 W. Douglas Robinson
W. Douglas Robinson Maria Nanau
Maria Nanau William Kirsch
William Kirsch Caleb T. Centanni
Caleb T. Centanni Nolan M. Clements
Nolan M. Clements- Department of Fisheries, Wildlife and Conservation Sciences, Oregon State University, Corvallis, OR, United States
Evening Grosbeak (Coccothraustes vespertinus) is a species of North American Fringillid finch thought to be one of the fastest declining songbirds across North America. It has been divided into five groups, potentially distinct lineages, based in part on structure of their flight calls. The primary flight calls of each type exhibit structural variation that has not been described and the degree to which that variation might lead to identification errors has not been quantified. We describe the variation in call structure of type 1 Evening Grosbeaks recorded at a spring migratory stopover site (Corvallis, Oregon) and nearby areas in the Pacific Northwest, USA. We recorded grosbeaks weekly from April through early June 2023. We reviewed more than 10,000 recorded call notes to characterize the variety of calls and their configurations. We found a high diversity of call notes including at least 11 recurring readily identifiable variants of the primary flight calls, all of which were attributed to individuals thought to be type 1 birds. Geographically, the nearest neighbors of type 1 Evening Grosbeaks are type 2s, which have uncommonly been recorded in our study area but whose breeding range appears to overlap that of type 1 in southern Oregon. We quantified recordings of type 2 flight calls and compared them with type 1 flight calls, finding that linear discriminant function analyses correctly identified >95% of recordings to type. Inclusion of a metric of asymmetry in call shape improved correct classification to 98.5%. We also found that the other dominant calls given by both types, buzzy trills, could be identified correctly to type with a high level of confidence. The sufficiently different characteristics of flight calls and trills indicate that types 1 and 2 are identifiable spectrographically in most cases, providing confidence that the geographic distribution and migratory movements of call-type populations, despite being essentially identical in plumage, can be documented effectively by characteristics of call notes.
1 Introduction
Mobile organisms pose challenges for understanding their ecology and evolution as well as their conservation (Robinson et al., 2010; Bridge et al., 2011). The intersection of ecology with conservation policy is also affected as policies often depend on operational designations of evolutionary lineages such as species, subspecies and evolutionarily significant units (Moritz, 1994; Haig et al., 2006; Winker, 2010). When such lineages are difficult to recognize because of morphological similarity with other lineages, tools that improve our ability to study and monitor those lineages are needed. Variation among infraspecific groups in vocalizations has been used to characterize distributional patterns and habitat choice (Groth, 1993; Parchman et al., 2006; Martin et al., 2020). A particular challenge arises when birds engage in irruptive movements, occurring in different geographic regions from year to year on an unpredictable schedule. Tracking the movements of such cryptic infraspecific populations and studying details of variation in calls can be facilitated through collaborative data collection efforts such as the growing enterprise of citizen scientists recording bird calls and archiving them in public online databases (Sullivan et al., 2009; Pérez-Granados, 2023).
Many species of Fringillid finches give distinctive call notes during flight. Such flight calls, when they are shared within populations or subspecies and are different from flight calls in other infraspecific groups, are referred to as call types (Groth, 1988). Interest in the diversity of call types within species has increased in recent decades, particularly after intensive study of Red Crossbill (Loxia recurvirostra) flight call variation led to identification of more than two dozen call-type groups across their Holoarctic geographic range (Groth, 1993; Parchman et al., 2006; Martin et al., 2019; Martin et al., 2020). Exploration of the diversity of flight calls given by other finch species has uncovered further infraspecific variation across the geographic distributions of those species. In some cases, such as Evening Grosbeak (Coccothraustes vespertinus; Figure 1), the distinctive flight call types map approximately onto the geographic ranges of subspecies described previously (Grinnell, 1917; Sewall et al., 2004). Those subspecies designations were based largely on slight differences in morphological characteristics such as plumage coloration and bill size. When plumages vary among subspecies in subtle ways not readily diagnosable during field observations but calls of subspecies vary diagnostically, the possibility of mapping movements across geography and time by recording calls creates opportunities to learn more about the dynamic movements of call-type populations.

Figure 1 Evening Grosbeaks are currently divided into five call types whose geographic ranges have been mapped approximately but where the exact locations of range boundaries remain unknown. In particular, the dynamics of year to year variation in geographic ranges of these often highly irruptive call-type populations have yet to be studied carefully. Illustration by Tara Kate Designs.
Five subspecies of Evening Grosbeak aligning approximately with five flight call types have been described (Sewall et al., 2004). The structure of each call type, as displayed in spectrograms, appears to be consistently different from other call types (Sewall et al., 2004). However, the degree to which the primary flight call notes vary structurally within each call type has yet to be quantified. Within-call-type variation is important to quantify because 1) most finch species with infraspecific call type diversity utter more than the stereotypical flight calls, so recognition of the range of vocabulary elements finches possess may ensure that call types are identified correctly based on examination of the most informative call elements; and 2) studies of call type variation in Red Crossbills revealed that many birds can be correctly typed based on examination of flight call characteristics, but geographic variability in the structural characteristics of flight calls is non-trivial suggesting accurate identification of call types can sometimes be difficult and nuanced (Tanttu et al., 2006). In the case of Evening Grosbeaks, their vocabulary is incompletely described and the variation in structure of their primary flight calls has not yet been fully explored.
