
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Bee Sci., 03 April 2024
Sec. Bees in Pollination
Volume 2 - 2024 | https://doi.org/10.3389/frbee.2024.1383589
 Herbeson O. J. Martins1*
Herbeson O. J. Martins1* Marcos C. Dórea2Vinina S. Ferreira3Cláudia I. Silva4
Marcos C. Dórea2Vinina S. Ferreira3Cláudia I. Silva4 Isabel Alves-dos-Santos5
Isabel Alves-dos-Santos5Plant-pollinator interactions constitute complex mutualistic relationships responsible for the maintenance of the reproduction of flowering plants in different biomes. The genus Centris stands out as one of the main groups of pollinators in the Neotropical region. These bees have a close relationship with floral oil-producing plants, using this resource in the construction of their nests and/or larval feeding. The aim of the present study was to compare the resources exploited by Centris perforator and Centris xanthomelaena and assess the established trophic relationships of these two species in a fragment of dry forest (Caatinga domain). For such, direct observations were made at the flowers and analyses were performed of the pollen load on the body of female bees and the content of the larval provisions in the nests. A strong association was found between the two bee species and plants with poricidal anthers of the genus Chamaecrista, which accounted for more than half of the larval diet in both species. Samples of the pollen load of C. xanthomelaena exhibited a greater diversity of pollen types. While Rhaphiodon echinus was a constant source of nectar for C. xanthomelaena, C. perforator females collected nectar from plants of short, rapid flowering, such as Cenostigma microphyllum and Tabebuia aurea. Malpighia emarginata was the main source of floral oil for both species. However, C. xanthomelaena also interacted with Angelonia cornigera and Krameria sp. to obtain this resource. The similarity in the trophic niche of the species was 57% in the subsamples of pollen on the body of females and 46% for larval provision. The results showed that areas of native vegetation are essential for bee populations in regions with agricultural activity, ensuring a regular supply of ecological resources for these bees.
For an estimated 90% of flowering plants, including diverse species of crops, depend on the pollination service provided by animals (Ollerton et al., 2011). The mutualistic relationships established between plants and pollinators are critical to the maintenance of ecosystems and are likely one of the most important animal-plant interactions (Klein et al., 2007; Ollerton et al., 2011). Bees (Apiformes) play a central role in this context, with solitary species constituting an important component of the community of pollinating insects (Roubik, 1992). The stability of these interactions is susceptible to anthropogenic changes and can be negatively impacted by numerous factors, such as habitat fragmentation (Aizen and Feinsinger, 2003; Xiao et al., 2016). Thus, studies on plant-pollinator interactions can contribute to the understanding of the dynamics of ecosystems.
The guild of oil-collecting bees and the plants that produce this resource was first studied by Vogel in 1974 and soon drew the attention of other authors (Vogel, 1974; Neff and Simpson, 1981; Buchmann, 1987; Cocucci, 1991; Vogel and MaChado, 1991; Neff and Simpson, 2017). The collection of floral oil by bees evolved independently in groups belonging to six different tribes, with Centridini, Tapinostapidini and Tetrapediini representatives of the Neotropical region (Renner, 2006; Alves-dos-Santos et al., 2007; Moure et al., 2007). Females of these bees have morphological adaptations on the forelegs and/or middle legs that enable oil collection from structures named elaiophores, which may be trichomal or epithelial (Vogel, 1974; Neff and Simpson, 1981; Buchmann, 1987). Some of these species are adapted to oil collecting from a specific type of elaiophore, whereas others are able to exploit this resource from both types (Neff and Simpson, 1981). Floral oil collected by females is a resource of high energy value mixed with pollen and nectar in larval provision and/or used as a lining for brood cells (Vogel, 1974; Simpson et al., 1977; Cane et al., 1983; Buchmann, 1987; Sabino et al., 2019).
Within the tribe Centridini, the genus Centris Fabricius is composed of 14 subgenera and approximately 332 species distributed in the Neotropical region (Silveira et al., 2002; Moure et al., 2007; Vivallo, 2020, Vivallo, 2022a, Vivallo, 2022b; Vivallo et al., 2023). Centris females can nest on different substrates, but ground nesting is the predominant habit in this genus (Coville et al., 1983; Camillo et al., 1995; Aguiar et al., 2005; Martins et al., 2020a). Various morphological, physiological and behavioral characteristics make Centris bees stand out as the main pollinators of plants with mass flowering and species of economic interest, such as the West Indian cherry (Malpighia emarginata DC.) (Frankie et al., 1983; Schlindwein, 2000; Vilhena and Augusto, 2007; Vilhena et al., 2012; Silva et al., 2017). However, available information on the floral resources necessary to maintain natural populations of Centris species remains scarce, especially for ground-nesting species. For instance, the pollen content of larval provisions was analyzed for Centris (Centris) flavifrons (Fabricius) (Rêgo et al., 2006; Dórea et al., 2017), Centris (Centris) caxiensis Ducke (Ribeiro et al., 2008); Centris (Melacentris) conspersa Mocsáry (Rocha-Filho et al., 2017), Centris (Paracentris) burgdorfi Friese (Sabino et al., 2019) and Centris (Melacentris) collaris Lepeletier (Rocha-Filho et al., 2018), which correspond to approximately 2% of the species of the genus.
Although data on plants visited by bees are traditionally obtained through direct observations of flowers (e.g., Aguiar et al., 2003), palynology become increasingly important in this scenario, providing more detailed information on the interactions between bees and plants (Silva et al., 2020). A grain of pollen serves as an important taxonomic marker (Erdtman, 1960); thus, analyses can be performed of the pollen deposited on the scopa/corbicula of females, the provisions stocked in the brood cells and even larval feces (Dórea et al., 2010a, Dórea et al., 2010b; Silva et al., 2017).
As most floral visits by bees have the objective of collecting resources for feeding the offspring, palynological analyses of the provisions in nests can provide more in-depth information on the true spectrum of plants visited by females throughout their lives (Dó́rea et al., 2017; Silva et al., 2017). With regards to solitary bees, however, palynology has been used for provisions of few species, mainly due to the difficulty in finding the nests, especially of those that nest in the ground (Sabino et al., 2019). An alternative to the use of larval provisions has been found in the analysis of the pollen load on females, as the pollen collected from anthers is directed to the scopa/corbicula, transported to the nest and offered as larval provision. However, grains of pollen can also remain adhered to different parts of the body of females and are responsible for the reproduction of plants through the pollination process (Rech et al., 2014). Analyses of the pollen load adhered to the body of females has proved to be equally efficient for determining the trophic niche of these species, even in the absence of the nests (Silva et al., 2010, Sabino et al., 2019).
Centris (Relicthemisia) xanthomelaena Moure and Castro and Centris (Trachina) perforator Smith are two species broadly distributed in the Caatinga domain (dry forest) of Brazil (Zanella, 2000; Vivallo, 2020) and abundant in the study area. Centris xanthomelaena nests in a dense aggregation while C. perforator nests widely scattered (Martins et al., 2020a, Martins et al., 2020b), and occur in a sympatric manner. Considering the overlap in the period of activity, the principal aim of the present study was to compare the floral resources exploited by C. perforator and C. xanthomelaena and assess the trophic relationships established between the two species in a fragment of the Caatinga domain.
The sampling area of the study is located in a fragment of the Caatinga situated on the Campus of Agrarian Sciences of Universidade Federal do Vale do São Francisco (UNIVASF) (9°19’44,2’’S, 40°33’30,1W) in the municipality of Petrolina, state of Pernambuco, Brazil (Figure 1A). The Caatinga is one of the phytogeographical domains of Brazil. It has a high level of endemism and is classified as a wooded steppe savanna (Andrade-Lima, 1981). The climate in the region is considered semiarid, dry and warm (BSh according to the Köppen classification), with low rainfall (annual average of 433 mm) and two well-defined seasons: dry from May to October and rainy from November to April. The annual average temperature is 24.8°C (Alvares et al., 2013).
