
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Bee Sci. , 17 February 2025
Sec. Bee Protection and Health
Volume 3 - 2025 | https://doi.org/10.3389/frbee.2025.1510004
This article is part of the Research Topic Women in Bee Science View all 3 articles
 Patrícia Nunes-Silva1*
Patrícia Nunes-Silva1* André Luis Acosta2
André Luis Acosta2 Rafael Cabral Borges3
Rafael Cabral Borges3 Breno Magalhães Freitas4
Breno Magalhães Freitas4 Ricardo Caliari Oliveira5
Ricardo Caliari Oliveira5 Tereza Cristina Giannini3
Tereza Cristina Giannini3 Vera Lucia Imperatriz-Fonseca6
Vera Lucia Imperatriz-Fonseca6Introduction: Understanding how climate change affects the distribution of Amazonian bumblebee species is essential for their conservation and the pollination services they provide. This study focuses on two poorly known species, Bombus brevivillus and Bombus transversalis, evaluating how future climate scenarios may alter their suitable habitats in the Brazilian Amazon. Identifying potential refugia and vulnerable areas is crucial for developing targeted conservation strategies.
Methods: Species distribution models were applied using occurrence data from museum records and field collections. Climatic suitability was projected under the baseline period (1970–2000) and two future periods (2021–2040 and 2041–2060) using the high-emission scenario (SSP5-8.5) from the IPCC AR6 report. An ensemble modeling approach combining five different algorithms was used to predict areas of stability, habitat loss, and potential range expansion.
Results: By 2060, B. brevivillus is projected to lose 41.6% of its current suitable habitat, with significant reductions in northern and coastal regions. Conversely, B. transversalis is expected to retain 89.5% of its current range, showing a westward distribution shift. New climatically suitable areas may emerge for both species, particularly in the western Amazon, potentially serving as future refugia.
Discussion: The findings highlight species-specific responses to climate change, with B. brevivillus being more vulnerable than B. transversalis. These results emphasize the need for proactive conservation measures to protect critical habitats and mitigate the impacts of climate change. Future research should focus on assessing thermal tolerance and habitat connectivity to refine conservation strategies and ensure the persistence of these essential pollinators in changing environmental conditions.
Bumblebees (Bombus spp.) are widely distributed across Europe, America, and Asia, with the most remarkable diversity in temperate regions. Although they are primarily associated with cooler climates, some species are adapted to warmer areas, such as the Mediterranean region, the lowland tropics of Asia, and parts of Central and South America (Williams et al., 2008; Goulson, 2009). However, their species richness is the lowest in neotropical regions, including Brazil, which hosts only eight species: Bombus bahiensis Santos Júnior et al., 2015, Bombus bellicosus Smith, 1879, Bombus brasiliensis Lepeletier, 1836, Bombus brevivillus Franklin, 1913, Bombus morio (Swederus, 1787), Bombus pauloensis Friese, 1913, Bombus rubriventris Lepeletier, 1836 and Bombus transversalis (Olivier, 1789). None of these species are distributed across the entire Brazilian territory, and their populations are limited to specific habitats (Moure and Melo, 2023; Moure and Sakagami, 1962). While B. brevivillus and B. morio are entirely black, the other species have yellow pilosity in some areas of the body (Santos Júnior et al., 2015).
Although there are few species, the biology of tropical bumblebees is less well-studied compared to the species of temperate climates. This knowledge gap is partly due to the difficulty in locating their colonies or maintaining them in laboratory settings for extended periods (Garófalo, 2005; Oliveira et al., 2015). Additionally, these tropical bumblebees are much more aggressive than the ones from temperate climates, posing challenges to research efforts (Laroca, 1972, 1976; Garófalo, 2005; Oliveira et al., 2015). Notably, it has been reported that B. brevivillus, apart from stinging, may also engage in a defensive behavior spitting an unidentified substance that deters intruders, also hindering people´s affection for these large-body bees (Oliveira et al., 2015).
The nests are built in pre-existing underground cavities or on the ground, with or without a protective layer of litter or vegetation cut by the bumblebees, a variation that occurs between and within species (Laroca, 1972, 1976; Olesen, 1989; Taylor and Cameron, 2003; Oliveira et al., 2015). Bumblebee colonies in tropical regions generally follow an annual reproductive cycle similar to those of temperate species (Laroca, 1976; Oliveira et al., 2015; Paula and Melo, 2015). However, under favorable climatic conditions, it is also possible that the new gynes produced by colonies do not enter diapause, but start new colonies (Garófalo, 1979). Furthermore, the bumblebees can reactive the colonies at the end of each lifecycle, occasionally producing perennial nests (Garófalo, 1979; Oliveira et al., 2022; Garófalo, 2023). This phenomenon may be a survival strategy, enabling colonies to rebound quickly using remaining resources and workers (Oliveira et al., 2022).
The Brazilian species B. pauloensis and B. morio are better studied and primarily found in Southeast and Northeast Brazil, respectively (Garófalo, 2023). On the contrary, less information is available about B. brevivillus and B. transversalis, the only bumblebee species inhabiting the Brazilian Amazon (Moure and Sakagami, 1962; Santos Júnior et al., 2015; Françoso et al., 2016). This lack of information is particularly concerning because climate change is predicted to drastically impact the habitat suitability of various bee species in the eastern Amazon region in the coming decades (Giannini et al., 2020a). Furthermore, the predicted reduction of habitat due to climate change, in synergy with losses of land cover and in the size of protected areas, indicates that B. brevivillus is one of the most endangered species of bumblebees (Krechemer and Marchioro, 2020).
Given these vulnerabilities and their importance as pollinators for Brazilian agriculture (Giannini et al., 2020b), this study aims to evaluate the effects of climate change on the habitat suitability of two Amazonian bumblebee species, B. brevivillus and B. transversalis, using the most recent climate scenarios from the Sixth Assessment Report (AR6) of the Intergovernmental Panel on Climate Change (IPCC2023). By integrating current biological data and novel information on occurrence sites and plant interactions, this research seeks to provide a comprehensive understanding of the future distribution patterns of these species and identify potential climate refugia to inform conservation strategies.
Empirical data on the occurrence of B. transversalis and B. brevivillus was obtained from the following sources: (a) the speciesLink biodiversity database (http://splink.org.br/), (b) the Global Biodiversity Information Facility (GBIF, 2023), and (c) a newly compiled database with data from the entomological collections of the Museu Paraense Emilio Goeldi (MPEG) and the Bee Collection of the Federal University of Ceara, as well as recent fieldworks (from BFM). Records with inaccurate coordinates (e.g., missing numbers or signs), questionable locations (e.g., points over water bodies), and duplicate entries with identical coordinates for the species were excluded during the initial screening. The final dataset (“c”) was not applied in the modeling process, as it was obtained afterwards; it was used exclusively in subsequent analyses. The database on species occurrences and their respective pseudo-absences can be accessed in the Supplementary Material 1.
The latest IPCC report (IPCC-AR6, 2023) consolidated sixty phases of the Coupled Model Intercomparison Project (CMIP6) as the reference for Global Circulation Models (GCMs) for the following decades, improving the accuracy of climate effect projections with significant updates in emission trends and spatial distributions (Hausfather, 2019; McBride et al., 2021). Updated climatic variables directly influence analysis results on terrestrial species. This aspect highlights the need to renew or develop new climate suitability models for species to support precise decision-making on conservation, management and ecological restoration (McBride et al., 2021; Schramek, 2021).