We focused our analysis on type 1 Evening Grosbeaks, a group known to breed primarily in the Pacific Northwest and to wander widely across western North America, overlapping in occurrence with at least three of the remaining four call types, especially type 2 in the Sierra Nevada Mountains of California (Gillihan and Byers, 2020). Critical evaluation of variability in type 1 calls and comparisons with type 2 calls, however, are needed to minimize identification errors for several reasons. First, we still have a lack of clarity regarding location of the geographic range boundary between types 1 and 2, the extent to which each range overlaps and the degree of dynamism across years in those range boundaries. Second, type 1 birds are known to wander widely across western North America and overlap extensively with type 2 birds during the non-breeding seasons. Third, type 2 birds have been detected increasingly north of their traditional Sierra Nevada Mountains geographic range. Our objectives were to: 1) describe the variety of call notes given by Evening Grosbeaks by recording them during their annual spring migration through our study area; 2) characterize the range of variation in the primary flight calls of type 1 birds; 3) assess the degree to which their call notes might be confused with calls of type 2, the only other type known to co-occur in the Pacific Northwest USA; 4) determine if other call notes aside from the primary flight calls may be used to distinguish type 1 from type 2 birds; and 5) determine the level of confidence with which calls may be used to track movements across geography and time.
2 Methods
2.1 Study area
The campus of Oregon State University in Corvallis, Oregon (44.56570, -123.27890), is a well-known spring migratory stopover site where Evening Grosbeaks forage on tree seeds for several weeks before continuing their migration to breeding sites (Robinson et al., 2022). In the 1970s, 150,000 to 250,000 birds gathered on campus to consume elm seeds but in recent decades the numbers have declined to 10,000 or fewer birds. We surveyed campus two to four times per week from late March through late May, 2023, by walking 5 km of transects routed along paths passing by the 140 elm trees growing on campus. We also augmented our Corvallis recordings by searching nearby forests in the Coast Range mountains of Benton County, adjacent to Corvallis. As grosbeaks are social and noisy birds, we listened for their calls to locate them. Once discovered, we counted them (see Robinson et al., 2022 for further details) and recorded them.
2.2 Sources of recordings
Recordings were made by positioning ourselves beneath trees containing perched Evening Grosbeaks. Flying flocks were also recorded when possible. When birds were perched, we aimed to obtain 1-2 minutes of recordings per flock. We used several types of recorders, including microphones built into smart phones (Apple iPhone XR and Samsung Galaxy) running the Voice Record Pro app, Zoom H5 Handy recorder and a Sennheiser ME-420 with parabolic mic. The same devices were also used to record birds discovered during ad hoc surveys across Corvallis and the nearby Coast Range woodlands in western Oregon. All recordings were archived in the Macaulay Library (https://www.macaulaylibrary.org) in association with eBird checklists. We also downloaded recordings from eBird/Macaulay Library and quantified calls in recordings made by other observers across western Oregon. Type 2 calls were downloaded from Macaulay Library from sites in Oregon and the Sierra Nevada Mountains, California.
2.3 Data analysis
We used Raven Pro (Cornell Lab of Ornithology, 2023) to view spectrograms of each recording and explore the diversity of calls made by Evening Grosbeaks in our study area (Figure 2). We categorized calls as primary flight calls, trills and other calls. We then visually sorted repeating patterns we observed among the primary type 1 flight calls to capture the range of variation in those calls. We visually inspected spectrograms of more than 10,000 calls. We did not attempt to count exactly the number of calls reviewed. The number of calls per second in the 160 recordings we inspected ranged from 0.2 to 10, across the 250 minutes of recordings. Next, we grouped calls into categories based on subjective assessment of recurring structural shapes. Subsequently, we randomly selected 40 recordings for type 1 and type 2 birds then chose at random within each of those recordings a flight call to measure quantitatively. We measured maximum and minimum frequency (Hz), difference between the maximum and minimum frequency (Hz), duration (s) and asymmetry of call shape. Asymmetry was a proportion where the difference between the maximum frequency of each call and the initial starting low frequency was divided by the difference between the maximum frequency and terminal low frequency. Calls with shorter initial elements had lower ratios than calls with initial and terminal elements that were similar in frequency span. We used the same process for selection of trills and measured the same characteristics (except for asymmetry). We also included a characteristic unique to trills, a count of the number of frequency peaks in each trill call, which indexed the ‘speed’ of the trills (number of peaks/second). Other calls were rare so we did not quantify their structures.
