 Yongjiu Zhang
Yongjiu Zhang Shuxiao Yang
Shuxiao Yang Xianliang Zheng
Xianliang Zheng Xiaoming Tan
Xiaoming Tan
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Bioeng. Biotechnol., 27 February 2025
Sec. Synthetic Biology
Volume 13 - 2025 | https://doi.org/10.3389/fbioe.2025.1552030
Cyanobacteria, renowned for their photosynthetic capabilities, serve as efficient microbial chassis capable of converting carbon dioxide into a spectrum of bio-chemicals. However, conventional genetic manipulation strategies have proven incompatible with the precise and systematic modifications required in the field of cyanobacterial synthetic biology. Here, we present an in-depth analysis of endogenous CRISPR-Cas systems within cyanobacterial genomes, with a particular focus on the Type I systems, which are the most widely distributed. We provide a comprehensive summary of the reported DNA defense mechanisms mediated by cyanobacterial Type I CRISPR-Cas systems and their current applications in genome editing. Furthermore, we offer insights into the future applications of these systems in the context of cyanobacterial genome editing, underscoring their potential to revolutionize synthetic biology approaches.
Cyanobacteria, a phylum of prokaryotic organisms, are capable of fixing carbon dioxide and releasing oxygen into the atmosphere through the process of photosynthesis. As one of the most ancient forms of life on Earth, they have been a dominant force in Earth’s ecosystems for approximately a billion years, markedly enhancing the levels of atmospheric oxygen (Demoulin et al., 2019). In current aquatic ecosystems, cyanobacteria continue to play a pivotal role in the biogeochemical cycling of nutrients across various habitats (Bhardwaj et al., 2024).
Because of their rapid autotrophic growth rate, genetic amenability, and photosynthetic capabilities, cyanobacteria have been considered as a promising photosynthetic chassis for biotechnological applications (Ducat et al., 2011; Angermayr et al., 2015). Metabolic engineering efforts have enabled the biosynthesis of a broad spectrum of products from CO2 in cyanobacteria, thereby demonstrating the substantial potential of cyanobacterial photosynthetic biotechnology (Ni et al., 2018). Nonetheless, the low product yields remains a critical barrier to the commercial feasibility of these technologies (Wang et al., 2020).
Systematic metabolic engineering is an effective approach to boost the efficiency of microbial biosynthesis. However, it is challenging to realize genome-scale modifications in cyanobacteria by using conventional genetic manipulation methods which reply on a limited numbers of drug markers. In contrast, CRISPR-Cas dependent genome editing facilitates the targeted modification of genomic loci in a marker-free manner and enables simultaneous modifications across multiple genomic loci, making it an ideal tool for the systematic metabolic engineering of cyanobacteria (Sun et al., 2018). Thus, many efforts have been invested in the introduction of CRISPR-Cas9 or CRISPR-Cas12a genome editing systems for cyanobacteria, as summarized in some recent reviews (Carroll et al., 2018; Vijay et al., 2019; Pattharaprachayakul et al., 2020; Dong et al., 2024).
In recent years, the utilization of native type I CRISPR-Cas systems for genome editing has emerged as a promising strategy in prokaryotes (Zheng et al., 2020; Xu et al., 2021). Unlike the widely employed exogenous Cas9 and Cas12a effectors, endogenously encoded Cas effectors in type I systems do not require additional over-expression and exhibit better compatibility with the host (Zheng et al., 2019; Du et al., 2022). Notably, the majority of cyanobacterial genomes encode endogenous CRISPR-Cas systems, which serve as immune systems against foreign DNA invasions (Doron et al., 2018; Strecker et al., 2019; Ziemann et al., 2023). The widespread distribution of type I CRISPR-Cas systems in cyanobacteria (Makarova et al., 2019; Pattharaprachayakul et al., 2020) holds significant potential for endogenous CRISPR-based genome engineering.
Recently, some studies have been performed to elucidate the adaptation and interference mechanisms of these native CRISPR-Cas systems in cyanobacteria (Jesser et al., 2019; McBride et al., 2020; Schwartz et al., 2022). Furthermore, several cyanobacterial CRISPR-Cas systems have been harnessed for genetic manipulations in mammalian cells or higher plants (Keishi Osakabe, 2020; Osakabe et al., 2021). We have successfully reprogrammed the native Type I CRISPR-Cas system of Synechococcus sp. PCC 7002 to target its own genome, thereby achieving marker-less gene deletions (Yang et al., 2024).
In this review, we present a comprehensive analysis of the native CRISPR-Cas systems within cyanobacteria. Our examination encompasses the prevalence of these systems across cyanobacterial genomes. Additionally, we summarize the underlying immune mechanisms of type I CRISPR-Cas systems, their current application in genome editing, and offer insights into the potential future use of type I CRISPR-Cas systems in cyanobacterial biotechnology.
In an analysis of 126 cyanobacterial genomes, it was predicted that the majority of these genomes harbored the CRISPR-Cas system (Cai et al., 2013). Additionally, a prevalence of CRISPR-Cas systems was identified in 171 genomes of multicellular cyanobacteria (Hou et al., 2018). To elucidate the distribution of CRISPR-Cas systems within cyanobacteria, we conducted a re-analysis of endogenous CRISPR-Cas systems in publicly available complete cyanobacterial genomes using the CRISPRCasTyper pipeline (Russel et al., 2020).