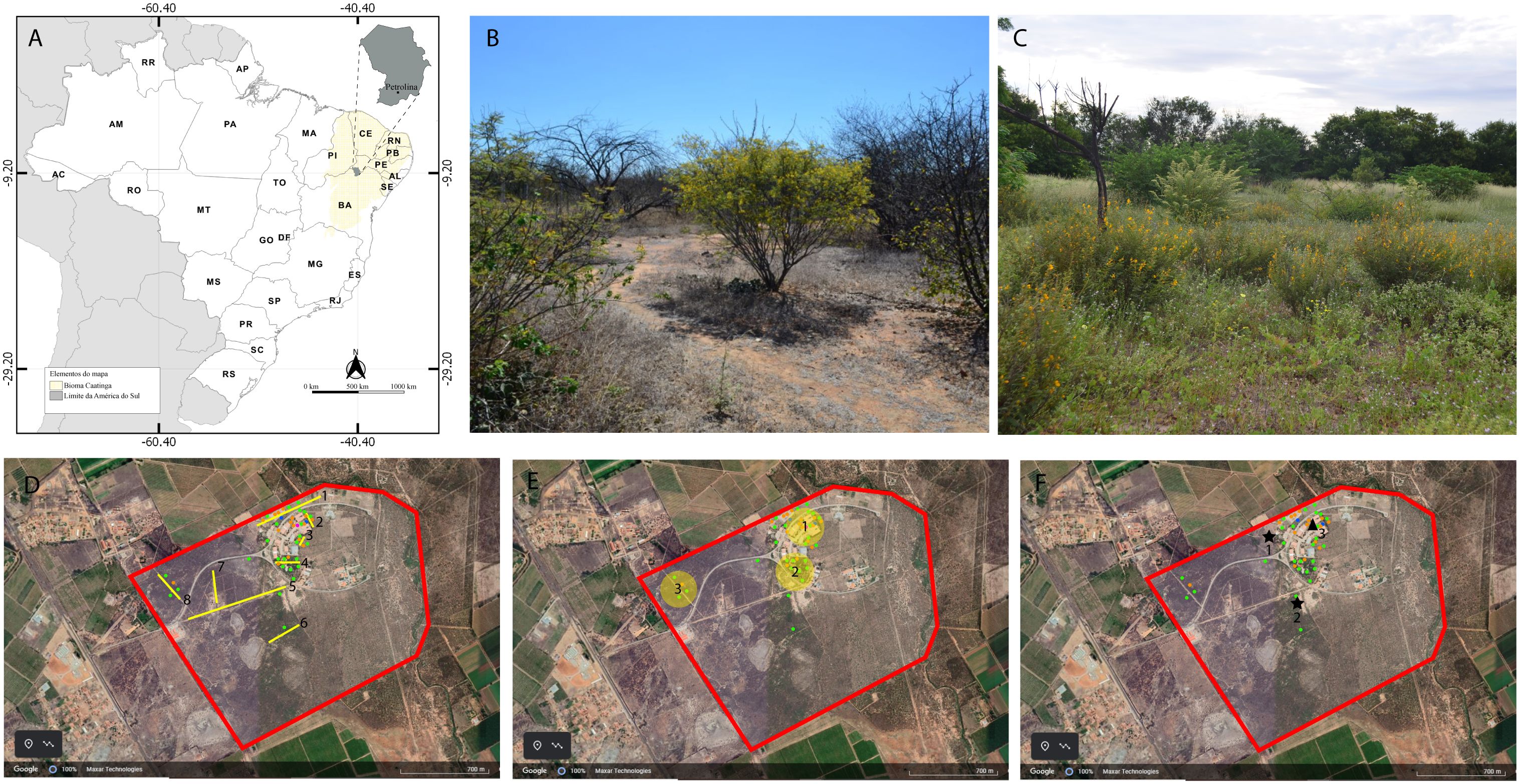
Figure 1 (A) Study area where samples were collected in the Caatinga environment, (B) open shrub layer with Cenostigma microphyllum blooming during the dry season, (C) herbaceous layer with the presence of Chamaecrista, (D–F) aerial view delimiting the Caatinga fragment area (red line): (D) transects traveled (the lines in yellow) to observe the plants visited by the species, (E) places where pollen load collections were carried out, (F) places where the brood cell samples were obtained (★ 1 and 2 represent the aggregations of C. xanthomelaena and ▲ represents the main nesting site for C. perforator).
The area is in the Senator Nilo Coelho Irrigation Project (C1), which is one of the main fruit farming centers in Brazil (Bustamante, 2009). Orchards of plants of agricultural interest surround the area, such as mangos (Mangifera indica L.), grapes (Vitis vinifera L.), coconut trees (Cocos nucifera L.) and acerola (West Indian cherry) (Malpighia emarginata DC.) (Figure 1).
The vegetation in the area is classified as low Caatinga (Andrade-Lima, 1981), with a predominance of bush vegetation, such as Cenostigma microphyllum (Mart. ex G. Don) Gagnon and G.P.Lewis and Senegalia tenuiflora (L.) Britton and Rose (Figure 1B), as well as an herbaceous stratum composed mainly of species of Lamiaceae, Leguminosidae, Malvaceae and Rubiaceae (Figure 1C), with the presence of large trees being rare.
Monthly samples of bees were collected between November 2017 and October 2018 to record plants visited by the species. Collections were performed on two consecutive days from 6:00 to 17:00 h, totaling 22 hours per month, in eight preestablished transects of different distances (Figure 1D). One collector carefully inspected each flowering plant for maximum five minutes and both female and male bees were caught randomly with an insect net during visits to the flowers. Whenever possible, the resource collected by the bee was observed. To avoid recapture, bees were marked on the thorax with a nontoxic pen (UniPosca ®) and released. Specimens of both species were sacrificed, mounted and deposited in the Entomological Collection of the UNIVASF Entomology Lab.
To characterize the composition of the pollen load on the females, monthly visits were performed in two perimeters (Figure 1E) on two consecutive days between the years 2018 and 2021. In this period, a maximum of five samples were collected per month based on the presence of females, totaling 60 samples (30 for each species). Females with pollen adhered to the scopa were caught with an insect net during flight. The pollen on the scopa was removed using a rinsing process following the method proposed by Silva et al. (2010) and transferred to 15-ml Falcon test tubes containing 1.5 ml of distilled water. The females were then dried with paper towel, marked on the thorax with nontoxic pens (UniPosca ®) of different colors to avoid recapture and released. The pollen removed was stored in 70% alcohol until acetolysis.
To understand the composition of the larval diet, samples of larval provisions were collected from 60 brood cells (30 from each species) from 30 nests of C. xanthomelaena and 27 nests of C. perforator (Figure 1F) between the years 2017 and 2021. Due to the low availability of C. perforator nests, no minimum number of samples per month was established for this species. Additionally, more than one brood cell was used of the nests built by two C. perforator females that had more than 12 cells/nest, with cells selected from different stages of construction of the nests. All samples were collected from fresh provisions soon after the conclusion of the nests by the females and stored in 70% alcohol until acetolysis.
The pollen samples were submitted to chemical treatment (acetolysis), as proposed by Erdtman (1960). At the end of the process, slides were mounted for optical microscopic analysis. Four slides were mounted with Kisser glycerol gelatin (Kisser, 1935) for each sample. After mounting, qualitative analysis was performed for the identification of the pollen types on each slide and quantitative analysis was performed, with counts of 1000 grains of pollen per sample (250 per slide) (Montero and Tormo, 1990). Pollen types that appeared at a frequency of less than 0.01% were considered contaminants and excluded from the quantitative analysis.
Based on the quantitative analysis, the frequency of occurrence was calculated for the different types of pollen in each sample and mean frequencies were calculated for each type among the samples (Villanueva-Gutiérrez and Roubik, 2004; Silva, 2009). The percentage of each pollen type was determined and categorized using the classification proposed by Jones and Bryant (1996): very frequent (> 50% of all pollen on the slide), frequent (20 to 50%), infrequent (10 to 20%) and rare (< 10%).
In the period from 2018 to 2019, monthly monitoring was performed of plants in bloom in the study area (UNIVASF Campus of Agrarian Sciences), with a total of 153 species sampled. A reference slide collection was created of the pollen obtained from the flowers of the species, following the method proposed by Erdtman (1960). The pollen collection contributed to a more precise identification of the pollen types in the samples from the bees. Vouchers of the plants were identified by specialists and deposited in the Vale do São Francisco Herbarium.
The Shannon-Weaver diversity index (H’) (Shannon, 1948) was used to assess the diversity of plants that composed the diet, considering the richness and abundance of pollen types. The uniformity of the floral resources was tested by calculating Pielou’s evenness index (J’) (Pielou, 1977). The dominance (D) of pollen types was determined using the Berger–Parker index (Magurran, 2004). The number of the most important pollen types in the diet of the species was calculated from the apparent S (apparent S = ). The analysis of trophic niche overlap of the two species was performed using the Jaccard similarity index (Sj) and the proportion of plant species in the diets was determined using the percentage similarity index (PS). (The formulas are available in the Supplementary Data 1).
The bipartite package of the R program 3.4.1 (R Core Team, 2017) was used for the representation of interaction networks in Eulerian graphs. To visualize the relationship of the plants on the genus level and the bee species, a weighted network was created through the analysis of quantitative matrices using the Pajek program version 5.13 (De Nooy et al., 2018) and the ‘Kamada-Kawai free’ method, making the subgroups and centrality of each vertex more evident.
A total of 433 individuals of the two species were collected: 209 specimens of Centris perforator and 224 specimens of Centris xanthomelaena (Table 1). Together both species visited 32 plant species (C. perforator = 22 species and C. xanthomelaena 19 species). Leguminosae was the botanical family with the largest number of interactions (n = 274), the majority of which involved the subfamily Caesalpinioideae (n = 226). Lamiaceae was the family with the second largest number of interactions (n = 104), despite the small number of species recorded in the area.
