
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Bioeng. Biotechnol., 24 February 2025
Sec. Synthetic Biology
Volume 13 - 2025 | https://doi.org/10.3389/fbioe.2025.1478413
 Thais Torquato Sales1,2†
Thais Torquato Sales1,2† Marco Antônio de Oliveira1,2†
Marco Antônio de Oliveira1,2† Lilian Hasegawa Florentino1,2
Lilian Hasegawa Florentino1,2 Rayane Nunes Lima1
Rayane Nunes Lima1 Elibio Rech1*
Elibio Rech1*Serine integrases (Ints) have gained prominence and have been extensively used in Synthetic Biology due to their ability to modify DNA sequences. Ints are recombinases encoded by the phage genome and have been used to unidirectionally catalyze an insertion, excision, or inversion of a specific DNA sequence between the two attachment sites (att) attB (bacterial attachment site) and attP (phage attachment site). The entire process is highly specific and accurate; therefore, Ints are widely used in genetic engineering and have been extensively studied due to their unique site-specific recombination properties and potential genome editing applications. Furthermore, new recombinational factors (RDFs) and their determinants are constantly being discovered, underlining the need to update progress in research involving Ints in eukaryotic cells. In this way, this review aims to provide an overview of Ints in eukaryotic cells and highlight how Ints can be used in innovative ways to advance genetic engineering applications in health, agriculture, and environmental sciences.

GRAPHICAL ABSTRACT | Moving beyond bacteria: unraveling the mechanisms of phage serine integrases in eukaryotic cells to drive biotechnological research, genome editing and therapeutic applications. Created with Canva Pro.
Viruses are the most numerous microorganisms and phages are the reservoir of most of the genetic diversity in the sea, with about 1030 particles present in the oceans alone (Suttle, 2007). In addition, they can infect a wide variety of prokaryotes and can be found in soil, ocean, and extreme environments (Chibani-Chennoufi et al., 2004; Mills et al., 2013). Phages have great biotechnological relevance due to the multifunctionality of their enzymes and environmental relevance by altering cellular metabolism and influencing the cycling of chemical elements such as nitrogen, oxygen, and carbon (Warwick-Dugdale et al., 2019; Wang et al., 2022; Bisen et al., 2024). Moreover, these viruses have different characteristics in size, morphology, and genomic organization (Hatfull and Hendrix, 2011; Simmonds and Aiewsakun, 2018; Zhu et al., 2024). Phages are classified according to their morphological characteristics, their genetic material content, the location where they can be found and the bacterial species they can infect (Hatfull and Hendrix, 2011). They all comprise a DNA genome encased in a shell of phage-encoded capsid proteins, which protect the genetic material and mediate its delivery to the next host cell (Grigson et al., 2023; Turner et al., 2024). With the advancement of technology, electron microscopy allowed the detailed visualization of several types of phages, some of which appear to have “heads”, “legs” and “tails” (Figure 1A). Despite this appearance, phages are immobile and rely on Brownian motion or pedesis to reach their target host (Kasman and Porter, 2022). As a result of billions of years of coevolution between phages and their prokaryotic hosts, various mechanisms of attack and defense have been developed by both (Bernheim and Sorek, 2018). In nature, this coevolution leads to a constant “conflict” between the two groups, with the bacteriophages trying to find ways to infect the bacteria and the bacteria trying to defend themselves against the phages (Piel et al., 2022; Butt et al., 2024; Patel et al., 2024; Siedentop et al., 2024), although their dynamic interactions are still not fully understood (Sabino et al., 2020). This can result in a wide variety of bacteriophages and bacteria, each adapted to a particular environment or host. Phages are identified into two groups: lytic, or virulent, and lysogenic, or temperate, phages (Baaziz et al., 2024). Its biological cycle involves bacterial adhesion and invasion. As an obligatory intracellular parasite of a bacterial cell, phages can replicate through the biological cycles in the prokaryotic cell: lytic and lysogenic (Figure 1). The bacteriophage lytic cycle significantly affects bacterial mortality and nutrient cycling, as at the end of the process virions are released that can infect other bacterial cells and the host cell is destroyed. However, the size of the virion burst can vary significantly depending on the characteristics of the phage, the bacterium against which the bacteriophage is directed, and the environments in which the bacteriophage-host relationship occurs (Weinbauer, 2004). Lysogenic activity, on the other hand, is characterized by the integration of the viral genetic material (prophage) into the host chromosome and, when the cell divides, the transmission of the viral genetic material to the daughter cells and the host can lead to the distribution of beneficial characteristics for their hosts, such as resistance to antibiotics and phages or increased virulence (Bailey et al., 2024; Rostøl et al., 2024). However, under certain conditions, temperate phages can either multiply via the lytic cycle or remain dormant in the host cell (Young, 2013; Fogg et al., 2014; Doore and Fane, 2016). Lysogenic activity is less common than the lytic cycle, but it can be important in some cases, such as in the spread of genes between bacteria. The phage genome integration mechanism is mediated by phage-encoded integrases which, with no need of any other phage-encoded factors, catalyze highly site-specific, unidirectional recombination reactions (Figure 1B). Both integration and excision require Large Serine Recombinase (LSRs), the enzyme that mediates site-specific DNA recombination. LSRs are characterized as bacteriophage integrases responsible for inserting viral DNA into the bacterial chromosome and excising host DNA (Groth et al., 2004; Smith, 2015). This review is designed to deliver a comprehensive exploration of Large Serine Integrases (Ints), encompassing their origins, discovery, functionality, structure, and applications. The fundamental objective is to amplify the existing repertoire of proteins within this class capable of regulating gene expression in eukaryotic cells. Lastly, by providing a robust and inclusive analysis, this review will offer vital insights, engender further research, and potentially pave the way for novel practical applications of LSRs in the realm of gene expression control.
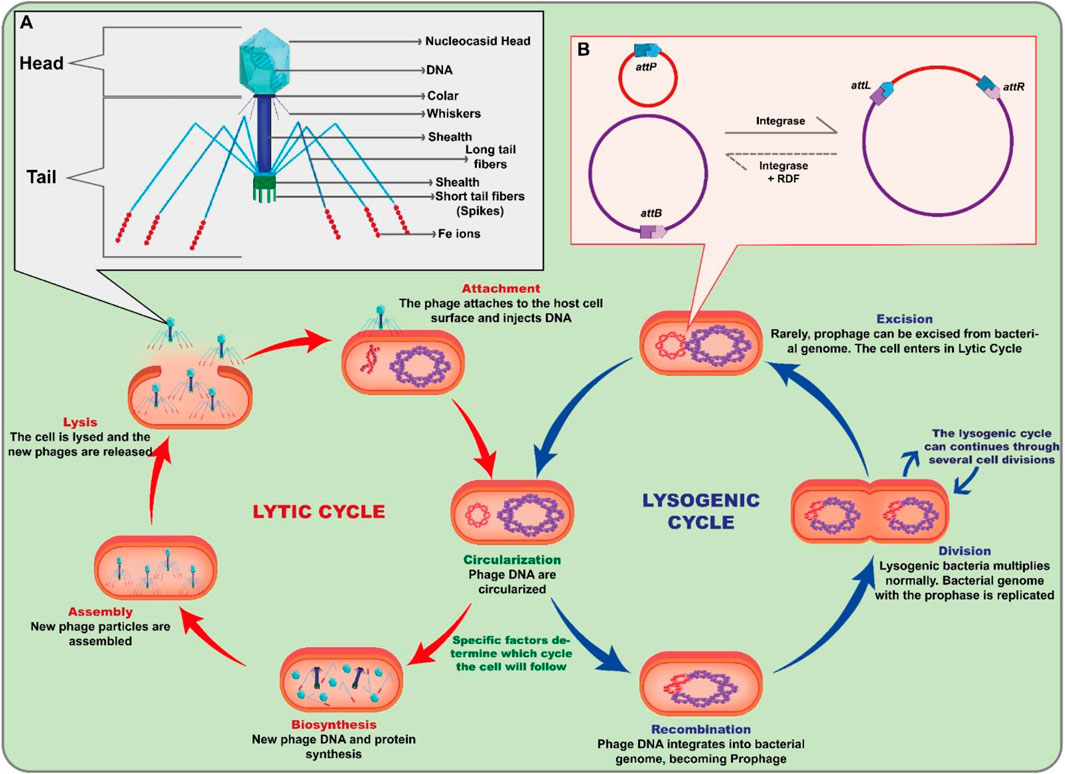
Figure 1. Bacteriophage life cycle and importance of serine-integrases in the process (A) The bacteriophage has a structure composed of a protein capsule that surrounds its genetic material. The tail of the phage functions in specific binding and entry of the virus into the host bacterium. It consists of a long tubular structure called a sheath, which connects to the phage head. At the end of the tail, specialized structures such as fibers or fibrils are found, allowing the phage to bind to specific receptors on the surface of the host bacterium. During the lytic cycle, the bacteriophage attaches to the host cell and injects its genetic material, which takes control of the cellular machinery, resulting in the production of new viruses and lysis of the host cell, releasing the newly formed viruses. In the lysogenic cycle, instead of immediately initiating the lytic process, the genetic material of the bacteriophage integrates into the bacterial genome, forming a structure called a prophage. (B) Scheme of site-specific recombination mediated by the phage in the bacterial host. The integrase performs precise recombination between an attB (bacterial attachment site) and an attP (phage attachment site). The result is the integration of the phage into the host genome and the formation of hybrid sequences attL (attachment Left site) and attR (attachment Right site).
Unlike homologous recombinases that facilitate recombination between similar sequences, site-specific recombinases (SSRs) are a group of enzymes specialized in promoting recombination exclusively between specific recognition sites. SSRs encompass a series of recombination processes involving mutual exchanges between defined sites in the DNA sequence (Grindley et al., 2006; Wang et al., 2011). These recombinases are present in various hosts and play crucial regulatory roles. Among the most commonly used are those found in phages, such as Cre, λ integrase, phiC31 integrase, Bxb1 integrase, and Flippase recombinase (FLP) in the yeast Saccharomyces cerevisiae. SSR’s (Site-specific recombinase systems) have been discovered in bacteria and yeast to promote a number of functions, including the phase shift of certain bacterial virulence factors and the integration of bacteriophages into the host genome (Wang et al., 2011). The recombinase superfamily can be divided into two fundamental groups, based on the active amino acid within the catalytic domain of the enzymes in each family: the tyrosine (Tyr) and the serine (Ser) recombinases (Gaj et al., 2014). Families can be further subdivided into members based on size or the mechanisms used (Wang et al., 2011). The family of tyrosine recombinases are the most widespread among prokaryotes (Bacteria and Archaea), and are also found in eukaryotes, sharing a catalytic domain with an easily recognized motif. Tyrosine recombinases can be bidirectional or unidirectional. Bidirectional recombination occurs between two identical sites, while unidirectional ones act at different attB and attP sites, resulting in an irreversible recombination. This reaction can be reversed in the presence of accessory (helper) proteins known as excisionases (Wang et al., 2011). The excision mechanism of this type of integrase is based on the formation of an intermediate state of the four DNA strands known as Holliday junction, similar to what occurs in homologous recombination (Grindley et al., 2006). The most studied phage tyrosine recombinases are the integrases present in lambda phage, Cre-lox, R-RS, HK101, pSAM2 and FLP-FRT systems (Wang et al., 2011). The serine family integrases that perform unidirectional recombination need accessory proteins present on the phage to reverse the reaction. This integrase group binds to the phage DNA strand at the attP site and bacterial DNA at the attB site and thus promotes double-strand breakage concomitantly. In this way the viral DNA is integrated into the bacterial genome with the formation at the left end of the attL site and the attR site at the right end. Several serine-type integrases have been described, such as: phiC31, Bxb1, phiBT1, TP901, R4, MR11, A118, phiK38, Wβ e SPBC (Brown et al., 2011).