We retrieved 335 complete cyanobacterial genomes from the NCBI Genome database (29 October 2024). Among the 315 unique genomes, at least one CRISPR-Cas system was identified in 197 genomes, representing 62.5% of the cyanobacterial genomes examined (Supplementary Table S1). The distribution of CRISPR-Cas systems was found to be significantly variable across different orders of cyanobacteria. A substantial majority (84.7%) of cyanobacteria lacking CRISPR-Cas systems were classified under the order Synechococcales. Upon excluding the Synechococcales order, 180 (90.9%) of the remaining 198 cyanobacterial genomes were predicted to contain the CRISPR-Cas system. Notably, nearly all genomes from the Nostocales (96.7%) and Leptolyngbyales (100%) orders, as well as over 80% of genomes from the Acaryochloridales, Chroococcales, and Oscillatoriales, were found to contain CRISPR-Cas systems (Figure 1A).
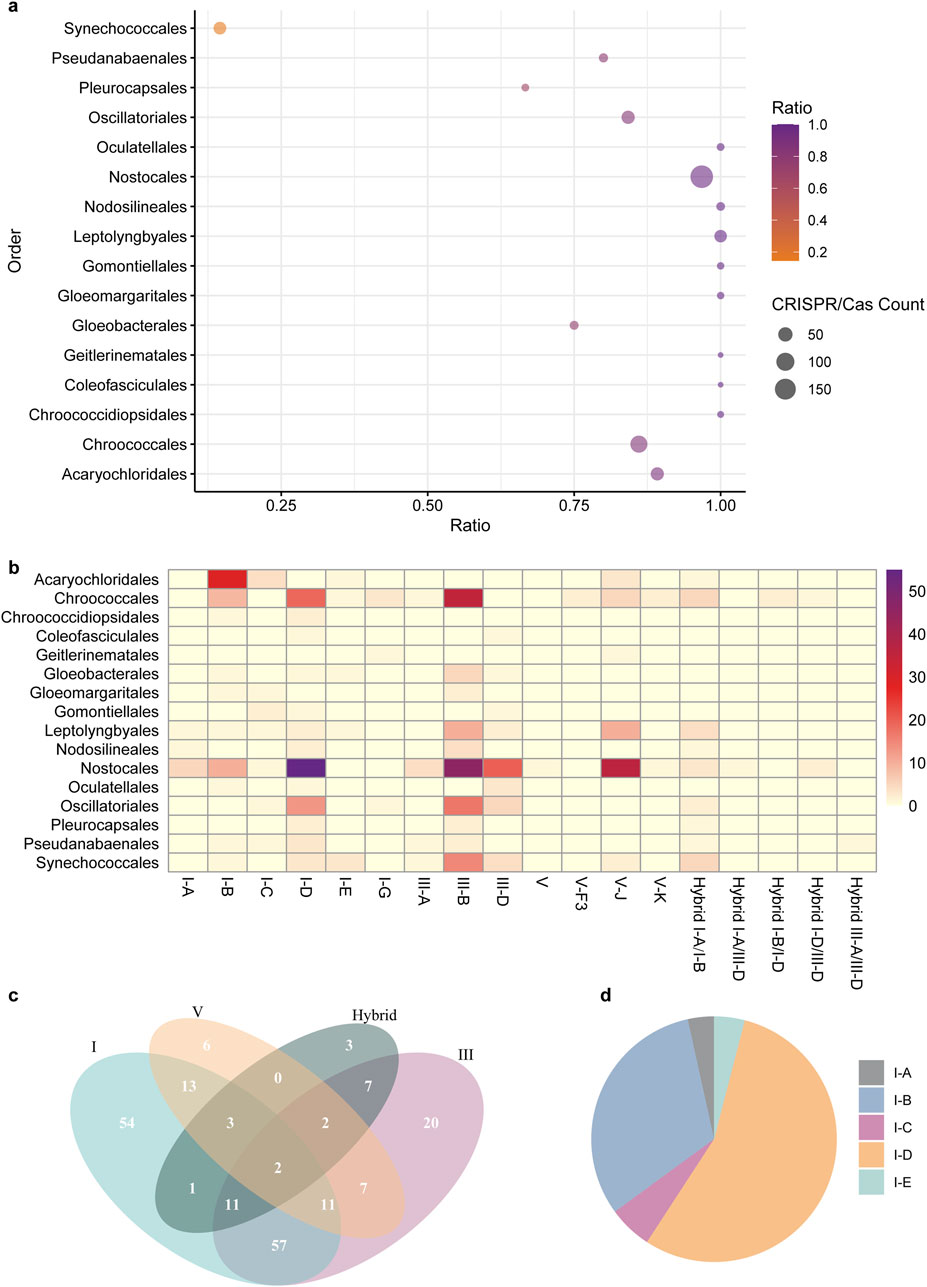
Figure 1. Distribution and prevalence of CRISPR-Cas Systems across cyanobacterial Orders. (A) The scatter plot presents the numbers of CRISPR-Cas systems across different cyanobacterial orders and the ratio of genomes containing the system to the total number of genomes in the specific orders. The x-axis represents the ratio, while the y-axis lists the cyanobacterial orders. The size of each point corresponds to the count of CRISPR-Cas systems within that order, with larger points indicating a higher count, as detailed in the legend. The color gradient from orange to purple corresponds the ratio of genomes with CRISPR-Cas systems. (B) The heatmap visualizes the abundance of different types of CRISPR-Cas systems across various cyanobacterial orders. The rows represent distinct CRISPR-Cas types (I-A, I-B, I-C, etc.), while the columns list the cyanobacterial orders. The color intensity corresponds to the number of CRISPR-Cas systems identified within each order, as indicated by the color legend. (C) The Venn diagram illustrates the co-existence of different CRISPR-Cas system types: I, III, and V, along with hybrid systems, in cyanobacterial genomes. Each circle represents a unique system type, with the numbers inside the overlapping areas indicating the count of genomes containing both of the systems. (D) The pie chart illustrates the proportion of five distinct subtypes of cyanobacterial Type I CRIPSR-Cas system. Each color corresponds to a specific subtype.
