
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Behav. Neurosci. , 07 July 2017
Sec. Emotion Regulation and Processing
Volume 11 - 2017 | https://doi.org/10.3389/fnbeh.2017.00117
This article is part of the Research Topic The Impact of Stress on Cognition and Motivation View all 11 articles
 Laura A. León1,2,3
Laura A. León1,2,3 Vitor Castro-Gomes4Santiago Zárate-Guerrero5
Vitor Castro-Gomes4Santiago Zárate-Guerrero5 Karen Corredor5†Antonio P. Mello Cruz6
Karen Corredor5†Antonio P. Mello Cruz6 Marcus L. Brandão1
Marcus L. Brandão1 Fernando P. Cardenas5†
Fernando P. Cardenas5† J. Landeira-Fernandez2*
J. Landeira-Fernandez2*The role of serotonin (5-hydroxytryptamine [5-HT]) and 5-HT2A receptors in anxiety has been extensively studied, mostly without considering individual differences in trait anxiety. Our laboratory developed two lines of animals that are bred for high and low freezing responses to contextual cues that are previously associated with footshock (Carioca High-conditioned Freezing [CHF] and Carioca Low-conditioned Freezing [CLF]). The present study investigated whether ketanserin, a preferential 5-HT2A receptor blocker, exerts distinct anxiety-like profiles in these two lines of animals. In the first experiment, the animals received a systemic injection of ketanserin and were exposed to the elevated plus maze (EPM). In the second experiment, these two lines of animals received microinjections of ketanserin in the infralimbic (IL) and prelimbic (PL) cortices and were exposed to either the EPM or a contextual fear conditioning paradigm. The two rat lines exhibited bidirectional effects on anxiety-like behavior in the EPM and opposite responses to ketanserin. Both systemic and intra-IL cortex injections of ketanserin exerted anxiolytic-like effects in CHF rats but anxiogenic-like effects in CLF rats. Microinjections of ketanserin in the PL cortex also exerted anxiolytic-like effects in CHF rats but had no effect in CLF rats. These results suggest that the behavioral effects of 5-HT2A receptor antagonism might depend on genetic variability associated with baseline reactions to threatening situations and 5-HT2A receptor expression in the IL and PL cortices.
Highlights
- CHF and CLF rats are two bidirectional lines that are based on contextual fear conditioning.
- CHF rats have a more “anxious” phenotype than CLF rats in the EPM.
- The 5-HT2A receptor antagonist ketanserin had opposite behavioral effects in CHF and CLF rats.
- Systemic and IL injections either decreased (CHF) or increased (CLF) anxiety-like behavior.
- PL injections either decreased (CHF) anxiety-like behavior or had no effect (CLF).
Several studies indicate that contextual fear conditioning represents one of the simplest animal models of investigating anticipatory anxiety (Brandão et al., 2008). It involves placing an animal (e.g., a rat) in a novel environment and delivering a brief unsignaled footshock several minutes later. The next day, the animal freezes when it is returned to the same chamber in the absence of footshock (Landeira-Fernandez, 1996). Conditioned freezing is a direct function of shock intensity (Sigmundi et al., 1983) and depends on the association between the contextual cues associated with the experimental chamber and footshock (Landeira-Fernandez et al., 2006). Classic anxiolytic benzodiazepines, such as midazolam and diazepam (Fanselow and Helmstetter, 1988), and non-benzodiazepine anxiolytics, such as the serotonin (5-hydroxytryptamine [5-HT])-1A receptor agonist ipsapirone (Inoue et al., 1996) and 5-HT reuptake inhibitors citalopram and fluvoxamine (Hashimoto et al., 1996), reduced conditioned freezing. Anxiogenic substances, such as the benzodiazepine inverse agonist dimethoxy-β-carboline, induced freezing behavior similarly to fear conditioning (Fanselow et al., 1991).
Two lines of Wistar rats, termed Carioca High-conditioned Freezing and Low-conditioned Freezing (CHF and CLF, respectively), were selectively bred for high and low levels of freezing in response to contextual cues that were previously associated with footshock (Castro-Gomes and Landeira-Fernandez, 2008). The results of our ongoing breeding program have shown a clear divergence of the conditioned freezing phenotype after only three generations (Castro-Gomes and Landeira-Fernandez, 2008). The presence of different levels of anxiety-like behavior, that are characteristic of each line, can be assessed using several behavioral tests, including the elevated plus maze (EPM), the social interaction test and defensive responses that are induced by electrical stimulation of the dorsal periaqueductal gray (Dias et al., 2009; Galvão et al., 2010; Castro-Gomes et al., 2011, 2014; Salviano et al., 2014). These two lines represent an important tool for investigating the anxiogenic/anxiolytic pharmacological profiles of various compounds (Castro-Gomes et al., 2013).
5-HT is an indoleamine that is intimately connected to the neurocircuitry that underlies anxiety (for review see Millan, 2003; Graeff, 2004). It exerts its behavioral and physiological effects by acting at different receptor subtypes that are distributed into seven G-protein-coupled receptor families (Hoyer et al., 2002; Hannon and Hoyer, 2008). The 5-HT2 receptor family (5-HT2A, 5-HT2B and 5-HT2C) has been the focus of much research interest because of its critical role in modulating anxiety-like behavior in animals and humans (Naughton et al., 2000; Graeff, 2002; Wood, 2003; Gordon and Hen, 2004). The predominant effect of 5-HT in this metabotropic receptor family (especially the 5-HT2A subtype) is excitatory and appears to mediate depolarizing effects (Davie et al., 1988; Eison and Mullins, 1996; Hasuo et al., 2002), but it may also have inhibitory activity or even interact with a number of other inhibitory interneurons (Avesar and Gulledge, 2012; Halberstadt, 2015; Wang et al., 2016).
Paradoxically, the effects of 5-HT2-acting drugs on anxiety have been highly inconsistent in animals and humans. For example, systemic administration of 5-HT2A,2C agonists exerts both anxiogenic-like (Charney et al., 1987; Lowy and Meltzer, 1988; Bastani et al., 1990; Rodgers et al., 1992; Gibson et al., 1994; Setem et al., 1999; Jones et al., 2002; Bull et al., 2003; Durand et al., 2003) and anxiolytic-like (Ripoll et al., 2006; Hughes et al., 2012) effects. Similarly, 5-HT2A,2C antagonists have been reported to exert anxiolytic-like effects (Critchley and Handley, 1987; Motta et al., 1992; Kennett et al., 1994; Nic Dhonnchadha et al., 2003), anxiogenic-like effects (Pellow et al., 1987), or no effects (Chaouloff et al., 1997; Setem et al., 1999). Similar inconsistencies have also been reported following direct infusions of 5-HT2 agonists and antagonists in anxiety-related postsynaptic brain sites (for review see Menard and Treit, 1999; Graeff, 2002, 2004). For example, 5-HT2A/2C agonist administration in the amygdala (Campbell and Merchant, 2003; de Mello Cruz et al., 2005) and ventral hippocampus (Alves et al., 2004) has been shown to be anxiogenic, whereas microinjections in the dorsal periaqueductal gray have been reported to be anxiolytic (Graeff et al., 1993).
Different experimental procedures, brain sites and selectivity for 5-HT2 receptors might contribute to these discrepancies. Moreover, the role of the 5-HT2 receptor family in anxiety might depend on genetic variables that are associated with different trait levels of defensive reactions. One of the purposes of the present study was to investigate the effects of systemic injections of a 5-HT2A receptor antagonist in CHF and CLF animals in the EPM. Ketanserin was chosen in this study because of our 25 years’ experience with the use of this drug as a pharmacological tool for the study of the neurobiology of anxiety and fear (Motta et al., 1992; de Luca et al., 2003; Oliveira et al., 2007; Almada et al., 2009). Ketanserin is a 5-HT2A/2C receptor antagonist that has higher affinity for 5-HT2A receptors than 5-HT2C receptors (Kristiansen and Dahl, 1996; López-Giménez et al., 1997; Knight et al., 2004).
EPM is based on rodents’ innate fear of open spaces (Treit et al., 1993). This model has been, behaviorally, physiologically and pharmacologically, validated as an animal model of anxiety in rats (Pellow et al., 1985; Pellow and File, 1986; Reibaud and Böhme, 1993). Factor analyses have also indicated that this test reliably dissociates the anxiety-related effects (open arm entries) from locomotor effects (closed arm entries) of several anxiolytic and anxiogenic agents (File, 1992; Cruz et al., 1994).
The role of 5-HT2A receptors in anxiety might also depend on serotonergic activity in different brain regions. The ventromedial prefrontal cortex (vmPFC) is composed of the infralimbic (IL) and prelimbic (PL) subregions. Serotonergic neurons in both the dorsal and medial raphe nuclei send robust projections to the vmPFC (Azmitia and Segal, 1978; Steinbusch, 1981; Blue et al., 1988). Moreover, 5-HT2 receptors, but mainly the 5-HT2A receptor subtype, are widely and densely distributed in both the PL and IL (Pazos et al., 1985; Pompeiano et al., 1994; Santana et al., 2004).
Behavioral results that have been generated with different animal models of anxiety indicate that the IL and PL play distinct and complex roles in conditioned and innate defensive reactions. The IL appears to inhibit the expression of anxiety-like behavior, whereas the PL facilitates its expression through descending projection to the basolateral complex of the amygdala (Likhtik et al., 2014). For example, stimulation of the IL reduced the expression of auditory fear conditioning (Vidal-Gonzalez et al., 2006) but produced anxiety-like behavior in the EPM (Bi et al., 2013). Moreover, inhibition of the IL impaired the acquisition, consolidation and expression of conditioned fear extinction (Sierra-Mercado et al., 2006, 2011; Corcoran and Quirk, 2007). Inactivation of the IL had an anxiolytic-like effect in the EPM (Bi et al., 2013). Stimulation of the PL increased the occurrence of conditioned fear (Vidal-Gonzalez et al., 2006) and anxiety-like behavior in the EPM (Wang et al., 2015). Inactivation of the PL reduced the expression of conditioned fear (Corcoran and Quirk, 2007) and anxiety-like behavior in the EPM (Wang et al., 2015), but PL blockade did not have any effects on innate fear reactions (Corcoran and Quirk, 2007). Neither stimulation nor inactivation of the PL caused any changes in the extinction of contextual (Laurent and Westbrook, 2009) or auditory (Sierra-Mercado et al., 2011) fear conditioning.
These results suggest that the IL and PL play opposite roles in fear conditioning that might depend on nature (i.e., innate or learned) of the threatening stimulus (Lisboa et al., 2010). Therefore, considering the nature of anxiety as well as the presence of 5-HT2A receptors in the PL and IL cortices, the present study also compared in two other experiments the effects of microinjections of the preferential 5-HT2A receptor antagonist ketanserin in the IL and PL in CHF and CLF rats in both innate (EPM) and learned (contextual fear conditioning) models of anxiety.
