
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Bioeng. Biotechnol. , 22 September 2020
Sec. Bioprocess Engineering
Volume 8 - 2020 | https://doi.org/10.3389/fbioe.2020.557572
This article is part of the Research Topic Novel Technologies for Microalgae Utilization to Achieve Global Sustainable Development Goals (SDGs) View all 12 articles
 Roland Wirth1,2
Roland Wirth1,2 Bernadett Pap1
Bernadett Pap1 Tamás Böjti2
Tamás Böjti2 Prateek Shetty1
Prateek Shetty1 Gergely Lakatos1Zoltán Bagi2
Gergely Lakatos1Zoltán Bagi2 Kornél L. Kovács2,3
Kornél L. Kovács2,3 Gergely Maróti1,4*
Gergely Maróti1,4*Microalgae-based bioenergy production is a promising field with regard to the wide variety of algal species and metabolic potential. The use of liquid wastes as nutrient clearly improves the sustainability of microalgal biofuel production. Microalgae and bacteria have an ecological inter-kingdom relationship. This microenvironment called phycosphere has a major role in the ecosystem productivity and can be utilized both in bioremediation and biomass production. However, knowledge on the effects of indigenous bacteria on microalgal growth and the characteristics of bacterial communities associated with microalgae are limited. In this study municipal, industrial and agricultural liquid waste derivatives were used as cultivation media. Chlorella vulgaris green microalgae and its bacterial partners efficiently metabolized the carbon, nitrogen and phosphorous content available in these wastes. The read-based metagenomics approach revealed a diverse microbial composition at the start point of cultivations in the different types of liquid wastes. The relative abundance of the observed taxa significantly changed over the cultivation period. The genome-centric reconstruction of phycospheric bacteria further explained the observed correlations between the taxonomic composition and biomass yield of the various waste-based biodegradation systems. Functional profile investigation of the reconstructed microbes revealed a variety of relevant biological processes like organic acid oxidation and vitamin B synthesis. Thus, liquid wastes were shown to serve as valuable resources of nutrients as well as of growth promoting bacteria enabling increased microalgal biomass production.
Biofuels derived from microalgae are alternative second-generation biofuels having no significant impact on agriculture (Klassen et al., 2016; Rizwan et al., 2018; Wirth et al., 2018). Microalgae have a higher biomass productivity than that of terrestrial crops and can be cultivated on marginal land area all year round. Additionally, the use of microalgae have the potential to directly reduce greenhouse gas emissions (GHG) through the replacement of fossil fuels and by photosynthetic CO2 fixation in their biomass (Lam and Lee, 2012; Yen et al., 2013). Water and nutrients are identified as important limiting resources for microalgae production. The nutrients for microalgae cultivation are readily available in various types of wastewater. Using photoheterotrophic microalgae in biological wastewater treatment represents a dual exploitation of green algae, removing dissolved organic and inorganic pollutants is combined with the production of sustainable bioresource for biofuel production (Mujtaba et al., 2015; Guldhe et al., 2017; Cheah et al., 2018; Vo Hoang Nhat et al., 2018; Li et al., 2019; Shetty et al., 2019). Microalgae have an evolutionary determined ecological relationship with bacteria in natural aquatic environments representing an important interkingdom association (Fuentes et al., 2016). These interactions are strongly influenced by nutrient cycling which regulates the productivity and stability of natural aquatic food webs. The intimate relationship between microalgae and bacteria represents the phycosphere, a key microenvironment ultimately mediating the ecosystem productivity (Cho et al., 2015; Seymour et al., 2017). The exchange of micro- and macronutrients defines the relationship of the interactive partners, which are influenced by a number of key aspects. Firstly, the pH level and the available nutrients determine the surrounding chemical environment, which has a central role in chemotaxis, the motility of bacteria, which enables microbial colonization (Medipally et al., 2015). Secondly, the bacterial communities in the specific ecosystem have important roles in shaping the phycosphere. The most frequently observed bacteria in wastewaters are affiliated with the phyla of the Bacteroidetes and Alpha-, Beta-, and Gammaproteobacteria (with Plant Growth Promoting Bacteria (PGPB) among them) (Guo and Tong, 2014; Kouzuma and Watanabe, 2015; Calatrava et al., 2018). Thirdly, the available microalgae and bacteria synergistically affect each other’s physiology and metabolism. Microalgae produce O2 through photosynthesis for consumption by the actively respiring aerobic bacteria, while bacteria release CO2, which improves the photosynthetic efficiency of green microalgae (Mouget et al., 1995). Another important interkingdom interaction is observed between vitamin-synthetizing bacteria and vitamin auxotrophic microalgae. Most microalgae are auxotrophic for vitamin B derivatives, which are essential for growth and provided by bacteria in exchange for organic carbon (Croft et al., 2005, 2006). Fourthly, the competition for available nutrients, algicidal activities or related defense mechanisms of microalgae are important factors in phycosphere development. Similarly to other natural symbiotic settings, there is only a thin line separating mutualistic and antagonistic associations between microalgae and bacteria (Santos and Reis, 2014; Ramanan et al., 2016).
There are three main sources of wastewater intensively studied in alternative microalgal cultivation; municipal, industrial and agricultural wastewater (Chiu et al., 2015; Guldhe et al., 2017). Utilization of natural microalgal-bacterial communities is a highly promising recycle solution for liquid wastes. This inexpensive and environment-friendly system can contribute to the sustainable management of water resources (Liu J. et al., 2017; Qi et al., 2018). The green microalgae Chlorella vulgaris is the most investigated eukaryotic algae species in wastewater treatment (Chiu et al., 2015; Otondo et al., 2018; Shetty et al., 2019). C. vulgaris is a common eukaryotic microalgae species found in various natural and engineered freshwater and soil habitats. C. vulgaris has a relatively small cell size, thin cell wall, fast growth rate and short reproduction time. This alga is a robust strain that can easily accommodate to changing physico-chemical conditions. Under nutrient limitation and stress C. vulgaris often accumulate high amount of lipids as store materials. These features make this microalgae suitable to cultivate in wastewater, thereby using it for combined wastewater treatment and bioenergy generation (Mussgnug et al., 2010; Collet et al., 2011; Mahdy et al., 2014; Klassen et al., 2016, 2017). It was observed that high nitrogen and phosphorus removal efficiency can be reached with Chlorella species (Chiu et al., 2015; Guldhe et al., 2017; Chen et al., 2018).
A number of studies examined municipal wastewater treatment efficiency using Chlorella-bacteria mixed cultures (Mujtaba et al., 2015; Otondo et al., 2018). More efficient nutrient removal was observed from settled domestic wastewater compared to the commonly used activated sewage process, which indicated the potential of microalgae in the activated sludge process potentially as a secondary step for further nutrient reduction and concomitant biomass production (Otondo et al., 2018). Besides, CO2 originated from the degradation of carbonaceous matter in an activated sludge process is released freely into the atmosphere, thus promoting GHG accumulation. In contrast, microalgae can assimilate CO2 into cellular components such as lipid and carbohydrate, thus achieving pollutant reduction in a more environmental-friendly way (Santos and Reis, 2014; Gonçalves et al., 2017).
In the bioenergy industry biogas is used as a source for generation of heat and/or electricity (Mao et al., 2015; Ullah Khan et al., 2017). Besides biogas, digestate is another important byproduct of anaerobic degradation of organic wastes. Digestate processing is a major bottleneck in the development of the biogas industry. Digestate can be separated into solid (10–20%) and liquid (80–90%) fractions (Xia and Murphy, 2016). Solid digestate is easily stored and transported, and can be used as an agricultural biofertilizer. However, liquid phase processing is more difficult mostly due to its relatively high ammonia content (Uggetti et al., 2014). Digestate is continuously produced, while land application is dependent on the growth stage of the crop and the period of the year. Therefore, digestate needs to be stored, which can increase GHG emission and the general costs as well (Xia and Murphy, 2016; Zhu et al., 2016). Previous studies reported that Chlorella species can be applied to treat liquid digestate (Collet et al., 2011; Skorupskaite et al., 2015; Uggetti et al., 2016). The performance of treatment is dependent on the algae access to carbon, nitrogen and phosphorous as well as on the availability of photosynthetically active light, which indicates a mixotrophic algae growth (Skorupskaite et al., 2015; Zhu et al., 2016).
The rapid growth of the poultry industry in agriculture has raised the need for poultry waste treatment (Sakar et al., 2009). The runoff coming from the chicken farms is highly harmful for the environment through altering the nitrogen and phosphorus balance (Liu Q. et al., 2017). One possible treatment of chicken manure is the anaerobic degradation (Anjum et al., 2017). Chicken manure can be used in small quantities in biogas producing anaerobic fermenters. High dosage of chicken manure cause ammonia accumulation and process failure (Nie et al., 2015; Sun et al., 2016). Water extraction is one possible solution for this problem (Böjti et al., 2017). The supernatant liquid waste still contains high amount of nitrogen and phosphorus, thereby represents suitable medium for microalgal biomass production (Han et al., 2017).
From the biotechnological process point of view the goal is to strengthen the mutually beneficial algal-bacterial interactions to achieve higher biomass growth (beside the bioremediation of liquid wastes). The present study examined and compared different types of wastewater recycling processes using microalgae and their specific bacterial partners. This investigation mainly focused on the interacting bacterial members in specific liquid wastes. The ubiquitous relationship between eukaryotic microalgae and bacteria should be taken into account when designing innovations in microalgal biotechnology (Cooper and Smith, 2015; Gonçalves et al., 2017; Quijano et al., 2017; Lian et al., 2018).
The Chlorella vulgaris MACC-360 microalgae was obtained from the Mosonmagyaróvár Algal Culture Collection (MACC) of Hungary. C. vulgaris was maintained and cultivated on TAP (Tris-acetate-phosphate) plates, then TAP liquid medium (500 mL) was used for the pre-growth of microalgal biomass. The TAP plates and liquid media were incubated at 50 μmol m–2 s–1 light intensity at 25°C for 4 days (OD750: 4.00 ± 0.20). The microalgal stock solution was equally distributed in 17–17 mL portions into 50 mL Falcon tubes with a final optical density (OD750) of 0.70 ± 0.10. Microalgal biomass was separated by centrifugation from the medium and used for inoculation (microalgal dry mass content: ∼100 mg/L). TAP medium was an internal control during the experiment. Different wastewater types were prepared as follows:
Chicken manure (CM) was collected from a commercial broiler poultry farm (Hungerit Corp.) located at Csengele, Hungary. The free-range poultry houses use wheat straw bedding. Water extraction comprised of soaking 2,5 g; 5 g; 10 g and 20 g CM in 100 mL distilled water (v/v %: 2,5; 5; 10 and 20) at room temperature followed by separation of the liquid (CMS: chicken manure supernatant) and solid phases by centrifugation (10,000 rpm for 8 min).
Inoculum sludge was obtained from an operating biogas plant (Zöldforrás Ltd) using pig manure and maize silage mixture as feedstock. The liquid and solid phases were separated by centrifugation (10,000 rpm for 8 min). Distilled water was used to dilute FE (2, 5, 10 and 20 mL effluent in 100 mL distilled water, respectively), to the final concentrations of 2; 5; 10 and 20% (v/v %), respectively.
