 Canan G. Nebigil
Canan G. Nebigil- CNRS-University of Strasbourg, UMR 7242, Illkirch, France
Obesity is a fast growing epidemic event worldwide. Fatness is associated with a number of comorbidities, including cardiovascular diseases (CVDs). Although obesity can be heredity in 30–70% cases, the environmental contributions also play an important role in the increasing prevalence of obesity. The relationship between development of obesity and CVD is poorly characterized. Obesity and CVD can also be resulted from a common mechanism such as metabolic, inflammatory, and neurohormonal changes. Prokineticins are defined as cytokines (immunoregulatory proteins), adipokines (adipocyte-secreted hormone), angiogenic (increasing vessel formation), or aneroxic (lowering food intake) hormones. Prokineticin-mediated signaling plays a key role in the development of obesity and CVD. Two forms of prokineticins exist in circulation and in various tissues including the brain, heart, kidney, and adipose. Prokineticins act on the two G protein-coupled receptors, namely, PKR1 and PKR2. Prokineticin-2 (PK2) via PKR1 receptor controls food intake and prevents adipose tissue expansion. The anti-adipocyte effect of PKR1 signaling is due to suppression of preadipocyte proliferation and differentiation capacity into adipocytes. PK2/PKR1 signaling promotes transcapillary passages of insulin and increases insulin sensitivity. It also plays an important role in the heart and kidney development and functions. Here, we discuss PK2 as a new adipocytokine in the association between obesity and CVD. We also highlight targeting PKR1 can be a new approach to treat obesity and CVD.
Introduction
Obesity is a major health problem in worldwide regardless of sex and age (1). It is both an independent risk factor and a risk marker for the development of asymptomatic and symptomatic cardiovascular disease (CVD) (2). Common pathways may involve in the pathogenesis of obesity and CVD. Indeed, CVD can occur due to structural and functional changes of the myocardium through excess fat deposition and constant and unremitting metabolic stress related to obesity (2). Interestingly, anti-obesity therapies with anorexic peptides improve cardiovascular function and reduce cardiovascular morbidity and mortality (3). Recent evident showed that some brain regions is involved in food intake regulation and also play an important role in regulation of cardiovascular-blood homeostasis (4). Therefore, it is important to delineate the common mechanisms regulating both obesity and cardiovascular events for development of novel therapeutics. Here, we outlined the current information on the role of anorexic and angiogenic peptide prokineticin signaling in obesity and CV-renal diseases.
Prokineticins and Their Receptors
Prokineticins are released by monocytes, macrophages, and reproductive organs (5). A high level of prokineticins has been found in obese human AT (6), as well as heart and kidney (7, 8). Two isoforms of prokineticins have been identified: prokineticin-1 and prokineticin-2 (PK2).
PK2 is an Anorexic Peptide
The regulation of food intake is a complex process involving reciprocal signals between the central nervous system and the periphery. The region in the hypothalamus governing a feeding and energy homeostasis is called as arcuate nucleus (ARC). The ARC contains primary neurons that express neuropeptides with opposing effects on food intake. ARC neurons release anorexic peptide such as the proopiomelanocortin (POMC)-derived peptide, alpha-melanocyte-stimulating hormone, and cocaine, amphetamine-regulated transcript (CART) peptide (9). However, in the ARC, neuropeptide Y (NPY)-producing neurons have been shown to stimulate food intake.
Prokineticin-2 controls food intake and fat tissue expansion through actions in the ARC in the hypothalamus (10). Mainly, PKR1 receptors are expressed in the NPY/AgRP and POMC/CART neurons. Intracranial injection of PK2 in rats abolishes food intake, whereas anti-PK2 antibody increases food intake. Anorexic effect of PK2 is mediated at least partly via the hypothalamic ARC melanocortin system. Indeed, PKR1 is the first non-melanocortin G protein-coupled receptors to be regulated by the melanocortin receptor accessory protein 2 that inhibits specifically PKR1 signaling (11).
Peripheral administration of PK2 reduces food intake and body weight in both lean mice and diet-induced obesity models (12). Global ablation of PK2 in mice leads to obesity. Hypothalamic PK2 levels were found extremely high in the early neonatal period. However, a low level of PK2 was observed under fasting conditions (13). The inactivating mutations of PK2 gene and the obesity have been correlated in human (12, 14). Anorexic effect of PK2 was completely absent in the PKR1 deficient mice (12), indicating that the anorexic effects of PK2 are mediated by PKR1 in the hypothalamus.