Integration and excision reactions are remarkable because of their simplicity and high level of directionality. Serine integrases recombine substrates containing a phage attachment site (attP, attachment Phage or phage binding site) and a bacterial site (attB, attachment Bacteria or bacterium attachment site), with each integrase being able to act only on their specific recombination site pairs. Integrase recombination of att sites results in two hybrid sites, attL (attachment Left or left binding site) and attR (attachment Right or right binding site), each consisting of half attP sequence and half attB sequence. Depending on the orientation and location of the recombination sites within the same DNA molecule or in separate molecules, whether they are linear or circular, it is possible to perform various reactions. These reactions include integration, which involves the insertion of a DNA sequence (cassette) into a specific site within a DNA molecule; deletion, which involves the excision of a DNA sequence from a molecule; inversion, which refers to the inversion of a specific DNA sequence, altering its orientation; and RMCE (Recombinase-Mediated Cassette Exchange), which allows the replacement of a previously inserted DNA cassette with a new sequence from a circular DNA containing the gene of interest flanked by two copies of the same att site, allowing integration of fragments while still preserving the integrity of the remaining genome sequence. A diagram showing the recombination steps and outcomes for different designs is presented in Figure 2.
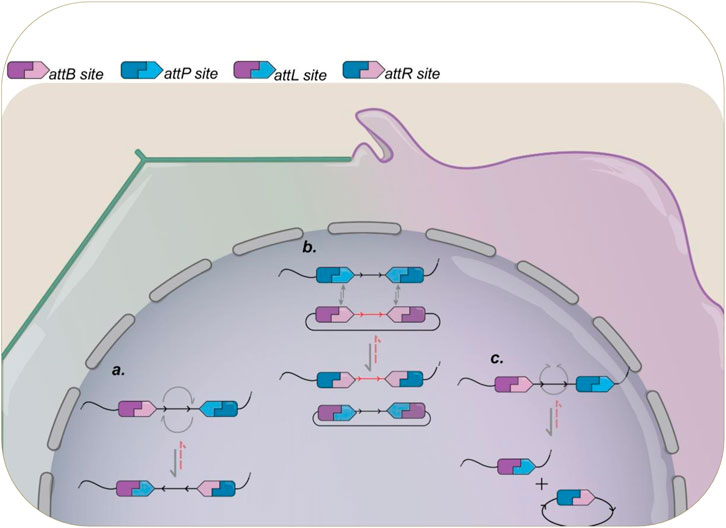
Figure 2. Schematic illustration of recombination activity promoted by serine integrases and the different possible outcomes. Already used in a variety of organisms, the outcome of a serine-integrase rearrangement can be controlled by the synthetic design of their attB/P sites. (A) when both sites are present in the same molecule, but with opposing orientation, their recombination will lead to an 180° inversion of the DNA sequence flanked by them. (B) Recombinase Mediated Cassette Exchange (RMCE) can be achieved when duplicates of each att site is present in different molecules. Upon recombination, the DNA sequences flanked will be swapped between the two constructions. (C) Different from DNA inversion, when both att sites are present in the same molecule, but presenting the same orientation, the recombination will result in excision of the target sequence, with formation of attL in the original molecule plus a circular DNA containing the excised DNA and the attR site formed. In all instances, the reverse reaction (red dotted lines) is only possible in the presence of cognate RDF.
Although initially unidirectional, the presence of an additional protein encoded by the phage, called recombination directionality factor (RDF) can reverse the reaction by activating attL x attR recombination and inhibiting attP x attB recombination (Olorunniji et al., 2016; Olorunniji et al., 2017; Olorunniji et al., 2019). The identification of RDF proteins has proven to be a complex challenge. Studies have used artificial intelligence to analyze the degree of orthogonality in integrative and excisive responses of serine integrases, such as ϕC31, ϕBT1, and TG1, along with their respective RDFs. The results highlighted TG1 as the most active integrase with greater directionality (MacDonald et al., 2024). Among the characterized proteins, there are few homologous sequences, size variation, and a wide diversity in gene loci (Fogg et al., 2017). Some of the serine integrases and their respective known RDFs are phiC31 – gp3 (Fogg et al., 2018); Bxb1 – gp47 (Ghosh et al., 2006); TP901-1 – orf7 (Breüner et al., 1999); phiRv1 – Rv1584c (Bibb and Hatfull, 2002); SPBc – SprB (Abe et al., 2014); phiJoe – gp52 (Fogg et al., 2017); A118 – gp44 (Mandali et al., 2017); phiBT1 – gp3BT1 (Zhang et al., 2013). Moreover, a variety of Ints were identified, allowing the use of several of them concomitantly, and in this way, allowing the construction of genetic circuits (Bonnet et al., 2013). In this article, the authors report the construction of six logic gates (Boolean gates) in Escherichia coli using serine integrases Bxb1 and TP901 to control the orientation of terminators and promoters and, thus, allow or block the expression of the gfp reporter gene. A robust database search for other integrases was performed by Yang et al. (2014), finding 34 new ones and their cognate attP and attB sites, of which 11 were functional in E. coli DH10b, with orthogonal property, that is, one integrase does not recognize the site of another (Yang et al., 2014). Furthermore, they demonstrated that several integrases can be used sequentially in the same DNA segment to change the state of a transcriptional logic gate (Yang et al., 2014). A software for the design of genetic circuits, named Cello, was developed by Nielsen et al. (2016), which made it possible to test 60 serine integrases in E. coli, of which 45 worked as expected (Nielsen et al., 2016). However, most works are focused on prokaryotic cells. Although the natural environment of serine integrases is prokaryotic, these enzymes have been shown to be active in a variety of eukaryotic organisms (Gomide et al., 2020).
The large serine recombinase family includes a diverse range of enzymes (Grindley et al., 2006; Smith, 2015). There are over 500 types of serine-integrase structures identified so far, and many domains remain unknown (DUFF domain) (Wang et al., 2023). However, the serine integrases commonly used in biotechnology have three well-known domains: Ser_recom [pfam 00239]; Recombinase [pfam 07508] and Zn-ribbon [pfam13408] (Figure 3). Furthermore, note that only some integrases have the Zn-ribbon domain (Figure 3). This is due to the presence or absence of four zinc-binding cysteine residues (blue stars), but all integrases are functional (Figure 4) (McEwan et al., 2011). The structural and biochemical features of these recombinases have been studied in detail to understand the mechanisms of their action and to develop novel tools for genetic engineering (Keravala et al., 2006; Bonnet et al., 2012; Van Duyne and Rutherford, 2013; Fogg et al., 2014; Rutherford and Van Duyne, 2014; Fan et al., 2016; Fogg et al., 2017; Stark, 2017). The tridimensional protein structures of a few serine integrases have been determined using X-ray crystallography and electron microscopy techniques (Ghosh et al., 2006; Rutherford et al., 2013; Gupta et al., 2017; Li et al., 2018; Mandali and Johnson, 2021). These structures reveal that the integrases adopt a homodimeric conformation with a conserved catalytic domain that contains the active site serine residue. The catalytic domain is connected to a DNA-binding domain, which recognizes the specific recombination sites in the DNA molecule. The enzyme is composed of two domains: N-terminal catalytic domain (NTD) and a C-terminal DNA binding domain (CTD) (McEwan et al., 2011). The catalytic domain contains a conserved serine residue that is essential for the recombination reaction, while the DNA binding domains recognize specific DNA sequences for recombination (att sites) (McEwan et al., 2009). In Figure 5A it is possible to observe the presence of conserved residues (yellow triangles): the N-terminal catalytic motif RxS … (S/D) RxxR. Moreover, these integrases have a flexible hinge region that allows for the formation of different conformations during the catalytic cycle (Figure 5B). This flexibility enables the integrase to adopt different configurations for the recognition and binding of DNA substrates, leading to the formation of different recombination products. Thus, serine integrases recognize and binds both attB and attP sites, then catalyzes a reaction that results in the integration, inversion or excision of the adjacent DNA. Further studies on the structure and function of these integrases will help in the development of new technologies for genetic manipulation and gene therapy.

Figure 3. The phylogenetic tree and schematic representation of the domain architecture of the serine integrases. Phylogenetic tree of prophage integrases inferred using the Serine recombinase domain amino acid sequence and schematic representation of the conserved domains of the full-length integrases. Each domain is represented on scale. Ser_recom represents the serine recombinase domain and Zn_rib represents the recombinase zinc beta ribbon domain.
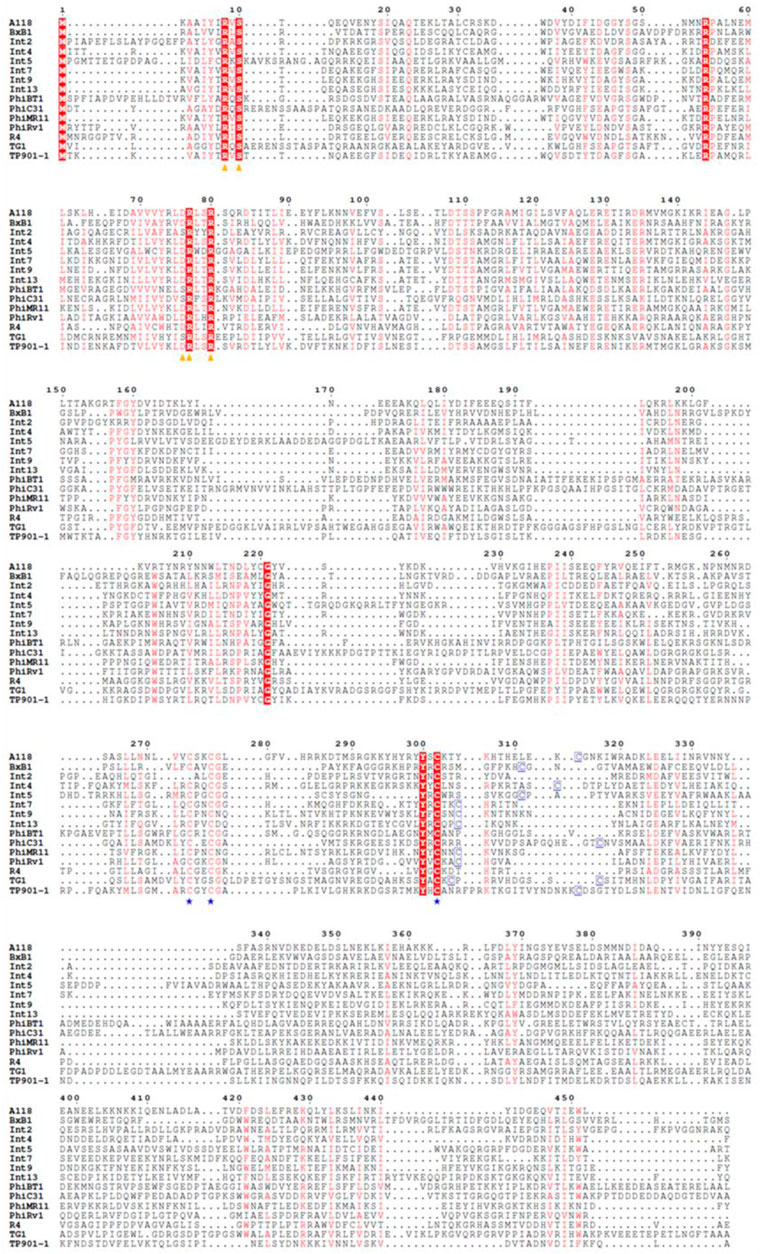
Figure 4. Amino acid sequence alignment of distinct Serine Integrases. Important catalytic domain residues are indicated with yellow triangles and cysteine residues are indicated with blue stars/squares. Conserved residues are highlighted in red.
