 Sophie M. Banas
Sophie M. Banas Silvina L. Diaz
Silvina L. Diaz Stéphane Doly
Stéphane Doly Arnauld Belmer
Arnauld Belmer Luc Maroteaux
Luc Maroteauxby Hertz, L., Rothman, D. L., Li, B., and Peng, L. (2015). Front. Behav. Neurosci. 9:25. doi: 10.3389/fnbeh.2015.00025
Working on the serotonin (5-hydroxytryptamine, 5-HT) 5-HT2B receptor since several years, we have read with high interest the review by Hertz et al. (2015). Previous studies from our group demonstrated that a direct injection in mouse raphe nucleus of the 5-HT2B agonist BW723C86 has the ability to increase extracellular levels of serotonin, which can be blocked by the selective 5-HT2B receptor antagonist RS127445 (Doly et al., 2008, 2009). We also reported that an acute injection of paroxetine 2 mg/kg in mice knocked out for the 5-HT2B receptor gene or in wild type mice injected with RS127445 (0.5 mg/kg) triggers a strong reduction in extracellular accumulation of 5-HT in hippocampus (Diaz et al., 2012). Following these observations, we showed that acute and chronic BW723C86 injection (3 mg/kg) can mimic the fluoxetine (3 mg/kg) and paroxetine (1 mg/kg) behavioral and biochemical antidepressant effects in mice (Diaz and Maroteaux, 2011; Diaz et al., 2012).
Hertz and coworkers used some of our published data on mice to support that fluoxetine and other SSRIs are acting as direct 5-HT2B receptor agonists independently of the serotonin transporter (SERT), based on their work on astrocytes. These authors forgot other important informations provided in Diaz's paper (Diaz et al., 2012), i.e., the absence of antidepressant effects of either fluoxetine or BW723C86 in mice lacking either the serotonin transporter (knockout for SERT) or differentiated serotonin neurons (knockout for Pet1). These data (i) rule out that the antidepressant effects of the 5-HT2B agonist BW723C86 or of fluoxetine could be independent of SERT; (ii) they indicate that serotonin neurons expressing SERT (and 5-HT2B receptors) are necessary for the 5-HT2B receptor effects independently of other cell types; (iii) they rule out the possibility that SSRIs mediate antidepressant effects only by stimulating directly putative astrocytic 5-HT2B receptors, which should be intact in these two mutant mice (SERT KO and Pet1 KO). A previous work (Launay et al., 2006) demonstrated a 5-HT2B receptor-mediated control of SERT activity in primary neurons from raphe nuclei, in which the BW723C86/5-HT2B receptor coupling promotes phosphorylations of SERT that can easily explain our findings in mice by a direct regulation of SERT by 5-HT2B receptors in a cell autonomous manner.
Furthermore, Hertz and coworkers state “The ability of all five currently used SSRIs to stimulate the 5-HT2B receptor equipotentially in cultured astrocytes has been known for several years.” This information originates from one of their previous observation on astrocyte cultures that “One micromolar of paroxetine, fluvoxamine or sertraline increased cPLA2a expression during chronic treatment; citalopram had a similar effect at 0.1–0.5 μM” (Zhang et al., 2010). Hertz and coworkers validated these data in one of their former review (Hertz et al., 2012), which reproduced their own earlier data (Kong et al., 2002), showing an half maximum effect (competition of mesulergine by fluoxetine on astrocytes) of 1 μM. However, in this review, they say that this initial calculation was “incorrect” and indicated that the Ki value of fluoxetine was in fact 70 nM on putative 5-HT2B astrocytic receptors (Hertz et al., 2012). These values, which are not supported by other published experimental evidence by these authors, are divergent from all published pharmacological values obtained with identified receptors: Bonhaus et al. (1997) and Knight et al. (2004) found Ki values for cloned human 5-HT2B receptors over 10 μM for fluoxetine, norfluoxetine and paroxetine (Table 1).