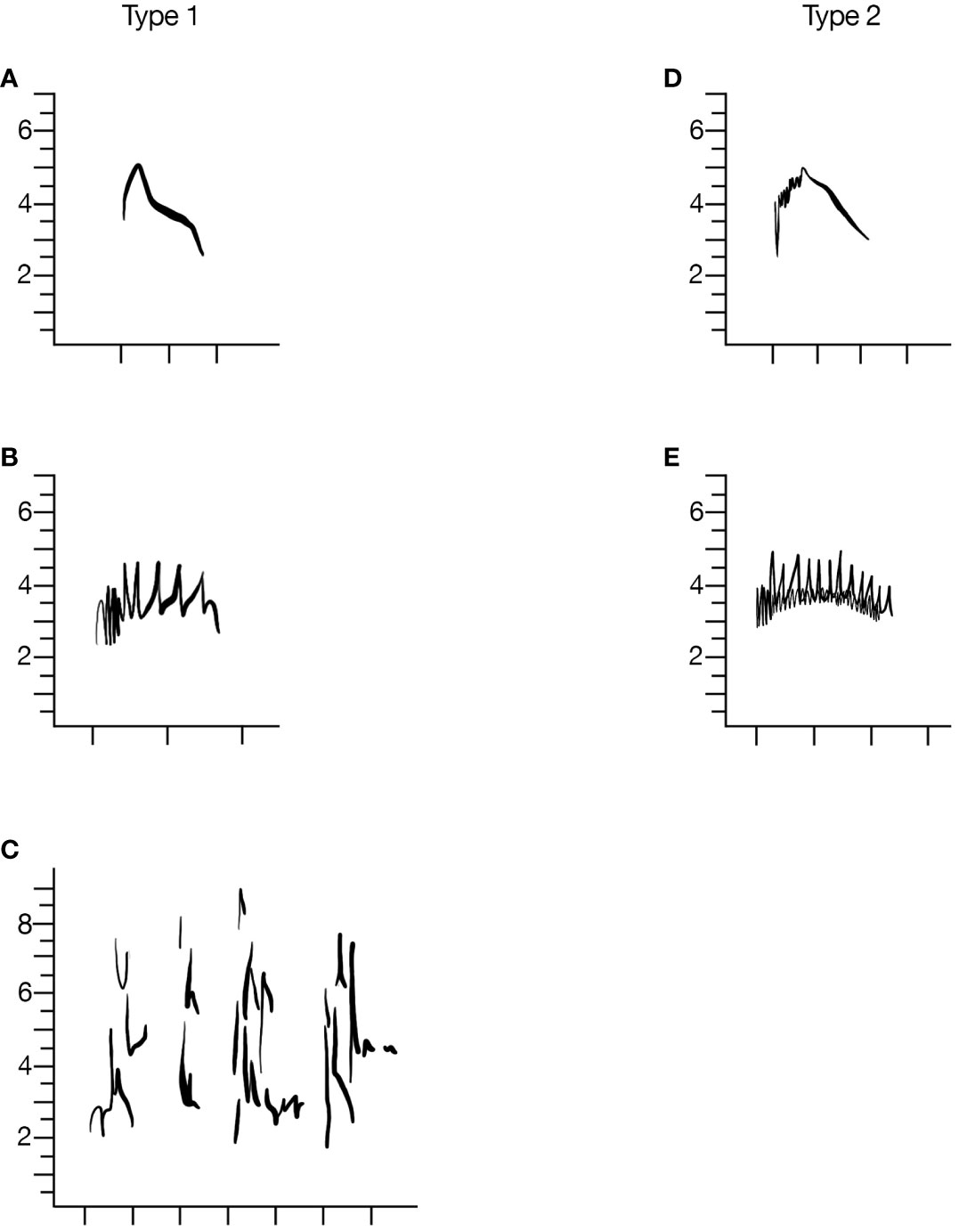
Figure 2 Evening Grosbeaks have two major categories of call types, including flight calls and trills. Left column of spectrograms are all Type 1 calls and right column is Type 2. Examples of typical flight calls (A, D) and trills (B, E) are depicted. Additional calls, such as the click of type 1 birds (C) were rarely encountered. X-axis is time and tick marks are spaced at 0.1 sec intervals.
We compared characteristics of type 1 and type 2 calls to determine the degree to which the variability of calls in each call type might lead to inaccurate identifications. To do so, we compared with one-way analysis of variance the duration, low and high frequencies and the change in frequencies of flight calls and trills, plus the number of trills per unit time in trill calls and our measure of asymmetry. We used discriminant function analysis (DFA) to assess accuracy of identifications of each set of calls to type. In the DFA, we explored the combinations of our measurements that allowed the highest accuracy of identifications, utilizing the linear, common covariance approach. We used all possible combinations of our variables with exceptions. High and low frequencies were uncorrelated but low frequencies were negatively related to delta frequency (high minus low frequency within each call) and high frequencies were positively related to delta frequency. Therefore, we did not combine low or high frequency measurements with delta frequency in our DFA. All analyses were conducted with JMP (JMP, 2018).
3 Results
We observed two main categories of call notes in Evening Grosbeaks in our study area (Figure 2). Primary flight calls and trills composed almost all of the calls but a variety of other rare vocalizations, which we did not characterize further, were recorded.
We recognized 11 recurring shapes of type 1 flight calls (Figure 3). These corresponded to 3 of the call shapes categorized by Sewall et al. (2004) plus 8 other variants. The primary flight calls were dominated by descending constrained whistles lasting approximately 0.15 sec, initiated at a mean high frequency of 4625 Hz and ending at mean frequency of 2540 Hz. Typically, the flight calls descended and included an inflection near the midpoint of the calls where the rate of decline in the frequency slowed briefly. Elaborations of the simple descending note were responsible for the range of variation we observed. Most often a brief (less than 0.05 sec) upward inflection was appended immediately prior to and continuous with the main descending call note. Extensions of the duration of the midpoint frequency stabilization were noted in many variations and other calls were structured with longer initial sections that rapidly modulated frequency in a narrow band and so resembled brief trills. Occasionally, we observed calls we categorized as flight calls that included similar initial sections but largely lacked or had greatly reduced down-slurred whistles, which caused the images to appear spectrographically similar to typical flight calls of type 2 birds (Figures 3J, K).