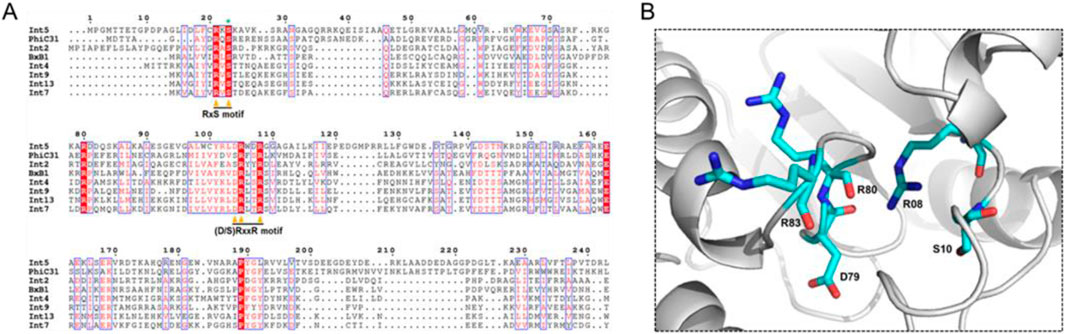
Figure 5. Representation of the three-dimensional structure of the N-terminal catalytic domain of Integrase 13 (Int13). (A) Amino acid sequence alignment of the N-terminal portion of different Serine Integrases. Important catalytic domain residues are indicated with triangles. Serine nucleophile (S10) is indicated with the green asterisk. Conserved residues are highlighted in red. (B) Structural homology modeling of the catalytic motif architecture: RxS… (D/S) RxxR of Int13.
Among serine recombinases, serine integrases are considered a particularly fascinating subfamily. Pioneering studies with these enzymes were conducted to modify the genomes of Streptomyces bacteria at specific sites known as attB. Using serine integrase, researchers were able to stably integrate the pSAM2 plasmid into the bacterial chromosome at a predetermined location. This successful integration enabled the transmission of the integrated plasmid to descendant cells, expanding the possibilities for the application of these biotechnological tools (Merrick et al., 2018; Abioye et al., 2023). Over the course of more than 30 years, a wide variety of applications utilizing serine integrases in various eukaryotic organisms, including human cells can be found in the literature (Fogg et al., 2014; Snoeck et al., 2019). The use of these proteins has been applied in various ways, such as the removal of transgenes or the insertion of desired sequences at specific sites, referred to in these cases as “anchoring sites” or “landing platforms.” Additionally, these proteins are employed in cassette exchange mechanisms, DNA assembly, and in the activation or deactivation of gene expression through sequence inversion, enabling the construction of genetic switches (Groth et al., 2000; Stark, 2017). The main serine-integrases used for DNA editing in various eukaryotic organisms and the rearrangement mechanism applied are presented in Table 1.
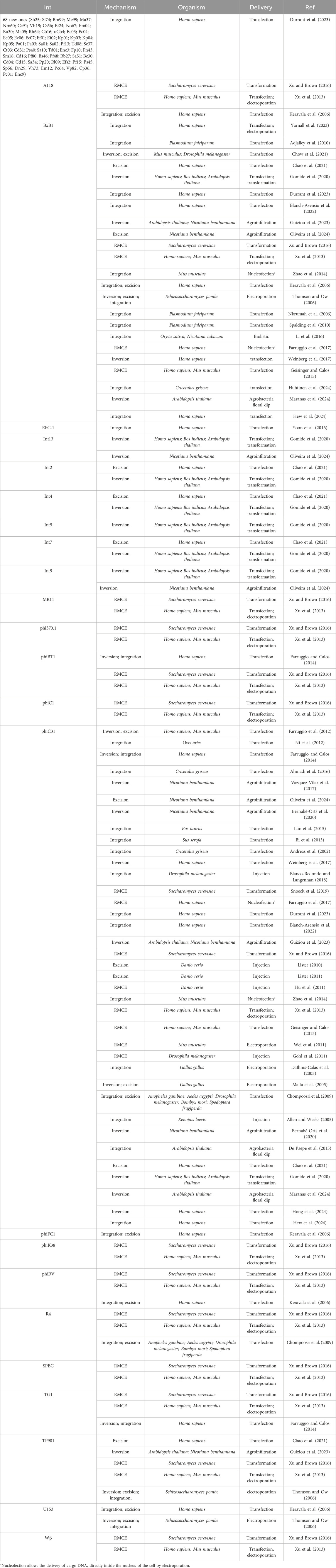
Table 1. The main serine-integrases used for DNA editing in distinct eukaryotic cells.
Saccharomyces cerevisiae is a widely studied model organism in biotechnology due to its ease of use, short life cycle, and expression capabilities. It can be used for the production of biofuels, metabolic engineering, and as a model for studying human diseases. phiC31, a type of serine integrase, has been utilized in the yeast S. cerevisiae to reintroduce specific markers, such as HIS5 and LEU2, through a technique called SIRE (Serine Integrase Recombinational Engineering). However, it has been observed that the efficiency of this system is restricted to low-copy vectors that carry the phiC31 integrase gene, likely because the enzyme exhibits toxic effects (Snoeck et al., 2019). In addition to the serine integrase phiC31 in yeast cells, the integrases ϕBT1, TP901, R4, Bxb1, MR11, A118, ϕK38, phiC31, Wβ, and SPBC demonstrated activity when expressed by S. cerevisiae. The authors conducted tests with fourteen serine integrases, some containing a nuclear localization signal (NLS), and observed that these enzymes were more efficient in promoting genomic integration reactions than simple homologous recombination. The group also noted that the presence of a nuclear localization signal (NLS) caused toxicity in the recombinases TG1 and BxB1. The most plausible explanation for these results was that the integrase in question was interacting with the nuclear genome in some way, becoming toxic. Finally, in the case of the recombinases Wβ and BL3, it was observed that the effect of the nuclear localization signal (NLS) was to reduce the toxicity of the integrase. This can be explained by the fact that these integrases do not rely on the NLS to enter the nucleus, and the presence of the NLS may compromise their ability to interact with the genome. This may occur due to a reduction in the integrase’s binding to ectopic recombination sites (Xu and Brown, 2016). Thomson and Ow (2006) selected three integrases from the extensive serine subfamily: Bxb1, TP901-1, and U153 (Thomson and Ow, 2006). This system has been shown capable of performing excision, inversion, and integration reactions in strains of Schizosaccharomyces pombe, such as FY527. In conclusion of the study, the functionality of these recombination systems in yeast could be applied, for instance, to invert DNA sequences and regulate gene expression. Given the potential of these systems in yeast, authors suggested they could be developed as tools for performing specific rearrangements in the genomes of plants and animals (Thomson and Ow, 2006).
An important work with serine integrases in mammalian organisms involved experiments with fifteen integrases in human fibrosarcoma HT1080 cells and mouse embryonic stem cells. Initially, authors performed assays with E. coli in the presence of a reporter plasmid to confirm the activity of each integrase and its cognate binding sites by expressing each integrase gene in bacterial strains. The results confirmed that all the integrases and their binding sites were active in E. coli (phiC31, Bxb1, φBT1, φC1, MR11, TP901–1, R4, A118, φRV, TG1, φ370.1, Wβ, BL3, SPBc and K38). However, only four integrases (Bxb1, phiC31, R4, and φBT1) have demonstrated the ability to mediate precise integration at a specific site of the genomic DNA in HT1080 and ES cells. The results indicated that the Bxb1 integrase is the most efficient and precise, followed by phiC31, while the R4 and φBT1 integrases exhibited lower efficiency and precision (Xu et al., 2013). Weinberg et al. (2017), using different recombinase enzymes, designed 113 genetic circuits for eukaryotic cells, such as human embryonic kidney cells (HEK293T) and Jukart T cells, achieving 96.5% functionality. These data demonstrate the ability of serine integrates as biological tools for diverse applications, from genome engineering to the construction of biocomputers (Brown et al., 2011; Weinberg et al., 2017). Due to their directionality, recombination efficiency and simple DNA sequence requirements, they have been adopted for applications in the fields of molecular genetics, biotechnology and synthetic biology (Fogg et al., 2014). Yoon et al., (2016) characterized EFC-1 as a new serine integrase that can be used for site-specific recombination. Assays were conducted both in vitro and in vivo with HEK293 cells. PCR analysis and confocal microscopy confirmed the efficiency of integration in cells and determined the minimum lengths of the attP and attB sites. Recently, a study conducted by Gomide et al. (2020) investigated the use of six serine integrases utilizing EGFP fluorescence as a measure and molecular analysis of attL/attR site formation after integrase activation in HEK 293T cells. Additionally, as a clinically relevant proof of concept, evaluations of one-way gene switches using these integrases were also performed in peripheral blood mononuclear cells (PBMCs), neural stem cells (NSCs) differentiated from induced pluripotent stem cells, and undifferentiated human embryonic stem cells (hES, BR-1 cell line) (Gomide et al., 2020). The phiC31 integrase was also utilized for integration into endogenous “pseudosites”, regions with similar sequence of an target site that despite lacking perfect match to the correct site can still be rearranged by an integrase in specific cellular contexts. The researchers assessed the occurrence of homologous recombination mediated by the phiC31 integrase between attB and pseudo attP sites in sheep cells (Ni et al., 2012). In this study, a cellular therapy strategy was implemented using genetically corrected induced pluripotent stem cells (iPSCs). The research group explored a mouse model called mdx, which exhibits Duchenne muscular dystrophy, a recessive X-linked disease (Xp21.1 locus). To achieve this, they utilized phiC31 integrase to reprogram the genome at a safe site in mdx fibroblast cells, and the Bxb1 integrase to insert the complete therapeutic dystrophin cDNA into the iPSCs genome (Zhao et al., 2014). In the study conducted by Luo et al. (2015), the aim was to produce transgenic cattle with high expression of recombinant human serum albumin (HSA) in milk. This was achieved by using the phiC31 integrase system and somatic cell nuclear transfer (SCNT). During the study, bovine fetal fibroblast cells (BFFs) were co-transfected with the specific mammary expression plasmid pIACH(-), which contained the attB recognition site for the phiC31 integrase, along with the integrase expression plasmid pCMVInt (Luo et al., 2015). Six serine integrases, namely, Int2, Int4, Int5, Int7, Int9, and Int13, were evaluated by measuring EGFP fluorescence and analyzing the formation of attL/attR sites after integrase functionality in bovine primary fibroblasts. This study aimed to broaden the repertoire of proteins within this class that are capable of activating or deactivating genetic switches in mammalian cells (Gomide et al., 2020).
