 André Krahner
André Krahner Michael Maixner
Michael Maixner Matthias Porten3
Matthias Porten3 Thomas Schmitt
Thomas Schmitt- 1Julius Kühn Institute (JKI) – Federal Research Centre for Cultivated Plants, Institute for Bee Protection, Braunschweig, Germany
- 2Julius Kühn Institute (JKI) – Federal Research Centre for Cultivated Plants, Institute for Plant Protection in Fruit Crops and Viticulture, Siebeldingen, Germany
- 3Public Service Centre for Rural Development in the Region Mosel (DLR Mosel), Bernkastel-Kues, Germany
- 4Senckenberg German Entomological Institute, Müncheberg, Germany
- 5Zoology, Institute of Biology, Faculty of Natural Sciences I, Martin Luther University Halle-Wittenberg, Halle (Saale), Germany
- 6Entomology and Biogeography, Institute of Biochemistry and Biology, Faculty of Science, University of Potsdam, Potsdam, Germany
Insect conservation through habitat management is particularly important in regions of high biodiversity potential, such as steep-slope viticultural landscapes. Therefore, we studied the effects of realistic options for the conservation of open and flower-rich vineyard fallows on local wild bee communities. Using a fully-crossed factorial design, we sampled bees in 10 vineyard fallows in the Moselle region (SW Germany), which were partially subjected to different treatments (mulching, sowing of wild flower strips, combination, and untreated control). Over two years, we sampled 9323 individuals from 140 bee species with hand netting and pan traps. Based on the hand-netting samples, AICc-based model selection (Generalized Linear Mixed Models) revealed effects of fallow treatment, interacting with sampling year, on the number of sampled bee individuals and on the number of sampled bee species. In the second sampling year, we sampled significantly more individuals in wild flower strips (WFS) as well as in the combination treatment than in the untreated control. Also in the second year, the number of sampled species was significantly higher in the combination treatment than in the mulching treatment, and significantly higher than in the untreated control. Vineyard fallow management can promote bee communities in steep-slope viticultural landscapes, and options involving WFS, especially in combination with an annual mulching, should be preferred over options solely relying on repeated mulching. Moreover, the findings highlight that management options involving WFS need some time after initiation for unfolding their positive effects on bee communities.
1 Introduction
Declining insect populations have been reported worldwide (Wagner, 2020). While agricultural land-use intensification has been identified as a major driver of recent insect decline (Habel et al., 2019), abandonment of agricultural management can also result in habitat degradation (Caraveli, 2000; Giupponi et al., 2006). This is especially true for wild bee populations in steep-slope vineyards, which are often xerothermophilic, preferring open and sun-exposed habitats (Krahner et al., 2018; Böhm et al., 2024).
In order to preserve suitable bee habitats within steep-slope viticulture landscapes, different management options are available, which involve the provision of food resources and measures to maintain the openness of habitats. In general, bee communities in agroecosystems benefit from additional supply of floral resources in the form of managed wildflower strips (Lowe et al., 2021). This also applies to managed vineyards, where wild bee abundance and species richness are positively correlated with floral cover of entomophilous plant species (Kratschmer et al., 2018) and floral diversity (Böhm et al., 2024). However, compared to floral enhancements in managed vineyards, our knowledge about the effects of such treatment in set-aside areas within viticultural landscapes (vineyard fallows) on bee communities is still limited.
For maintaining suitable bee habitats on vineyard fallows, prevention of bush encroachment is pivotal. Without management, vineyard fallows are generally overgrown by shrubby vegetation (Wersebeckmann et al., 2023). This may negatively affect floral resources, because important food plants are replaced by dominant, less bee attractive plants (Kirmer et al., 2018). Likewise, bush encroachment may also have a negative impact on nesting resources (Boetzl et al., 2022), especially because most ground-nesting bees characteristic for the region prefer open habitats for nesting (Westrich, 2019; Böhm et al., 2024). Repeated mulching is a common practice in managed vineyards (Pardini et al., 2002), as well as in sown flower strips and field margins with spontaneous vegetation (Kirmer et al., 2018). However, our understanding of how repeated mulching affects bee communities in vineyard fallows remains incomplete.
In order to close remaining knowledge gaps and to provide recommendations for insect conservation efforts in vineyard landscapes on steep slopes, we investigated the short-term effects of wild flower strips and repeated mulching, including their interaction, on the abundance and species richness of wild bees. Our main hypotheses were, 1) that mulching and flower strips would be associated with a higher number of bee individuals and species compared to the no-treatment control, 2) that the combination of mulching and wild flower strips would be associated with a greater bee abundance and species diversity compared to single treatments, and 3) that these effects would increase over the years.
