
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Bee Sci., 02 July 2024
Sec. Bee Protection and Health
Volume 2 - 2024 | https://doi.org/10.3389/frbee.2024.1384846
This article is part of the Research TopicApplied Bee Science and Technology TransferView all 8 articles
 Pablo Juri1*
Pablo Juri1* Enrique Nogueira1José Anzola1Valentina Rodríguez-Batista1
Enrique Nogueira1José Anzola1Valentina Rodríguez-Batista1 Belén Branchiccela2Ciro Invernizzi3
Belén Branchiccela2Ciro Invernizzi3Introduction: Varroa destructor stands as the primary global pest of honey bees, inflicting direct harm on bees while also acting as a vector for a multitude of viruses. Integrated pest management is widely recognized as the optimal strategy for minimizing acaricide application. Designed bottom boards that can limit the growth of the mite population by removing them from the hive and preventing their return and reinfestation of the colony show promising potential.
Methods: The aim of this study was to compare Varroa population growth by employing three different types of bottom boards. An apiary of 55 colonies was randomly divided into five groups according to the bottom board they had: 1) tubular bottom board, 2) screen bottom board, 3) sticky bottom board, 4) conventional bottom board with a permanent treatment with acaricide (positive control), and 5) conventional bottom board (negative control). Varroa infestation level in bees, colony strength (bee and brood population), and honey reserves were determined between April and December (between autumn and spring in the southern hemisphere).
Results: Colonies that had sticky bottom boards had approximately 50% less Varroa infestation than colonies with conventional bottom boards at the end of the study. Varroa infestation levels did not differ significantly between the tubular, screen, and conventional bottom boards. Colony strength was similar in all groups. No differences were observed in honey production between colonies from different groups.
Discussion: The use of sticky bottom boards was the most promising board to limit Varroa population growth and prevent colonies' reinfestation. This result encourages future studies to analyze the incorporation of this strategy in integrated pest management programs.
The ectoparasitic mite Varroa destructor is the main sanitary pest of honey bees worldwide (Nazzi and Le Conte, 2016; Noël et al., 2020; Traynor et al., 2020). This mite reproduces in brood cells by feeding on the hemolymph of larvae and pupae, and has a phoretic phase where it feeds on bees fat bodies (Rosenkranz et al., 2010; Nazzi and Le Conte, 2016; Ramsey et al., 2019). In addition to the direct damage caused by V. destructor it also acts as a vector of different RNA viruses and favors their replication by suppressing the bees immune response (Yang and Cox-Foster, 2005; Beaurepaire et al., 2020). The virus mostly associated with V. destructor is the Deformed Wing Virus (DWV) (De Miranda and Genersch, 2010; Wilfert et al., 2016). The infestation of worker pupae by V. destructor decreases honey bees’ longevity (Amdam et al., 2004; Aldea and Bozinovic, 2020), increases the titers of the DWV (De Miranda and Genersch, 2010; Beaurepaire et al., 2020), and suppresses the immune response exposing honey bees to infection by other organisms (Yang and Cox-Foster, 2005; Annoscia et al., 2019). At the colony level, infected colonies reduce their population and productivity (Currie and Gatien, 2006; Emsen et al., 2014) and produce fewer swarms (Fries et al., 2003). In many regions of the world, colonies that do not receive acaricides often collapse (Rosenkranz et al., 2010). The synthetic molecules widely used to control V. destructor are the organophosphate coumaphos, the pyrethroids tau-fluvalinate and flumethrin, and the formamidine amitraz (Rosenkranz et al., 2010). However, mites can develop resistance to all of them and residues can persist in hive products (Elzen et al., 2000; Maggi et al., 2011; Mitton et al., 2022).
Integrated pest management is a practice that is in growing development to address diseases in animals, allowing for a reduction in the use of conventional zootherapeutics. Regarding honey bees colonies, it is considered the most effective approach to mitigate the damage caused by Varroa and minimize the use of acaricides (Imdorf et al., 2003; Noël et al., 2020; Jack and Ellis, 2021). Among the resistance behaviors exhibited by honey bees against Varroa, grooming holds particular significance (reviewed by Pritchard, 2016). Autogrooming is the ability of parasitized bees to dislodge mites by themselves, while allogrooming refers to this behavior with assistance from other bees (Boecking and Spivak, 1999; Pritchard, 2016; Mondet et al., 2020). Since, in a standard Langstroth hive with wooden bottom boards, the 50% of the mites fallen are still alive and can reinfest new bees (Lobb and Martin, 1997), the effectiveness of grooming behavior in controlling Varroa could be further increased if hives are equipped with bottom boards that prevent detached mites from reinfesting the colony (Harbo and Harris, 2004; Araneda and Calzadilla, 2011).
The aim of this study was to compare the effect of different bottom boards in V. destructor population and in colony strength.