All cyanobacterial CRISPR-Cas candidates can be categorized into two major classes with three types, including types I and III in the Class 1, and types V in the Class 2 (Figure 1B). Type I, III, V and certain hybrid CRISPR-Cas systems were found in 152, 117, 44 and 29 cyanobacterial genomes, respectively, accounting for 48.3%, 37.1%, 14.0% and 9.2% of the surveyed genomes. Notably, type I systems were found in all 16 orders of cyanobacteria (Figure 1B), indicating that type I CRIPSR-Cas systems are most prevalent in cyanobacteria.
Additionally, some cyanobacterial genomes harbor more than one type of CRISPR-Cas. Eighty-one cyanobacterial genomes (25.7%) were found to contain both type 1 and III systems, while thirteen genomes contained all three types (type I, III and V) of CRISPR-Cas systems (Figure 1C). Among the identified type I systems, the type I-D system is the most prevalent, accounting for 63.2% of the surveyed genomes, followed by I-B, I-C, I-A, and I-E (Figure 1D).
Normally, the natural type I CRISPR-Cas system’s response to invading DNA can be categorized into three distinct stages: adaptation, processing of CRISPR RNA (crRNA), and interference (Zheng et al., 2020). Synechocystis sp. PCC 6803 (hereafter Syn6803) was chosen as a model for most fundamental researches on cyanobacterial type I system.
During the adaptation stage, a short DNA fragment is captured, processed and integrated into the host’s CRISPR arrays (Figure 2A). The Cas1-Cas2 complex is universally implicated in this process. Besides, Cas4, encoded by a gene typically located adjacent to cas1 and cas2, is also required for the selection of spacers with a specific protospacer adjacent motif (PAM) in certain type I systems (Kieper et al., 2018; Lee et al., 2018; Zhang et al., 2019). A recent structural analysis of the type I-D CRISPR-Cas in Syn6803 showed a two-step assembly mechanism involving these three Cas proteins during the adaptation stage (Wu et al., 2021). Initially, Cas4 forms a stable complex with Cas1, which processed prespacer precursors into their mature form in a PAM-dependent manner. This maturation of prespacers leads to the disassembly of the Cas1-Cas4-prepacer complex and facilitates the assembly of Cas1–Cas2–prespacer complex. Finally, the prespacer, now associated with the Cas1-Cas2 complex, is integrated into the host’s CRISPR array as a new spacer (Wu et al., 2021).
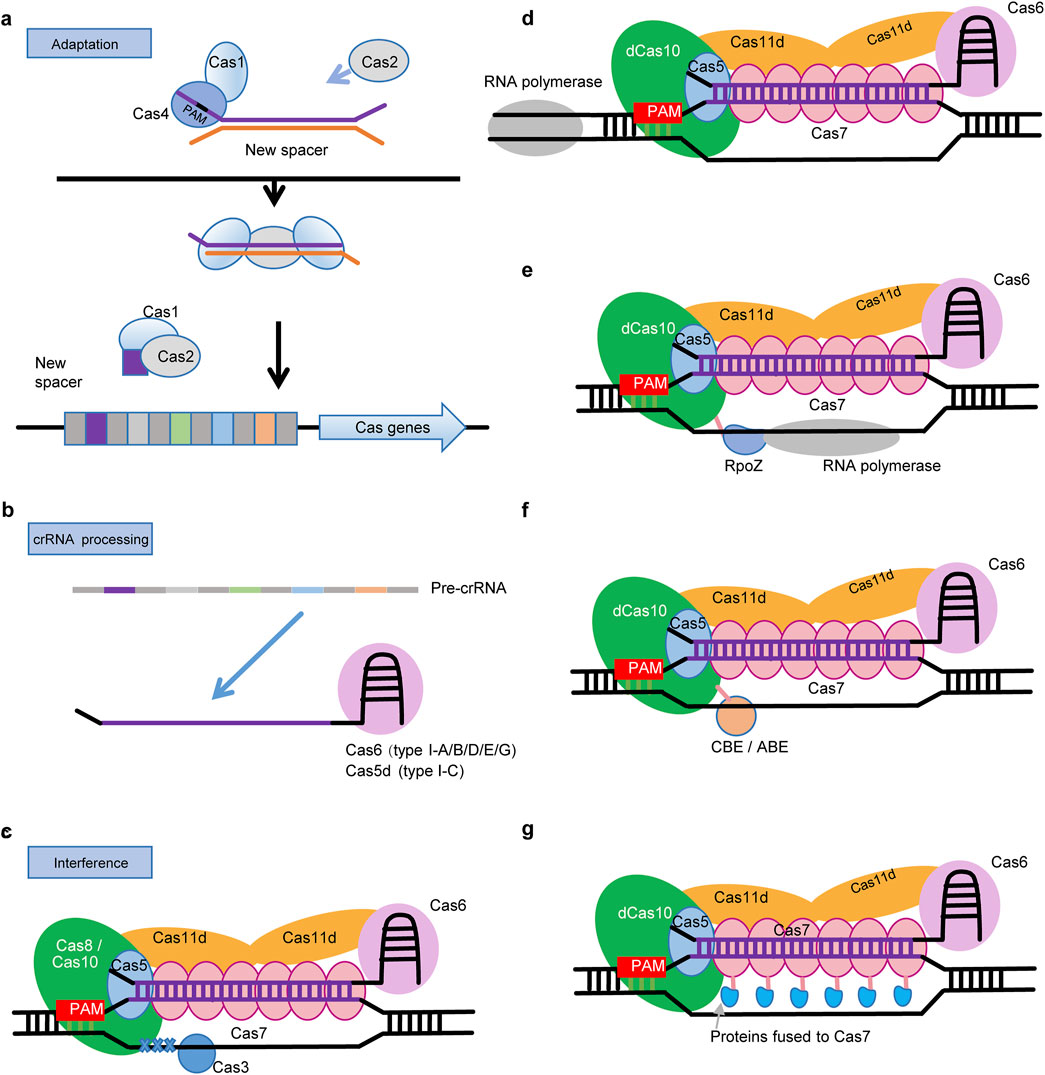
Figure 2. Mechanisms and applications of cyanobacterial type I CRISPR-Cas in genome editing. (A) Adaptation: Cas1 and Cas2 proteins capture a short sequence from an exogenous DNA and integrate it into the CRISPR array as a new spacer. In some type I systems, Cas4 is also required for the selection of spacers. (B) crRNA Processing: Transcription of the CRISPR array generates pre-crRNA with multiple hairpin structures. Cas5d (type I-C) or Cas6 (other type I systems) process the pre-crRNA by binding to its 3′end. (C) Interference: Subsequently, the Cas5 binds to the 5′-handle, and the Cas7 subunit associates with the crRNA to form the structural backbone of the crRNA. In most type I systems, Cas8 binds to Cas5 and recognizes the PAM, Cas3 then initiates cleavage through its nuclease activity. In contrast, the type I-D system lacks Cas8, with Cas10 responsible for both PAM recognition and DNA cleavage. Here, Cas3 exhibits only helicase activity. (D) CRISPRi: In cyanobacterial type I systems, when Cas3 or Cas10 is inactivated, it can bind but not cleave DNA, preventing RNA polymerase progression and resulting in gene silencing. (E) CRISPRa: Inactivated Cas10 (type I-D) or Cas3 (other type I systems) is fused with the ω-subunit (RpoZ) of RNA polymerase. The latter recruits the core complex of RNA polymerase, thus activating the expression of targeted gene. (F) Base Editing: Inactivated Cas10 (type I-D) or Cas3 (other type I systems), without cleavage activity, fused with a cytosine deaminase (CBE) or an adenosine (ABE), targets specific sites for base editing. (G) Besides deficient nucleases, Cas7 should be another choice for fusion expression of RpoZ or deaminases.