Three sets of digital layers were acquired from WorldClim (Fick and Hijmans, 2017) at 2.5 arc minutes of resolution, each set representing 19 bioclimatic variables in different periods and altitudes. These bioclimatic variables capture geographic patterns of averages, seasonality, and extremes in precipitation and temperature, along with their combinations. The first set (baseline scenario) represents historical climate data for 1970-2000 based on interpolated empirical conditions (Hijmans et al., 2005). The remaining two sets are projections for future climate scenarios under climate change effects: 2021-2040 and 2041-2060. For both periods, the latest CMIP6-GCMs are produced by the Japanese Agency for Marine-Earth Science and Technology (MIROC6-GCM), based on the SSP585 scenario. All variables were cropped to the extent of South America, as both species are endemic to this neotropical region.
Among the Shared Socioeconomic Pathways (SSPs), the SSP5-8.5 represents the most intensive global greenhouse gas emission trajectories projected by AR6 (IPCC-AR6, 2023). While no scenario can be guaranteed as the definitive future climate outcome, SSP5-8.5 is considered the most appropriate for this study because considering the worst possible scenario will ensure that conservation actions will be most effective under any potential future conditions, including the most pessimistic climate outcome. The model’s predictive accuracy was enhanced by assessing the initial set of 19 bioclimatic variables from the baseline period (1970–2000) to identify and exclude those with high inter-correlation, as multicollinearity can negatively impact the predictive performance of regression-based models. A Variance Inflation Factor (VIF) analysis was performed using the `vifcor` function from the R package `usdm` (Naimi et al., 2014) to avoid deleterious multicollinearity effects on regression algorithms. The refined set of variables was subsequently used in the modeling process, with future climate variables selected accordingly to match the baseline set. The variables chosen for species after VIF can be accessed in the SM-B.
A habitat suitability approach was employed (Hirzel and Le Lay, 2008) using the Biomod2 package version 3.5.1 (Thuiller et al., 2021) in R (R Core Team, 2021) to map the climatic suitability of the species under the baseline conditions (1970-2000). Five algorithms were selected based on their high performance in similar studies (Aguirre-Gutiérrez et al., 2013; Acosta et al., 2016): GLM – Generalized Linear Model (glm package; Hastie and Pregibon, 1992), GBM – Generalized Boosting Model (gbm package; Greenwell et al., 2020), GAM – Generalized Additive Model (mgcv package; Hastie and Tibshirani, 1990), RF – Random Forest (randomForest package; Breiman, 2001), and MAXENT – Maximum Entropy (maxent package; Phillips et al., 2021). Algorithm parameterization followed recommendations from the literature (Guisan and Thuiller, 2005; Phillips et al., 2006; Thuiller et al., 2009; Thuiller et al., 2021).
This modeling approach requires presence and absence data; however, confirmed absence data for the target species were unavailable. To address this, five pseudo-absence datasets, each containing ten times the number of presence points (as suggested by Chefaoui and Lobo (2008) for improved predictive performance), were generated randomly outside predefined exclusion zones (20 km buffer radius) around presence points. These buffers represent twice the estimated maximum dispersal distance of individuals from their colonies (Borges et al., 2020), ensuring that pseudo-absences were not placed within these areas.
Each modeling run involved randomly splitting the presence dataset into 80% for training and 20% for evaluation using True Skill Statistics (TSS; Allouche et al., 2006). This random division and replacement of presence and pseudo-absence datasets were repeated for each run. 125 models were generated for each Bombus species under the baseline scenario, resulting from the combination of five algorithms, five pseudo-absence datasets, and five random partitions of the presence data (5 algorithms × 5 pseudo-absences × 5 runs = 125 models).
Only the models with the highest predictive performance (TSS > 0.8) were retained for further analysis. These top-performing models were used to construct a Baseline Ensemble Forecast Model (BEFM) using the Committee Averaging method, which converts probabilistic predictions into binary outcomes by maximizing specificity and sensitivity thresholds and then averaging them (Wisz et al., 2008; Hao et al., 2019; Thuiller et al., 2021). The selected models to build the BEFM were also projected onto future climate scenarios using the Biomod2 projection function and ensembled. Each ensemble forecast model (baseline, 2040, and 2060) was subsequently reclassified in binary values for further comparison: areas with suitability values ≥ 75% in continuous models were reclassified as binary=1 (suitable), and regions with < 75% were reclassified as binary=0 (unsuitable).
We then generated delta models by comparing the baseline with each future scenario, achieved by concatenating binary values side-by-side. This approach allowed us to classify four categories: (A) consistently suitable (potential climate sanctuary and/or refuge) - areas currently suitable (baseline zones with value = 1) that remain suitable (value = 1) in future scenarios (delta value = 11); (B) loss of suitability - areas currently suitable (baseline value = 1) that become unsuitable (value = 0) in future scenarios (delta value = 10); (C) gain of suitability (potential climate refuge) - areas currently unsuitable (baseline value = 0) that become suitable (value=1) in future scenarios (delta value = 01); and (D) consistently unsuitable - areas consistently unsuitable (baseline value = 0) that remain unsuitable (value=0) in future scenarios (combined value = 00). It is important to note that the calculation of areas lost in terms of climatic suitability is not offset by areas gained, as there is no guarantee that the species will be able to occupy these new areas in future. This is due to non-climatic factors not accounted for in the model, such as dispersal capacity relative to the pace of climate change, the availability of food resources and shelter, competition from other species, diseases, and the lack of ecological/landscape connectivity between currently suitable patches and new projected suitable zones in the future. For details on modeling outputs, parameters and input data, see Supplementary Material 1.
The suitable climate range of B. transversalis is expected to significantly change the geospatial distribution of suitability levels over the coming decades (2040, 2060) within its near-current range (Figure 1A). A visual comparison between the continuous models from the baseline to future scenarios (Figures 1A–C) indicates that, while there is a reduction in suitability in marginal areas to the southeast of the Amazon Biome, there is a concurrent shift in suitability towards the northwest and west. This shift is accompanied by increased homogeneity in the coverage of areas with higher suitability.
