 Joana Sá1
Joana Sá1 Simone Sá1
Simone Sá1 Hélène Leménager2
Hélène Leménager2 Raquel Costa1
Raquel Costa1 Brigitte Onteniente2
Brigitte Onteniente2 Raquel Soares3,4
Raquel Soares3,4 Viviana P. Ribeiro1*
Viviana P. Ribeiro1* Ana L. Oliveira1*
Ana L. Oliveira1*- 1Universidade Católica Portuguesa, CBQF-Centro de Biotecnologia e Química Fina–Laboratório Associado, Escola Superior de Biotecnologia, Porto, Portugal
- 2Phenocell S.A.S., Grasse, France
- 3Department of Biomedicine, Faculty of Medicine, University of Porto, Porto, Portugal
- 4i3S, Institute of Research and Innovation in Health, University of Porto, Porto, Portugal
In 2021, approximately 537 million people worldwide, primarily in low- and middle-income countries, were affected by diabetes, leading to approximately 6.7 million deaths annually or severe secondary complications including life-threatening hyperglycemia. For nearly 50 years, current therapeutic approaches include full pancreas transplantation and isolated pancreatic islets, more recently, cell therapy such as in vitro generated islets and stem cell derived. The transplantation of pancreatic islet cells can be less invasive than full organ transplantation, however, does not achieve the same rate of functional success due to the low survival of the engrafted cells. Tissue-engineered bioartificial pancreas has been designed to address such issues, improving cell engraftment, survival, and immune rejection problems, with the added advantage that the tissue produced in vitro has an unlimited source of material.
1 Introduction
Diabetes is a critical public health issue and one of the four key noncommunicable diseases that global leaders are focusing on. The incidence and prevalence of diabetes has been consistently rising over the past several decades. In 2021, approximately 537 million adults, aged between 20 and 79 years, were living with diabetes, representing about 10.5% of the global adult population. That year, diabetes contributed to approximately 6.7 million deaths worldwide (Collaboration, 2010). By 2030, it is estimated that the number of adults with diabetes will increase to 643 million, and by 2045, this number could reach 783 million, meaning that 1 in 8 adults will have diabetes (Saeedi et al., 2019). According to the International Diabetes Federation, the number of people with type 1 diabetes in 2021 was approximately 8.75 million worldwide, and the number of people with type 2 diabetes was approximately 483 million worldwide, being 90% of the total number of diabetics. Additionally, it is estimated that the number of people with type 2 diabetes who are in a more advanced stage of the disease and require insulin treatment is between 20% and 30% of patients with this type of diabetes (IDF, 2025; Ogle et al., 2022; Basu et al., 2018). Diabetes is one of the leading causes of death, being characterized by elevated blood glucose levels that significantly contribute to other severe complications, e.g., cardiovascular diseases and kidney failure (IDF, 2021). This has resulted in significant economic losses for individuals and their families, as well as for health systems and national economies due to direct medical expenses and lost productivity. During pregnancy, poorly managed diabetes can raise the risk of fetal death and other issues (Collaboration, 2010; Skyler et al., 2016). Hyperglycemia occurs when pancreas fails to efficiently produce insulin and the cells in our body become unresponsive to insulin, a condition known as insulin resistance, leading to elevated blood sugar levels (Skyler et al., 2016). This condition can be identified through specific methods, including glucose concentration test in venous plasma; glycated hemoglobin (HbA1c) analysis, which reflects the average levels of glycemia over the past 3 months; fructosamine test, which provides the average blood sugar levels over the past 2–3 weeks. Elevated levels of glucose and markers indicate potential or persistent hyperglycemia (ElSayed et al., 2025a). Consequently, different types of diabetes can arise according to different etiologies, including type 1 and type 2 diabetes, gestational diabetes, genetic predisposition, exocrine pancreatic disorders, and medications. When diabetes is not well managed, complications are developed. Abnormally high blood glucose can have a life-threatening impact if it triggers conditions such as diabetic ketoacidosis in types 1 and 2, and hyperosmolar coma in type 2. Abnormally low blood glucose can occur in all types of diabetes and may result in seizures or loss of consciousness (Atkinson et al., 2015; Roglic Gojka, 2016).
The prevalent type 2 diabetes, often seen in adults, results from insulin resistance or inadequate insulin production, with a notable surge in cases over the last 3 decades. Different pharmacological therapies are available to treat type 2 diabetes, offering benefits aimed at managing blood glucose levels and mitigating the long-term complications associated with the disease. However, difficulties in adhering to lifelong medication regimens, the need for frequent adjustments or combination therapies to maintain blood glucose control are significant challenges (ElSayed et al., 2025a).
Type 1 diabetes is characterized by minimal or no insulin production, resulting from autoimmune destruction of the insulin-producing beta cells in pancreas. In this condition, the exogenous administration of insulin is essential to maintain glycemic control and prevent acute complications (Skyler et al., 2016; Atkinson et al., 2015). Thus, is at level of type 1 diabetes that the greatest challenge arises with more affordable treatments and of higher risk to the patients. Recent technological advancements improved glycemic control and reduced the number of manual insulin injections, i.e., continuous subcutaneous insulin infusion (CSII), continuous glucose monitoring (CGM), and automated insulin delivery (AID) systems. Nevertheless, patients still face the risk of sever hypoglycaemia and diabetic ketoacidosis with the possibility of technical failures, errors in insulin calculation, or unpredictable variations in absorption and metabolism can lead to dangerous metabolic imbalances, requiring constant vigilance (ElSayed et al., 2025b). Pancreas and islet transplantation offer benefits aimed at managing blood glucose levels and mitigating the long-term complications associated insulin administration. However, there is a lack of donors, surgical complications may occur, as well as the risk of organ rejection and the lifelong requirement for immunosuppressive therapy (Aloke et al., 2022).
Thus, cellular therapies will be crucial treatments for diabetes, especially in preventing associated complications and improving patients’ quality of life. These therapies represent a significant advancement in diabetes care, providing an unlimited therapeutic source, offering hope for better glycemic control, preventing cases associated with hypoglycemia, and consequently reducing long-term complications related to poor glycemic control and leading healthier lives for those affected (Abadpour et al., 2021a).
2 Current available therapies for diabetes
Until recently, the primary goal of a diabetes treatment involved to restore the body’s natural ability to regulate blood glucose levels. Nevertheless, current needs in diabetes management have sought the application of more personalized therapies that can meet patients’ specific needs. This is achieved through the application of drugs that enhance insulin sensitivity, or in more complex cases, surgical interventions involving islet cell and pancreas transplantation. This section explores these treatments options, reenforcing that the optimal treatment choice is influenced by a multitude of factors, including diabetes type, patient age, the presence of comorbidities, and individualized treatment objectives (Figure 1).
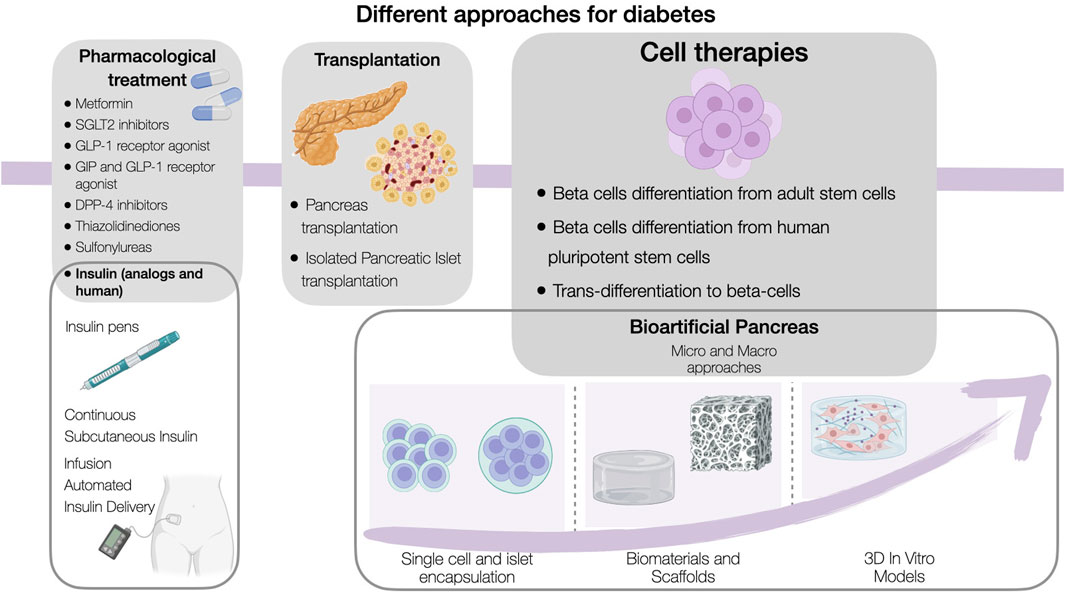
Figure 1. Schematic diagram of the different approaches for diabetes.
2.1 Pharmacological therapies
Antidiabetic pharmacotherapy has evolved significantly in recent decades, offering a wide range of therapeutic options. The selection of the ideal drug depends on several factors, including the drug’s pharmacokinetic and pharmacodynamic profile, individual tolerability, and the presence of comorbidities (ElSayed et al., 2025b; Mudaliar, 2023).
The pharmacological therapy for type 1 diabetes is based only on insulin administration, which is essential for glycemic control. Additionally, pramlintide is introduced as a pharmacological adjuvant, serving as a complementary therapy to insulin that helps improve postprandial glycemic control by reducing significant fluctuations in glucose levels after meals (ElSayed et al., 2025b).
Currently, there are several types of insulin available for managing type 1 diabetes, including different categories tailored to patients’ needs. The main differences between the various types of insulin lie in their onset, peak, and duration of action. Ultra-rapid-acting insulins (lispro, aspart, and glulisine) have an onset of action approximately 15 min after administration, peak at 1 h, and remain active for 2–4 h, making them ideal for immediate postprandial control. Long-acting insulins (glargine and detemir) provide stable glycemic control for up to 24 h, whereas ultra-long-acting insulins, such as degludec, last for more than 24 h, allowing greater flexibility in administration and reduced glycemic fluctuations. Intermediate-acting insulins, such as NPH, act within 2–4 h, peak between 4 and 12 h, and have a duration of 12–18 h, often being used in mixed regimens. Short-acting insulins (regular) have an onset of action at 30 min, peak between 2 and 3 h, and last for 3–6 h, being utilized in specific glycemic control contexts. Finally, inhaled insulins, such as Afrezza, offer a rapid-acting option administered via inhalation, particularly indicated for postprandial glycemic control in selected patients (ElSayed et al., 2025b).