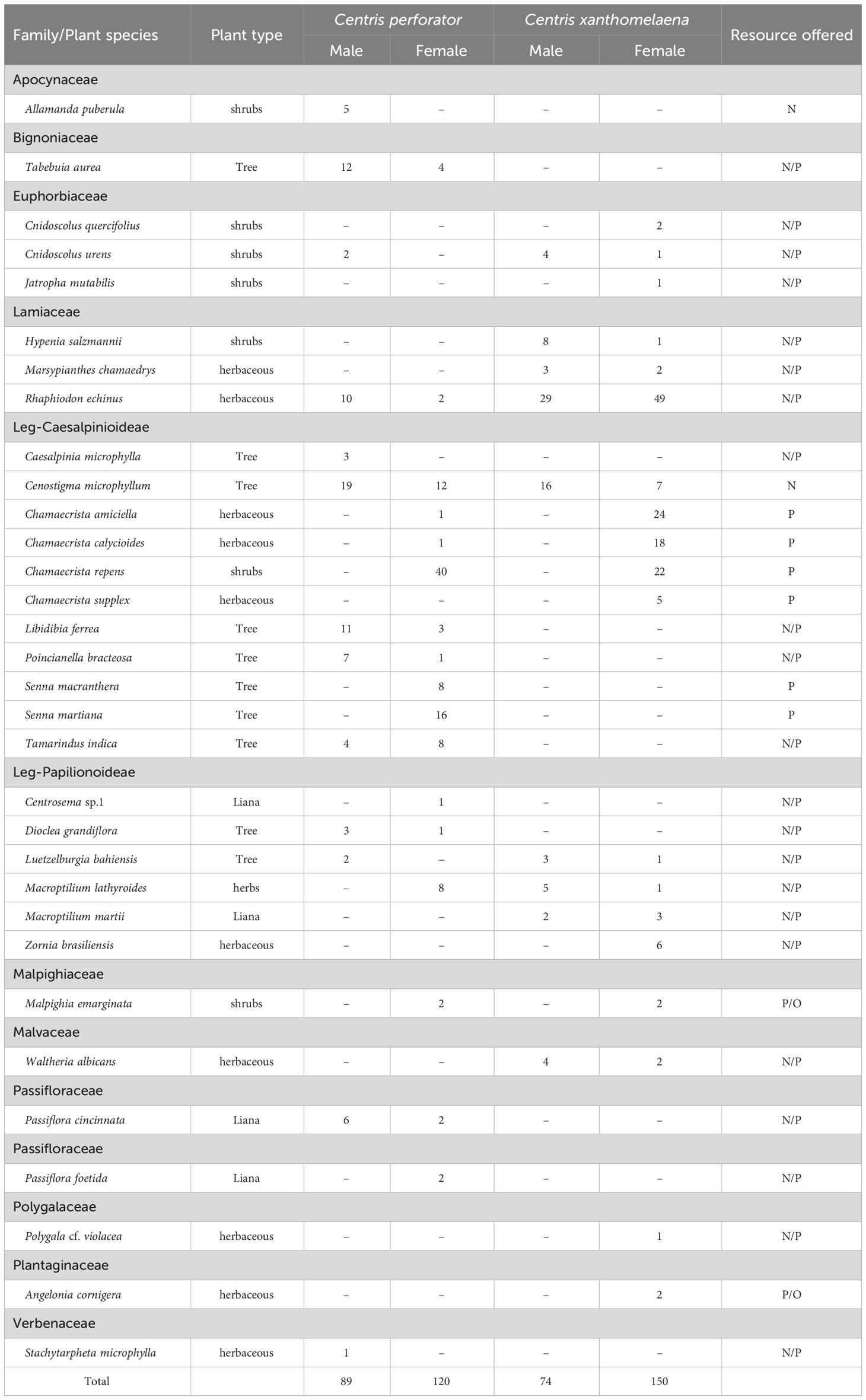
Table 1 Plants visited by females and males of Centris perforator and Centris xanthomelaena in a fragment of Caatinga in the municipality of Petrolina, PE, Brazil, and main resource offered (P = pollen; N = nectar; O = floral oil).
Centris perforator females interacted with the flowers of 19 plant species and males visited 14 species. The largest part of the pollen sources used for larval feeding was nearly exclusively from plants of the family Leguminosae, with Chamaecrista and Senna as the genera with the largest number of visits (n = 73 and 24, respectively). Cenostigma microphyllum (Mart. ex G.Don) Gagnon and G.P.Lewis was the main source of nectar visited by both male and females of C. perforator. However, Rhaphiodon echinus (Nees and Mart.) Schauer and Libidibia ferrea (Mart. ex Tul.) L.P.Queiroz also received a number of visits by males of the species. The only source of floral oil visited by the females was Malpighia emarginata, which was also used sporadically as a source of pollen. During the flowering period of Tabebuia aurea (Silva Manso) Benth. and Hook.f. ex S.Moore (September-November), nectar collecting by both males and females seems to concentrate on this plant.
Females of the species C. xanthomelaena interacted with the same number of plants as those of C. perforator (n = 19). However, C. xanthomelaena males restricted their visits to smaller number of plants (n = 9). Females restricted pollen collecting to four species of Chamaecrista (L.) Moench (n = 69), followed by Zornia brasiliensis Vogel (n = 6). Both males and females were regularly seen visiting R. echinus, which was the main species visited by females to collect nectar. During pollen collection, females also interacted with R. echinus. Malpighia emarginata and Angelonia cornigera Hook.f. were the two oil sources visited by the females. Table 1 displays the complete list of plants on which males and females of both species were observed or collected.
For both sample types (pollen load on females and larval provisions), 47 pollen types were found, corresponding to at least 16 families and 26 genera of plants (Table 2).
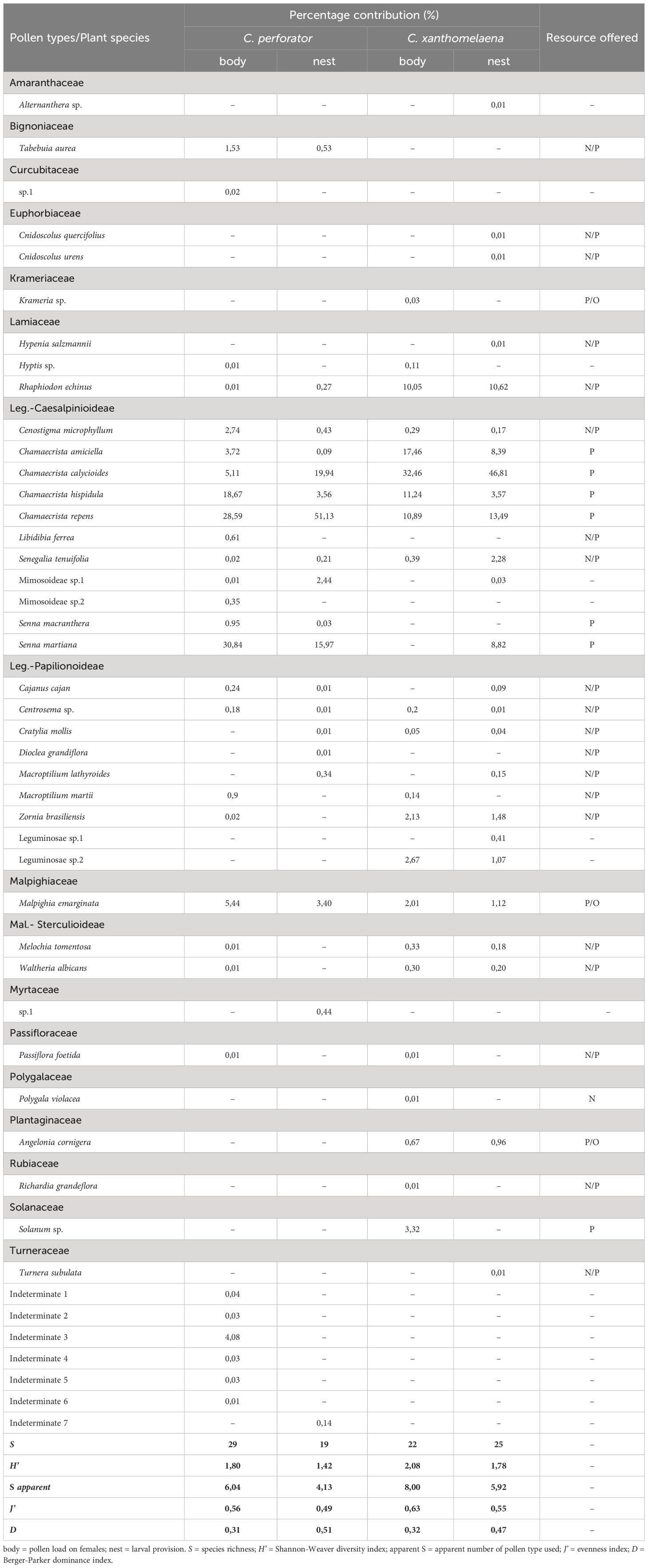
Table 2 Frequency of pollen types/plants recorded in samples from the body of females and nests of Centris perforator and Centris xanthomelaena.
The qualitative analyses of the pollen load from females (n = 30) revealed the presence of 25 pollen types belonging to seven botanical families, six of which could not be identified (Figure 2). The greatest richness of pollen types was found in the family Leguminosae (n = 15 species). The pollen types with the greatest representativeness were Senna martiana, Chamaecrista repens and Chamaecrista hispidula, which together accounted for 78.1% of the pollen types represented in the samples and were the three main sources of pollen visited by the females (Table 2). Other pollen types that had representation greater than 1% were Malpighia emarginata, Chamaecrista calycioides, Chamaecrista amiciella, Cenostigma microphyllum and Tabebuia aurea, which were classified as rare pollens, accounting together for 18.54% of the pollen types found in the samples. Cenostigma microphyllum and T. aurea were the main sources of nectar and M. emarginata was the only source of floral oil exploited by the species in the sample.