The study was carried out from April to December, 2022 (autumn to spring in the southern hemisphere) at Campo Experimental No. 2 of the Facultad de Veterinaria, San José, Uruguay. An apiary with 55 colonies with sister queens in a single brood box (Langstroth hives) was established. Queens were obtained from a colony without prior selection in any sense in a region where beekeepers must control Varroa infestation throughout the year. Bee population and brood area were standardized on average 20,000 bees and 4,000 cells with brood (Delaplane et al., 2013). All colonies were treated with acaricide (Flumethrin) 80 days prior to the beginning of the experiment to minimize the infestation level with Varroa and the population dynamic during the assay was monitored.
The colonies were sampled five times (every sixty days), estimating the following parameters: bee population, brood area and Varroa infestation level. The worker bee population was estimated by counting the number of frames covered by bees and converting this to the total number of bees (Delaplane et al., 2013). Brood population was estimated as the percentage of comb face with brood and converted to number of brood cells (Delaplane et al., 2013). Finally, the infestation level with Varroa was analyzed as the percentage of phoretic mites in a sample of 200–300 bees. The mites were removed from the bees with ethanol 75%, and the percentage of infected bees was determined (Dietemann et al., 2013).
The consumption of honey reserves of the colonies was estimated monthly from April to August, when the rape (Brassica napus) crops bloomed, and the bees start collecting nectar significantly. To do this, each hive was weighed, and the weight of the roof, box, frames, and bottom board was subtracted. As the colonies required it, honey suppers were added to honey storage. To estimate the honey collected in spring, three harvests were done, weighing the frames with honey from each colony before and after honey extraction (Büchler et al., 2024).
Three different bottom boards were used. The tubular bottom boards were constructed as described in Brouard (2005). Briefly, ten tubes (450 mm in length and 32 mm in diameter) were positioned on a wooden frame (70 mm high) with an open surface matching the inner dimensions of the hive body (450 mm by 350 mm) and were spaced 3.5 mm each apart. These tubes were securely affixed beneath the open surface by two slats. The screen bottom board consisted of a wooden frame (450 mm by 420 mm and 40 mm high) with a metal screen (400 mm x 230 mm) with a 3 mm grid pattern in the central part (Pettis and Shimanuki, 1999). The sticky bottom board has a wooden frame and metal mesh that covers the entire bottom surface (440 x 370 mm) and has a tray below where a sheet of cardboard smeared with petroleum jelly is placed to retain fallen mites (Figure 1).

Figure 1 (A) Tubular bottom board, (B) Screen bottom board and (C) Sticky bottom board evaluated in the study.
The colonies were randomly allocated into five groups separated by twenty meters to minimize the drift between groups. Each group of colonies belonged to one of the following treatments according to the bottom board they had: 1) tubular bottom board (n=11), 2) screen bottom board (n=10), 3) sticky bottom board (n=11), 4) conventional bottom board with a permanent treatment with acaricide (Flumethrin, three applications throughout the study period) (positive control) (n=12), and 5) conventional bottom board (negative control) (n=11).
Generalized linear mixed models (GLMM) were used to assess the relationship between treatments (positive control, tubular, sticky and mesh bottom boards) and sampling (as fixed effects), with colony`s strength (bee and brood population), Varroa infestation levels and honey reserves as response variables (package {lme4}). The identity of the colonies was considered as repeated measures in the samplings as a random effect. In the case of bee and brood population, a GLMM with Poisson distribution and a log link function was used. In the case of V. destructor infestation levels, a GLMM with Gamma distribution and a log link function were used. In addition, the effects of the treatments were analyzed particularly at the beginning (to ensure equivalent groups of colonies, February, 2022) and at the end of the study (December, 2022). To do that, ANOVA and Scheffe test or Kruskal-Wallis and Mann-Whitney tests (corrected for multiple comparisons with Bonferroni test) were used when the variables fitted or not the assumptions of parametric statistics, respectively. Generalized linear models (GLM) were used to analyze the effect of the treatments on honey production as fixed effect and response variable, respectively, with a Gamma distribution and a log link function.
In all cases, R Studio software was used and p-values under 0.05 were considered statistically significant.
At the beginning of the experiment, all the colonies showed equal bee and brood population across the five groups (bee population Kruskal Wallis, p=0.93; brood population ANOVA, p=0.62). These parameters followed the natural dynamics of the colonies in this period of the year with higher bee and brood populations in early spring (Table 1; Figure 2). These parameters did not differ between treatments (Table 1).