One of the greatest developments and transformations in type 1 diabetes care is the advancement of technologies such as Continuous Subcutaneous Insulin Infusion (CSII), Continuous Glucose Monitoring (CGM), and Automated Insulin Delivery (AID). CSII uses insulin pumps to deliver continuous doses and bolus insulin, ensuring more precise control of the patient’s needs. CGM monitors glucose levels in real time, allowing for quick adjustments and early detection of hypoglycemia or hyperglycemia. Finally, AID systems, which combine insulin pumps and CGM, automatically adjust insulin doses based on glucose readings, providing more efficient glucose control and reducing the need for manual intervention. These advanced technologies significantly improve patients’ quality of life by optimizing glucose control and reducing the risk of complications associated with type 1 diabetes (Marfil-Garza et al., 2021; Larsen, 2004).
Although technologies like CSII, CGM, and AID offer significant benefits in managing type 1 diabetes, they also present some risks and disadvantages. The high cost of these devices and the lack of access in some healthcare systems can be barriers for patients. Technical failures, such as issues with calibration or communication between devices, can affect treatment effectiveness. Additionally, sensor inaccuracies may delay the detection of glucose fluctuations, increasing the risk of hypoglycemia or diabetic ketoacidosis. The complexity of use and potential for human error in device management may also lead to complications, requiring continuous monitoring and proper training to minimize risks. Because of that education on insulin dose adjustment based on carbohydrate intake, physical activity, and glycemic trends is considered crucial. Glucagon is typically prescribed for those at high risk of hypoglycemia, and treatment plans should be regularly reviewed and adjusted (Atkinson et al., 2015; ElSayed et al., 2025b; Diabetes Technology et al., 2025).
In the management of type 2 diabetes, a patient-centered approach is highly emphasized, considering individualized glycemic, weight goals and the possibility of hypoglycemia. Associated comorbidities, especially cardiovascular disease and chronic kidney disease should also be part of the equation. Lifestyle modifications and pharmacotherapy are therefore both important. There are various therapeutic drug classes available, including metformin, sodium-glucose cotransporter-2 (SGLT2) inhibitors, glucagon-like peptide-1 (GLP-1) receptor agonists, gastric inhibitory polypeptide (GIP) and GLP-1 receptor agonists, alpha-glucosidase inhibitors, dipeptidyl peptidase 4 (DPP4) inhibitors, thiazolidinediones, sulfonylureas, and insulin human and analogues (ElSayed et al., 2025b).
The therapeutic plan should be regularly reviewed and adjusted every 3–6 months. Insulin therapy is effective for severe hyperglycemia and can be used in combination with other antidiabetic therapeutic classes (ElSayed et al., 2025a; ElSayed et al., 2025b).
2.2 Pancreas transplantation and clinical islet transplantation
Pancreas transplantation and pancreatic islet transplantation represent innovative approaches in the treatment of type 1 diabetes, with the potential to significantly transform patients’ quality of life. These interventions aim to restore natural insulin production and glycemic regulation, providing alternatives to continuous exogenous insulin use (Marfil-Garza et al., 2021; Larsen, 2004).
Pancreas transplantation is a complex surgical procedure used specially for type 1 diabetes treatment. This procedure involves surgical implantation of a healthy donor pancreas, allowing for the resumption of normal insulin secretion and glucose regulation, and eliminating the need to administrate exogenous insulin (Gruessner and Gruessner, 2013; Rickels, 2012). Pancreas transplants are more often performed together with kidneys, known as simultaneous pancreas-kidney (SPK) transplants. With the best 1-year pancreas graft survival rate, SPK is recommended for DM1 patients and/or with end-stage renal disease (ESRD) (Larsen, 2004; van der Boog et al., 2004; Gupta and Sharma, 2012; Larsen et al., 2002). Pancreas transplantation alone (PTA) or pancreas after kidney transplantation (PAK) are alternative approaches for patients without ESRD or for those who have previously received a kidney transplant (Larsen, 2004; Larson et al., 2004). Despite high 1-year post-transplant patient survival (>95%) and graft (almost 85%) rates, lifelong immunosuppressive therapy is mandatory to prevent graft rejection, with typical regimens including tacrolimus, mycophenolate mofetil, and corticosteroids (Larsen, 2004; Kalluri, 2012). Despite a relatively high success rate, complications such as graft thrombosis, infection, and both acute and chronic rejection remain significant challenges (Humar et al., 2003). Successful pancreas transplantation can lead to insulin independence, improved glycemic control, and reduced long-term diabetic complications, enhancing the patient’s quality of life. However, the procedure requires rigorous candidate selection and ongoing postoperative care to ensure long-term graft function and patient survival (Larsen, 2004).
Langerhans islets are clusters of insulin-producing beta cells in the pancreas. When isolated from donor pancreases and infused into the patient’s liver via the portal vein, they can function as replacement cells to potentially restore normoglycemia. Clinical islet transplantation is an emerging and effective therapy for type 1 diabetic patients who suffer from severe hypoglycemia and glucose instability despite insulin therapy (Marfil-Garza et al., 2021; Cayabyab et al., 2021). Recent studies underscore the importance of islets in glucose homeostasis and their potential to improve outcomes in diabetes treatment (Marfil-Garza et al., 2021; Félix-Martínez et al., 2024; Briggs et al., 2022). Studies indicate that for patients with type 1 diabetes or pancreatogenic diabetes, islet transplantation can stabilize glycemic instability, improve quality of life, and reduce insulin dependence (Rodriguez et al., 2020; Moon et al., 2024), (Langlois et al., 2024). Furthermore, auto islet transplantation after pancreatectomy helps prevent postsurgical diabetes (Arce et al., 2016). The studies also suggest that using islets from a single well major histocompatibility complex (MHC) matched donor may improve graft survival. However, challenges remain, especially in isolating large quantities of islets, with collagenase digestion being a critical limiting factor that needs further research (London, 1995). Although clinical islet transplantation holds promise, its application remains limited to specific patient groups due to its complexity, cost, and the need for donor pancreases. Additionally, the need for long-term immunosuppressant drugs to prevent the body from rejecting the transplanted islets puts patients at a higher risk of infections and other complications. The surgical procedure itself carries risks like bleeding, infection, and pancreatitis. Finally, the limited lifespan of the transplanted islets and the possibility of graft failure require additional treatments and can impact a patient’s quality of life (London, 1995). Despite considerable advancements, pancreas transplantation and islet transplantation remain challenging due to surgical complications, the need for prolonged immunosuppression, and limitations in donor availability. In this sense, new approaches offer potential future advances in diabetes treatment, including the development of new cell therapies, namely, using stem cell-derived sources, cells obtained from reprogramming (Wang et al., 2022; Domínguez-Bendala and Ricordi, 2012), or their combination with different biomaterials or extracellular matrices to recreate some of the pancreatic tissue functionality (Hering et al., 2006; Kroon et al., 2008a; Rezania et al., 2012; Rickels and Robertson, 2019).
3 Cell therapies based in pancreatic beta cells
Cell-based therapies have become a groundbreaking strategy for treating diabetes. Despite the early-stage of the research in pancreatic cell-based therapies, the introduction of stem cells as a renewable cell source to restore the beta cell population has raised attention. Innovative cell-based strategies are focused on creating insulin-producing cells from different cell sources, with the potential to make diabetes treatment more effective. This section focuses on the recent advancements in cell-based therapies designed to achieve insulin producing beta cells for diabetes treatment.
3.1 Trans-differentiation to beta cells
Trans-differentiation involves the re-programming of a differentiated cell lineage into another differentiated cell lineage without the need to pass from a stemness stage. This is possible because the differentiated cells retain the capacity of switching some genes to others resulting in the change of the cells’ phenotype. In case of pancreas, trans-differentiation of liver, stomach and small intestine cells has been established sharing the same lineage origin of pancreatic cells (Okere et al., 2016; Nair et al., 2020).
In vertebrates, liver and pancreatic cells originate from a shared pool of progenitor stem cells located in the extra-hepatic biliary tree. These cells differentiate into hepatic and pancreatic tissues in response to chemical signals released by the developing heart (Meivar-Levy and Ferber, 2019). Thus, several groups have successfully trans-differentiated hepatic cells into insulin-producing cells (Meivar-Levy and Ferber, 2019; Ferber et al., 2000). The first approach involved the recombinant adenovirus-mediated gene transfer to insert pancreatic and duodenal homeobox-1(PDX1) in mice liver, which is a master regulator of pancreas organogenesis and activate insulin production (Ferber et al., 2000). In the meantime, liver to pancreas trans-differentiation evolved being investigated in vitro in rodent, pig human-derived hepatic cells, and primary human liver cultures (Meivar-Levy and Ferber, 2019). Despite the interesting outcomes of these approaches, several limitations came up including the need of using adenovirus for gene transfer or the achievement of hybrid hepatocyte-beta cells phenotypes during culture of these cells which limited the balance between insulin production and glucose levels.
The intestinal and antral stomach are rich in endocrine cells similar to pancreatic beta cells (Ariyachet et al., 2006). Spadoni et al. (Spadoni et al., 2017) indicated that it is possible to reprogramming insulin-producing cells in the small intestine of patients with type 1 diabetes with no evidence of immune rejection. Ariyachet et al. (Ariyachet et al., 2006) also showed that the expression of β-cell reprogramming factors in a diabetic mouse model in vivo effectively converted antral stomach cells into insulin-producing cells with significant molecular and functional similarities to β-cells.