2 Material and methods
We investigated the effect of different options for managing vineyard fallows on wild bee communities. Using a fully-crossed factorial design, we sampled bees in 10 vineyard fallows in the Moselle valley in SW Germany (49.916469°N, 6.995033°E; Figure 1; for exact locations of study sites, refer to https://doi.org/10.5073/20240408-104443-0). Eight of these vineyard fallows were clustered in one study site, with nearest neighbors being separated by 160 to 370 m, while the other two vineyard fallows had a distance of 5045 m to the nearest neighbor. The fallows varied in size (1060 to 4109 m²), were adjacent to managed vineyards, and were partially subjected to four different treatments: mulching (one fall mulching event per year), sowing of a wild flower seed mixture in spring 2013 to create wildflower strips (WFS; Saaten-Zeller GmbH & Co. KG, Eichenbühl-Riedern, seed provenance south-western Germany; Table 1), combination treatment of sowing and mulching, and untreated control. Within a vineyard fallow, differently treated areas were the same area size. We sampled bees from April to October in 2013 and 2014, under favorable weather conditions (no or little wind, cloud cover 50% or less, temperatures ≥15 °C), by hand netting (all 10 vineyard fallows), and using pan traps as a complementary method (6 vineyard fallows). Hand netting was standardized to a sampling time of 10 min per treatment variant and sampling event, along a 1 m wide non-permanent transect line that covered as much of the treatment area as possible, while maintaining a minimum distance of 2 m from the edge of the treatment area. For hand netting, the sampling sequence of transects was randomized for each sampling day so that the individual transects were sampled at different times of the day, with the restriction that sampling along shaded transects was avoided. In total, we completed 13 sampling events (5 in 2013, 8 in 2014). For pan trapping, we used triplets of fluorescent blue, white and yellow traps (Rondo-Gelbfangschale, Temmen GmbH, Hattersheim, Germany; upper diameter: 23 cm; volume: 2.3 L; Krahner et al., 2021). These traps were made by priming with white spray paint (sparvar 1315), followed by application of fluorescent spray paints (sparvar 3107, 3104, and 3103 for blue, yellow, and white colors, respectively; Spray-Color GmbH, Merzenich, Germany). In total, we had 9 sampling events (5 in 2013, 4 in 2014). Pan trapping events varied between 48 and 96 hours, but within a sampling event, pan traps were exposed the same time across all vineyard fallows and treatment variants. Some of the sampled data have already been used for analysis in a comparison of different methods for sampling wild bees (Krahner et al., 2021).
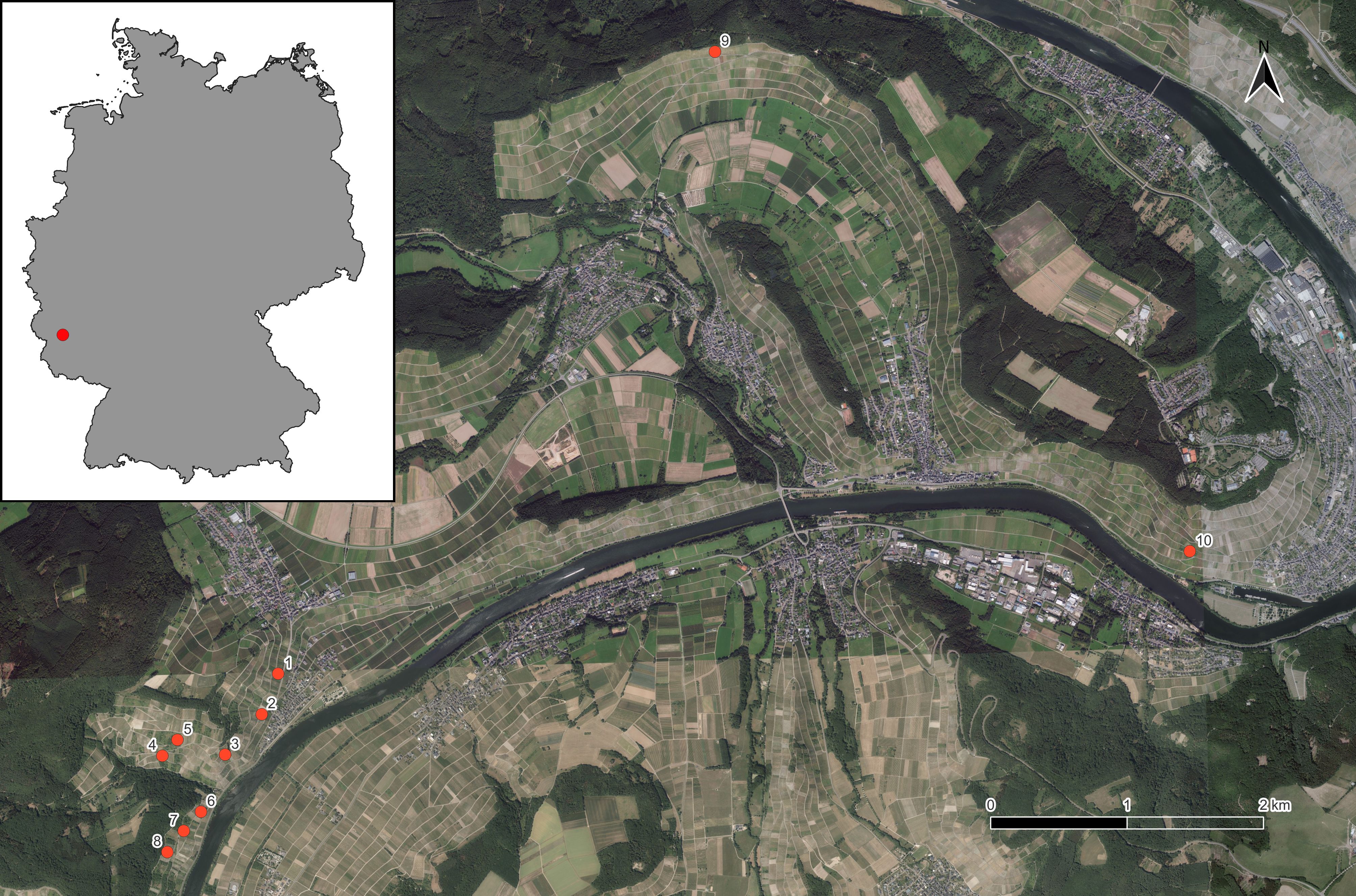
Figure 1 Location of the studied vineyard fallows in the Moselle river valley (1–10, red dots representing fallow centroids), inset graphic showing the location of the study area (red dot) within Germany (grey). Digital orthophoto (DOP): © GeoBasis-DE/BKG 2024; Terms of use: http://sg.geodatenzentrum.de/web_public/nutzungsbedingungen.pdf. Geodata for Germany modified from Hijmans et al. (2015).
