 Camila Maia-Silva1,2*
Camila Maia-Silva1,2* Michael Hrncir3
Michael Hrncir3 Tereza Cristina Giannini2
Tereza Cristina Giannini2 Manuel Toledo-Hernández4
Manuel Toledo-Hernández4 Vera L. Imperatriz-Fonseca2,3
Vera L. Imperatriz-Fonseca2,3- 1Faculdade de Economia, Administração, Contabilidade e Atuária, Universidade de São Paulo, São Paulo, Brazil
- 2Instituto Tecnológico Vale, Belém, Brazil
- 3Instituto de Biociências, Universidade de São Paulo, São Paulo, Brazil
- 4Sustainable Agricultural Systems and Engineering Laboratory, School of Engineering, Westlake University, Hangzhou, Zhejiang, China
Cocoa (Theobroma cacao) is a multi-billion-dollar business. This tropical crop strongly depends on animal pollination for fruit development and seed production. The lack or inefficiency of natural pollinators in cocoa plantations has driven farmers to search for alternatives, such as laborious pollination by hand. A so far untested alternative, which has received increasing attention during the past couple of years, is targeted crop pollination through managed social bees. However, owing to the smallness of the flowers of T. cacao as well as structural barriers that impede large insects to access the stigma, only tiny bees may be a viable option for targeted cocoa pollination. In the present study, we asked whether small stingless bees (Apidae, Meliponini) could come into consideration as managed cocoa crop-pollinators, especially in shadowy agroforests. Among the 188 meliponine species native to the Brazilian Amazon region, which comprises an important portion of the center of origin of T. cacao, we selected 52 species based on morphological criteria (intertegular distance ≤ 1.4 mm; body length: 2.2–6.0 mm). Important for cocoa production, some of these Meliponini have an ample geographic distribution, occurring both in the center of origin of T. cacao and beyond (centers of cocoa production in Brazil: Pará: 35 spp., Bahia: 10 spp.). Presumably all species may be active at illuminance levels below those found in heavily shaded cocoa plantations, at least at times when anthers show full dehiscence and during maximum receptivity of the stigma. The bees’ potential to forage under reduced light regimes is corroborated by the finding that between 20 and 60% of the naturally exploited food sources are understory vegetation, including herbs, subshrubs, shrubs, and lianas. Many of the selected Meliponini build their nests, at least facultatively, in tree cavities, which facilitates their transfer to rational hives and, hence, the use of managed colonies in directed crop pollination. Important next steps for validating the potential of these small stingless bees in targeted cocoa pollination should comprise detailed studies on their foraging behavior and olfactory learning capacities.
1 Introduction
The cocoa tree (Theobroma cacao, Malvaceae) is native to the Amazon region (Cornejo et al., 2018; Zarrillo et al., 2018), and is a key global export commodity, providing a source of income for millions of smallholder farmers in global tropical biodiversity hotspots (Klein et al., 2008; Beg et al., 2017; Tscharntke et al., 2023). Despite the existence of self-compatible wild genotypes (Branco et al., 2018; Morán et al., 2021), the hermaphroditic flowers of many cocoa varieties, particularly of those of the upper Amazon region, are self-incompatible (Glendinning, 1972). Thus, fruit set highly depends on cross-pollination (Branco et al., 2018). Cocoa trees produce small, whitish flowers directly from the trunk and branches (Young et al., 1987) that produce only vestigial amounts of nectar in epidermal trichomes (Young et al., 1984) and, thus, offer pollen as main attractant for flower visitors. The anthers (masculine parts) are concealed within shell-shaped petal hoods, and style, stigma, and ovary (feminine parts) are fenced by five staminodes (Wolcott et al., 2023). Both flower size (10-15 mm in diameter) and structural barriers (petal shell, staminode fence) impede access to the female reproductive parts of the flower by large insects (Frimpong-Anin et al., 2014; Wolcott et al., 2023). Thus, being small is an imperative characteristic of a successful cocoa pollinator. However, flower visitors may likewise be too minute to interact effectively with the reproductive structures of a flower. Species with a body size of less than 2 mm usually fail to touch the style while crawling along the inner surface of the staminodes, thus rendering pollen deposition improbable (Kaufmann, 1975a; Frimpong-Anin et al., 2014).