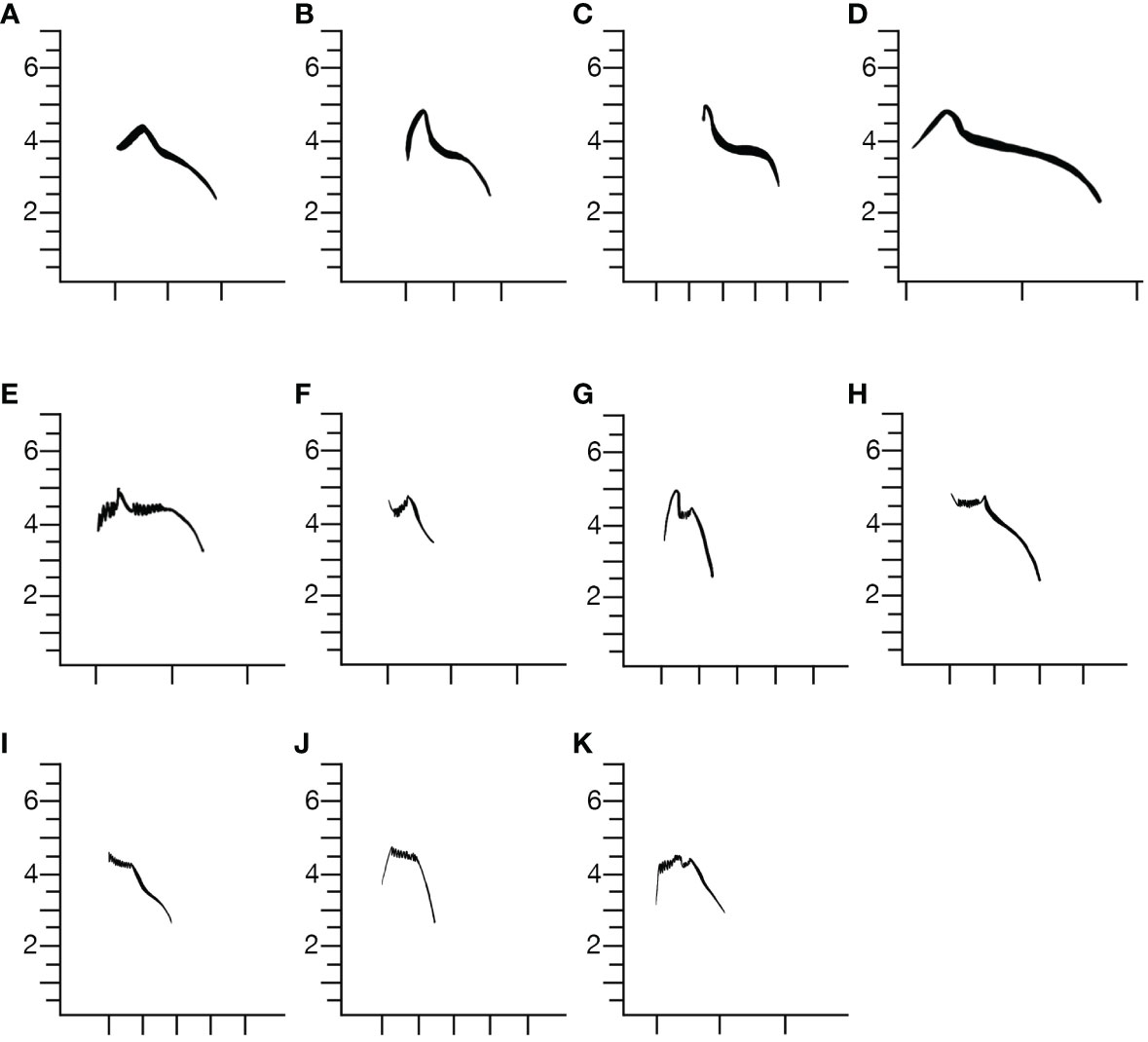
Figure 3 Variation in Type 1 flight calls revealed at least 11 repeated variants within the approximately 10,000 calls we reviewed. Of the 11 variants shown, (A–G) were common while (H–K) were rare. X-axis is time and tick marks are spaced at 0.1 sec intervals.
When statistically comparing the type 1 and type 2 flight calls, all measurements overlapped (Table 1). A comparison of means, however, showed individual type 1 calls were briefer in average duration, spanned a wider range of frequencies, and extended to higher and lower frequencies than type 2 calls (Table 1). Type 1 flight calls were also more asymmetric in shape than type 2 calls. The initial upward element of type 1 calls begins at a higher frequency (and is often completely missing) than the initial element of type 2 calls but both end at similar frequencies as the final elements terminate. Overall, despite extensive overlap in most characteristics of the type 1 and type 2 primary flight calls, they were statistically significantly different, particularly with respect to duration and asymmetry of shape.

Table 1 Duration (s), frequency and asymmetry characteristics of the primary flight calls of type 1 and type 2 Evening Grosbeaks.
Trills were also quite variable within type 1 birds (Figure 4). Comparing structure of trills with type 2 birds, we found patterns similar to those when comparing flight calls except for duration (Table 2). Mean duration differed by only 0.01 sec. Frequency peaks during each cycle of trill calls also overlapped between type 1 and 2 birds but type 2 birds averaged 3 more peaks per unit time and so had faster trills. The frequency range of individual type 2 trill calls had lower low and higher high frequencies than those of type 1 trills, which also produced a wider difference in low to high (delta) frequencies, on average, within type 2 trills.

Figure 4 Trills of Type 1 birds (A–I) were also quite variable in duration, number of frequency peaks per unit time, and frequencies spanned but were largely distinct from trills of Type 2 birds (e.g., Figure 2), which averaged more peaks per unit time. X-axis is time and tick marks are spaced at 0.1 sec intervals.