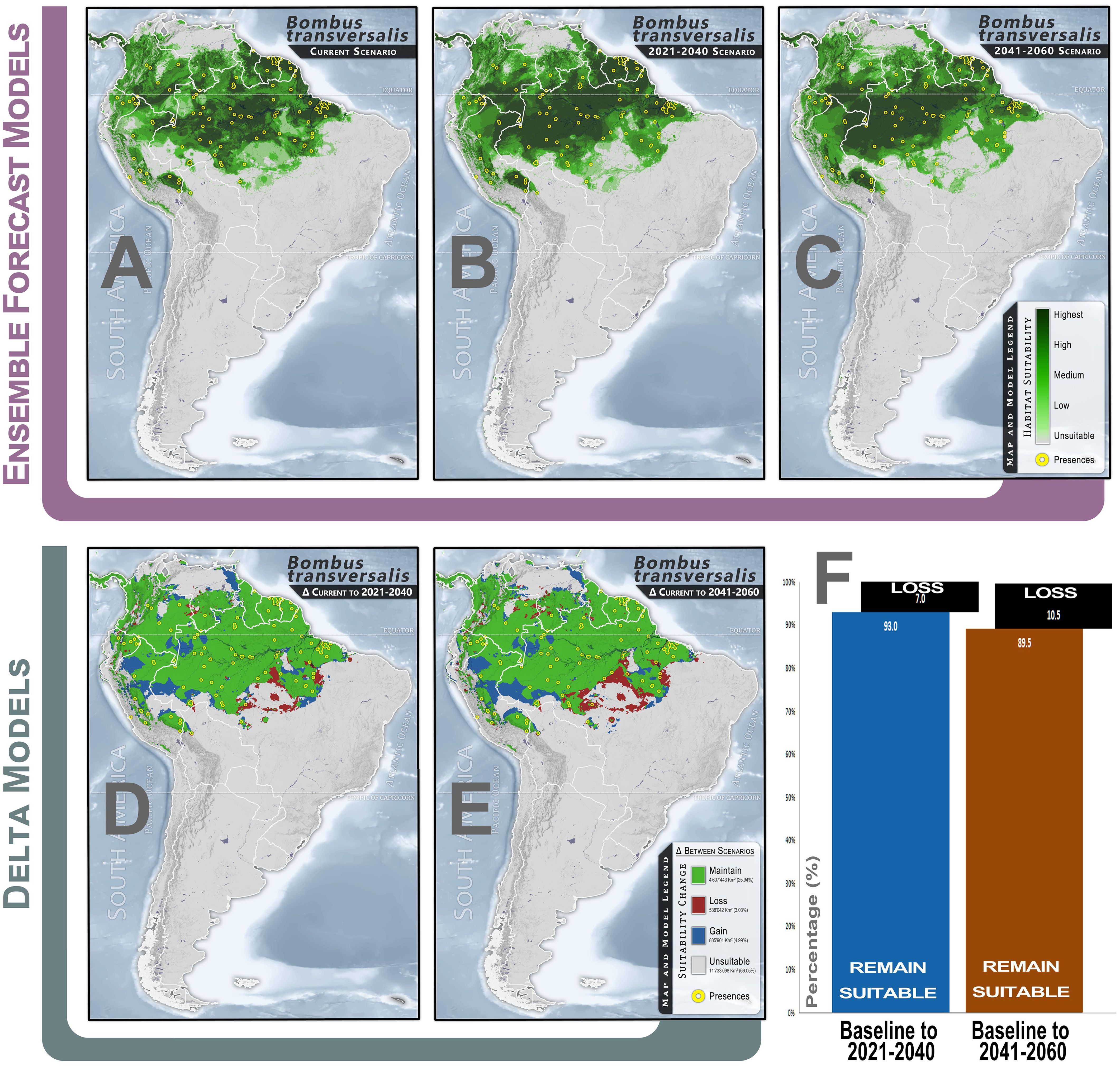
Figure 1. Climate habitat suitability and climate change effects for Bombus transversalis. (A) Continuous climate suitability for the species in the baseline scenario, considering near-current climate conditions (1970-2000); (B) Continuous climate suitability projection based on MIROC6-SSP-858 GCMs for 2021-2040; (C) Continuous climate suitability projection based on MIROC6-SSP-858 GCMs for 2041-2060; (D) Delta model comparing changes from baseline to the 2021-2040 scenario; (E) Delta model comparing changes from baseline to the 2041-2060 scenario; (F) Bar plot showing the proportion (%) of changes (loss and maintenance) in the climatically suitable range for the species.
The models predict a 7% reduction in climatically suitable habitat extension for B. transversalis from the baseline period (1970–2000) to the 2021–2040 scenario and a 10.5% from baseline to the 2041–2060 scenario. In all future scenarios, consistent habitat suitability losses are detected for this species, predominantly in the southeastern region of the Amazon biome (Figures 1D–F; Supplementary Table S1). These losses are relatively minor in area compared to the regions that maintain suitability. An immense suitable zone of species’ climate suitability will persist, stretching from the Atlantic coast to the Andean Mountain ranges. New suitable areas, mainly concentrated in the western Amazon and near the Andean region, are projected to emerge. This matches the prediction that the species’ range will shift westward.
Habitats with the highest levels of climate suitability for B. brevivillus (dark green zones) were detected primarily in spots concentrated in northern and northeastern Brazil, with a relatively wide but scattered distribution across several other parts of the continent (Figure 2A). The species is mainly found in coastal zones (Mata Atlantica Biome), large river basins (riparian forests and surroundings), and areas of relatively higher altitudes, notably plateaus and highlands (Araripe plateau, Ibiapaba plateau, Mirador plateau, Meruoca mountains, Baturité mountains and Carajás mountain range).
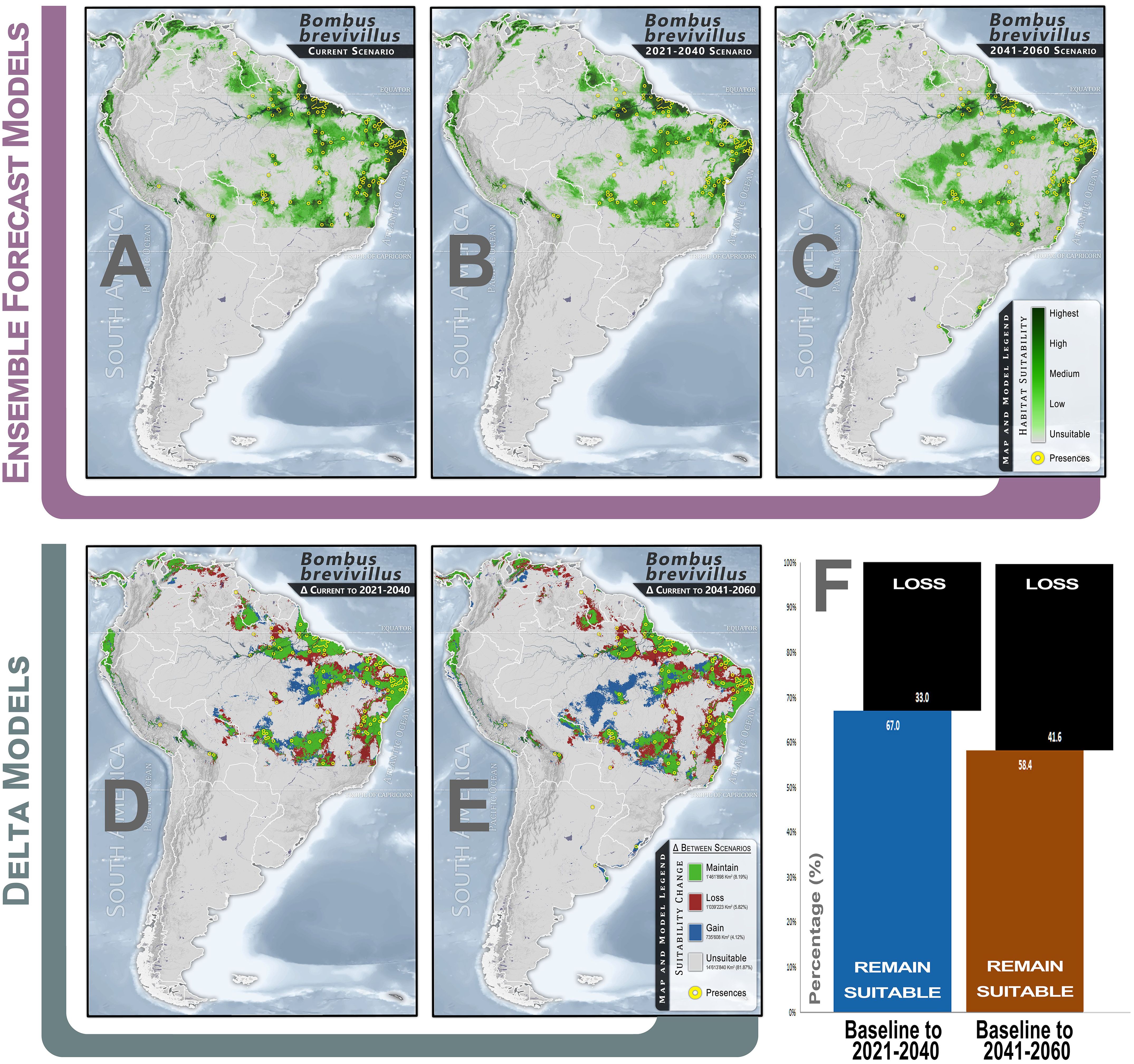
Figure 2. Climate habitat suitability and climate change effects for Bombus brevivillus. (A) Continuous climate suitability for the species in the baseline scenario, considering near-current climate conditions (1970-2000); (B) Continuous climate suitability projection based on MIROC6-SSP-858 GCMs for 2021-2040; (C) Continuous climate suitability projection based on MIROC6-SSP-858 GCMs for 2041-2060; (D) Delta model comparing changes from baseline to the 2021-2040 scenario; (E) Delta model comparing changes from baseline to the 2041-2060 scenario; (F) Bar plot showing the proportion (%) of changes (loss and maintenance) in the climatically suitable range for the species.