Recently, researchers have focused on alpha cells as a potential source for replacing beta cells in trans-differentiation approaches for diabetes treatment (Saleh et al., 2021). Alpha-cells (glucagon producers) and beta cells (insulin producers) share a common precursor lineage which have shown to simplify reprogramming efforts for insulin-producing cells development. Furuyama et al. (Furuyama et al., 2019) showed that reprogramming alpha-cells with MAFA and PDX1 genes induced these cells to produce insulin while retaining alpha-cells features. Conversely, the deletion of PDX1 in adult mice beta cells resulted in increased alpha-cells and altered islets morphology, suggesting inherent plasticity among alpha- and beta cells. The fact that both cells possess functional similarities in terms of glucose transporters and hormone secretion, as well as anatomic proximity, sharing islet location, blood supply and innervation, may enhance the trans-differentiation process with a minimum impact on cells metabolism. In fact, alpha-cells to beta cells trans-differentiation were reported as naturally occurring in both healthy and diabetic human and mouse pancreatic islets, which holds great potential as a pathway to regenerate beta cells in diabetic patients (Saleh et al., 2021).
3.2 Beta cells differentiation from adult stem cells
Over the years, regenerative medicinal approaches for type 1 diabetes have been actively exploiting adult stem cell-based therapies envisioning the repopulation of the injured pancreatic islets.
Adult mesenchymal stem cells (MSCs), mostly derived from bone marrow, adipose tissue and dental pulp, have good self-renewal and multi-differentiation capacity and possess the ability to secrete immunomodulatory and trophic mediators (Caplan, 2015; Maqsood et al., 2020), rendering them a great potential for cell-based therapies in regenerative medicine. One of the most promising strategies using adult MSCs as a source for pancreatic islets consists in the generation of adult stem cell-derived insulin-secreting beta cells that can be transplanted into patients preventing autoimmune process and restoring beta cell function (Luo et al., 2024). MSCs are prone to be differentiated in vitro, using inductors namely nicotinamide, BMP7 and exendin4, and become insulin productive cells that, when transplanted in vivo, are able to improve glucose tolerance and reduce hyperglycemia (Singh et al., 2023). Additionally, other pre-clinical study conducted with a distinct approach to guide differentiation using agents as activin A, BMP4, nicotinic acid and GSK3b inhibitor, were able to create insulin-producing cells expressing beta cell markers, including insulin, PDX1, and NKX6.1. The resulting IPCs exhibited glucose-stimulated insulin secretion, suggesting functional beta cell activity (Thakur et al., 2020). Besides some challenges regarding the differentiation protocols that vary depending on the cell source and culture conditions, the scientific community is reaching good outcomes with MSC-derived cells, exhibiting a morphological and molecular resemble of the native pancreatic beta cells. However, they are less functional than the native pancreatic beta cells, producing lower levels of insulin and reduced expression of maturation factors, such as MAFA and SIX3 (Nair et al., 2019; Veres et al., 2019a; Karimova et al., 2022).
Nonetheless, interesting findings also arise from adult pancreatic stem cells which are found both inside the islets and in the epithelium of pancreatic ducts (Ramiya et al., 2000; Abraham et al., 2002; Ramiya et al., 2000; Zulewski et al., 2001).
Preclinical studies reveal the existence of a population of cells within islets that express a stem cell-specific marker, nestin, and can differentiate into either a ductal or an endocrine pancreatic phenotype, with a huge capacity for proliferation in vitro. These islet-derived progenitor cells are a distinct population of cells that reside within islets and could be used in the neogenesis of islet endocrine cells (Zulewski et al., 2001; Ji et al., 2022).
Additionally, Ramiya and collaborators successfully generate functional insulin-producing cells by the differentiation of pancreatic stem cells extracted from the ducts, using animal models (Ramiya et al., 2000). Later on, another study focused on the isolation and characterization of pancreas-derived multipotent progenitor cells, found at low frequency (nearly 0.025%) from both islet and ductal isolates derived from adult mouse pancreas. Upon differentiation, the colonies produce distinct populations of pancreatic cells, among them β-like cells with glucose-dependent responsiveness and insulin secretion, constituting a promising candidate for diabetic therapeutic avenues (Seaberg et al., 2004).
Adult stem and progenitor cell-based therapies have made remarkable progress reversing insulin-dependent diabetes after transplantation. However, many challenges remain, being imperative to optimize cell differentiation protocols, other sources of stem cells, explore new strategies to improve cell survival and functionality and to generate cells that meet the quality and safety to be applied in clinical trials.
3.3 Beta cells differentiation from hPSCs
The use of human pluripotent stem cells (hPSCs) in cell therapies is revolutionizing diabetes treatment. Both embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) share the ability to differentiate into the three embryonic germ layers, including insulin-producing pancreatic beta cells (Pagliuca et al., 2014; Kroon et al., 2008b). Moreover, hPSCs possess unlimited proliferation capability in vitro, raising hopes for high-scalable cell therapies (Kropp et al., 2016). The challenges lie in the step-by-step control of the differentiation pathway to ensure production of pure and functional hPSC-derived beta cells, which can be grafted in vivo without triggering immune rejection or teratoma formation (Faleo et al., 2017).
Significant advances in beta cell differentiation protocols have improved the efficiency and functionality of hPSC-derived beta cells while reducing teratoma risk (Pagliuca et al., 2014; Kroon et al., 2008b; D’Amour et al., 2006; Rezania et al., 2014; Veres et al., 2019b; Yoshihara et al., 2020). These protocols generally involve the time-controlled regulation of specific signalling pathways to mimic the embryonic development of the pancreas (Jin and Jiang, 2022). Recent procedures have produced insulin-producing beta cells with proven in vivo efficacy, inhibiting a glucose challenge in 73% of immunodeficient mice 2 weeks after transplantation (Pagliuca et al., 2014). Insulin levels in the bloodstream were maintained 18 weeks after transplantation.
To prevent in vivo immune rejection of the graft, hPSC-based therapies can be combined with systemic immunosuppression, though carrying significant side effects (Petrus-Reurer et al., 2021). Alternative approaches have focused on IFNγ-stimulation of PD-L1 expression, a key determinant of immune tolerance in beta cells, or the encapsulation of the hPSC-derived beta cells using materials such as hydrogels (Yoshihara et al., 2020; Henry et al., 2018; Ma et al., 2016). To date, hESC-derived beta cells launched phase 1/2 clinical trials in three studies (ClinicalTrials.gov). Viacyte’s two trials, started in 2014 and 2017, involved encapsulated hESC-derived pancreatic endoderm (NCT02239354, NCT03163511) (Shapiro et al., 2021; Ramzy et al., 2021; Henry et al., 2018). The trials yielded promising results in terms of safety and efficacy, even though insulin production was insufficient for full insulin independence (Shapiro et al., 2021). Vertex’s trial, initiated in 2021, uses hESC-derived islet cells (NCT04786262).
Although promising, the ethical concerns surrounding the embryonic origin of ESCs limit their widespread utilization (Robertson, 2010). A more ethical alternative resides in iPSC, as they are obtained by reprogramming adult somatic cells and do not involve the destruction of embryos (Takahashi et al., 2007). Additionally, the use of iPSCs in research and clinic is more straightforward than with hESCs (Daley et al., 2016). Finally, generating iPSC from the patient could reduce the risk of immune rejection and enable personalized therapies.
3.4 Maturation and development of functional beta cells
The maturation and development of functional beta cells, essential for insulin production and glucose regulation, is a complex multi-stage process. It begins in the embryonic pancreas, where progenitor cells differentiate into beta cells under transcription factors like PDX1 and NGN3 (Timmons and Boyle, 2022). During fetal development, beta cells start expressing insulin at non-functional levels. In the neonatal period (infants <1 year), they undergo significant changes, enabling insulin secretion at low glucose concentrations and establishing glucose-stimulated insulin secretion (GSIS) (Nair et al., 2020; Barsby and Otonkoski, 2022). In contrast, juvenile beta cells (ages 1–9) behave more like adult beta cells in GSIS but release less insulin, influenced by diet and hormonal changes. They also respond to secretagogues, e.g., sulfonylureas and meglitinides, that stimulate insulin production by the cells and thereby indicating their maturation stage by 1 year after birth (Timmons and Boyle, 2022; Matthews and Wallace, 2005). Nevertheless, there are some key factors that distinguish juvenile beta cells from adult beta cells. They have higher expression of the MAFB marker, which regulates glucose sensing and insulin secretion (Artner et al., 2010). In contrast, most adult beta cells show low levels of proliferative markers and high expression of transcription factors like SIX2 and SIX3, crucial for insulin secretion (Tremmel et al., 2023). Additionally, adult beta cells contain lipid droplets essential for normal insulin secretion, which are absent in juvenile beta-cells. As a result, adult beta cells are considered fully differentiated and the first to glucose stimulation and insulin secretion.
Despite their differentiation degree, adult beta cells exhibit diverse subtypes with functional heterogeneity and plasticity, enabling them to adapt to various physiological needs and metabolic stresses (Rutter et al., 2024). “Hub” cells serve as coordinators within the islet, synchronizing the activity of other beta cell subtypes for stable insulin secretion (Satin et al., 2020). FLTP− beta cells possess high proliferative capacity, essential for regeneration during metabolic changes like pregnancy or obesity. Stress-resistant beta cells are vital for maintaining function under metabolic and immune stress, such as in diabetes (Satin et al., 2020; Benninger and Kravets, 2022). Leader beta cells use distinct calcium signalling pathways to guide other subtypes during insulin secretion, adjusting their response based on the islet’s physiological state (Avrahami et al., 2017).
The functional heterogeneity among the adult beta cells subtypes is crucial for maintaining islet functionality during metabolic stresses and disease states. However, the underlying mechanisms that control interconversion among adult beta cells subtypes remain unclear, hindering the development of effective methods for generating functional beta cells from progenitor stem cells for cell-based therapies (Liu and Hebrok, 2017).
4 Engineering bioartificial pancreas
Engineered bioartificial pancreas have been proposed as innovative therapeutic solutions and modelling systems for diabetes screening and treatment, by integrating cells, biomaterials and advanced technologies. The development of a bioartificial pancreas has emerged as a promising concept for the treatment of insulin-deficient patients, offering a potential solution to overcome the limitations of current treatments (Chen et al., 2025). Furthermore, researchers have been creating biomimicking environments that support the growth and function of beta cells, replicating the pancreatic extracellular matrix for studying disease mechanisms and developing advanced models for diabetes research (Salg et al., 2019).