The detection of variability in integrase activity and its impact on the expression of downstream genes could have significant implications for the design of genetic circuits and for the use of these enzymes in gene therapy or other applications of Synthetic biology. Recently, a study established a characterization framework that includes an experimental protocol, mathematical models, and a computational pipeline for evaluating the main enzymatic parameters of serine integrases, using real-time measurements of fluorescent protein levels in HEK293 cells. The integrases evaluated were: int7, int2, int4, TP901, Bxb1, and phiC31 (Chao et al., 2021). The model identified three independent parameters: expression level, catalytic rate, and substrate affinity, which together explain a substantial amount of the differences observed in enzymatic activities. Among the results, the Bxb1 integrase showed a higher catalytic rate than phiC31, despite phiC31 being expressed at higher levels. The researchers propose that recombinases possess diverse characteristics that influence recombination efficiencies. As an example, int7, whose activity is virtually imperceptible, is expressed at a high level, indicating that its low final recombination efficiency is due to the protein’s functionality, not merely deficient expression. Conversely, phiC31, the recombinase with the highest final recombination efficiency, seemingly catalyzes recombination relatively slowly and relies on extremely high expression for its high efficiency, while int4 had a high catalytic rate but low expression. The team also tested expression vectors using a strong promoter (CMV) or a weak promoter (Ub) to enhance the expression of the integrases in cells. The researchers concluded that the expression level and binding affinity of serine integrases affect the expression of the downstream reporter gene, which could skew the results of the endpoint assay from directly reflecting the actual occurrence of DNA recombination (Chao et al., 2021).
Another study was the first to investigate the potential use of phiC31 to identify and utilize specific integration sites, called pseudo attP sites, in the porcine genome (Bi et al., 2013). The group conducted experiments in the PK15 cell line, transfecting plasmids containing phiC31 attB and attP, and obtained a maximum measured frequency of intramolecular recombination in the cells. They concluded that the selected integrase functioned safely to modify the cellular genome. Furthermore, through the Alamar Blue assay, they demonstrated that the integration of the transgene at these specific sites had little impact on cell proliferation, as no aberrant morphology or abnormal proliferation were observed in the transgenic cell lines. The group established an ideal model to study the effect of the position of an identical transgene in different chromosomal contexts. These findings also serve as a foundation for targeted porcine genomic engineering and can be used to produce genetically modified pigs for agricultural and biomedical purposes (Bi et al., 2013).
The Cre/loxP systems derived from bacteriophage and the FLP/FRT recombinase system derived from yeast have been utilized for introducing genetic modifications in various cell models. In 2002, Andreas S. et al. conducted experiments to compare the efficiency of the phiC31 with the Cre and FLPe recombinases in mammalian cell recombination. Chinese hamster ovary (CHO) cells were transiently transfected with the enzymes (phiC31, Cre, and FLPe) to evaluate their capability in catalyzing DNA recombination. Additionally, reporter cells were generated by stable transfection of NIH 3T3 cells to determine their efficiency in stable integrated targets (Andreas et al., 2002). The phiC31 integrase was modified by adding a nuclear localization signal (NLS) to investigate whether the fusion of NLS could further enhance the efficiency of the recombination process. Two different NLS fusions were created: one with an N-terminal NLS called (NLS-phiC31) and another version with a C-terminal NLS called (phiC31-NLS) to examine if the position of the NLS would influence recombinase activity. The research results indicated that the modified phiC31 integrase, with the addition of a C-terminal NLS, exhibited higher efficiency compared to the Cre and FLPe recombinases in mammalian cell recombination (Andreas et al., 2002).
Rheumatoid Arthritis (RA) is a widely recognized systemic autoimmune disease that is incurable. The etiology of RA remains uncertain, but it is commonly associated with genetic and environmental factors (Ding et al., 2023). Keravala et al. (2006) explored the use of the phiC31 integrase to mediate genetic integration in cultured synovial cells and to enhance genetic expression in rabbit joints. They utilized Hig82 cells, an adherent cell line derived from rabbit synovium, and primary human synovial cells obtained from RA patients for in vitro and in vivo experiments (Keravala et al., 2006). Donor plasmids that expressed the marker gene and contained the attB site, along with the plasmid expressing the phiC31 integrase, were injected, through the patellar tendon, into the hind joints of female rabbits. The results showed that the phiC31 integrase was effective in integrating into synovial cells and led to a significant increase in gene expression in the rabbits’ joints. This suggests that phiC31 integrase could be a promising strategy for gene therapy and the treatment of joint diseases (Keravala et al., 2006).
In this review article, we highlight the integrases Bxb1 and phiC31, regarded as the primary serine recombinases to the date. However, the reduced efficiency and the limited number of known serine integrases substantially restrict their utility as tools for DNA integration in mammalian genome engineering and other eukaryotic cells. Durrant, M.G. et al., (2023), carried out the systematic identification and characterization of a vast number of large serine recombinases (LSRs), derived from mobile genetic elements. In this study, a computational method was developed to identify thousands of new serine integrases and their attachment sites, expanding the known diversity of LSR by over 100 times and enabling the prediction of their site-specificities (Durrant et al., 2023). Tests in human cells, including K-562 lymphoblastoid cells and HEK293 cells, were conducted, classifying them according to genomic targeting. In total, 146,028 genomes from bacterial isolates available in the RefSeq database of the National Center for Biotechnology (NCBI) were analyzed. A series of new LSRs were identified, including Kp03 and Pa01, which outperform the previously characterized Bxb1 by 2–7 times in episomal and chromosomal integration efficiency. The LSR database may also include candidates capable of targeting non-human genomes, encompassing plants, eukaryotic microbes, insects, birds, among other model or non-model study organisms, which could facilitate stable transgenesis in a variety of organisms (Durrant et al., 2023). The group demonstrated that their findings are more extensive compared to previous techniques that relied solely on prophage annotations (Yang et al., 2014). The first study of hybridization with serine integrases to demonstrate chimeric activity of serine integrases addresses the engineering of chimeras to obtain activity in both E. coli cells and mammalian cells, specifically HeLa cells (Farruggio and Calos, 2014). Chimeras were created from three characterized members of the serine integrase family: phiC31, phiBT1, and TG1, by combining their amino and carboxy-terminal portions. The authors constructed several binary hybrids using arrangements that involve some portion of the phiC31 integrase and tested for activity in E. coli and/or HeLa cells with inversion reporter assays. The chimeras constructed were phiC31-phiBT1 (CB), phiBT1-phiC31 (BC), phiC31-TG1 (CT), and TG1-phiC31 (TC). Hybrids of three out of the four tested architectures—BC, CT, and TC chimeras—showed activity in E. coli at hybrid and/or parental att sites. The BC chimeras also demonstrated efficiency in HeLa cells, both in extra-chromosomal assays and at pseudo-sites. This study concluded on the degree of structural compatibility between the catalytic and C-terminal domains of phiC31, phiBT1, and TG1 integrases, and established foundations for using hybridization to create serine integrases with new specificities. Farruggio et al. (2012) were also the first to demonstrate the activity of the serine integrase on attL x attR sites in mammalian cells, including human cell lines (HeLa and HEK293) and mouse cells (NIH3T3). They validated phiC31 RDF as a new tool that would enable future studies to explore phiC31 integrase recombination in both direct and reverse directions (Farruggio et al., 2012). Farruggio et al. (2017) developed an RMCE protocol, called dual integrase cassette exchange (DICE), using phiC31 and Bxb1 integrases in human induced pluripotent stem cells (hiPSCs) to efficiently exchange specific genetic cassettes (H11 safe harbor site in the human genome) (Farruggio et al., 2017). The DICE system aims to allow specific and efficient exchange of genetic cassettes between different genomic locations. In addition, it has the ability to perform several genetic modifications in parallel, using different integrations and recombination sites. This facilitates the analysis of multiple genetic factors in a single experiment, increasing efficiency and reducing experimental variability (Farruggio et al., 2017; Zhu et al., 2014). The group also developed an RMCE protocol using phiC31 and Bxb1 integrases with the DICE method in other cell types, including the H9 female human embryonic stem cell line, gamma-irradiated CF1 mouse embryonic fibroblast feeder cells, and gamma-irradiated DR4 mouse embryonic fibroblast feeder cells (Geisinger and Calos, 2015). Overcoming the challenges of inserting multiple transgenes or genomic fragments of unlimited size into specific locations in mammalian cells, an innovative platform called STRAIGHT-IN (Serine and Tyrosine Recombinase Assisted Integration of Genes for High-Throughput Investigation) was developed to enable targeted genomic integration of large DNA payloads in human-induced pluripotent stem cells (hiPSCs). This methodology combines serine integrases (Bxb1 and phiC31) with CRISPR/Cas9-mediated homologous recombination to achieve precise and site-specific replacement of large genomic regions. The authors demonstrated that there is no restriction on the size of the DNA payload that can be integrated, overcoming one of the main limitations of other commonly used DNA integration systems, such as viral vectors and programmable nuclease knock-ins. Results showed the efficient integration of DNA sequences ranging from 14 to 50 kb in length. Thus, the STRAIGHT-IN platform can significantly simplify the generation of cell lines or animal models containing large and complex genetic circuits, as well as facilitate multiplex genetic assays (Blanch-Asensio et al., 2022). Wei et al. (2011) used the RMCE method to generate a strain of mice called p53 Platform. These mice contain specific attP sequences for the phiC31 integrase at the Trp53 locus. The Trp53 or TP53 gene encodes the p53 protein, which plays a crucial role in cell cycle regulation and cancer prevention. To produce somatic cell lines with endogenously controlled expression of mutant p53, cassette exchange mediated by the phiC31 integrase was performed in embryonic stem (ES) cell lines from mice. This approach allowed precise insertion of DNA sequences at the Trp53 locus, facilitating the investigation of the effects of different mutations in the p53 gene (Wei et al., 2011). Present programmable addition via site-specific targeting elements (PASTE) system combines the precise targeting of CRISPR-Cas9 with the Bxb1 integrase in primary human hepatocytes and T cells. This method was developed to eliminate a defective gene and replace it with a functional gene without inducing double-strand breaks in the DNA. The PASTE system achieved programmable integration of genetic payloads of up to approximately 36 kb in a single delivery reaction (Yarnall et al., 2023). Chow et al. (2021) presented an innovative system called integrase-editable memory by engineered mutagenesis with optical in situ readout (intMEMOIR) (Chow et al., 2021). This system uses the serine integrase Bxb1 to perform irreversible nucleotide edits and fluorescence in situ hybridization (FISH) to read these mutations. This design generates various digital and permanent edit states that can be stably inherited over many cell cycles. The synthetic barcode enabled the in situ reconstruction of lineage relationships in mouse embryonic stem cells (mES) and in fly neural development. The editing deletes or inverts its target region, encoding information in three-state memory elements: “not mutated,” “inverted,” or “deleted.” To create a “trit,” the authors flanked a barcode sequence with an inverted pair of attP sites at one end and an attB site at the other, allowing Bxb1-mediated recombination to produce irreversible deletion or inversion of the barcode (Chow et al., 2021). The ability to visualize cell lineage relationships, distribution, and interactions between cells in native tissue provides deep insight into cell fate determination during development, homeostasis, and disease.
