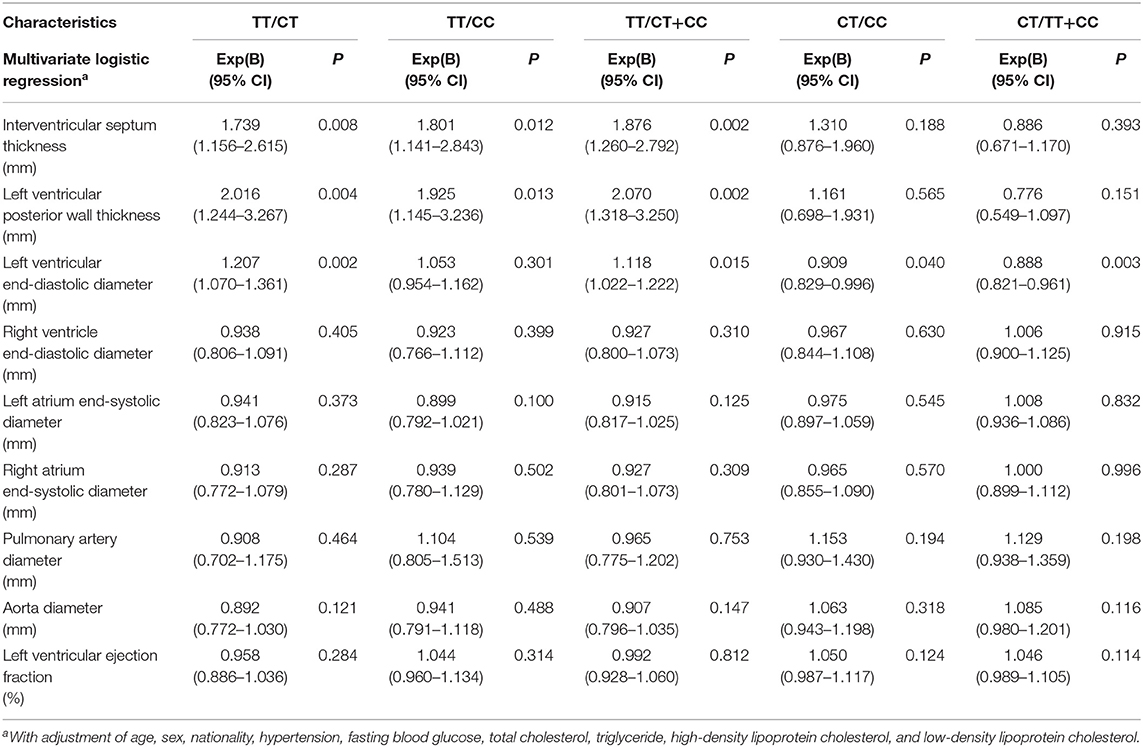
Table 6. Associations of cardiac structure and function with different genotypes of rs189037 in the middle-aged and elderly.
Discussion
In middle-aged and elderly and centenarian participants, the current study demonstrated that CT genotype of SNP rs189037 was significantly associated with reduced LVEDD, while TT genotype was significantly associated with increased IVST, LVPWT, and LVEDD. For SNP rs189037 in ATM gene, heterozygous mutation might result in LV concentric development, whereas homozygous mutation might further induce LV eccentric hypertrophy. Moreover, the frequency of TT genotype was similar between young and centenarian participants; however, middle-aged and elderly participants showed significantly higher frequency of TT genotype. SNP rs189037 without mutation might be an indicator of youth health and successful aging, whereas mutant rs189037 might hinder successful aging and human longevity.
Human lifespan is the outcome of multiple processes involving genetic factors; thus, several genetic studies have investigated putative associations between genes and longevity, but to the best of our knowledge, no studies have been conducted to assess the influence of the genes across human lifespan. The current study identified SNP rs189037 without mutation as a biomarker of youth health and successful aging by analyzing genotypic change in ATM across human lifespan. ROS can lead to DSBs and telomere dysfunction, which are critical causes of DNA lesions (8). Once DNA lesions occur, the protein encoded by ATM gene phosphorylates numerous downstream effectors involved in the G1/S, intra-S, and G2/M checkpoint responses and additional factors involved in DNA repair, including tumor suppressor proteins P53, ChK1, and ChK2 (10). Therefore, ATM mutations possibly make individuals susceptible to cancer and are harmful for physiological activity. This suggests that ATM gene plays a significant role in prolonging human lifespan.
Accumulating evidence has illustrated that SNPs in gene regulate gene expression, and people with different SNPs show distinct phenotypes (16, 17). A previous study has shown the presence of an AP-2a-binding site around SNP rs189037 in ATM gene, which is a transcriptional factor involved in both physiological and pathological processes, such as tumorigenesis and morphogenesis (18). It has been verified in human cells that TT genotype of SNP rs189037 increases ATM mRNA expression, and AP-2a represses ATM expression in humans by binding to CC genotype (6). It is also believed that balance exists in every activity of the physiological process; mutant SNP in gene alters protein phenotypes, disrupts physiological balance, and causes pathological changes. For example, mutant SNP rs189037 in ATM gene or consistently high activity of P53 results in abnormal cardiac structure or premature senescence of individuals (19). The current study reported that SNP rs189037 without mutation might prevent the development of an abnormal cardiac structure and death at an early age; hence, it is not only an indicator and biomarker of youth health and successful aging but also promotes successful aging and prolongs human lifespan.