4.1 Macro- and micro-approaches for beta cells delivery
Two main approaches to beta cell therapies have been developed, namely macro-scale and micro-scale delivery systems (Skyler and Ricordi, 2011; Toftdal et al., 2024). Although the concept of “nanoencapsulation” has been proposed to define the individual encapsulation of cells, we find it less suitable, as cells inherently exist at the micro scale. Therefore, we consider microencapsulation as the individual encapsulation of cells or their encapsulation within a matrix. Following this rational, we focus only on macro and micro-scale encapsulation approaches.
Macro-scale devices are beneficial because they can be removed if safety issues arise. However, their small surface area limits the diffusion of essential molecules like oxygen and insulin, which reduces therapy effectiveness (Abadpour et al., 2021a; Toftdal et al., 2024). Macroencapsulation devices (MEDs) provide a promising treatment for type 1 diabetes by protecting insulin-secreting pancreatic beta cells from the immune system. However, traditional MEDs have low cell capacity and poor nutrient delivery, leading to cell death and decreased insulin output. Convection-enhanced MEDs (ceMEDs) improve nutrient flow and cell survival. Devices like Encaptra, βAir Bio, and the Artificial Pancreas are in trials but still struggle with nutrient transport. Ongoing research aims to improve their long-term effectiveness for diabetes management (Yang et al., 2021).
Micro-scale devices offer a significantly larger surface area compared to macro-scale, enhancing mass transfer and oxygenation for the encapsulated cells. However, these devices are challenging to retrieve and tend to disperse uncontrollably within the body (Abadpour et al., 2021a; Toftdal et al., 2024). M. Hamid et al. compared insulin release from microencapsulated BRIN-BD11 insulin-secreting beta cells to non-encapsulated cells. They found that while encapsulated cells released approximately 60% of the insulin produced by non-encapsulated cells, the functional responses to different stimuli such as glucose and amino acids were comparable, indicating preserved cell functionality. Microencapsulation also protects transplanted cells from immune destruction, suggesting it is a promising approach to improve diabetes treatment by preserving pancreatic beta cell function (Hamid et al., 2001).
Macro and micro-approaches have shown promise, with future perspectives focusing on creating hybrid devices that combine the immune protection of macro-devices with the nutrient exchange efficiency of micro-devices, incorporating biocompatible materials and promoting vascularization to improve graft viability and function in clinical applications (Abadpour et al., 2021b).
4.2 Biomaterials and scaffolds for bioartificial pancreas development
To increase cell therapies’ efficacy, it is necessary to have viable platforms for facilitating cell growing, transporting and implantation, avoiding cell migration from the targeted site. This has been fostering many advancements in the biomaterial field to find solutions that can mimic pancreatic tissue environment.
Several synthetic and natural-based biomaterials have been playing an essential role for beta cells encapsulation, ensuring their long-term viability and acting as an immunoshield against the natural body environment (Table 1) (Toftdal et al., 2024; Kaur et al., 2017).
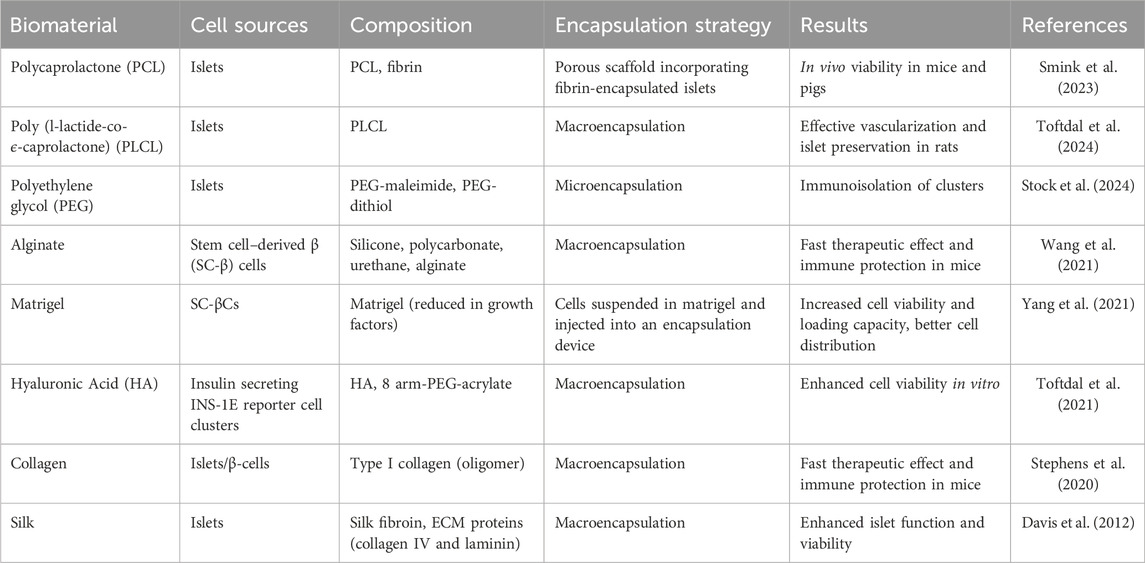
Table 1. Key biomaterials used in recent beta cell encapsulation strategies.
Synthetic-based biomaterials are versatile and offer a tailored control over physicochemical properties of cell-encapsulating materials in terms of porosity, flexibility and stability. Moreover, the inert properties and high reproducibility of synthetic-based biomaterials allows for more efficient cell/islet-encapsulation performances with reduced risks of immune response after encapsulation (Toftdal et al., 2024). PEG hydrogels, known for their immunoprotected properties, create a protective barrier around islets, shielding them from the immune system and promoting long-term survival (Davis et al., 2012; Smink et al., 2018; An et al., 2018). Polycaprolactone (PCL) is widely used in clinical settings due to its favourable degradation rate and biocompatibility. It has also been employed as a scaffold for beta cell transplantation, showing excellent islet survival (Smink et al., 2023; Stock et al., 2024). Finally, poly (L-lactic-co-caprolactone) (PLCL), a co-polymer of PCL and polylactic acid (PLA), offers adjustable degradation and mechanical properties based on the PCL-to-PLA ratio. PLCL is also biocompatible, cost-effective, and holds significant potential for soft tissue engineering (Toftdal et al., 2024). To conclude, these materials have the potential to address the challenges associated especially with islet transplantation, such as immune rejection and graft failure, and improve clinical outcomes for patients with type 1 diabetes (Smink et al., 2018).
Natural-based biomaterials have emerged as promising candidates due to their inherent biocompatibility and ability to mimic the extracellular matrix (ECM) of the pancreas (Toftdal et al., 2024). Alginate, a polysaccharide derived from seaweed, is widely used due to its quick gelation properties, low cytotoxicity, and ability to encapsulate cells effectively (An et al., 2018). Hyaluronic acid (HA), a component of the pancreatic ECM, is another natural material known for its positive influence on cell adhesion, survival, and proliferation (Toftdal et al., 2021; Cañibano-Hernández et al., 2019). Collagen, a major structural protein in various tissues, is also used due to its exceptional biocompatibility and ability to be cross-linked in various ways (Stephens et al., 2020). Silk fibroin is also a promising material for cell therapy, supporting cell growth and differentiation while maintaining its structural integrity and biocompatibility over time (Davis et al., 2012).
4.3 3D in vitro models of diabetes
3D in vitro models are gaining significance in diabetes and cell therapy research. Unlike traditional 2D cultures, these models provide a more accurate representation of the physiological environment, enabling more effective simulation of human body conditions (Li et al., 2020).
In vitro 3D models for diabetes, such as organoids and spheroids, more accurately mimic the structure and microenvironment of pancreatic islets, resulting in better functionality and insulin production by beta cells. These models are valuable for replicating healthy and diabetic states, providing important insights into the progression of diabetes and the effects of potential treatments. Additionally, they have great potential for use in large-scale drug testing, allowing for the evaluation of their efficacy and safety in a more relevant biological context (Mohandas et al., 2023; Zhang et al., 2022). Companies like Readily3D (Printing Industry, 2025) and Aspect Biosystems (Printing Industry, 2025) are at the forefront of this research, developing bioprinted models for diabetes drug testing, which helps in creating more accurate and relevant testing platforms.
Advances in tissue engineering have enabled the development of 3D scaffolds that support the growth and differentiation of stem cells into insulin-producing cells, offering promises for cell replacement therapies in diabetes. The incorporation of extracellular matrix components into these models improves cell survival, proliferation, and function, making them more robust and reliable for research and therapeutic applications (Zhao et al., 2021).
The studies presented by Pagliuca et al., Candiello et al., and Wang et al., have explored various strategies to optimize the culture of islet organoids, aiming to obtain more physiologically relevant models for the study of diabetes and cell therapy (Zhang et al., 2022). Pagliuca et al. demonstrated that suspension culture, using a shaking system, can generate cell clusters with characteristics like native islets (Millman et al., 2016; Zhao et al., 2021). Candiello et al. developed a hydrogel that facilitated the self-aggregation of pancreatic progenitor cells, resulting in an increase in the population of cells co-expressing endocrine differentiation markers (Candiello et al., 2018). Wang et al. used a combined collagen and Matrigel scaffold (C-M scaffold) to promote β-cell maturation and increase insulin production (Tao et al., 2019).
Promising approaches include the generation of pancreatic beta cells from stem cells in 3D cultures, which can be used for transplants in diabetic patients. Organoids, which replicate the multicellular complexity of the pancreas, have been used to study both type 1 and type 2 diabetes, offering an advanced platform for research and treatment development. Progress in 3D in vitro models is leading to a deeper understanding of diabetes and the development of more effective treatments for the disease (Shahjalal et al., 2018).
5 Progress in clinical studies challenges ahead and market expansion
Recent advancements in clinical studies have shown significant progress in human beta cell technology and bioartificial pancreas development. A key focus has been generating functional pancreatic beta cells from hPSCs. These cells can potentially replace damaged beta cells in diabetic patients, offering a more sustainable treatment option. Some clinical studies have also shown that islet transplantation can benefit patients with diabetes, though challenges like limited donor availability and the need for immunosuppressive therapy remain (Berishvili et al., 2024; Raoufinia et al., 2024).