Prokineticin in Obesity
Obesity can be resulted from adipocyte hypoplasia/hyperthrophy accompanied with inflammation of AT, defective of extracellular matrix remodeling, fibrosis, and an altered secretion or expression of adipokines (15). PK2 releases from AT in obese individuals; however, it suppresses AT expansion by two distinct mechanisms: the central regulation of food intake and limiting preadipocyte function.
In isolated preadipocytes, PKR1 activation suppresses proliferation and adipogenic differentiation (6). Indeed, an abnormally excessive abdominal fat mass accumulation was observed in adipose tissue-specific PKR1-deficient (PKR1ad−/−) mice (6). The expansion of AT in both PKR1 null and PKR1ad−/− mice was due to formation of new adipocytes. These mice displayed an acceleration of preadipocyte proliferation and differentiation. Despite PKR1null and PKR1ad−/− mice display abdominal obesity, only PKR1null mice have peripheral obesity with a diabetes-like syndrome (6). Thus, non-adipocyte PKR1-mediated events may contribute to the development of a diabetes-like syndrome. Angiogenesis has important roles in the modulation of insulin sensitivity and expansion of AT (16). Indeed, endothelial-specific PKR1 knockout mice (PKR1ec−/−) had insulin resistance in adipocytes (17). Insulin cannot promote normal fat storage, resulting in excess circulating free fatty acids that, in turn, further contribute into insulin resistance in muscle, leading to diabetes-like syndrome in PKR1ec−/− adipocytes.
The expansion of AT in obesity is also required a shift in the polarized states of macrophages from the M2 to the pro-inflammatory M1 form (18). PK2 promotes inflammatory phenotype of mouse macrophages (19) and reduces IL-10 and IL-4 production in mice splenocytes (20). In contrast, PKR1ad−/− mice displayed substantial infiltration of macrophage in the AT. Whether PKR1 signaling retains an M2 polarization, or triggers the phenotypic switch from M1 to M2 to preserve adequate adipocyte function in obesity is unknown.
Prokineticin in Insulin Resistance
The transcapillary delivery of insulin from endothelial cells (ECs) to the skeletal muscle is the rate-limiting step in insulin-stimulated glucose uptake (21). The defect in insulin delivery process via ECs contributes to insulin resistance (22). Thus, the vascular endothelium is considered as a potential therapeutic target for prevention of insulin resistance and related complications (23).
Endothelial cell-specific PKR1 knockout (PKR1ec−/−) mice exhibited impaired capillary formation and low transcapillary insulin uptake, which was rescued by PKR1 gene transfection by adenovirus (17). Overexpressing PKR1 in EC promotes insulin transendothelial uptake (24) and angiogenesis (25). These data highlight the role of PKR1 as a positive regulator of insulin uptake (26). In concert with this in vitro finding, PKR1ec−/− mice exhibit hyperphagia and severe lipodystrophy due to poor capillary formation in the AT. Lipodystrophies, involving a loss of AT, are known to induce hyperphagia and peripheral insulin resistance (27). Impaired insulin delivery and signaling in ECs have also been observed in human patients with type 2 diabetes and obesity with insulin resistance (28).
Therapeutic strategies targeting PKR1 could be important to treat obesity and obesity-associated insulin resistance, since PKR1 signaling suppresses appetite, reduces adipocyte expansion, promotes normal fat storage, and increases insulin sensitivity.
Prokineticin in Heart Development and Function
PKR1 regulates epicardial–mesenchymal transition to form epicardial-derived progenitor cell (EPDC) during cardiogenesis (29). Genetic ablation of PKR1 in epicardium (PKR1wt1−/−) leads to a ventricular hypoplasia, septal defects, and deficient vascularization, leading to embryonic lethality. Epicardial PKR1 contributes to cardiomyocyte proliferation in a paracrine pathway that is required for the development of ventricular wall. Epicardial PKR1 is also a key signaling for EPDC proliferation and differentiation into vasculogenic cell type, involved in formation of coronary circulation (30).
Altered expression of prokineticins and their receptors has been implicated in heart failure (31) and aortic rupture (32). Prokineticin signaling plays an important role especially in cardiac progenitor cell commitment and cell-to-cell communications (30).