The municipal wastewater was originated from the Municipal Wastewater Plant of Szeged, Hungary and sampled from the secondary settling tank. The liquid phase was separated from the solid phase by centrifugation (10,000 rpm 8 min). Final concentrations were set at 20 and 50 v/v % using distilled water. Non-diluted (100 v/v %) MW was also used for cultivation.
Cultivation was performed in 250 mL serum bottles (Wheaton glass serum bottle, WH223950) with liquid volume of 200 mL and stirred on a magnetic stirrer tray. Cultivation time was 4 days. Bottles were sealed with paper plugs. Different media were incubated at 50 μmol m–2 s–1 light intensity at 25°C. The OD750 values of the different wastewater media were summarized in Supplementary Information.
The dry matter (DM) content was quantified by drying the biomass at 105°C overnight and weighing the residue. Further heating of this residue at 550°C for 1 h provided the organic dry mass (oDM) content.
To determine C/N (both liquid and biomass), an Elementar Analyzer Vario MAX CN (Elementar Group, Hanau, Germany) was employed. The approach is based on the principle of catalytic tube combustion under O2 supply at high temperatures (combustion temperature: 900°C, post-combustion temperature: 900°C, reduction temperature: 830°C, column temperature: 250°C). The desired components were separated from each other using specific adsorption columns (containing Sicapent (Merck, Billerica, MA, United States), in C/N mode) and were determined in succession with a thermal conductivity detector. Helium served as flushing and carrier gas.
For the determination of NH4+ ion content, the Merck Spectroquant Ammonium test (1.00683.0001) (Merck, Billerica, MA, United States) was used.
Total phosphate content of the different types of wastewater were measured by the standard 4500-PE ascorbic acid, molybdenum blue method (APHA-AWWA-WPCF, 1998).
The VOAs measurement process was carried out using a Pronova FOS/TAC 2000 Version 812-09.2008 automatic titrator (Pronova, Berlin, Germany).
The samples were centrifuged (13,000 rpm for 10 min) and the supernatant was filtered through polyethersulfone (PES) centrifugal filter (PES 516-0228, VWR) at 16,000 g for 20 min. The concentrations of volatile organic acids were measured with HPLC (Hitachi LaChrome Elite) equipped with refractive index detector L2490. The separation was performed on an ICSep ICE-COREGEL—64H column. The temperature of the column and detector was 50 and 41°C, respectively. 0.01 M H2SO4 (0.8 mL min–1) was used as eluent. Acetate, propionate, and butyrate were determined in a detection range of 0.01–10 g L–1. Propionate and butyrate were present in traces relative to acetate and therefore these are not reported in the results section.
To measure the biochemical oxygen demand of the wastewater samples a 5-day BOD test was applied (OxiTop OC 110, Wissenschaftlich-Technische Werkstätten GmbH). In the parallel 500 mL BOD-sample bottles 43 mL of wastewater solution were placed. The results were read after 5 days in mg O2/L.
Experiments were carried out in 160 mL reactor vessels (Wheaton glass serum bottle, Z114014 Aldrich) containing 60 mL liquid phase at mesophilic temperature (37 ± 0.50°C). All fermentations were done in triplicates. The inoculum sludge was filtered to remove particles larger than 1 mm and was used according to the VDI 4630 protocol (Vereins Deutscher Ingenieure 4630, 2006). Each batch fermentation experiment lasted for 30 days in triplicates.
The CH4 content was determined with an Agilent 6890N GC (Agilent Technologies) equipped with an HP Molesive 5 Å (30 m × 0.53 mm × 25 μm) column and a TCD detector. The temperature of the injector was 150°C and split mode 0.2:1 was applied. The column temperature was maintained at 60°C. The carrier gas was Linde HQ argon 5.0 with the flow rate set at 16.80 mL/min. The temperature of TCD detectore was set to 150°C.
In this study data originated from the most effective cultivations under illumination are summarized and highlighted (MW: 100 v/v %, FE: 10 v/v % and CMS: 5 v/v %). All data collected under the various dilution parameters are shown in Supplementary Information.
The composition of the microbial community was investigated two times during the experimental period from each wastewater type and control (TAP), i.e., at the starting point (inoculation) and at the end of cultivation. For total community DNA isolation 2 mL of samples were used from each cultivation media type. DNA extraction and quality estimation were performed according Wirth et al. (2019).
The Ion Torrent PGMTM platform was used for shotgun sequencing, the manufacturer’s recommendations were followed (Life Technologies, United States). Sample preparation, quantification and barcoding were described previously (Wirth et al., 2019). Sequencing was performed with Ion PGM 200 Sequencing kit (4474004) on Ion Torrent PGM 316 chip. The characteristic fragment parameters are summarized in Supplementary Table 1. Raw sequences are available on NCBI Sequence Read Archive (SRA) under the submission number: PRJNA625695.
Galaxy Europe server was employed to pre-process the raw sequences (i.e., sequence filtering, mapping, quality checking) (Afgan et al., 2016). Low-quality reads were filtered by Prinseq (Schmieder and Edwards, 2011) (min. length: 60; min. score: 15; quality score threshold to trim positions: 20; sliding window used to calculated quality score:1). Filtered sequences were checked with FastQC (Supplementary Table 1).
After filtering and checking the passed sequences were further analized by Kaiju applying default greedy run mode on Progenomes2 database (Menzel et al., 2016; Mende et al., 2017). MEGAN6 was used to investigate microbial communities and export data for statistical calculation (Huson et al., 2016). Statistical Analysis of Metagenomic Profiles (STAMP) was used to calculate principal component analysis (PCA) employing ANOVA statistical test (Parks and Beiko, 2010). The distribution of abundant microbial classes between cultivation media were presented with Circos (Krzywinski et al., 2009).
The filtered sequences produced by Prinseq were co-assembled with Megahit (Li et al., 2015) (min. contig length: 2000; min k-mer size: 21; max k-mer size: 141). Bowtie 2 was equipped to mapped back the original sequences to the contigs (Langmead and Salzberg, 2012). Then Anvi’o V5 was used following the “metagenomics” workflow (Eren et al., 2015). Briefly, during contig database generation GC content, k-mer frequencies were computed, open reading frames were identified by Prodigal (Hyatt et al., 2010) and Hidden Markov Modell (HMM) of single-copy genes were aligned by HMMER on each contig (Finn et al., 2011; Campbell et al., 2013; Rinke et al., 2013; Simão et al., 2015). InterProScan v5.31-70 was used on Pfam and Kaiju on NCBInr database for the functional and taxonomic annotation of contigs (Finn et al., 2014, 2017; Jones et al., 2014; Menzel et al., 2016). The taxonomic and functional data were imported into the contig database. BAM files made by Bowtie2 were used to profile contig database, in this way sample-specific information was obtained about the contigs (i.e., mean coverage of contigs) (Langmead and Salzberg, 2012). Three automated binning programs, namely CONCOCT, METABAT2 and MAXBIN2 were employed to reconstruct microbial genomes from the contigs (Alneberg et al., 2013; Kang et al., 2015; Wu et al., 2015). The Anvi’o human-guided binning option was used to refine MAGs Anvi’o interactive interface was employed to visualize and summarize the data. Binning statistics is summarized in Supplementary Table 1. Figure finalization was made by open-source vector graphics editor Gimp 2.10.81. Prokka was employed to translate and map protein sequences (create protein FASTA file of the translated protein coding sequences) (Seemann, 2014). For the calculation of module completion ratio (MCR) MAPLE 2.3.2 (Metabolic And Physiological potentiaL Evaluator) was used (Arai et al., 2018). This automatic system is mapping genes on an individual genome and calculating the MCR in each functional module defined by Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa and Guto, 2000) (Supplementary Table 2).
The bioremediation efficiency of Chlorella vulgaris microalgae and its phycosphere was characterized through the assessment of carbon, nitrogen, phosphate and BOD removal capability of the algal-bacterial biomass (Figure 1). The performance of microalgal-bacterial dry biomass was monitored in three liquid waste types i.e., municipal wastewater (MW), fermentation effluent (FE) and chicken manure supernatant (CMS) over 4 days. The light conditions in the cultivating media are of key importance for microalgal biomass generation. The applied wastewater types are typically dark liquids; therefore, different dilutions with distilled water were prepared in order to increase light penetration to the cultures. Only the experimental data of the most effective dilutions (non-diluted MW, 10 v/v % FE and 5 v/v % CMS) are shown and discussed in the main text of the article (efficiency was defined by the obtained yield of microalgal biomass). However, the nutrient composition of all dilutions for each liquid waste were measured and detailed in Supplementary Information. TAP medium was used as control during the experiments. Significant nutrient removal was observed in all three types of investigated wastewater indicating an active metabolism of the C. vulgaris microalgae and its bacterial partners. However, due to the specific features of the various liquid wastes serving as growth media the algal-bacterial nutrient removal and bioremediation capability was strongly varying. There is a clear correlation between the available nutrients (phosphate, nitrogen and acetate) and the algal-bacterial biomass yield.