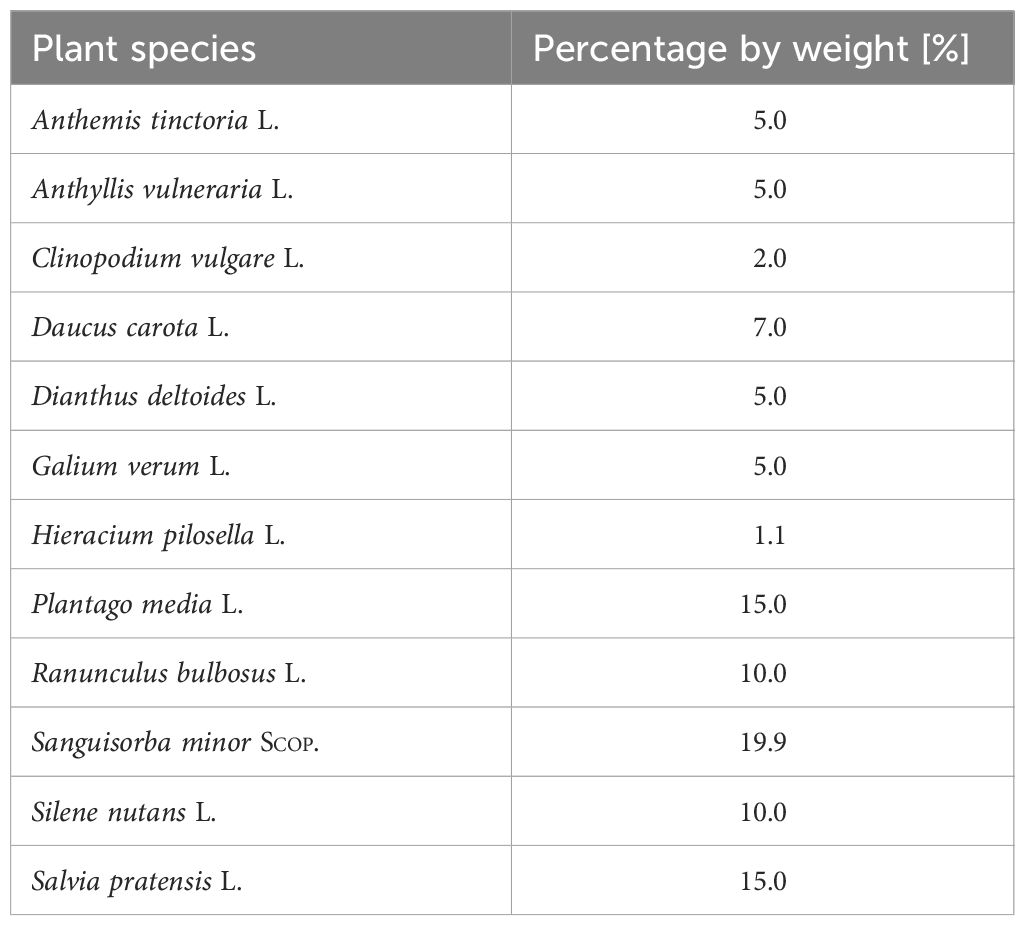
Table 1 Composition of the perennial wild flower seed mixture used as a treatment variant.
Following sampling, we stored insects in 70% Ethanol. We dried, pinned and identified all bee individuals to species level following the taxonomy in Westrich et al. (2011) and using the following literature: Ebmer (1969, 1970, 1971); Dathe (1980); Koster (1986); Dylewska (1987); Dorn and Weber (1988); Mauss (1992); Amiet (1996); Amiet et al. (1999, 2001, 2004, 2007, 2010); Schmid-Egger and Scheuchl (1997); Banaszak and Romasenko (1998); Scheuchl (2000, 2006); Gusenleitner and Schwarz (2002); Straka and Bogusch (2011); and Gokzekade et al. (2015). We did not separate some morphologically similar species, combining Andrena ovatula, A. wilkella, A. intermedia, A. similis, and A. gelriae as Andrena ovatula agg.; Bombus cryptarum, B. lucorum, B. magnus, and B. terrestris as Bombus terrestris agg.; Bombus hortorum and B. ruderatus as Bombus hortorum agg.; Halictus eurygnathus, H. langobardicus and H. simplex as Halictus simplex agg.; Lasioglossum nitidulum and L. smeathmanellum as Lasioglossum smeathmanellum agg. In the analysis, we treated these species groups as single species. Voucher specimens are deposited in the collection of the corresponding author.