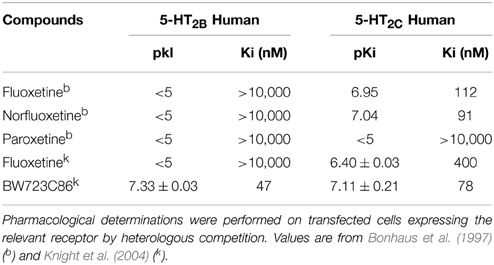
Table 1. Affinity constants (pKi) for different binding compounds to Human 5-HT2C and 5-HT2B receptors.
To sum up, it is clear from our experiments in mice that acute or chronic fluoxetine (and paroxetine) cannot act by a direct 5-HT2B receptor stimulation independently of SERT and serotonergic neurons. Furthermore, our pharmacological determination in mice is in accordance with affinity of SSRIs for human 5-HT2B receptors with Ki values over 5 μM (Diaz et al., 2012) and with no agonist activity (unpublished), while SSRI Ki values for SERT are in nanomolar range. Many SSRIs have published Ki values around 1 μM for muscarinic acetylcholine and histamine receptors or other monoamine transporters. The contribution of these other receptors/transporters in astrocytes could explain the findings of Hertz and coworkers independently of putative direct agonist effects at 5-HT2B receptors. Finally, the published evidence for 5-HT2B receptor expression in microglia (Krabbe et al., 2011; Kolodziejczak et al., 2015) add another level of complexity, as the concept of the tripartite synapse has recently been expanded to the monoaminergic systems to explain antidepressant drug responses (Quesseveur et al., 2013). A full set of research is still needed to understand the putative role for 5-HT in glial cells including astrocytes and/or microglia in respect to the relationship between SSRIs, serotonergic neurons and 5-HT2B receptors. The needs for further research is warranted to determine how this receptor subtype contribute to SSRI antidepressant effects.
Sources of Funding
Funding for this study was provided by the CNRS, the INSERM, the UPMC, and by grants from the Fondation de France, the Fondation pour la Recherche Médicale “Equipe FRM DEQ2014039529,” the French Ministry of Research (Agence Nationale pour la Recherche ANR-12-BSV1-0015-01 and the Investissements d'Avenir programme ANR-11-IDEX-0004-02). ML's team is part of the École des Neurosciences de Paris Ile-de-France network and of the Bio-Psy Labex. SD has been supported by fellowships from IBRO and then from Region Ile de France DIM STEM.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Bonhaus, D. W., Weinhardt, K. K., Taylor, M., Desouza, A., Mcneeley, P. M., Szczepanski, K., et al. (1997). RS-102221: a novel high affinity and selective, 5-HT2C receptor antagonist. Neuropharmacology 36, 621–629. doi: 10.1016/S0028-3908(97)00049-X
Diaz, S., Doly, S., Narboux-Nême, N., Fernandez, S., Mazot, P., Banas, S., et al. (2012). 5-HT2B receptors are required for serotonin-selective antidepressant actions. Mol. Psychiatry 17, 154–163. doi: 10.1038/mp.2011.159
Diaz, S., and Maroteaux, L. (2011). Implication of 5-HT2B receptors in the serotonin syndrome. Neuropharmacology 61, 495–502. doi: 10.1016/j.neuropharm.2011.01.025
Doly, S., Bertran-Gonzalez, J., Callebert, J., Bruneau, A., Banas, S. M., Belmer, A., et al. (2009). Role of serotonin via 5-HT2B receptors in the reinforcing effects of MDMA in mice. PLoS ONE 4:e7952. doi: 10.1371/journal.pone.0007952
Doly, S., Valjent, E., Setola, V., Callebert, J., Hervé, D., Launay, J. M., et al. (2008). Serotonin 5-HT2B receptors are required for 3,4-methylenedioxymethamphetamine-induced hyperlocomotion and 5-HT release in vivo and in vitro. J. Neurosci. 28, 2933–2940. doi: 10.1523/JNEUROSCI.5723-07.2008