Research in the field of genetic engineering has been dedicated to exploring techniques for insect control. The goal is to develop safe and efficient approaches to reduce disease transmission in animals and humans, protect public health, and combat agricultural pests. In a study involving the model organism Drosophila melanogaster, a genomic integration method using the integrase enzyme phiC31 was employed to direct transgenes to specific locations in the genome. Two transgenic strains of D. melanogaster were generated using the serine integrase phiC31, both in cultured cells (S2) and in embryos (Groth et al., 2004). In studies conducted by Chompoosr et al. (2009), it was found that the integrases phiC31 and R4 have the ability to facilitate site-specific directional recombination in various cell lines, such as Aedes aegypti (Aag2), Anopheles gambiae (Sua5B), Drosophila melanogaster (S2), the silkworm Bombyx mori (BmN4), and the fall armyworm Spodoptera frugiperda (Sf9 cells) (Chompoosri et al., 2009). An alternative approach to enable repeated and independent manipulation of two linked genetic loci in Drosophila was achieved through phiC31 integration. A second pair of attB/attP targeting and transgenesis vectors was introduced, operating in parallel and independently of existing tools. Two synthetic orthologous genes were genetically modified to include orthogonal attB/attP sites. The results demonstrated that the specificity, directionality, and efficiency of recombination of the introduced orthogonal attB CC /attP CC sites can be used in conjunction with the canonical attB TT /attP TT system for independent manipulation and analysis of two transgenesis targets simultaneously in the same organism (Blanco-Redondo and Langenhan, 2018). The InSITE (integrase swappable in vivo targeting element) system using RMCE with phiC31 integrase allowed the replacement of the GAL4/UAS binary expression system with analogous systems (such as hemi-driver split GAL4, GAL80, LexA, or QF) in fruit fly D. melanogaster. With the InSITE system, if a fly line with the desired expression pattern contains the wrong genetic element, the correct element can be swapped in with just a few simple crosses, without the need to inject any plasmids (Gohl et al., 2011).
Model organisms, such as Danio rerio and Xenopus laevis, have also been used in research with the phiC31 integrase. Ints were employed as a mediator for the recombination of transgenes containing attP and attB sites within cis-regulatory sequences, aiming to excise specific elements in zebrafish (Lister, 2010; Lister, 2011). RMCE was also evaluated in the model, using phiC31 integrase to efficiently mediate cassette exchange in both somatic and germline cells of zebrafish (Hu et al., 2011). Additionally, it was used to generate transgenic Xenopus embryos through co-injection of mRNA encoding the phiC31 integrase along with plasmid encoding gfp (Allen and Weeks, 2005). In the chicken DT40 cell line, the phiC31 integrase was employed to facilitate the efficient integration of plasmids and fragments up to 100 kb into vertebrate chromosomes. The results of this research demonstrated that the phiC31 integrase can be successfully used in the generation of transgenic chickens (Dafhnis-Calas et al., 2005). The nuclear import signal and the position within a prokaryotic recombinase can play a crucial role in its efficiency in mammalian cells. Previous studies with the phiC31 integrase in eukaryotic cells have also revealed a variable requirement for a nuclear localization signal (NLS). In chicken DT40 cells, this requirement is absolute, while in D. melanogaster, there appears to be no requirement at all (Groth et al., 2004; Dafhnis-Calas et al., 2005). The group also conducted experiments with chicken DT40 cells to design a minichromosome derived from the human Y chromosome, containing attB and attP sites flanking the centromeric alphoid DNA. They used the phiC31 integrase to promote the deletion or inversion of the centromeric interval. The results showed that the phiC31 integrase promotes efficient, irreversible, and site-specific long-range chromosomal rearrangement in vertebrate cells. Furthermore, they demonstrated that no pseudo-sites were detected in the chicken genome (Malla et al., 2005).
Li et al. (2016) developed a protocol for biolistic transformation for site-specific integration mediated by Bxb1 in tobacco (Nicotiana tabacum) and rice (Oryza sativa), demonstrating efficient gene insertions at these target sites (Li et al., 2016). To implement this system in rice, they generated lines through conventional transformation mediated by Agrobacterium, and various precise target sites in the rice genome were tracked. The GoldenBraid 3.0 (GB3.0) system was developed as a platform for assembling reusable genetic components for Plant Synthetic Biology, incorporating features of its synthetic parts (Sarrion-Perdigones et al., 2013). In this context, a GB3.0 database was established based on the GB2.0 assembly system. For the GB3.0 system, transient Luciferase/Renilla (Luc/Ren) agroinfiltration experiments were carried out in N. benthamiana. The group developed two movements to control transcription. The first one is based on the phiC31 integrase fused to activation (Gal4 or VP64) or repression phiC31 fused to an RD (BRD) domains (Vazquez-Vilar et al., 2017). In the second one, the phiC31 integrase and its counterpart RDF were used to switch between active or inactive states of reporter genes by inverting regulatory parts in Nicotiana benthamiana (Bernabé-Orts et al., 2020). This research marked the first reversible memory swap in entire plants based on the serine phiC31 integrase and its corresponding RDF. De Paepe et al. (2013) demonstrated that T-DNA integration occurred in the progeny of 9% of the T-DNA transformants obtained in Arabidopsis using the method called Iterative Site-Specific Integration (ISSI), which combines the activities of CRE recombinase and phiC31 integrase for efficient T-DNA integration (De Paepe et al., 2013). In another study, the functionality of six integrases: Int2, Int4, Int5, Int7, Int9, and Int13, as well as the phiC31 and Bxb1 integrases in a plasmid cotransfection system was proven to perform the 180° rotation of coding and promoter sequences of the designed genetic switches, thus controlling the expression of the gfp reporter in Arabidopsis thaliana protoplasts (Gomide et al., 2020). The gene expression control systems mentioned above in plants open a wide range of combinatorial possibilities for genetic circuits specifically designed for plants. Such systems allow for the monitoring of agricultural applications, as well as the activation of synthetic genetic networks that respond to biotic factors, such as diseases and pathogens, and abiotic factors. The development of lateral roots in plants serves as an excellent model for the application and study of gene expression, thanks to its well-defined transcriptional control mechanisms. In addition to their positioning, lateral roots exhibit structural characteristics linked to environmental adaptability, making them ideal candidates. To construct synthetic circuits in transgenic Arabidopsis lines, Guiziou et al. (2023) tested the efficiency of serine integrases phiC31, Bxb1 and Tp901. Their results indicated that phiC31 and Bxb1 integrations were constitutively expressed in all plant tissues and were orthogonal to each other (Guiziou et al., 2023). In contrast, the Tp901 integrase did not induce any changes in the targets that carried its recognition sequences, even with strong promoters and codon optimization. phiC31 integration was selected for further experiments in lateral root development. To facilitate this, the researchers developed an integrase toolbox, selecting promoters for several well-trained transcription factors expressed during the initial stages of lateral root initiation: ARF7, ARF19, LBD16 and GATA23. In addition, N. benthamiana was used for transient assays. The team also characterized two methods for adjusting the timing and level of integrase activity: the split-intein integrase and the estradiol-inducible integrase. These methods allow greater control over gene expression, increasing the potential for sophisticated synthetic biology applications in plants. Lastly, Oliveira et al. (2024) integrated a bimodular DNA excision and inversion circuit driven by four distinct serine integrases into the genome of N. benthamiana. The system, denoted Int-Plex@, demonstrated to be functional, allowing the precise excision and inversion of genomic DNA sequences. Through the application of BxB1, phiC31, Int9 and Int13, it was possible to activate specific modules to control the removal or inversion of DNA segments, showing the potential of the system as a tool for engineering genetic circuits and modulating metabolic pathways in plants in a controlled and reversible manner (Oliveira et al., 2024).
Plasmodium falciparum is the causative agent of malaria. The protozoan invades human erythrocytes, inducing a cascade of changes in these cells for its development and proliferation (Zheng et al., 2024). In this scenario, science has developed various tools for genetic manipulation of this organism. Nkrumah et al. (2006) developed P. falciparum strains with the attB site integrated into the cg6 gene, which encodes an important protein in the parasite’s life cycle, using the Bxb1 integrase (Nkrumah et al., 2006). The use of this integrase allowed for more efficient and precise integration compared to traditional genetic transformation methods, enabling the generation of transgenic parasites with controlled genetic insertions. Building on this work, Spalding et al. (2010) sought to validate the method by modifying Bxb1 integrase-mediated recombination in P. falciparum, using the H protein from the glycine cleavage complex (GCV) located in the mitochondria as an indicator (Spalding et al., 2010). They generated integrated cell lines expressing the H-GFP protein (HDd2 protein) and demonstrated the localization of this protein in the parasite’s mitochondria, thereby validating the efficacy of the recombination method. Balabaskaran-Nina and Desai (2018) described a new application strategy using the Bxb1 integrase to allow targeted gene substitutions through an intronic attB sequence within the gene of interest (Balabaskaran-Nina and Desai, 2018). The authors argue that this approach enables rapid specific site integration, encompassing the full spectrum of native genetic modifications and offering distinct advantages compared to other methods, including CRISPR-Cas9.