PKR1 signaling protects cardiomyocytes against hypoxia-mediated apoptosis by activating Akt signaling pathway (7). Transgenic mice-overexpressing PKR1 in the cardiomyocytes exhibits an increased number of EPDCs, associated with increased number of vessels (30). Indeed, the cardiac PKR1 signaling up-regulates its own ligand PK2 to stimulate the EPDC differentiation into endothelial and smooth muscle cells to promote neovasculogenesis (30). Interestingly, PKR1null mice displayed cardiomyocyte contractile defects and apoptosis partially due to lack of PKR1 signaling in cardiomyocytes (8). These data indicate that cardiomyocyte PKR1 is essential for cardiomyocyte survival and contractility with a cell autonomous way. However, cardiomyocyte PKR1 derives EPDCs proliferation and differentiation.
In cardiac ECs, PKR1 activates Akt and MAPK to promote proliferation, migration, and angiogenesis (25). Accordingly, loss of PKR1 in ECs leads to defective angiogenesis (17). The posterior walls of PKR1ec−/− hearts were thinner due to the loss of capillary formation and a high level of apoptosis (17). Indeed, PKR1ec−/− hearts displayed ectopic lipid deposition and abnormal insulin signaling together with capillary defects, resulting in impaired diastolic function. Abnormal insulin signaling was due to defective transcapillary transport of insulin in the vascular wall of PKR1ec−/−mice. In accord with this in vivo findings, isolated ECs from the mutant cardiac and renal tissues exhibited very little insulin uptake, confirming that the loss of PKR1 from ECs decreased insulin transport (17). Indeed, activation of PKR1 in ECs promoted FITC-insulin passage. Nitric oxide deficiency in the ECs is associated with the insulin resistance and endothelial dysfunction (33). Similarly, in the endothelium of patients with diabetes mellitus, insulin-mediated eNOS activation is altered (34). In agreement, insulin uptake and insulin-mediated eNOS activation were impaired in PKR1-deficient ECs. Impaired transcapillary insulin delivery leads to defective eNOS activation, affecting endothelium-dependent relaxation in PKR1ec−/−aortas (17). These impairments in PKR1ec−/− mice resulted in hypertension at the later age. These mice models should facilitate studies of both pathogenesis and therapy of cardiac disorders in humans.
Prokineticin in Renal Development and Function
In contrast to developing heart, PKR1 is necessary for renal mesenchymal–epithelial transition (MET) that is involved in formation of renal progenitors, regulating glomerulogenesis toward forming nephrons during kidney development (29). Indeed, PKR1 activates NFATc3 and modifies MET processing involved in the development of nephron. Mutant mice with targeted PKR1 gene disruptions in nephron progenitors has been shown to exhibit partial embryonic and postnatal lethality due to hypoplastic kidneys with premature glomeruli and necrotic nephrons. Kidney developmental defects in these mice were manifested in the adult stage as renal atrophy with glomerular defects, nephropathy, and uremia (29).
PKR1 knockout mice also displayed renal tubular dilation, reduced glomerular capillaries, urinary phosphate excretion, and proteinuria (8). Similarly, PKR1ec−/− mice displayed enlarged tubular structures with a swollen necrotic nucleus, abnormal mitochondria, and aberrant organization of podocytes, dilatation of the Bowman’s spaces in the glomeruli, a compact glomerulus, and fibrosis. Defects in tubular and glomerular structures are associated with functional abnormalities such as high levels of creatinine clearance and proteinuria (8). Indeed, increased excretion of absolute renal phosphate (Pi) in the PKR1ec−/− mice is due to lower levels of sodium–calcium and sodium–phosphate exchanger. The morphological changes in the PKR1ec−/− kidneys were accompanied with apoptosis impaired insulin signaling and lipid accumulation. Endothelial dysfunction resulted from loss of PKR1 signaling underlies the pathological features of heart and kidney.