All statistical analyses were conducted in the R environment (v4.2.2; R Core Team, 2022). We analysed the number of sampled wild bee individuals and species (excluding managed Apis mellifera) with Generalized Linear Mixed Models (GLMMs), using vineyard fallow as random intercept factor to account for among-fallow differences, a log-link function and a negative binomial (bee individuals) or a Poisson (bee species) error distribution. Because replication (number of vineyard fallows) varied between sampling methods, we conducted separate analyses for hand net and pan trap samples. We created full models with treatment variant, year and the treatment-by-year interaction as fixed effects. GLMMs were fit using the ‘glmmTMB’ package (v1.1.5; Brooks et al., 2017). For selection of most parsimonious models, we compared the full models to all possible reduced models using Akaike Information Criterion (AICc), holding the random effect structure constant. We used the ‘MuMIn’ package (v1.47.1; Bartoń, 2022) for model selection, reducing the fixed effect terms of the full models to yield best-fitting final models. We validated final models by visually inspecting the residual plots, using function ‘simulateResiduals’ from the ‘DHARMa’ package (v0.4.6; Hartig, 2020). We used the ‘emmeans’ package (v1.8.3; Lenth, 2022) for post hoc tests, and corrected P values via the Tukey method. We determined significant statistical differences at alpha = 0.05.
3 Results
In total, we sampled 9323 individuals (pan traps: 7867, hand netting: 1456) from 140 bee species. Based on the hand-netting samples, AICc-based model selection (lowest AICc) revealed effects of fallow treatment, interacting with sampling year, on the number of sampled bee individuals (Figure 2) and on the number of sampled bee species (Figure 3).

Figure 2 Effect of treatment variants, interacting with study year, on the number of bee individuals sampled with hand netting. Red dots and error bars represent model fit (mean and 95% CI), while grey dots represent observations from 10 vineyard fallows. Different letters indicate significant differences (α = 0.05). Treatments: untreated control (ctrl), annual mulching (m), sowing of wild flower strip (s), and combination of annual mulching and sowing (ms). Treatments were initiated in spring 2013.
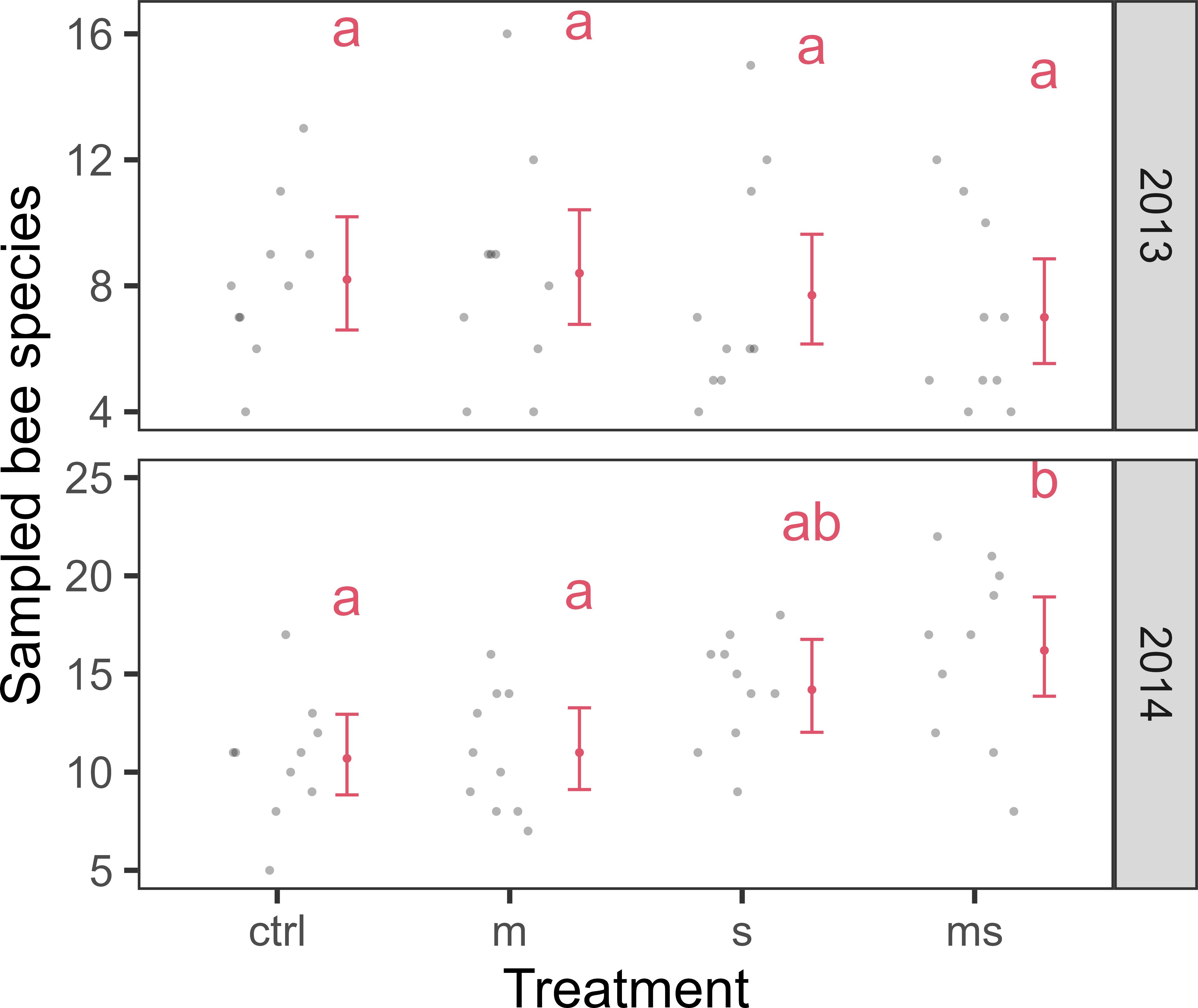
Figure 3 Effect of treatment variants, interacting with study year, on the number of bee species sampled with hand netting. Red dots and error bars represent model fit (mean and 95% CI), while grey dots represent observations from 10 vineyard fallows. Different letters indicate significant differences (α = 0.05). Treatmens: untreated control (ctrl), annual mulching (m), sowing of wild flower strip (s), and combination of annual mulching and sowing (ms). Treatments were initiated in spring 2013.