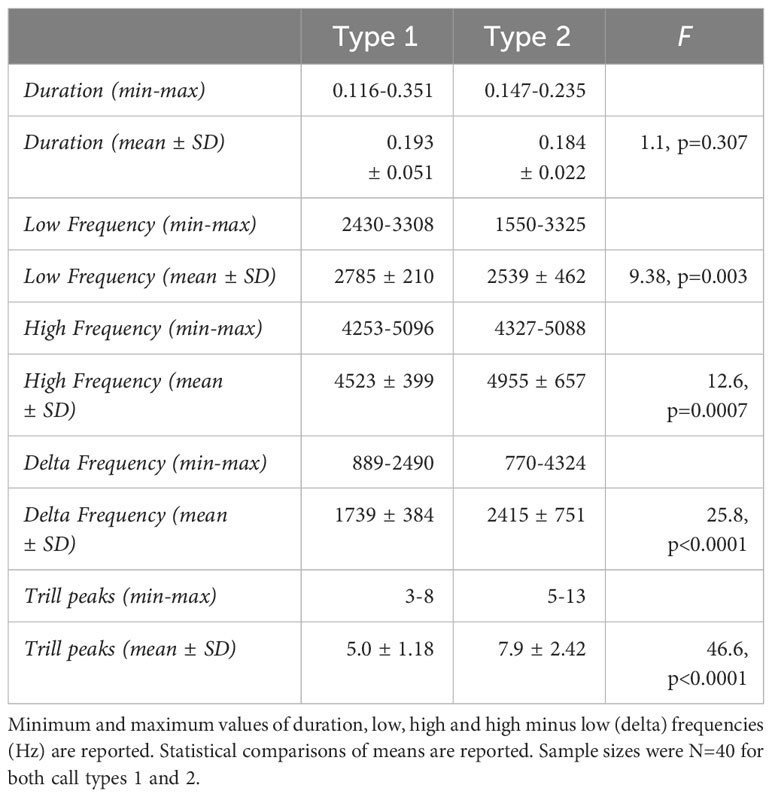
Table 2 Duration (s) and frequency characteristics of the trills of type 1 and type 2 Evening Grosbeaks.
Discriminant function analyses revealed misclassification rates as high as 55% for flight calls and 42% for trills when single characteristics were inspected (Table 3). Dramatic reduction of misclassification errors was achieved when multiple call components were combined. Misclassification of flight calls to correct call type were lowest (1.3%) when duration was combined with asymmetry. If asymmetry information was excluded, misclassification rates based on only frequency and duration measurements were still robust at 2.5-5%. Using duration alone, only 3.8% of flight calls were misclassified to call type. Trills were more often incorrectly classified to call type based on our set of characteristics but the combination of number of trill peaks, duration and delta frequency produced the lowest misclassification rate (8.8%).
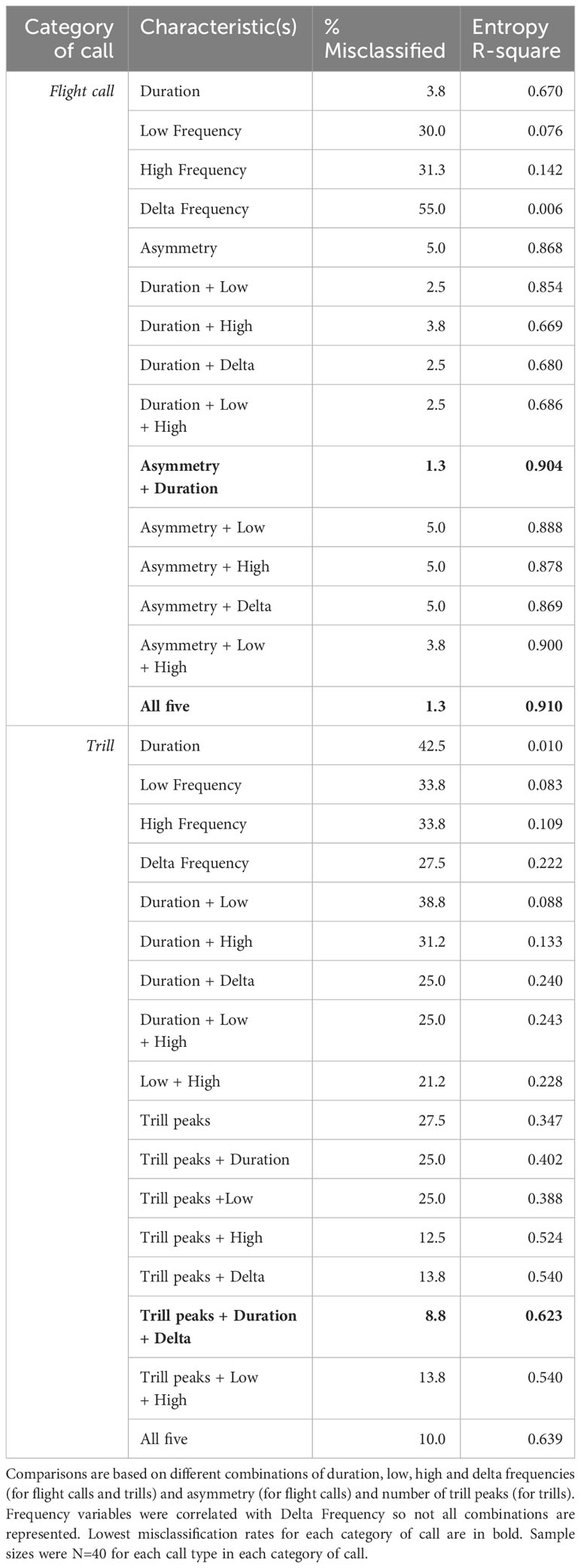
Table 3 Rates of correct vs incorrect classifications of Evening Grosbeak call types as determined from discriminant function analysis.