For the coming decades, significant shifts will occur in the distribution of currently suitable areas (Figures 2A–C), with a decrease in suitability levels in northern and coastal zones (evidenced by the decrease in green intensity across scenarios) and an expansion of suitability coverage towards the eastern regions. The models detected a 33% reduction in the extent of climatically suitable habitats from the baseline to the 2021-2040 scenario and a 41.6% reduction for the 2041-2060 (Figures 2D–F; Supplementary Table S1). Losses will be more pronounced in the marginal zones of the baseline suitable areas, indicating a contraction of the overall suitable extent (Red zones in Figures 2D, E). However, gain zones were also detected in the most central areas of the climatically suitable distribution area for the species, although it cannot be guaranteed that the species will be able to occupy these zones in the future. (Blue zones in Figures 2D, E).
Overall, we found 206 B. transversalis and 388 specimens of Bombus brevivilus in the entomological collections (MPEG and CAUFC). Bees from both species have been collected in all 12 months of the year from 1877 to 2024. Although both species appear to be active throughout the year, more than 65% of the B. brevivilus specimens were collected in the first five months of the year (January to May), 30% of individuals were collected between June and October and about 4% were collected between November and December (Figure 3). In a dissimilar trend, 29% of the B. transversalis specimens were collected between January and May, more than 62% were collected between June and October, and 5% of the bees were collected between November and December (Figure 3A).
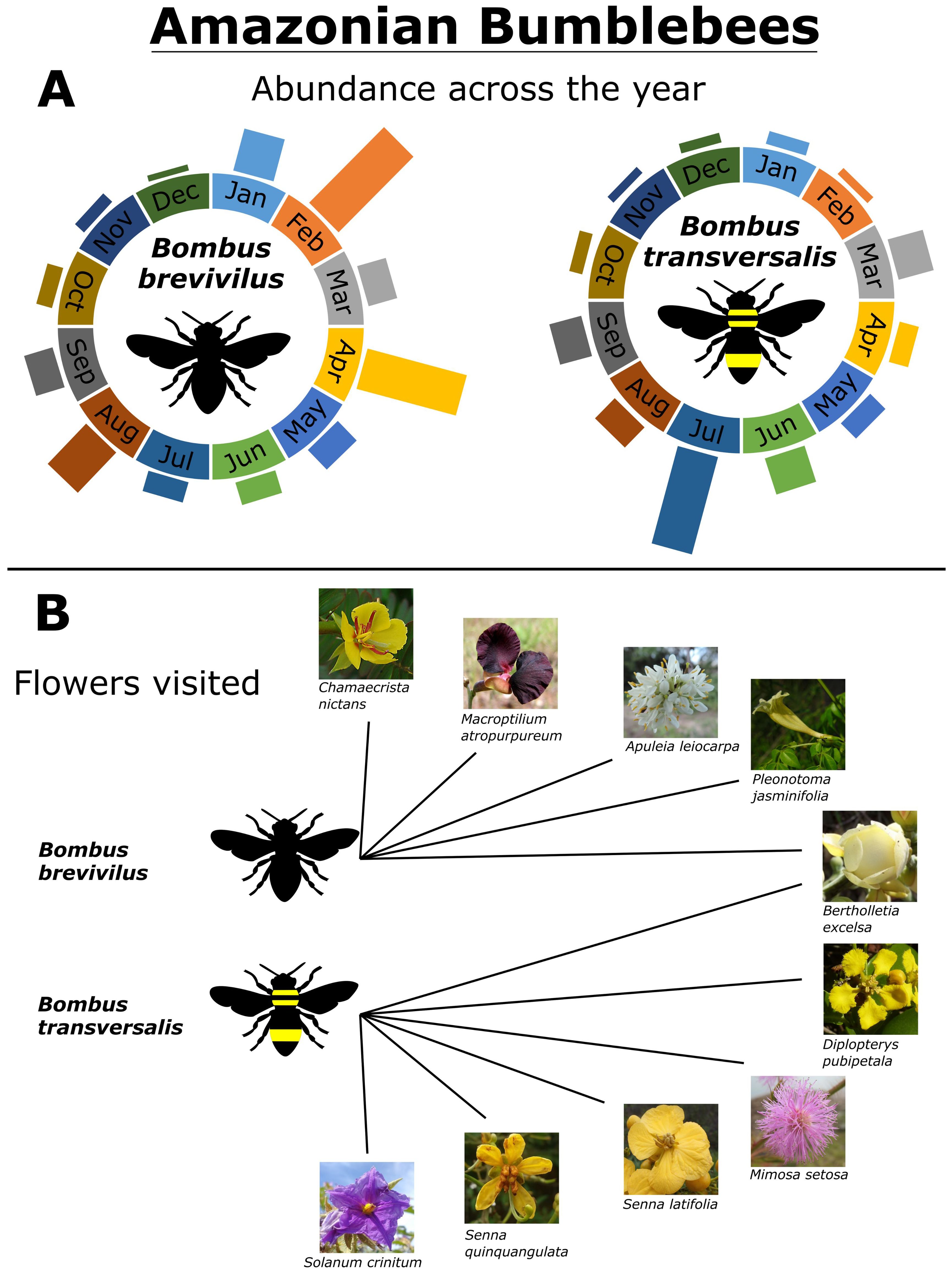
Figure 3. Amazonian bumblebees (Bombus brevivillus and B. transversalis). (A) Relative abundance of Amazonian bumblebees according to the historical data from entomological collections (MPEG and CAUFC). The total abundance of B. brevivillus is 383 specimens, B. transversalis 206 specimens. (B) Amazonian bumblebee interactions with local flowers. Data was obtained from specimen labels and the analysis of the pollen grains adhered to the specimen body (Romeiro et al., 2023).
Although most of the specimens found in the collections were probably collected when interacting with flowers, these are not described in the specimens´ labels. Only specimens collected between 2018 and 2024 provide an identification of the plants visited. The two species interacted with ten plant species (Figure 3B), all plants with flowers with matching sizes and exposed rewards.
The models predicted that both B. brevivillus and B. transversalis will experience shifts in their geographic distribution due to climate change in future decades, with variations in the extent of suitable habitats over time. By 2060, B. brevivillus is expected to lose more than 40% of its suitable climate zone extent, while B. transversalis will maintain its large climatically suitable zone, with limited losses (10.5%) and a westward shift. Previous predictions have suggested that B. brevivillus may lose between 57.2% to 67.2% of climatically suitable habitat by 2050, and between 66% and 78.4% by 2070, depending on the scenario (Krechemer and Marchioro, 2020). Our results indicate lower losses, likely due to the use of updated future climate scenarios (IPCC-AR6, 2023). In contrast, B. transversalis is projected to maintain a significant portion of its current habitat and shift its range westward, suggesting potential stability in its core distribution (Krechemer and Marchioro, 2020). The limited losses predicted for B. transversalis are consistent with previous studies that estimated a 9.1% to 19% decrease in climatically suitable areas by 2050 and 20% to 24.7% by 2070 (Krechemer and Marchioro, 2020). This relative stability is likely due to the predicted climate stability for the central and western portions of the Brazilian Amazon (Bottino et al., 2024), which also supports the observed westward shift.