Hertz, L., Li, B., Song, D., Ren, J., Dong, L., Chen, Y., et al. (2012). Astrocytes as a 5-HT2B-mediated SERT-independent SSRI target, slowly altering depression-associated genes and function. Curr. Signal Transduct. Ther. 7, 65–80. doi: 10.2174/1574362799278154
Hertz, L., Rothman, D. L., Li, B., and Peng, L. (2015). Chronic SSRI stimulation of astrocytic 5-HT2B receptors change multiple gene expressions/editings and metabolism of glutamate, glucose and glycogen: a potential paradigm shift. Front. Behav. Neurosci. 9:25. doi: 10.3389/fnbeh.2015.00025
Knight, A. R., Misra, A., Quirk, K., Benwell, K., Revell, D., Kennett, G., et al. (2004). Pharmacological characterisation of the agonist radioligand binding site of 5-HT2A, 5-HT2B and 5-HT2C receptors. Naunyn Schmiedebergs. Arch. Pharmacol. 370, 114–123. doi: 10.1007/s00210-004-0951-4
Kolodziejczak, M., Bechade, C., Gervasi, N., Irinopoulou, T., Banas, S. M., Cordier, C., et al. (2015). Serotonin modulates developmental microglia via 5-HT receptors: potential implication during synaptic refinement of retinogeniculate projections. ACS Chem. Neurosci. 6, 1219–1230. doi: 10.1021/cn5003489
Kong, E. K., Peng, L., Chen, Y., Yu, A. C., and Hertz, L. (2002). Up-regulation of 5-HT2B receptor density and receptor-mediated glycogenolysis in mouse astrocytes by long-term fluoxetine administration. Neurochem. Res. 27, 113–120. doi: 10.1023/A:1014862808126
Krabbe, G., Matyash, V., Pannasch, U., Mamer, L., Boddeke, H. W. G. M., and Kettenmann, H. (2011). Activation of serotonin receptors promotes microglial injury-induced motility but attenuates phagocytic activity. Brain Behav. Immun. 26, 419–428. doi: 10.1016/j.bbi.2011.12.002
Launay, J. M., Schneider, B., Loric, S., Da Prada, M., and Kellermann, O. (2006). Serotonin transport and serotonin transporter-mediated antidepressant recognition are controlled by 5-HT2B receptor signaling in serotonergic neuronal cells. FASEB J. 20, 1843–1854. doi: 10.1096/fj.06-5724com
Quesseveur, G., Gardier, A. M., and Guiard, B. P. (2013). The monoaminergic tripartite synapse: a putative target for currently available antidepressant drugs. Curr. Drug Targets 14, 1277–1294. doi: 10.2174/13894501113149990209
Zhang, S., Li, B., Lovatt, D., Xu, J., Song, D., Goldman, S. A., et al. (2010). 5-HT2B receptors are expressed on astrocytes from brain and in culture and are a chronic target for all five conventional 'serotonin-specific reuptake inhibitors'. Neuron Glia Biol. 6, 113–125. doi: 10.1017/S1740925X10000141
Keywords: serotonin transporter gene, serotonin, neurotransmitter reuptake, depression, mouse models, receptors, cell surface
Citation: Banas SM, Diaz SL, Doly S, Belmer A and Maroteaux L (2015) Commentary: Chronic SSRI stimulation of astrocytic 5-HT2B receptors change multiple gene expressions/editings and metabolism of glutamate, glucose and glycogen: a potential paradigm shift. Front. Behav. Neurosci. 9:207. doi: 10.3389/fnbeh.2015.00207
Received: 18 May 2015; Accepted: 20 July 2015;
Published: 05 August 2015.
Edited by:
Allan V. Kalueff, ZENEREI Institute, USA and Guangdong Ocean University, ChinaReviewed by:
Matthew O. Parker, Queen Mary University of London, UKTatyana Strekalova, Maastricht University, Netherlands
Copyright © 2015 Banas, Diaz, Doly, Belmer and Maroteaux. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luc Maroteaux,bHVjLm1hcm90ZWF1eEB1cG1jLmZy