4 Discussion
Type 1 Evening Grosbeaks have primary flight calls composed of a core element, a descending, frequency-inflected whistled teew. Deviations from the core element are diverse and include addition of initial ascending elements, addition of brief (<0.03 sec) bursts of rapidly frequency-modulated notes prefacing the downward whistle and reduction of the differences between the maximum and minimum frequencies within notes. Despite the substantial variation in these notes, primary flight calls produced by type 1 birds were accurately separated from the most likely other call type to occur with them in our region, type 2, in 98% of cases. Furthermore, we discovered that trills, calls commonly given by both types 1 and 2 Evening Grosbeaks, were identified accurately to type based on a small set of measured characteristics in 91% of cases. Although all characteristics of flight calls and trills that we measured overlapped extensively among the two types, average differences and differences of multiple characteristics assessed in combination permit accurate identification to type in nearly all cases. The key differences were duration, which averages shorter in type 1 birds, and asymmetry in call shape. The first half of most type 1 calls spans a narrower range of frequencies than the second half whereas in type 2 calls the first and second halves of primary flight calls appear to span similar frequency ranges quite consistently.
Several aspects of vocal characteristics require further study to build our understanding of sources of variation in call structures and potential additional opportunities or limits to application of audio recordings for tracking grosbeaks by call type. First, we analyzed recordings but rarely knew additional information about identity of the calling birds such as sex or age. Careful observation of specific individuals might reveal relationships we could not detect. We did note a small tendency for frequency characteristics of type 2 flight calls and trills to be bimodal, which could possibly relate to sexual dimorphism. We did not see evidence of bimodal distributions in type 1 birds.
Second, the question of whether primary flight calls are limited to flying birds or also given by perched birds remains to be rigorously answered. Anecdotally, we observed that the full range of variants of flight calls in type 1 birds were recorded in the large foraging (perched) flocks at our primary study site. We consider it possible that flying birds give a more limited set of calls while commuting but note that sample sizes of flying birds may be naturally smaller because birds quickly move out of range of microphones. Again anecdotally, the most frequently heard type 1 flight call variant at our study site was the pure descending whistle (Figures 3B, C), which is quite different from any type 2 call. We have no evidence type 2 birds ever give descending whistles like that type 1 call. This observation, if supported by additional quantitative analysis, would indicate that separation of type 1 from type 2 birds when detected as fly-overs, for example, with automatic recording units should not cause concern about misclassification errors.
We found that a limited and easily measured set of variables aids in the correct classification of more than 98% of primary flight calls and 90% of trills to type 1 versus type 2. We are not aware of any confirmed records in our study area of the other three types reported to occur in North America. Nevertheless, additional quantitative comparisons of the range of variability in types 3 and 4, especially, should be conducted as type 1 may co-occur at least in some years with each. Additional quantitative analysis of variation of types 2 versus 4 should be conducted as those types appear to be the most similar (Sewall et al., 2004). Given evidence that both types 3 and 4 wander widely across the boreal and eastern North America (type 3) or the Rocky Mountains (type 4) correct classification of those types will be enhanced by a fuller understanding of the diversity of calls those types produce. Overall, our evidence from analysis of variability in types 1 and 2 aligns with the previous conclusion that the four main types in North America should be diagnosable with confidence (Sewall et al., 2004) but we encourage additional quantitative comparisons. Such analyses may reveal a previously unappreciated diversity of variants in flight calls and, as we found, that other calls (such as trills) given by each call type may also provide additional evidence for correct identification to call type.
In conclusion, accurate identification by sound provides confidence that these different populations, otherwise inseparable in the field based on morphology alone, can be identified and tracked through time and across their ranges with a high level of confidence. The rapidly increasing number of recordings collected by community scientists and archived in public databases such as the Macaulay Library (eBird) and xeno-canto.org (Figure 5), provides an unparalleled opportunity to monitor the movements and map the geographic ranges of Evening Grosbeaks. Type 1 grosbeaks, in particular, appear to engage in fairly consistent spring migration patterns, appearing in Pacific Northwestern woodlands each spring (Robinson et al., 2022). However, dynamism exists in their patterns of migratory movement and selection of breeding range, neither of which have been well quantified. We conclude that confidence in correct identifications from audio recordings and the widespread coverage of western North America by birders contributing recordings should help reveal in finer detail the irruptions and interannual variation in distribution of type 1 birds. Such data are a first step toward unraveling potential causes of distributional shifts of type 1 populations. Improvements in knowledge of movements, causes of irruptions and drivers of population fluctuations will also inform on-going efforts to conserve and restore populations of Evening Grosbeaks, such as with the Road 2 Recovery Project in Canada and the United States. To our knowledge, aside from the widely studied Red Crossbills, Evening Grosbeaks are among a unique group of birds that can now be tracked by call type. Given that each call type of Evening Grosbeak may be directly linked to subspecific status and each appears to largely occupy a distinct geographic region for breeding, monitoring to inform conservation and management of the different populations appears feasible.