Regarding changes in geographic distribution, B. brevivillus will undergo significant range shifts: while expanding into eastern regions, northern and coastal zones will experience a drastic reduction in suitability levels. Krechemer and Marchioro (2020) also predicted this eastward shift of distribution. Meanwhile, B. transversalis is projected to expand westward into newly suitable areas, potentially using these regions as future climate refugia. However, whether B. transversallis can effectively colonize these areas depends on other limiting ecological and environmental factors not accounted for in this study. The reduction in suitability along the southeastern margins of its range within the Amazon Biome, as observed here, aligns with the predictions of Krechemer and Marchioro (2020), who predicted that B. transversalis would lose regions that are climatically suitable in the southern range of its distribution.
Climate change impacts on four other South American bumblebee species have been predicted by Krechemer and Marchioro (2020): Bombus bellicosus, Bombus brasiliensis, Bombus morio and Bombus pauloensis. The expected effect varied with the species, but B. bellicosus, B. brevivillus and B. brasiliensis were considered the most vulnerable due to significant habitat losses, limited protected areas, and habitat fragmentation (Krechemer and Marchioro, 2020). Similarly, Martins et al. (2015) have also pointed out the vulnerability of B. bellicosus because of climate change, among other factors. Losses in climatically suitable areas have also been predicted for B. morio (Elias et al., 2017; Françoso et al., 2019) and B. pauloensis (Françoso et al., 2019). In the Neotropics, changes in distribution range were also predicted for Bombus funebris in the western region of South America (Nascimento et al., 2022). Populations of this species are expected to become restricted to high-altitude areas in the Andean region by 2070 (Nascimento et al., 2022).
In contrast, climate change does not appear to play a significant role in the decline of the southernmost bumblebee, Bombus dahlbomii and will probably have a small impact on its geographic range, which may decrease by 13% to 14% between 2080 and 2100 due to this stressor alone (Morales et al., 2022). These contrasting responses highlight that species-specific traits, such as thermal tolerance, nesting behavior, ecological interactions, and flexibility, influence how different bumblebee species respond to changing climates.
As discussed above, species-specific responses to climate change have been reported for other bumblebee species (Jackson et al., 2022; Singh et al., 2023), often linked to differences in their ecology and physiology. There is both inter- and intraspecific variation in heat resistance among bumblebees (Martinet et al., 2021). Intraspecific variation can occur between populations in different geographic areas (Oyen et al., 2016; Martinet et al., 2021), but is also species-specific and associated with geographically widespread species (Martinet et al., 2021). Furthermore, Martinet et al. (2021) pointed out that bumblebees are generally more tolerant to cold than to heat, and the capacity to tolerate extreme temperatures may limit their distributions, which affect, at least partially, species population trends for expansion or contraction. However, existing studies do not include tropical bumblebee species. B. brevivillus and B. transversalis may be more tolerant to heat than bumblebees from temperate and polar regions, and there may be interspecific variation in heat tolerance that could explain the difference in the predicted range shifts and suitable habitat losses and gains. Studying the thermal tolerance of those bumblebees would contribute to understanding and making more precise predictions of the effects of climate change on them.
Nest temperature and architecture may also play a role in the species-specific responses of bumblebees to climate change. Recently, Kevan et al. (2024) emphasized that the nest temperature, especially in and around it, are critical factors in comprehending the consequences of heat stress on bumblebees, but nest architecture and the thermal characteristics of nest substrates have yet to be studied. Their review indicated that, in general, the optimum temperature for brood rearing ranges from 28 to 32°C for species that live from the cold High Arctic to tropical environments, and it is conserved among bumblebee species. However, the authors have found data on only around 25 bumblebee species in 20 studies (Kevan et al., 2024), and among them, the only bumblebee from South America was Bombus atratus (now B. pauloensis, Moure and Melo, 2023) (Vega et al., 2011).
Although B. transversalis was not included in the review, Taylor and Cameron (2003) measured the internal temperature of four nests and found a mean of 30.1°C, which is around 4.5°C warmer than the ambient temperature. The nest humidity and temperature were maintained regardless of ambient conditions, which was attributed to the insulating nest cover (built with leaves and pieces of wood) and the fanning behavior of workers under direct sunlight on the nest (Taylor and Cameron, 2003). Still, no experimental study on the role of these two factors has been conducted. Furthermore, fanning may not be effective under extreme temperatures. Although there may be differences between tropical and temperate species, the temperate bumblebee B. impatiens fails to maintain nest temperature even by increasing fanning at 35°C and workers abandon the colony at this temperature. This suggests that heatwaves exceeding 30°C negatively impact on colony health (Bretzlaff et al., 2024).
The nests of B. transversalis are built on the ground surface, on a pile of leaves and sticks, and the bumblebees use vegetation (cut woven leaven) to cover it (Taylor and Cameron, 2003). B. brevivillus also constructs its nests on the ground surface but nests underground, covering the nests with vegetation (Oliveira et al., 2015). A thin layer of wax is constructed over the brood area (Oliveira et al., 2015), which is not frequent for B. transversalis, that may occasionally construct a thin layer of wax that covers part of the brood area (Taylor and Cameron, 2003). The vegetation cover (as also suggested by Taylor and Cameron, 2003) and the layers of wax and old brood cells may act in passive thermoregulation. Thus, investigating these mechanisms is crucial to predict how these species will respond to heat stress and climate change.
Furthermore, there is an urgent need to study the thermal tolerance of bumblebee species, to understand and predict the effects of climate change more precisely, in particular of South American bumblebees. The few species of bumblebees (B. terrestris and B. impatiens) tested for thermal tolerance were not capable of acclimation after prolonged heat and heat waves, decreasing their ability to cope with heat stress (Oyen and Dillon, 2018; Quinlan et al., 2023; Sepúlveda and Goulson, 2023). In addition, nutritional stress (e.g. starvation) lowers the survival of B. impatiens after heat stress compared to bumblebees with access to food, indicating that nutrition directly affects heat tolerance (Quinlan et al., 2023; but see Oyen and Dillon, 2018). In summary, heat stress can negatively affect survival, foraging behavior, reproduction, colony development, and fertility (White and Dillon, 2023). For example, thermal stress affects the viability and integrity of the DNA of the sperm of the cold-adapted species like Bombus magnus and Bombus jonellus (both declining), but not of the warm-adapted and widespread B. terrestris (Martinet et al., 2021).
Species-specific responses to extreme temperatures and thermal tolerances must be further investigated, as they drive changes in bumblebee community composition. These changes occur as some species’ populations increase while others decline, leading to shifts in population ranges (e.g. Kerr et al., 2015; Martinet et al., 2015; Arnóczkyné Jakab et al., 2023). For instance, some species’ geographic distribution may change as a result of warmer winters (Biella et al., 2021). Unfortunately, monitoring of bumblebee communities in the Neotropical region, especially in the Amazon region, is virtually nonexistent. This lack of data makes it impossible to track population changes, highlighting the urgent need for year-round monitoring.
This monitoring is urgent and should be carried out throughout the year because both B. brevivillus and B. transversalis have been observed throughout the year, although their activity peaks at specific times (B. brevivilus: January to May; B. transversalis: June to October). The colony cycle of B. transversalis aligns closely with the wet-dry seasons in the Amazon: colonies establish during the wet season and mature in the dry season. Traditional communities have noted that most nests in pristine areas appear perennial, as new queens reuse old nest structures rather than building new ones (Taylor and Cameron, 2003). However, this trait could make this species vulnerable under global changes scenarios. Nest construction involves significant investment, and the inability to establish new nests in response to climate and land use change may threaten their populations. Similarly, B. brevivillus, exhibits a colony cycle where, during periods of high worker population, colonies prioritize producing males and gynes, which will disperse in search of mates just before and during the characteristic colony collapse observed in Bombus colonies (Oliveira et al., 2015). Reactivation of B. brevivillus nests have been observed (Oliveira et al., 2022), possibly as a survival strategy to avoid predation. Reactivated nests retain some workers and resources, allowing for rapid colony growth and the early establishment of a defensive worker force.