Despite the global economic importance of cocoa, we know only little regarding the identity of the main cocoa pollinators and their suitable habitats so far. In cocoa-growing areas of the Americas, West Africa, and Southeast Asia, small dipterans (hereafter “midges”) from the families Ceratopogonidae and Cecidomyiidae account for nearly 50% of cocoa flower visits (Toledo-Hernández et al., 2017). Several of these species have been considered pollinators of T. cacao (Bigger, 2012) despite the lack of strong experimental evidence (e.g.: Winder, 1977; Young, 1985a). In any case, there are two major caveats to efficient cocoa pollination by midges. First, these small dipterans have a limited flight capacity, with foraging ranges of usually less than 10 m (Chumacero de Schawe et al., 2018). Hence, their movement within plantations is rather restricted (Klein et al., 2008). Second, midges are biting insects that only occasionally visit cocoa flowers to feed on pollen (Kaufmann, 1974, 1975; Claus et al., 2018) or chew on stomate-type nectaries located on the petals (Young et al., 1984; Young, 1985a). Only a small portion actually carries pollen grains between flowers (Kaufmann, 1975a; Winder, 1977; Jaramillo et al., 2024). Not surprisingly, therefore, pollination success by these small dipterans is rather low. In studies investigating natural cocoa pollination, researchers only occasionally found a sufficient amount of pollen grains on the female flower parts after midge visits (> 35 pollen grains, considered the threshold for pollination success, in 14.4% of flowers visited by Ceratopogonidae: Kaufmann, 1975b; > 35 pollen grains in 5.1% of flowers visited by Ceratopogonidae and Cecidomyiidae: Winder, 1977; at least one pollen grain in 29.0% of flowers visited by potential pollinators: Vansynghel et al., 2022). Even in those rare cases of effective pollen deposition, fruit set is still far from guaranteed. In their study, Vansynghel et al. (2022) observed that only about three percent of the flowers that had received more than 35 pollen grains produced young fruits. Hence, the estimated contribution of midges to crop yield may actually be less than one percent (Vansynghel et al., 2022). This value is astonishingly close to the fruit set of some self-incompatible cocoa varieties after experimental self-pollination by hand (0.0-7.4%, average = 2.3%; Branco et al., 2018).
The reduced pollination efficiency of pollinators that naturally occur in cocoa plantations has been considered the main cause for low fruit set (Groeneveld et al., 2010; Toledo-Hernández et al., 2017, 2021; Vansynghel et al., 2022). For cocoa farmers, economically more relevant than fruit set is the number of seeds produced per pod, which increases with the number of successfully deposited pollen grains (Falque et al., 1995). At least 150 pollen grains are necessary to achieve top yields of between 50 and 60 seeds/pod (Falque et al., 1995). Thus, since common arthropods hardly deposit more than 30 to 40 pollen grains (Vansynghel et al., 2022), cocoa producers invest in labor-intensive hand pollination to increase their yield (Brazil: Toledo-Hernández et al., 2023; Ghana: Wongnaa et al., 2021; Indonesia: Toledo-Hernández et al., 2020). Yet, while hand pollination appears profitable to farmers, further empirical research is required to gain a deeper understanding of long-term yield stability, as well as of the associated socioeconomic and environmental trade-offs (Wanger et al., 2021).