In bioartificial pancreas development, encapsulation techniques have been one of the major research areas. Encapsulation of islets in a semi-permeable membrane protects them from the immune system while allowing the exchange of nutrients and insulin, aiming to reduce or eliminate the need for immunosuppressive drugs (Raoufinia et al., 2024; Hwang et al., 2016; Pareta et al., 2013). Advances in 3D bioprinting have enabled the creation of complex, tissue-like structures that mimic the function of a natural pancreas. Although still experimental, this technology shows promise for future clinical applications (Xu et al., 2022).
Despite these advancements, several challenges persist. In human beta cell technology, preventing the immune system from attacking transplanted cells is a primary issue. Current strategies involve immunosuppressive drugs, which have significant side effects. Ensuring the long-term viability and functionality of transplanted beta cells is another major hurdle, with ongoing research aimed at improving their survival and insulin-producing capabilities (Abadpour et al., 2021a). Teratoma formation is another issue in beta cell clusters derived from hPSCs due to the inefficiency of the differentiation process. Despite strategies to enrich beta cell populations, the risk remains because even a single proliferating cell can initiate teratoma formation (Tan et al., 2022).
Similarly, bioartificial pancreas development faces challenges such as finding a reliable and scalable source of islets and developing biocompatible, durable encapsulation materials that allow necessary nutrient and insulin exchange (Abadpour et al., 2021a; Pareta et al., 2013).
The market for diabetes treatment technologies is expanding rapidly due to the increasing prevalence of diabetes, technological advancements, and significant investments from both public and private sectors. These advancements hold the potential to revolutionize diabetes treatment, offering more effective and sustainable solutions for patients worldwide (Gruessner and Gruessner, 2013).
6 Conclusion and future perspectives
Current therapies for diabetes have evolved significantly in the recent years, moving well beyond the pharmacological treatments which traditionally rely on the control of blood glucose levels. While insulin therapy remains essential for individuals with Type 1 diabetes, many patients face challenges with medication side effects, dosing difficulties, and general treatment burden, which impact quality of life and adherence. Pancreas transplants can be an alternative has they offer a more lasting solution. However, they are limited by donor shortage and by the surgical and immunosuppressive risks associated to it. Clinical islet transplantation has emerged as a promising alternative, offering the potential for insulin independence. Yet, this procedure is still dealing with challenges such as limited islet availability, potential autoimmune rejection, and long-term graft viability, which restricts its use in the general patient population.
In response to these limitations, researchers are intensifying efforts in cell-based therapies, which promise a sustainable and personalized approach to diabetes management. The production of mature, functional beta cells outside the body represents a significant breakthrough in cell therapy. This endeavour involves either reprogramming cells from other pancreatic types or differentiating adult stem cells directly into insulin-producing beta cells. These approaches offer promise, as they could circumvent some immune compatibility issues associated with traditional allogeneic transplants. The differentiation of beta cells from hPSCs stands out as the most exciting development, holding the potential for an unlimited supply of patient-specific insulin-producing cells. The most significant study up to now was performed by Z. Shen and co-workers, who developed GMP-compliant personalized pluripotent stem-cell-derived islets allowing, for the first time, its clinical autologous transplantation (Wang et al., 2024). Around 1.5 million islets where implanted into a woman’s abdominal muscles, allowing for easy monitorization using magnetic resonance imaging and quick removal if necessary. The woman was able to produce enough insulin to live without insulin administration for more than a year. While this study pushes the boundaries of personalized therapies for diabetes treatment, it is still too early to understand its full potential. Issues related with safety and efficacy in the long-term and the scale up to a higher number of patients remains to be evaluated. Also, challenges remain in refining the differentiation protocols, and controlling production costs, which are essential for moving these cells into full clinical application. In this regard, engineering a bioartificial pancreas, which can ensure long-term cell function, presents another Frontier in diabetes treatment.
Advances in biomaterials, as well as in scaffold design and processing, aim to create robust, glucose-responsive living implants that enhance the delivery and survival of beta cells. Researchers are currently focused on developing bioartificial pancreas models that closely emulate the structure and functionality of native pancreatic tissue. Decellularized pancreatic scaffolds, in particular, provide a naturally ECM-rich environment that supports both cell integration and insulin production, helping to replicate the in vivo conditions necessary for beta cell endurance in the living body. The applicability of these bioengineering strategies extends beyond clinical use, as they can also serve as physiologically relevant in vitro models for studying diabetes. These tools can transform preclinical drug testing and advance the development of bioartificial pancreas prototypes, moving researchers closer to create functional, transplantable pancreatic substitutes. With continued advancements, bioengineered constructs can potentially transform diabetes treatment, offering regenerative, patient-specific therapies, that could provide lasting therapeutic options to a wide range of patients.
Author contributions
JS: Writing–original draft, Writing–review and editing. SS: Writing–original draft, Writing–review and editing. HL: Writing–original draft, Writing–review and editing. RC: Writing–original draft, Writing–review and editing. BO: Writing–original draft, Writing–review and editing. RS: Writing–original draft, Writing–review and editing. VR: Writing–original draft, Writing–review and editing. AO: Writing–original draft, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. National Funds from Fundação para a Ciência e a Tecnologia (FCT), through the projects LESSisMORE under FCT ERC-Portugal Program and UIDB/50016/2020. IBEROS + POCTEP. Individual Junior Research contract 2023.07374.CEECIND attributed to VR and the Doctoral Research Grant (2024.00955.BDANA) attributed to SS by FCT.
Conflict of interest
Authors HL and BO were employed by Phenocell S.A.S.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abadpour, S., Wang, C., Niemi, E. M., and Scholz, H. (2021a). Tissue engineering strategies for improving beta cell transplantation outcome. TISSUE Eng. Regen. 8, 205–219. doi:10.1007/s40472-021-00333-2
Abadpour, S., Wang, C., Niemi, E. M., and Scholz, H. (2021b). Tissue engineering strategies for improving beta cell transplantation outcome. Curr. Transpl. Rep. 8 (3), 205–219. doi:10.1007/s40472-021-00333-2
Abraham, E. J., Leech, C. A., Lin, J. C., Zulewski, H., and Habener, J. F. (2002). Insulinotropic hormone glucagon-like peptide-1 differentiation of human pancreatic islet-derived progenitor cells into insulin-producing cells. Endocrinology 143 (8), 3152–3161. doi:10.1210/endo.143.8.8973
Aloke, C., Egwu, C. O., Aja, P. M., Obasi, N. A., Chukwu, J., Akumadu, B. O., et al. (2022). Current advances in the management of diabetes mellitus. Biomedicines 10 (10), 2436. doi:10.3390/biomedicines10102436. Available online at: https://www.mdpi.com/2227-9059/10/10/2436
American Diabetes Association Professional Practice Committee (2025). Diabetes Technology: Standards of Care in Diabetes. Diabetes care. 48, S146–66. doi:10.2337/dc25-s007
An, D., Chiu, A., Flanders, J. A., Song, W., Shou, D., Lu, Y. C., et al. (2018). Designing a retrievable and scalable cell encapsulation device for potential treatment of type 1 diabetes. Proc. Natl. Acad. Sci. 115 (2), E263–E272. doi:10.1073/pnas.1708806115
Arce, K. M., Lin, Y. K., Stevens, T., Walsh, R. M., and Hatipoglu, B. A. (2016). Total pancreatectomy and islet cell autotransplantation: definitive treatment for chronic pancreatitis. Cleve Clin. J. Med. 83 (6), 435–442. doi:10.3949/ccjm.83a.15056. Available online at: http://www.ccjm.org/content/83/6/435.abstract
Ariyachet, C., Tovaglieri, A., Xiang, G., Lu, J., Shah, M. S., Richmond, C. A., et al. (2006). Reprogrammed stomach tissue as a renewable source of functional β cells for blood glucose regulation. Cell Stem Cell 18 (3), 410–421. doi:10.1016/j.stem.2016.01.003
Artner, I., Hang, Y., Mazur, M., Yamamoto, T., Guo, M., Lindner, J., et al. (2010). MafA and MafB regulate genes critical to β-cells in a unique temporal manner. Diabetes 59 (10), 2530–2539. doi:10.2337/db10-0190
Atkinson, M. A., von Herrath, M., Powers, A. C., and Clare-Salzler, M. (2015). Current concepts on the pathogenesis of type 1 diabetes—considerations for attempts to prevent and reverse the disease. Diabetes Care 38 (6), 979–988. doi:10.2337/dc15-0144
Avrahami, D., Klochendler, A., Dor, Y., and Glaser, B. (2017). Beta cell heterogeneity: an evolving concept. Diabetologia 60 (8), 1363–1369. doi:10.1007/s00125-017-4326-z
Barsby, T., and Otonkoski, T. (2022). Maturation of beta cells: lessons from in vivo and in vitro models. Diabetologia 65 (6), 917–930. doi:10.1007/s00125-022-05672-y
Basu, S., Yudkin, J. S., Kehlenbrink, S., Women’ S Hospital, B., Davies, J., Wild, S. H., et al. (2018). ESTIMATION of global insulin use for type 2 estimation of global insulin utilisation for type 2 diabetes mellitus, 2018 to 2030. Available online at: www.haiweb.org.
Benninger, R. K. P., and Kravets, V. (2022). The physiological role of β-cell heterogeneity in pancreatic islet function. Nat. Rev. Endocrinol. 18 (1), 9–22. doi:10.1038/s41574-021-00568-0
Berishvili, E., Peloso, A., Tomei, A., and Pepper, A. (2024). The future of beta cells replacement in the era of regenerative medicine and organ bioengineering. Transpl. Int. 37, 12885. doi:10.3389/ti.2024.12885
Briggs, J. K., Schonblum, A., Landsman, L., and Benninger, R. K. P. (2022). Going with the flow: pericyte-regulated islet blood flow influences glucose homeostasis. Diabetes 71 (8), 1611–1613. doi:10.2337/dbi22-0016
Candiello, J., Grandhi, T. S. P., Goh, S. K., Vaidya, V., Lemmon-Kishi, M., Eliato, K. R., et al. (2018). 3D heterogeneous islet organoid generation from human embryonic stem cells using a novel engineered hydrogel platform. Biomaterials 177, 27–39. doi:10.1016/j.biomaterials.2018.05.031. Available online at: https://www.sciencedirect.com/science/article/pii/S0142961218303740.