In the crRNA processing stage, mature crRNAs are cleaved from pre-crRNA which results from the transcription of CRISPR array (Figure 2B). CRISPR repeats in the pre-cRNA form stable stem-loop structures that are recognized and processed by a RNA endonuclease. Type I-C systems use a unique Cas5 variant, Cas5d (Punetha et al., 2014), while the rest of type I systems use Cas6 for crRNA processing. Each subtype of type I CRISPR-Cas has its own specific stem-loop structures, with variations in stem and loop sizes and sequences. After crRNA processing, Cas6 proteins remain bound to the 3′hairpin structure of the mature crRNA (Zheng et al., 2020). Base on both in vivo and in vitro analyses, Cas6-1 is found to be a specialized RNA endonuclease critical for the maturation of crRNA in the type I-D CRISPR-Cas of Syn6803, exhibiting a single turnover mechanism and requiring a conserved histidine residue for enzymatic activity and positive residues for RNA binding (Scholz et al., 2013; Reimann et al., 2016; Jesser et al., 2019).
In the interference phase, Cas6-crRNA assembles with Cas7, Cas5, and Cas8 to from a CRISPR associated complex for antiviral defense (Cascade) (Figure 2C). In a typical type I Cascade complex (Lu et al., 2024), multiple Cas7 proteins binds around the central part of crRNA, while Cas5 and Cas6 bind to 5′ and 3′-end of crRNA respectively. In most type I systems, a large subunit Cas8 which is normally located adjacent to Cas5 is responsible for recognizing PAM sequences. Besides, the C-terminus of Cas8 and several small subunits (SSU) constitute the inner “belly” of Cascade.
In response to invading DNA, the Cascade complex, guided by base pairing between the embedded crRNA and protospacer, selectively targets invading DNA molecules, thereby forming an R-loop structure. The binding of dsDNA to the Cascade complex induces a conformational change, facilitating the recruitment of Cas3 which is usually composed of an N-terminal HD phosphohydrolase domain and a C-terminal helicase domain (Sinkunas et al., 2011). Finally, Cas3 nicks and then degrades the invading DNA in the presence of ATP.
However, in the cyanobacterial type I-D system, Cas3d lacks the HD nuclease domain and Cas8 is absent. Instead, Cas10d, which harbors a domain of the nuclease, was proved to be responsible for DNA cleavage (Osakabe et al., 2021) and also compensates for the roles of Cas8, mediating PAM recognition instead (Schwartz et al., 2022). Furthermore, the type I-D Cascade incorporates Cas11d small subunits that are derived from an alternative translation initiation site within Cas10d. Cas11d is essential for the specific binding of the I-D Cascade to target double-stranded DNA (dsDNA), as demonstrated by the significantly reduced DNA-binding capacity of the complex lacking Cas11d. And the structure of the I-D Cascade, featuring a helical backbone of Cas7d subunits and a boot-shaped Cas10d large subunit, aligns more closely with the architecture of Type III systems rather than the more curved Cas7 backbone and smaller Cas8 large subunits typical of other Type I systems (McBride et al., 2020).