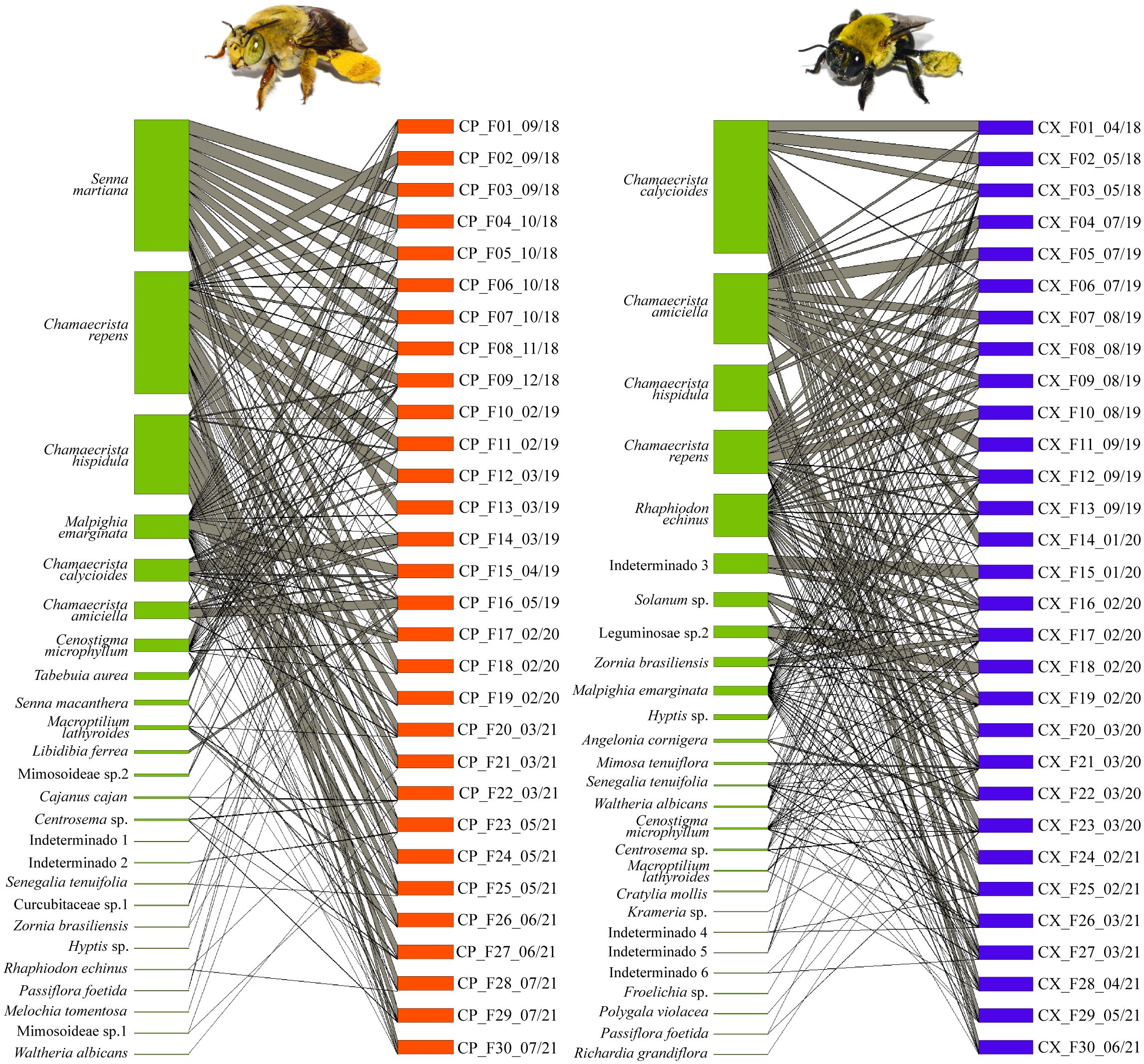
Figure 2 Bipartite graphs representing interaction networks of Centris perforator (left) and Centris xanthomelaena (right) based on samples from pollen load of females. Pollen types/plants represented on left and individuals on right, with interactions represented by grey lines. Width of lines represents number of interactions established from the number of pollen types/plants found in samples. Codes of samples represent species (CP, C. perforator; CX, C. xanthomelaena), type and number of sample (F, females) and collection date (month/year).
Eighteen pollen types were found in the larval provisions (n = 30), belonging to five botanical families, with Leguminosae again the family with the greatest richness of pollen types (n = 14 species). Chamaecrista repens was the predominant pollen type in the samples and, together with the pollen types Chamaecrista calycioides and Senna martiana, represented the main sources of pollen, accounting for 88.05% of the pollen types found in the samples (Figure 3, Table 2). Chamaecrista hispidula and Malpighia emarginata were rare pollen types and the latter was the probable source of floral oil.
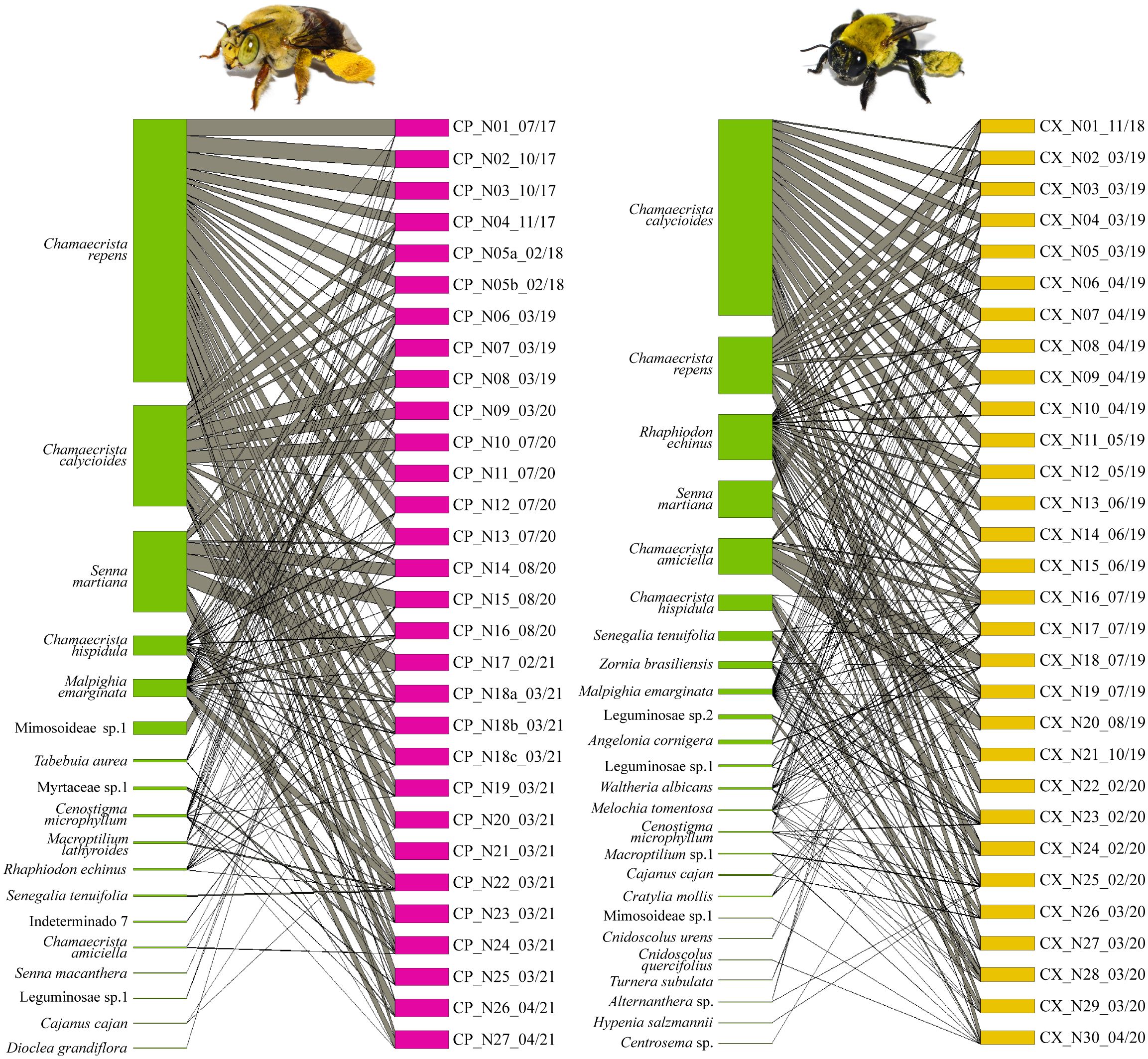
Figure 3 Bipartite graphs representing interaction networks of Centris perforator (left) and Centris xanthomelaena (right) based on samples from larval provisions. Pollen types/plants represented on left and individuals on right, with interactions represented by grey lines. Width of lines represents number of interactions established from the number of pollen types/plants found in samples. Codes of samples represent species (CP, C. perforator; CX, C. xanthomelaena), type and number of sample (N, nests) and collection date (month/year).
Twenty-seven pollen types from 10 botanical families were found in the pollen loads on the body of the females (n = 30). The main sources of pollen corresponded to four pollen types from the genus Chamaecrista (Figure 2): Chamaecrista calycioides, Chamaecrista amiciella, Chamaecrista hispidula and Chamaecrista repens. Rhaphiodon echinus was an infrequent nectar source, but was present in all samples analyzed, corresponding to 10.07% of the diet. Melochia tomentosa and Waltheria albicans were alternative sources of nectar, which, despite being rare pollen types, were frequent in the samples. Three sources of oil were recorded in the samples: Malpighia emarginata, Angelonia cornigera and Krameria sp. Among the other pollen types found in the samples, only Solanum sp. (3.32%), Leguminosae sp.2 and Zornia brasiliensis had contributions higher than 1% (Table 2).
Twenty-five pollen types belonging to eight botanical families were found in the larval provisions (n = 30). Once again, the genus Chamaecrista was predominant in the sample (Figure 3), accounting for 72.28%: Chamaecrista calycioides, Chamaecrista repens, Chamaecrista amiciella and Chamaecrista hispidula. Senna martiana was a rare source of pollen, accounting for 8.82% of the sample. Rhaphiodon echinus was the most representative source of nectar, corresponding to 10.63% of the pollen grains found. Pollen from two floral oil-producing species was identified with rare frequency in the samples: Malpighia emarginata and Angelonia cornigera (Table 2).
Centris xanthomelaena use a greater diversity of pollen types (H’nest = 2.08; H’body = 1.78) compared to C. perforator (H’nest = 1,42; H’body = 1,81). The number of species considered important in the diet of the immature forms (apparent S) varied with the type of sample, ranging from four to eight (Table 2). Three to nine pollen types were found in each subsample for C. perforator ( = 5.4 ± 1.44; n = 60), whereas three to 13 pollen types were found in each subsample for C. xanthomelaena ( = 7 ± 2.35; n = 60).
The two species used plants of the genus Chamaecrista as the main source of pollen (Figure 4), diverging on the species level. While Ch. repens was used by C. perforator as the main source of pollen, C. xanthomelaena collected the large portion of pollen from Ch. calycioides. Moreover, Senna martiana composed a considerable fraction of the larval diet for C. perforator. A difference was also found with regards to the sources of nectar, with Ce. microphyllum as the main source for C. perforator and R. echinus as the most important source for C. xanthomelaena. The Jaccard index (Sj) revealed 36.8% similarity in the pollen samples from the body of all females and 43.3% in the larval provision samples. The proportions of plant species in the diet (PS) were similar, with 43.52% overlap in the samples from the body of the females and 47.89% in the samples from the nests.