Aging is a complicated process affected by several genetic and non-genetic factors (1). Genes are the predominant factors responsible for aging, and SNPs associated with aging might exist. Mice with deficient ATM gene have been shown to experience accelerated aging (20). As a regulator of oxidative stress and sensor of DNA damage, ATM gene prevents cellular senescence, death, or cell cycle arrest; in addition, it protects individuals from ROS-induced DNA damage. Ding et al. have described an association of SNP rs189037 in ATM gene with the proportion of coronary artery disease in Chinese Han population and demonstrated a low proportion of coronary artery disease in participants harboring TT genotype (21). In the current study, the young and centenarian participants showed significantly lower frequency of mutant rs189037 in ATM gene than middle-aged and elderly participants. Individuals without mutant rs189037 in ATM gene may age later and live long, and reversing mutant rs189037 might be a suitable strategy to delay human aging and promote human longevity.
Conclusions
The current study demonstrated that mutant SNP rs189037 in ATM gene was more commonly identified in the middle-aged and elderly participants than in young participants and centenarians. It was significantly associated with increased LV thickness and volume and might induce LV eccentric hypertrophy and shorten human lifespan.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethics Committee of the Hainan Hospital of the Chinese People's Liberation Army General Hospital (Sanya, Hainan; No: 301hn11201601). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
SF, JH, XC, BL, HS, JD, YJZ, YY, and YLZ contributed to the study design, conducted the data collection and analyses, and drafted the paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by grants from the National Natural Science Foundation of China (81900357, 81903392, 81941021, 81901252, 82001476), the Military Medical Science and Technology Youth Incubation Program (20QNPY110), the National Key R&D Program of China (2018YFC2000400), the National S&D Resource Sharing service platform Project of China (YCZYPT[2018]07), the General Hospital of PLA Medical Big Data R&D Project (MBD2018030), the China Postdoctoral Science Foundation-funded project (2019M650359), the National Geriatric Disease Clinical Medicine Research Center Project (NCRCG-PLAGH-2017-014), the Central Health Care Scientific Research Project (W2017BJ12), the Hainan Medical and Health Research Project (16A200057), the Sanya Medical and Health Science and Technology Innovation Project (2016YW21, 2017YW22, 2018YW11), the Military Medicine Youth Program of Chinese PLA General Hospital (QNF19069), and the Clinical Scientific Research Supporting Fund of Chinese PLA General Hospital (2017FC-CXYY-3009).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate all the staff of CHCCS for their continued cooperation and contribution in fieldwork.
Abbreviations
ApoE, apolipoprotein E; ATM, ataxia-telangiectasia mutated; ChK, checkpoint kinases; CHCCS, China Hainan Centenarian Cohort Study; DBP, diastolic blood pressure; DNA, deoxyribonucleic acid; DSBs, DNA double-strand breaks; FBG, fasting blood glucose; HDL-C, high-density lipoprotein cholesterol; IVST, interventricular septum thickness; LDL-C, low-density lipoprotein cholesterol; LVEDD, left ventricular end-diastolic diameter; LVPWT, left ventricular posterior wall thickness; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; RVEDD, right ventricle end-diastolic diameter; SBP, systolic blood pressure; SNP, single nucleotide polymorphism; TC, total cholesterol; TG, triglyceride.