Cas9 and Cas12a, derived from type II and V CRISPR-Cas systems, are the most widely utilized CRISPR-associated (Cas) effectors for genome editing. Despite their popularity, type I CRISPR-Cas systems are more prevalent in bacteria and archaea (Makarova et al., 2019) and exhibit a more aggressive interaction with DNA targets, which enables them with a distinct advantage in facilitating large-fragment deletions within host genomes (Cameron et al., 2019; Morisaka et al., 2019; Li J. et al., 2024). Up to now, only a limited number of cyanobacterial type I systems have been employed for genome editing in heterologous hosts. Notably, the type I-B system from Synechocystis sp. PCC 6714 has been successfully utilized for genome editing in human T cells, underscoring its potential in human genome editing. This system is characterized by its ability to induce long-spectrum, unidirectional deletions within the human T cells (Lu et al., 2024). Furthermore, the Cascasde of type I-D CRISPR-Cas system, comprising five Cas proteins (Cas3, 5, 6, 7, and 10) from Microcystis aeruginosa, has been heterologously expressed in mammalian cells for targeted genome editing, enabling the introduction of both small deletions and bidirectional large-fragment deletions ranging from 2.5 to 18.5 kb (Osakabe et al., 2021). This system has also demonstrated the capacity to induce short indels and bi-directional long-range deletions in tomato cells (Keishi Osakabe, 2020). Recently, we have successfully developed a genome editing tool based on the endogenous type I-D CRISPR-Cas system of Synechococcus sp. PCC 7002, enabling precise genetic modifications in this organism. Utilizing this tool, we deleted the glgA1 gene and created a double mutant of the glgA1 and glgA2 genes, thereby demonstrating the system’s capability for targeted genome editing. Additionally, we exhibited the tool’s ability to knock out large genomic fragments and to cure the endogenous pAQ5 plasmid, highlighting its potential for genome streamlining (Yang et al., 2024).
These reports prove the potential of endogenous type I CRISPR-Cas systems for precise genome editing. Notably, these native systems alleviate concerns regarding cell toxicity associated with Cas effectors, which often limit the application of the widely used exogenous CRISPR-Cas9 systems (Wendt et al., 2016). Given the prevalence of type I systems in cyanobacterial genomes (Supplementary Table S1), these native systems hold promise for repurposing as efficient genome editing tools for their respective cyanobacterial hosts.
In contrast to genome editing via DNA double-strand breaks (DSB) induced by Cas nucleases, several CRISPR-derived technologies, including CRISPR interference (CRISPRi) (Qi et al., 2013), CRISPR activation (CRISPRa) (Maeder et al., 2013), and base editing (BE) (Komor et al., 2016), can modulate transcriptional levels of target genes or introduce point mutations without DSB. These technologies typically rely on nuclease-deficient Cas9 (dCas9 or nCas9), which can bind but not cleave the double-strand of target DNA. To date, all reports on the application of these strategies in cyanobacteria (Gordon et al., 2016; Huang et al., 2016; Yao et al., 2016; Bourgade et al., 2024; Li X.-D. et al., 2024; Li X. et al., 2024) are based on heterologous type II or V Cas effectors.
To implement these strategies in cyanobacteria using endogenous type I CRISPR-Cas systems, it is necessary to modify cyanobacterial genomes to inactivate the key nuclease subunit of Cascade, specifically Cas10 in type I-D (Osakabe et al., 2021) and Cas3 in other type I systems (Zheng et al., 2019). The binding of nuclease-deficient Cascade impedes the passage of RNA polymerase, thereby achieving highly specific inhibition of gene transcription (CRISPRi) in cyanobacteria (Figure 2D). To enhance the expression levels of specific genes in cyanobacteria, the inactivated nucleases can be further modified to fuse with the omega subunit of RNA polymerase (RpoZ), which recruits the host’s RNA polymerase core enzymes. Thus, RNA polymerase will be guided to the specific gene region, enhancing the transcription of target genes (Figure 2E). For precise nucleotide substitution in specific genomic regions in cyanobacteria, the inactivated nucleases can be fused with a deaminase that mediates C-to-T (CBE) or A-to-G (ABE) substitutions (Figure 2F). It is worthy to note that, Cas7 can also serve as an alternative for fusing RpoZ or deaminases (Figure 2G), in addition to inactivated nucleases, considering the multiple copies of Cas7 within the Cascade complex of type I systems (Guo et al., 2024).
Cyanobacteria have been recognized as a promising class of photosynthetic chassis microorganisms that are able to convert CO2 to bio-chemicals. To improve CO2 conversion efficiency by recombinant cyanobacteria, efficient genetic tools compatible with systematic genetic modifications are essential. In this review, we have analyzed the distribution of CRISPR-Cas systems in completed cyanobacterial genomes to date, summarized the current understanding of the adapted immune mechanisms of native type I systems, and provided future perspectives on the application of these native systems for cyanobacterial genome editing.
The cyanobacteria within the order Synechococcales that lack CRISPR-Cas systems are primarily from the marine families Prochlorococcaceae and Synechococcaceae, which is consistent with the previous survey (Cai et al., 2013). In contrast, seven cyanobacteria from the Merismopediaceae family, which also belong to the Synechococcales order, such as the well-known model Synechocystis sp. PCC 6803 and PCC 6714, do possess CRISPR-Cas systems (Supplementary Table S1), suggesting a divergence within the Synechococcales order regarding the presence of CRISPR-Cas system. Furthermore, a recent survey highlighted a strong association between CRISPR-Cas systems and cyanobacteria thriving in eutrophic and hyper-eutrophic conditions (Park et al., 2024), indicating that the presence or absence of these systems may depend on the ecological niche and evolutionary pressures faced by the cyanobacteria.
Compared to the foreign CRISPR-Cas9 system, endogenous type I CRISPR-Cas systems offer advantages in terms of cell compatibility, long-range editing capabilities, and target specificity (Yoshimi and Mashimo, 2022), making them suitable as species-specific genetic tools for cyanobacterial species harboring these endogenous systems. However, several challenges remain to be addressed to fully realize their potential. First, for some endogenous type I CRISPR-Cas systems with PAM sequences yet to be identified, it is essential to characterize their PAM sequence requirements first. Second, successful gene editing depends critically on efficient gRNA delivery and expression. Both transcriptional levels of gRNA and its structural parameters (sequence design and length optimization) need to be systematically evaluated. Third, due to the polyploid nature of cyanobacterial genomes, careful screening for homozygous mutant strains is essential to ensure both genotypic and phenotypic stability in the edited strains. In addition, further fundamental research on native cyanobacterial CRISPR-Cas systems is necessary, including their off-target effects and responses to foreign DNA, to optimize these endogenous tools for future applications.