Cañibano-Hernández, A., Saenz del Burgo, L., Espona-Noguera, A., Orive, G., Hernández, R. M., Ciriza, J., et al. (2019). Hyaluronic acid enhances cell survival of encapsulated insulin-producing cells in alginate-based microcapsules. Int. J. Pharm. 557, 192–198. doi:10.1016/j.ijpharm.2018.12.062. Available online at: https://www.sciencedirect.com/science/article/pii/S0378517318309803.
Caplan, A. I. (2015). Adult mesenchymal stem cells: when, where, and how. Stem Cells Int. 2015 (1), 1–6. doi:10.1155/2015/628767
Cayabyab, F., Nih, L. R., and Yoshihara, E. (2021). Advances in pancreatic islet transplantation sites for the treatment of diabetes. Front. Endocrinol. (Lausanne) 12, 732431. doi:10.3389/fendo.2021.732431. Available online at: https://www.frontiersin.org/journals/endocrinology/articles/10.3389/fendo.2021.732431.
Chen, S., Wang, W., Shen, L., Liu, H., Luo, J., Ren, Y., et al. (2025). A 3D-printed microdevice encapsulates vascularized islets composed of iPSC-derived β-like cells and microvascular fragments for type 1 diabetes treatment. Biomaterials, 315, 122947. doi:10.1016/j.biomaterials.2024.122947
Collaboration, TERF (2010). Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: a collaborative meta-analysis of 102 prospective studies. Lancet 375 (9733), 2215–2222. doi:10.1016/S0140-6736(10)60484-9
Daley, G. Q., Hyun, I., Apperley, J. F., and Barker, R. A. (2016). Benvenisty N, Bredenoord AL, et al. Setting Global Standards for Stem Cell Research and Clinical Translation: The 2016 ISSCR Guidelines. Stem Cell Rep. 14 (6), 787–797. doi:10.1016/j.stemcr.2016.05.001
D’Amour, K. A., Bang, A. G., Eliazer, S., Kelly, O. G., Agulnick, A. D., Smart, N. G., et al. (2006). Production of pancreatic hormone–expressing endocrine cells from human embryonic stem cells. Nat. Biotechnol. 24 (11), 1392–1401. doi:10.1038/nbt1259
Davis, N. E., Beenken-Rothkopf, L. N., Mirsoian, A., Kojic, N., Kaplan, D. L., Barron, A. E., et al. (2012). Enhanced function of pancreatic islets co-encapsulated with ECM proteins and mesenchymal stromal cells in a silk hydrogel. Biomaterials 33 (28), 6691–6697. doi:10.1016/j.biomaterials.2012.06.015. Available online at: https://www.sciencedirect.com/science/article/pii/S0142961212006515.
Domínguez-Bendala, J., and Ricordi, C. (2012). Present and future cell therapies for pancreatic beta cell replenishment. World J. Gastroenterol. 18 (47), 6876–6884. doi:10.3748/wjg.v18.i47.6876
ElSayed, N. A., McCoy, R. G., Aleppo, G., Bajaj, M., Balapattabi, K., Beverly, E. A., et al. (2025b). 9. Pharmacologic approaches to glycemic treatment: standards of care in diabetes—2025. Diabetes Care 48 (Suppl. ment_1), S181–S206. doi:10.2337/dc25-s009
ElSayed, N. A., McCoy, R. G., Aleppo, G., Balapattabi, K., Beverly, E. A., Briggs Early, K., et al. (2025a). 2. Diagnosis and classification of diabetes: standards of care in diabetes—2025. Diabetes Care 48 (Suppl. ment_1). Available online at: https://diabetesjournals.org/care/article/48/Supplement_1/S27/157566/2-Diagnosis-and-Classification-of-Diabetes.
Faleo, G., Russ, H. A., Wisel, S., Parent, A. V., Nguyen, V., Nair, G. G., et al. (2017). Mitigating ischemic injury of stem cell-derived insulin-producing cells after transplant. Stem Cell Rep. 9 (3), 807–819. doi:10.1016/j.stemcr.2017.07.012
Félix-Martínez, G. J., Osorio-Londoño, D., and Godínez-Fernández, J. R. (2024). Impact of oxygen and glucose availability on the viability and connectivity of islet cells: a computational study of reconstructed avascular human islets. PLoS Comput. Biol. 20 (8). doi:10.1371/journal.pcbi.1012357
Ferber, S., Halkin, A., Cohen, H., Ber, I., Einav, Y., Goldberg, I., et al. (2000). Pancreatic and duodenal homeobox gene 1 induces expression of insulin genes in liver and ameliorates streptozotocin-induced hyperglycemia. Nat. Med. 6 (5), 568–572. doi:10.1038/75050
Furuyama, K., Chera, S., van Gurp, L., Oropeza, D., Ghila, L., Damond, N., et al. (2019). Diabetes relief in mice by glucose-sensing insulin-secreting human α-cells. Nature 567 (7746), 43–48. doi:10.1038/s41586-019-0942-8
Gruessner, R. W. G., and Gruessner, A. C. (2013). The current state of pancreas transplantation. Nat. Rev. Endocrinol. 9 (9), 555–562. doi:10.1038/nrendo.2013.138
Gupta, S. S., and Sharma, R. K. (2012). Renal transplantation in diabetic end-stage renal disease. Clin. Queries Nephrol. 1 (2), 159–162. doi:10.1016/s2211-9477(12)70012-2. Available online at: https://www.sciencedirect.com/science/article/pii/S2211947712700122.
Hamid, M., McCluskey, J. T., McClenaghan, N. H., and Flatt, P. R. (2001). Functional examination of microencapsulated bioengineered insulin-secreting beta-cells. Cell Biol. Int. 25 (6), 553–556. doi:10.1006/cbir.2000.0663
Henry, R. R., Pettus, J., Wilensky, J. O. N., Shapiro, A. M. J., Senior, P. A., Roep, B., et al. (2018). Initial clinical evaluation of VC-01TM combination product—a stem cell–derived islet replacement for type 1 diabetes (T1D). Diabetes 67 (Suppl. ment_1), 138–OR. doi:10.2337/db18-138-OR
Hering, B. J., Wijkstrom, M., Graham, M. L., Hårdstedt, M., Aasheim, T. C., Jie, T., et al. (2006). Prolonged diabetes reversal after intraportal xenotransplantation of wild-type porcine islets in immunosuppressed nonhuman primates. Nat. Med. 12 (3), 301–303. doi:10.1038/nm1369
Humar, A., Khwaja, K., Ramcharan, T., Asolati, M., Kandaswamy, R., Gruessner, R. W. G., et al. (2003). Chronic rejection: the next major challenge for pancreas transplant recipients. Transplantation 76 (6), 918–923. doi:10.1097/01.tp.0000079457.43199.76Available online at: https://journals.lww.com/transplantjournal/fulltext/2003/09270/chronic_rejection__the_next_major_challenge_for.7.aspx.
Hwang, P. T. J., Shah, D. K., Garcia, J. A., Bae, C. Y., Lim, D. J., Huiszoon, R. C., et al. (2016). Progress and challenges of the bioartificial pancreas. Nano Converg. 3 (1), 28. doi:10.1186/s40580-016-0088-4
IDF (2021). Diabetes atlas. Available online at: www.diabetesatlas.org.
IDF (2025). Diabetes atlas 10th edition. Available online at: www.diabetesatlas.org.
Ji, Z., Lu, M., Xie, H., Yuan, H., and Chen, Q. (2022). β cell regeneration and novel strategies for treatment of diabetes (Review). Biomed. Rep. 17 (3), 72. doi:10.3892/br.2022.1555
Jin, W., and Jiang, W. (2022). Stepwise differentiation of functional pancreatic β cells from human pluripotent stem cells. Vol. Cell Regen., 11, 24. doi:10.1186/s13619-022-00125-8
Kalluri, H. V. (2012). Current state of renal transplant immunosuppression: present and future. World J. Transpl. 2 (4), 51. doi:10.5500/wjt.v2.i4.51
Karimova, M. V., Gvazava, I. G., and Vorotelyak, E. A. (2022). Overcoming the limitations of stem cell-derived beta cells. Vol. Biomolecules, 12, 810. doi:10.3390/biom12060810
Kaur, G., Baino, F., Mauro, J. C., Kumar, V., Pickrell, G., Sriranganathan, N., et al. (2017). “Biomaterials for cell encapsulation: progress toward clinical applications,” in Clinical applications of biomaterials: state-of-the-art progress, trends, and novel approaches. Editor G. Kaur (Cham: Springer International Publishing), 425–458. doi:10.1007/978-3-319-56059-5_14
Kroon, E., Martinson, L. A., Kadoya, K., Bang, A. G., Kelly, O. G., Eliazer, S., et al. (2008a). Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat. Biotechnol. 26 (4), 443–452. doi:10.1038/nbt1393
Kroon, E., Martinson, L. A., Kadoya, K., Bang, A. G., Kelly, O. G., Eliazer, S., et al. (2008b). Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat. Biotechnol. 26 (4), 443–452. doi:10.1038/nbt1393
Kropp, C., Kempf, H., Halloin, C., Robles-Diaz, D., Franke, A., Scheper, T., et al. (2016). Impact of feeding strategies on the scalable expansion of human pluripotent stem cells in single-use stirred tank bioreactors. Stem Cells Transl. Med. 5 (10), 1289–1301. doi:10.5966/sctm.2015-0253
Langlois, A., Pinget, M., Kessler, L., and Bouzakri, K. (2024). Islet transplantation: current limitations and challenges for successful outcomes. Cells 13 (21), 1783. doi:10.3390/cells13211783. Available online at: https://www.mdpi.com/2073-4409/13/21/1783.