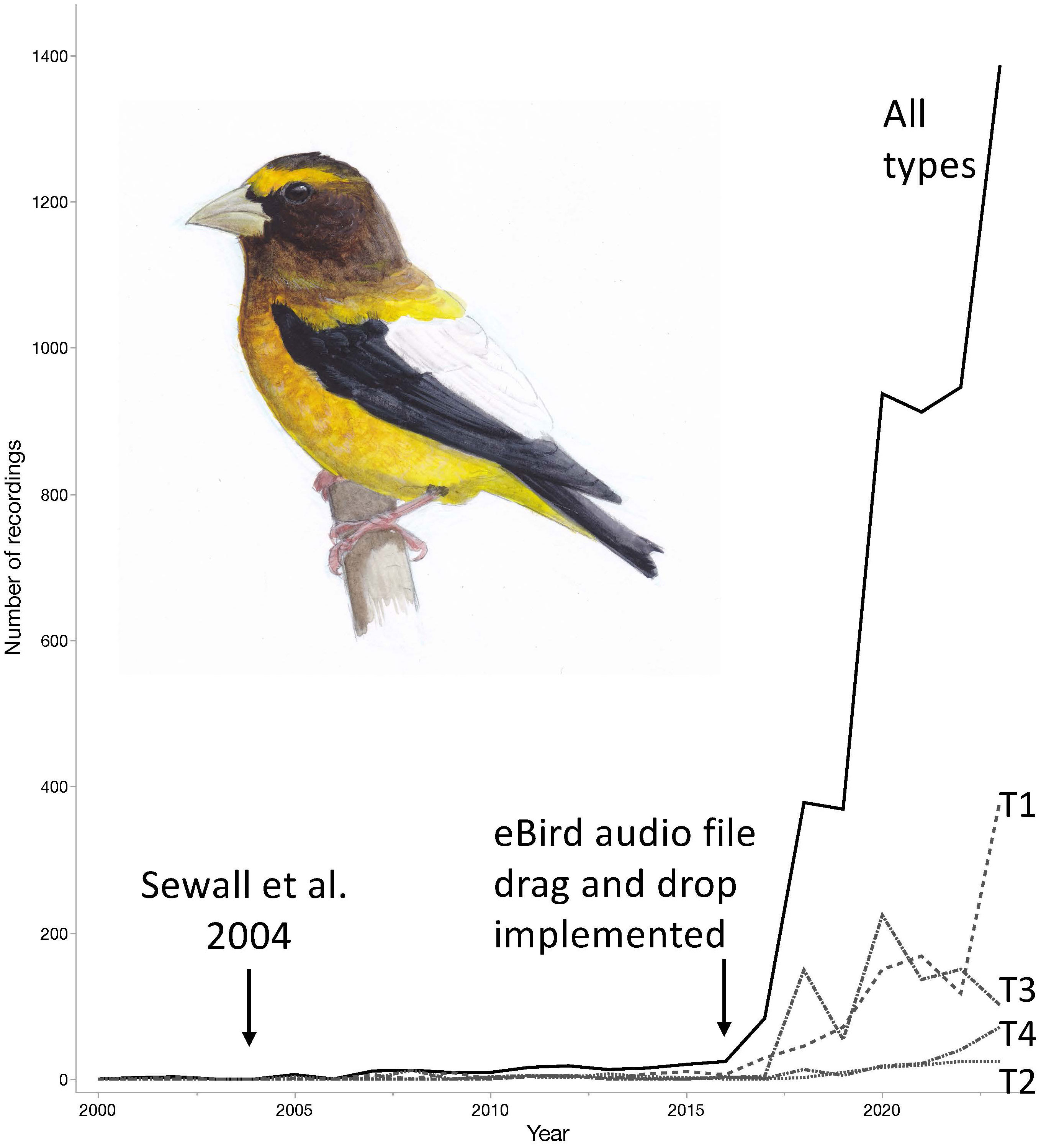
Figure 5 The availability of sound recordings of Evening Grosbeaks has increased dramatically since the publication of Sewall et al. (2004). The increase accelerated when eBird implemented drag and drop tools for contributors to add sound recordings to their eBird checklists. The recordings accumulated in the Macaulay Library much faster after that time. Differences across the call types (T1, T2, etc) are inconsistent with type 1 being the most frequently recorded and type 5 (not shown) containing less than 10 total recordings as of 2023. Illustration of Evening Grosbeak: © 2023 Jack Hobe | www.jackhobe.com.
As our confidence in the utility of call types for tracking movements of infraspecific-level groups grows, the variety of ecological questions we can address increases. How dynamic are the geographic ranges of each call type from year to year and how much overlap occurs? Are call-type groups simply geographically separated subdivisions or are they potentially also divided into ecotypes specializing on particular food resources as at least some Red Crossbill call types do (Centanni et al., 2024)? Are the call-type groups cohesive lineages, perhaps even cryptic species, or do mixed-group pairings occur at geographic range boundaries? Which call-type groups are the most irruptive and what drives the irruptions? Knowledge of the biology of this species of conservation concern can improve rapidly using audio recordings to achieve identification to call-type populations, putatively subspecies, in the field.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because Observational study only; no handling.
Author contributions
WR: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MN: Data curation, Funding acquisition, Investigation, Methodology, Writing – review & editing. WK: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Writing – review & editing. CC: Data curation, Investigation, Writing – review & editing. NC: Data curation, Investigation, Methodology, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Support was provided by the Bob and Phyllis Mace Watchable Wildlife Professorship (WR), the College of Agricultural Sciences Beginning Researcher Program (MN) and the Finch Research Network (WK).