YZ: Visualization, Writing–original draft, Writing–review and editing. SY: Writing–review and editing. XZ: Writing–review and editing. XT: Funding acquisition, Visualization, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by the National Natural Science Foundation of China [grant number 31871303 to XT], Open Funding Project of the State Key Laboratory of Biocatalysis and Enzyme Engineering [grant number SKLBEE20220028 to XZ] and Scientific Technology Research and Development Project of Yichang City [grant number A24-1-042 to XZ].
Author XZ was employed by AngelYeast Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fbioe.2025.1552030/full#supplementary-material
Angermayr, S. A., Gorchs Rovira, A., and Hellingwerf, K. J. (2015). Metabolic engineering of cyanobacteria for the synthesis of commodity products. Trends Biotechnol. 33 (6), 352–361. doi:10.1016/j.tibtech.2015.03.009
Bhardwaj, A., Singh, P., Gupta, N., Bhattacharjee, S., Srivastava, A., Parida, A., et al. (2024). “Chapter 20 - cyanobacteria: a key player in nutrient cycling,” in Cyanobacteria. Editors A. K. Mishra,, and S. S. Singh (Academic Press), 579–596.
Bourgade, B., Xie, H., Lindblad, P., and Stensjö, K. (2024). Development of a CRISPR activation system for targeted gene upregulation in Synechocystis sp. PCC 6803. doi:10.1101/2024.10.31.621247
Cai, F., Axen, S. D., and Kerfeld*, C. A. (2013). Evidence for the widespread distribution of CRISPR-Cas system in the Phylum Cyanobacteria. RNA Biol. 10 (5), 687–693. doi:10.4161/rna.24571
Cameron, P., Coons, M. M., Klompe, S. E., Lied, A. M., Smith, S. C., Vidal, B., et al. (2019). Harnessing type I CRISPR-Cas systems for genome engineering in human cells. Nat. Biotechnol. 37 (12), 1471–1477. doi:10.1038/s41587-019-0310-0
Carroll, A. L., Case, A. E., Zhang, A., and Atsumi, S. (2018). Metabolic engineering tools in model cyanobacteria. Metab. Eng. 50, 47–56. doi:10.1016/j.ymben.2018.03.014
Demoulin, C. F., Lara, Y. J., Cornet, L., François, C., Baurain, D., Wilmotte, A., et al. (2019). Cyanobacteria evolution: insight from the fossil record. Free Radic. Biol. Med. 140, 206–223. doi:10.1016/j.freeradbiomed.2019.05.007
Dong, Z., Chen, L., Wang, Y., Sun, T., and Zhang, W. (2024). Current advances in CRISPR-Cas-mediated gene editing and regulation in cyanobacteria. Blue Biotechnol. 1 (No.1), 9. doi:10.1186/s44315-024-00009-3
Doron, S., Melamed, S., Ofir, G., Leavitt, A., Lopatina, A., Keren, M., et al. (2018). Systematic discovery of antiphage defense systems in the microbial pangenome. Science 359 (No.6379), eaar4120. doi:10.1126/science.aar4120
Du, K., Gong, L., Li, M., Yu, H., and Xiang, H. (2022). Reprogramming the endogenous type I CRISPR-Cas system for simultaneous gene regulation and editing in Haloarcula hispanica. mLife 1 (1), 40–50. doi:10.1002/mlf2.12010
Ducat, D. C., Way, J. C., and Silver, P. A. (2011). Engineering cyanobacteria to generate high-value products. Trends Biotechnol. 29 (2), 95–103. doi:10.1016/j.tibtech.2010.12.003
Gordon, G. C., Korosh, T. C., Cameron, J. C., Markley, A. L., Begemann, M. B., and Pfleger, B. F. (2016). CRISPR interference as a titratable, trans-acting regulatory tool for metabolic engineering in the cyanobacterium Synechococcus sp. strain PCC 7002. Metab. Eng. 38, 170–179. doi:10.1016/j.ymben.2016.07.007
Guo, J., Gong, L., Yu, H., Li, M., An, Q., Liu, Z., et al. (2024). Engineered minimal type I CRISPR-Cas system for transcriptional activation and base editing in human cells. Nat. Commun. 15 (1), 7277. doi:10.1038/s41467-024-51695-x
Hou, S., Brenes-Álvarez, M., Reimann, V., Alkhnbashi, O. S., Backofen, R., Muro-Pastor, A. M., et al. (2018). CRISPR-Cas systems in multicellular cyanobacteria. RNA Biol. 16 (4), 518–529. doi:10.1080/15476286.2018.1493330
Huang, C., Shen, C., Li, H., Sung, L., Wu, M., and Hu, Y. (2016). CRISPR interference (CRISPRi) for gene regulation and succinate production in cyanobacterium S. elongatus PCC 7942. Microb. Cell Fact. 15 (No.1), 196–211. doi:10.1186/s12934-016-0595-3
Jesser, R., Behler, J., Benda, C., Reimann, V., and Hess, W. R. (2019). Biochemical analysis of the Cas6-1 RNA endonuclease associated with the subtype I-D CRISPR-Cas system in Synechocystis sp. PCC 6803. RNA Biol. 16 (4), 481–491. doi:10.1080/15476286.2018.1447742