References
1. Christensen K, Johnson TE Vaupel JW. The quest for genetic determinants of human longevity: challenges and insights. Nat Rev Genet. (2006) 7:436–48. doi: 10.1038/nrg1871
2. Sebastiani P, Solovieff N, Dewan AT, Walsh KM, Puca A, Hartley SW, et al. Genetic signatures of exceptional longevity in humans. PLoS ONE. (2012) 7:e29848. doi: 10.1371/journal.pone.0029848
3. Schächter F, Faure-Delanef L, Guénot F, Rouger H, Froguel P, Lesueur-Ginot L, et al. Genetic associations with human longevity at the APOE and ACE loci. Nat Genet. (1994) 6:29–32. doi: 10.1038/ng0194-29
4. Bostock CV, Soiza RL Whalley LJ. Genetic determinants of ageing processes and diseases in later life. Maturitas. (2009) 62:225–9. doi: 10.1016/j.maturitas.2008.12.012
5. Lee HC Wei YH. Mitochondria and aging. Adv Exp Med Biol. (2012) 942:311–27. doi: 10.1007/978-94-007-2869-1_14
6. Chen T, Dong B, Lu Z, Tian B, Zhang J, Zhou J, et al. A functional single nucleotide polymorphism in promoter of ATM is associated with longevity. Mech Ageing Dev. (2010) 131:636–40. doi: 10.1016/j.mad.2010.08.009
7. Lavin MF, Birrell G, Chen P, Kozlov S, Scott S, Güven N. ATM signaling and genomic stability in response to DNA damage. Mutat Res. (2005) 569:123–32. doi: 10.1016/j.mrfmmm.2004.04.020
8. Lombard DB, Chua KF, Mostoslavsky R, Franco S, Gostissa M, Alt FW. DNA repair, genome stability, and aging. Cell. (2005) 120:497–512. doi: 10.1016/j.cell.2005.01.028
9. Herbig U, Ferreira M, Condel L, Carey D, Sedivy JM. Cellular senescence in aging primates. Science. (2006) 311:1257. doi: 10.1126/science.1122446
10. Shiloh Y. ATM and related protein kinases: safeguarding genome integrity. Nat Rev Cancer. (2003) 3:155–68. doi: 10.1038/nrc1011
11. McKinnon PJ. ATM and ataxia telangiectasia. EMBO Rep. (2004) 5:772–6. doi: 10.1038/sj.embor.7400210
12. Puca AA, Daly MJ, Brewster SJ, Matise TC, Barrett J, Shea-Drinkwater M, et al. A genome-wide scan for linkage to human exceptional longevity identifies a locus on chromosome 4. Proc Natl Acad Sci USA. (2001) 98:10505–8. doi: 10.1073/pnas.181337598
13. Fu S, Li Y, Zhang F, Luan F, Lv F, Deng J, et al. Centenarian longevity is positively correlated with IgE levels but negatively correlated with C3/C4 levels, abdominal obesity and metabolic syndrome. Cell Mol Immunol. (2020) 17:1196–7. doi: 10.1038/s41423-020-0386-y
14. Fu S, Luo L, Ye P, Liu Y, Zhu B, Zheng J, et al. Overall and abdominal obesity indicators dad different association with central arterial stiffness and hemodynamics independent of age, sex, blood pressure, glucose, and lipids in Chinese community-dwelling adults. Clin Interv Aging. (2013) 8:1579–84. doi: 10.2147/CIA.S54352
15. Fu S, Yao Y, Wu S, Deng J, Lv F, Zhao Y. Inverse association between periumbilical fat and longevity mediated by complement C3 and cardiac structure. Aging. (2020) 12:23296–305. doi: 10.18632/aging.104113
16. Sun T, Gao Y, Tan W, Ma S, Shi Y, Yao J, et al. A six-nucleotide insertion-deletion polymorphism in the CASP8 promoter is associated with susceptibility to multiple cancers. Nat Genet. (2007) 39:605–13. doi: 10.1038/ng2030
17. Giacconi R, Cipriano C, Albanese F, Boccoli G, Saba V, et al. The-174G/C polymorphism of IL-6 is useful to screen old subjects at risk for atherosclerosis or to reach successful ageing. Exp Gerontol. (2004) 39:621–28. doi: 10.1016/j.exger.2003.12.013
18. Leask A, Byrne C Fuchs E. Transcription factor AP2 and its role in epidermal-specifific gene expression. Proc Natl Acad Sci USA. (1991) 15:7948–52. doi: 10.1073/pnas.88.18.7948
19. Tyner SD, Venkatachalam S, Choi J, Jones S, Ghebranious N, Igelmann H, et al. p53 mutant mice that display early ageing-associated phenotypes. Nature. (2002) 415:45–53. doi: 10.1038/415045a
20. Ito K, Hirao A, Arai F, Matsuoka S, Takubo K, Hamaguchi I, et al. Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature. (2004) 431:997–1002. doi: 10.1038/nature02989
Keywords: ataxia-telangiectasia mutated gene, human lifespan, mutant rs189037, ventricular thickness, human longevity
Citation: Fu S, Hu J, Chen X, Li B, Shun H, Deng J, Zhang Y, Yao Y and Zhao Y (2021) Mutant Single Nucleotide Polymorphism rs189037 in Ataxia-Telangiectasia Mutated Gene Is Significantly Associated With Ventricular Wall Thickness and Human Lifespan. Front. Cardiovasc. Med. 8:658908. doi: 10.3389/fcvm.2021.658908
Received: 26 January 2021; Accepted: 20 April 2021;
Published: 26 May 2021.
Edited by:
Christian Schulte, University Heart and Vascular Center Hamburg (UHZ), GermanyReviewed by:
Christoph Sinning, University Heart and Vascular Center Hamburg (UHZ), GermanyRalf Schubert, Goethe University Frankfurt, Germany
Copyright © 2021 Fu, Hu, Chen, Li, Shun, Deng, Zhang, Yao and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juelin Deng, anVlbGluZDIwMTImI3gwMDA0MDsxNjMuY29t; Yujie Zhang, ZG9jemhhbmd5dWppZSYjeDAwMDQwOzE2My5jb20=; Yao Yao, eWFveWFvJiN4MDAwNDA7bnNkLnBrdS5lZHUuY24=; Yali Zhao, emhhb3lsMzAxJiN4MDAwNDA7MTYzLmNvbQ==
†These authors have contributed equally to this work and share first authorship