A possibility to improve the yield of pollinator-dependent crops, so far untested in cocoa, is targeted crop pollination by social bees. This technique differs from other forms of managed or directed pollination, which rely on the introduction of managed bees into crops (Shivanna, 2015), by pre-training the colonies on a specific target scent (Farina et al., 2023a). This olfactory priming has been shown to increase the foragers’ visits to the target flowers, thus, eventually, enhancing crop yield (sunflower: Farina et al., 2020; pear and apple: Farina et al., 2022; almond: Farina et al., 2023b). To this day, only honey bees (Apis mellifera) have been studied concerning targeted crop pollination (Farina et al., 2023a). Although foragers of this bee species may occasionally visit flowers of T. cacao (Harland, 1925; Soria, 1975; see however: Erickson et al., 1988), their body size certainly hampers their access to the female parts of the flowers (body length = 10.3 mm; intertegular distance = 3.0 mm; Kendall et al., 2019). A promising group for targeted cocoa pollination, however, are the stingless bees (Apidae, Meliponini). This pantropic group of highly eusocial bees comprises more than 600 species that vary in body size from 2 to 12 mm (Engel et al., 2023; Roubik, 2023). Except for some obligatory cleptobiotic (specialized on robbing nests of other bee species) and necrophagous species (collecting flesh from dead animals), Meliponini are considered opportunistic generalist foragers that collect nectar and pollen from a wide variety of flowering plants (Biesmeijer et al., 2005; Biesmeijer and Slaa, 2006; Bueno et al., 2023). Several small species have been observed visiting cocoa flowers (Nannotrigona punctata, Nannotrigona sp., Paratrigona subnuda, Plebeia flavocincta, Plebeia mosquito, Tetragonisca angustula: Soria, 1975; Young, 1981, 1985b; Lemos, 2014; Jaramillo et al., 2024) or were attracted to Theobroma floral oils (species not identified: Young et al., 1987, 1989; Young and Severson, 1994). For pollen collection, foragers land on the ligule of a cocoa petal and insert their head into the petal hood, where they extract pollen from the anthers through head movements. After five to ten flower visits, the bees brush the pollen grains from the mouth parts to the corbiculae on their hind tibiae (Hernandez, 1965 apud Soria, 1975; Young, 1981). Most importantly for cross-pollination, foragers visit more than one flower per cocoa tree and more than one cocoa tree per foraging trip (Young, 1981). However, whether the bees may pass through the staminode fence to reach the stigma for pollen deposition or not has remained an open controversy (observations in favor: Jaramillo et al., 2024; observations against: Young, 1981).
A presumed limitation for cocoa crop-pollination by bees is the reduced light intensity under the dense canopy of shade-trees (Young, 1985b). Despite a worldwide trend towards full-sun cocoa monocultures (Franzen and Mulder, 2007; Clough et al., 2009; Armengot et al., 2016), most of today’s cocoa production comes from family-managed farms and agroforests (Vaast and Somarriba, 2014). Shade-trees are key elements to the success of both these forms of farming because they increase nutrient cycling, preserve soil moisture, and reduce the vulnerability to climate change and erosion (Bentley et al., 2004; Tscharntke et al., 2011; Blaser-Hart et al., 2021). In addition to the shade provided by these trees, the flowers of modern cocoa cultivars are predominantly on trunks and lower branches (Young et al., 1987), which drastically reduces illumination for flower visitors. Since the flight activity of small meliponine species is limited by light level (Streinzer et al., 2016), foraging might be restricted to cocoa trees in direct sunlight (Young, 1985b). However, whether bees prefer sunlit or shaded patches may be associated with morphological features beyond body size, such as body coloration (Biesmeijer et al., 1999) or the density of thoracic and abdominal hairs (Barrett and O’Donnell, 2023). Thus, visits of cocoa flowers by small stingless bees even under heavily shaded conditions should not be ruled out.
Stingless bee species native to the Amazon region are particularly interesting candidates for targeted cocoa pollination. Their common evolutionary history with ancestral T. cacao, which depended on long-distance movement of pollen among different patches of trees, presumably with the help of bees (Young and Severson, 1994), suggests that these Meliponini may have routinely visited wild-type cocoa flowers. Intriguingly, the flowers of T. cacao show several phenological features that hint at their ancestral dependency on diurnal pollinators, including progressive anther dehiscence between sunrise and noon, maximum receptivity of stigma and style as well as peak floral odor release around noon (Erickson et al., 1987; Young et al., 1987; Jaramillo et al., 2024).