Keishi Osakabe, N. W., Miyaji, T., Murakami, E., Marui, K., Ueta, R., Hashimoto, R., et al. (2020). Genome editing in plants using CRISPR type I-D nuclease. Commun. Biol. 3 (No.1), 648. doi:10.1038/s42003-020-01366-6
Kieper, S. N., Almendros, C., Behler, J., McKenzie, R. E., Nobrega, F. L., Haagsma, A. C., et al. (2018). Cas4 facilitates PAM-compatible spacer selection during CRISPR adaptation. Cell Rep. 22 (13), 3377–3384. doi:10.1016/j.celrep.2018.02.103
Komor, A. C., Kim, Y. B., Packer, M. S., Zuris, J. A., and Liu, D. R. (2016). Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 533 (7603), 420–424. doi:10.1038/nature17946
Lee, H., Zhou, Y., Taylor, D., and Sashital, D. (2018). Cas4-dependent prespacer processing ensures high-fidelity programming of CRISPR arrays. Mol. Cell. 70 (1), 48–59.e5. doi:10.1016/j.molcel.2018.03.003
Li, J., Zhao, D., Zhang, T., Xiong, H., Hu, M., Liu, H., et al. (2024a). Precise large-fragment deletions in mammalian cells and mice generated by dCas9-controlled CRISPR/Cas3. Sci. Adv. 10 (11), eadk8052. doi:10.1126/sciadv.adk8052
Li, X., Wei, Y., Wang, S.-Y., Wang, S.-G., and Xia, P.-F. (2024c). One-for-all gene inactivation via PAM-independent base editing in bacteria. J. Biol. Chem. 301, 108113. doi:10.1016/j.jbc.2024.108113
Li, X.-D., Liu, L.-M., Xi, Y.-C., Sun, Q.-W., Luo, Z., Huang, H.-L., et al. (2024b). Development of a base editor for convenient and multiplex genome editing in cyanobacteria. Commun. Biol. 7 (1), 994. doi:10.1038/s42003-024-06696-3
Lu, M., Yu, C., Zhang, Y., Ju, W., Ye, Z., Hua, C., et al. (2024). Structure and genome editing of type I-B CRISPR-Cas. Nat. Commun. 15 (1), 4126. doi:10.1038/s41467-024-48598-2
Maeder, M. L., Linder, S. J., Cascio, V. M., Fu, Y., Ho, Q. H., and Joung, J. K. (2013). CRISPR RNA-guided activation of endogenous human genes. Nat. Methods. 10 (10), 977–979. doi:10.1038/nmeth.2598
Makarova, K. S., Wolf, Y. I., Iranzo, J., Shmakov, S. A., Alkhnbashi, O. S., Brouns, S. J. J., et al. (2019). Evolutionary classification of CRISPR–Cas systems: a burst of class 2 and derived variants. Nat. Rev. Microbiol. 18 (No.2), 67–83. doi:10.1038/s41579-019-0299-x
McBride, T. M., Schwartz, E. A., Kumar, A., Taylor, D. W., Fineran, P. C., and Fagerlund, R. D. (2020). Diverse CRISPR-Cas complexes require independent translation of small and large subunits from a single gene. Mol. Cell. 80 (6), 971–979.e7. doi:10.1016/j.molcel.2020.11.003
Morisaka, H., Yoshimi, K., Okuzaki, Y., Gee, P., Kunihiro, Y., Sonpho, E., et al. (2019). CRISPR-Cas3 induces broad and unidirectional genome editing in human cells. Nat. Commun. 10 (1), 5302. doi:10.1038/s41467-019-13226-x
Ni, J., Tao, F., Xu, P., and Yang, C. (2018). Engineering cyanobacteria for photosynthetic production of C3 platform chemicals and terpenoids from CO2. Adv. Exp. Med. Biol. 1080, 239–259. doi:10.1007/978-981-13-0854-3_10
Osakabe, K., Wada, N., Murakami, E., Miyashita, N., and Osakabe, Y. (2021). Genome editing in mammalian cells using the CRISPR type I-D nuclease. Nucleic Acids Res. 49 (No.11), 6347–6363. doi:10.1093/nar/gkab348
Park, H., Bulzu, P. A., Shabarova, T., Kavagutti, V. S., Ghai, R., Kasalický, V., et al. (2024). Uncovering the genomic basis of symbiotic interactions and niche adaptations in freshwater picocyanobacteria. Microbiome 12 (1), 150. doi:10.1186/s40168-024-01867-0
Pattharaprachayakul, N., Lee, M., Incharoensakdi, A., and Woo, H. M. (2020). Current understanding of the cyanobacterial CRISPR-Cas systems and development of the synthetic CRISPR-Cas systems for cyanobacteria. Enzyme Microb. Technol. 140, 109619. doi:10.1016/j.enzmictec.2020.109619
Punetha, A., Sivathanu, R., and Anand, B. (2014). Active site plasticity enables metal-dependent tuning of Cas5d nuclease activity in CRISPR-Cas type I-C system. Nucleic Acids Res. 42 (6), 3846–3856. doi:10.1093/nar/gkt1335
Qi, L. S., Larson, M. H., Gilbert, L. A., Doudna, J. A., Weissman, J. S., Arkin, A. P., et al. (2013). Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 152 (5), 1173–1183. doi:10.1016/j.cell.2013.02.022
Reimann, V., Alkhnbashi, O. S., Saunders, S. J., Scholz, I., Hein, S., Backofen, R., et al. (2016). Structural constraints and enzymatic promiscuity in the Cas6-dependent generation of crRNAs. Nucleic Acids Res. 45 (2), 915–925. doi:10.1093/nar/gkw786
Russel, J., Pinilla-Redondo, R., Mayo-Muñoz, D., Shah, S. A., and Sørensen, S. J. (2020). CRISPRCasTyper: automated identification, annotation, and classification of CRISPR-Cas loci. CRISPR J. 3 (6), 462–469. doi:10.1089/crispr.2020.0059