Landscape also influences the distribution of bumblebee species and the composition of their communities; thus, land use should also be considered in the planning of bumblebee conservation strategies along with climate, and, when possible, their effects should be evaluated together (Christman et al., 2022). Colonies of B. terrestris, for example, produce more individuals as temperature increases, however, only when there is high availability of resources (Zaragoza-Trello et al., 2021). When the resource availability is low, the number of queens and males produced by colonies decreases, but not of workers, showing that the effect of climate interacts with resource availability and varies with caste (Zaragoza-Trello et al., 2021). Therefore, although producing more workers may be a strategy to survive under low resource availability, ensuring enough floral resources for colonies is essential for minimizing the effects of climate change (Zaragoza-Trello et al., 2021). For B. brevivillus and B. transversalis, further research is needed to identify the plants they rely on for foraging, as few records exist on their floral preferences.
Moreover, climate change may influence bumblebees by changing habitats, which should also be considered in the predictions. In Belgium, for example, the probability of species decline was linked to the kind of habitat preferred by the species, and species that choose to live in open habitats presented a higher chance to decline than species preferring forests (Rollin et al., 2020). This difference may explain why B. brevivillus appears more vulnerable than B. transversalis. While B. brevivillus occupies both forests and open habitats (Oliveira et al., 2015), B. transversalis constructs its nests on terra firme in the Amazon Forest (Olesen, 1989; Taylor and Cameron, 2003).
Unfortunately, besides the threats from climate change and deforestation reducing its suitable areas, B. brevivillus also faces the direct action of humans that usually destroy its colonies as soon as they are found in precaution against the highly defensive behavior shown by this species. This is markedly recurrent when nests are built close to urban areas and agricultural settings, especially banana and coffee plantations (Oliveira et al., 2015). Considering this scenario and the results reported in this study, one question is if there will be losses of the pollination services provided by B. brevivillus.
Conversely, B. transversalis may become an essential pollinator in the coming decades. Bumblebee species are important pollinators of commodities such as the Brazil nut (Cavalcante et al., 2018); with a large body size and long flight distance (Borges et al., 2020), efforts for the conservation of pollination services should consider the maintenance of protected areas for supporting bumblebee populations (Freitas et al., 2014). This is particularly important in that, at least in the Eastern Amazon (Carajás Forest), it is predicted that the populations of 95% of bee species will suffer a decrease in occurrence area (Giannini et al., 2020a). Climate change will potentially have negative consequences for crop pollination in Brazil (Elias et al., 2017; Giannini et al., 2020a; Sales et al., 2021), thus predicting the effects of climate change on bees, which are the leading group of crop pollinators in the country (Oliveira et al., 2024), is essential to conservation management policies. In conclusion, both B. brevivillus and B. transversalis are predicted to experience range shifts and habitats losses by 2060. These changes may reduce their geographic overlap and are likely tied to species-specific traits that require further investigation. Therefore, scientifically accurate predictions are essential for developing conservation and management plans to ensure the survival of these species and the pollination services they provide.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The manuscript presents research on animals that do not require ethical approval for their study.
PN-S: Visualization, Writing – original draft, Writing – review & editing. AA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. RB: Formal analysis, Visualization, Writing – original draft, Writing – review & editing. BF: Data curation, Funding acquisition, Resources, Writing – original draft, Writing – review & editing. RO: Conceptualization, Formal analysis, Investigation, Methodology, Writing – review & editing. TG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Writing – review & editing. VI-F: Conceptualization, Data curation, Funding acquisition, Methodology, Project administration, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. We thank Instituto Tecnológico Vale and Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq (process number 4443.384/2018-9) for financial support. BF was funded by CNPq (process number 308358/2019-8) and a cooperation agreement between BAYER AG. (Germany) and the Federal University of the State of Ceara (Brazil) on the Project “Breeding, multiplication and handling of native pollinators for crop pollination in Brazil”. CNPq also funded VLIF (315957/2023-9). RO is a Serra Húnter professor.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frbee.2025.1510004/full#supplementary-material
Acosta A. L., Giannini T. C., Imperatriz-Fonseca V. L., Saraiva A. M. (2016). Worldwide alien invasion: a methodological approach to forecast the potential spread of a highly invasive pollinator. PloS One 11 (1), e0148295. doi: 10.1371/journal.pone.0148295
Aguirre-Gutiérrez J., Carvalheiro L. G., Polce C., van Loon E. E., Raes N., Reemer M., et al. (2013). Fit-for-purpose: Species distribution model performance depends on evaluation criteria – dutch hoverflies as a case study. PloS One 8 (5), e63708. doi: 10.1371/journal.pone.0063708
Allouche O., Tsoar A., Kadmon R. (2006). Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43 (6), 1223–1232. doi: 10.1111/j.1365-2664.2006.01214.x
Arnóczkyné Jakab D., Tóth M., Szarukán I., Szanyi S., Józan Z., Sárospataki M., et al. (2023). Long-term changes in the composition and distribution of the Hungarian bumble bee fauna (Hymenoptera, Apidae, Bombus). J. Hymenopt. Res. 96, 207–237. doi: 10.3897/jhr.96.99002
Biella P., Ćetković A., Gogala A., Neumayer J., Sárospataki M., Šima P., et al. (2021). Northwestward range expansion of the bumblebee Bombus haematurus into Central Europe is associated with warmer winters and niche conservatism. Insect Sci. 28, 861–872. doi: 10.1111/1744-7917.12800
Borges R. C., Padovani K., Imperatriz-Fonseca V. L., Giannini T. C. (2020). A dataset of multi-functional ecological traits of Brazilian bees. Sci. Data 7, 120. doi: 10.1007/s10905-015-9502-8
Bottino M. J., Nobre P., Giarolla E., et al. (2024). Amazon savannization and climate change are projected to increase dry season length and temperature extremes over Brazil. Sci. Rep. 14, 5131. doi: 10.1038/s41598-024-55176-5
Breiman L. (2001). In Machine Learning Vol. 45. (Berlim/Heidelberg, Germany), 5–32. doi: 10.1023/a:1010933404324
Bretzlaff T., Kerr J. T., Darveau C. A. (2024). Handling heatwaves: balancing thermoregulation, foraging and bumblebee colony success. Conserv. Physiol. 12, 1. doi: 10.1093/conphys/coae006
Cavalcante M. C., Galetto L., Maués M. M., Pacheco A. J. S., Bomfim I. G. A., M. Freitas B. M. (2018). Nectar production dynamics and daily pattern of pollinator visits in Brazil nut (Bertholletia excelsa Bonpl.) plantations in Central Amazon: implications for fruit production. Apidologie 49, 505–516. doi: 10.1007/s13592-018-0578-y
Chefaoui R. M., Lobo J. M. (2008). Assessing the effects of pseudo-absences on predictive distribution model performance. Ecol. Model. 210, 478–486. doi: 10.1016/j.ecolmodel.2007.08.010
Christman M. E., Spears L. R., Strange J. P., Pearse W. D., Burchfield E. K., Ramirez R. A. (2022). Land cover and climate drive shifts in Bombus assemblage composition. Agric. Ecosyst. Environ. 339, 108113. doi: 10.1016/j.agee.2022.108113
Elias M. A. S., Borges F. J. A., Bergamini L. L., Franceschinelli E. V., Sujii E. R. (2017). Climate change threatens pollination services in tomato crops in Brazil. Agric. Ecosyst. Environ. 239, 257–264. doi: 10.1016/j.agee.2017.01.026
Fick S. E., Hijmans R. J. (2017). WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatology 37 (12), 4302–4315. doi: 10.1002/joc.5086
Françoso E., Zuntini A. R., Arias M. C. (2019). Combining phylogeography and future climate change for conservation of Bombus morio and B. pauloensis (Hymenoptera: Apidae). J. Insect Conserv. 23, 63–73. doi: 10.1007/s10841-018-0114-4
Françoso E., de Oliveira F. F., Arias M. C. (2016). An integrative approach identifies a new species of bumblebee (Hymenoptera: Apidae: Bombini) from northeastern Brazil. Apidologie 47, 171–185. doi: 10.1007/s13592-015-0385-7
Freitas B. M., Pacheco Filho A. J., Andrade P. B., Lemos C. Q., Rocha E. E. M., Pereira N. O., et al. (2014). Forest remnants enhance wild pollinator visits to cashew flowers and mitigate pollination deficit in NE Brazil. J. Poll. Ecol. 12, 22–30. doi: 10.26786/1920-7603(2014)10
Garófalo C. A. (1979). Observações preliminares sobre a fundação solitária de colônias de Bombus (Fervidobombus) atratus Franklin (Hymenoptera, Apidae). Boletim de Zoologia Univ. São Paulo 4, 53–64. doi: 10.11606/issn.2526-3358.bolzoo.1979.121826
Garófalo C. A. (2005). Bombus: as mamangavas de chão e sua importância como agentes polinizadores. Available online at: http://www.apacame.org.br/mensagemdoce/80/msg80.html (Accessed March 25, 2011).