The aims of the present study were to determine whether and to which extent stingless bees may de facto be considered potential pollinators of T. cacao, and to identify promising candidate species for targeted pollination of cocoa crop in Brazil. Specifically, we asked the following questions: (1) How many stingless bee species can be considered potential cocoa pollinators, based on their natural geographic distribution (occurrence in the Amazon region) and body size? (2) What is the extension of the geographic distribution of these species (species with broad geographic range could be more suitable for managed cocoa pollination)? (3) What do we know about the timing of foraging of these species (potential overlap with the reproductive period of cocoa flowers; light level necessary for foraging)? (4) Do these bee species show a preference for a specific stratum of vegetation (foraging in lower strata like shrubs, subshrubs, or herbs evidence the possibility to visit coca flowers under shade cover)? (5) What are their preferred nesting sites (species nesting in tree cavities may be more suitable for managed beekeeping than soil-nesting species)?
2 Materials and methods
2.1 Selection of candidate Meliponini for targeted cocoa pollination
The selection of candidate meliponine species for targeted cocoa pollination was based on two main species traits, (1) their natural occurrence in the Brazilian Amazon region and (2) their body size.
2.1.1 Selection criterion 1: natural occurrence in the Brazilian Amazon
Four hundred and forty-nine species of stingless bees have been registered in the Neotropics so far (Ascher and Pickering, 2020). We focused our research on Meliponini native to the Brazilian Amazon because this region comprises an important portion of the center of origin of Theobroma cacao (Zarrillo et al., 2018; Tscharntke et al., 2023) (Figure 1A), and comprehensive datasets on stingless bees are available for this biome. Moreover, many meliponine species of the Brazilian Amazon have also been documented in neighboring countries, some even in other biomes across the Neotropics. Our first stage of species selection was based on lists of Meliponini and their geographic occurrence by Pedro (2014) and Camargo et al. (2023) as well as on a public bee biodiversity dataset from the Carajás National Forest (5° 52′ 11″ to 6° 32′ 13″ S; 49° 53′ 28″ to 50° 44′ 29″ W) in the State of Pará (Giannini et al., 2020), which lies within the major cocoa producer regions in Brazil with an annual production of 128.9 thousand tons (FAOSTAT, 2020; Brainer, 2021).