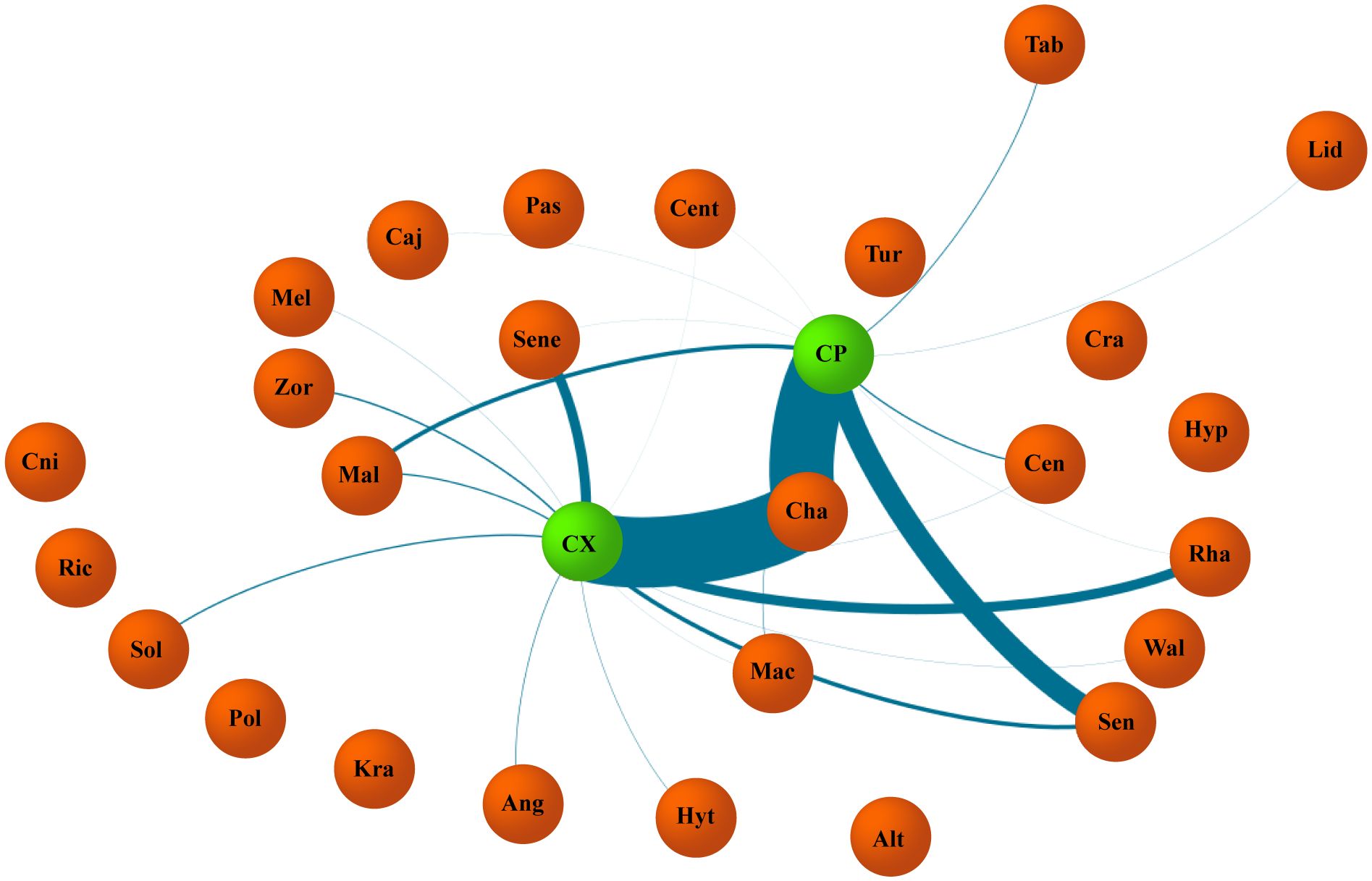
Figure 4 Weighted graph representing interactions established between Centris perforator, Centris xanthomelaena and genera of plants found in both samples of each species. Green vertices represent species of bee (CP, C. perforator; CX, C. xanthomelaena). Orange vertices represent plant genera found in samples. Lines in blue represent interactions established and width is proportional to frequency of pollen grains found in samples, demonstrating strength of the interaction.
The analyses of the pollen load on the females and the nests of Centris perforator and Centris xanthomelaena demonstrated a strong association with plants with poricidal anthers, such as the genus Chamaecrista, which corresponded to the largest part of the pollen types and was present in all samples analyzed. Pollen types from at least four species of Chamaecrista were identified in the samples, together corresponding to more than 45% of the pollen types found on the scopas and in the nests. Certainly, Chamaecrista is the main source of pollen for both species of Centris. The results of the present study lend strength to the hypothesis put forth by Aguiar et al. (2003) that species of Centris endemic to the Caatinga domain, such as Centris hyptidis Ducke and C. xanthomelaena, restrict pollen collecting to fewer sources in comparison to more widely distributed species of the genus. The description of pollen sources found in the nests of Centris analis (Fabricius), Centris tarsata Smith and Centris trigonoides Lepeletier studied in an area of the Caatinga demonstrated a strong association with species of Leguminosae, especially the genera Chamaecrista and Senna (Aguiar et al., 2003; Dórea et al., 2010a, Dórea et al., 2010b; Dórea et al., 2013; Cruz et al., 2015). Vivallo and Zanella (2021) in a collection of plants visited by C. xanthomelaena also pointed to Chamaecrista as the only source of pollen recorded to date. Moreover, Sabino et al. (2019) found that Chamaecrista was a constant or dominant source of pollen for Centris burgdorfi even in different types of vegetation.
The two studied bee species differ regarding to the species Chamaecrista exploited for pollen. Ch. calycioides was the predominant pollen type in the samples of C. xanthomelaena while Ch. repens was the dominant pollen types in the samples of C. perforator. This difference may be related to the size of the flowers, as Centris xanthomelaena, which is smaller, exhibited a preference for smaller Chamaecrista flowers (Figure 5), unlike the dominant species in the samples of C. perforator, thus diminishing competition among the females (see the details of measurements in Supplementary Table 1, Supplementary Figure 1).
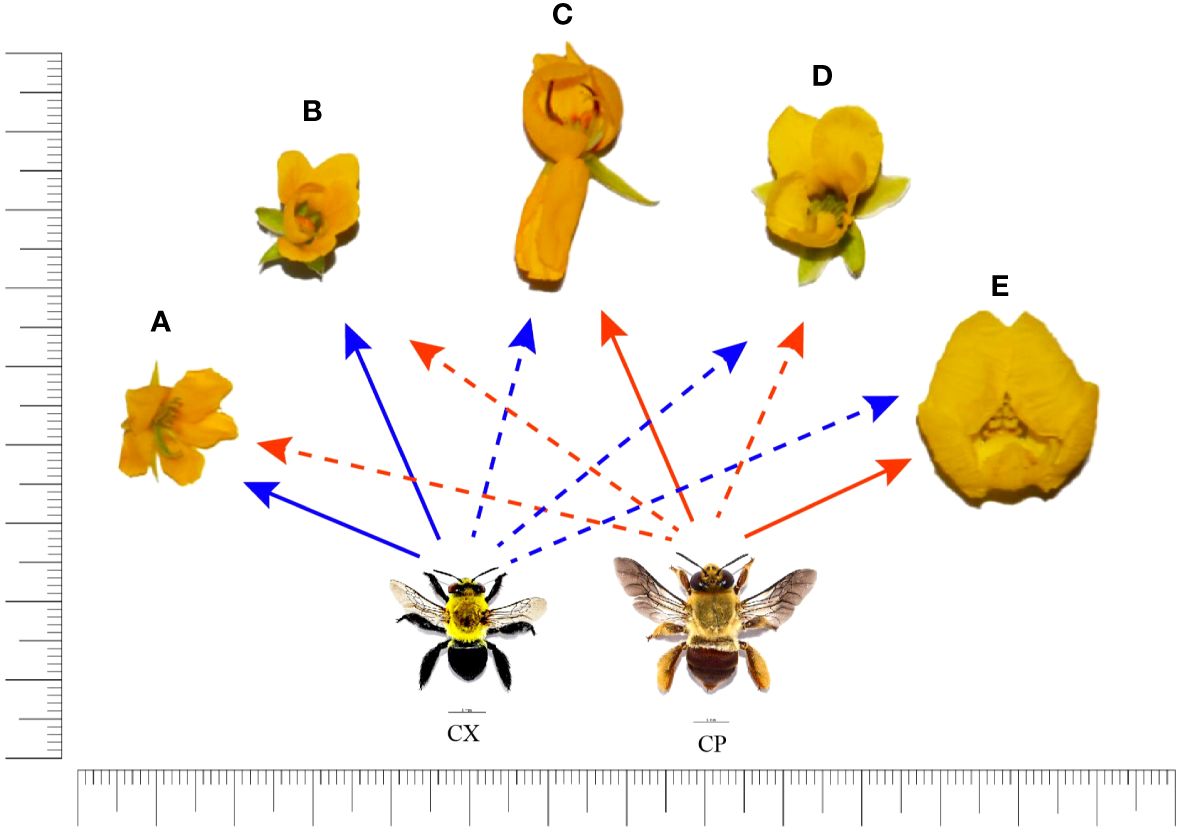
Figure 5 Relative sizes of Chamaecrista and Senna flowers used as a pollen source by Centris xanthomelaena (CX) and Centris perforator (CP) in their diets. The arrows indicate partial (dashed line) or priority (solid line) use of pollen. (A) Chamaecrista calycioides; (B) Chamaecrista amiciella; (C) Chamaecrista repens; (D) Chamaecrista hispidula; (E) Senna martiana. Scale = 5 mm.