There are several techniques used in gene editing research, such as the TALEN (Transcription activator-like effector nuclease), ZFN (Zinc Finger Nucleases), and the CRISPR/Cas9 systems. These methods use different proteins to direct restriction enzymes to cut DNA at specific locations. Each system has advantages and disadvantages, and the selection depends on the purpose of the genetic modification. However, it is still a difficult task to identify nucleases with high affinity and specificity. New methodologies are currently being used in Synthetic Biology to create new molecular approaches that lead to characterization, generating new components, cells, biomolecules, expression, and modification of genetic material (DNA/RNA). Genetic engineering offers promising applications in the areas of health, agriculture, and the environment, which requires the development of tools to improve the modulation of multiple genes. The use of serine integrases has been increasingly explored in Synthetic Biology studies because these enzymes can mediate specific recombination of DNA sequences, enabling the integration or excision of genes of interest at specific locations in the genome. A major advantage of using these recombinases is their high efficiency in inserting or removing DNA sequences, avoiding unwanted changes in the genome. This makes these enzymes a promising tool for various biotechnological applications, such as the production of transgenic organisms and gene therapies. It is essential to highlight that serine integrases of phage origin are valuable tools for promoting selective integration of genomic sequences into eukaryotic cells, combined with donor plasmids containing the attB recognition site with introduced genomic attP sites or pseudo-attP sites. Recent studies have shown that serine integrases can be successfully used in plants, human, and animal cells, paving the way for therapeutic and biotechnological applications. The applications may include various areas such as treatment of metabolic disorders, immune-mediated diseases, anticancer therapy, human or animal reproductive area, animal breeding and cloning in agriculture, among others. In addition, the requirement for recognition of two sites (attB/attP) for its action can be a minimizing factor for off-target gene editing. Despite the promising results observed in the use of these enzymes in eukaryotic cells, many questions remain to be investigated.
TS: Writing–original draft, Writing–review and editing. MO: Conceptualization, Writing–original draft, Writing–review and editing. LF: Conceptualization, Writing–review and editing. RL: Conceptualization, Writing–original draft. ER: Funding acquisition, Supervision, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by Embrapa Genetic Resources and Biotechnology/National Institute of Science and Technology in Synthetic Biology, National Council for Scientific and Technological Development/ Ministry of Agriculture Livestock and Supply (465603/2014-9; 400145/2023-5), Research Support Foundation of the Federal District (0193.001.262/2017), and Coordination for the Improvement of Higher Education Personnel.
This is a brief acknowledgment of the technological support provided by the Canva Pro IA software, which was used for creating planned and precise scientific visual representations.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abe, K., Kawano, Y., Iwamoto, K., Arai, K., Maruyama, Y., Eichenberger, P., et al. (2014). Developmentally-regulated excision of the SPβ prophage reconstitutes a gene required for spore envelope maturation in Bacillus subtilis. PLoS Genet. 10 (10), e1004636. doi:10.1371/journal.pgen.1004636
Abioye, J., Lawson-Williams, M., Lecanda, A., Calhoon, B., McQue, A. L., Colloms, S. D., et al. (2023). High fidelity one-pot DNA assembly using orthogonal serine integrases. Biotechnol. J. 18 (3), e2200411. doi:10.1002/biot.202200411
Adjalley, S. H., Lee, M. C., and Fidock, D. A. (2010). A method for rapid genetic integration into Plasmodium falciparum utilizing mycobacteriophage Bxb1 integrase. Methods Mol. Biol. 634, 87–100. doi:10.1007/978-1-60761-652-8_6
Ahmadi, M., Mahboudi, F., Akbari Eidgahi, M. R., Nasr, R., Nematpour, F., Ahmadi, S., et al. (2016). Evaluating the efficiency of phiC31 integrase-mediated monoclonal antibody expression in CHO cells. Biotechnol. Prog. 32 (6), 1570–1576. doi:10.1002/btpr.2362
Allen, B. G., and Weeks, D. L. (2005). Transgenic Xenopus laevis embryos can be generated using φC31 integrase. Nat. Methods 2 (12), 975–979. doi:10.1038/nmeth814
Andreas, S., Schwenk, F., Küter-Luks, B., Faust, N., and Kühn, R. (2002). Enhanced efficiency through nuclear localization signal fusion on phage PhiC31-integrase: activity comparison with Cre and FLPe recombinase in mammalian cells. Nucleic Acids Res. 30 (11), 2299–2306. doi:10.1093/nar/30.11.2299
Baaziz, H., Makhlouf, R., McClelland, M., and Hsu, B. B. (2024). Bacterial resistance to temperate phage is influenced by the frequency of lysogenic establishment. iScience 27 (4), 109595. doi:10.1016/j.isci.2024.109595
Bailey, Z. M., Igler, C., and Wendling, C. C. (2024). Prophage maintenance is determined by environment-dependent selective sweeps rather than mutational availability. Curr. Biol. 34, 1739–1749.e7. doi:10.1016/j.cub.2024.03.025
Balabaskaran-Nina, P., and Desai, S. A. (2018). Diverse target gene modifications in Plasmodium falciparum using Bxb1 integrase and an intronic attB. Parasites and Vectors 11 (1), 548. doi:10.1186/s13071-018-3129-5
Bernabé-Orts, J. M., Quijano-Rubio, A., Vazquez-Vilar, M., Mancheño-Bonillo, J., Moles-Casas, V., Selma, S., et al. (2020). A memory switch for plant synthetic biology based on the phage ϕC31 integration system. Nucleic Acids Res. 48 (6), 3379–3394. doi:10.1093/nar/gkaa104
Bernheim, A., and Sorek, R. (2018). Viruses cooperate to defeat bacteria. Nature 559 (7715), 482–484. doi:10.1038/d41586-018-05762-1
Bi, Y., Liu, X., Zhang, L., Shao, C., Ma, Z., Hua, Z., et al. (2013). Pseudo attP sites in favor of transgene integration and expression in cultured porcine cells identified by Streptomyces phage phiC31 integrase. BMC Mol. Biol. 14, 20. doi:10.1186/1471-2199-14-20
Bibb, L. A., and Hatfull, G. F. (2002). Integration and excision of the Mycobacterium tuberculosis prophage-like element, φRv1. Mol. Microbiol. 45 (6), 1515–1526. doi:10.1046/j.1365-2958.2002.03130.x
Bisen, M., Kharga, K., Mehta, S., Jabi, N., and Kumar, L. (2024). Bacteriophages in nature: recent advances in research tools and diverse environmental and biotechnological applications. Environ. Sci. Pollut. Res. Int. 31 (15), 22199–22242. doi:10.1007/s11356-024-32535-3
Blanch-Asensio, A., Grandela, C., Brandão, K. O., de Korte, T., Mei, H., Ariyurek, Y., et al. (2022). STRAIGHT-IN enables high-throughput targeting of large DNA payloads in human pluripotent stem cells. Cell Rep. Methods 2 (10), 100300. doi:10.1016/j.crmeth.2022.100300
Blanco-Redondo, B., and Langenhan, T. (2018). Parallel genomic engineering of two Drosophila genes using orthogonal attB/attP sites. G3 (Bethesda) 8 (9), 3109–3118. doi:10.1534/g3.118.200565
Bonnet, J., Subsoontorn, P., and Endy, D. (2012). Rewritable digital data storage in live cells via engineered control of recombination directionality. Proc. Natl. Acad. Sci. U. S. A. 109 (23), 8884–8889. doi:10.1073/pnas.1202344109
Bonnet, J., Yin, P., Ortiz, M. E., Subsoontorn, P., and Endy, D. (2013). Amplifying genetic logic gates. Science 340 (6132), 599–603. doi:10.1126/science.1232758
Breüner, A., Brøndsted, L., and Hammer, K. (1999). Novel organization of genes involved in prophage excision identified in the temperate lactococcal bacteriophage TP901-1. J. Bacteriol. 181 (23), 7291–7297. doi:10.1128/jb.181.23.7291-7297.1999
Brown, W. R., Lee, N. C., Xu, Z., and Smith, M. C. (2011). Serine recombinases as tools for genome engineering. Methods 53 (4), 372–379. doi:10.1016/j.ymeth.2010.12.031
Butt, L., Meyer, J. R., Lindsay, R. J., Beardmore, R. E., and Gudelj, I. (2024). Bacterial resistance response and resource availability mediate viral coexistence. J. Evol. Biol. 37 (4), 371–382. doi:10.1093/jeb/voae022
Chao, G., Travis, C., and Church, G. (2021). Measurement of large serine integrase enzymatic characteristics in HEK293 cells reveals variability and influence on downstream reporter expression. Febs J. 288 (22), 6410–6427. doi:10.1111/febs.16037
Chibani-Chennoufi, S., Bruttin, A., Dillmann, M. L., and Brüssow, H. (2004). Phage-host interaction: an ecological perspective. J. Bacteriol. 186 (12), 3677–3686. doi:10.1128/jb.186.12.3677-3686.2004
Chompoosri, J., Fraser, T., Rongsriyam, Y., Komalamisra, N., Siriyasatien, P., Thavara, U., et al. (2009). Intramolecular integration assay validates integrase phi C31 and R4 potential in a variety of insect cells. Southeast Asian J. Trop. Med. Public Health 40 (6), 1235–1253.