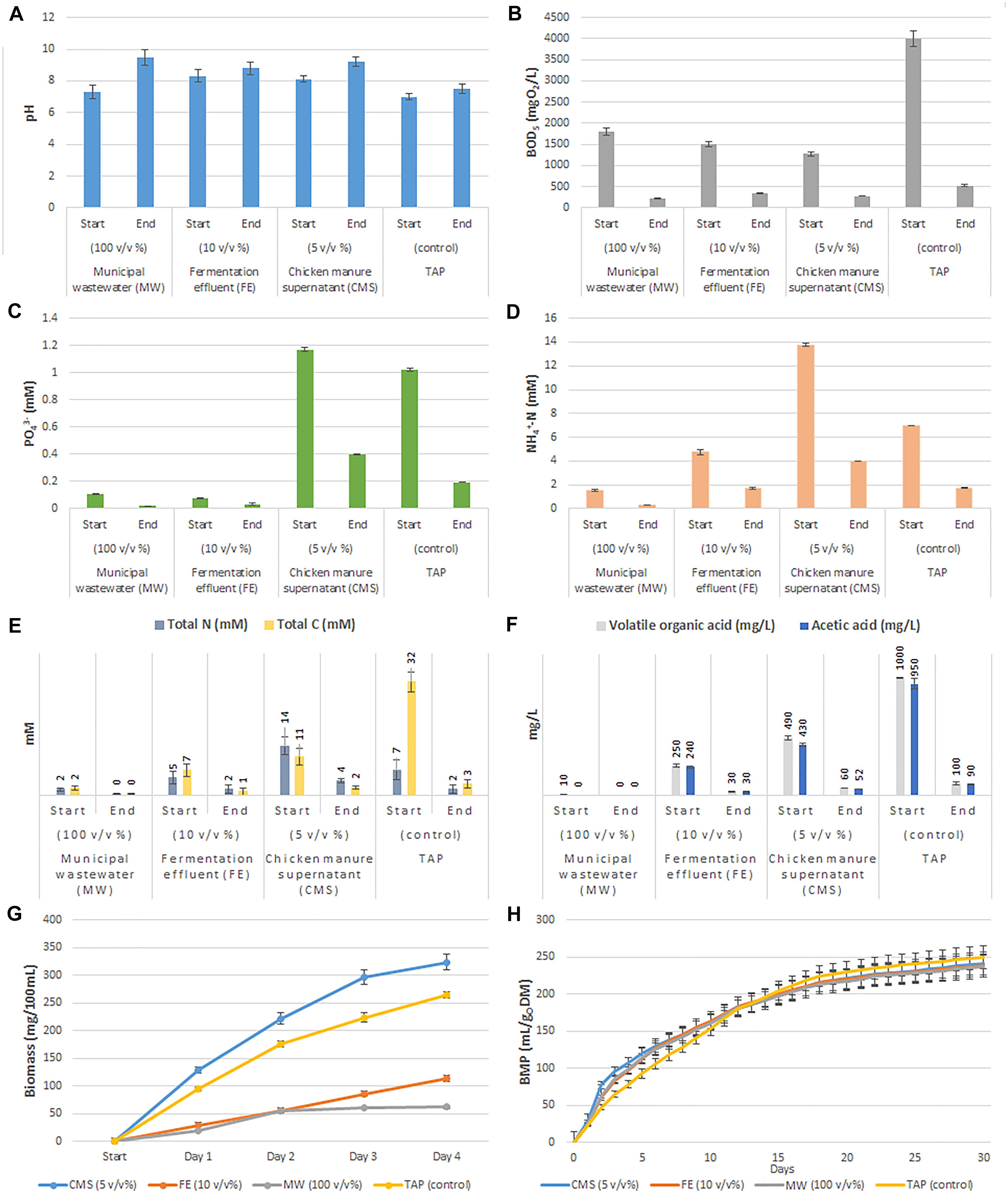
Figure 1. Summary of microalgal-bacterial bioremediation and cultivation efficiency on different types of wastewater. (A) Results of pH measurements. (B) Results of biological oxygen demand calculations. (C) Total phosphate measurements. (D) Ammonium ion measurement data. (E) Total carbon and nitrogen contents. (F) Volatile organic acid (VOAs) and acetic acid concentrations. (G) Biomass growth dynamics over time (days). (H) Cumulative biological methane potential of cultivated biomasses.
The non-diluted municipal wastewater (MW) originated from the second settling tank of a wastewater plant contained the lowest amount of nutrients (acetate and nitrogen) and had the lowest optical density (OD750: 0.02) compared to the 10 v/v % fermentation effluent (FE) originated from a production scale biogas digester (OD750: 0.72) and to the 5 v/v % chicken manure supernatant (CMS: OD750: 0.25) (Supplementary Information). The nutrient removal rate of phosphate and total nitrogen (mostly ammonium) was also shown to be dependent on the light penetration. The highest phosphate removal rate was observed in CMS (0.20 mM day–1), while only 0.02 mM day–1 and 0.01 mM day–1 phosphate uptake were monitored in MW and in FE, respectively (Figure 1C). The monitored phosphate consumption in CMS were comparable to that of measured in TAP medium (0.20 mM day–1). Moreover, in all tested media the microalgal-bacterial consortia removed nitrogen more effectively than phosphate. Total nitrogen removal rate was 0.32 mM day–1 in MW, 0.78 mM day–1 in FE and 2.46 mM day–1 in CMS, respectively (Figure 1E). Similar values were observed for the ammonium content (MW: 0.31 mM day–1, FE: 0.77 mM day–1) and CMS: 2.44 mM day–1) (Figure 1D). Significant organic carbon utilization was observed in all types of liquid wastes. The observed total nitrogen (and ammonium) removal rate were higher in CMS compared to TAP medium (CMS: 2.46 mM day–1 and in TAP: 1.31 mM day–1, respectively). Carbon removal rate was around 82% in all liquid wastes (CMS: 2.20 mM day–1, FE: 1.51 mM day–1, MW: 0.38 mM day–1) (Figure 1E). Likewise, considerable decrease in total VOAs (and acetic acid) was monitored through the experiment (FE: 2 mM day–1, MW and CMS: 3 and 108 mg L–1 day–1) (Figure 1F). As expected, the high C utilization capability of C. vulgaris and its phycosphere is in strong correlation with the BOD consumption (CMS: 78%, FE: 77% and MW: 88%) (Figure 1B). During cultivation pH increase was observed (Figure 1A). The increased pH correlated with the degradation of the organic substrates. The dry mass of the co-cultivated C. vulgaris biomass was the highest in CMS with 0.70–0.90 g DM L–1 day–1, while in FE it was 0.30–0.60 g DM L–1 day–1. The lowest microalgal-bacterial biomass was measured in MW with a value of 0.10–0.20 g DM L–1 day–1. The bacterial biomass was only ∼10% of the total biomass in MW, while these values were ∼38 and ∼27% in FE and CMS, respectively (Supplementary Information and Figure 1G). Highest biomass production was observed in CMS followed by TAP, FE and MW (Figure 1G). The cultivated total algal-bacterial biomass carbon to nitrogen ratio in MW, FE and CMS was 9:1, 7:1 and 6:1, respectively. The higher C/N ratio of MW compared to the TAP control (5:1) might indicate nitrogen limitation in MW. The biochemical methane potential (BMP) of the cultivated mixed biomasses show negligible differences compared to the TAP control (TAP: 249 ± 15 CH4 mLN g oDM–1; MW:236 ± 14 CH4 mLN g oDM–1; FE: 238 ± 14 CH4 mLN g oDM–1 and CMS: 241 ± 15 CH4 mLN g oDM–1) (Figure 1H).
An average of 271,721 sequence reads were generated for each sample, with a mean read length of 231 nucleotides using an Ion Torrent PGM sequencing platform. Sequence reads were quality filtered by Prinseq, this resulted in an average of 266,119 reads with a mean length of 232 nucleotides (Supplementary Table 1). The sequences were analyzed and bacterial partners of C. vulgaris were identified using the Kaiju software on Progenomes2 database. The comparison of the prokaryotic microbes using PCA showed significant community shifts between the different wastewater samples over cultivation time (Figure 2A). At the start point (T0) the CMS, FE and MW liquid wastes have diverse microbial community (Figure 2B). The most abundant classes in CMS were Actinobacteria (55%), Bacilli (27%) and Gammaproteobacteria (7%), while in FE Clostridia (33%), Bacteroidia (27%), Bacilli (8%), and in MW Beta- and Gammaproteobacteria (23–23%) as well as Actinobacteria (13%) dominated. The relative abundance of the observed taxa significantly changed over the cultivation period. The Alpha-, Beta- and Gammaproteobacteria and Bacilli classes dominated the prokaryotic community at the end point of the experiments (CMS: Gammaproteobacteria 74%, Alphaproteobacteria 11%, Betaproteobacteria 7%; FE: Alphaproteobacteria 60%, Gammaproteobacteria 17%, Betaproteobacteria 16%; MW: Alphaproteobacteria 52%, Bacilli 40%, Gammaproteobacteria 4%, respectively). The control TAP media showed the least microbial shift between the start and the end of the cultivation, where representatives of the Gammaproteobacteria class (T0: 100%; end: 95%, respectively), were the most abundant (Figures 2A,B).

Figure 2. (A) PCA of prokaryotic communities in various cultivation media. The variation represented by the first axis (PC1, 64% of overall variation) and the second axis (PC2, 19% of overall variation) indicating diverse phylogenetic structures. (B) Relative distribution of abundant microbial classes in different media (left side: classes, right side: cultivation media).
Metagenome assembly was carried out by Megahit. A total of 6,148 contigs with a minimum length of 2,000 nucleotides were generated. The contigs were then binned together using MAXBIN2, METABAT2 and CONCOCT automated binning programs. The generated bins were further refined by human guided binning process based on automated binning results with Anvi’o. The 7 bins accounted for a total of 20,038,573 nucleotides. Bins were checked for completion and contamination using CheckM.
Seven metagenome assembled genomes (MAGs) were generated by Anvi’o (Figure 3). Bin 1 contained the C. vulgaris genome fragments. Beside Bin 1 six bacterial MAGs were detected. From these six MAGs five belonged to partly unknown taxa, namely Pseudomonas, Exiguobacterium, Acinetobacter, Enterobacteriaceae and Bacteroidetes. The unknown Pseudomonas (Bin 2) showed a high degree of genome completeness (95%). This MAG included ribosomal maturation proteins (Supplementary Table 2), however, 16S rRNA sequences were not found by HMMER (Bowers et al., 2017). One species level bin (Bin 6) belonged to the Bacteroidetes bacterium 4484_276. By mapping back the original reads to the unknown Pseudomonas (Bin 2) and unknown Acinetobacter (Bin 3) bins it was observed, that these microbes were detected in all cultivation media at each time point. The unknown Enterobacteriaceae (Bin 5) was found in all liquid waste cultivations (i.e., MW, FE, CMS), while the unknown Exiguobacterium (Bin 4) occurred only in MW. The low quality Bacteroidetes bacterium 4484_276 (Bin 6) and the unknown Bacteroidetes (Bin 7) bins were detected only in FE.