Larsen, J., Lane, J., and Mack-Shipman, L. (2002). Pancreas and kidney transplantation. Curr. Diab Rep. 2 (4), 359–364. doi:10.1007/s11892-002-0027-0
Larsen, J. L. (2004). Pancreas transplantation: indications and consequences. Endocr. Rev. 25 (6), 919–946. doi:10.1210/er.2002-0036
Larson, T. S., Bohorquez, H., Rea, D. J., Nyberg, S. L., Prieto, M., Sterioff, S., et al. (2004). Pancreas-after-kidney transplantation: an increasingly attractive alternative to simultaneous pancreas-kidney transplantation. Transplantation 77 (6), 838–843. doi:10.1097/01.tp.0000114611.73689.3b. Available online at: https://journals.lww.com/transplantjournal/fulltext/2004/03270/pancreas_after_kidney_transplantation__an.9.aspx.
Li, Y., Liu, Z., Tang, Y., Fan, Q., Feng, W., Luo, C., et al. (2020). Three-dimensional silk fibroin scaffolds enhance the bone formation and angiogenic differentiation of human amniotic mesenchymal stem cells: a biocompatibility analysis. Acta Biochim. Biophys. Sin. (Shanghai) 52 (6), 590–602. doi:10.1093/abbs/gmaa042
Liu, J., and Hebrok, M. (2017). All mixed up: defining roles for β-cell subtypes in mature islets. Genes Dev. 31, 228–240. doi:10.1101/gad.294389.116
London, N. J. M. (1995). Clinical studies of human islet transplantation. Ann. R. Coll. Surg. Engl. 77, 263–269.
Luo, Y., Yu, P., and Liu, J. (2024). The efficiency of stem cell differentiation into functional beta cells for treating insulin-requiring diabetes: recent advances and current challenges. Endocrine 86 (1), 1–14. doi:10.1007/s12020-024-03855-8
Ma, D., Duan, W., Li, Y., Wang, Z., Li, S., Gong, N., et al. (2016). PD-L1 deficiency within islets reduces allograft survival in mice. PLoS One 11 (3). doi:10.1371/journal.pone.0152087
Maqsood, M., Kang, M., Wu, X., Chen, J., Teng, L., and Qiu, L. (2020). Adult mesenchymal stem cells and their exosomes: sources, characteristics, and application in regenerative medicine. Life Sci. 256, 118002. doi:10.1016/j.lfs.2020.118002. Available online at: https://www.sciencedirect.com/science/article/pii/S0024320520307529.
Marfil-Garza, B. A., Shapiro, A. M. J., and Kin, T. (2021). Clinical islet transplantation: current progress and new frontiers. J. Hepatobiliary Pancreat. Sci. 28 (3), 243–254. doi:10.1002/jhbp.891
Matthews, D. R., and Wallace, T. M. (2005). Review: sulphonylureas and the rise and fall of beta-cell function. Br. J. Diabetes Vasc. Dis. 5 (4), 192–196. doi:10.1177/14746514050050040301
Meivar-Levy, I., and Ferber, S. (2019). Liver to pancreas transdifferentiation. Curr. Diab Rep. 19 (9), 76. doi:10.1007/s11892-019-1198-2
Millman, J. R., Xie, C., Van Dervort, A., Gürtler, M., Pagliuca, F. W., and Melton, D. A. (2016). Generation of stem cell-derived β-cells from patients with type 1 diabetes. Nat. Commun. 7 (1), 11463. doi:10.1038/ncomms11463
Mohandas, S., Vijaya, G., Kumaran, K., Gopinath, V., Paulmurugan, R., and Ramkumar, K. (2023). New frontiers in three-dimensional culture platforms to improve diabetes research. Pharmaceutics 15, 725. doi:10.3390/pharmaceutics15030725
Moon, J. E., Lee, Y. N., Jeong, S., Jun, H. R., Hoang, M. H., Jo, Y., et al. (2024). Enhancing differentiation and functionality of insulin-producing cells derived from iPSCs using esterified collagen hydrogel for cell therapy in diabetes mellitus. Stem Cell Res. Ther. 15 (1), 374. doi:10.1186/s13287-024-03971-2
Mudaliar, S. (2023). The evolution of diabetes treatment through the ages: from starvation diets to insulin, incretins, SGLT2-inhibitors and beyond. J. Indian Inst. Sci. 103 (1), 123–133. doi:10.1007/s41745-023-00357-w
Nair, G. G., Liu, J. S., Russ, H. A., Tran, S., Saxton, M. S., Chen, R., et al. (2019). Recapitulating endocrine cell clustering in culture promotes maturation of human stem-cell-derived β cells. Nat. Cell Biol. 21 (2), 263–274. doi:10.1038/s41556-018-0271-4
Nair, G. G., Tzanakakis, E. S., and Hebrok, M. (2020). Emerging routes to the generation of functional β-cells for diabetes mellitus cell therapy. Nat. Rev. Endocrinol. 16 (9), 506–518. doi:10.1038/s41574-020-0375-3
Ogle, G. D., Wang, F., Gregory, G. A., and Maniam, J. (2022). Acknowledgements Type 1 diabetes numbers in children and adults. Available online at: www.diabetesatlas.org.
Okere, B., Lucaccioni, L., Dominici, M., and Iughetti, L. (2016). Cell therapies for pancreatic beta-cell replenishment. Ital. J. Pediatr. 42 (1), 62. doi:10.1186/s13052-016-0273-4
Pagliuca, F. W., Millman, J. R., Gürtler, M., Segel, M., Van Dervort, A., Ryu, J. H., et al. (2014). Generation of functional human pancreatic β cells in vitro. Cell 159 (2), 428–439. doi:10.1016/j.cell.2014.09.040
Pareta, R. A., Farney, A. C., and Opara, E. C. (2013). Design of a bioartificial pancreas. Pathobiology 80 (4), 194–202. doi:10.1159/000345873
Petrus-Reurer, S., Romano, M., Howlett, S., Jones, J. L., Lombardi, G., and Saeb-Parsy, K. (2021). Immunological considerations and challenges for regenerative cellular therapies. Commun. Biol. 4 (1), 798. doi:10.1038/s42003-021-02237-4
Printing Industry (2025). 3D printing Industry. Available online at: https://3dprintingindustry.com/news/readily3d-develops-3d-bioprinted-mini-pancreas-for-diabetes-drug-testing-191440/.
Ramiya, V. K., Maraist, M., Arfors, K. E., Schatz, D. A., Peck, A. B., and Cornelius, J. G. (2000). Reversal of insulin-dependent diabetes using islets generated in vitro from pancreatic stem cells. Nat. Med. 6 (3), 278–282. doi:10.1038/73128
Ramzy, A., Thompson, D. M., Ward-Hartstonge, K. A., Ivison, S., Cook, L., Garcia, R. V., et al. (2021). Implanted pluripotent stem-cell-derived pancreatic endoderm cells secrete glucose-responsive C-peptide in patients with type 1 diabetes. Cell Stem Cell 28 (12), 2047–2061.e5. doi:10.1016/j.stem.2021.10.003
Raoufinia, R., Rahimi, H. R., Saburi, E., and Moghbeli, M. (2024). Advances and challenges of the cell-based therapies among diabetic patients. J. Transl. Med. 22 (1), 435. doi:10.1186/s12967-024-05226-3
Rezania, A., Bruin, J. E., Arora, P., Rubin, A., Batushansky, I., Asadi, A., et al. (2014). Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 32 (11), 1121–1133. doi:10.1038/nbt.3033
Rezania, A., Bruin, J. E., Riedel, M. J., Mojibian, M., Asadi, A., Xu, J., et al. (2012). Maturation of human embryonic stem cell–derived pancreatic progenitors into functional islets capable of treating pre-existing diabetes in mice. Diabetes 61 (8), 2016–2029. doi:10.2337/db11-1711
Rickels, M. R. (2012). Recovery of endocrine function after islet and pancreas transplantation. Curr. Diab Rep. 12 (5), 587–596. doi:10.1007/s11892-012-0294-3
Rickels, M. R., and Robertson, R. P. (2019). Pancreatic islet transplantation in humans: recent progress and future directions. Endocr. Rev. 40 (2), 631–668. doi:10.1210/er.2018-00154
Robertson, J. A. (2010). Embryo stem cell research: ten years of controversy. J. Law, Med. and Ethics 38 (2), 191–203. doi:10.1111/j.1748-720X.2010.00479.x
Rodriguez, S., Alexander, M., and Lakey, J. R. T. (2020). “Pancreatic islet transplantation: a surgical approach to type 1 diabetes treatment,” in Obesity and diabetes: scientific advances and best practice. Editors J. Faintuch, and S. Faintuch (Cham: Springer International Publishing), 655–664. doi:10.1007/978-3-030-53370-0_48
Rutter, G. A., Gresch, A., Delgadillo Silva, L., and Benninger, R. K. P. (2024). Exploring pancreatic beta-cell subgroups and their connectivity. Nat. Metab. 6, 2039–2053. doi:10.1038/s42255-024-01097-6
Saeedi, P., Petersohn, I., Salpea, P., Malanda, B., Karuranga, S., Unwin, N., et al. (2019). Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: results from the international diabetes federation diabetes atlas, 9th edition. Diabetes Res. Clin. Pract. 157, 107843. doi:10.1016/j.diabres.2019.107843
Saleh, M., Gittes, G. K., and Prasadan, K. (2021). Alpha-to-beta cell trans-differentiation for treatment of diabetes. Biochem. Soc. Trans. 49 (6), 2539–2548. doi:10.1042/BST20210244
Salg, G. A., Giese, N. A., Schenk, M., Hüttner, F. J., Felix, K., Probst, P., et al. (2019). The emerging field of pancreatic tissue engineering: a systematic review and evidence map of scaffold materials and scaffolding techniques for insulin-secreting cells. J. Tissue Eng. 10, 2041731419884708. doi:10.1177/2041731419884708
Satin, L. S., Zhang, Q., and Rorsman, P. (2020). “Take me to your leader”: an electrophysiological appraisal of the role of hub cells in pancreatic islets. Diabetes 69 (5), 830–836. An Electrophysiological Appraisal of the Role of Hub Cells in Pancreatic Islets. doi:10.2337/dbi19-0012
Seaberg, R. M., Smukler, S. R., Kieffer, T. J., Enikolopov, G., Asghar, Z., Wheeler, M. B., et al. (2004). Clonal identification of multipotent precursors from adult mouse pancreas that generate neural and pancreatic lineages. Nat. Biotechnol. 22 (9), 1115–1124. doi:10.1038/nbt1004
Shahjalal, H.Md, Abdal Dayem, A., Lim, K. M., Jeon, T. il, and Cho, S. G. (2018). Generation of pancreatic β cells for treatment of diabetes: advances and challenges. Stem Cell Res. Ther. 9 (1), 355. doi:10.1186/s13287-018-1099-3
Shapiro, A. M. J., Thompson, D., Donner, T. W., Bellin, M. D., Hsueh, W., Pettus, J., et al. (2021). Insulin expression and C-peptide in type 1 diabetes subjects implanted with stem cell-derived pancreatic endoderm cells in an encapsulation device. Cell Rep. Med. 2 (12), 100466. doi:10.1016/j.xcrm.2021.100466. Available online at: https://www.sciencedirect.com/science/article/pii/S2666379121003384.