The animals that were used in the present study were selectively bred for high (CHF) and low (CLF) contextual fear conditioning according to procedures described in our previous work (Castro-Gomes and Landeira-Fernandez, 2008). Briefly, albino Wistar rats were selectively bred for differences in defensive freezing behavior in a contextual fear-conditioning paradigm. This protocol involved one acquisition session and one test session. During acquisition, each animal was placed in the observation chamber for 8 min. At the end of this period, three unsignaled 0.5 mA, 1 s electric footshocks were delivered with an intershock interval of 20 s. Three minutes after the last footshock, the animal was returned to its home cage. The test session was conducted approximately 24 h after training. This test consisted of placing the animal for 8 min in the same chamber where the three footshocks were delivered the previous day. No footshock or other stimulation occurred during this period. All of the animals were phenotyped before beginning each experiment. The phenotyping procedure consisted of evaluating the amount of conditioned freezing during the test session (Castro-Gomes and Landeira-Fernandez, 2008). The experiments began 2 months after the phenotyping procedure. The first experiment investigated the effects of systemic intraperitoneal ketanserin administration in CHF and CLF animals of the 10th generation. The second experiment investigated the effects of ketanserin injections in the IL in CHF and CLF animals of the 15th generation. The third experiment investigated the effects of ketanserin injections in the PL in CHF and CLF animals of the 20th generation. Male rats from both the CHF and CLF lines were 4–6 months old at the beginning of each of the three experiments in the present study.
All the animals were born and maintained in the colony room of the PUC-Rio Psychology Department at controlled room temperature (24 ± 1°C) and a 12 h/12 h light/dark cycle (lights on 7:00 AM–7:00 PM). They were housed in groups of 3–5 according to their respective lines in polycarbonate cages (18 cm × 31 cm × 38 cm) with food and water available ad libitum. All of the behavioral experiments were conducted during the light phase of the light/dark cycle. The animals were handled once daily for 2 min for 5 days before the beginning of each experiment. The experimental procedures were performed in accordance with the guidelines for experimental animal research that were established by the Brazilian Society of Neuroscience and Behavior (SBNeC) and National Institutes of Health Guide for the Care and Use of Laboratory Animals. Animal handling and the methods of sacrifice were reviewed and approved by the Committee for Animal Care and Use of PUC-Rio (protocol no. 20/2009).
Contextual fear conditioning was performed in four observational chambers (25 cm × 20 cm × 20 cm) inside sound-attenuating boxes. A video camera was mounted on the back of each observational chamber so that the animal’s behavior could be observed on a monitor in an adjacent room. Background noise (78 dB) was supplied by a white-noise generator. The chamber had a grid floor (15 stainless steel rods spaced 1.5 cm apart) connected to a shock generator (0.5 mA, 1 s duration) and scrambler (AVS, SCR04; São Paulo, Brazil). An interface with four channels (Insight, Ribeirão Preto, Brazil) connected the shock generator to a computer, which allowed the experimenter to apply an electric footshock. A digital multimeter was used to calibrate the shock intensities before each experiment. A 5% ammonium hydroxide solution was used to clean the chamber before and after each test.
The EPM was elevated 50 cm above the floor and had two open arms (50 cm × 10 cm, with 1 cm high edges) and two closed arms (50 cm × 10 cm, with 40 cm high walls) arranged so that arms of the same type were opposite each other. All of the arms were connected by an open central area (10 cm × 10 cm). The tests were performed in a room that was illuminated by a 100-W light bulb that was suspended 1.75 m above the central part of the maze. A 20% alcohol solution was used to clean the maze between trials.
Ketanserin tartrate 97% (Sigma-Aldrich, St. Louis, MO, USA) was dissolved in 2% dimethylsulfoxide (DMSO) vehicle solution. Both the systemic (0.5 mg/kg) and central (5 nmol/μl) doses were selected according to our previous work (Motta et al., 1992; de Luca et al., 2003; Oliveira et al., 2007). Rats in the control group received 2% DMSO dissolved in saline (vehicle).
The rats in Experiments 2 and 3 were implanted with bilateral cannulae in the IL or PL. The rats were anesthetized with an intraperitoneal injection of 75 mg/kg ketamine hydrochloride (ketamine 50®, Holliday-Scott SA) and 10 mg/kg xylazine (Seton®, Calier) and mounted in a stereotaxic instrument with the incisor bar set 3.3 mm below the interaural line. Each rat was bilaterally implanted with stainless steel guide cannulae (outer diameter, 0.7 mm) aimed 0.5 mm above the target area. With bregma as the reference for each plane (Paxinos and Watson, 1986), the coordinates for IL cannula implantation were the following: anterior/posterior, 3.2 mm; lateral, ± 0.4 mm; depth, 4.2 mm. The coordinates for PL cannula implantation were the following: anterior/posterior, 3.2 mm; lateral, ± 0.5 mm; depth, 2.4 mm. The guide cannulae were anchored to the skull by dental acrylic and one stainless steel screw. After implantation, the guide cannulae were sealed with a stainless steel wire to prevent blockage. Immediately after cannula implantation, the animals received 0.1 ml of a combination of antibiotics (benzylpenicillin benzathine, 600,000 UI; benzylpenicillin procaine, 300,000 UI; potassium benzylpenicillin, 300,000 UI; dihydrostreptomycin sulfate, 250 mg; streptomycin sulfate, 250 mg, intramuscular) and an analgesic (flunixin, 50 mg/kg, subcutaneous) to prevent infections and decrease post-surgical pain.
At the end of the experiments, the animals were bilaterally injected with 2 μl of 1% Evan’s Blue dye in the IL and PL to verify the accuracy of the injections. The animals were then sacrificed with an overdose of urethane (1.25 g/kg, intraperitoneal; Sigma-Aldrich, St. Louis, MO, USA) and intracardially perfused with 0.9% saline and 4% formalin through the left ventricle. The brains were removed and stored in 10% formalin for at least 2 weeks and then sectioned using a cryostat (CM-1900, Leica, Germany) into 60 μm sections. The injection sites were identified using the rat brain atlas of Paxinos and Watson (1986). Only rats with bilateral cannula sites in the IL or PL were considered for the statistical analysis.
The CHF and CLF animals were randomly assigned to the ketanserin (0.5 mg/kg) or vehicle group. Thirty minutes after the intraperitoneal injection, each animal was placed in the center of the EPM facing one of the closed arms. The experimental session lasted 5 min. A highly trained observer who remained blind to the treatment conditions recorded the number of entries into and time spent on the open and closed arms with the aid of computer software. The percentage of open arm entries (100 × open arm entries total arm entries) and percentage of time spent on the open arms (100 × time open/[time open + time closed]) were calculated for each animal as indices of anxiety-like behavior. Based on factor analysis of the rats’ behavior in the EPM (File, 1992; Cruz et al., 1994), the absolute number of closed arm entries was interpreted as a reliable index of general locomotor activity.
One week after surgery, CHF and CLF animals were randomly assigned to the ketanserin (5 nmol/μl) or vehicle group. Experiment 2 investigated the behavioral effects of ketanserin microinjections in the IL. Experiment 3 investigated the behavioral effects of ketanserin microinjections in the PL. Bilateral infusions were performed through an internal cannula (outer diameter, 0.3 mm) that extended 0.5 mm beyond the guide cannula tip. The cannula was attached to a 10 μl Hamilton syringe via polyethylene-10 tubing. Confirmation of a successful infusion was achieved by monitoring the movement of a small air bubble inside the polyethylene-10 tubing. A volume of 0.5 μl/side was delivered over approximately 2 min through the 10 μl Hamilton syringe, driven by a Harvard syringe pump. Following the infusion, the internal cannula was left in place for an additional 2 min to minimize reflux up the cannula shaft. Previous results from our laboratory demonstrated the effectiveness of these parameters when ketanserin was injected locally into brain structures (Motta et al., 1992; de Luca et al., 2003; Oliveira et al., 2007; Almada et al., 2009). Other results suggest that the drug may diffuse approximately 0.5–1.0 mm from the tip of the infusion cannula (Allen et al., 2008).
Five minutes after the injection, each animal was tested in the EPM. At the end of the EPM test, each animal was returned to the same chamber where contextual fear conditioning occurred to test the duration of freezing as a measure of the conditioned fear response. The animal stayed there for 8 min with no footshock or other stimulation. A time-sampling procedure was used to assess fear conditioning in response to contextual cues. Every 2 s, a well-trained observer recorded episodes of freezing, which were defined as the total absence of movement of the body or vibrissa, with the exception of movement required for respiration.
The results were statistically analyzed by two-way analysis of variance (ANOVA) to detect overall differences. One independent factor was treatment (ketanserin and vehicle), and the other independent factor was rat line (CLF and CHF). Tukey’s Honestly Significant Difference test was used for post hoc pairwise comparisons between groups. In all cases, values of p < 0.05 were considered statistically significant. All analysis were performed with SPSS software.
The number of animals in each experimental condition in this experiment was the following: CLF animals injected with vehicle (n = 8), CLF animals injected with ketanserin (n = 10), CHF animals injected with vehicle (n = 8), and CHF animals injected with ketanserin (n = 8). Figure 1 shows the behavioral effects in CHF and CLF that received systemic injections of ketanserin or vehicle in the EPM. The two-way ANOVA of the percentage of open arm entries (Figure 1A) indicated a main effect of rat line (F(1,30) = 4.71, p < 0.05) and a rat line × treatment interaction (F(1,30) = 21.66, p < 0.01). No main effect of treatment was found (F(1,30) = 0.01, p > 0.90). The post hoc comparisons revealed that systemic ketanserin administration significantly increased the percentage of open arm entries in CHF animals compared with the vehicle but significantly decreased this measure in CLF animals (both p < 0.05).
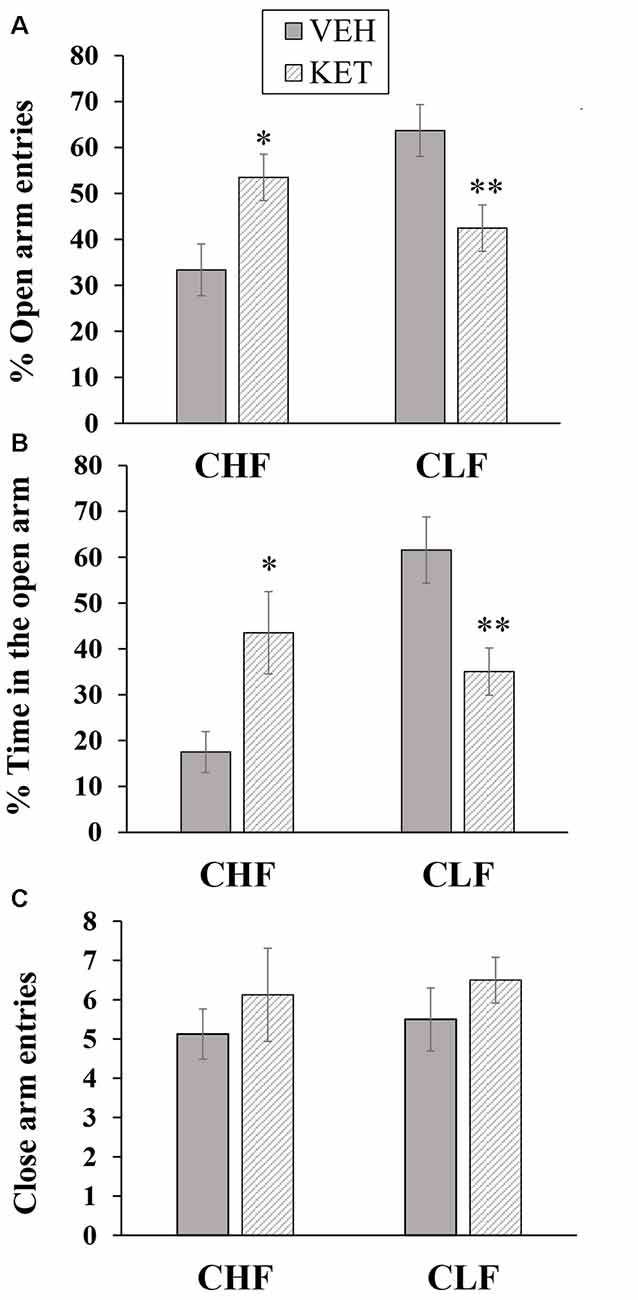
Figure 1. Mean + SEM percentage of open arm entries (A), percent time spent on the open arms (B) and closed arm entries (C) in the elevated plus maze (EPM) in Carioca High-conditioned Freezing (CHF) and Carioca Low-conditioned Freezing (CLF) animals that received systemic ketanserin (KET) or vehicle (VEH) injections. *p < 0.05 KET vs. VEH among CHF animals; **p < 0.05 KET vs. VEH among CLF animals.