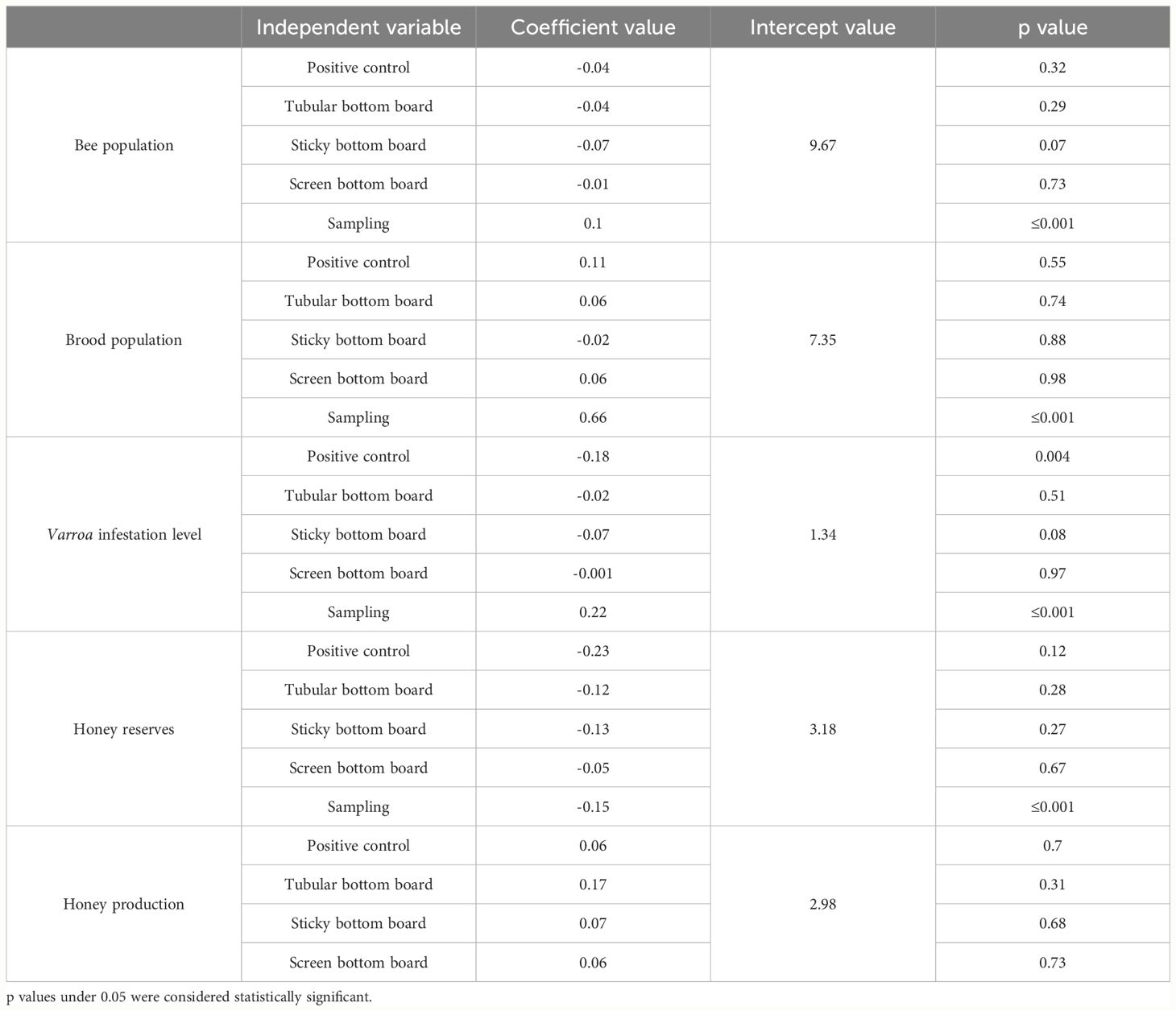
Table 1 Effect of treatment and sampling time on bee and brood population, on infestation level with Varroa destructor and in honey reserves, evaluated by generalized linear mixed models.
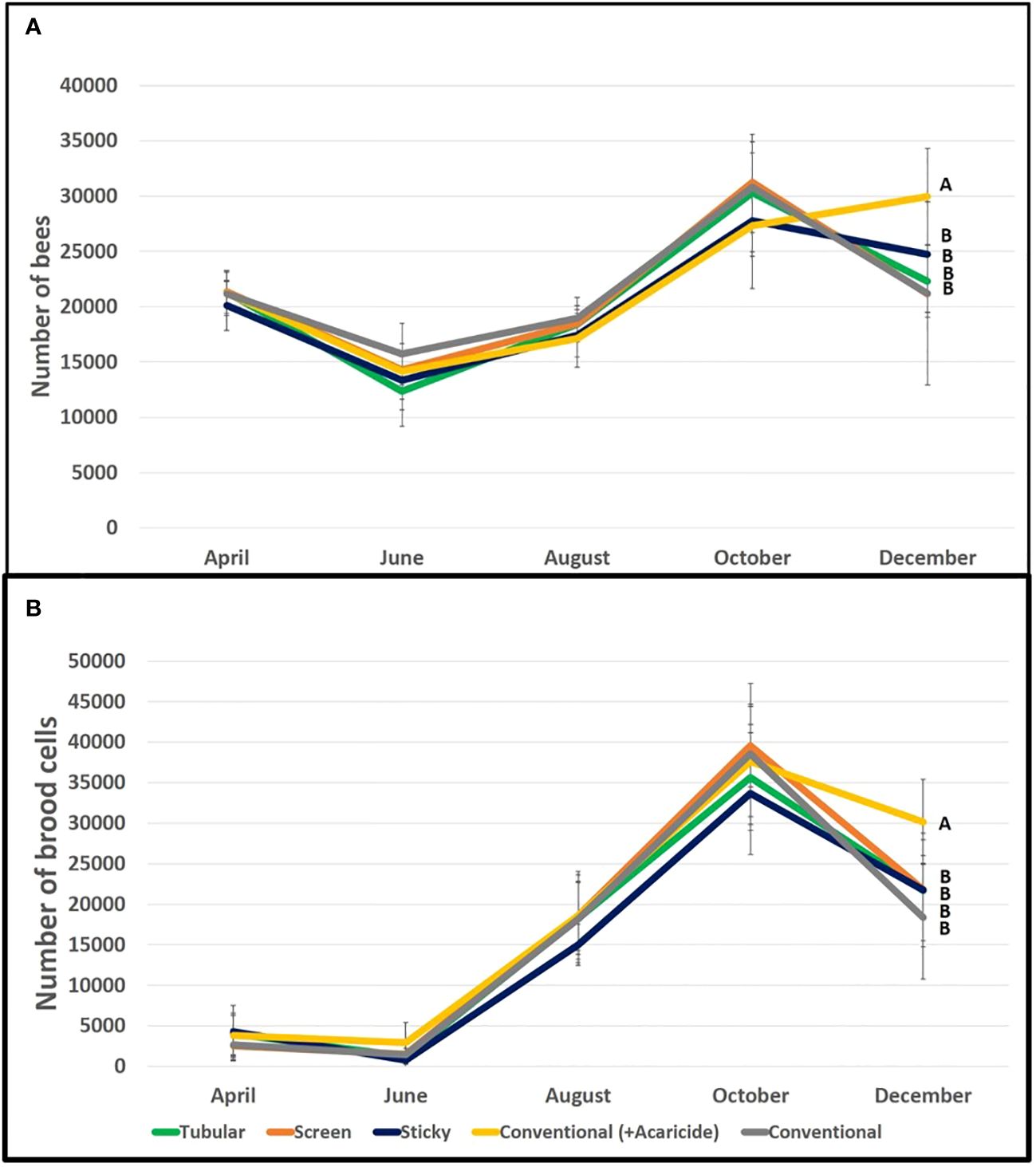
Figure 2 (A) Bee population in the colonies of different groups. Only statistical results of the final record are shown, the only one that showed significant differences. Different letters indicate significant differences (P < 0.05). (B) Brood population of the colonies in different groups. Only the statistical results of the final record are shown, the only one that showed significant differences. Different letters indicate significant differences (P < 0.05).
At the beginning of the experiment all the colonies had no Varroa and the infestation levels of this mite was close to zero between April and June. In addition, colonies treated with acaricides were Varroa-free throughout the entire observation period being significantly lower in comparison to the other treatment groups (Table 1; Figure 3). However, from August to December, Varroa population increased in the other groups (Table 1), with sticky bottom boards showing less mites (6%) in comparison to the negative control group (13%) and tubular bottom board (12%) (Table 1; December, Mann Whitney, negative control - sticky bottom board p=0.03, tubular bottom board - sticky bottom board p=0.01, screen bottom board - sticky bottom board p=0.99).
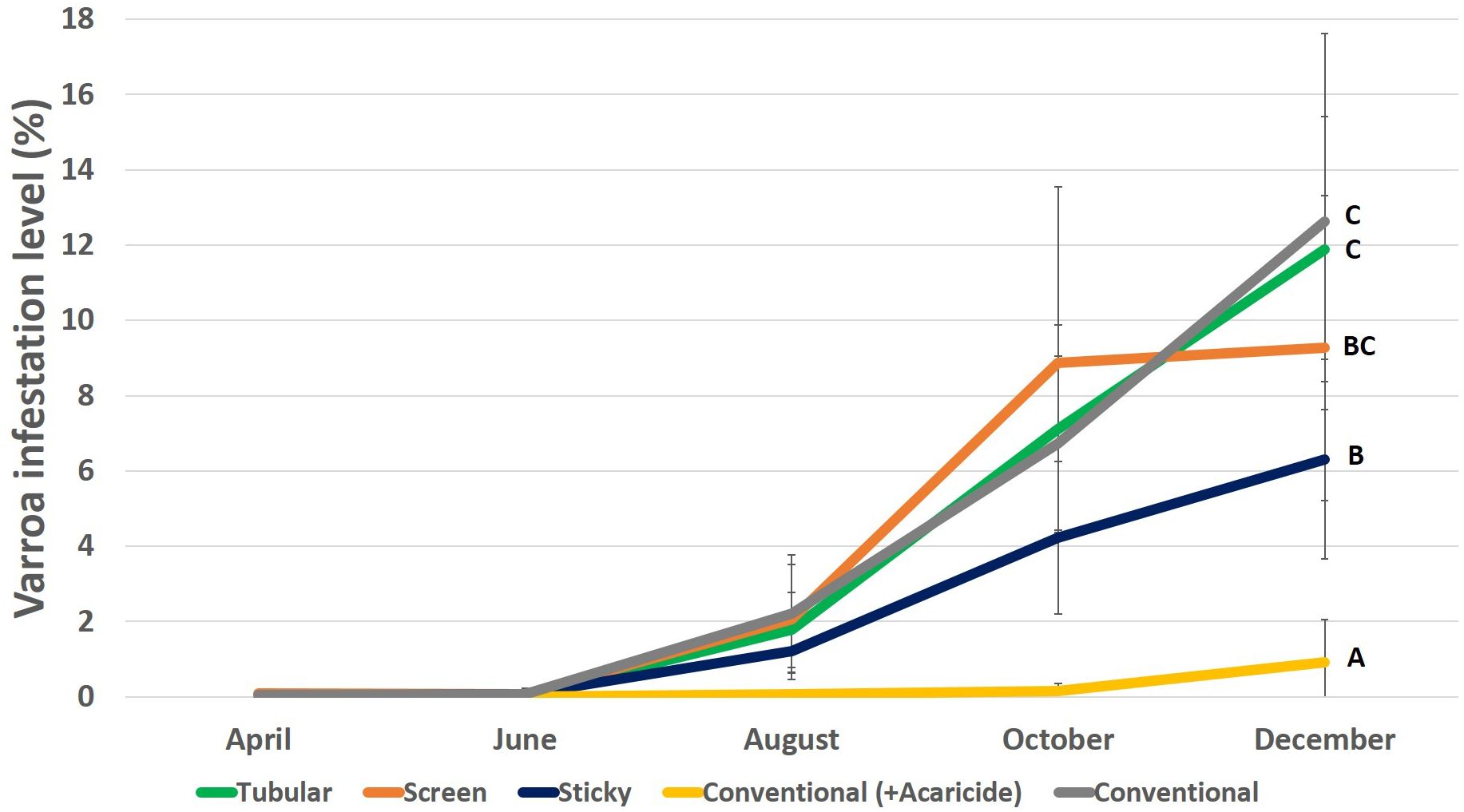