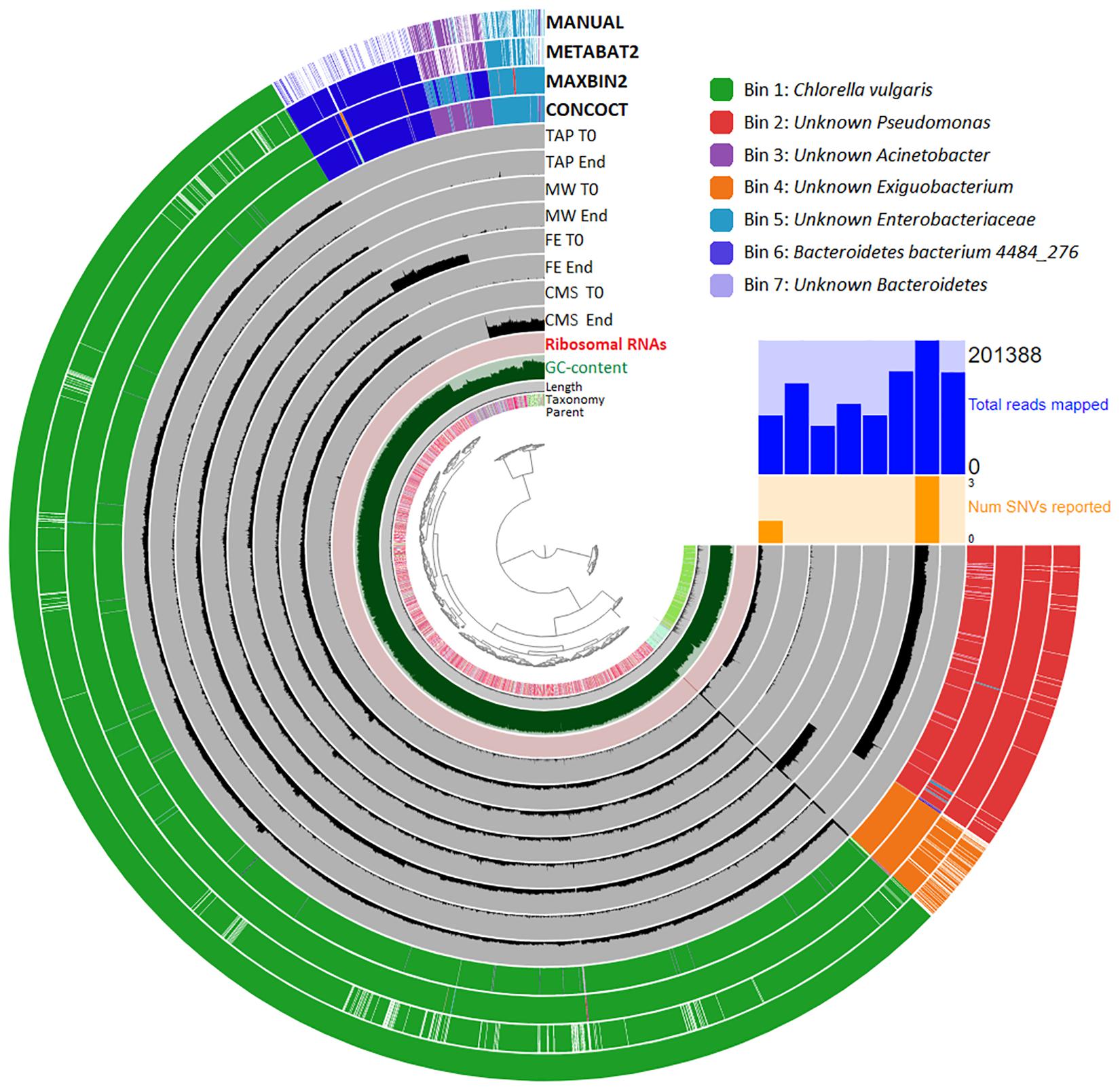
Figure 3. Visualization of the genome-centric metagenomics data. The hierarchical dendrogram of the contigs based on its tetra-nucleotide frequency is in the center of the picture. The taxonomy of the individual contigs is color-coded. The length, GC content, and the presence of ribosomal RNAs of contigs is displayed at the next level. The inner circles show the frequency of the contigs in each wastewater type (CMS: chicken manure supernatant, FE: fermentation effluent, MW: municipal wastewater) in time (T0: start point, End: end point). The outer layer shows the CONCOCT, MAXBIN2, METABAT2 and manual binning results (color-code: upper right corner).
To predict protein pathways, the translated protein coding sequences created by Prokka were further analyzed to calculate module completion ratio (MCR) by MAPLE 2.3.2 using the Kegg database (Kanehisa and Guto, 2000; Seemann, 2014; Arai et al., 2018). The unknown Pseudomonas (Bin 2) bin genom harbored complete pathways of gluconeogenesis, Entner-Doudoroff pathway, pyruvate-oxidation, beta-oxidation, sulphate reduction, pentose phosphate pathway, fatty acid, amino acid, cofactor and vitamin metabolism (Supplementary Table 2). The MCR of vitamin B biosynthesis was also found at high percentage in the unknown Pseudomonas MAG. Among vitamin B variants, the complete biotin (B7) biosynthesis pathway was detected (100%) in Bin 2, while the completeness of cobalamin (B12) and thiamin (B1) biosynthesis pathways were 86% and 60%, respectively. Between the MAGs showing low degree of genome completeness the unknown Acinetobacter (Bin 3) and the unknown Enterobacteriaceae bin (Bin 5) had complete MCRs for acetate kinase pathway, while the unknown Exiguobacterium (Bin 4) and Bacteroidetes bacterium 4484_276 (bin 6) bins had complete phospho-ribose-diphosphate pathway. The unknown Bacteroidetes (Bin 7) had the lowest genome completeness among the detected MAGs, therefore complete pathways could not be detected in this bin (Supplementary Table 2).
Microalgae and their phycosphere represent powerful natural associations, which can be exploited in bioremediation and biofuel production (Gonçalves et al., 2017; Guldhe et al., 2017). Using liquid wastes for alternative algae cultivation has emerged as a potential cost effective strategy to make microalgae biotechnology more sustainable and economically feasible. It is essential to understand the nature of microalgal-bacterial relationships in order to develop combined bioremediation and biofuel production systems. Therefore, the main objective in this study was the assessment of nutrient removal and microalgal-bacterial biomass production efficiency using different types of wastewater sources (i.e., chicken manure supernatant, fermentation effluent and municipal wastewater). Furthermore, bioremediation and production efficiency data were supported by applying read-based and novel genome-centric approach for the identification of the phycosphere components and their functional profiles.
THE following major bioremediation process parameters were measured during the experiments: pH, biomass yield, carbon, nitrogen and phosphorous content. The biomass’ carbon/nitrogen ratio and biochemical methane potential were also characterized. The experiments were designed for 4 days, since previous literature data indicated that C. vulgaris entered stationary growth phase by the 4th–5th day, no significant biomass production could be observed thereafter (Mujtaba et al., 2015, 2017; Otondo et al., 2018; Qi et al., 2018) (Supplementary Information).
The total carbon (TC), total nitrogen (TN) and phosphate (PO43–) concentrations of the applied liquid wastes substantially varied (Figures 1E,C and Supplementary Information). The major nutrients required for microalgal growth are nitrogen and phosphorus incorporated to the cells via active transport. Ammonium is among the most common forms of nitrogen that can easily be utilized by most microalgal species (Gonçalves et al., 2017). Thus, liquid wastes represent a cheap source of nitrogen for microalgal cultivation (Razzak et al., 2013). Previously it was observed, that the optimal ammonium concentration for microalgal cultivation was around 8–10 mM (Uggetti et al., 2014; Chen et al., 2018), higher concentration might inhibit microalgal growth (Källqvist and Svenson, 2003). Another important element required for microalgae growth and metabolism is phosphorus primarily occurring in the form of phosphate (PO43–) in wastewater. Phosphorus is an essential ingredient of ATP and nucleic acids in the cells. Phosphate availability has a large impact on microalgal photosynthesis as well (Razzak et al., 2013). Optimal phosphate concentration was found around ∼1 mM (Chiu et al., 2015). The concentration of ammonium and phosphate were relatively low in the applied non-diluted MW (NH4+-N: 1.6 mM; PO43–: 0.1 mM) (Figures 1C,D and Supplementary Information). In the diluted FE (10 v/v%) the amount phosphate was low (PO43–: 0.1 mM), while the ammonium content was approximately half of the optimum (NH4+-N: 4.8 mM). The diluted CMS (5 v/v%) contained high amount of both nutrients (NH4+-N: 13.7 mM; PO43–: 1.2 mM) (Figures 1C,D and Supplementary Information). The ammonium and phosphate removal rates were also high in CMS (NH4+-N: 2,44 mM day–1; PO43–: 0.20 mM day–1), while lower in FE (NH4+-N: 0.77 mM day–1; PO43–: 0.01 mM day–1) and MW (NH4+-N: 0.31 mM day–1; PO43–: 0.02 mM day–1). The experimental data indicated that mostly C. vulgaris was responsible for the removal of ammonium and phosphate, and the biomass yield strongly correlated with the removal efficiencies. The results also implied to the dependency of microalgae growth on the available nitrogen sources, which is in good correlation with previous studies (Chiu et al., 2015). The observed low nitrogen content of the biomass generated on MW compared to the TAP control might be explained by the nitrogen limitation (Klassen et al., 2015; Seger et al., 2019).
Microalgae can fix CO2 derived from flue gas emission through photosynthesis (Sayre, 2010; Pires et al., 2012). Additionally, microalgae are able to uptake soluble carbonates as a source of CO2 (Thomas et al., 2016; Sydney et al., 2019). This uptake depends on the environmental pH. At low pH values the CO2 uptake occurs through diffusion (pH 7 ± 1), while in the case of bicarbonate, which is the common form of inorganic carbon under high pH (10 ± 1), the microalgal cells use active transport (Gonçalves et al., 2017). Microalgal photosynthesis raises pH by consumption of CO2 and HCO3–. It was observed that microalgal growth rate is affected by the pH as pH affects the availability of inorganic carbon. When pH is around or over 10, CO2 is limiting and bicarbonate is used as a carbon source (Otondo et al., 2018). The pH is slightly increased during the microalgal-bacterial biomass generation in all type of liquid wastes indicating effective photosynthetic activity of microalgae. At the end point of the biomass production in MW the pH was high, this might have been an inhibitory on microalgal biomass growth beside the limited nutrient source (Figure 1).
Although microalgae are mainly autotrophic, C. vulgaris is able to grow in a mixotrophic/photoheterotrophic way using organic carbon source (e.g., acetate, glucose) in addition to CO2 (Skorupskaite et al., 2015; Zuñiga et al., 2016). Typically both respiratory and photosynthetic processes occur in darkish wastewater (Morales-Sánchez et al., 2015; Skorupskaite et al., 2015; Zuñiga et al., 2016). Microalgae also consume the CO2 released from bacterial respiration, in turn the algae provide the O2 necessary for the phycospheric bacteria to degrade organic carbon sources (Fuentes et al., 2016; Liu J. et al., 2017). Therefore, organic carbon source of liquid wastes is readily reduced by both microalgal and bacterial metabolic activities. Furthermore, it was observed earlier that microalgae could improve the energy efficiency of BOD removal (Mujtaba and Lee, 2016). These observations were confirmed, significant carbon loss was detected in all type of applied wastewaters (over 80%), which was in clear correlation with the BOD removal rate.
Using microalgae and its phycosphere to utilize nutrients from wastewater for biomass production and the combined use of the generated biomass for biofuel generation is a promising and promoted way to build circular economy (Chiu et al., 2015; Zhu et al., 2016). The advantage of the algal biomass-based biogas production is that the microalgal-bacterial biomass can be directly applied in the biogas reactor, the total biomass is degraded and converted to methane and CO2 by a complex microbial community in a well-controlled manner (Guldhe et al., 2017). Microalgal dry biomass productivity was found to be the most effective in CMS (18% higher compared to TAP) followed by FE (CMS: 0.70–0.90 g DM/L/day; FE: 0.30–0.60 g DM/L/day), while the lowest biomass was detected when using MW (0.10–0.20 g DM/L/day) (Supplementary Information and Figure 1G). Similarly, bacterial content was found to be higher in biomass generated in CMS and FE (27 and 38%), while only 10% in MW. The high nutrient content (including acetate, phosphate and ammonium) of CMS explains its effectiveness in biomass production. The biochemical methane potential (BMP) of the biomass generated in the alternative media were comparable to the methane potential of the biomass produced on TAP control (ranging from 236 to 241 CH4 mLN/g oDM in CMS, FE, and MW, while 249 ± 15 CH4 mLN/g oDM in TAP). Differences in BMP might be caused by the biomass carbon to nitrogen ratio and by bacterial content of the biomass (Arcila and Buitrón, 2016; Molinuevo-Salces et al., 2016; Jankowska et al., 2017). The presence of bacteria also explains the relatively higher C/N ratio of biomass cultivated in FE and CMS compared to that of TAP. However, in the aspect of anaerobic digestion this ratios are far from the optimal range (C/N: 20–30:1) (Ward et al., 2014). Thus, the long-term effects of the low C/N ratio and the bacterial content of the biomass on the anaerobic digestion and on the decomposing microbial community need to be further investigated (Wirth et al., 2015a,b, 2018).