Scholz, I., Lange, S. J., Hein, S., Hess, W. R., and Backofen, R. (2013). CRISPR-Cas systems in the cyanobacterium Synechocystis sp. PCC6803 exhibit distinct processing pathways involving at least two Cas6 and a Cmr2 protein. PLoS One 8 (2), e56470. doi:10.1371/journal.pone.0056470
Schwartz, E. A., McBride, T. M., Bravo, J. P. K., Wrapp, D., Fineran, P. C., Fagerlund, R. D., et al. (2022). Structural rearrangements allow nucleic acid discrimination by type I-D Cascade. Nat. Commun. 13 (1), 2829. doi:10.1038/s41467-022-30402-8
Sinkunas, T., Gasiunas, G., Fremaux, C., Barrangou, R., Horvath, P., and Siksnys, V. (2011). Cas3 is a single-stranded DNA nuclease and ATP-dependent helicase in the CRISPR/Cas immune system. EMBO J. 30 (No.7), 1335–1342. doi:10.1038/emboj.2011.41
Strecker, J., Ladha, A., Gardner, Z., Schmid-Burgk, J. L., Makarova, K. S., Koonin, E. V., et al. (2019). RNA-guided DNA insertion with CRISPR-associated transposases. Science 365 (No.6448), 48–53. doi:10.1126/science.aax9181
Sun, T., Li, S., Song, X., Diao, J., Chen, L., and Zhang, W. (2018). Toolboxes for cyanobacteria: recent advances and future direction. Biotechnol. Adv. 36 (4), 1293–1307. doi:10.1016/j.biotechadv.2018.04.007
Vijay, D., Akhtar, M. K., and Hess, W. R. (2019). Genetic and metabolic advances in the engineering of cyanobacteria. Curr. Opin. Biotechnol. 59, 150–156. doi:10.1016/j.copbio.2019.05.012
Wang, L., Chen, L., Yang, S., and Tan, X. (2020). Photosynthetic conversion of carbon dioxide to oleochemicals by cyanobacteria: recent advances and future perspectives. Front Microbiol. 11, 634. doi:10.3389/fmicb.2020.00634
Wendt, K. E., Ungerer, J., Cobb, R. E., Zhao, H., and Pakrasi, H. B. (2016). CRISPR/Cas9 mediated targeted mutagenesis of the fast growing cyanobacterium Synechococcus elongatus UTEX 2973. Microb. Cell Fact. 15 (1), 115. doi:10.1186/s12934-016-0514-7
Wu, C., Tang, D., Cheng, J., Hu, D., Yang, Z., Ma, X., et al. (2021). Mechanisms of spacer acquisition by sequential assembly of the adaptation module in Synechocystis. Nucleic Acids Res. 49 (5), 2973–2984. doi:10.1093/nar/gkab105
Xu, Z., Li, Y., Li, M., Xiang, H., and Yan, A. (2021). Harnessing the type I CRISPR-Cas systems for genome editing in prokaryotes. Environ. Microbiol. 23 (2), 542–558. doi:10.1111/1462-2920.15116
Yang, S., Zhang, Y., Li, C., and Tan, X. (2024). Repurposing endogenous Type I-D CRISPR-Cas system for genome editing in Synechococcus sp. PCC7002. Microbiol. Res. 288, 127884. doi:10.1016/j.micres.2024.127884
Yao, L., Cengic, I., Anfelt, J., and Hudson, E. P. (2016). Multiple gene repression in cyanobacteria using CRISPRi. Acs Synth. Biol. 5 (3), 207–212. doi:10.1021/acssynbio.5b00264
Yoshimi, K., and Mashimo, T. (2022). Genome editing technology and applications with the type I CRISPR system. Gene. Genome. 3, 100013. doi:10.1016/j.ggedit.2022.100013
Zhang, Z., Pan, S., Liu, T., Li, Y., and Peng, N. (2019). Cas4 nucleases can effect specific integration of CRISPR spacers. J. Bacteriol. 201 (12), e00747. doi:10.1128/jb.00747-18
Zheng, Y., Han, J., Wang, B., Hu, X., Li, R., Shen, W., et al. (2019). Characterization and repurposing of the endogenous Type I-F CRISPR-Cas system of Zymomonas mobilis for genome engineering. Nucleic Acids Res. 47 (21), 11461–11475. doi:10.1093/nar/gkz940
Zheng, Y., Li, J., Wang, B., Han, J., Hao, Y., Wang, S., et al. (2020). Endogenous type I CRISPR-Cas: from foreign DNA defense to prokaryotic engineering. Front. Bioeng. Biotech. 8, 62. doi:10.3389/fbioe.2020.00062
Keywords: type I CRISPR-Cas, cyanobacterium, genome editing, synthetic biology, metabolic engineering
Citation: Zhang Y, Yang S, Zheng X and Tan X (2025) Cyanobacterial type I CRISPR-Cas systems: distribution, mechanisms, and genome editing applications. Front. Bioeng. Biotechnol. 13:1552030. doi: 10.3389/fbioe.2025.1552030
Received: 27 December 2024; Accepted: 10 February 2025;
Published: 27 February 2025.
Edited by:
Stephan Klähn, Helmholtz Association of German Research Centres (HZ), GermanyReviewed by:
Tao Sun, Tianjin University, ChinaCopyright © 2025 Zhang, Yang, Zheng and Tan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoming Tan, eGlhb21pbmcudGFuQGh1YnUuZWR1LmNu; Xianliang Zheng, emhlbmd4bEBhbmdlbHllYXN0LmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.