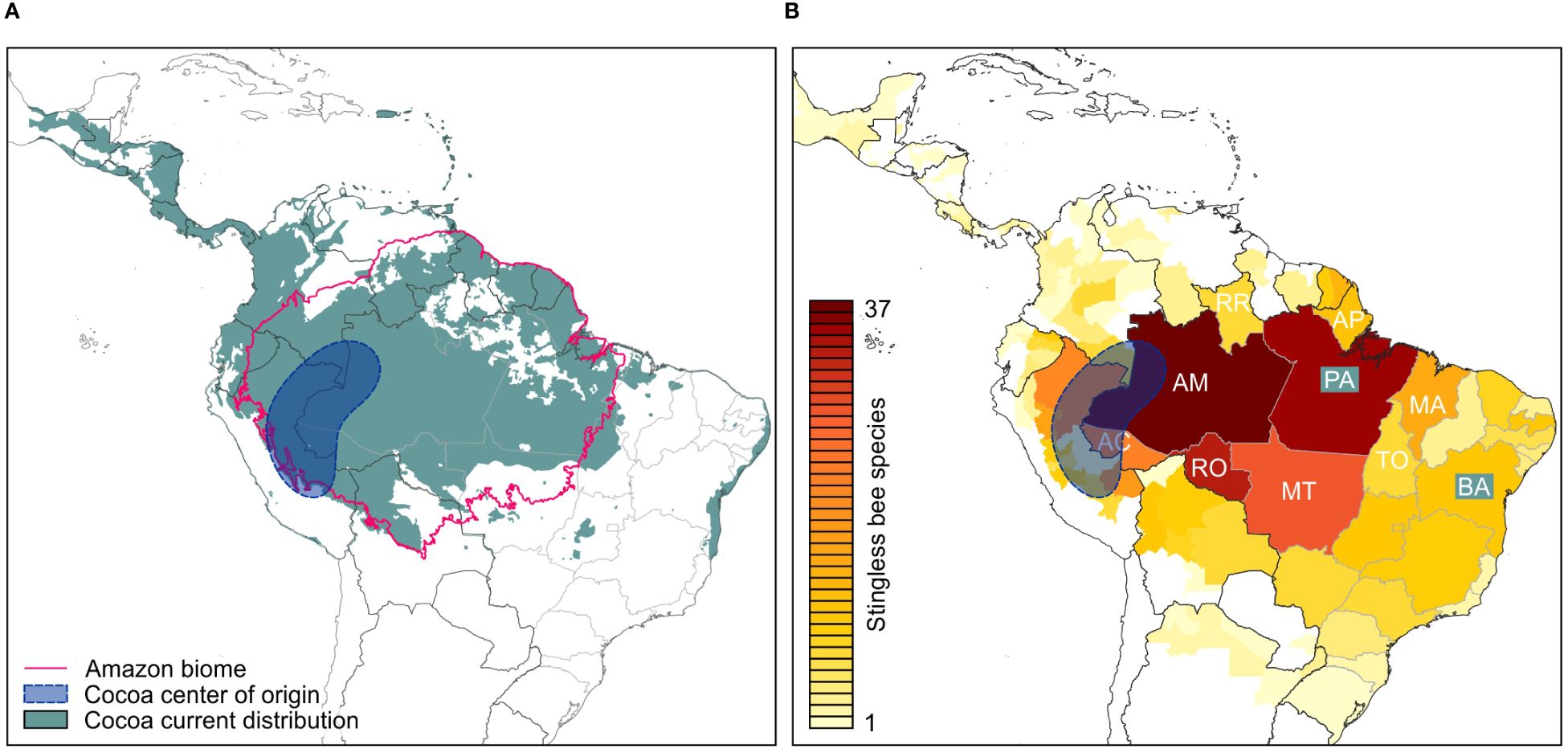
Figure 1 Cocoa distribution and richness of small stingless bee species in the Neotropics. (A) Given is the estimated distribution of Theobroma cacao (grey-shaded background) and its presumed center of origin (blue shaded area), based on Zarrillo et al. (2018), the range of the Amazon biome (pink line), country borders (black lines) and state/province borders (grey lines) of Brazil. (B) Number of small stingless bee species native to the Amazon region documented for each Brazilian state or province of other Latin American countries. Species numbers are color coded (see color scale). Brazilian states entirely or partially in the Amazon biome: AC, Acre; AM, Amazonas; AP, Amapá; MA, Maranhão; MT, Mato Grosso; PA, Pará; RO, Rondônia; RR, Roraima; TO, Tocantins. Emphasized are states with highest cocoa production (Pará, PA: 53%; Bahia, BA: 40% of Brazilian production).
2.1.2 Selection criterion 2: body size
Body size, presumably, is a key factor for effective cocoa pollination. Minute midges (Ceratopogonidae, Cecidomyiidae) with a body length of approximately 3 mm (Kaufmann, 1975b) have been proposed as the main cocoa pollinators worldwide (Glendinning, 1972; Kaufmann, 1975b; Young, 1982; Claus et al., 2018; Wolcott et al., 2023). Based on the midge scale and a recent study that points to small stingless bees, particularly Tetragonisca angustula (body length ≈ 4–5 mm; Grüter, 2020), as promising cocoa pollinators in Colombia (Jaramillo et al., 2024), we selected meliponine species of genera classified as minute or small in terms of body length according to Grüter (2020). Among the Meliponini that naturally occur in the Brazilian Amazon (see 2.1.1 Selection criterion 1), we chose bee species with an intertegular distance (ITD) of up to 1.4 mm (approximate distance between staminodes halfway to the top, estimated from images published in Wolcott et al., 2023) and a body length (BL) ranging from 2 mm to 6.5 mm. If available, ITD and BL data were taken from the datasets by Mayes et al. (2019) and Giannini et al. (2020). ITDs and BLs for species not included in these datasets were taken from relevant scientific literature (Table 1).