We did not observe significant differences with regard to sampled individuals and species in the first sampling year. In the second sampling year, we sampled significantly more individuals in the WFS treatment (ratio = 1.484, SE = 0.221, df = 78, p = 0.047) than in the untreated control. Also in the second year, we sampled significantly more bee individuals in the combination treatment compared to the untreated control (ratio = 1.679, SE = 0.248, df = 78, p = 0.004) and compared to the mulching treatment (ratio = 1.555, SE = 0.227, df = 78, p = 0.018). Regarding the number of sampled bee species, we sampled significantly more species in the combination treatment compared to the untreated control (ratio = 1.514, SE = 0.189, df = 79, p = 0.007) and compared to the mulching treatment (ratio = 1.473, SE = 0.182, df = 78, p = 0.013). Based on pan trap samples, the treatment variable was incorporated neither in the most parsimonious model explaining sampled bee individuals nor in the most parsimonious model explaining sampled bee species (in both cases, ΔAICc > 4).
4 Discussion
The 140 wild bee species sampled over two years in this study correspond to one quarter of the total number of bee species found in Germany. This is roughly comparable to results from a two-year study of managed vineyards from the same study region (Böhm et al., 2024: 1013 individuals from 110 species), and also to results from a one-year study of vineyard landscapes from the Upper Middle Rhine Valley in SW Germany (Wersebeckmann et al., 2023: 3385 individuals from 115 species). These figures highlight the value of viticultural landscapes with steep slopes, and especially of vineyard fallows within these landscapes, as habitats for wild bees. To our best knowledge, our study is the first to investigate the effects of bee habitat management in vineyard fallow on steep slopes.
Our experiment covered only the first two years after establishment of the treatment variants. Because perennial flower strips may unfold their effects on pollinator communities over a longer period of several years after establishment (Albrecht et al., 2021; Schmied et al., 2022), longer-term observations are needed to assess the longer-term conservation value of the treatments investigated here. Still, the present results are relevant for bee conservation in vineyard landscapes, because vineyard fallows often exist for only one to a few years, before they are replanted with wine grapes. Moreover, the study design has an emphasis on a high temporal sampling intensity rather than a wide sampling area. This approach is favorable in terms of yielding sampling results representative of the real species richness within a bee community (Levenson et al., 2024). Our approach also means our results directly apply to a relatively small study area. However, since we chose our study area to represent common landscape context and management practices in central Europe, we are confident that the implications for conservation measures we derive from our study can be extrapolated to vineyard landscapes on steep slopes across central Europe.
In contrast to hand net samples, we observed no difference in sampled bee abundance and species richness between treatment variants, based on pan trap samples. This may be a consequence of the lower replication in this method (6 vineyard fallows sampled with pan traps, compared to 10 fallows sampled with hand netting). Moreover, although pan traps are an efficient method for sampling bees (Westphal et al., 2008), especially in steep-slope viticultral landscapes (Krahner et al., 2021), they have a strong bias for particular taxa (Portman et al., 2020) and trait groups of bees (Klaus et al., 2024). It is also possible that pan traps were unable to resolve differences in bee activity between treatments, due to the close proximity of the differently treated fallow areas. While the competition between neighboring pan traps levels off at distances of 3–5 m between traps (Droege et al., 2010), pan traps in one treatment variant may have attracted bees from adjacent treatment variants, especially in absence of surrounding floral resources (Wilson et al., 2008; Prendergast and Hogendoorn, 2021). In the following, we therefore focus the discussion on the results from hand netting. More generally, our results caution against close distances when replicating pan traps within a landscape.
Based on hand netting data, both treatment variants involving WFS increased the number of sampled bee individuals compared to the untreated control in the second study year, but not so in the first study year. This is in line with previous research, which showed positive effects of perennial WFS to increase over the years following sowing (Lowe et al., 2021). The effect may be driven by the sown perennials, which increase in floral abundance over the first two years following sowing, accompanied by an increase in bee abundance (Carvell et al., 2022). Likewise, Kehinde et al. (2018) observed that an increase in floral abundance resulted in an increased bee abundance in viticultural landscapes. Thus, although we did not survey floral resources in this study, floral abundance would be a plausible explanation for the treatment effect on bee abundance we observed in our study.