Chow, K. K., Budde, M. W., Granados, A. A., Cabrera, M., Yoon, S., Cho, S., et al. (2021). Imaging cell lineage with a synthetic digital recording system. Science 372 (6538), eabb3099. doi:10.1126/science.abb3099
Dafhnis-Calas, F., Xu, Z., Haines, S., Malla, S. K., Smith, M. C., and Brown, W. R. (2005). Iterative in vivo assembly of large and complex transgenes by combining the activities of phiC31 integrase and Cre recombinase. Nucleic Acids Res. 33 (22), e189. doi:10.1093/nar/gni192
De Paepe, A., De Buck, S., Nolf, J., Van Lerberge, E., and Depicker, A. (2013). Site-specific T–DNA integration in A mediated by the combined action of CRE recombinase and ϕC31 integrase. Plant J. 75 (1), 172–184. doi:10.1111/tpj.12202
Ding, Q., Hu, W., Wang, R., Yang, Q., Zhu, M., Li, M., et al. (2023). Signaling pathways in rheumatoid arthritis: implications for targeted therapy. Signal Transduct. Target Ther. 8 (1), 68. doi:10.1038/s41392-023-01331-9
Doore, S. M., and Fane, B. A. (2016). The microviridae: diversity, assembly, and experimental evolution. Virology 491, 45–55. doi:10.1016/j.virol.2016.01.020
Durrant, M. G., Fanton, A., Tycko, J., Hinks, M., Chandrasekaran, S. S., Perry, N. T., et al. (2023). Systematic discovery of recombinases for efficient integration of large DNA sequences into the human genome. Nat. Biotechnol. 41 (4), 488–499. doi:10.1038/s41587-022-01494-w
Fan, H. F., Hsieh, T. S., Ma, C. H., and Jayaram, M. (2016). Single-molecule analysis of ϕC31 integrase-mediated site-specific recombination by tethered particle motion. Nucleic Acids Res. 44 (22), 10804–10823. doi:10.1093/nar/gkw861
Farruggio, A. P., Bhakta, M. S., and Calos, M. P. (2017). Use of the DICE (dual integrase cassette exchange) system. Methods Mol. Biol. 1642, 69–85. doi:10.1007/978-1-4939-7169-5_5
Farruggio, A. P., and Calos, M. P. (2014). Serine integrase chimeras with activity in E. coli and HeLa cells. Biol. Open 3 (10), 895–903. doi:10.1242/bio.20148748
Farruggio, A. P., Chavez, C. L., Mikell, C. L., and Calos, M. P. (2012). Efficient reversal of phiC31 integrase recombination in mammalian cells. Biotechnol. J. 7 (11), 1332–1336. doi:10.1002/biot.201200283
Fogg, P. C., Colloms, S., Rosser, S., Stark, M., and Smith, M. C. (2014). New applications for phage integrases. J. Mol. Biol. 426 (15), 2703–2716. doi:10.1016/j.jmb.2014.05.014
Fogg, P. C. M., Haley, J. A., Stark, W. M., and Smith, M. C. M. (2017). Genome integration and excision by a new Streptomyces bacteriophage, ϕJoe. Appl. Environ. Microbiol. 83 (5), e02767. doi:10.1128/aem.02767-16
Fogg, P. C. M., Younger, E., Fernando, B. D., Khaleel, T., Stark, W. M., and Smith, M. C. M. (2018). Recombination directionality factor gp3 binds ϕC31 integrase via the zinc domain, potentially affecting the trajectory of the coiled-coil motif. Nucleic Acids Res. 46 (3), 1308–1320. doi:10.1093/nar/gkx1233
Gaj, T., Sirk, S. J., and Barbas, C. F. (2014). Expanding the scope of site-specific recombinases for genetic and metabolic engineering. Biotechnol. Bioeng. 111 (1), 1–15. doi:10.1002/bit.25096
Geisinger, J. M., and Calos, M. P. (2015). Using phage integrases in a site-specific dual integrase cassette exchange strategy. Methods Mol. Biol. 1239, 29–38. doi:10.1007/978-1-4939-1862-1_3
Ghosh, P., Wasil, L. R., and Hatfull, G. F. (2006). Control of phage Bxb1 excision by a novel recombination directionality factor. PLoS Biol. 4 (6), e186. doi:10.1371/journal.pbio.0040186
Gohl, D. M., Silies, M. A., Gao, X. J., Bhalerao, S., Luongo, F. J., Lin, C. C., et al. (2011). A versatile in vivo system for directed dissection of gene expression patterns. Nat. Methods 8 (3), 231–237. doi:10.1038/nmeth.1561
Gomide, M. S., Sales, T. T., Barros, L. R. C., Limia, C. G., de Oliveira, M. A., Florentino, L. H., et al. (2020). Genetic switches designed for eukaryotic cells and controlled by serine integrases. Commun. Biol. 3 (1), 255. doi:10.1038/s42003-020-0971-8
Grigson, S. R., Giles, S. K., Edwards, R. A., and Papudeshi, B. (2023). Knowing and naming: phage annotation and nomenclature for phage therapy. Clin. Infect. Dis. 77 (Suppl. 5), S352–s359. doi:10.1093/cid/ciad539
Grindley, N. D., Whiteson, K. L., and Rice, P. A. (2006). Mechanisms of site-specific recombination. Annu. Rev. Biochem. 75, 567–605. doi:10.1146/annurev.biochem.73.011303.073908
Groth, A. C., Fish, M., Nusse, R., and Calos, M. P. (2004). Construction of transgenic Drosophila by using the site-specific integrase from phage φC31. Genetics 166 (4), 1775–1782. doi:10.1093/genetics/166.4.1775
Groth, A. C., Olivares, E. C., Thyagarajan, B., and Calos, M. P. (2000). A phage integrase directs efficient site-specific integration in human cells. Proc. Natl. Acad. Sci. U. S. A. 97 (11), 5995–6000. doi:10.1073/pnas.090527097
Guiziou, S., Maranas, C. J., Chu, J. C., and Nemhauser, J. L. (2023). An integrase toolbox to record gene-expression during plant development. Nat. Commun. 14 (1), 1844. doi:10.1038/s41467-023-37607-5
Gupta, K., Sharp, R., Yuan, J. B., Li, H., and Van Duyne, G. D. (2017). Coiled-coil interactions mediate serine integrase directionality. Nucleic Acids Res. 45 (12), 7339–7353. doi:10.1093/nar/gkx474
Hatfull, G. F., and Hendrix, R. W. (2011). Bacteriophages and their genomes. Curr. Opin. Virol. 1 (4), 298–303. doi:10.1016/j.coviro.2011.06.009
Hew, B. E., Gupta, S., Sato, R., Waller, D. F., Stoytchev, I., Short, J. E., et al. (2024). Directed evolution of hyperactive integrases for site specific insertion of transgenes. Nucleic Acids Res. 52 (14), e64. doi:10.1093/nar/gkae534
Hong, J., Sohn, K. C., Park, H. W., Jeon, H., Ju, E., Lee, J. G., et al. (2024). All-in-one IQ toggle switches with high versatilities for fine-tuning of transgene expression in mammalian cells and tissues. Mol. Ther. Methods Clin. Dev. 32 (1), 101202. PMID: 38374964; PMCID: PMC10875299. doi:10.1016/j.omtm.2024.101202
Hu, G., Goll, M. G., and Fisher, S. (2011). ΦC31 integrase mediates efficient cassette exchange in the zebrafish germline. Dev. Dyn. 240 (9), 2101–2107. doi:10.1002/dvdy.22699
Huhtinen, O., Prince, S., Lamminmäki, U., Salbo, R., and Kulmala, A. (2024). Increased stable integration efficiency in CHO cells through enhanced nuclear localization of Bxb1 serine integrase. BMC Biotechnol. 24, 44. doi:10.1186/s12896-024-00871-4
Kasman, L. M., and Porter, L. D. (2022). “Bacteriophages,” in StatPearls [Internet]. Treasure Island, FL: StatPearls Publishing.
Keravala, A., Groth, A. C., Jarrahian, S., Thyagarajan, B., Hoyt, J. J., Kirby, P. J., et al. (2006). A diversity of serine phage integrases mediate site-specific recombination in mammalian cells. Mol. Genet. Genomics 276 (2), 135–146. doi:10.1007/s00438-006-0129-5
Li, H., Sharp, R., Rutherford, K., Gupta, K., and Van Duyne, G. D. (2018). Serine integrase attP binding and specificity. J. Mol. Biol. 430 (21), 4401–4418. doi:10.1016/j.jmb.2018.09.007
Li, R., Han, Z., Hou, L., Kaur, G., Yin, Q., and Ow, D. W. (2016). Method for biolistic site-specific integration in plants catalyzed by Bxb1 integrase. Methods Mol. Biol. 1469, 15–30. doi:10.1007/978-1-4939-4931-1_2
Lister, J. A. (2010). Transgene excision in zebrafish using the phiC31 integrase. Genesis 48 (2), 137–143. doi:10.1002/dvg.20595
Lister, J. A. (2011). Use of phage φC31 integrase as a tool for zebrafish genome manipulation. Methods Cell Biol. 104, 195–208. doi:10.1016/b978-0-12-374814-0.00011-2
Luo, Y., Wang, Y., Liu, J., Lan, H., Shao, M., Yu, Y., et al. (2015). Production of transgenic cattle highly expressing human serum albumin in milk by phiC31 integrase-mediated gene delivery. Transgenic Res. 24 (5), 875–883. doi:10.1007/s11248-015-9898-0
MacDonald, A. I., Baksh, A., Holland, A., Shin, H., Rice, P. A., Stark, W. M., et al. (2024). Variable orthogonality of RDF - large serine integrase interactions within the ϕC31 family. nature, 587898. doi:10.1101/2024.04.03.587898
Malla, S., Dafhnis-Calas, F., Brookfield, J. F., Smith, M. C., and Brown, W. R. (2005). Rearranging the centromere of the human Y chromosome with phiC31 integrase. Nucleic Acids Res. 33 (19), 6101–6113. doi:10.1093/nar/gki922
Mandali, S., Gupta, K., Dawson, A. R., Van Duyne, G. D., and Johnson, R. C. (2017). Control of recombination directionality by the Listeria phage A118 protein Gp44 and the coiled-coil motif of its serine integrase. J. Bacteriol. 199 (11), e00019. doi:10.1128/jb.00019-17
Mandali, S., and Johnson, R. C. (2021). Control of the serine integrase reaction: roles of the coiled-coil and helix E regions in DNA site synapsis and recombination. J. Bacteriol. 203 (16), e0070320. doi:10.1128/jb.00703-20
Maranas, C. J., George, W., Scallon, S. K., VanGilder, S., Nemhauser, J. L., and Guiziou, S. (2024). A history-dependent integrase recorder of plant gene expression with single-cell resolution. Nat. Commun. 15, 9362. doi:10.1038/s41467-024-53716-1
McEwan, A. R., Raab, A., Kelly, S. M., Feldmann, J., and Smith, M. C. (2011). Zinc is essential for high-affinity DNA binding and recombinase activity of ΦC31 integrase. Nucleic Acids Res. 39 (14), 6137–6147. doi:10.1093/nar/gkr220
McEwan, A. R., Rowley, P. A., and Smith, M. C. (2009). DNA binding and synapsis by the large C-terminal domain of C31 integrase. Nucleic Acids Res. 37 (14), 4764–4773. doi:10.1093/nar/gkp485
Merrick, C. A., Zhao, J., and Rosser, S. J. (2018). Serine integrases: advancing synthetic biology. ACS Synth. Biol. 7 (2), 299–310. doi:10.1021/acssynbio.7b00308
Mills, S., Shanahan, F., Stanton, C., Hill, C., Coffey, A., and Ross, R. P. (2013). Movers and shakers: influence of bacteriophages in shaping the mammalian gut microbiota. Gut Microbes 4 (1), 4–16. doi:10.4161/gmic.22371
Ni, W., Hu, S., Qiao, J., Wang, Y., Shi, H., Wang, Y., et al. (2012). ΦC31 integrase mediates efficient site-specific integration in sheep fibroblasts. Biosci. Biotechnol. Biochem. 76 (11), 2093–2095. doi:10.1271/bbb.120439