The read-based metagenomics approach revealed a diverse microbial composition at the start point of cultivations in different type of liquid wastes (Supplementary Information). The PCA of the prokaryotic communities showed significant alterations during the cultivation period (Figure 2A). At the starting point the highest diversity was observed in FE, where Clostridia, Bacteroidia and Bacilli were the most abundant classes. Beta,- Gammaproteobacteria and Bacilli dominated the microbial communities in MW. Actinobacteria, Bacilli and Gammaproteobacteria were the most abundant classes in CMS (Figure 2B). The observed microbial classes are typical for chicken manure, municipal wastewater and anaerobic digesters (Lu et al., 2007; Ju et al., 2014; Campanaro et al., 2020). The starting communities were significantly altered by the end of the cultivation period. Mainly Alpha-, Beta-, Gammaproteobacteria and Bacilli became the most dominant classes (Figure 2B). In previous studies similar changes were observed in the prokaryotic microbial community composition in microalgal-seeded systems (Krustok et al., 2015; Chen et al., 2019; Paquette et al., 2020). The TAP medium (control) showed the lowest composition change, in this medium the representatives of Gammaproteobacteria class were the dominant bacterial partners of C. vulgaris microalgae throughout the cultivation. Two further interesting aspects were observed in the microbial communities. On one hand the prokaryotic community of CMS at the end point was the most similar to that of the TAP medium (Figure 2A). On the other hand the dominance of the class Gammaproteobacteria is in close correlation with the biomass yield (Figures 1, 2B).
The genome-centric metagenomics results further explain these interesting observations. The human-guided binning approach resulted one medium (Bin 2) and six low quality (Bin 1, 3–7) Metagenome-Assembled Genomes (MAGs) (Bowers et al., 2017). These bins are identified as one eukaryotic algae MAG (Bin 1) and six bacterial MAGs (Bin 2–7). The unknown Pseudomonas (Bin 2), unknown Acinetobacter (Bin 3) and unknown Enterobacteriaceae (Bin 5) belong to the class Gammaproteobacteria within the phylum Proteobacteria. Two bins were found as representatives of the phylum Bacteroidetes, these are the Bacteroidetes bacterium 4484-246 MAG (Bin 6) and an unknown Bacteroidetes MAG (Bin 7), while the unknown Exiguobacterium MAG (Bin 4) belongs to the phylum Firmicutes (Figure 3).
Multiple members of the class Gammaproteobacteria and the phylum Bacteroidetes are considered as Plant Growth Promoting Bacteria (PGPB) interacting with microalgae trough metabolite exchange and by enhancing the microalgal biomass yield and lipid production (Seymour et al., 2017; Calatrava et al., 2018; Cho et al., 2019). The representatives of class Gammaproteobacteria, the phylum Bacteroidetes and the genus Exiguobacterium are commonly found in the phycosphere of C. vulgaris cultivated on liquid wastes strengthening the hypothesis, that there are a specific interactions between microalgae and bacteria (Guo and Tong, 2014; Kouzuma and Watanabe, 2015; Mujtaba et al., 2017; Cheah et al., 2018; Qi et al., 2018). It was reported that the representatives of the genus Pseudomonas are capable of increasing the growth rate of Chlorella microalgae species through the reduction of photosynthetic oxygen tension (Berthold et al., 2019) beside their decomposing activities (Mujtaba et al., 2017; Cheah et al., 2018). The presence of Pseudomonas sp. resulted higher Chlorella cell concentrations in a given period compared to that observed in axenic microalgae culture (Guo and Tong, 2014; Mujtaba and Lee, 2016). Certain Pseudomonas and Acinetobacter sp. also promoted the Chlorella microalgae growth when cultivated on palm oil mill effluent (Cheah et al., 2018). A symbiotic relationship between Chlorella and Bacteroidetes species was described recently, the abundance of Bacteroidetes specifically increased during pre-treatment of dairy-derived liquid digestate (Zhu et al., 2019). In another study Proteobacteria and Bacteroidetes induced growth promotion of three microalgae, Chlamydomonas reinhardtii, C. vulgaris and Euglena gracilis in wastewater and swine manure effluent (Toyama et al., 2018). The genus Exiguobacterium was previously described among the dominant bacteria during domestic wastewater treatment, this specific bacterium was shown to promote Chlorella biomass accumulation and chlorophyll synthesis (Qi et al., 2018; Ren et al., 2019).
The read coverage of bins indicated that the unknown Pseudomonas (Bin 2) and unknown Acinetobacter (Bin 3) were presented in all types of wastewater media. The unknown Enterobacteriaceae (Bin 5) was detected in CMS, FE and MW, while Bacteroidetes bacterium 4484_276 (Bin 6) and the unknown Bacteroidetes (Bin 7) were present only in FE. These data indicated that some of the bacteria were in strong interaction with the Chlorella algae while the others were specific to the applied wastewater type. It was reported that many bacteria are able to survive together with microalgae in algae culture collections for long term (Krohn-Molt et al., 2017). The unknown Pseudomonas (Bin 2) and the unknown Acinetobacter (Bin 3) seem to belong this category, they had a strong interaction with Chlorella and might have been inoculated together into the examined waste liquids. The unknown Enterobacteriaceae and Exiguobacterium, furthermore the representatives of Bacteroidetes are likely to be wastewater-specific bacterial strains (Toyama et al., 2018).
Multiple factors influence the presence of bacterial partners of eukaryotic microalgae. A highly important factor is the algal photosynthesis, through which microalgae can increase the dissolved oxygen concentration and the pH of the medium (Seymour et al., 2017). Also the microalgal products having bactericidal effect are important in shaping the phycosphere. The C. vulgaris are able to produce a mixture of polyunsaturated fatty acids exhibiting antibiotic activity, i.e., chlorellin (Fergola et al., 2007). Chlorellin is produced in small amount in stationary growth phase, and it exerts different inhibitory effects on different bacteria (DellaGreca et al., 2010; Alwathnani and Perveen, 2017). The effect of chlorellin might have been limited on the development of the phycosphere due to the applied short cultivation time (4 days). Nevertheless, bacteria are also able to influence microalgal growth through nutrient competition (Guldhe et al., 2017). Based on the measurement of the key nutrients and binning results, microalgae and bacteria are competing for VOAs (i.e., acetate). C. vulgaris is able to use acetate in photoheterotrophic cultivation mode via active transport (Zuñiga et al., 2016; Huang et al., 2017; Cecchin et al., 2018). The functional profiling of the unknown Pseudomonas (Bin 2), unknown Acinetobacter (Bin 3) and unknown Enterobacteriaceae (Bin 5) resulted in pathways with complete module completion ratio (MCR). These pathways are linked to fatty acid metabolism (Supplementary Table 2). Therefore, it is assumed that these bacteria were mainly responsible for the fatty acid consumption, while the microalgae had only minor role in this metabolic activity. They degrade the fatty acids and release CO2 during their metabolic activity, this CO2 is consumed by microalgae which in turn produce photosynthetic oxygen essential for the bacteria for fatty acid oxidation. According to MCR calculations the unknown Exiguobacterium (Bin 4) and the Bacteroidetes bacterium 4484-246 (Bin 6) have complete phospho-ribo-biphosphate biosynthesis pathway indicating their carbohydrate metabolic activity. It is not clear, whether these bacteria use the microalgal carbohydrate by-products or possibly degrade algal cell wall components. However, it is very likely that these bacteria also produce CO2, thereby increase microalgal photosynthetic activity and growth. Since the genome completeness of these bacteria is low, similarly to the unknown Acinetobacter (Bin 3) and the unknown Enterobacteriaceae (Bin 5), the knowledge on their detailed roles in the phycosphere is limited.
Vitamins like cobalamin, thiamin, biotin are needed in the lipid biosynthesis pathway in microalgae and higher plants (Croft et al., 2006; Smith et al., 2007). Although C. vulgaris is not auxotroph for vitamin B derivatives, the addition of these ingredients still have a positive effect for Chlorella growth (Croft et al., 2005). Previous studies involving 306 microalgal species showed that more than half of the examined species (51%) required exogenous cobalamin (vitamin B12), 22% required thiamin (vitamin B1) and 5% required biotin (vitamin B7) for better growth (Croft et al., 2006). It was reported that vitamin supplementation increased the lipid production and intracellular vitamin concentration of the Chlorella species, which ultimately resulted in increased growth rate and biomass yield (Fazeli Danesh et al., 2018). It is possible to supply these vitamins by the addition of bacterial partners. It is especially beneficial at industrial scale algae farms to increase sustainability and economic feasibilty. The genome-centric binning results showed that the unknown Pseudomonas (Bin 2) showed high MCR for biotin (100%), cobalamin (80%) and thiamin (60%) biosynthesis. The capability of this specific MAG to synthesize these important vitamin B derivatives further supports the close relationship between this bacterium and the C. vulgaris microalgae.
The applied microalgae and its phycosphere effectively reduced the carbon, nitrogen and phosphorus content as well as decreased the BOD of the applied liquid wastes. The nitrogen and phosphorus losses were predominantly caused by the microalgal activity. Nitrogen had the greatest effect on the growth of microalgae, however, the algal consumption of this nutrient depended on the transparency of the medium (light penetration) implying to the significance of the photosynthetic algae growth. The fatty acid content of the liquid wastes was used by both the microalgae and the bacterial partners, however, microalgae had limited importance in this activity. The CO2 produced by the phycospheric bacteria was consumed by microalgae and in exchange the photosynthetically produced oxygen was respired by the phycospheric bacteria during the oxidation of organic acids. CMS proved to be the most efficient for microalgal dry mass production, while FE and MW had medium and low efficiency in this term, respectively. However, the lowest bacterial content was detected in the dry biomass grown in MW. Diverse prokaryotic microbial community featured the used liquid wastes at the start point of cultivation, which compositions are typical to the given wastewater type. These were significantly changed at the endpoint. The genom-centric approach revealed that the unknown Pseudomonas (Bin 2) and the unknown Acinetobacter (Bin 3) strongly interacted with Chlorella. Such genome-level investigations may reveal bacterial indicators of culture status, which could be useful for monitoring the health of microalgae in complex bioremediating communities (Seger et al., 2019). The explorations on microalgae-bacteria associations in wastewater contribute to the better understanding of phycosphere activities and help their applications in bioremediation and combined next-generation biofuel production.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number (s) can be found in the article/ Supplementary Material.
RW designed and performed the bioinformatics analyses and composed the manuscript. BP, TB, GL, and ZB performed the wastewater cultivation experiments and analytical measurements. PS contributed to the metagenome analyses. KK and GM designed the study, composed the manuscript and thoroughly discussed the relevant literature. All authors read and approved the final manuscript.