In general, pollinator richness in WFS increases over the first three years (Lowe et al., 2021), which might explain why we did not observe treatment effects of WFS before the second study year. We observed a positive effect of WFS on bee species richness only in combination with annual mulching, while mulching in the first year is likely to positively influence establishment of sown plants (Carvell et al., 2022). Therefore, we assume that annually mulched WFS had a positive effect on bee species richness through an increase in species diversity of plants providing floral resources to bees, and/or through an increase in nesting resources. Species richness of wild bees in viticultural landscapes and the diversity of flowering plants are correlated (Böhm et al., 2024; but see Wersebeckmann et al., 2023). However, because we did not survey flowering plant species richness in this study, we cannot validate this assumption with our data. Since the soil of the vineyard fallows remained undisturbed after sowing of the WFS, and because wild bees use WFS as nesting habitats under low soil disturbance (Boetzl et al., 2022), it is likely that the annually mulched WFS served as a nesting habitat for wild bees. Most native bee species in viticultural landscapes are ground nesting (Krahner et al., 2018; Wersebeckmann et al., 2023), and creation of sun exposed ground spots with scarce vegetation is an important conservation measure for promoting bees (Gardein et al., 2022).
Based on our study, the mechanisms behind the observed effects of the combination treatment remain obscure. The increase in bee species richness and abundance may be a consequence of individuals dispersing into the fallow due to increasing resources, or represent a population build-up of bees nesting within the study site, or both. Future studies could involve resource intensive nest surveys and mark-recapture methodology in order to investigate further the role of these two mechanisms.
In summary, our study shows that WFS are an effective conservation measure for increasing bee abundance and species richness in vineyard fallows in the short term. Corroborating the recommendations by Carvell et al. (2022), an annual mulching of WFS is highly advisable. Vineyard fallows often exist for several to many years, and the habitat value of WFS may change over the years (Albrecht et al., 2021), with decreases in wild bee abundance and diversity from one and two years after establishment, respectively. Therefore, we suggest studies with a longer observation period in order to investigate whether our findings hold true in the longer term.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://doi.org/10.5073/20240408-104443-0.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements. The permission required for the collection of bees and butterflies was granted by the responsible authority (Struktur- und Genehmigungsdirektion Nord, Koblenz).
Author contributions
AK: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. MM: Conceptualization, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing. MP: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing. TS: Conceptualization, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was financially supported by the German Federal Ministry of Food and Agriculture (BMEL) based on a decision of the Parliament of the Federal Republic of Germany, granted by the Federal Office for Agriculture and Food (BLE; grant number 2811HS003). The APC was funded by Julius Kühn Institute (JKI) – Federal Research Centre for Cultivated Plants.
Acknowledgments
We thank T. Volkmer for sampling and initial determination of some of the wild bee individuals, A.W. Ebmer, O. Diestelhorst, G. Reder, H.H. Dathe, and J. Esser for supporting bee species identification, and the Moselle valley wine growers for providing access to their vineyards. The permission required for the collection of bees and butterflies was granted by the responsible authority (Struktur- und Genehmigungsdirektion Nord, Koblenz).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Albrecht M., Knecht A., Riesen M., Rutz T., Ganser D. (2021). Time since establishment drives bee and hoverfly diversity, abundance of crop-pollinating bees and aphidophagous hoverflies in perennial wildflower strips. Basic Appl. Ecol. 57, 102–114. doi: 10.1016/j.baae.2021.10.003
Amiet F. (1996). Hymenoptera. Apidae, 1. Teil. Allgemeiner Teil, Gattungsschlüssel, die Gattungen Apis, Bombus and Psithyrus (Neuchâtel: Centre Suisse de Cartographie de la Faune and Schweizerische Entomologische Gesellschaft).
Amiet F., Herrmann M., Müller A., Neumeyer R. (2001). Apidae 3 (Neuchâtel: Centre Suisse de Cartographie de la Faune and Schweizerische Entomologische Gesellschaft)
Amiet F., Herrmann M., Müller A., Neumeyer R. (2004). Apidae 4 (Neuchâtel: Centre Suisse de Cartographie de la Faune and Schweizerische Entomologische Gesellschaft)
Amiet F., Herrmann M., Müller A., Neumeyer R. (2007). Apidae 5 (Neuchâtel: Centre Suisse de Cartographie de la Faune and Schweizerische Entomologische Gesellschaft)
Amiet F., Herrmann M., Müller A., Neumeyer R. (2010). Apidae 6 (Neuchâtel: Centre Suisse de Cartographie de la Faune and Schweizerische Entomologische Gesellschaft)
Amiet F., Müller A., Neumeyer R. (1999). Apidae 2 (Neuchâtel: Centre Suisse de Cartographie de la Faune and Schweizerische Entomologische Gesellschaft)
Banaszak J., Romasenko L. (1998). Megachilid Bees of Europe (Hymenoptera, Apoidea, Megachilidae). (Bydgoszcz: Pedagogical University).
Bartón K. (2022). “MuMIn,” in Multi-model interference. Version 1.47.1. Available at: https://CRAN.R-project.org/package=MuMIn.