Figure 3 Varroa infestation level in the colonies of different groups. Only the statistical results of the final record are shown, in the others there were only differences between the conventional bottom board with acaricide (positive control) group and the other groups. Different letters indicate significant differences (P < 0.05).
The tubular bottom board, the screen bottom board and negative control groups showed similar infestation level with Varroa at the end of the experiment (December, Mann Whitney, tubular bottom board - screen bottom board p=0.99, tubular bottom board - negative control p=0.99, screen bottom board - negative control p=0.99) (Figure 3).
Honey reserves decreased over time in all the colonies from 18.3 ± 3.6 kg to 10.2 ± 3.6 kg, while treatments did not affect this parameter (Table 1) (Figure 4). Colonies that had a conventional bottom board and were treated with acaricides (positive control) presented fewer reserves from April to August (April, Scheffe, positive control - tubular bottom board p=0.03; May, Scheffe, positive control - tubular bottom board p=0.001, positive control - screen bottom board p=0.002, positive control - sticky bottom board p=0.003, positive control - negative control p=0.0001; June, Scheffe, positive control - negative control p=0.003; July, Scheffe, positive control - tubular bottom board p=0.002, positive control - screen bottom board p=0.005, positive control - sticky bottom board p=0.001, positive control - negative control p=0.0002; August, Scheffe, positive control - tubular bottom board p=0.01, positive control - sticky bottom board p=0.001, positive control - negative control p=0.0002). In the remaining comparisons, no significant differences were found (p>0.05) (Figure 4).