Singh, A., Afshan, N., Singh, A., Singh, S. K., Yadav, S., Kumar, M., et al. (2023). Recent trends and advances in type 1 diabetes therapeutics: a comprehensive review. Eur. J. Cell Biol. 102 (2), 151329. doi:10.1016/j.ejcb.2023.151329. Available online at: https://www.sciencedirect.com/science/article/pii/S0171933523000444.
Skyler, J. S., Bakris, G. L., Bonifacio, E., Darsow, T., Eckel, R. H., Groop, L., et al. (2016). Differentiation of diabetes by pathophysiology, natural history, and prognosis. Diabetes 66 (2), 241–255. doi:10.2337/db16-0806
Skyler, J. S., and Ricordi, C. (2011). Stopping type 1 diabetes: attempts to prevent or cure type 1 diabetes in man. Diabetes 60 (1), 1–8. doi:10.2337/db10-1114
Smink, A. M., de Haan, B. J., Lakey, J. R. T., and de Vos, P. (2018). Polymer scaffolds for pancreatic islet transplantation — progress and challenges. Am. J. Transplant. 18 (9), 2113–2119. doi:10.1111/ajt.14942. Available online at: https://www.sciencedirect.com/science/article/pii/S1600613522096848.
Smink, A. M., Rodriquez, S., Li, S., Ceballos, B., Corrales, N., Alexander, M., et al. (2023). Successful islet transplantation into a subcutaneous polycaprolactone scaffold in mice and pigs. Transpl. Direct 9 (1), e1417. doi:10.1097/txd.0000000000001417. Available online at: https://journals.lww.com/transplantationdirect/fulltext/2023/01000/successful_islet_transplantation_into_a.11.aspx.
Spadoni, I., Fornasa, G., and Rescigno, M. (2017). Organ-specific protection mediated by cooperation between vascular and epithelial barriers. Nat. Rev. Immunol. 17 (12), 761–773. doi:10.1038/nri.2017.100
Stephens, C. H., Morrison, R. A., McLaughlin, M., Orr, K., Tersey, S. A., Scott-Moncrieff, J. C., et al. (2020). Oligomeric collagen as an encapsulation material for islet/β-cell replacement: effect of islet source, dose, implant site, and administration format. Am. J. Physiology-Endocrinology Metabolism 319 (2), E388–E400. doi:10.1152/ajpendo.00066.2020
Stock, A. A., Gonzalez, G. C., Pete, S. I., De Toni, T., Berman, D. M., Rabassa, A., et al. (2024). Performance of islets of Langerhans conformally coated via an emulsion cross-linking method in diabetic rodents and nonhuman primates. Sci. Adv. 8 (26), eabm3145. doi:10.1126/sciadv.abm3145
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., et al. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131 (5), 861–872. doi:10.1016/j.cell.2007.11.019
Tan, L. S., Chen, J. T., Lim, L. Y., and Teo, A. K. K. (2022). Manufacturing clinical-grade human induced pluripotent stem cell-derived beta cells for diabetes treatment. Cell Prolif. 55, e13232. doi:10.1111/cpr.13232
Tao, T., Wang, Y., Chen, W., Li, Z., Su, W., Guo, Y., et al. (2019). Engineering human islet organoids from iPSCs using an organ-on-chip platform. Lab Chip, 948–58.
Thakur, G., Lee, H. J., Jeon, R. H., Lee, S. L., and Rho, G. J. (2020). “Small molecule-induced pancreatic β-like cell development: mechanistic approaches and available strategies,”, 21. Int. J. Mol. Sci., 2388. doi:10.3390/ijms21072388
Timmons, J., and Boyle, J. (2022). “Sulfonylureas and meglitinides,” in Diabetes drug notes, 49–66. doi:10.1002/9781119785033.ch3
Toftdal, M. S., Grunnet, L. G., and Chen, M. (2024). Emerging strategies for beta cell encapsulation for type 1 diabetes therapy. Adv. Healthc. Mater 13 (19), 2400185. doi:10.1002/adhm.202400185
Toftdal, M. S., Taebnia, N., Kadumudi, F. B., Andresen, T. L., Frogne, T., Winkel, L., et al. (2021). Oxygen releasing hydrogels for beta cell assisted therapy. Int. J. Pharm. 602, 120595. doi:10.1016/j.ijpharm.2021.120595. Available online at: https://www.sciencedirect.com/science/article/pii/S0378517321004002.
Tremmel, D. M., Mikat, A. E., Gupta, S., Mitchell, S. A., Curran, A. M., Menadue, J. A., et al. (2023). Validating expression of beta cell maturation-associated genes in human pancreas development. Front. Cell Dev. Biol. 11, 1103719. doi:10.3389/fcell.2023.1103719
van der Boog, P. J. M., Ringers, J., Paul, L. C., Jukema, J. W., Baranski, A., Lemkes, HHPJ, et al. (2004). Simultaneous kidney-pancreas transplantation: the preferred option for patients with type I diabetes mellitus and approaching end-stage renal disease. Transpl. Rev. 18 (3), 129–138. doi:10.1016/j.trre.2004.04.003. Available online at: https://www.sciencedirect.com/science/article/pii/S0955470X04000436.
Veres, A., Faust, A. L., Bushnell, H. L., Engquist, E. N., Kenty, J. H. R., Harb, G., et al. (2019a). Charting cellular identity during human in vitro β-cell differentiation. Nature 569 (7756), 368–373. doi:10.1038/s41586-019-1168-5
Veres, A., Faust, A. L., Bushnell, H. L., Engquist, E. N., Kenty, J. H. R., Harb, G., et al. (2019b). Charting cellular identity during human in vitro β-cell differentiation. Nature 569 (7756), 368–373. doi:10.1038/s41586-019-1168-5
Wang, S., Du, Y., Zhang, B., Meng, G., Liu, Z., Liew, S. Y., et al. (2024). Transplantation of chemically induced pluripotent stem-cell-derived islets under abdominal anterior rectus sheath in a type 1 diabetes patient. Cell 187 (22), 6152–6164.e18. doi:10.1016/j.cell.2024.09.004
Wang, X., Gao, M., Wang, Y., and Zhang, Y. (2022). The progress of pluripotent stem cell-derived pancreatic β-cells regeneration for diabetic therapy. Front. Endocrinol. (Lausanne)., 13, 927324. doi:10.3389/fendo.2022.927324
Wang, X., Maxwell, K. G., Wang, K., Bowers, D. T., Flanders, J. A., Liu, W., et al. (2021). A nanofibrous encapsulation device for safe delivery of insulin-producing cells to treat type 1 diabetes. Sci. Transl. Med. 13 (596), eabb4601. doi:10.1126/scitranslmed.abb4601
Xu, Y., Song, D., and Wang, X. (2022). 3D bioprinting for pancreas engineering/manufacturing. Polym. (Basel) 14, 5143. doi:10.3390/polym14235143
Yang, K., O’Cearbhaill, E. D., Liu, S. S., Zhou, A., Chitnis, G. D., Hamilos, A. E., et al. (2021). A therapeutic convection–enhanced macroencapsulation device for enhancing β cell viability and insulin secretion. Proc. Natl. Acad. Sci. U. S. A. 118 (37), e2101258118. doi:10.1073/pnas.2101258118
Yoshihara, E., O’Connor, C., Gasser, E., Wei, Z., Oh, T. G., Tseng, T. W., et al. (2020). Immune-evasive human islet-like organoids ameliorate diabetes. Nature 586 (7830), 606–611. doi:10.1038/s41586-020-2631-z
Zhang, X., Ma, Z., Song, E., and Xu, T. (2022). Islet organoid as a promising model for diabetes. Protein Cell 13 (4), 239–257. doi:10.1007/s13238-021-00831-0
Zhao, X., Li, Q., Guo, Z., and Li, Z. (2021). Constructing a cell microenvironment with biomaterial scaffolds for stem cell therapy. Stem Cell Res. Ther. 12 (1), 583. doi:10.1186/s13287-021-02650-w
Zulewski, H., Abraham, E. J., Gerlach, M. J., Daniel, P. B., Moritz, W., Müller, B., et al. (2001). Multipotential nestin-positive stem cells isolated from adult pancreatic islets differentiate ex vivo into pancreatic endocrine, exocrine, and hepatic phenotypes. Diabetes 50 (3), 521–533. doi:10.2337/diabetes.50.3.521
Keywords: diabetes, cell therapies, beta cells, pancreas, tissue engineering
Citation: Sá J, Sá S, Leménager H, Costa R, Onteniente B, Soares R, Ribeiro VP and Oliveira AL (2025) Advancing diabetes treatment: from human beta cell technology to bioartificial pancreas development . Front. Biomater. Sci. 4:1524518. doi: 10.3389/fbiom.2025.1524518
Received: 08 November 2024; Accepted: 28 February 2025;
Published: 19 March 2025.
Edited by:
Rajakrishnan Veluthakal, City of Hope National Medical Center, United StatesReviewed by:
João Paulo Monteiro Carvalho Mori Cunha, University of Copenhagen, DenmarkCopyright © 2025 Sá, Sá, Leménager, Costa, Onteniente, Soares, Ribeiro and Oliveira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Viviana P. Ribeiro, dnByaWJlaXJvQHVjcC5wdA==; Ana L. Oliveira, YWxvbGl2ZWlyYUB1Y3AucHQ=