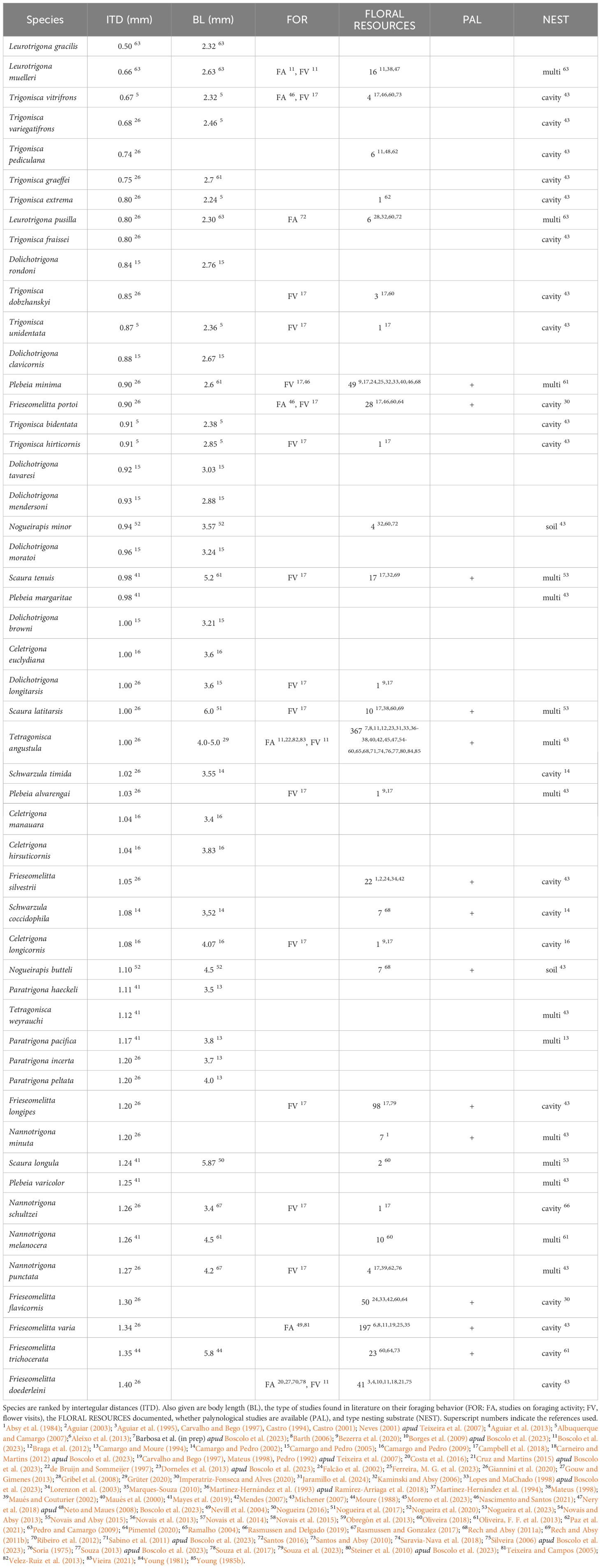
Table 1 Small Amazonian Meliponini with potential to visit cocoa flowers.
2.2 Distribution of the selected species
To determine the geographic range of occurrence of the meliponine species selected as candidates for targeted cocoa pollination (see 2.1), we consulted the online databases SpeciesLink (https://specieslink.net/), Discover Life (https://www.discoverlife.org/), and GBIF – Global Biodiversity Information Facility (https://www.gbif.org/), as well as scientific literature that provided the locations of data collection (Rasmussen and Gonzalez, 2009; Oliveira, F. F. et al., 2013; Brown and Oliveira, 2014; Misiewicz et al., 2014; Alvarez, 2015; Rasmussen and Delgado, 2019; Giannini et al., 2020; Raudales, 2022). In case georeferenced data were missing, whenever possible, we estimated the geographic position of the collected specimens from the verbatim description of the location indicated in the respective references. Distribution maps were elaborated in QGIS 3.32 (Free Software Foundation, Boston, USA).