Figure 4 Honey reserves in the colonies of different groups of the colonies between April and August. Different letters indicate significant differences (P < 0.05).
Similar amount of honey was produced by the colonies from the different treatment groups between September to December, with an average of 21.4 ± 8.6 kg of honey per colony (Table 1).
Several studies have been conducted to explore alternative methods for Varroa control, aiming to include them in integrated pest management and minimize the use of acaricides (reviewed in van der Steen and Vejsnæs, 2021). However, despite these efforts, maintaining mite populations below the damage threshold for bee colonies remains challenging and costly. This study focused on the potential use of tubular, screen, and sticky bottom boards to limit the growth of mite populations to prevent detached mites from re-establish contact with the bees.
In this study, the tubular bottom boards and the screen bottom boards perform similarly to the conventional bottom boards, failing to significantly control the mite population growth. These results contrast with those found by Araneda and Calzadilla (2011) who reported that the screen bottom board and the tubular bottom board exhibited an effectiveness (ratio between varroas fallen in one month in the evaluated bottom boards and total varroas fallen in the following month after applying an acaricide) of 20% and 16%, respectively. Pettis and Shimanuki (1999) showed that Varroa infestation level did not experience noteworthy reductions in colonies equipped with modified screen bottom boards, which is in line with the results obtained in this study.