Nielsen, A. A., Der, B. S., Shin, J., Vaidyanathan, P., Paralanov, V., Strychalski, E. A., et al. (2016). Genetic circuit design automation. Science 352 (6281), aac7341. doi:10.1126/science.aac7341
Nkrumah, L. J., Muhle, R. A., Moura, P. A., Ghosh, P., Hatfull, G. F., Jacobs, W. R., et al. (2006). Efficient site-specific integration in Plasmodium falciparum chromosomes mediated by mycobacteriophage Bxb1 integrase. Nat. Methods 3 (8), 615–621. doi:10.1038/nmeth904
Oliveira, M. A. d., Lima, R. N., Florentino, L. H., Almeida, M. M. S. d., Melo, F. L., Bonnet, R. V., et al. (2024). Development of Int-Plex@ binary memory switch system: plant genome modulation driven by large serine-integrases. J. bioRxiv 2024, 575089. doi:10.1101/2024.01.11.575089
Olorunniji, F. J., Lawson-Williams, M., McPherson, A. L., Paget, J. E., Stark, W. M., and Rosser, S. J. (2019). Control of ϕC31 integrase-mediated site-specific recombination by protein trans-splicing. Nucleic Acids Res. 47 (21), 11452–11460. doi:10.1093/nar/gkz936
Olorunniji, F. J., Merrick, C., Rosser, S. J., Smith, M. C. M., Stark, W. M., and Colloms, S. D. (2017). Multipart DNA assembly using site-specific recombinases from the large serine integrase family. Methods Mol. Biol. 1642, 303–323. doi:10.1007/978-1-4939-7169-5_19
Olorunniji, F. J., Rosser, S. J., and Stark, W. M. (2016). Site-specific recombinases: molecular machines for the Genetic Revolution. Biochem. J. 473 (6), 673–684. doi:10.1042/bj20151112
Patel, P. H., Taylor, V. L., Zhang, C., Getz, L. J., Fitzpatrick, A. D., Davidson, A. R., et al. (2024). Anti-phage defence through inhibition of virion assembly. Nat. Commun. 15 (1), 1644. doi:10.1038/s41467-024-45892-x
Piel, D., Bruto, M., Labreuche, Y., Blanquart, F., Goudenège, D., Barcia-Cruz, R., et al. (2022). Phage-host coevolution in natural populations. Nat. Microbiol. 7 (7), 1075–1086. doi:10.1038/s41564-022-01157-1
Rostøl, J. T., Quiles-Puchalt, N., Iturbe-Sanz, P., Lasa, Í., and Penadés, J. R. (2024). Bacteriophages avoid autoimmunity from cognate immune systems as an intrinsic part of their life cycles. Nat. Microbiol. 9, 1312–1324. doi:10.1038/s41564-024-01661-6
Rutherford, K., and Van Duyne, G. D. (2014). The ins and outs of serine integrase site-specific recombination. Curr. Opin. Struct. Biol. 24, 125–131. doi:10.1016/j.sbi.2014.01.003
Rutherford, K., Yuan, P., Perry, K., Sharp, R., and Van Duyne, G. D. (2013). Attachment site recognition and regulation of directionality by the serine integrases. Nucleic Acids Res. 41 (17), 8341–8356. doi:10.1093/nar/gkt580
Sabino, J., Hirten, R. P., and Colombel, J. F. (2020). Review article: bacteriophages in gastroenterology-from biology to clinical applications. Aliment. Pharmacol. Ther. 51 (1), 53–63. doi:10.1111/apt.15557
Sarrion-Perdigones, A., Vazquez-Vilar, M., Palací, J., Castelijns, B., Forment, J., Ziarsolo, P., et al. (2013). GoldenBraid 2.0: a comprehensive DNA assembly framework for plant synthetic biology. Plant Physiol. 162 (3), 1618–1631. doi:10.1104/pp.113.217661
Siedentop, B., Rüegg, D., Bonhoeffer, S., and Chabas, H. (2024). My host's enemy is my enemy: plasmids carrying CRISPR-Cas as a defence against phages. Proc. Biol. Sci. 291 (2015), 20232449. doi:10.1098/rspb.2023.2449
Simmonds, P., and Aiewsakun, P. (2018). Virus classification - where do you draw the line? Arch. Virol. 163 (8), 2037–2046. doi:10.1007/s00705-018-3938-z
Smith, M. C. M. (2015). Phage-encoded serine integrases and other large serine recombinases. Microbiol. Spectr. 3 (4). doi:10.1128/microbiolspec.MDNA3-0059-2014
Snoeck, N., De Mol, M. L., Van Herpe, D., Goormans, A., Maryns, I., Coussement, P., et al. (2019). Serine integrase recombinational engineering (SIRE): a versatile toolbox for genome editing. Biotechnol. Bioeng. 116 (2), 364–374. doi:10.1002/bit.26854
Spalding, M. D., Allary, M., Gallagher, J. R., and Prigge, S. T. (2010). Validation of a modified method for Bxb1 mycobacteriophage integrase-mediated recombination in Plasmodium falciparum by localization of the H-protein of the glycine cleavage complex to the mitochondrion. Mol. Biochem. Parasitol. 172 (2), 156–160. doi:10.1016/j.molbiopara.2010.04.005
Stark, W. M. (2017). Making serine integrases work for us. Curr. Opin. Microbiol. 38, 130–136. doi:10.1016/j.mib.2017.04.006
Suttle, C. A. (2007). Marine viruses-major players in the global ecosystem. Nat. Rev. Microbiol. 5 (10), 801–812. doi:10.1038/nrmicro1750
Thomson, J. G., and Ow, D. W. (2006). Site-specific recombination systems for the genetic manipulation of eukaryotic genomes. Genesis 44 (10), 465–476. doi:10.1002/dvg.20237
Turner, D., Adriaenssens, E. M., Lehman, S. M., Moraru, C., and Kropinski, A. M. (2024). Bacteriophage taxonomy: a continually evolving discipline. Methods Mol. Biol. 2734, 27–45. doi:10.1007/978-1-0716-3523-0_3
Van Duyne, G. D., and Rutherford, K. (2013). Large serine recombinase domain structure and attachment site binding. Crit. Rev. Biochem. Mol. Biol. 48 (5), 476–491. doi:10.3109/10409238.2013.831807
Vazquez-Vilar, M., Quijano-Rubio, A., Fernandez-Del-Carmen, A., Sarrion-Perdigones, A., Ochoa-Fernandez, R., Ziarsolo, P., et al. (2017). GB3.0: a platform for plant bio-design that connects functional DNA elements with associated biological data. Nucleic Acids Res. 45 (4), 2196–2209. doi:10.1093/nar/gkw1326
Wang, J., Chitsaz, F., Derbyshire, M. K., Gonzales, N. R., Gwadz, M., Lu, S., et al. (2023). The conserved domain database in 2023. Nucleic Acids Res. 51 (D1), D384–d388. doi:10.1093/nar/gkac1096
Wang, S., Yang, Y., and Jing, J. (2022). A synthesis of viral contribution to marine nitrogen cycling. Front. Microbiol. 13, 834581. doi:10.3389/fmicb.2022.834581
Wang, Y., Yau, Y. Y., Perkins-Balding, D., and Thomson, J. G. (2011). Recombinase technology: applications and possibilities. Plant Cell Rep. 30 (3), 267–285. doi:10.1007/s00299-010-0938-1
Warwick-Dugdale, J., Buchholz, H. H., Allen, M. J., and Temperton, B. (2019). Host-hijacking and planktonic piracy: how phages command the microbial high seas. Virol. J. 16 (1), 15. doi:10.1186/s12985-019-1120-1
Wei, Q. X., Odell, A. F., van der Hoeven, F., and Hollstein, M. (2011). Rapid derivation of genetically related mutants from embryonic cells harboring a recombinase-specific Trp53 platform. Cell Cycle 10 (8), 1261–1270. doi:10.4161/cc.10.8.15303
Weinbauer, M. G. (2004). Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 28 (2), 127–181. doi:10.1016/j.femsre.2003.08.001
Weinberg, B. H., Pham, N. T. H., Caraballo, L. D., Lozanoski, T., Engel, A., Bhatia, S., et al. (2017). Large-scale design of robust genetic circuits with multiple inputs and outputs for mammalian cells. Nat. Biotechnol. 35 (5), 453–462. doi:10.1038/nbt.3805
Xu, Z., and Brown, W. R. (2016). Comparison and optimization of ten phage encoded serine integrases for genome engineering in Saccharomyces cerevisiae. BMC Biotechnol. 16, 13. doi:10.1186/s12896-016-0241-5
Xu, Z., Thomas, L., Davies, B., Chalmers, R., Smith, M., and Brown, W. (2013). Accuracy and efficiency define Bxb1 integrase as the best of fifteen candidate serine recombinases for the integration of DNA into the human genome. BMC Biotechnol. 13, 87. doi:10.1186/1472-6750-13-87
Yang, L., Nielsen, A. A., Fernandez-Rodriguez, J., McClune, C. J., Laub, M. T., Lu, T. K., et al. (2014). Permanent genetic memory with >1-byte capacity. Nat. Methods 11 (12), 1261–1266. doi:10.1038/nmeth.3147
Yarnall, M. T. N., Ioannidi, E. I., Schmitt-Ulms, C., Krajeski, R. N., Lim, J., Villiger, L., et al. (2023). Drag-and-drop genome insertion of large sequences without double-strand DNA cleavage using CRISPR-directed integrases. Nat. Biotechnol. 41 (4), 500–512. doi:10.1038/s41587-022-01527-4
Yoon, B., Kim, I., Nam, J. A., Chang, H. I., and Ha, C. H. (2016). In vivo and in vitro characterization of site-specific recombination of a novel serine integrase from the temperate phage EFC-1. Biochem. Biophys. Res. Commun. 473 (1), 336–341. doi:10.1016/j.bbrc.2016.03.106
Young, R. (2013). Phage lysis: do we have the hole story yet? Curr. Opin. Microbiol. 16 (6), 790–797. doi:10.1016/j.mib.2013.08.008
Zhang, L., Zhu, B., Dai, R., Zhao, G., and Ding, X. (2013). Control of directionality in Streptomyces phage φBT1 integrase-mediated site-specific recombination. PLoS One 8 (11), e80434. doi:10.1371/journal.pone.0080434
Zhao, C., Farruggio, A. P., Bjornson, C. R., Chavez, C. L., Geisinger, J. M., Neal, T. L., et al. (2014). Recombinase-mediated reprogramming and dystrophin gene addition in mdx mouse induced pluripotent stem cells. PLoS One 9 (4), e96279. doi:10.1371/journal.pone.0096279
Zheng, K., Li, Q., Jiang, N., Zhang, Y., Zheng, Y., Zhang, Y., et al. (2024). Plasmodium falciparum selectively degrades α-spectrin of infected erythrocytes after invasion. mBio 15 (4), e0351023. doi:10.1128/mbio.03510-23
Zhu, F., Gamboa, M., Farruggio, A. P., Hippenmeyer, S., Tasic, B., Schüle, B., et al. (2014). DICE, an efficient system for iterative genomic editing in human pluripotent stem cells. Nucleic Acids Res. 42 (5), e34. doi:10.1093/nar/gkt1290
Keywords: serine integrase, eukaryotic cell, phage (bacteriophage), synthetic biology, genome edition tools
Citation: Sales TT, de Oliveira MA, Florentino LH, Lima RN and Rech E (2025) There and turn back again: the application of phage serine integrases in eukaryotic systems. Front. Bioeng. Biotechnol. 13:1478413. doi: 10.3389/fbioe.2025.1478413
Received: 09 August 2024; Accepted: 03 February 2025;
Published: 24 February 2025.
Edited by:
Jose Ruben Morones-Ramirez, Autonomous University of Nuevo León, MexicoReviewed by:
Shaobo Yang, Dana–Farber Cancer Institute, United StatesCopyright © 2025 Sales, de Oliveira, Florentino, Lima and Rech. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elibio Rech, ZWxpYmlvLnJlY2hAZW1icmFwYS5icg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.