Boetzl F. A., Krimmer E., Holzschuh A., Krauss J., Steffan-Dewenter I. (2022). Arthropod overwintering in agri-environmental scheme flowering fields differs among pollinators and natural enemies. Agricul Ecosyst. Environ. 330, 107890. doi: 10.1016/j.agee.2022.107890
Böhm L., Krahner A., Porten M., Maixner M., Schäffer J., Schmitt T. (2024). Crossing old concepts: the ecological advantages of new vineyard types. Diversity 16, 44. doi: 10.3390/d16010044
Brooks M. E., Kristensen K., van Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378. doi: 10.32614/RJ-2017-066
Caraveli H. (2000). A comparative analysis on intensification and extensification in mediterranean agriculture: dilemmas for LFAs policy. J. Rural Stud. 16, 231–242. doi: 10.1016/S0743-0167(99)00050-9
Carvell C., Mitschunas N., McDonald R., Hulmes S., Hulmes L., O’Connor R. S., et al. (2022). Establishment and management of wildflower areas for insect pollinators in commercial orchards. Basic Appl. Ecol. 58, 2–14. doi: 10.1016/j.baae.2021.11.001
Dathe H. H. (1980). Die Arten der Gattung Hylaeus F. in Europa (Hymenoptera: Apoidea, Colletidae). Mitteilungen aus dem Zoologischen Museum Berlin 56, 207–294.
Dorn M., Weber D. (1988). Die Luzerne-Blattschneiderbiene and ihre Verwandten in Mitteleuropa (Wittenberg Lutherstadt: A. Ziemsen)
Droege S., Tepedino V. J., LeBuhn G., Link W., Minckley R. L., Chen Q., et al. (2010). Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conserv. Diversity 3, 15–23. doi: 10.1111/j.1752-4598.2009.00074.x
Dylewska M. (1987). Die Gattung Andrena Fabricius (Andrenidae, Apoidea) in Nord- und Mitteleuropa. Acta Zoologica Cracoviensia 30, 359–708.
Ebmer P. A. W. (1969). Die Bienen des Genus Halictus Latr. s. l. im Großraum von Linz (Hymenoptera, Apidae). Systematik, Biogeographie, Ökologie und Biologie mit Berücksichtigung aller bisher aus Mitteleuropa bekannten Arten. Teil I. Naturkundliches Jahrbuch der Stadt Linz. 15, 133–183.
Ebmer P. A. W. (1970). Die Bienen des Genus Halictus Latr. s. l. im Großraum von Linz (Hymenoptera, Apidae). Teil II. Naturkundliches Jahrbuch der Stadt Linz 16, 19–82.
Ebmer P. A. W. (1971). Die Bienen des Genus Halictus Latr. s. l. im Großraum von Linz (Hymenoptera, Apidae). Teil III. Naturkundliches Jahrbuch der Stadt Linz 17, 63–156.
Gardein H., Fabian Y., Westphal C., Tscharntke T., Hass A. (2022). Ground-nesting bees prefer bare ground areas on calcareous grasslands. Global Ecol. Conserv. 39, e02289. doi: 10.1016/j.gecco.2022.e02289
Giupponi C., Ramanzin M., Sturaro E., Fuser S. (2006). Climate and land use changes, biodiversity and agri-environmental measures in the Belluno province, Italy. Environ. Sci. Policy 9, 163–173. doi: 10.1016/j.envsci.2005.11.007
Gokzekade J. F., Gereben-Krenn B.-A., Neumayer J., Krenn H. W. (2015). Feldbestimmungsschlüssel für die Hummeln Österreichs, Deutschlands and der Schweiz (Hymenoptera, Apidae). Linzer biologische Beiträge 47, 5–42
Gusenleitner F., Schwarz M. (2002). Weltweite Checkliste der Bienengattung Andrena mit Bemerkungen and Ergänzungen zu paläarktischen Arten (Hymenoptera, Apidae, Andreninae, Andrena). Entomofauna Supplement 10, 1–1280
Habel J. C., Samways M. J., Schmitt T. (2019). Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodiversity Conserv. 28, 1343–1360. doi: 10.1007/s10531-019-01741-8
Hartig F. (2022). “DHARMa,” in Residual diagnostics for hierarchical (multi-level / mixed) regression models. Version 0.4.6. Availabe at: https://CRAN.R-project.org/package=DHARMa (Accessed October 5, 2023).
Hijmans R., Kapoor J., Wieczorek J., Garcia N., Maunahan A., Rala A., et al. (2015). GADM database of Global Administrative Areas. Version 2.8. Available at: http://gadm.org/country (Accessed February 24, 2016).
Kehinde T., von Wehrden H., Samways M., Klein A.-M., Brittain C. (2018). Organic farming promotes bee abundance in vineyards in Italy but not in South Africa. J. Insect Conserv. 22, 61–67. doi: 10.1007/s10841-017-0038-4
Kirmer A., Rydgren K., Tischew S. (2018). Smart management is key for successful diversification of field margins in highly productive farmland. Agricul Ecosyst. Environ. 251, 88–98. doi: 10.1016/j.agee.2017.09.028
Klaus F., Ayasse M., Classen A., Dauber J., Diekötter T., Everaars J., et al. (2024). Improving wild bee monitoring, sampling methods, and conservation. Basic Appl. Ecol. 75, 2–11. doi: 10.1016/j.baae.2024.01.003
Koster A. (1986). Het genus Hylaeus in Nederland (Hymenoptera, Colletidae). Zool Bijdragen 36, 1–120
Krahner A., Dathe H. H., Schmitt T. (2018). Wildbienen (Hymenoptera, Aculeata: Apiformes) des Mittleren Moseltals: Die Weinbausteillagen im Klotten-Treiser Moseltal. Beiträge zur Entomol = Contribut to Entomology 68, 107–131. doi: 10.21248/contrib.entomol.68.1.107-131
Krahner A., Schmidt J., Maixner M., Porten M., Schmitt T. (2021). Evaluation of four different methods for assessing bee diversity as ecological indicators of agro-ecosystems. Ecol. Indic. 125, 107573. doi: 10.1016/j.ecolind.2021.107573
Kratschmer S., Pachinger B., Schwantzer M., Paredes D., Guernion M., Burel F., et al. (2018). Tillage intensity or landscape features: What matters most for wild bee diversity in vineyards? Agricul Ecosyst. Environ. 266, 142–152. doi: 10.1016/j.agee.2018.07.018
Lenth R. V. (2022). “emmeans,” in Estimated marginal means, aka least-squares means. Version 1.8.3. Available at: https://CRAN.R-project.org/package=emmeans (Accessed October 5, 2023).