This study has been supported in part by the Hungarian National Research, Development and Innovation Fund projects GINOP-2.2.1-15-2017-00081, GINOP-2.2.1-15-2017-00033, and EFOP-3.6.2-16-2017-00010. RW and GM received support from the Hungarian NKFIH fund projects PD121085 and FK123899. This work was also supported by the János Bolyai Research Scholarship (GM) of the Hungarian Academy of Sciences and by a Bolyai+ Grant UNKP-19-4-SZTE-70 (GM) and by the Lendület-Programme (GM) of the Hungarian Academy of Sciences (LP2020-5/2020).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fbioe.2020.557572/full#supplementary-material
SUPPLEMENTARY INFORMATION | Bioremediation data, biomass production and biochemical methane production (BMP) measurements.
TABLE S1 | Sequence statistics, read-based and genome-centric data.
TABLE S2 | Results of module completion ratio (MCR) calculations.
BMP, biochemical methane potential (test); BOD, biological oxygene demand (test); C/N, carbon to nitrogen ratio; CMS, chicken manure supernatant (medium); DM and oDM, dry mass and organic dry mass; FE, anaerobe fermentation effluent (medium); GHG, Green house gas (emission); MAGs, metagenome assembled genomes; MCR, module completion ratio; MW, municipal wastewater (medium); PCA, principal component analysis; PGPB, plant growth promoting bacteria; TAP, tris-acetate-phosphate (medium); TC and TN, total carbon and total nitrogen; VOAs, volatile organic acids.
Afgan, E., Baker, D., van den Beek, M., Blankenberg, D., Bouvier, D., Čech, M., et al. (2016). The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 44, W3–W10. doi: 10.1093/nar/gkw343
Alneberg, J., Bjarnason, B. S., de Bruijn, I., Schirmer, M., Quick, J., Ijaz, U. Z., et al. (2013). CONCOCT: clustering contigs on coverage and composition. arXiv 1–28. doi: 10.1371/journal.pone.0005299
Alwathnani, H., and Perveen, K. (2017). Antibacterial activity and morphological changes in human pathogenic bacteria caused by chlorella vulgaris extracts. Biomed. Res. 28, 1610–1614.
Anjum, R., Grohmann, E., and Krakat, N. (2017). Anaerobic digestion of nitrogen rich poultry manure: impact of thermophilic biogas process on metal release and microbial resistances. Chemosphere 168, 1637–1647. doi: 10.1016/j.chemosphere.2016.11.132
APHA-AWWA-WPCF (1998). Standard Methods for the Examination of Water and Wastewater, 20th Edn. Washington, DC: American Public Health Association.
Arai, W., Taniguchi, T., Goto, S., Moriya, Y., Uehara, H., Takemoto, K., et al. (2018). MAPLE 2.3.0: an improved system for evaluating the functionomes of genomes and metagenomes. Biosci. Biotechnol. Biochem. 82, 1515–1517. doi: 10.1080/09168451.2018.1476122
Arcila, J. S., and Buitrón, G. (2016). Microalgae–bacteria aggregates: effect of the hydraulic retention time on the municipal wastewater treatment, biomass settleability and methane potential. J. Chem. Technol. Biotechnol. 91, 2862–2870. doi: 10.1002/jctb.4901
Berthold, D. E., Shetty, K. G., Jayachandran, K., Laughinghouse, H. D., and Gantar, M. (2019). Enhancing algal biomass and lipid production through bacterial co-culture. Biomass Bioenergy 122, 280–289. doi: 10.1016/j.biombioe.2019.01.033
Böjti, T., Kovács, K. L., Kakuk, B., Wirth, R., Rákhely, G., and Bagi, Z. (2017). Pretreatment of poultry manure for efficient biogas production as monosubstrate or co-fermentation with maize silage and corn stover. Anaerobe 46, 138–145. doi: 10.1016/j.anaerobe.2017.03.017
Bowers, R. M., Kyrpides, N. C., Stepanauskas, R., Harmon-Smith, M., Doud, D., Reddy, T. B. K., et al. (2017). Minimum information about a single amplified genome (MISAG) and a metagenome-assembled genome (MIMAG) of bacteria and archaea. Nat. Biotechnol. 35, 725–731. doi: 10.1038/nbt.3893
Calatrava, V., Hom, E. F. Y., Llamas, Á, Fernández, E., and Galván, A. (2018). Ok, thanks! A new mutualism between chlamydomonas and methylobacteria facilitates growth on amino acids and peptides. FEMS Microbiol. Lett. 365:fny021. doi: 10.1093/femsle/fny021/4828328
Campanaro, S., Treu, L., Rodriguez-R, L. M., Kovalovszki, A., Ziels, R. M., Maus, I., et al. (2020). New insights from the biogas microbiome by comprehensive genome-resolved metagenomics of nearly 1600 species originating from multiple anaerobic digesters. Biotechnol. Biofuels 13, 1–18. doi: 10.1186/s13068-020-01679-y
Campbell, J. H., O’Donoghue, P., Campbell, A. G., Schwientek, P., Sczyrba, A., Woyke, T., et al. (2013). UGA is an additional glycine codon in uncultured SR1 bacteria from the human microbiota. Proc. Natl. Acad. Sci. U.S.A. 110, 5540–5545. doi: 10.1073/pnas.1303090110
Cecchin, M., Benfatto, S., Griggio, F., Mori, A., Cazzaniga, S., Vitulo, N., et al. (2018). Molecular basis of autotrophic vs mixotrophic growth in Chlorella sorokiniana. Sci. Rep. 8:6465. doi: 10.1038/s41598-018-24979-8
Cheah, W. Y., Show, P. L., Juan, J. C., Chang, J. S., and Ling, T. C. (2018). Waste to energy: the effects of Pseudomonas sp. on Chlorella sorokiniana biomass and lipid productions in palm oil mill effluent. Clean Technol. Environ. Policy 20, 2037–2045. doi: 10.1007/s10098-018-1505-7
Chen, X., Hu, Z., Qi, Y., Song, C., and Chen, G. (2019). The interactions of algae-activated sludge symbiotic system and its effects on wastewater treatment and lipid accumulation. Bioresour. Technol. 292:122017. doi: 10.1016/j.biortech.2019.122017
Chen, X., Li, Z., He, N., Zheng, Y., Li, H., Wang, H., et al. (2018). Nitrogen and phosphorus removal from anaerobically digested wastewater by microalgae cultured in a novel membrane photobioreactor. Biotechnol. Biofuels 11:190. doi: 10.1186/s13068-018-1190-0
Chiu, S. Y., Kao, C. Y., Chen, T. Y., Chang, Y., Bin, Kuo, C. M., et al. (2015). Cultivation of microalgal Chlorella for biomass and lipid production using wastewater as nutrient resource. Bioresour. Technol. 184, 179–189. doi: 10.1016/j.biortech.2014.11.080
Cho, D. H., Ramanan, R., Heo, J., Lee, J., Kim, B. H., Oh, H. M., et al. (2015). Enhancing microalgal biomass productivity by engineering a microalgal-bacterial community. Bioresour. Technol. 175, 578–585. doi: 10.1016/j.biortech.2014.10.159
Cho, K., Heo, J., Cho, D.-H., Tran, Q.-G., Yun, J.-H., Lee, S.-M., et al. (2019). Enhancing algal biomass and lipid production by phycospheric bacterial volatiles and possible growth enhancing factor. Algal Res. 37, 186–194. doi: 10.1016/j.algal.2018.11.011
Collet, P., Hélias Arnaud, A., Lardon, L., Ras, M., Goy, R. A., and Steyer, J. P. (2011). Life-cycle assessment of microalgae culture coupled to biogas production. Bioresour. Technol. 102, 207–214. doi: 10.1016/j.biortech.2010.06.154
Cooper, M. B., and Smith, A. G. (2015). Exploring mutualistic interactions between microalgae and bacteria in the omics age. Curr. Opin. Plant Biol. 26, 147–153. doi: 10.1016/j.pbi.2015.07.003
Croft, M. T., Lawrence, A. D., Raux-Deery, E., Warren, M. J., and Smith, A. G. (2005). Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature 438, 90–93. doi: 10.1038/nature04056
Croft, M. T., Warren, M. J., and Smith, A. G. (2006). Algae need their vitamins. Eukaryot. Cell 5, 1175–1183. doi: 10.1128/ec.00097-06
DellaGreca, M., Zarrelli, A., Fergola, P., Cerasuolo, M., Pollio, A., and Pinto, G. (2010). Fatty acids released by Chlorella vulgaris and their role in interference with Pseudokirchneriella subcapitata: experiments and modelling. J. Chem. Ecol. 36, 339–349. doi: 10.1007/s10886-010-9753-y
Eren, A. M., Esen, ÖC., Quince, C., Vineis, J. H., Morrison, H. G., Sogin, M. L., et al. (2015). Anvi’o: an advanced analysis and visualization platform for ‘omics data. PeerJ 3:e1319. doi: 10.7717/peerj.1319
Fazeli Danesh, A., Mooij, P., Ebrahimi, S., Kleerebezem, R., and van Loosdrecht, M. (2018). Effective role of medium supplementation in microalgal lipid accumulation. Biotechnol. Bioeng. 115, 1152–1160. doi: 10.1002/bit.26548
Fergola, P., Cerasuolo, M., Pollio, A., Pinto, G., and DellaGreca, M. (2007). Allelopathy and competition between Chlorella vulgaris and Pseudokirchneriella subcapitata: experiments and mathematical model. Ecol. Modell. 208, 205–214. doi: 10.1016/j.ecolmodel.2007.05.024
Finn, D. R., Clements, J., and Eddy, R. S. (2011). HMMER web server: interactive sequence similarity searching. Nucleic Acids Res. 39, W29–W37.