In contrast to this research, where no significant differences were found in bee population and brood area, the studies by Harbo and Harris (2004) reports that the introduction of screen bottom board in the colonies led to an increase in brood. Sammataro et al. (2004) found that screen bottom boards are most effective in reducing Varroa infestation when used in colonies with selected hygienic queens. Recently, Liu et al. (2020) conducted a meta-analysis from several studies that included 145 colonies and found that the Varroa population in colonies with screen bottom boards is significantly lower compared to those with conventional bottom boards.
The use of the sticky bottom board seems to be a promising strategy to be implemented in integrated pest management programs, since it allowed to limit Varroa infestation levels by nearly 50% in comparison to the conventional boards. Pettis and Shimanuki (1999) also evaluated sticky bottom boards, but only obtained a 28% reduction in mite infestation in an intermediate sampling, without preventing infection levels above the economic threshold. However, the limitations of these sticky bottom boards must be considered since in December the colonies reached a significant level of infestation (6%), enough to begin to affect them.
The difference in effectiveness in limiting Varroa growth between the three analyzed bottom boards could be due to the percentage of surface area where the detached mites would exit the hive. In tubular bottom board this space is approximately 10% of the surface, so many of the detached mites will fall on the tubes and might reinfest the bees. The sticky and screen bottom boards have a 3 mm mesh where the wires occupy a smaller proportion of the surface. In this case the difference between both bottom boards may not lie in the use of an adhesive to retain the fallen mites, but in the surface of the mesh, which in the screen bottom boards is approximately 60% of that of the sticky bottom boards. The results obtained in colonies with screen bottom boards in the last sampling, where Varroa infestation does not increase, is striking. Perhaps with a high mite load the differences between the screen and the sticky bottom boards will be reduced. These factors justify future studies to optimize the design of the devices.
This study shows significant differences in Varroa infestation levels, particularly when mite populations were high. A decline in colony strength parameters (bee and brood population) was observed across all groups in comparison to the treated group of colonies. A phenomenon resembling the “Parasitic Mite Syndrome,” characterized by scattered brood and bee loss as described by Rosenkranz et al. (2010), was evident in certain colonies. It is conceivable that if the trial had extended over a longer period, many colonies would have eventually depopulate and collapse due to this syndrome as Varroa infestation increased.
Two of the evaluated bottom boards, the tubular and the screen bottom boards, allow the entry of air from the environment, which could lead to higher consumption of honey reserves to maintain the appropriate temperature in the brood nest during the autumn and winter. However, this phenomenon was not observed, so the use of these bottom boards does not imply a cost in honey reserves. The greater consumption of reserves of the colonies treated with acaricides with conventional bottom boards in relation to the other colonies is an unexpected result that it is difficult to explain since this group of colonies did not differ from the others in bee population and brood area. On the other hand, the colonies of the different groups did not differ in honey production. Therefore, the use of the three bottom boards evaluated does not negatively affect the consumption of reserves or honey production.
Previous studies have shown that mite-resistant bee populations exhibit higher grooming and hygienic behaviors compared to mite-susceptible colonies (Guzman-Novoa et al., 2012; Hunt et al., 2016; Locke, 2016; Mendoza et al., 2020; Russo et al., 2020; Mendoza et al., 2022). The benefit of these bottom boards, especially the sticky ones, can be enhanced when populations of bees selected for high grooming behavior or Varroa resistance characteristics are obtained. The combination of these strategies can be synergistic, thus achieving effective mite control with minimal acaricide use. Therefore, further investigation of these designed bottom boards is needed to assess their effectiveness under such conditions.
In conclusion this study shows a significant effectiveness of the sticky bottom board in limiting Varroa population growth, particularly when mite infestation levels are high. The use of an adhesive to retain detached mites offers a practical and promising approach to managing high Varroa levels and complement other strategies for the mite control in integrated pest management programs.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
The manuscript presents research on animals that do not require ethical approval for their study.
PJ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. EN: Conceptualization, Investigation, Validation, Visualization, Writing – review & editing. JA: Investigation, Methodology, Validation, Visualization, Writing – review & editing. VR-B: Investigation, Methodology, Validation, Visualization, Writing – review & editing. BB: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. CI: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was carried out thanks to funding from the Instituto Nacional de Investigación Agropecuaria (project FPTA 370) and a postgraduate scholarship from Universidad de la República for Pablo Juri (Master’s scholarship granted by the Comisión Académica de Posgrado).