In the Caatinga domain, Leguminosae is the best represented botanical family and is an important resource for various species of bees (Queiroz, 2002). The high availability and proximity of plants in this family to nesting sites optimizes the provisioning phase and, consequently, diminishes the energy expenditure of females in search of resources. Despite being an abundant resource, pollen collection from various species of Leguminoseae, especially those of the subtribe Cassiinea, is limited to bees with the capacity to perform collection by vibration, which is known as “buzz pollination”, such as those of the genera Bombus, Centris, Epicharis, Melipona and Xylocopa (Nunes-Silva et al., 2010). With buzz pollination, pollen is released from the poricidal anthers through the vibration executed by the thoracic muscles of the bees (Buchmann, 1983). The high protein content of the pollen produced by plants with poricidal anthers may result in the preference on the part of females due to the quality of the resource (Roulston et al., 2000; Vaudo et al., 2020).
Richness in the pollen samples ranged from 18 to 27 species. The diversity indices in the samples of C. perforator varied mainly due to the presence of contaminant species, with percentages lower than 1% (12 species) on the body of females. We believe that various of these species were visited for nectar consumption (even the undetermined ones) or only represent contamination during foraging of the females. The number of species and Shannon’s diversity index (H’) were lower in the nests of C. perforator (S = 18; H’=1.42) and the brood cells are evidently supplied with pollen from Leguminosae – Caesalpineaceae (> 75% Chamaecrista + 16% Senna), thus explaining a higher dominance index in comparison to pollen from the scopa. The richness of pollen types used by C. xanthomelaena was slightly higher in the samples from the scopa than those from the nests. In contrast, dominance was greater in the samples from the brood cells of the two species.
The evenness indices (J’) were similar in all samples (difference of less than 10%) and slightly higher in the sample from the nests of the two species of Centris. The Jaccard indices (Sj) indicated 36.8% similarity in the samples of pollen from the body of the females of the two species and 43.3% for the samples from nests. The proportions of plants species in the diet (PS) varied, with 43.52% overlap in the samples from the scopas of the females and 47.89% in the samples from the nests of the two species.
Despite that close similarity in the sources of pollen exploited by the two species of Centris, a difference was found in the plants visited to obtain nectar. Rhaphiodon echinus was the main source of nectar recorded in all samples of C. xanthomelaena, with M. tomentosa and W. albicans as secondary sources. Apparently, R. echinus is an annual source of nectar composing the diet of females and immature forms, even in periods of resource scarcity, and accounted for approximately 10% of the pollen types in the samples. Martins et al. (2020b) found that nectar is one of the constituents of the larval provisions of C. xanthomelaena and added together with the oil soon after pollen collection trips. Rhaphiodon echinus was also reported as a source of nectar for other species of Centris in the Caatinga domain, such as C. analis (Dórea et al., 2010a), C. tarsata (Dórea et al., 2010b) and C. trigonoides (Dórea et al., 2013). Besides being endemic to the Caatinga, R. echinus is one of the main plants of the herbaceous stratum in the study area (Harley et al., 2015). Floral visitation data confirm that this herbaceous plant is a key species for the permanence of C. xanthomelaena in the area.
The sources of nectar for C. perforator were more diverse. The pollen from Cenostigma microphylum appears with greater frequency in the samples, but the main source of nectar alternated with the period of the year. In samples from September to November, Tabebuia aurea was the main source of nectar visited by the females. This species has mass flowering in the dry season (Barros, 2001), when resource scarcity naturally occurs.
In the same way as the females, the males of both species diverged in their trophic niche with regards to the main sources of nectar exploited. Males of C. xanthomelaena visited flowers of R. echinus more intensely, whereas the two main sources of nectar visited by males of C. perforator were Ce. microphylum and T. aurea. Moreover, males of this species demonstrated territorial behavior at these plants, underscoring their importance for the trophic niche. To maximize reproductive success, patrolling for virgin females on flowers is one of the tactics employed by males of different species (Paxton, 2005), which would explain the preference of males of C. xanthomelaena and C. perforator for the main sources of nectar used by the females.
Bees of the genus Centris have a strong association with plants of the family Malpighiaceae (Anderson, 1990) and this interaction is reflected in the source of floral oil exploited by C. perforator and C. xanthomelaena. The analysis of the samples revealed that M. emarginata is the main source of oil used by both species. Despite appearing to be the exclusive source of oil for C. perforator, the C. xanthomelaena samples indicate that Agelonia cornigera and Krameria sp. are alternative sources of oil used by the females. Besides floral oil, some studies indicate that Malphigiaceae also furnishes pollen as a reward, such as the genus Epicharis, in which pollen loads can contain more than 90% of the pollen from Byrsonima (Gaglianone, 2005). However, only two samples of C. perforator had M. emarginata as the dominant pollen type.
The presence of pollen from Angelonia cornigera in the samples occurred simultaneously to the rainy period (February to May), in which peak flowering of this plant also occurred. The flowers were found in sites with sandy soil very close to the nesting sites. In the samples containing pollen from A. cornigera, the frequency of grains of pollen from M. emarginata was very low or absent.
Although most species of Centris are adapted to collect oil from epithelial elaiophores (Vogel, 1974; Neff and Simpson, 1981), some species have morphological modifications that enable collecting oil from flowers with trichomal elaiophores (Vogel, 1974; Simpson et al., 1990; Cocucci, 1991; MaChado et al., 2002). This modified “four-legged” pattern (Neff and Simpson, 1981) is found in C. xanthomelaena, C. burgdorfi, C. tricolor and all species of the subgenus Hemisiella (Taniguchi, 2010), enabling these species to expand their sources of oil (Neff and Simpson, 1981). Sabino et al. (2019) studied three populations of C. burgdorfi in different types of vegetation and found plasticity with regards to the sources of oil exploited in flowers with epithelial and trichomal elaiophores of the genera Krameria and Angelonia, respectively. Although pollen from Krameria was found in a low frequency in the samples of C. xanthomelaena, no plant belonging to this genus was found within a 500-meter radius of the nesting sites of the species. There are reports of C. xanthomelaena serving as an effective pollinator of Krameria grandiflora, which also received sporadic visits from C. perforator (Carneiro et al., 2019).
The females of Centris perforator and Centris xanthomelaena demonstrated high dependence on the vegetation mainly in the herbaceous and shrub strata. These strata are fundamental to the survival and permanence of pollinating agents in agricultural areas (Wolowski et al., 2019), but are constantly eliminated within these areas. Acerola (West Indian cherry) requires pollinators for cross fertilization (Freitas et al., 1999; Sazan et al., 2014), which makes good management practices all the more necessary, ensuring not only productivity, but also the quality of the fruit (Magalhães and Freitas, 2013). As the visits of females of C. perforator and C. xanthomelaena are directed to the collection of the floral oil in acerola, the possibility of the management of females in orchards depends on the presence of the surrounding herbaceous and shrub strata, ensuring the availability of sources of pollen and nectar essential to the survival and permanence of the populations of the species in the area.
The availability of floral resources is one of the main factors responsible for the regulation and abundance of native bee populations (Roulston and Goodell, 2011), especially those resources that are indispensable to species survival, such as the case of floral oil for Centris (Silva et al., 2017). Silva et al. (2019) call attention to the change in the regularity of floral resources in response to climate change events, such as long periods of drought, especially in dry forests like the Caatinga. Consequently, the reduction in these floral resources for bees can cause the decline in these pollinators, thus affecting the biome as a whole (Kearns et al., 1998; Kevan and Imperatriz-Fonseca, 2002). The herbaceous stratum constitutes one of the main groups of plants that can suffer these effects more intensely, especially plants endemic to the Caatinga (Silva et al., 2019). In this scenario, populations of C. perforator and C. xanthomelaena could be affected, as both species are highly dependent on plants of this stratum, such as the endemic species Cenostigma microphyllum and Raphiodun echinus.
Despite visiting numerous species of plants, few of these species contributed to the major fraction of the diet of immature forms. The genus Chamaecrista is a key group for the maintenance of populations of Centris perforator and Centris xanthomelaena in the area of the Caatinga domain. However, differences in the trophic niches are found on the species level, thus avoiding interspecific competition. Most females of both species interacted with acerola (West Indian cherry) (Malpighia emarginata) only for the collection of floral oil, which underscores the importance of maintaining plants that provide nectar and pollen in areas surround the orchards, especially species with poricidal anthers, like Chamaecrista. Lastly, areas of native forest are fundamental to the maintenance of bee populations in zones of agricultural activity, ensuring the regularity of important resources for the feeding of adults and the diet of immature forms.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The manuscript presents research on animals that do not require ethical approval for their study.
HM: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. MD: Formal analysis, Methodology, Writing – original draft, Writing – review & editing. VF: Conceptualization, Writing – original draft, Writing – review & editing. CS: Methodology, Writing – original draft, Writing – review & editing. IA-D-S: Conceptualization, Formal analysis, Investigation, Methodology, Resources, Supervision, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. We thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for granting the scholarship 134121/2019-8 to the first author.
The authors would like to thank Dr. Felipe Vivallo from the National Museum of Rio de Janeiro to identify the species of bees and Vale do São Francisco Herbarium (Hvasf) for identifying the species of plants. We also thank the USP Graduate Program in Entomology and LEAV-USP for their support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frbee.2024.1383589/full#supplementary-material
Aguiar C. M., Garófalo C. A., Almeida G. F. (2005). Trap-nesting bees (Hymenoptera, Apoidea) in areas of dry semideciduous forest and caatinga, Bahia, Brazil. Rev. Bras. Zool. 22, 1030–1038. doi: 10.1590/S0101-81752005000400031
Aguiar C. M. L., Zanella F. C. V., Martins C. F., Carvalho C. A. L. D. (2003). Plantas visitadas por Centris spp. (Hymenoptera: Apidae) na Caatinga para obtenção de recursos florais. Neotrop. Entomol. 32, 247–259. doi: 10.1590/S1519566X2003000200009
Aizen M. A., Feinsinger P. (2003). “Bees not to be? Responses of insect pollinator faunas and flower pollination to habitat fragmentation,” in How landscapes change (Springer, Berlin, Heidelberg), 111–129. doi: 10.1007/978-3-662-05238-9_7
Alvares C. A., Stape J. L., Sentelhas P. C., Gonçalves J. L. M., Sparovek G. (2013). Köppen’s climate classification map for Brazil. Meteorol. Zeitschrift. 22, 711–728. doi: 10.1127/0941-2948/2013/0507
Alves-dos-Santos I., MaChado I. C., Gaglianone M. C. (2007). História natural das abelhas coletoras de óleo. Oecologia Brasiliensis 11, 544–557. doi: 10.4257/oeco.2007.1104.06
Anderson W. R. (1990). The origin of the Malpighiaceae - the evidence from morphology. Mem. New York Bot. Gard. 64, 210–224.