Finn, R. D., Attwood, T. K., Babbitt, P. C., Bateman, A., Bork, P., Bridge, A. J., et al. (2017). InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 45, D190–D199. doi: 10.1093/nar/gkw1107
Finn, R. D., Bateman, A., Clements, J., Coggill, P., Eberhardt, R. Y., Eddy, S. R., et al. (2014). Pfam: the protein families database. Nucleic Acids Res. 42, 222–230. doi: 10.1093/nar/gkt1223
Fuentes, J. L., Garbayo, I., Cuaresma, M., Montero, Z., González-Del-Valle, M., and Vílchez, C. (2016). Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 14:100. doi: 10.3390/md14050100
Gonçalves, A. L., Pires, J. C. M., and Simões, M. (2017). A review on the use of microalgal consortia for wastewater treatment. Algal Res. 24, 403–415. doi: 10.1016/j.algal.2016.11.008
Guldhe, A., Kumari, S., Ramanna, L., Ramsundar, P., Singh, P., Rawat, I., et al. (2017). Prospects, recent advancements and challenges of different wastewater streams for microalgal cultivation. J. Environ. Manage. 203, 299–315. doi: 10.1016/j.jenvman.2017.08.012
Guo, Z., and Tong, Y. W. (2014). The interactions between Chlorella vulgaris and algal symbiotic bacteria under photoautotrophic and photoheterotrophic conditions. J. Appl. Phycol. 26, 1483–1492. doi: 10.1007/s10811-013-0186-1
Han, X., Rusconi, N., Ali, P., Pagkatipunan, K., and Chen, F. (2017). Nutrients extracted from chicken manure accelerate growth of microalga Scenedesmus obliquus HTB1. Green Sustain. Chem. 07, 101–113. doi: 10.4236/gsc.2017.72009
Huang, A., Sun, L., Wu, S., Liu, C., Zhao, P., Xie, X., et al. (2017). Utilization of glucose and acetate by Chlorella and the effect of multiple factors on cell composition. J. Appl. Phycol. 29, 23–33. doi: 10.1007/s10811-016-0920-6
Huson, D. H., Beier, S., Flade, I., Górska, A., El-Hadidi, M., Mitra, S., et al. (2016). MEGAN community edition - interactive exploration and analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 12:e1004957. doi: 10.1371/journal.pcbi.1004957
Hyatt, D., Chen, G. L., LoCascio, P. F., Land, M. L., Larimer, F. W., and Hauser, L. J. (2010). Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 11:119. doi: 10.1186/1471-2105-11-119
Jankowska, E., Sahu, A. K., and Oleskowicz-Popiel, P. (2017). Biogas from microalgae: review on microalgae’s cultivation, harvesting and pretreatment for anaerobic digestion. Renew. Sustain. Energy Rev. 75, 692–709. doi: 10.1016/j.rser.2016.11.045
Jones, P., Binns, D., Chang, H. Y., Fraser, M., Li, W., McAnulla, C., et al. (2014). InterProScan 5: genome-scale protein function classification. Bioinformatics 30, 1236–1240. doi: 10.1093/bioinformatics/btu031
Ju, F., Guo, F., Ye, L., Xia, Y., and Zhang, T. (2014). Metagenomic analysis on seasonal microbial variations of activated sludge from a full-scale wastewater treatment plant over 4 years. Environ. Microbiol. Rep. 6, 80–89. doi: 10.1111/1758-2229.12110
Källqvist, T., and Svenson, A. (2003). Assessment of ammonia toxicity in tests with the microalga, Nephroselmis pyriformis, Chlorophyta. Water Res. 37, 477–484. doi: 10.1016/S0043-1354(02)00361-5
Kanehisa, M., and Guto, S. (2000). Kegg: kyoto encyclopedia of genes and genomes. Infect. Genet. Evol. 44, 313–317. doi: 10.1016/j.meegid.2016.07.022
Kang, D. D., Froula, J., Egan, R., and Wang, Z. (2015). MetaBAT, an efficient tool for accurately reconstructing single genomes from complex microbial communities. PeerJ 3:e1165. doi: 10.7717/peerj.1165
Klassen, V., Blifernez-Klassen, O., Hoekzema, Y., Mussgnug, J. H., and Kruse, O. (2015). A novel one-stage cultivation/fermentation strategy for improved biogas production with microalgal biomass. J. Biotechnol. 215, 44–51. doi: 10.1016/j.jbiotec.2015.05.008
Klassen, V., Blifernez-Klassen, O., Wibberg, D., Winkler, A., Kalinowski, J., Posten, C., et al. (2017). Highly efficient methane generation from untreated microalgae biomass. Biotechnol. Biofuels 10:186. doi: 10.1186/s13068-017-0871-4
Klassen, V., Blifernez-Klassen, O., Wobbe, L., Schlüter, A., Kruse, O., and Mussgnug, J. H. (2016). Efficiency and biotechnological aspects of biogas production from microalgal substrates. J. Biotechnol. 234, 7–26. doi: 10.1016/j.jbiotec.2016.07.015
Kouzuma, A., and Watanabe, K. (2015). Exploring the potential of algae/bacteria interactions. Curr. Opin. Biotechnol. 33, 125–129. doi: 10.1016/j.copbio.2015.02.007
Krohn-Molt, I., Alawi, M., Förstner, K. U., Wiegandt, A., Burkhardt, L., Indenbirken, D., et al. (2017). Insights into microalga and bacteria interactions of selected phycosphere biofilms using metagenomic, transcriptomic, and proteomic approaches. Front. Microbiol. 8:1941. doi: 10.3389/fmicb.2017.01941
Krustok, I., Truu, J., Odlare, M., Truu, M., Ligi, T., Tiirik, K., et al. (2015). Effect of lake water on algal biomass and microbial community structure in municipal wastewater-based lab-scale photobioreactors. Appl. Microbiol. Biotechnol. 99, 6537–6549. doi: 10.1007/s00253-015-6580-7
Krzywinski, M., Schein, J., Birol, I., Connors, J., Jones, S. J., Gascoyne, R., et al. (2009). Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–1645. doi: 10.1101/gr.092759.109
Lam, M. K., and Lee, K. T. (2012). Microalgae biofuels: a critical review of issues, problems and the way forward. Biotechnol. Adv. 30, 673–690. doi: 10.1016/j.biotechadv.2011.11.008
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Li, D., Liu, C. M., Luo, R., Sadakane, K., and Lam, T. W. (2015). MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 31, 1674–1676. doi: 10.1093/bioinformatics/btv033
Li, K., Liu, Q., Fang, F., Luo, R., Lu, Q., Zhou, W., et al. (2019). Microalgae-based wastewater treatment for nutrients recovery: a review. Bioresour. Technol. 291:121934. doi: 10.1016/j.biortech.2019.121934
Lian, J., Wijffels, R. H., Smidt, H., and Sipkema, D. (2018). The effect of the algal microbiome on industrial production of microalgae. Microb. Biotechnol. 11, 806–818. doi: 10.1111/1751-7915.13296
Liu, J., Wu, Y., Wu, C., Muylaert, K., Vyverman, W., Yu, H. Q., et al. (2017). Advanced nutrient removal from surface water by a consortium of attached microalgae and bacteria: a review. Bioresour. Technol. 241, 1127–1137. doi: 10.1016/j.biortech.2017.06.054
Liu, Q., Wang, J., Bai, Z., Ma, L., and Oenema, O. (2017). Global animal production and nitrogen and phosphorus flows. Soil Res. 55:451. doi: 10.1071/sr17031
Lu, J., Santo Domingo, J., and Shanks, O. C. (2007). Identification of chicken-specific fecal microbial sequences using a metagenomic approach. Water Res. 41, 3561–3574. doi: 10.1016/j.watres.2007.05.033
Mahdy, A., Mendez, L., Ballesteros, M., and González-Fernández, C. (2014). Algaculture integration in conventional wastewater treatment plants: anaerobic digestion comparison of primary and secondary sludge with microalgae biomass. Bioresour. Technol. 184, 236–244. doi: 10.1016/j.biortech.2014.09.145
Mao, C., Feng, Y., Wang, X., and Ren, G. (2015). Review on research achievements of biogas from anaerobic digestion. Renew. Sustain. Energy Rev. 45, 540–555. doi: 10.1016/j.rser.2015.02.032
Medipally, S. R., Yusoff, F. M., Banerjee, S., Shariff, M., Medipally, S. R., Yusoff, F. M., et al. (2015). Microalgae as sustainable renewable energy feedstock for biofuel production. Biomed Res. Int. 2015:519513. doi: 10.1155/2015/519513
Mende, D. R., Letunic, I., Huerta-Cepas, J., Li, S. S., Forslund, K., Sunagawa, S., et al. (2017). ProGenomes: a resource for consistent functional and taxonomic annotations of prokaryotic genomes. Nucleic Acids Res. 45, D529–D534. doi: 10.1093/nar/gkw989
Menzel, P., Ng, K. L., and Krogh, A. (2016). Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 7:11257. doi: 10.1038/ncomms11257
Molinuevo-Salces, B., Mahdy, A., Ballesteros, M., and González-Fernández, C. (2016). From piggery wastewater nutrients to biogas: microalgae biomass revalorization through anaerobic digestion. Renew. Energy 96, 1103–1110. doi: 10.1016/j.renene.2016.01.090
Morales-Sánchez, D., Martinez-Rodriguez, O. A., Kyndt, J., and Martinez, A. (2015). Heterotrophic growth of microalgae: metabolic aspects. World J. Microbiol. Biotechnol. 31, 1–9. doi: 10.1007/s11274-014-1773-2
Mouget, J. L., Dakhama, A., Lavoie, M. C., and de la Noüe, J. (1995). Algal growth enhancement by bacteria: is consumption of photosynthetic oxygen involved? FEMS Microbiol. Ecol. 18, 35–43. doi: 10.1016/0168-6496(95)00038-C
Mujtaba, G., and Lee, K. (2016). Advanced treatment of wastewater using symbiotic co-culture of microalgae and bacteria. Appl. Chem. Eng. 27, 1–9. doi: 10.14478/ace.2016.1002
Mujtaba, G., Rizwan, M., and Lee, K. (2015). Simultaneous removal of inorganic nutrients and organic carbon by symbiotic co-culture of Chlorella vulgaris and Pseudomonas putida. Biotechnol. Bioprocess. Eng. 20, 1114–1122. doi: 10.1007/s12257-015-0421-5
Mujtaba, G., Rizwan, M., and Lee, K. (2017). Removal of nutrients and COD from wastewater using symbiotic co-culture of bacterium Pseudomonas putida and immobilized microalga Chlorella vulgaris. J. Ind. Eng. Chem. 49, 145–151. doi: 10.1016/j.jiec.2017.01.021
Mussgnug, J. H., Klassen, V., Schlüter, A., and Kruse, O. (2010). Microalgae as substrates for fermentative biogas production in a combined biorefinery concept. J. Biotechnol. 150, 51–56. doi: 10.1016/j.jbiotec.2010.07.030
Nie, H., Jacobi, H. F., Strach, K., Xu, C., Zhou, H., and Liebetrau, J. (2015). Mono-fermentation of chicken manure: ammonia inhibition and recirculation of the digestate. Bioresour. Technol. 178, 238–246. doi: 10.1016/j.biortech.2014.09.029
Otondo, A., Kokabian, B., Stuart-Dahl, S., and Gude, V. G. (2018). Energetic evaluation of wastewater treatment using microalgae, Chlorella vulgaris. J. Environ. Chem. Eng. 6, 3213–3222. doi: 10.1016/j.jece.2018.04.064
Paquette, A. J., Sharp, C. E., Schnurr, P. J., Allen, D. G., Short, S. M., and Espie, G. S. (2020). Dynamic changes in community composition of Scenedesmus-seeded artificial, engineered microalgal biofilms. Algal Res. 46:101805. doi: 10.1016/j.algal.2020.101805