A similar pattern was observed for the percentage of time spent in the open arms (Figure 1B). The two-way ANOVA revealed a main effect of rat line (F(1,30) = 7.31, p < 0.05) and a rat line × treatment interaction (F(1,30) = 15.91, p < 0.01). No main effect of treatment was found (F(1,30) = 0.01, p > 0.90). The post hoc comparisons revealed that systemic ketanserin administration significantly increased the percentage of time spent on the open arms in CHF animals compared with vehicle but significantly decreased this measure in CLF animals (both p < 0.05).
Figure 1C shows the effects of ketanserin and vehicle on the absolute number of closed arm entries in CHF and CLF animals. No differences were observed among groups. The two-way ANOVA revealed no main effect of rat line or treatment and no interaction between factors (all p > 0.05).
The histological analysis of the cannula placements confirmed that the infusions were made in the IL region in all animals that were included in the statistical analysis. Four of forty rats in the experiment were excluded because their cannula missed the IL. Figure 2 shows the bilateral microinjection sites in the IL. The final sample size for each group was the following: CLF animals injected with vehicle (n = 9), CLF animals injected with ketanserin (n = 11), CHF animals injected with vehicle (n = 7), and CHF animals injected with ketanserin (n = 9).
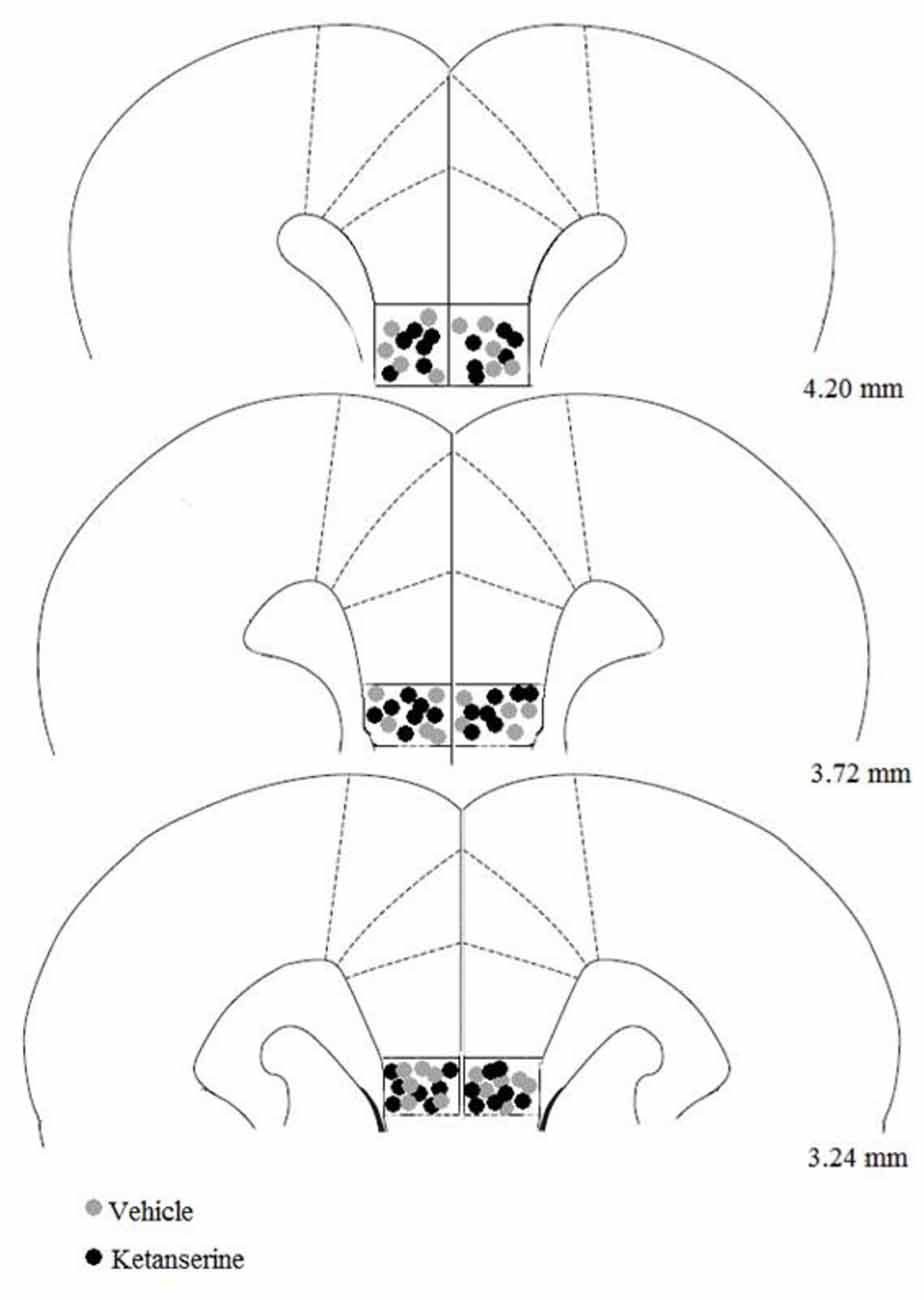
Figure 2. Site of microinjection tips in the infralimbic (IL) cortex. Gray square indicates site of vehicle injection. Black circle indicates site of ketanserin injections. Plates are taken from Paxinos and Watson (1986) and the numbers on the right side of each plate indicate the distance (in millimeters) from bregma.
Figure 3 shows the mean ± SEM percentage of open arm entries, percent time spent on the open arms and closed arm entries in the EPM in CHF and CLF animals that received ketanserin or vehicle microinjections in the IL.
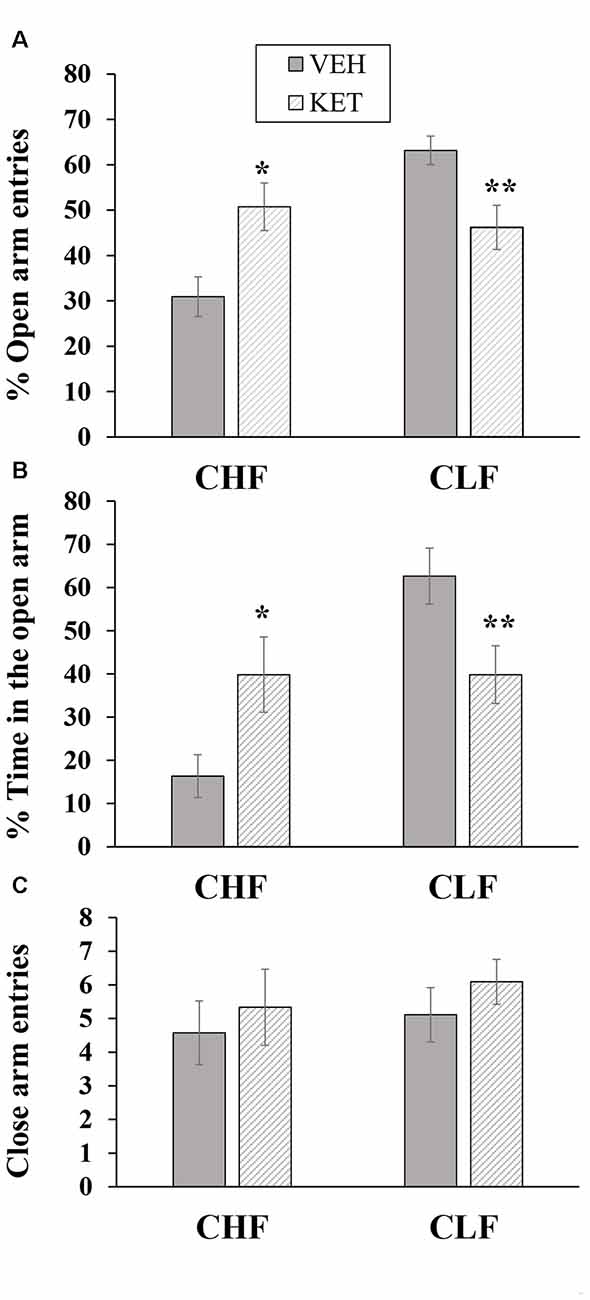
Figure 3. Mean + SEM percentage of open arm entries (A), percent time spent on the open arms (B) and closed arm entries (C) in the EPM in CHF and CLF animals that received ketanserin (KET) or vehicle (VEH) microinjections in the IL cortex. *p < 0.05 KET vs. VEH among CHF animals; **p < 0.05 KET vs. VEH among CLF animals.
The two-way ANOVA of the percentage of open arm entries (Figure 3A) indicated a main effect of rat line (F(1,32) = 8.81, p < 0.05) and a rat line × treatment interaction (F(1,32) = 15.61, p < 0.01). No main effect of treatment was found (F(1,32) = 0.09, p > 0.76). The post hoc comparisons revealed that ketanserin microinjections in the IL significantly increased the percentage of open arm entries in CHF animals compared with the vehicle but significantly decreased this measure in CLF animals (both p < 0.05).
The two-way ANOVA of the percentage of time spent on the open arms (Figure 3B) revealed a main effect of rat line (F(1,32) = 10.41, p < 0.05) and a rat line × treatment interaction (F(1,32) = 10.41, p < 0.05). No main effect of treatment was found (F(1,32) = 0.01, p > 0.90). The post hoc comparisons revealed that ketanserin significantly increased the percentage of time spent on the open arms in CHF animals compared with the vehicle but significantly decreased this measure in CLF animals (both p < 0.05).
The two-way ANOVA of the absolute number of closed arm entries in CHF and CLF animals that received microinjections of ketanserin or vehicle (Figure 3C) revealed no main effect of rat line or treatment and no interaction between these factors (all p > 0.05).
Figure 4 shows the mean ± SEM percentage of time spent freezing in CHF and CLF animals that received microinjections of ketanserin or vehicle in the IL. The two-way ANOVA indicated main effects of rat line (F(1,32) = 69.12, p < 0.01), treatment (F(1,32) = 70.61, p < 0.01) and a significant rat line × treatment interaction (F(1,32) = 66.15, p < 0.01). The post hoc comparisons indicated that ketanserin microinjections in the IL significantly decreased the percentage of time spent freezing in CHF animals compared with the vehicle but significantly increased this measure in CLF animals (both p < 0.05).