Barros M. G. (2001). Pollination ecology of Tabebuia aurea (Manso) Benth. and Hook. and T. ochracea (Cham.) Standl. (Bignoniaceae) in Central Brazil cerrado vegetation. Rev. Bras. Bot. 24, 255–261. doi: 10.1590/S0100-84042001000300003
Buchmann S. L. (1983). “Buzz pollination in angiosperms,” in Handbook of experimental pollination biology. Eds. Jones C. E., Little R. J. (Van Nostrand-Rheinhold, New York), 73–113.
Buchmann S. L. (1987). The ecology of oil flowers and their bees. Annu. Rev. Ecol. Evol. Syst. 18, 343–369. doi: 10.1146/annurev.es.18.110187.002015
Bustamante P. M. A. C. (2009). A fruticultura no Brasil e no Vale do São Francisco: vantagens e desafios. Rev. Econ. Nordeste. 40, 153–171. doi: 10.61673/ren.2009.344
Camillo E., Garofalo C. A., Serrano J. C., Muccillo G. (1995). Diversidade e abundância sazonal de abelhas e vespas solitarias em ninhos armadilhas (Hymenoptera, Apocrita, Aculeata). Rev. Bras. Entomol. 39, 459–470.
Cane J. H., Eickwort G. C., Wesley F. R., Spielholz J. (1983). Foraging, grooming and mate-seeking behaviors of Macropis nuda (Hymenoptera, Melittidae) and use of Lysimachia ciliata (Primulaceae) oils in larval provisions and cell linings. Am. Midl. Nat. 110, 257–264. doi: 10.2307/2425267
Carneiro L. T., André C. B. D. S., Takahasi A., Alves-dos-Santos I. (2019). Interactions between oil-collecting bees and Krameria grandiflora (Krameriaceae) with emphasis on the role of specialized floral traits in the mutual fit. Arthropod Plant Interact. 13, 213–226. doi: 10.1007/s11829-019-09689-w
Cocucci A. A. (1991). Pollination biology of nierembergia (Solanaceae). Plant Syst. Evol. 174, 17–35. doi: 10.1007/BF00937691
Coville R. E., Frankie G. W., Vinson S. B. (1983). Nests of Centris segregata (Hymenoptera: Anthophoridae) with a review of the nesting habits of the genus. J. Kans. Entomol. Soc. 56, 109–122.
Cruz A. P. A. D., Dórea M. D. C., Lima L. C. L. (2015). Pollen types used by Centris (Hemisiella) tarsata Smith, (1874) (Hymenoptera, Apidae) in the provisioning of brood cells in an area of Caatinga. Acta Bot. Bras. 29, 282–284. doi: 10.1590/0102-33062015abb0005
De Nooy W., Mrvar A., Batagelj V. (2018). Exploratory social network analysis with Pajek: Revised and expanded edition for updated software Vol. 46 (Cambridge university press).
Dórea M. C., Aguiar C. M. L., Figueroa L. E. R., Lima L. C. L., Santos F. A. R. (2010a). Residual pollen in nests of Centris analis (Hymenoptera, Apidae, Centridini) in an area of caatinga vegetation from Brazil. Oecologia Aust. 14, 232–237. doi: 10.4257/oeco.2010.1401.13
Dórea M. C., Aguiar C. M. L., Figueroa L. E. R., Lima L. C. L., Santos F. A. R. (2010b). Pollen residues in nests of Centris tarsata Smith (Hymenoptera, Apidae, Centridini) in a tropical semiarid area in NE Brazil. Apidologie 41, 557–567. doi: 10.1051/apido/2010005
Dórea M. D. C., Aguiar C. M. L., Figueroa L. E. R., Lima L. C. L., Santos F. D. A. R. (2013). A study of pollen residues in nests of Centris trigonoides Lepeletier (Hymenoptera, Apidae, Centridini) in the Caatinga vegetation, Bahia, Brazil. Grana 52, 122–128. doi: 10.1080/00173134.2012.745595
Dórea M. C., Santos F. D. A. R., Aguiar C. M. L., Martins C. F. (2017). Bee life in the city: an analysis of the pollen provisions of Centris (Centris) flavifrons (Centridini) in an urban area. Sociobiology 64, 166–173. doi: 10.13102/sociobiology.v64i2.1277
Frankie G. W., Haber W. A., Opler P. A., Bawa K. S. (1983). “Characteristics and organization of the large bee pollination system in the Costa Rican dry forest,” in Handbook of experimental pollination biology. Eds. Jones C. E., Little R. J. (Van Nostrand Reinhold, New York, New York, USA), 411–447.
Freitas B. M., Alves J. E., Brandão G. F., Araújo Z. B. (1999). Pollination requirements of West Indian cherry (Malpighia emarginata) and its putative pollinators, Centris bees, in NE Brazil. J. Agric. Sci. 133, 303–311. doi: 10.1017/S0021859699006930
Gaglianone M. C. (2005). Nesting biology, seasonality, and flower hosts of Epicharis nigrita (Friese 1900)(Hymenoptera: Apidae: Centridini), with a comparative analysis for the genus. Stud. Neotrop. Fauna Environ. 40, 191–200. doi: 10.1080/01650520500250145
Harley R., França F., Santos E. P., Santos J. S., Pastore J. F. (2015). Lamiaceae in Lista de Espécies da Flora do Brasil (Rio de Janeiro: jardim botânico do Rio de Janeiro). Available at: http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB8294.
Jones G. D., Bryant V. M. (1996). Melissopalynology. Chapter 23D. In: Jansonius J., McGregor D. C., editors. Palynology: principles and applications. Dallas (TX): American Association of Stratigraphic Palynologists Foundation. 3, 933–938.
Kearns C. A., Inouye D. W., Waser N. M. (1998). Endangered mutualisms: the conservation of plant-pollinator interactions. Annu. Rev. Ecol. Evol. Syst. 29, 83–112. doi: 10.1146/annurev.ecolsys.29.1.83
Kevan P., Imperatriz-Fonseca V. L. (2002). Pollinating bees: the conservation link between Agriculture and Nature (Brasília: Ministério do Desenvolvimento).
Klein A. M., Vaissière B., Cane J. H., Steffan-Dewenter I., Cunningham S. A., Kremen C., et al. (2007). Importance of pollinators in changing landscapes for world crops. Proc. R. Soc B: Biol. Sci. 274, 303–313. doi: 10.1098/rspb.2006.3721
MaChado I. C., Vogel S., Lopes A. V. (2002). Pollination of Angelonia cornigera Hook. (Scrophulariaceae) by long-legged, oil-collecting bees in NE Brazil. Plant Biol. 4, 352–359. doi: 10.1055/s-2002-32325
Magalhães C. B., Freitas B. M. (2013). Introducing nests of the oil-collecting bee Centris analis (Hymenoptera: Apidae: Centridini) for pollination of acerola (Malpighia emarginata) increases yield. Apidologie 44, 234–239. doi: 10.1007/s13592-012-0175-4
Martins H. O. D. J., Alves-dos-Santos I., Vivallo F., Ferreira V. S. (2020a). Biological notes on Centris (TraChina) perforator with a comparative analysis of the species with ground-nesting habits of the genus (Apidae: Centridini). J. Apic. Res. 60, 123–134. doi: 10.1080/00218839.2020.1828238
Martins H. O. J., Oliveira-Rebouças P. L., Aguiar C. M. L., Silva C. I., Ferreira V. S. (2020b). Nesting biology of Centris (Paracentris) xanthomelaena (Hymenoptera: Apidae: Centridini). J. Apic. Res. 62, 222–232. doi: 10.1080/00218839.2020.1828234
Montero I., Tormo R. (1990). Discriminación del origen geográfico de mieles de Extremadura por métodos estadísticos. Actas VIII Simp (Puerto de Santa Cruz (Tenerife, España): Palinol), 397–403.
Moure J. S., Melo G. A. R., Vivallo F. (2007). “Centridini Cockerell & Cockerel,” in Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical region. Eds. Moure J. S., Urban D., Melo G. A. R. (Curitiba: Sociedade Brasileira de Entomologia), 83–142.
Neff J. L., Simpson B. B. (1981). Oil-Collecting structures in the anthophoridae (Hymenoptera): morphology, function, and use in systematics. J. Kans. Entomol. Soc 54, 95–123.
Neff J. L., Simpson B. B. (2017). Vogel’s great legacy: The oil flower and oil-collecting bee syndrome. Flora 232, 104–116. doi: 10.1016/j.flora.2017.01.003
Nunes-Silva P., Hrncir M., Fonseca V. L. I. (2010). A polinização por vibração. Oecologia Australis 14, 140–151. doi: 10.4257/oeco.2010.1401.07
Ollerton J., Winfree R., Tarrant S. (2011). How many flowering plants are pollinated by animals? Oikos 120, 321–326. doi: 10.1111/j.1600-0706.2010.18644.x
Paxton R. J. (2005). Male mating behaviour and mating systems of bees: an overview. Apidologie 36, 145–156. doi: 10.1051/apido:2005007
Pielou E. C. (1977). The latitudinal spans of seaweed species and their patterns of overlap. J. Biogeogr. 4, 299–311. doi: 10.2307/3038189
Queiroz L. D. (2002). “Distribuição das espécies de Leguminosae na Caatinga,” in Vegetação e flora da caatinga (APNE/CNIP, Recife), 141–153.