Acknowledgments
We thank the many contributors to eBird and xeno-canto who augmented our recordings with their own. Undergraduates at Oregon State University (especially A. Cemy) and one high school student (S. Neal) joined our field trips and participated in data collection. We thank Tara Kate Designs for Figure 1 and for constructing Figures 2-4 for us. Jack Hobe shared his Evening Grosbeak illustration for Figure 5.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bridge E. S., Thorup K., Bowlin M. S., Chilson P. B., Diehl R. H., Fléron R. W., et al. (2011). Technology on the move: recent and forthcoming innovations for tracking migratory birds. BioScience 61, 689–698. doi: 10.1525/bio.2011.61.9.7
Centanni R., Robinson J., Young R. (2019). Is resource specialization the key? Some, but not all Red Crossbill call types associate with their key conifers in a diverse North American landscape. Front. Bird Sci. 3, 1321583. doi: 10.3389/fbirs.2024.1321583
Cornell Lab of Ornithology (2023). Raven Pro: Interactive Sound Analysis Software (version 2.0) (Ithaca, New York: Cornell Lab of Ornithology).
Gillihan S. W., Byers B. E. (2020). “Evening Grosbeak (Coccothraustes vespertinus), version 1.0,” in Birds of the World. Eds. Poole A. F., Gill F. B. (Cornell Lab of Ornithology, Ithaca, New York, USA). doi: 10.2173/bow.evegro.01
Grinnell J. (1917). The subspecies of hesperiphona vespertina. Condor 19, 17–22. doi: 10.2307/1362452
Groth J. G. (1988). Resolution of cryptic species in appalachian red crossbills. Condor 90, 745–760. doi: 10.2307/1368832
Groth J. G. (1993). Evolutionary Differentiation in Morphology, Vocalizations, and Allozymes Among Nomadic Sibling Species in the North American Red Crossbill (Loxia Curvirostra) Complex. (Berkeley and Los Angeles, California: University of California Press).
Haig S. M., Beever E. A., Chambers S. M., Draheim H. M., Dugger B. D., Dunham S., et al. (2006). Taxonomic considerations in listing subspecies under the U.S. Endangered species act. Conserv. Biol. 20, 1584–1594. doi: 10.1111/j.1523-1739.2006.00530.x
Martin R., Rochefort J., Mundry R., Segelbacher G. (2019). Delimitation of call types of Red Crossbill (Loxia curvirostra) in the Western Palearctic. Écoscience 26, 177–194. doi: 10.1080/11956860.2018.1564483
Martin R., Rochefort J., Mundry R., Segelbacher G. (2020). On the relative importance of ecology and geographic isolation as drivers for differentiation of call types of red crossbill Loxia curvirostra in the Palearctic. J. Avian Biol. 51, 1–15. doi: 10.1111/jav.02358
Moritz C. (1994). Defining ‘Evolutionarily significant units’ for conservation. Trends Ecol. Evol. 9, 373–375. doi: 10.1016/0169-5347(94)90057-4
Parchman T. L., Benkman C. W., Britch S. C. (2006). Patterns of genetic variation in the adaptive radiation of New World crossbills (Aves: Loxia). Mol. Ecol. 15, 1873–1887. doi: 10.1111/j.1365-294X.2006.02895.x
Pérez-Granados C. (2023). BirdNET: applications, performance, pitfalls and future opportunities. Ibis 165, 1068–1075. doi: 10.1111/ibi.13193
Robinson W. D., Bowlin M. S., Bisson I., Shamoun-Baranes J., Thorup K., Diehl R. H., et al. (2010). Integrating concepts and technologies to advance the study of bird migration. Front. Ecol. Environ. 8, 354–361. doi: 10.1890/080179
Robinson W. D., Greer J., Masseloux J., Hallman T. A., Curtis J. R. (2022). Dramatic declines of evening grosbeak numbers at a spring migration stop-over site. Diversity 14, 496. doi: 10.3390/d14060496
Sewall K., Kelsey R., Hahn T. P. (2004). Discrete variants of Evening Grosbeak flight calls. Condor 06, 161–165. doi: 10.1093/condor/106.1.161
Sullivan B. L., Wood C. L., Iliff M. J., Bonney R. E., Fink D., Kelling S. (2009). eBird: A citizen-based bird observation network in the biological sciences. Biol. Conserv. 142, 2282–2292. doi: 10.1016/j.biocon.2009.05.006
Tanttu J. T., Turunen J., Selin A., Ojanen M. (2006). Automatic feature extraction and classification of crossbill (loxia spp.) flight calls. Bioacoustics 15, 251–269. doi: 10.1080/09524622.2006.9753553
Keywords: bird flight call, Coccothraustes, irruptive behavior, sound analysis, spectrogram
Citation: Robinson WD, Nanau M, Kirsch W, Centanni CT and Clements NM (2024) Flight calls and trills of Evening Grosbeaks can be used to map movements and ranges of call types 1 and 2. Front. Bird Sci. 3:1340750. doi: 10.3389/fbirs.2024.1340750
Received: 18 November 2023; Accepted: 30 January 2024;
Published: 13 February 2024.
Edited by:
Michal Budka, Adam Mickiewicz University in Poznań, PolandReviewed by:
Emilia Grzędzicka, Polish Academy of Sciences, PolandGianpasquale Chiatante, University of Tuscia, Italy
Copyright © 2024 Robinson, Nanau, Kirsch, Centanni and Clements. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: W. Douglas Robinson, ZG91Z2xhcy5yb2JpbnNvbkBvcmVnb25zdGF0ZS5lZHU=