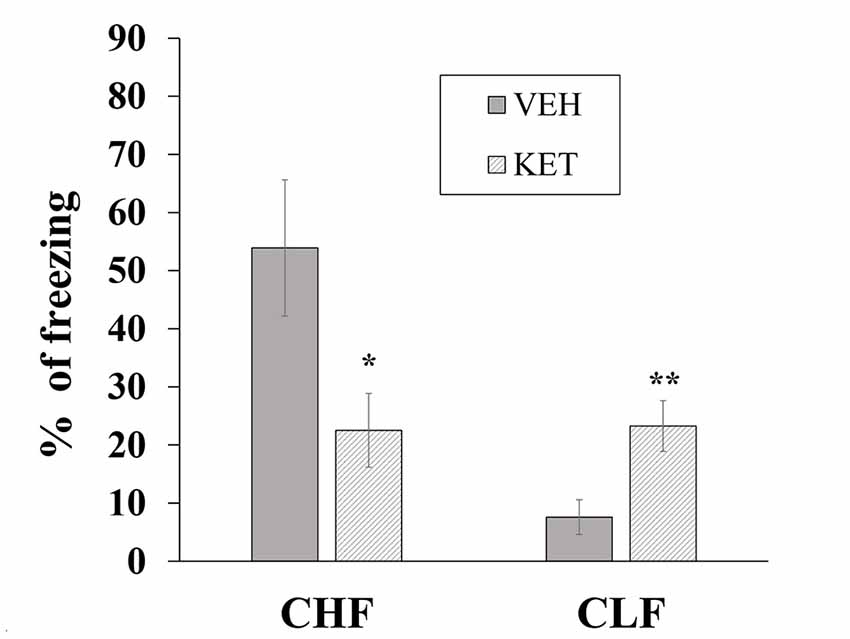
Figure 4. Mean + SEM percentage of time spent freezing in CHF and CLF animals that received ketanserin (KET) or vehicle (VEH) microinjections in the IL cortex. *p < 0.05 KET vs. VEH among CHF animals; **p < 0.05 KET vs. VEH among CLF animals.
Figure 5 shows coronal sections of the injection sites in the PL. The injections were distributed throughout the entire rostral-caudal extent of the target area within the PL. Three of thirty-nine rats in the experiment were excluded because their cannula missed the PL. The final sample size for each group was the following: CLF animals injected with vehicle (n = 10), CLF animals injected with ketanserin (n = 9), CHF animals injected with vehicle (n = 8) and CHF animals injected with ketanserin (n = 9).
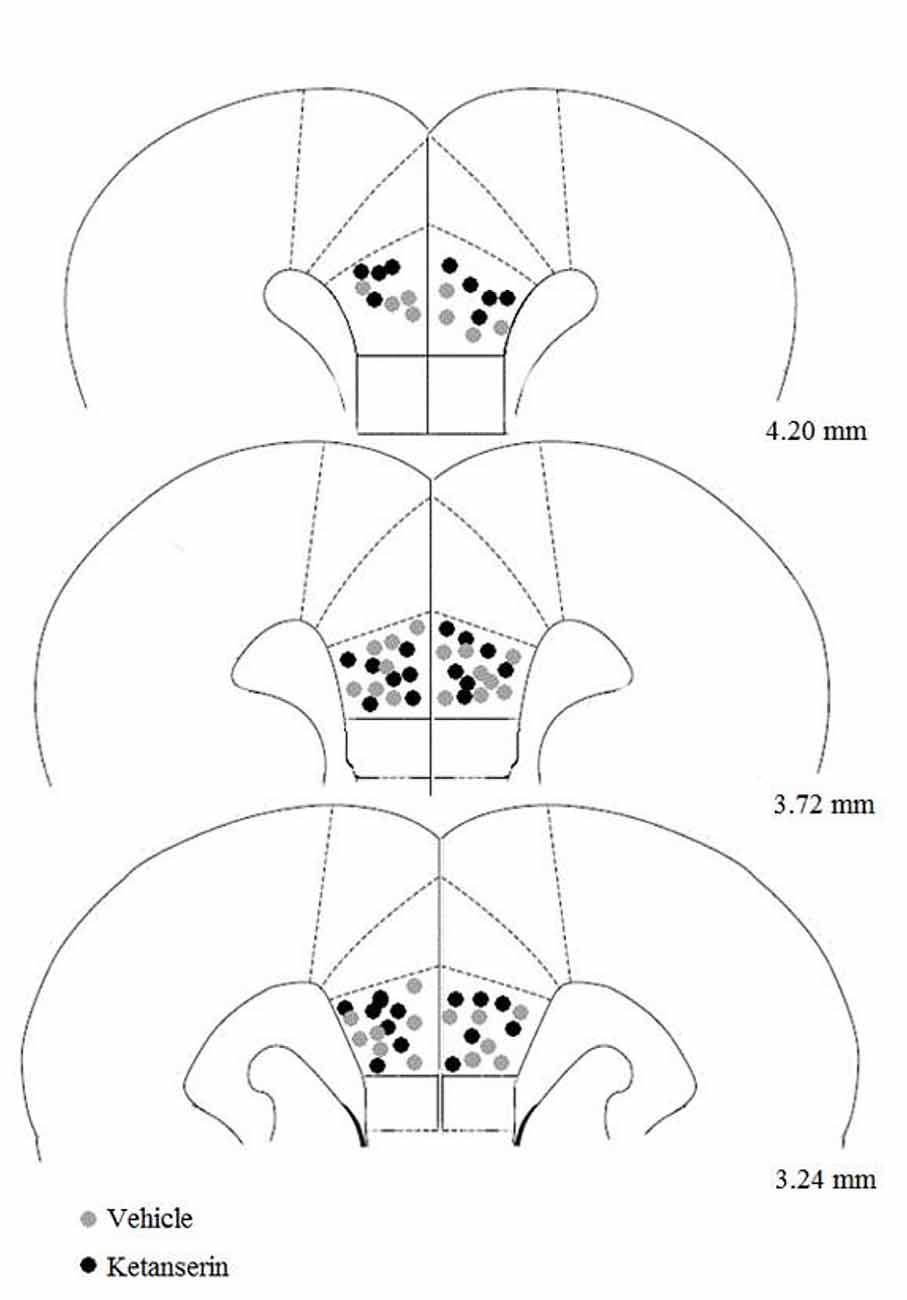
Figure 5. Site of microinjection tips in the prelimbic (PL) cortex. Gray square indicates site of vehicle injection. Black circle indicates site of ketanserin injections. Plates are taken from Paxinos and Watson (1986) and the numbers on the right side of each plate indicate the distance (in millimeters) from bregma.
Figure 6 shows the mean ± SEM percentage of open arm entries, percent time spent on the open arms and closed arm entries in the EPM in CHF and CLF animals that received microinjections of ketanserin or vehicle in the PL.
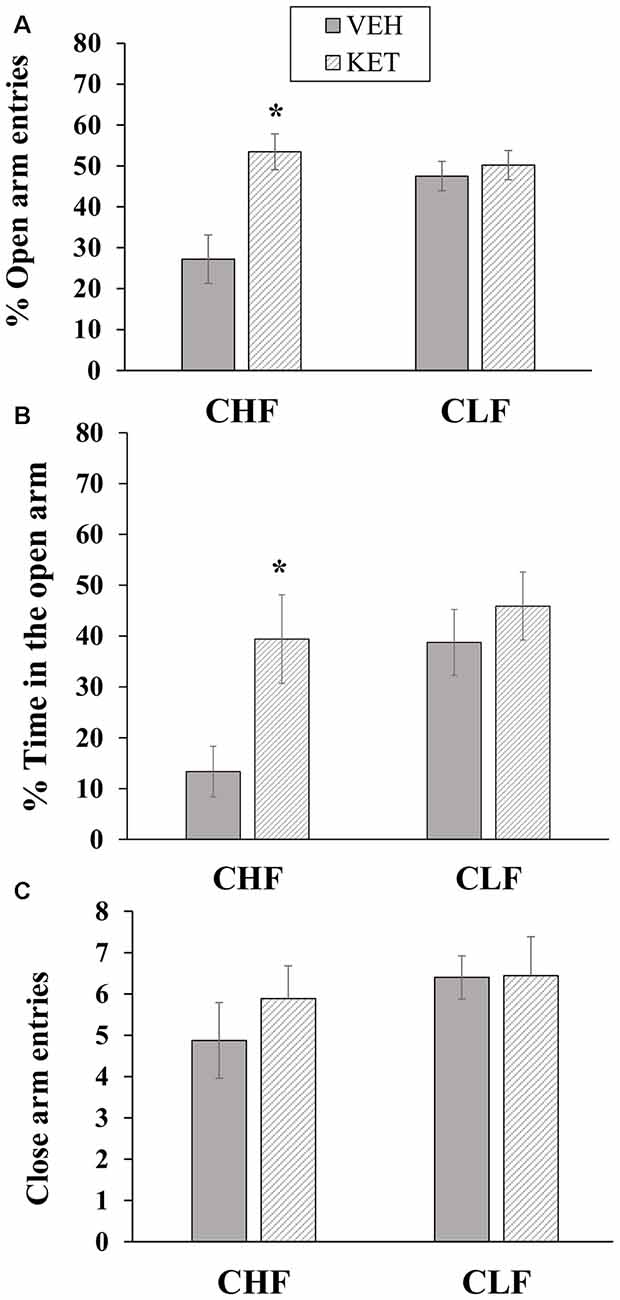
Figure 6. Mean + SEM percentage of open arm entries (A), percent time spent on the open arms (B) and closed arm entries (C) in the EPM in CHF and CLF animals that received ketanserin (KET) or vehicle (VEH) microinjections in the PL cortex. *p < 0.05 KET vs. VEH among CHF animals.
The two-way ANOVA of the percentage of open arm entries (Figure 6A) revealed a main effect of treatment (F(1,32) = 11.01, p < 0.05) and a rat line × treatment interaction (F(1,32) = 7.32, p < 0.01). No main effect of treatment was found (F(1,32) = 3.81, p > 0.05). The post hoc comparisons revealed that ketanserin microinjections in the PL significantly increased the percentage of open arm entries in CHF animals compared with vehicle (p < 0.05) but had no effect on this measure in CLF animals (p > 0.05).
The two-way ANOVA of the percentage of time spent on the open arms (Figure 6B) revealed main effects of treatment (F(1,32) = 6.47, p < 0.05) and rat line (F(1,32) = 5.98, p < 0.05). No interaction between rat line and treatment was found (F(1,32) = 2.10, p > 0.15).
Figure 6C shows the absolute number of closed arm entries in the EPM maze in CHF and CLF animals that received microinjections of ketanserin or vehicle. The two-way ANOVA revealed no main effect of rat line or treatment and no interaction between these factors (all p > 0.05).