R Core Team. (2017). R: A language and environment for statistical computing (Version 3.4.0). R Foundation Stat. Computing. http://www.R-project.org.
Rech A. R., Agostini K., Oliveira P. E., MaChado I. C. (2014). Biologia da polinização (Rio de Janeiro: Projecto Cultural), 524.
Rêgo M. M. C., Albuquerque P. M. C., Ramos M. C., Carreira L. M. (2006). Aspectos da biologia de nidificação de Centris flavifrons (Friese) (Hymenoptera: Apidae, Centridini), um dos principais polinizadores do murici (Byrsonima crassifolia L. Kunth, Malpighiaceae), no Maranhão. Neotrop. Entomol. 35, 579–587. doi: 10.1590/S1519-566X2006000500003
Renner S. S. (2006). “Rewardless flowers in the angiosperms and the role of insect cognition in their evolution,” in Plant-pollinator interactions: from specialization to generalization (University of Chicago Press, Chicago), 123–144.
Ribeiro E. K. M. D., Rêgo M. M. C., MaChado I. C. S. (2008). Cargas polínicas de abelhas polinizadoras de Byrsonima chrysophylla Kunth. (Malpighiaceae): fidelidade e fontes alternativas de recursos florais. Acta Bot. Brasilica 22, 165–171. doi: 10.1590/S0102-33062008000100017
Rocha-Filho L. C., Ferreira-Caliman M. J., Garófalo C. A., Augusto S. C. (2018). “A specialist in an urban area: are cities suitable to harbour populations of the oligolectic bee Centris (Melacentris) collaris (Apidae: Centridini)?,” in Annales Zoologici Fennici, vol. 55. (Finnish Zoological and Botanical Publishing Board), 135–150.
Rocha-Filho L. C. D., Ferreira-Caliman M. J., Serrano J. C., Camargo J. M. F., Garófalo C. A. (2017). Nesting ecology of the oil-collecting bee Centris (Melacentris) conspersa Mocsáry and its potential association with the cleptoparasite Cyphomelissa diabolica Friese (Apidae: Centridini, Ericrocidini). J. Apic. Res. 56, 489–496. doi: 10.1080/00218839.2017.1351906
Roulston T. H., Cane J. H., Buchmann S. L. (2000). What governs protein content of pollen: pollinator preferences, pollen–pistil interactions, or phylogeny? Ecol. Monogr. 70, 617–643. doi: 10.2307/2657188
Roulston T. A. H., Goodell K. (2011). The role of resources and risks in regulating wild bee populations. Annu. Rev. Entomol. 56, 293–312. doi: 10.1146/annurev-ento-120709-144802
Sabino W. O., Alves-dos-Santos I., Silva C. I. (2019). Versatility of the trophic niche of Centris (Paracentris) burgdorfi (Apidae, Centridini). Arthropod Plant Interact. 13, 227–237. doi: 10.1007/s11829-018-9654-5
Sazan M. S., Queiroz E. P., Ferreira-Caliman M. J., Parra-Hinojosa A., Silva C. I., Garófalo C. A. (2014). Manejo dos polinizadores da aceroleira (Ribeirão Preto: Holos). 54p.
Schlindwein C. (2000). “A importância de abelhas especializadas na polinização de plantas nativas e conservação do meio ambiente,” in Anais do IV Encontro sobre Abelhas (Universidade de São Paulo, Ribeirão Preto), 131–141.
Shannon C. E. A. (1948). Mathematical theory of communication. Bell System Tech. J. 27, 379–423. doi: 10.1002/bltj.1948.27.issue-3
Silva C. I. D. (2009). Distribuição espaço-temporal de recursos florais utilizados por espécies de Xylocopa (Hymenoptera, Apidae) e interação com plantas do Cerrado sentido restrito no Triângulo Mineiro. Universidade Federal de Uberlândia, Uberlândia, 294.
Silva J. L. S. E., Cruz-Neto O., Peres C. A., Tabarelli M., Lopes A. V. (2019). Climate change will reduce suitable Caatinga dry forest habitat for endemic plants with disproportionate impacts on specialized reproductive strategies. PLoS One 14, e0217028. doi: 10.1371/journal.pone.0217028
Silva C. I., Ballesteros P. L. O., Palmero M. A., Bauermann S. G., Evaldt A. C. P., Oliveira P. E. (2010). Catálogo polínico: Palinologia aplicada em estudos de conservação de abelhas do gênero Xylocopa no Triângulo Mineiro. Edufu (Brazil).
Silva C. I., Hirotsu C. M., Filho A. J. S. P., Queiroz E. P., Garófalo C. A. (2017). Is the maximum reproductive rate of Centris analis (Hymenoptera, Apidae, Centridini) associated with floral resource availability? Arthropod Plant Interac. 11, 389–402. doi: 10.1007/s11829-017-9513-9
Silva C. I., Radaeski J. N., Arena M. V. N., Bauermann S. G. (2020). Atlas of pollen and plants used by bees (Rio Claro: Consultoria Inteligente en servicios ecosistemicos).
Silveira F. A., Melo G. A. R., Almeida E. A. B. (2002). Abelhas brasileiras: sistemática e identificação (Brasil: Belo Horizonte). 253p.
Simpson B. B., Neff J. L., Dieringer G. (1990). The production of floral oils by Monttea (scrophulariaceae) and the function of tarsal pads in Centris bees. Plant Syst. Evol. 173, 209–222. doi: 10.1007/BF00940864
Simpson B. B., Neff J. L., Seigler D. (1977). Krameria, free fatty acids and oil collecting bees. Nature 267, 150–151. doi: 10.1038/267150a0
Taniguchi M. (2010). Morfologia funcional das estruturas envolvidas na coleta e transporte de óleo floral por fêmeas do gênero Centris Fabricius 1804 (Hymenoptera, Apidae: Centridini). Universidade de São Paulo, Ribeirão Preto.
Vaudo A. D., Tooker J. F., Patch H. M., Biddinger D. J., Coccia M., Crone M. K., et al. (2020). Pollen protein: Lipid macronutrient ratios may guide broad patterns of bee species floral preferences. Insects 11, 132. doi: 10.3390/insects11020132
Vilhena A. M. G. F., Augusto S. C. (2007). Polinizadores da aceroleira Malpighia emarginata DC (Malpighiaceae) em área de cerrado no Triângulo Mineiro. Biosci. J. 23, 14–23.
Vilhena A. M. G. F., Rabelo L. S., Bastos E. M., Augusto S. C. (2012). Acerola pollinators in the savanna of Central Brazil: temporal variations in oilcollecting bee richness and a mutualistic network. Apidologie 43, 51–62. doi: 10.1007/s13592-011-0081-1
Villanueva-Gutiérrez R., Roubik D. W. (2004). Why are African honey bees and not European bees invasive? Pollen diet diversity in community experiments. Apidologie 35, 481–491. doi: 10.1051/apido:2004041
Vivallo F. (2020). Lectotype designations taxonomic notes and new synonymies in some species of the bee genus Centris Fabricius 1804 described by Frederick Smith (Hymenoptera: Apidae). Zootaxa 4729, 151–176. doi: 10.11646/zootaxa.4729.2
Vivallo F. (2022a). The oil-collecting bees of Centris (Melanocentris) (Hymenoptera: Apidae) (Rio de Janeiro). 514 pp.
Vivallo F. (2022b). The oil-collecting bees of Centris (Aphemisia)/ (Schisthemisia) (Hymenoptera: Apidae) (Rio de Janeiro). 281 pp.
Vivallo F., Vásquez-Lenis E., Mérida-Rivas J. (2023). New species of centris (Xanthemisia) moure from the mesoamerican dominion and the mexican transition zone (Hymenoptera: apidae). Zootaxa 5351, 341–355. doi: 10.11646/zootaxa.5351.3
Vivallo F., Zanella F. C. V. (2021). Relicthemisia, a new subgenus of the oil-collecting bee genus Centris Fabricius 1804 with notes on distribution and host plants of C. xanthomelaena Moure & Castro 2001 (Hymenoptera: Apidae). Eur. J. Taxon 760, 1–15. doi: 10.5852/ejt.2021.760.1435
Vogel S., MaChado I. C. (1991). Pollination of four sympatric species of Angelonia (Scrophulariaceae) by oil-collecting bees in NE. Brazil. Plant Syst. Evol. 178, 153–178. doi: 10.1007/BF00937962
Wolowski M., Agostini K., Rech A. R., Varassin I. G., Maués M., Freitas L., et al. (2019). Relatório temático sobre polinização, polinizadores e produção de alimentos no Brasil. 1st edn (São Carlos: Editora Cubo). doi: 10.4322/978-85-60064-83-0
Xiao Y., Li X., Cao Y., Dong M. (2016). The diverse effects of habitat fragmentation on plant–pollinator interactions. Plant Ecol. 217, 857–868. doi: 10.1007/s11258-016-0608-7
Keywords: solitary bee, Centris, diet range, Caatinga, plant-pollinator interactions
Citation: Martins HOJ, Dórea MC, Ferreira VS, Silva CI and Alves-dos-Santos I (2024) Range and trophic niche overlap of two sympatric species of floral oil collecting bees in a fragment of dry forest. Front. Bee Sci. 2:1383589. doi: 10.3389/frbee.2024.1383589
Received: 07 February 2024; Accepted: 11 March 2024;
Published: 03 April 2024.
Edited by:
Susan E. Fahrbach, Wake Forest University, United StatesReviewed by:
Ujjwal Layek, Rampurhat College, IndiaCopyright © 2024 Martins, Dórea, Ferreira, Silva and Alves-dos-Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Herbeson O. J. Martins, aGVyYmVzb24uYmlvQGhvdG1haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.