Conclusion
Multiple biological mechanisms linking obesity and CVD events have been identified. Identification of signaling pathways linking obesity and CVD is important for development of novel therapeutics. PKR1 signaling plays an important role in central regulation of appetite, the suppression of adipocyte mass and insulin sensitizing effects on skeletal muscle and other tissues, cardiac regeneration, and kidney development and function (Figure 1). Whether PKR1 signaling regulates heart and kidney function via vagus nerve remains to be study. Recently, PKR1 non-peptide agonist has been discovered (35), which it prevents cardiac lesion formation and improves cardiac function after myocardial infarction in mice, promoting proliferation of cardiac progenitor cells and neovasculogenesis. Targeting PKR1 can be a novel therapeutic approach to treat obesity, diabetes and CVD.
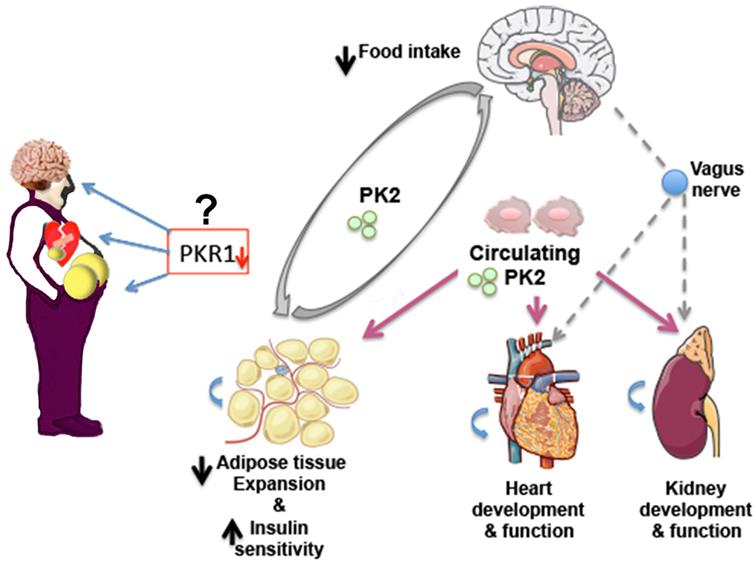
Figure 1. Prokineticin-2 (PK2)/PKR1 signaling may act as a new connector between development of obesity, diabetes and cardiovascular diseases (CVDs). PKR1 deficiency promotes WAT expansion and insulin resistance, CVD, and alters food intake in mice. Whether reduced level of PKR1 or functional mutated PKR1 involves these disorders in human needs to be studied. PK2/PKR1 signaling in central nervous system (CNS) regulates food intake. PK2 released from adipocytes controls preadipocyte conversion to adipocyte via PKR1 signaling and may affect food intake via CNS. Circulating or local PK2 signaling via PKR1 contributes development and function of heart and kidney. Whether this signaling involves heart and kidney regulation via CNS remains to be study.
Author Contributions
The author confirms being the sole contributor of this work and approved it for publication.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
The publication was supported in part by grants from Fondation pour la Recherche Médicale (Equipe Labellisée), Centre National de la Recherche Scientifique, and Université de Strasbourg. This work has also been published within the LABEX ANR-10-LABX-0034_Medalis and received a financial support from the French government managed by Agence Nationale de la Recherche (ANR) under “Programme d’investissement d’avenir.”
References
2. Mandviwala T, Khalid U, Deswal A. Obesity and cardiovascular disease: a risk factor or a risk marker? Curr Atheroscler Rep (2016) 18:21. doi:10.1007/s11883-016-0575-4
3. Valassi E, Scacchi M, Cavagnini F. Neuroendocrine control of food intake. Nutr Metab Cardiovasc Dis (2008) 18:158–68. doi:10.1016/j.numecd.2007.06.004
4. Mikulaskova B, Maletinska L, Zicha J, Kunes J. The role of food intake regulating peptides in cardiovascular regulation. Mol Cell Endocrinol (2016) 436:78–92. doi:10.1016/j.mce.2016.07.021
5. Kaser A, Winklmayr M, Lepperdinger G, Kreil G. The AVIT protein family. Secreted cysteine-rich vertebrate proteins with diverse functions. EMBO Rep (2003) 4:469–73. doi:10.1038/sj.embor.embor830
6. Szatkowski C, Vallet J, Dormishian M, Messaddeq N, Valet P, Boulberdaa M, et al. Prokineticin receptor 1 as a novel suppressor of preadipocyte proliferation and differentiation to control obesity. PLoS One (2013) 8:e81175. doi:10.1371/journal.pone.0081175
7. Urayama K, Guilini C, Messaddeq N, Hu K, Steenman M, Kurose H, et al. The prokineticin receptor-1 (GPR73) promotes cardiomyocyte survival and angiogenesis. FASEB J (2007) 21:2980–93. doi:10.1096/fj.07-8116com