Figure 7 shows the mean ± SEM percentage of time spent freezing in CHF and CLF animals that received microinjections of ketanserin or vehicle in the PL. The two-way ANOVA indicated main effects of rat line (F(1,32) = 26.02, p < 0.01), treatment (F(1,32) = 27.30, p < 0.01) and a significant rat line × treatment interaction (F(1,32) = 28.42, p < 0.01). The post hoc comparisons indicated that ketanserin microinjections in the PL significantly decreased conditioned freezing in CHF animals compared with vehicle (p < 0.05) but had no effect in CLF animals (p > 0.05).
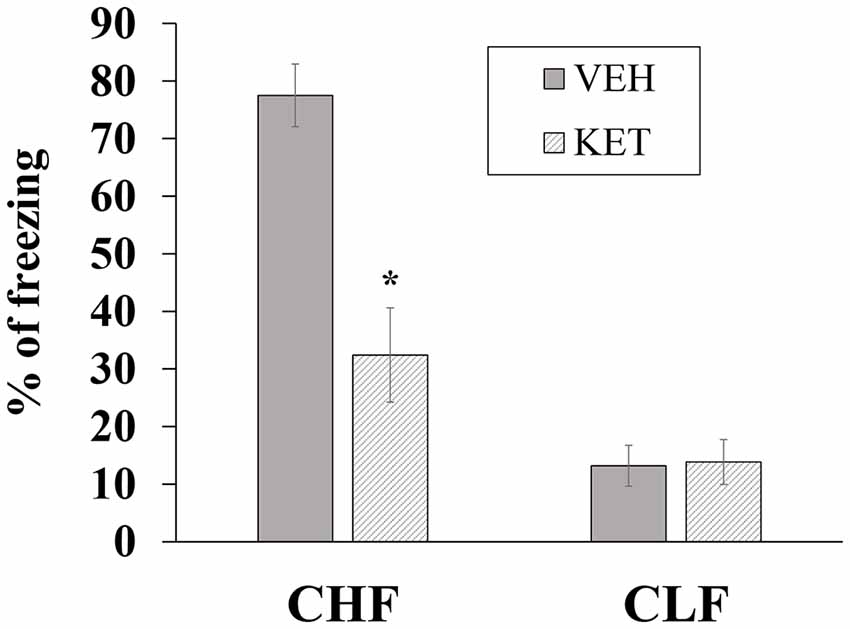
Figure 7. Mean + SEM percentage of time spent freezing in CHF and CLF animals that received ketanserin (KET) or vehicle (VEH) microinjections in the PL cortex. *p < 0.05 KET vs. VEH among CHF animals.
Although there are several animal models of anxiety, very few have highlighted individual differences in the vulnerability to threatening situations as an important variable for studying behavioral responses to pharmacological compounds (Steimer, 2011; Beckers et al., 2013). Selectively bred models of trait anxiety represent an important tool for investigating neural pathophysiological mechanisms associated with anxiety disorders (Castro-Gomes et al., 2013). The present study evaluated the behavioral response to the preferential 5-HT2A receptor antagonist ketanserin in two lines of animals that were selectively bred for high (CHF) and low (CLF) freezing responses to contextual cues that were previously associated with footshock.
The present results from these two lines of animals from the 10th, 15th and 20th generations consistently showed that CHF rats displayed a significantly more “anxious” phenotype, reflected by open arm parameters in the EPM compared with CLF animals. No differences were found between CHF and CLF rats in the number of closed arm entries, suggesting that the anxiety-like profile of CHF animals was not attributable to locomotor impairment but rather to increases in aversion to the open arms. These behavioral differences between CHF and CLF rats in the EPM are consistent with previous studies from our laboratory and indicate that the conditioned freezing parameter that is used for our breeding program remained stable in a different threatening situation across different generations (Hassan et al., 2013, 2015; Dias et al., 2014; Mousovich-Neto et al., 2015).
Importantly, a biological phenomenon known as genetic drift (Falconer and Mackay, 1996) might represent a confounding factor in our breeding program, in which allele frequencies significantly increase or decrease, possibly leading to differential fixation of the alleles in the CHF and CFL lines as a result of the small size of the two populations. Nevertheless, the present results and past findings from our group strongly indicate that our artificial selection program has indeed resulted in contrasting phenotypes between our two lines that are not attributable to random genetic effects, which cannot be entirely excluded in any evolutionary process that involves small breeding groups.
Systemic injections of ketanserin induced opposite effects in these two lines of rats when the animals were evaluated in the same animal model of anxiety. Ketanserin administration in CHF animals increased both the percentage of entries into and time spent on the open arms of the EPM, without changing the absolute number of closed arm entries. This indicates a selective anxiolytic-like effect without locomotor interference in this test. Conversely, CLF animals that received systemic ketanserin injections exhibited a behavioral pattern suggestive of an anxiogenic-like action in the EPM, reflected by a decrease in open arm exploration and no changes in closed arm entries. These results are consistent with a previous study that reported a bidirectional effect of systemic ketanserin administration in the EPM in female rats with different basal levels of anxiety associated with hormonal states (Díaz-Véliz et al., 1997). Ketanserin produced an anxiogenic-like effect in low-anxiety females that had high estrogen levels (proestrus) but produced an anxiolytic-like effect in high-anxiety females that had low estrogen levels (diestrus). León et al. (2009) showed that methylenedioxymethamphetamine and fluoxetine administration also had opposite effects, depending on whether the subjects were pre-exposed to chronic mild stress, suggesting differential effects of the drug that depended on basal conditions (León et al., 2009). Therefore, differences in baseline levels of anxiety appear to be important for determining the behavioral effects of serotonergic manipulations in the EPM. We previously reported in the rat EPM that the anxiogenic-like effects of the 5-HT2 receptor agonist TFMPP administered systemically were prevented by intra-amygdala infusions of the mixed 5-HT2A/2C-receptor antagonist ritanserin, which does not affect basal levels of anxiety in this animal model (de Mello Cruz et al., 2005).
The behavioral effects of pharmacological interventions are a dynamic process that involves both environmental and genetic factors. The present results highlight the importance of considering the underlying heterogeneity of a given experimental animal population. This is particularly important in basic research that investigates the role of serotonergic activity in anxiety. Conflicting results might be found because most experiments use very heterogeneous populations with considerable variations in anxiolytic-like responses to the same threatening situation. Ignoring the impact of individual differences in behavioral pharmacology research that is performed with heterogeneous populations of animals might mask behavioral effects due to an average result across different levels of defensive responses that these animals might present to cope with a threatening situation (Veenema et al., 2004; Beerling et al., 2011; Castro et al., 2012; Duclot and Kabbaj, 2013). Human and animal studies have shown individual differences in ways of coping with environmental challenges (Blanchard et al., 2001; de Kloet et al., 2005; Bardi et al., 2012; Metna-Laurent et al., 2012; Coppens et al., 2013). Using similar populations, some individuals display higher vulnerability to the development of anxiety-related disorders when faced with threatening situations, whereas other individuals seem to be more resilient to the development these types of pathologies (Bolger and Zuckerman, 1995; Hammen, 2005; Aisa et al., 2008; Uchida et al., 2008; Sandi and Richter-Levin, 2009; Oitzl et al., 2010). Therefore, individual variability needs to be taken into account in animal models of anxiety. Using animal lines that express an anxious-like phenotype is an important methodology for investigating the ways in which the underlying neuropharmacology might contribute to the observed behavioral differences.
The present study also investigated the participation of 5-HT2 receptors in the IL and the PL subregions of the vmPFC in CHF and CLF animals in the EPM and contextual fear conditioning paradigm. Intra-IL acute infusions of ketanserin induced the same bidirectional behavioral effects in the EPM as systemic injections (i.e., an anxiolytic-like effect in CHF rats but an anxiogenic-like effect in CLF rats), and these effects extended to contextual fear conditioning. Intra-PL acute infusions of ketanserin also had an anxiolytic-like effect in CHF animals but no effect in CLF rats in both models of anxiety. These results suggest that participation of the IL and PL on anxiety-like behavior might not depend solely on the nature (i.e., innate or learned) of the threatening stimulus, as suggested by Corcoran and Quirk (2007) and Sierra-Mercado et al. (2006), but rather on the genetic vulnerability of specific animals to the threatening situation. Moreover, ketanserin injections in either the IL or PL produced the same anxiolytic-like effect in CHF animals. Behavioral differences between ketanserin injections in the IL and PL were only observed in CLF animals, in which an anxiogenic-like effect was only observed when it was injected in the IL and not in the PL.
The IL sends descending projection to the basolateral complex of the amygdala (Likhtik et al., 2014). These projections seem to play an important role in the extinction of conditioned fear (McDonald, 1998; Vertes, 2004). Inactivation of the IL impaired the consolidation and retrieval of extinction of fear conditioning (Quirk et al., 2000; Laurent and Westbrook, 2009). The present study only found an anxiogenic-like effect in CLF rats when the IL region was microinjected with ketanserin. Surprisingly, CHF animals displayed an anxiolytic-like response to microinjection of the same 5-HT2 antagonist. This could be related to differences in 5-HT2 receptor expression in this subregion of the vmPFC in both lines of animals. 5-HT2 receptor downregulation after chronic treatment with antidepressants (Peroutka and Snyder, 1980) and serotonin receptor agonists has been reported (Conn and Sanders-Bush, 1986; Leysen et al., 1987a,b; Smith et al., 1999).
Neurons in the PL play an excitatory role in conditioned fear behavior. This excitatory effect appears also to be mediated by descending projections to the basolateral complex of the amygdala (Likhtik et al., 2014). Previous results indicated that pharmacological inhibition of the PL reduced the expression of conditioned fear (Corcoran and Quirk, 2007). Furthermore, immunohistochemistry indicated that the PL exhibits greater activation when animals are reexposed to contextual cues that were previously associated with footshock (Lemos et al., 2010). Our results in CHF animals are consistent with these previous studies and highlight the participation of postsynaptic 5-HT2A receptors in the PL in contextual fear conditioning. However, ketanserin did not induce any reduction in conditioned fear among CLF animals, which might be attributable to a floor effect because the animals that received vehicle injections already had very low levels of the freezing response.
Importantly, the present study has several limitations that need to be considered when interpreting the findings. One of the limitations refers to the selectivity of ketanserin. Although this drug has been extensively used as a pharmacological tool to efficiently block 5-HT2A receptors (Kristiansen and Dahl, 1996; López-Giménez et al., 1997; Knight et al., 2004), it also has high binding affinity for both histamine H1 (Wouters et al., 1985; Ghoneim et al., 2006) and α1-adrenergic (Hoyer et al., 1987; Israilova et al., 2002) receptors. Thus, the participation of vmPFC 5-HT2A receptors in bidirectional behavioral changes in CHF and CLF rats should not be considered in isolation. Although beyond the scope of the present study, consideration should be given to the possible influence of H1 and α1-adrenergic receptors in the present results. Future studies should further investigate the involvement of 5-HT2A receptors in the vmPFC in CHF and CLF animals.
Substantial contributions to the conception or design of the work; or the acquisition, analysis, or interpretation of data for the work; and drafting the work or revising it critically for important intellectual content; and final approval of the version to be published; and agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors are thankful to Dr. André Ramos (UFSC) for helpful comments and discussions. This work was supported by grants from the National Council for Scientific and Technological Development (CNPq) and Rio de Janeiro State Research Foundation (FAPERJ) to JL-F. LAL had a doctoral fellowship from CAPES. SZ-G and KC had doctoral fellowships from the Colombian government. VC-G had a postdoctoral fellowship from CNPq.