Levenson H. K., Metz B. N., Tarpy D. R. (2024). Effects of study design parameters on estimates of bee abundance and richness in agroecosystems: a meta-analysis. Ann. Entomol Soc. Am. 117, 92–106. doi: 10.1093/aesa/saae001
Lowe E. B., Groves R., Gratton C. (2021). Impacts of field-edge flower plantings on pollinator conservation and ecosystem service delivery – A meta-analysis. Agricul Ecosyst. Environ. 310, 107290. doi: 10.1016/j.agee.2020.107290
Mauss V. (1992). Bestimmungsschlüssel für Hummeln der Bundesrepublik Deutschland (Hamburg: Deutscher Jugendbund für Naturbeobachtung).
Pardini A., Faiello C., Snowball R., Mancuso S., Longhi F. (2002). Cover crop species and their management in vineyards and olive groves. Adv. Hortic. Sci. [rivista dell’ortofloroftutticol italiana]. 16, 225–234. doi: 10.1400/14122
Portman Z. M., Bruninga-Socolar B., Cariveau D. P. (2020). The state of bee monitoring in the United States: A call to refocus away from bowl traps and towards more effective methods. Ann. Entomol Soc. America 113, 337–342. doi: 10.1093/aesa/saaa010
Prendergast K. S., Hogendoorn K. (2021). FORUM: Methodological shortcomings and lack of taxonomic effort beleaguer Australian bee studies. Austral Ecol. 46, 880–884. doi: 10.1111/aec.12998
R Core Team (2022). R: A language and environment for statistical computing (Vienna: R Foundation for Statistical Computing)
Scheuchl E. (2000). Illustrierte Bestimmungstabellen der Wildbienen Deutschlands and Österreichs. Band I: Schlüssel der Gattungen and der Arten der Familie Anthophoridae (Velden: Eigenverlag).
Scheuchl E. (2006). Illustrierte Bestimmungstabellen der Wildbienen Deutschlands and Österreichs. Band II: Schlüssel der Arten der Familien Megachilidae and Melittidae (Stenstrup: Apollo Books)
Schmid-Egger C., Scheuchl E. (1997). Illustrierte Bestimmungstabellen der Wildbienen Deutschlands and Österreichs. Band III: Schlüssel der Arten der Familie Andrenidae (Velden/Vils: Eigenverlag)
Schmied H., Getrost L., Diestelhorst O., Maaßen G., Gerhard L. (2022). Between perfect habitat and ecological trap: even wildflower strips mulched annually increase pollinating insect numbers in intensively used agricultural landscapes. J. Insect Conserv. 26, 425–434. doi: 10.1007/s10841-022-00383-6
Straka J., Bogusch P. (2011). Contribution to the taxonomy of the Hylaeus gibbus species group in Europe (Hymenoptera, Apoidea and Colletidae). Zootaxa 2932, 51–67. doi: 10.11646/zootaxa.2932.1.6
Wagner D. L. (2020). Insect declines in the anthropocene. Annu. Rev. Entomology 65, 457–480. doi: 10.1146/annurev-ento-011019-025151
Wersebeckmann V., Warzecha D., Entling M. H., Leyer I. (2023). Contrasting effects of vineyard type, soil and landscape factors on ground- versus above-ground-nesting bees. J. Appl. Ecol. 60, 601–613. doi: 10.1111/1365-2664.14358
Westphal C., Bommarco R., Carré G., Lamborn E., Morison N., Petanidou T., et al. (2008). Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 78, 653–671. doi: 10.1890/07-1292.1
Westrich P., Frommer U., Mandery K., Riemann H., Ruhnke H., Saure C., et al. (2011). “Rote Liste und Gesamtartenliste der Bienen (Hymenoptera, Apidae) Deutschlands," in Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands: Band 3: Wirbellose Tiere (Teil 1) (Bundesamt für Naturschutz, Bonn-Bad Godesberg), 373–417.
Keywords: wild bees (Apiformes), vineyard fallows, wild flower strips, mulching, insect conservation, set-aside, viticulture, pollinators
Citation: Krahner A, Maixner M, Porten M and Schmitt T (2024) Annually mulched wild flower strips increase the observed wild bee (Apiformes) species richness and abundance in vineyard fallows in the short term. Front. Bee Sci. 2:1391789. doi: 10.3389/frbee.2024.1391789
Received: 26 February 2024; Accepted: 29 May 2024;
Published: 13 June 2024.
Edited by:
Katja Hogendoorn, University of Adelaide, AustraliaReviewed by:
Michael C. Orr, Institute of Zoology (CAS), ChinaJan Brus, Palacký University, Olomouc, Czechia
Copyright © 2024 Krahner, Maixner, Porten and Schmitt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: André Krahner, YW5kcmUua3JhaG5lckBqdWxpdXMta3VlaG4uZGU=