The authors would like to express their gratitude to the staff of Campo Experimental No. 2 at the Facultad de Veterinaria, Universidad de la República, for their permission to conduct the study on their premises. Also want to thank other collaborators and researchers who have worked on this global study and are not part of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aldea P., Bozinovic F. (2020). The energetic and survival costs of Varroa parasitism in honeybees. Apidologie 51, 997–1005. doi: 10.1007/s13592-020-00777-y
Amdam G. V., Hartfelder K., Norberg K., Hagen A., Omholt S. W. (2004). Altered physiology in worker honey bees (Hymenoptera: Apidae) infested with the mite Varroa destructor (Acari: Varroidae): A factor in colony loss during overwintering. J. Econ. Entomol. 97, 741–747. doi: 10.1603/0022-0493(2004)097[0741:APIWHB]2.0.CO;2
Annoscia D., Brown S. P., Di Prisco G., De Paoli E., Del Fabbro S., Frizzera D., et al. (2019). Haemolymph removal by Varroa mite destabilizes the dynamical interaction between immune effectors and virus in bees, as predicted by Volterra’s model. Proc Biol Sci. 286(1901), 20190331. doi: 10.1098/rspb.2019.0331
Araneda X., Calzadilla A. (2011). Evaluación de dos modelos de pisos trampa para el control del ácaro Varroa destructor Oud. sobre la abeja Apis mellifera L. Idesia 29, 99–104. doi: 10.4067/S0718–34292011000300015
Beaurepaire A., Piot N., Doublet V., Antunez K., Campbell E., Chantawannakul P., et al. (2020). Diversity and global distribution of viruses of the western honey bee, Apis mellifera. Insects 11, 239. doi: 10.3390/insects11040239
Boecking O., Spivak M. (1999). Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidologie 30, 141–158. doi: 10.1051/apido:19990205
Brouard I. (2005). Trial report on the Happykeeper bottom board. Exploitation du Lycée agricole de Toulouse-Auzeville. E.P.L.E.A. Toulouse-Auzeville 25.
Büchler R., Andonov S., Bernstein R., Bienefeld K., Costa C., et al. (2024). Standard methods for rearing and selection of Apis mellifera queens 2.0. J. Apic. Res. 63, 65–75. doi: 10.1080/00218839.2023.2295180
Currie R. W., Gatien P. (2006). Timing acaricide treatments to prevent Varroa destructor (Acari: Varroidae) from causing economic damage to honey bee colonies. Can. Entomol. 138, 238–252. doi: 10.4039/n05-024
Delaplane K., van der Steen J., Guzman-Novoa E. (2013). Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 52, 1–12. doi: 10.3896/IBRA/1.52.1.03
De Miranda J. R., Genersch E. (2010). Deformed wing virus. J. Invertebr. Pathol. 103, S48–S61. doi: 10.1016/j.jip.2009.06.012
Dietemann V., Nazzi F., Martin S., Anderson D., Locke B., et al. (2013). Standard methods for varroa research. J. Apic. Res. 52, 1–54. doi: 10.3896/IBRA.1.52.1.09
Elzen P. J., Baxter J. R., Spivak M., Wilson W. T. (2000). Control of Varroa jacobsoni Oud resistant to fluvalinate and amitraz using coumaphos. Apidologie 31, 437–441. doi: 10.1051/apido:2000134
Emsen B., Guzman-Novoa E., Kelly P. G. (2014). Honey production of honey bee (Hymenoptera: Apidae) colonies with high and low Varroa destructor (Acari: Varroidae) infestation rates in eastern Canada. Can. Entomol. 146, 236–240. doi: 10.4039/tce.2013.68
Fries I., Hansen H., Imdorf A., Rosenkranz P. (2003). Swarming in honey bees (Apis mellifera) and Varroa destructor population development in Sweden. Apidologie 34(4), 389–397. doi: 10.1051/apido:2003032
Guzman-Novoa E., Emsen B., Unger P., Espinosa Montaño L. G., Petukhova T. (2012). Genotypic variability and relationships between mite infestation levels, mite damage, grooming intensity, and removal of Varroa destructor mites in selected strains of worker honey bees (Apis mellifera L.). J. Invertebr. Pathol. 110, 314–320. doi: 10.1016/j.jip.2012.03.020
Harbo J. R., Harris J. W. (2004). Effect of screen floors on populations of honey bees and parasitic mites (Varroa destructor). J. Apic. Res. 43, 114–117. doi: 10.1080/00218839.2004.11101120
Hunt G., Given J. K., Tsuruda J. M., Andino G. K. (2016). Breeding mite-biting bees to control Varroa. Bee Culture 8, 41–47.
Imdorf A., Charriere J. D., Kilchenmann V., Bogdanov S., Fluri P. (2003). Alternative strategy in central Europe for the control of Varroa destructor in honey bee colonies. Apiacta 38, 258–278.