Aisa, B., Tordera, R., Lasheras, B., Del Rio, J., and Ramírez, M. J. (2008). Effects of maternal separation on hypothalamic-pituitary-adrenal responses, cognition and vulnerability to stress in adult female rats. Neuroscience 154, 1218–1226. doi: 10.1016/j.neuroscience.2008.05.011
Allen, T. A., Narayanan, N. S., Kholodar-Smith, D. B., Zhao, Y., Laubach, M., and Brown, T. H. (2008). Imaging the spread of reversible brain inactivations using fluorescent muscimol. J. Neurosci. Methods 171, 30–38. doi: 10.1016/j.jneumeth.2008.01.033
Almada, R. C., Borelli, K. G., Albrechet-Souza, L., and Brandão, M. L. (2009). Serotonergic mechanisms of the median raphe nucleus-dorsal hippocampus in conditioned fear: output circuit involves the prefrontal cortex and amygdala. Behav. Brain Res. 203, 279–287. doi: 10.1016/j.bbr.2009.05.017
Alves, S. H., Pinheiro, G., Motta, V., Landeira-Fernandez, J., and Cruz, A. P. M. (2004). Anxiogenic effects in the rat elevated plus-maze of 5-HT2C agonists into ventral but not dorsal hippocampus. Behav. Pharmacol. 15, 37–43. doi: 10.1097/00008877-200402000-00005
Avesar, D., and Gulledge, A. T. (2012). Selective serotonergic excitation of callosal projection neurons. Front. Neural Circuits 6:12. doi: 10.3389/fncir.2012.00012
Azmitia, E. C., and Segal, M. (1978). An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat. J. Comp. Neurol. 179, 641–667. doi: 10.1002/cne.901790311
Bardi, M., Rhone, A. P., Franssen, C. L., Hampton, J. E., Shea, E. A., Hyer, M. M., et al. (2012). Behavioral training and predisposed coping strategies interact to influence resilience in male Long-Evans rats: implications for depression. Stress 15, 306–317. doi: 10.3109/10253890.2011.623739
Bastani, B., Nash, J. F., and Meltzer, H. Y. (1990). Prolactin and cortisol responses to MK-212, a serotonin agonist in obsessive-compulsive disorder. Arch. Gen. Psychiatry 47, 833–839. doi: 10.1001/archpsyc.1990.01810210041006
Beckers, T., Krypotos, A. M., Boddez, Y., Effting, M., and Kindt, M. (2013). What’s wrong with fear conditioning? Biol. Psychol. 92, 90–96. doi: 10.1016/j.biopsycho.2011.12.015
Beerling, W., Koolhaas, J. M., Ahnaou, A., Bouwknecht, J. A., de Boer, S. F., Meerlo, P., et al. (2011). Physiological and hormonal responses to novelty exposure in rats are mainly related to ongoing behavioral activity. Physiol. Behav. 103, 412–420. doi: 10.1016/j.physbeh.2011.03.014
Bi, L. L., Wang, J., Luo, Z. Y., Chen, S. P., Geng, F., Chen, Y. H., et al. (2013). Enhanced excitability in the infralimbic cortex produces anxiety-like behaviors. Neuropharmacology 72, 148–156. doi: 10.1016/j.neuropharm.2013.04.048
Blanchard, D. C., Hynd, A. L., Minke, K. A., Minemoto, T., and Blanchard, R. J. (2001). Human defensive behaviors to threat scenarios show parallels to fear- and anxiety-related defense patterns of non-human mammals. Neurosci. Biobehav. Rev. 25, 761–770. doi: 10.1016/s0149-7634(01)00056-2
Blue, M. E., Yagaloff, K. A., Mamounas, L. A., Hartig, P. R., and Molliver, M. E. (1988). Correspondence between 5-HT2 receptors and serotonergic axons in rat neocortex. Brain Res. 453, 315–328. doi: 10.1016/0006-8993(88)90172-2
Bolger, N., and Zuckerman, A. (1995). A framework for studying personality in the stress process. J. Pers. Soc. Psychol. 69, 890–902. doi: 10.1037//0022-3514.69.5.890
Brandão, M. L., Zanoveli, J. M., Ruiz-Martinez, R. C., Oliveira, L. C., and Landeira-Fernandez, J. (2008). Different patterns of freezing behavior organized in the periaqueductal gray of rats: association with different types of anxiety. Behav. Brain Res. 188, 1–13. doi: 10.1016/j.bbr.2007.10.018
Bull, E. J., Huston, P. H., and Fone, K. C. (2003). Reduced social interaction following 3,4-methylenedioxymethamphetamine is not associated with enhanced 5-HT2C receptor responsivity. Neuropharmacology 44, 439–448. doi: 10.1016/s0028-3908(02)00407-0
Campbell, B. M., and Merchant, K. M. (2003). Serotonin 2C receptors within the basolateral amygdala induce acute fear-like responses in an open-field environment. Brain Res. 993, 1–9. doi: 10.1016/s0006-8993(03)03384-5
Castro, J. E., Diessler, S., Varea, E., Márquez, C., Larsen, M. H., Cordero, M. I., et al. (2012). Personality traits in rats predict vulnerability and resilience to developing stress-induced depression-like behaviors, HPA axis hyper-reactivity and brain changes in pERK1/2 activity. Psychoneuroendocrinology 37, 1209–1223. doi: 10.1016/j.psyneuen.2011.12.014
Castro-Gomes, V., Hassan, W., Maisonnette, S., Johnson, L. R., Ramos, A., and Landeira-Fernandez, J. (2013). Behavioral evaluation of eight rat lines selected for high and low anxiety-related responses. Behav. Brain Res. 257, 39–48. doi: 10.1016/j.bbr.2013.09.028
Castro-Gomes, V., and Landeira-Fernandez, J. (2008). Amygdaloid lesions produced similar contextual fear conditioning disruption in the Carioca High- and Low-conditioned Freezing rats. Brain Res. 1233, 137–145. doi: 10.1016/j.brainres.2008.07.044
Castro-Gomes, V., León, L. A., Mograbi, D., Cardenas, L. F., and Landeira-Fernandez, J. (2014). Contextual fear extinction and re-extinction in carioca high- and low-conditioned freezing rats. World J. Neurosci. 4, 247–252. doi: 10.4236/wjns.2014.43028
Castro-Gomes, V., Silva, C. E. B., and Landeira-Fernandez, J. (2011). “The carioca high- and low-conditioned freezing lines: a new animal model of generalized anxiety disorder,” in Anxiety Disorders, ed. V. Kalinin (Rijeka: Intech), 121–134.
Chaouloff, F., Kulikov, A., and Mormède, P. (1997). Repeated DOI and SR 46349B treatments do not affect elevated plus-maze anxiety despite opposite effects on cortical 5-HT2A receptors. Eur. J. Pharmacol. 334, 25–29. doi: 10.1016/s0014-2999(97)01197-7
Charney, D. S., Woods, S. W., Goodman, W. R., and Heninger, G. R. (1987). Serotonin function in anxiety. II. Effects of the serotonin agonist mCPP on panic disorder patients and healthy subjects. Psychopharmacology 92, 14–21. doi: 10.1007/BF00215473
Conn, P. J., and Sanders-Bush, E. (1986). Regulation of serotonin-stimulated phosphoinositide hydrolysis: relation to the serotonin 5-HT-2 binding site. J. Neurosci. 6, 3669–3675.
Coppens, C. M., de Boer, S. F., Steimer, T., and Koolhaas, J. M. (2013). Correlated behavioral traits in rats of the Roman selection lines. Behav. Genet. 43, 220–226. doi: 10.1007/s10519-013-9588-8
Corcoran, K. A., and Quirk, G. J. (2007). Activity in prelimbic cortex is necessary for the expression of learned, but not innate, fears. J. Neurosci. 27, 840–844. doi: 10.1523/JNEUROSCI.5327-06.2007
Critchley, M. A., and Handley, S. L. (1987). Effects in the X-maze model of agents acting at 5-HT1 and 5-HT2 receptors. Psychopharmacology 93, 502–506. doi: 10.1007/bf00207243
Cruz, A. P. M., Frei, F., and Graeff, F. G. (1994). Ethopharmacological analysis of rat behavior on the elevated plus-maze. Pharmacol. Biochem. Behav. 49, 171–176. doi: 10.1016/0091-3057(94)90472-3
Davie, M., Wilkinson, L. S., and Roberts, M. H. (1988). Evidence for excitatory 5-HT2-receptors on rat brainstem neurones. Br. J. Pharmacol. 94, 483–491. doi: 10.1111/j.1476-5381.1988.tb11551.x
de Kloet, E. R., Joels, M., and Holsboer, F. (2005). Stress and the brain: from adaptation to disease. Nat. Rev. Neurosci. 6, 463–475. doi: 10.1038/nrn1683
de Luca, M. C., Brandão, M. L., Motta, V. A., and Landeira-Fernandez, J. (2003). Antinociception induced by stimulation of ventrolateral periaqueductal gray at the freezing threshold is regulated by opioid and 5-HT2A receptors as assessed by the tail-flick and formalin tests. Pharmacol. Biochem. Behav. 75, 459–466. doi: 10.1016/s0091-3057(03)00145-x
de Mello Cruz, A. P., Pinheiro, G., Alves, S. H., Ferreira, G., Mendes, M., Faria, L., et al. (2005). Behavioral effects of systemically administered MK-212 areprevented by ritanserinmicroinfusion into the basolateral amygdala of rats exposed to the elevated plus-maze. Psychopharmacology 182, 345–354. doi: 10.1007/s00213-005-0108-2
Dias, G. P., Bevilaqua, M. C. N., da Luz, A. C. D. S., Fleming, R. L., de Carvalho, L. A., Cocks, G., et al. (2014). Hippocampal biomarkers of fear memory in an animal model of generalized anxiety disorder. Behav. Brain Res. 263, 34–45. doi: 10.1016/j.bbr.2014.01.012
Dias, G. P., Bevilaqua, M. C., Silveira, A. C., Landeira-Fernandez, J., and Gardino, P. F. (2009). Behavioral profile and dorsal hippocampal cells in Carioca High-conditioned Freezing rats. Behav. Brain Res. 205, 342–348. doi: 10.1016/j.bbr.2009.06.038
Díaz-Véliz, G., Alarcón, T., Espinoza, C., Dussaubat, N., and Mora, S. (1997). Ketanserin and anxiety levels: influence of gender, estrous cycle, ovariectomy and ovarian hormones in female rats. Pharmacol. Biochem. Behav. 58, 637–642. doi: 10.1016/s0091-3057(97)90004-6
Duclot, F., and Kabbaj, M. (2013). Individual differences in novelty seeking predict subsequent vulnerability to social defeat through a differential epigenetic regulation of brain-derived neurotrophic factor expression. J. Neurosci. 33, 11048–11060. doi: 10.1523/JNEUROSCI.0199-13.2013
Durand, M., Mormèd, P., and Chaouloff, F. (2003). Wistar-Kyoto rats are sensitive to the hypomotor and anxiogenic effects of mCPP. Behav. Pharmacol. 14, 173–177. doi: 10.1097/00008877-200303000-00010
Eison, A. S., and Mullins, U. L. (1996). Regulation of central 5-HT2A receptors: a review of in vivo studies. Behav. Brain Res. 73, 177–181. doi: 10.1016/0166-4328(96)00092-7
Falconer, D. S. M., and Mackay, T. F. C. (1996). Introduction to Quantitative Genetics. Essex: Longman.