Parks, D. H., and Beiko, R. G. (2010). Identifying biologically relevant differences between metagenomic communities. Bioinformatics 26, 715–721. doi: 10.1093/bioinformatics/btq041
Pires, J. C. M., Alvim-Ferraz, M. C. M., Martins, F. G., and Simões, M. (2012). Carbon dioxide capture from flue gases using microalgae: engineering aspects and biorefinery concept. Renew. Sustain. Energy Rev. 16, 3043–3053. doi: 10.1016/j.rser.2012.02.055
Qi, W., Mei, S., Yuan, Y., Li, X., Tang, T., Zhao, Q., et al. (2018). Enhancing fermentation wastewater treatment by co-culture of microalgae with volatile fatty acid- and alcohol-degrading bacteria. Algal Res. 31, 31–39. doi: 10.1016/j.algal.2018.01.012
Quijano, G., Arcila, J. S., and Buitrón, G. (2017). Microalgal-bacterial aggregates: applications and perspectives for wastewater treatment. Biotechnol. Adv. 35, 772–781. doi: 10.1016/j.biotechadv.2017.07.003
Ramanan, R., Kim, B. H., Cho, D. H., Oh, H. M., and Kim, H. S. (2016). Algae-bacteria interactions: evolution, ecology and emerging applications. Biotechnol. Adv. 34, 14–29. doi: 10.1016/j.biotechadv.2015.12.003
Razzak, S. A., Hossain, M. M., Lucky, R. A., Bassi, A. S., and De Lasa, H. (2013). Integrated CO2 capture, wastewater treatment and biofuel production by microalgae culturing - a review. Renew. Sustain. Energy Rev. 27, 622–653. doi: 10.1016/j.rser.2013.05.063
Ren, H. Y., Zhu, J. N., Kong, F., Xing, D., Zhao, L., Ma, J., et al. (2019). Ultrasonic enhanced simultaneous algal lipid production and nutrients removal from non-sterile domestic wastewater. Energy Convers. Manag. 180, 680–688. doi: 10.1016/j.enconman.2018.11.028
Rinke, C., Schwientek, P., Sczyrba, A., Ivanova, N. N., Anderson, I. J., Cheng, J. F., et al. (2013). Insights into the phylogeny and coding potential of microbial dark matter. Nature 499, 431–437. doi: 10.1038/nature12352
Rizwan, M., Mujtaba, G., Memon, S. A., Lee, K., and Rashid, N. (2018). Exploring the potential of microalgae for new biotechnology applications and beyond: a review. Renew. Sustain. Energy Rev. 92, 394–404. doi: 10.1016/j.rser.2018.04.034
Sakar, S., Yetilmezsoy, K., and Kocak, E. (2009). Anaerobic digestion technology in poultry and livestock waste treatment - a literature review. Waste Manag. Res. 27, 3–18. doi: 10.1177/0734242X07079060
Santos, C. A., and Reis, A. (2014). Microalgal symbiosis in biotechnology. Appl. Microbiol. Biotechnol. 98, 5839–5846. doi: 10.1007/s00253-014-5764-x
Sayre, R. (2010). Microalgae: the potential for carbon capture. Bioscience 60, 722–727. doi: 10.1525/bio.2010.60.9.9
Schmieder, R., and Edwards, R. (2011). Quality control and preprocessing of metagenomic datasets. Bioinformatics 27, 863–864. doi: 10.1093/bioinformatics/btr026
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Seger, M., Unc, A., Starkenburg, S. R., Holguin, F. O., and Lammers, P. J. (2019). Nutrient-driven algal-bacterial dynamics in semi-continuous, pilot-scale photobioreactor cultivation of Nannochloropsis salina CCMP1776 with municipal wastewater nutrients. Algal Res. 39:101457. doi: 10.1016/j.algal.2019.101457
Seymour, J. R., Amin, S. A., Raina, J. B., and Stocker, R. (2017). Zooming in on the phycosphere: the ecological interface for phytoplankton-bacteria relationships. Nat. Microbiol. 2:17065. doi: 10.1038/nmicrobiol.2017.65
Shetty, P., Boboescu, I. Z., Pap, B., Wirth, R., and Kovács, K. L. (2019). Exploitation of algal-bacterial consortia in combined biohydrogen generation and wastewater treatment. Front. Energy Res. 7:52. doi: 10.3389/fenrg.2019.00052
Simão, F. A., Waterhouse, R. M., Ioannidis, P., Kriventseva, E. V., and Zdobnov, E. M. (2015). BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 31, 3210–3212. doi: 10.1093/bioinformatics/btv351
Skorupskaite, V., Makareviciene, V., and Levisauskas, D. (2015). Optimization of mixotrophic cultivation of microalgae Chlorella sp. for biofuel production using response surface methodology. Algal Res. 7, 45–50. doi: 10.1016/j.algal.2014.12.001
Smith, A. G., Croft, M. T., Moulin, M., and Webb, M. E. (2007). Plants need their vitamins too. Curr. Opin. Plant Biol. 10, 266–275. doi: 10.1016/j.pbi.2007.04.009
Sun, C., Cao, W., Banks, C. J., Heaven, S., and Liu, R. (2016). Biogas production from undiluted chicken manure and maize silage: a study of ammonia inhibition in high solids anaerobic digestion. Bioresour. Technol. 218, 1215–1223. doi: 10.1016/j.biortech.2016.07.082
Sydney, E. B., Sydney, A. C. N., de Carvalho, J. C., and Soccol, C. R. (2019). “Potential carbon fixation of industrially important microalgae,” in Biofuels From Algae, eds A. Pandey, J.-S. Chang, C. R. Soccol, D.-J. Lee, and Y. Chisti, (Amsterdam: Elsevier), 67–88.
Thomas, D. M., Mechery, J., and Paulose, S. V. (2016). Carbon dioxide capture strategies from flue gas using microalgae: a review. Environ. Sci. Pollut. Res. 23, 16926–16940. doi: 10.1007/s11356-016-7158-3
Toyama, T., Kasuya, M., Hanaoka, T., Kobayashi, N., Tanaka, Y., Inoue, D., et al. (2018). Growth promotion of three microalgae, Chlamydomonas reinhardtii, Chlorella vulgaris and Euglena gracilis, by in situ indigenous bacteria in wastewater effluent. Biotechnol. Biofuels 11:176. doi: 10.1186/s13068-018-1174-0
Uggetti, E., Passos, F., Solé, M., Garfí, M., and Ferrer, I. (2016). Recent achievements in the production of biogas from microalgae. Waste Biomass Valoriza. 8, 129–139. doi: 10.1007/s12649-016-9604-3
Uggetti, E., Sialve, B., Latrille, E., and Steyer, J. P. (2014). Anaerobic digestate as substrate for microalgae culture: the role of ammonium concentration on the microalgae productivity. Bioresour. Technol. 152, 437–443. doi: 10.1016/j.biortech.2013.11.036
Ullah Khan, I., Hafiz Dzarfan, Othman, M., Hashim, H., Matsuura, T., Ismail, A. F., et al. (2017). Biogas as a renewable energy fuel – a review of biogas upgrading, utilisation and storage. Energy Convers. Manag. 150, 277–294. doi: 10.1016/j.enconman.2017.08.035
Vereins Deutscher Ingenieure 4630, (2006). Fermentation of Organic Materials – Characterisation of the Substrate, Sampling, Collection of Material Data, Fermentation Tests, VDI Guideline 4630. Düsseldorf: Verein Deutscher Ingenieure.
Vo Hoang Nhat, P., Ngo, H. H., Guo, W. S., Chang, S. W., Nguyen, D. D., Nguyen, P. D., et al. (2018). Can algae-based technologies be an affordable green process for biofuel production and wastewater remediation? Bioresour. Technol. 256, 491–501. doi: 10.1016/j.biortech.2018.02.031
Ward, A. J., Lewis, D. M., and Green, F. B. (2014). Anaerobic digestion of algae biomass: a review. Algal Res. 5, 204–214. doi: 10.1016/j.algal.2014.02.001
Wirth, R., Böjti, T., Lakatos, G., Maróti, G., Bagi, Z., Rákhely, G., et al. (2019). Characterization of core microbiomes and functional profiles of mesophilic anaerobic digesters fed with Chlorella vulgaris green microalgae and maize silage. Front. Energy Res. 7:111. doi: 10.3389/fenrg.2019.00111
Wirth, R., Lakatos, G., Böjti, T., Maróti, G., Bagi, Z., Kis, M., et al. (2015a). Metagenome changes in the mesophilic biogas-producing community during fermentation of the green alga Scenedesmus obliquus. J. Biotechnol. 215, 52–61. doi: 10.1016/j.jbiotec.2015.06.396
Wirth, R., Lakatos, G., Maróti, G., Bagi, Z., Minárovics, J., Nagy, K., et al. (2015b). Exploitation of algal-bacterial associations in a two-stage biohydrogen and biogas generation process. Biotechnol. Biofuels 8:59. doi: 10.1186/s13068-015-0243-x
Wirth, R., Lakatos, G., Böjti, T., Maróti, G., Bagi, Z., Rákhely, G., et al. (2018). Anaerobic gaseous biofuel production using microalgal biomass - a review. Anaerobe 52, 1–8. doi: 10.1016/j.anaerobe.2018.05.008
Wu, Y. W., Simmons, B. A., and Singer, S. W. (2015). MaxBin 2.0: an automated binning algorithm to recover genomes from multiple metagenomic datasets. Bioinformatics 32, 605–607. doi: 10.1093/bioinformatics/btv638
Xia, A., and Murphy, J. D. (2016). Microalgal cultivation in treating liquid digestate from biogas systems. Trends Biotechnol. 34, 264–275. doi: 10.1016/j.tibtech.2015.12.010
Yen, H. W., Hu, I. C., Chen, C. Y., Ho, S. H., Lee, D. J., and Chang, J. S. (2013). Microalgae-based biorefinery - from biofuels to natural products. Bioresour. Technol. 135, 166–174. doi: 10.1016/j.biortech.2012.10.099
Zhu, L., Yan, C., and Li, Z. (2016). Microalgal cultivation with biogas slurry for biofuel production. Bioresour. Technol. 220, 629–636. doi: 10.1016/j.biortech.2016.08.111
Zhu, S., Feng, S., Xu, Z., Qin, L., Shang, C., Feng, P., et al. (2019). Cultivation of Chlorella vulgaris on unsterilized dairy-derived liquid digestate for simultaneous biofuels feedstock production and pollutant removal. Bioresour. Technol. 285:121353. doi: 10.1016/j.biortech.2019.121353
Zuñiga, C., Li, C.-T., Huelsman, T., Levering, J., Zielinski, D. C., McConnell, B. O., et al. (2016). Genome-Scale metabolic model for the green alga Chlorella vulgaris UTEX 395 accurately predicts phenotypes under autotrophic, heterotrophic, and mixotrophic growth conditions. Plant Physiol. 172, 589–602. doi: 10.1104/pp.16.00593
Keywords: wastewater, green algae, phycosphere, algal-bacterial interactions, metagenomics
Citation: Wirth R, Pap B, Böjti T, Shetty P, Lakatos G, Bagi Z, Kovács KL and Maróti G (2020) Chlorella vulgaris and Its Phycosphere in Wastewater: Microalgae-Bacteria Interactions During Nutrient Removal. Front. Bioeng. Biotechnol. 8:557572. doi: 10.3389/fbioe.2020.557572
Received: 30 April 2020; Accepted: 28 August 2020;
Published: 22 September 2020.
Edited by:
Raul Muñoz, University of Valladolid, SpainReviewed by:
Peter J. Lammers, Arizona State University, United StatesCopyright © 2020 Wirth, Pap, Böjti, Shetty, Lakatos, Bagi, Kovács and Maróti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gergely Maróti, bWFyb3RpLmdlcmdlbHlAYnJjLmh1; bWFyb3RpLmdlcmdlbHlAYnJjLm10YS5odQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.