Garófalo C. A. (2023). “Padrões comportamentais de rainhas de Bombus (Thoracobombus) morio Swederus, (1787) (Hymenoptera, Apidae),” in Estudos sobre abelhas e vespas brasileiras [livro eletrônico]: uma homenagem ao Professor Lúcio Campos. Eds. Resende H. C., Werneck H. A. (Laboratório de Genética da Conservação de Abelhas, Universidade Federal de Viçosa, Florestal, MG), 101–108.
GBIF. (2023). The global biodiversity information facility. Available online at: https://www.gbif.org/. (Accessed January 2023).
Giannini T. C., Alves D. A., Alves R., Cordeiro G. D., Campbell A. J., Awade M., et al. (2020b). Unveiling the contribution of bee pollinators to Brazilian crops with implications for bee management. Apidologie 51, 406–421. doi: 10.1007/s13592-019-00727-3
Giannini T. C., Costa W. F., Borges R. C., Miranda L., da Costa C. P. W., Saraiva A. M., et al. (2020a). Climate change in the Eastern Amazon: crop-pollinator and occurrence-restricted bees are potentially more affected. Reg. Environ. Change 20, 1. doi: 10.1007/s10113-020-01611-y
Goulson D. (2009). Bumblebees: behaviour, ecology, and conservation (New York: Oxford University Press).
Greenwell B., Boehmke B., Cunningham J., GBM-Developers. (2020). gbm Generalized Boosted Regression Models. Available at: https://cran.rproject:web/packages/gbm/index.html.
Guisan A., Thuiller W. (2005). Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 8, 993–1009. doi: 10.1111/j.1461-0248.2005.00792.x
Hao T., Elith J., Guillera-Arroita G., Lahoz-Monfort J. J. (2019). A review of evidence about use and performance of species distribution modeling ensembles like BIOMOD. Diversity Distributions 25, 839–852. doi: 10.1111/ddi.12892
Hastie T. J., Pregibon D. (1992). “Generalized linear models,” in Statistical Models S (Chapter 6). Eds. Chambers M., Hastie T. J. (Pacific Grove, CA, U.S.A: Wadsworth & Brooks/Cole).
Hastie T. J., Tibshirani R. J. (1990). “Generalized additive models.” in Chapman & Hall/CRC Monographs on Statistics & Applied Probability, Vol. 43. (Boca Raton, FL, U.S.A).
Hausfather Z. (2019). CMIP6: the next generation of climate models explained. Available at: www.carbonbrief:cmip6-the-next-generation-of-climate-models-explained (Accessed January 2023).
Hijmans R. J., Cameron S. E., Parra J. L., Jones P. G., Jarvis A. (2005). Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978. doi: 10.1002/joc.1276
Hirzel A. H., Le Lay G. (2008). Habitat suitability modeling and niche theory. J. Appl. Ecol. 45, 1372–1381. doi: 10.1111/j.1365-2664.2008.01524.x
IPCC-AR6. (2023). Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Eds. Core Writing Team, Lee H., Romero J. (Geneva, Switzerland: IPCC), 35–115. doi: 10.59327/IPCC/AR6-9789291691647
Jackson H. M., Johnson S. A., Morandin L. A., Richardson L. L., Guzman L. M., M’Gonigle L. K. (2022). Climate change winners and losers among North American bumblebees. Biol. Lett. 18, 20210551. doi: 10.1098/rsbl.2021.0551
Kerr J. T., Pindar A., Galpern P., Packer L., Potts S. G., Roberts S. M., et al. (2015). Climate change impacts on bumblebees converge across continents. Science 349, 177–180. doi: 10.1126/science.aaa7031
Kevan P. G., Rasmont P., Martinet B. (2024). Thermodynamics, thermal performance and climate change: temperature regimes for bumblebee (Bombus spp.) colonies as examples of superorganisms. Front. Bee Sci. 2. doi: 10.3389/frbee.2024.1351616
Krechemer FdS., Marchioro C. A. (2020). Past, present and future distributions of bumblebees in South America: Identifying priority species and areas for conservation. J. Appl. Ecol. 57, 1829–1839. doi: 10.1111/1365-2664.13650
Laroca S. (1972). Sobre a bionomia de Bombus brasiliensis (Hymenoptera, Apoidea). Acta Biol. Parana. 1, 7–28. doi: 10.5380/abpr.v1i0.636
Laroca S. (1976). Sobre a bionomia de Bombus morio (Hymenoptera, Apoidea). Acta Biol. Parana. 5, 107–127. doi: 10.5380/abpr.v5i0.904
Martinet B., Dellicour S., Ghisbain G., Przybyla K., Zambra E., Lecocq T., et al. (2021). Global effects of extreme temperatures on wild bumblebees. Conserv. Biol. 35, 1507–1518. doi: 10.1111/cobi.13685
Martinet B., Rasmont P., Cederberg B., Evrard D., Ødegaard F., Paukkunen J., et al. (2015). Forward to the north: Two Euro-Mediterranean bumblebee species now cross the Arctic Circle. Ann. Soc Entomol. 51, 303–309. doi: 10.1080/00379271.2015.1118357
Martins A. C., Silva D. P., de Marco P., Melo G. A. R. (2015). Species conservation under future climate change: the case of Bombus bellicosus, a potentially threatened South American bumblebee species. J. Insect. Conserv. 19, 33–43. doi: 10.1007/s10841-014-9740-7
Mcbride L. A., Hope A. P., Canty T. P., Bennett B. F., Tribett W. R., Salawitch R. J. (2021). Comparison of CMIP6 historical climate simulations and future projected warming to an empirical model of global climate. Earth System Dynamics 12, 545–579. doi: 10.5194/esd-12-545-2021
Morales C. L., Montalva J., Arbetman M. P., Aizen M. A., Martins A. C., Silva D. P. (2022). Does climate change influence the current and future projected distribution of an endangered species? The case of the southernmost bumblebee in the world. J. Insect. Conserv. 26, 257–269. doi: 10.1007/s10841-022-00384-5
Moure J. S., Melo G. A. R. (2023). Bombini Latreill. In: Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region - online version. Available online at: https://www.moure.cria.org.br/catalogue (Accessed October 8, 2024).