Fanselow, M. S., and Helmstetter, F. J. (1988). Conditional analgesia, defensive freezing, and benzodiazepines. Behav. Neurosci. 102, 233–243. doi: 10.1037//0735-7044.102.2.233
Fanselow, M. S., Helmstetter, F. J., and Calcagnetti, D. J. (1991). “Parallels between the behavioral effects of dimethoxy-carboline (DMCM) and conditional fear stimuli,” in Current Topics in Animal Learning: Brain, Emotion and Cognition, eds L. Dachowski and C. F. Flaherty (Hillsdale, NJ: Lawrence Erlbaum), 187–206.
File, S. E. (1992). “Behavioural detection of anxiolytic action,” in Experimental Approaches to Anxiety and Depression, eds J. M. Elliottm, D. J. Heal and C. A. Marsden (Chichester: Wiley), 25–44.
Galvão, B. O., Larrubia, B. C., Hommes, W. J., Cardenas, L. F., de Mello Cruz, A. P., and Landeira-Fernandez, J. (2010). Effects of contextual fear conditioning and pentylenetetrazol on panic-like reactions induced by dorsal periaqueductal gray stimulation with N-methyl-D-aspartate. Psychol. Neurosci. 3, 67–72. doi: 10.3922/j.psns.2010.1.008
Ghoneim, O. M., Legere, J. A., Golbraikh, A., Tropsha, A., and Booth, R. G. (2006). Novel ligands for the human histamine H1 receptor: synthesis, pharmacology and comparative molecular field analysis studies of 2-dimethylamino-5–(6)-phenyl-1,2,3,4-tetrahydronaphthalenes. Bioorganic and Medicinal Chemistry 14, 6640–6658. doi: 10.1016/j.bmc.2006.05.077
Gibson, E. L., Barnfield, A. M., and Curzon, G. (1994). Evidence that mCPP-induced anxiety in the plus-maze is mediated by postsynaptic 5-HT2C receptors but not by sympathomimetic effects. Neuropharmacology 33, 457–465. doi: 10.1016/0028-3908(94)90076-0
Gordon, J. A., and Hen, R. (2004). The serotonergic system and anxiety. Neuromolecular Med. 5, 27–40. doi: 10.1385/nmm:5:1:027
Graeff, F. G. (2002). On serotonin and experimental anxiety. Psychopharmacology 163, 467–476. doi: 10.1007/s00213-002-1112-4
Graeff, F. G. (2004). Serotonin, the periaquedutalgray and panic. Neurosci. Biobehav. Rev. 28, 239–259. doi: 10.1016/j.neubiorev.2003.12.004
Graeff, F. G., Silveira, M. C. L., Nogueira, R. L., Audi, E. A., and Oliveira, R. M. W. (1993). Role of the amygdala and periaquedutal gray in anxiety and panic. Behav. Brain Res. 58, 123–131. doi: 10.1016/0166-4328(93)90097-a
Halberstadt, A. L. (2015). Recent advances in the neuropsychopharmacology of serotonergic hallucinogens. Behav. Brain Res. 277, 99–120. doi: 10.1016/j.bbr.2014.07.016
Hammen, C. (2005). Stress and depression. Annu. Rev. Clin. Psychol. 1, 293–319. doi: 10.1146/annurev.clinpsy.1.102803.143938
Hannon, J., and Hoyer, D. (2008). Molecular biology of 5-HT receptors. Behav. Brain Res. 195, 198–213. doi: 10.1016/j.bbr.2008.03.020
Hashimoto, S., Inoue, T., and Koyama, T. (1996). Serotonin reuptake inhibitors reduce conditioned fear stress-induced freezing behavior in rats. Psychopharmacology 123, 182–186. doi: 10.1007/bf02246175
Hassan, W., de Castro Gomes, V., Pinton, S., Batista Teixeira da Rocha, J., and Landeira-Fernandez, J. (2013). Association between oxidative stress and contextual fear conditioning in carioca high- and low-conditioned freezing rats. Brain Res. 1512, 60–67. doi: 10.1016/j.brainres.2013.03.039
Hassan, W., Silva, C. E., Rocha, J. B., and Landeira-Fernandez, J. (2015). Modulatory effect of diphenyl diselenide in carioca high- and low-conditioned freezing rats. Eur. J. Pharmacol. 761, 341–344. doi: 10.1016/j.ejphar.2015.05.069
Hasuo, H., Matsuoka, T., and Akasu, T. (2002). Activation of presynaptic 5-Hydroxytryptamine 2A Receptors facilitates excitatory synaptic transmission via protein kinase C in the dorsolateral septal nucleus. J. Neurosci. 22, 7509–7517.
Hoyer, D., Hannon, J. P., and Martin, G. R. (2002). Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 71, 533–554. doi: 10.1016/s0091-3057(01)00746-8
Hoyer, D., Vos, P., Closse, A., Palacios, J. M., Engel, G., and Davies, H. (1987). [3H]ketanserin labels serotonin 5-HT2 and alpha 1-adrenergic receptors in human brain cortex. J. Cardiovasc. Pharmacol. 10, S48–S50. doi: 10.1097/00005344-198700103-00012
Hughes, C. R., Tran, L., and Keele, B. (2012). 5-HT2A receptor activation normalizes exaggerated fear behavior in p-Chlorophenylalanine (PCPA)-treated rats. J. Behav. Brain Sci. 2, 454–462. doi: 10.4236/jbbs.2012.24053
Inoue, T., Tsuchiya, K., and Koyama, T. (1996). Effects of typical and atypical antipsychotic drugs on freezing behavior induced by conditioned fear. Pharmacol. Biochem. Behav. 55, 195–201. doi: 10.1016/s0091-3057(96)00064-0
Israilova, M., Suzuki, F., Tanaka, T., Nagatomo, T., Taniguchi, T., and Muramatsu, I. (2002). Binding and functional affinity of sarpogrelate, its metabolite m-1 and ketanserin for human recombinant alpha-1-adrenoceptor subtypes. Pharmacology 65, 69–73. doi: 10.1159/000056189
Jones, N., Duxon, M. S., and King, S. M. (2002). 5-HT2C receptor mediation of unconditioned escape behaviour in the unstable elevated exposed plus maze. Psychopharmacology 164, 214–220. doi: 10.1007/s00213-002-1197-9
Kennett, G. A., Pittaway, K., and Blackburn, T. P. (1994). Evidence that 5-HT2C receptor antagonists are anxiolytic in the Geller-Seifter model of anxiety. Psychopharmacology 114, 90–96. doi: 10.1007/bf02245448
Knight, A. R., Misra, A., Quirk, K., Benwell, K., Revell, D., Kennett, G., et al. (2004). Pharmacological characterisation of the agonist radioligand binding site of 5-HT2A, 5-HT2B and 5-HT2C receptors. Naunyn Schmiedebergs Arch. Pharmacol. 370, 114–123. doi: 10.1007/s00210-004-0951-4
Kristiansen, K., and Dahl, S. G. (1996). Molecular modeling of serotonin, ketanserin, ritanserin and their 5-HT2C receptor interactions. Eur. J. Pharmacol. 306, 195–210. doi: 10.1016/0014-2999(96)00180-X
Landeira-Fernandez, J. (1996). Context and Pavlovian conditioning. Braz. J. Med. Biol. Res. 29, 149–173.
Landeira-Fernandez, J., DeCola, J. P., Kim, J. J., and Fanselow, M. S. (2006). Immediate shock deficit in fear conditioning: effects of shock manipulations. Behav. Neurosci. 120, 873–879. doi: 10.1037/0735-7044.120.4.873
Laurent, V., and Westbrook, R. F. (2009). Inactivation of the infralimbic but not the prelimbic cortex impairs consolidation and retrieval of fear extinction. Learn. Mem. 16, 520–529. doi: 10.1101/lm.1474609
Lemos, J. I., Resstel, L. B., and Guimarães, F. S. (2010). Involvement of the prelimbic prefrontal cortex on cannabidiol-induced attenuation of contextual conditioned fear in rats. Behav. Brain Res. 207, 105–111. doi: 10.1016/j.bbr.2009.09.045
León, L. A., Landeira-Fernandez, J., and Cardenas, F. P. (2009). Effects of chronic intracerebroventricular 3,4-methylenedioxy-N-methamphetamine (MDMA) or fluoxetine on the active avoidance test in rats with or without exposure to mild chronic stress. Brain Res. 205, 259–264. doi: 10.1016/j.bbr.2009.06.039
Leysen, J. E., Van Gompel, P., de Chaffoy de Courcelles, D., and Niemegeers, C. J. (1987a). Opposite regulation of serotonin-S2 and dopamine-D2 receptors in rat brain following chronic receptor blockade. J. Recept. Res. 7, 223–239. doi: 10.3109/10799898709054987
Leysen, J. E., Van Gompel, P., Gommeren, W., and Laduron, P. M. (1987b). Differential regulation of dopamine-D2 and serotonin-S2 receptors by chronic treatment with the serotonin-S2 antagonists, ritanserin, and setoperone. Psychopharmacol. Ser. 3, 214–224. doi: 10.1007/978-3-642-71288-3_25
Likhtik, E., Stujenske, J. M., Topiwala, M. A., Harris, A. Z., and Gordon, J. A. (2014). Prefrontal entrainment of amygdala activity signals safety in learned fear and innate anxiety. Nat. Neurosci. 17, 106–113. doi: 10.1038/nn.3582
Lisboa, S. F., Stecchini, M. F., Corrêa, F. M., Guimarães, F. S., and Resstel, L. B. (2010). Different role of the ventral medial prefrontal cortex on modulation of innate and associative learned fear. Neuroscience 171, 760–768. doi: 10.1016/j.neuroscience.2010.09.048
López-Giménez, J. F., Mengod, G., Palacios, J. M., and Vilaró, M. T. (1997). Selective visualization of rat brain 5-HT2A receptors by autoradiography with [3H]MDL 100,907. Naunyn Schmiedebergs Arch. Pharmacol. 356, 446–454. doi: 10.1007/pl00005075
Lowy, M. T., and Meltzer, H. Y. (1988). Stimulation of cortisol and prolactin secretion in humans by MK-212, a centrally active serotonin agonist. Biol. Psychiatry 23, 818–828. doi: 10.1016/0006-3223(88)90070-4
McDonald, A. J. (1998). Cortical pathways to the mammalian amygdala. Prog. Neurobiol. 55, 257–332. doi: 10.1016/s0301-0082(98)00003-3
Menard, J., and Treit, D. (1999). Effects of centrally administered anxiolytic compounds in animal models of anxiety. Neurosci. Biobehav. Rev. 23, 591–613. doi: 10.1016/s0149-7634(98)00056-6
Metna-Laurent, M., Soria-Gómez, E., Verrier, D., Conforzi, M., Jégo, P., Lafenêtre, P., et al. (2012). Bimodal control of fear-coping strategies by CB1 cannabinoid receptors. J. Neurosci. 32, 7109–7118. doi: 10.1523/JNEUROSCI.1054-12.2012
Millan, M. J. (2003). The neurobiology and control of anxious states. Prog. Neurobiol. 70, 83–244. doi: 10.1016/s0301-0082(03)00087-x
Motta, V., Maisonnette, S., Morato, S., Castrechini, P., and Brandão, M. L. (1992). Effects of blockade of 5-HT2 receptors and activation of 5-HT1A receptors on the exploratory activity of rats in the elevated plus-maze. Psychopharmacology 107, 135–139. doi: 10.1007/bf02244978
Mousovich-Neto, F., Lourenço, A. L., Landeira-Fernandez, J., and Corrêa da Costa, V. M. (2015). Endocrine and metabolic function in male Carioca High-conditioned Freezing rats. Physiol. Behav. 142, 90–96. doi: 10.1016/j.physbeh.2015.01.028
Naughton, M., Mulrooney, J. B., and Leonard, B. E. (2000). A review of the role of serotonin receptors in psychiatric disorders. Hum. Psychopharmacol. 15, 397–415. doi: 10.1002/1099-1077(200008)15:6<397::aid-hup212>3.0.co;2-l
Nic Dhonnchadha, B. A., Bourin, M., and Hascoët, M. (2003). Anxiolytic-like effects of 5-HT2 ligands on three mouse models of anxiety. Behav. Brain Res. 140, 203–214. doi: 10.1016/s0166-4328(02)00311-x
Oitzl, M. S., Champagne, D. L., van der Veen, R., and de Kloet, E. R. (2010). Brain development under stress: hypotheses of glucocorticoid actions revisited. Neurosci. Biobehav. Rev. 34, 853–866. doi: 10.1016/j.neubiorev.2009.07.006
Oliveira, L. C., Broiz, A. C., de Macedo, C. E., Landeira-Fernandez, J., and Brandão, M. L. (2007). 5-HT2 receptor mechanisms of the dorsal periaqueductal gray in the conditioned and unconditioned fear in rats. Psychopharmacology 191, 253–262. doi: 10.1007/s00213-006-0653-3
Paxinos, G., and Watson, C. (1986). The Rat Brain in Stereotaxic Coordinates. 2nd Edn. New York, NY: Academic Press.