Jack C. J., Ellis J. D. (2021). Integrated pest management control of Varroa destructor (Acari: Varroidae), the most damaging pest of Apis mellifera L. (Hymenoptera: Apidae) colonies. J. Insect Sci. 21, 6. doi: 10.1093/jisesa/ieab058
Liu F., Xu X., Zhang Y., Zhao H., Huang Z. Y. (2020). A meta-analysis shows that screen bottom boards can significantly reduce Varroa destructor population. Insects 11, 624. doi: 10.3390/insects11090624
Lobb N., Martin S. (1997). Mortality of Varroa jacobsoni Oudemans during or soon after emergence of worker and drone honeybees Apis mellifera L. Apidologie 28, 367–374. doi: 10.1051/apido:19970604
Locke B. (2016). Natural Varroa mite-surviving Apis mellifera honeybee populations. Apidologie 47, 467–482. doi: 10.1007/s13592-015-0412-8
Maggi M. D., Ruffinengo S. R., Mendoza Y., Ojeda P., Ramallo G., et al. (2011). Susceptibility of Varroa destructor (Acari: Varroidae) to synthetic acaricides in Uruguay: Varroa mites potential to develop acaricide resistance. Parasitol. Res. 108, 815–821. doi: 10.1007/s00436-010-2122-5
Mendoza Y., Santos E., Clavijo-Baquett S., Invernizzi C. (2022). A reciprocal transplant experiment confirmed mite-resistance in a honey bee population from Uruguay. Vet. Sci. 9, 596. doi: 10.3390/vetsci9110596
Mendoza Y., Tomasco I. H., Antúnez K., Castelli L., Branchiccela B., et al. (2020). Unraveling honey bee-Varroa destructor interaction: multiple factors involved in differential resistance between two Uruguayan populations. Vet. Sci. 7, 116. doi: 10.3390/vetsci7030116
Mitton G., Meroi Arcerito F., Cooleyc H., Fernández de Landa G., Eguaras M., Ruffinengo S. R., et al. (2022). More than sixty years living with Varroa destructor: a review of acaricide resistance. Int. J. Pest Manag. 1–18. doi: 10.1080/09670874.2022.2094489
Mondet F., Beaurepaire A., McAfee A., Locke B., Alaux C., et al. (2020). Honey bee survival mechanisms against the parasite Varroa destructor: A systematic review of phenotypic and genomic research reports. Int. J. Parasitol. 50, 433–447. doi: 10.1016/j.ijpara.2020.03.005
Nazzi F., Le Conte Y. (2016). Ecology of Varroa destructor, the major ectoparasite of the Western honey bee, Apis mellifera. Annu. Rev. Entomol. 61, 417–432. doi: 10.1146/annurev-ento-010715-023731
Noël A., Le Conte Y., Mondet F. (2020). Varroa destructor: how does it harm Apis mellifera honey bees and what can be done about it? Emerg. Top. Life Sci. 4, 45–57. doi: 10.1042/ETLS20190125
Pettis J., Shimanuki H. (1999). A hive modification to reduce varroa populations. Am. Bee J. 139, 471–473.
Pritchard D. J. (2016). Grooming by honey bees as a component of varroa resistant behavior. J. Apic. Res. 55, 38–48. doi: 10.1080/00218839.2016.1196016
Ramsey S. D., Ochoa R., Bauchan G., Gulbronson C., Mowery J. D., et al. (2019). Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. U.S.A. 116, 1792–1801. doi: 10.1073/pnas.1818371116
Rosenkranz P., Aumeier P., Ziegelmann B. (2010). Biology and control of Varroa destructor. J. Invertebr. Pathol. 103, S96–119. doi: 10.1016/j.jip.2009.07.016
Russo R. M., Liendo M. C., Landi L., Pietronave H., Merke J., et al. (2020). Grooming behavior in naturally Varroa-resistant Apis mellifera colonies from north-central Argentina. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.590281
Sammataro D., Hoffman G. D., Wardell G., Finley J., Ostiguy N. (2004). Testing a combination of control tactics to manage Varroa destructor (Acari: Varroidae) population levels in honey bee (Hymenoptera: Apidae) colonies. Int. J. Acarol. 30, 71–76. doi: 10.1080/01647950408684371
Traynor K. S., Mondet F., de Miranda J. R., Techer M., Kowallik V., et al. (2020). Varroa destructor: A complex parasite, crippling honey bees worldwide. Trends Parasitol. 36, 592–606. doi: 10.1016/j.pt.2020.04.004
van der Steen J., Vejsnæs F. (2021). Varroa control: A brief overview of available methods. Bee World 98, 50–56. doi: 10.1080/0005772X.2021.1896196
Wilfert L., Long G., Leggett H. (2016). Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science 351, 594–597. doi: 10.1126/science.aac9976
Keywords: Varroa destructor, colony losses, bottom boards, integrated pest management, beekeeping innovations, beekeeping sustainability
Citation: Juri P, Nogueira E, Anzola J, Rodríguez-Batista V, Branchiccela B and Invernizzi C (2024) Evaluation of three different bottom boards in honeybee hives for the control of Varroa destructor. Front. Bee Sci. 2:1384846. doi: 10.3389/frbee.2024.1384846
Received: 11 February 2024; Accepted: 07 June 2024;
Published: 02 July 2024.
Edited by:
M. Marta Guarna, Agriculture and Agri-Food Canada (AAFC), CanadaReviewed by:
Alejandra Carla Scannapieco, National Scientific and Technical Research Council (CONICET), ArgentinaCopyright © 2024 Juri, Nogueira, Anzola, Rodríguez-Batista, Branchiccela and Invernizzi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pablo Juri, cGp1cmk4QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.