8. Boulberdaa M, Turkeri G, Urayama K, Dormishian M, Szatkowski C, Zimmer L, et al. Genetic inactivation of prokineticin receptor-1 leads to heart and kidney disorders. Arterioscler Thromb Vasc Biol (2011) 31:842–50. doi:10.1161/ATVBAHA.110.222323
9. Coll AP, Farooqi IS, Challis BG, Yeo GS, O’Rahilly S. Proopiomelanocortin and energy balance: insights from human and murine genetics. J Clin Endocrinol Metab (2004) 89:2557–62. doi:10.1210/jc.2004-0428
10. Gardiner JV, Bataveljic A, Patel NA, Bewick GA, Roy D, Campbell D, et al. Prokineticin 2 is a hypothalamic neuropeptide that potently inhibits food intake. Diabetes (2010) 59:397–406. doi:10.2337/db09-1198
11. Chaly AL, Srisai D, Gardner EE, Sebag JA. The melanocortin receptor accessory protein 2 promotes food intake through inhibition of the prokineticin receptor-1. Elife (2016) 5:e12397. doi:10.7554/eLife.12397
12. Beale K, Gardiner JV, Bewick GA, Hostomska K, Patel NA, Hussain SS, et al. Peripheral administration of prokineticin 2 potently reduces food intake and body weight in mice via the brainstem. Br J Pharmacol (2013) 168:403–10. doi:10.1111/j.1476-5381.2012.02191.x
13. Iwasa T, Matsuzaki T, Munkhzaya M, Tungalagsuvd A, Kawami T, Murakami M, et al. Changes in the responsiveness of hypothalamic prokineticin 2 mRNA expression to food deprivation in developing female rats. Int J Dev Neurosci (2014) 34:76–8. doi:10.1016/j.ijdevneu.2014.02.001
14. Sarfati J, Guiochon-Mantel A, Rondard P, Arnulf I, Garcia-Pinero A, Wolczynski S, et al. A comparative phenotypic study of Kallmann syndrome patients carrying monoallelic and biallelic mutations in the prokineticin 2 or prokineticin receptor 2 genes. J Clin Endocrinol Metab (2010) 95:659–69. doi:10.1210/jc.2009-0843
15. Hill JO, Wyatt HR, Peters JC. Energy balance and obesity. Circulation (2012) 126:126–32. doi:10.1161/CIRCULATIONAHA.111.087213
16. Cao Y. Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell Metab (2013) 18:478–89. doi:10.1016/j.cmet.2013.08.008
17. Dormishian M, Turkeri G, Urayama K, Nguyen TL, Boulberdaa M, Messaddeq N, et al. Prokineticin receptor-1 is a new regulator of endothelial insulin uptake and capillary formation to control insulin sensitivity and cardiovascular and kidney functions. J Am Heart Assoc (2013) 2:e000411. doi:10.1161/JAHA.113.000411
18. Sorisky A, Molgat AS, Gagnon A. Macrophage-induced adipose tissue dysfunction and the preadipocyte: should I stay (and differentiate) or should I go? Adv Nutr (2013) 4:67–75. doi:10.3945/an.112.003020
19. Martucci C, Franchi S, Giannini E, Tian H, Melchiorri P, Negri L, et al. Bv8, the amphibian homologue of the mammalian prokineticins, induces a proinflammatory phenotype of mouse macrophages. Br J Pharmacol (2006) 147:225–34. doi:10.1038/sj.bjp.0706467
20. Franchi S, Giannini E, Lattuada D, Lattanzi R, Tian H, Melchiorri P, et al. The prokineticin receptor agonist Bv8 decreases IL-10 and IL-4 production in mice splenocytes by activating prokineticin receptor-1. BMC Immunol (2008) 9:60. doi:10.1186/1471-2172-9-60
21. Kubota T, Kubota N, Kumagai H, Yamaguchi S, Kozono H, Takahashi T, et al. Impaired insulin signaling in endothelial cells reduces insulin-induced glucose uptake by skeletal muscle. Cell Metab (2011) 13:294–307. doi:10.1016/j.cmet.2011.01.018