Pazos, A., Cortés, R., and Palacios, J. M. (1985). Quantitative autoradiographic mapping of serotonin receptors in the rat brain: II. Serotonin-2 receptors. Brain Res. 346, 231–249. doi: 10.1016/0006-8993(85)90857-1
Pellow, S., Chopin, P., File, S. E., and Briley, M. (1985). Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 14, 149–167. doi: 10.1016/0165-0270(85)90031-7
Pellow, S., and File, S. E. (1986). Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: a novel test of anxiety in the rat. Pharmacol. Biochem. Behav. 24, 525–529. doi: 10.1016/0091-3057(86)90552-6
Pellow, S., Johnston, A. L., and File, S. E. (1987). Selective agonists and antagonists for 5-hydroxytryptamine receptor subtypes and interactions with yohimbine and FG 7142 using the elevated plus-maze test in the rat. J. Pharm. Pharmacol. 39, 917–928. doi: 10.1111/j.2042-7158.1987.tb03129.x
Peroutka, S. J., and Snyder, S. H. (1980). Regulation of serotonin2 (5-HT2) receptors labeled with [3H]spiroperidol by chronic treatment with the antidepressant amitriptyline. J. Pharmacol. Exp. Ther. 215, 582–587.
Pompeiano, M., Palacios, J. M., and Mengod, G. (1994). Distribution of the serotonin 5-HT2 receptor family mRNAs: comparison between 5-HT2A and 5-HT2C receptors. Mol. Brain Res. 23, 163–178. doi: 10.1016/0169-328x(94)90223-2
Quirk, G. J., Russo, G. K., Barron, J. L., and Lebron, K. (2000). The role of ventromedial prefrontal cortex in the recovery of extinguished fear. J. Neurosci. 20, 6225–6231.
Reibaud, M., and Böhme, G. A. (1993). “Evaluation of putative anxiolytics in the elevated plus-maze test,” in Paradigms for the Study of Behavior (Series Title: Methods in Neurosciences), ed. M. Conn (San Diego, CA: Academic Press), 230–239.
Ripoll, N., Hascoët, M., and Bourin, M. (2006). Implication of 5-HT2A subtype in DOI activity in the four-plates test-retest paradigm in mice. Behav. Brain Res. 166, 131–139. doi: 10.1016/j.bbr.2005.07.013
Rodgers, R. J., Cole, J. C., Cobain, M. R., Daly, P., Doran, P. J., Eells, J. R., et al. (1992). Anxiogenic-like effects of fluprazine and eltoprazine in the mouse elevated plus-maze: profile comparisons with 8-OH-DPAT, CG 12066b, TFMPP and mCPP. Behav. Pharmacol. 3, 621–634. doi: 10.1097/00008877-199212000-00009
Salviano, M., Ferreira, G., Greidinger, M., Couto, K., Landeira-Fernandez, J., and Cruz, A. P. M. (2014). Behavioral evaluation of male and female carioca high- and low-freezing rats. Trends Psychol. 22, 663–675. doi: 10.9788/tp2014.3-11
Sandi, C., and Richter-Levin, G. (2009). From high anxiety trait to depression: a neurocognitive hypothesis. Trends Neurosci. 32, 312–320. doi: 10.1016/j.tins.2009.02.004
Santana, N., Bortolozzi, A., Serrats, J., Mengod, G., and Artigas, F. (2004). Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb. Cortex 14, 1100–1109. doi: 10.1093/cercor/bhh070
Setem, J., Pinheiro, A. P., Motta, V. A., Morato, S., and Cruz, A. P. (1999). Ethopharmacological analysis of 5-HT ligands on the rat elevated plus-maze. Pharmacol. Biochem. Behav. 62, 515–521. doi: 10.1016/s0091-3057(98)00193-2
Sierra-Mercado, D. Jr., Corcoran, K. A., Lebrón-Milad, K., and Quirk, G. J. (2006). Inactivation of the ventromedial prefrontal cortex reduces expression of conditioned fear and impairs subsequent recall of extinction. Eur. J. Neurosci. 24, 1751–1758. doi: 10.1111/j.1460-9568.2006.05014.x
Sierra-Mercado, D., Padilla-Coreano, N., and Quirk, G. J. (2011). Dissociable roles of prelimbic and infralimbic cortices, ventral hippocampus and basolateral amygdala in the expression and extinction of conditioned fear. Neuropsychopharmacology 36, 529–538. doi: 10.1038/npp.2010.184
Sigmundi, R. A., Bouton, M. E., and Bolles, R. C. (1983). Conditioned freezing in the rat as a function of shock intensity and CS modality. Bull. Psychonom. Soc. 15, 254–256. doi: 10.3758/bf03334524
Smith, R. L., Barrett, R. J., and Sanders-Bush, E. (1999). Mechanism of tolerance development to 2,5-dimethoxy-4-iodoamphetamine in rats: down-regulation of the 5-HT2A, but not 5-HT2C, receptor. Psychopharmacology 144, 248–254. doi: 10.1007/s002130051000
Steimer, T. (2011). Animal models of anxiety disorders in rats and mice: some conceptual issues. Dialogues Clin. Neurosci. 13, 495–506.
Steinbusch, H. W. (1981). Distribution of serotonin-immunoreactivity in the central nervous system of the rat—cell bodies and terminals. Neuroscience 6, 557–618. doi: 10.1016/0306-4522(81)90146-9
Treit, D., Menard, J., and Royan, C. (1993). Anxiogenic stimuli in the elevated plus-maze. Pharmacol. Biochem. Behav. 44, 463–469. doi: 10.1016/0091-3057(93)90492-c
Uchida, S., Nishida, A., Hara, K., Kamemoto, T., Suetsugi, M., Fujimoto, M., et al. (2008). Characterization of the vulnerability to repeated stress in Fischer 344 rats: possible involvement of microRNA-mediated down-regulation of the glucocorticoid receptor. Eur. J. Neurosci. 27, 2250–2261. doi: 10.1111/j.1460-9568.2008.06218.x
Veenema, A. H., Koolhaas, J. M., and de Kloet, E. R. (2004). Basal and stress-induced differences in HPA axis, 5-HT responsiveness, and hippocampal cell proliferation in two mouse lines. Ann. N Y Acad. Sci. 1018, 255–265. doi: 10.1196/annals.1296.030
Vertes, R. P. (2004). Differential projections of the infralimbic and prelimbic cortex in the rat. Synapse 51, 32–58. doi: 10.1002/syn.10279
Vidal-Gonzalez, I., Vidal-Gonzalez, B., Rauch, S. L., and Quirk, G. J. (2006). Microstimulation reveals opposing influences of prelimbic and infralimbic cortex on the expression of conditioned fear. Learn. Mem. 13, 728–733. doi: 10.1101/lm.306106
Wang, G. Q., Cen, C., Li, C., Cao, S., Wang, N., Zhou, Z., et al. (2015). Deactivation of excitatory neurons in the prelimbic cortex via Cdk5 promotes pain sensation and anxiety. Nat. Commun. 6:7660. doi: 10.1038/ncomms8660
Wang, H., Hu, L., Liu, C., Su, Z., Wang, L., Pan, G., et al. (2016). 5-HT2 receptors mediate functional modulation of GABAa receptors and inhibitory synaptic transmissions in human iPS-derived neurons. Sci. Rep. 6:20033. doi: 10.1038/srep20033
Wood, M. D. (2003). Therapeutic potential of 5-HT2C receptor antagonists in the treatment of anxiety disorders. Curr. Drug Targets CNS Neurol. Disord. 2, 383–387. doi: 10.2174/1568007033482698
Keywords: breeding lines, freezing, contextual fear conditioning, elevated plus maze, serotonin, 5-HT2A receptors, medial prefrontal cortex
Citation: León LA, Castro-Gomes V, Zárate-Guerrero S, Corredor K, Mello Cruz AP, Brandão ML, Cardenas FP and Landeira-Fernandez J (2017) Behavioral Effects of Systemic, Infralimbic and Prelimbic Injections of a Serotonin 5-HT2A Antagonist in Carioca High- and Low-Conditioned Freezing Rats. Front. Behav. Neurosci. 11:117. doi: 10.3389/fnbeh.2017.00117
Received: 17 March 2017; Accepted: 31 May 2017;
Published: 07 July 2017.
Edited by:
Pedro Morgado, University of Minho, PortugalReviewed by:
Antoine Besnard, Massachusetts General Hospital, United StatesCopyright © 2017 León, Castro-Gomes, Zárate-Guerrero, Corredor, Mello Cruz, Brandão, Cardenas and Landeira-Fernandez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: J. Landeira-Fernandez, bGFuZGVpcmFAcHVjLXJpby5icg== orcid.org/0000-0002-8395-8008
† Karen Corredor orcid.org/0000-0001-5576-7784
† Fernando P. Cardenas orcid.org/0000-0002-8826-6211
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.