Moure M. S., Sakagami S. F. (1962). As mamangabas sociais do Brasil (Bombus Ltr.) (Hymenoptera, Apoidea). Studia Ent. Rio 5, 65–194.
Naimi B., Hamm N. A. S., Groen T. A., Skidmore A. K., Toxopeus A. G. (2014). Where is positional uncertainty a problem for species distribution modeling? Ecography 37, 191–203. doi: 10.1111/j.1600-0587.2013.00205.x
Nascimento A. C., Montalva J., Ascher J. S., Engel M. S., Silva D. P. (2022). Current and future distributions of a native Andean bumble bee. J. Insect. Conserv. 26, 559–569. doi: 10.1007/s10841-022-00395-2
Olesen J. M. (1989). Behaviour and nest structure of the Amazonian Bombus transversalis in Ecuador. J. Trop. Ecol. 5, 243–246. doi: 10.1017/S0266467400003540
Oliveira M. O., Cavalcante M. C., Freitas B. M. (2015). Nesting Behavior and colony description of the neotropical bombus (Thoracobombus) brevivillus in northeastern Brazil. J. Insect Behav. 28, 297–302. doi: 10.1007/s10905-015-9502-8
Oliveira W., Colares L. F., Porto R. G., Viana B. F., Tabarelli M., Lopes A. V. (2024). Food plants in Brazil: origin, economic value of pollination and pollinator shortage risk. Sci. Total Environ. 912. doi: 10.1016/j.scitotenv.2023.169147
Oliveira M. O., Meneses H. M., Nogueira D. S., Gomes A. M. S., Cavalcante M. C., Freitas B. M. (2022). Evidence of nest reactivation and perennial colonies in the Neotropical Bumble Bee Bombus brevivillus (Hymenoptera: Apidae: Bombini). Neotrop. Entomol. 51, 886–893. doi: 10.1007/s13744-022-00992-5
Oyen K. J., Dillon M. E. (2018). Critical thermal limits of bumblebees (Bombus impatiens) are marked by stereotypical behaviors and are unchanged by acclimation, age or feeding status. J. Exp. Biol. 221. doi: 10.1242/jeb.165589
Oyen K. J., Giri S., Dillon M. E. (2016). Altitudinal variation in bumble bee (Bombus) critical thermal limits. J. Therm. Biol. 59, 52–57. doi: 10.1016/j.jtherbio.2016.04.015
Paula G. A. R., Melo G. A. R. (2015). Inferring sex and caste seasonality patterns in three species of bumblebees from Southern Brazil using biological collections. Neotrop. Entomol. 44, 10–20. doi: 10.1007/s13744-014-0251-8
Phillips S. J., Anderson R. P., Schapire R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Model. 190 (3–4), 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Phillips S. J., Dudík M., Schapire R. E. (2021). Maxent software for modeling species niches and distributions (Version 3.4.1). Available online at: http://biodiversityinformatics.amnh.org/open_source/maxent/.
Quinlan G. M., Feuerborn C., Hines H. M., Grozinger C. M. (2023). Beat the heat: thermal respites and access to food associated with increased bumble bee heat tolerance. J. Exp. Biol. 226 (17), jeb245924. doi: 10.1242/jeb.245924
R Core Team. (2021). R: A language and environment for statistical computing (Vienna, Austria: The R Foundation for Statistical Computing).
Rollin O., Vray S., Dendoncker N., Denis Michez M., Ne D., Rasmont P. (2020). Drastic shifts in the Belgian bumblebee community over the last century. Biodiversity Conserv. 29 (8), 2553–2573. doi: 10.1007/s10531-020-01988-6
Romeiro L. A., Borges R. C., da Silva E. F., Guimarães J. T. F., Giannini T. C. (2023). Assessing entomological collection data to build pollen interaction networks in the tropical Amazon forest. Arthropod-Plant Inte. 17, 313–325. doi: 10.1007/s11829-023-09968-7
Sales L. P., Rodrigues L., Masiero R. (2021). Climate change drives spatial mismatch and threatens the biotic interactions of the Brazil nut. Glob. Ecol. Biogeogr. 30, 117–127. doi: 10.1111/geb.13200
Santos Júnior J. E., Santos F. R., Silveira F. A. (2015). Hitting an unintended target: phylogeography of Bombus brasiliensis Lepeletier 1836 and the first new Brazilian bumblebee species in a century (Hymenoptera: Apidae). PloS One 10, e0125847. doi: 10.1371/journal.pone.0125847
Schramek C. (2021). New Scenarios and Greater Certainty in IPCC AR6. Available online at: https://greencentralbanking.com/2021/08/10/new-scenarios-and-greater-certainty-in-ipcc-ar6/.
Sepúlveda Y., Goulson D. (2023). Feeling the heat: bumblebee workers show no acclimation capacity of upper thermal tolerance to simulated heatwaves. J. Therm. Biol. 116. doi: 10.1016/j.jtherbio.2023.103672
Singh A. P., Chandra A., De K., Uniyal V. P., Sathyakumar S. (2023). Decreasing potential suitable habitat of bumble bees in the Great Himalayan National Park Conservation area. Orient. Insects 57, 36–53. doi: 10.1080/00305316.2022.2040631
Taylor O. M., Cameron S. A. (2003). Nest construction and architecture of the Amazonian bumble bee (Hymenoptera: Apidae). Apidologie 34, 321–331. doi: 10.1051/apido:2003035
Thuiller W., Georges D., Gueguen M., Engler R., Breiner F. (2021) biomod2: Ensemble platform for species distribution modeling. Available online at: https://cran.r-project.org/package=biomod2 (Accessed January 2023).
Thuiller W., Lafourcade B., Engler R., Araújo M. B. (2009). BIOMOD – a platform for ensemble forecasting of species distributions. Ecography 32 (3), 369–373. doi: 10.1111/j.1600-0587.2008.05742.x
Vega L., Torres A., Hoffmann W., Lamprecht I. (2011). Thermal investigations associated with the behaviour patterns of resting workers of Bombus atratus (Hymenoptera: Apidae). J. Therm. Anal. Calorim. 104, 233–237. doi: 10.1007/s10973-011-1373-4
Wisz M. S., Hijmans R. J., Li J., Peterson A. T., Graham C. H., Guisan A. (2008). Effects of sample size on the performance of species distribution models. Diversity Distributions 14, 763–773. doi: 10.1111/j.1472-4642.2008.00482.x
White S. A., Dillon M. E. (2023). Climate warming and bumble bee declines: the need to consider sub-lethal heat, carry-over effects, and colony compensation. Front. Physiol. 14. doi: 10.3389/fphys.2023.1251235
Williams P. H., Cameron S. A., Hines H. M., Cederberg B., Rasmont P. (2008). A simplified subgeneric classification of the bumblebees (genus Bombus). Apidologie 39, 46–74. doi: 10.1051/apido:2007052
Keywords: Bombus, species distribution modeling, conservation, decision-making, priority zones
Citation: Nunes-Silva P, Acosta AL, Borges RC, Freitas BM, Oliveira RC, Giannini TC and Imperatriz-Fonseca VL (2025) Climate change will alter Amazonian bumblebees’ distribution, but effects are species-specific. Front. Bee Sci. 3:1510004. doi: 10.3389/frbee.2025.1510004
Received: 11 October 2024; Accepted: 29 January 2025;
Published: 17 February 2025.
Edited by:
Luca Ruiu, University of Sassari, ItalyReviewed by:
Moses Chemurot, National Agricultural Research Organisation, UgandaCopyright © 2025 Nunes-Silva, Acosta, Borges, Freitas, Oliveira, Giannini and Imperatriz-Fonseca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patrícia Nunes-Silva, cG5zaWx2YUBhbHVtbmkudXNwLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.