22. Genders AJ, Frison V, Abramson SR, Barrett EJ. Endothelial cells actively concentrate insulin during its transendothelial transport. Microcirculation (2013) 20:434–9. doi:10.1111/micc.12044
23. Cao Y. Angiogenesis as a therapeutic target for obesity and metabolic diseases. Chem Immunol Allergy (2014) 99:170–9. doi:10.1159/000353254
24. LeCouter J, Ferrara N. EG-VEGF and the concept of tissue-specific angiogenic growth factors. Semin Cell Dev Biol (2002) 13:3–8. doi:10.1006/scdb.2001.0284
25. Guilini C, Urayama K, Turkeri G, Dedeoglu DB, Kurose H, Messaddeq N, et al. Divergent roles of prokineticin receptors in the endothelial cells: angiogenesis and fenestration. Am J Physiol Heart Circ Physiol (2010) 298:H844–52. doi:10.1152/ajpheart.00898.2009
26. Von Hunolstein JJ, Nebigil CG. Can prokineticin prevent obesity and insulin resistance? Curr Opin Endocrinol Diabetes Obes (2015) 22:367–73. doi:10.1097/MED.0000000000000185
27. Guo T, Bond ND, Jou W, Gavrilova O, Portas J, McPherron AC. Myostatin inhibition prevents diabetes and hyperphagia in a mouse model of lipodystrophy. Diabetes (2012) 61:2414–23. doi:10.2337/db11-0915
28. Kolka CM, Bergman RN. The endothelium in diabetes: its role in insulin access and diabetic complications. Rev Endocr Metab Disord (2013) 14:13–9. doi:10.1007/s11154-012-9233-5
29. Arora H, Boulberdaa M, Qureshi R, Bitirim V, Gasser A, Messaddeq N, et al. Prokineticin receptor-1 signaling promotes epicardial to mesenchymal transition during heart development. Sci Rep (2016) 6:25541. doi:10.1038/srep25541
30. Urayama K, Guilini C, Turkeri G, Takir S, Kurose H, Messaddeq N, et al. Prokineticin receptor-1 induces neovascularization and epicardial-derived progenitor cell differentiation. Arterioscler Thromb Vasc Biol (2008) 28:841–9. doi:10.1161/ATVBAHA.108.162404
31. Urban JD, Clarke WP, von Zastrow M, Nichols DE, Kobilka B, Weinstein H, et al. Functional selectivity and classical concepts of quantitative pharmacology. J Pharmacol Exp Ther (2007) 320:1–13. doi:10.1124/jpet.106.104463
32. Choke E, Cockerill GW, Laing K, Dawson J, Willson WRW, Loftus IM, et al. Whole genome-expression profiling reveals a role for immune and inflammatory response in abdominal aortic aneurysm rupture. Eur J Vasc Endovasc (2009) 37:305–10. doi:10.1016/j.ejvs.2008.11.017
33. Duncan ER, Crossey PA, Walker S, Anilkumar N, Poston L, Douglas G, et al. Effect of endothelium-specific insulin resistance on endothelial function in vivo. Diabetes (2008) 57:3307–14. doi:10.2337/db07-1111
34. Tabit CE, Shenouda SM, Holbrook M, Fetterman JL, Kiani S, Frame AA, et al. Protein kinase C-beta contributes to impaired endothelial insulin signaling in humans with diabetes mellitus. Circulation (2013) 127:86–95. doi:10.1161/CIRCULATIONAHA.112.127514
Keywords: prokineticin, G protein-coupled receptors, obesity, diabetes, anorexic, angiogenic hormones
Citation: Nebigil CG (2017) Prokineticin Is a New Linker between Obesity and Cardiovascular Diseases. Front. Cardiovasc. Med. 4:20. doi: 10.3389/fcvm.2017.00020
Received: 02 March 2017; Accepted: 27 March 2017;
Published: 12 April 2017
Edited by:
Ichiro Sakuma, Hokko Memorial Hospital, JapanReviewed by:
Naoki Ishimori, Hokkaido University Hospital, JapanKoichi Okita, Hokusho University, Japan
Masato Furuhashi, Sapporo Medical University, Japan
Copyright: © 2017 Nebigil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Canan G. Nebigil, bmViaWdpbEB1bmlzdHJhLmZy