2.3 Behavioral traits of the selected species
To estimate whether and to which extent the stingless bees, selected based on their geographic occurrence and size (see 2.1), may de facto be candidates for targeted cocoa pollination, we analyzed the scientific literature available concerning relevant behavioral traits. Particularly important characteristics in this context are the timing of the species’ foraging activity and their stratum preference. The timing of foraging, especially the foraging onset and peak activity, provide insight into whether the bees may visit cocoa flowers during the period of maximum anther dehiscence and stigma/style receptivity. Moreover, the time of foraging onset and the stratum preference indicate the capacity of a species to forage under reduced light conditions and, thus, the potential to visit cocoa flowers under dense shade as is the case in agroforests. Information on the species’ nesting behavior is relevant to estimate the possibility and facility of management.
2.3.1 Timing of foraging activity
We searched the scientific literature for data on foraging activity. We found a total of 13 studies investigating either the flight activity at nest entrances, observations of flower visits, or bee samplings at flowers (Supplementary Material 2). From studies on changes in flight activity at the nest entrance (exit and/or entrance of foragers) over time, we extracted the times of foraging onset, peak foraging activity (whenever given), and foraging end. Most commonly, observations in these studies had been performed hourly. Thus, we could not determine the exact times of first exit/peak/last entrance. In these cases, we used the full hour given in the study as reference time. Studies on flower visits (observations at flowers or bee sampling), sometimes, provided the exact moment of a given event. Most frequently, however, the time was given to the nearest hour, 30-minute, or 10-minute interval. In any case, we used the time reported in the respective study as reference time. Since “clock time” is not a very accurate information concerning foraging activity, particularly foraging onset and end, we calculated the time relative to sunrise. To obtain information on sunrise times, we used the georeferences and dates of data collection provided by the studies whenever possible. In case georeferences were missing, we estimated the approximate geographic position from the verbatim description of the location indicated in the respective study. In case study periods were provided only to the nearest month, we used the 15th day of a given month as reference date. For longer time periods, we took the median day as reference date. Sunrise times were obtained from the online app SunCalc (https://www.suncalc.org).
To understand the relation between foraging activity and light level, we used data on light intensity threshold available for two of the selected bee species (Frieseomelitta doederleini: Gouw and Gimenes, 2013; Tetragonisca angustula: Velez-Ruiz et al., 2013) and complemented these with data on a Trigonisca species from Costa Rica (Trigonisca pipioli: Streinzer et al., 2016), which has a body size similar to the Trigonisca species in our study (Albuquerque and Camargo, 2007). Nonlinear regression analysis (SigmaPlot for Windows 2013: Systat Software Inc., Richmond, USA) was used to determine the relationship between body size (ITD) and light intensity threshold. Data on average illuminance at canopy level and in the understory of tropical forests were approximated using measurements of a weather station at Mossoró in northeastern Brazil (MH, unpublished data) as well as data provided in scientific literature (Yoda, 1974; Becek and Salim, 2019; Matuso et al., 2021). Again, the time was calculated as time relative to sunrise.
To get an insight into the potential temporal overlap between bee activity and reproductive activity of cocoa flowers, we used phenological data on T. cacao flowers provided by Young et al. (1987; for similar data see: Montoya and Cruzatty, 2014; Jaramillo et al., 2024). For pollen sac dehiscence: 53.3% dehiscent at 06h15, 68.5% at 08h30, 100% at 09h15 and thereafter. For maximum stigma/style receptivity, estimated through a score of oxygen production by the structures (score includes estimates of the number of bubbles, the rate of bubbling, and the size of bubbles) after application of hydrogen peroxide: score = 19.5 at 6h15, 20.5 at 08h30, 24.5 at 09h15, 28.0 at 12h15, 26.5 at 15h15, and 23.0 at 20h00, respectively (Young et al., 1987). Considering an average cocoa-pollen longevity of three to four hours (Soria, 1975; Talledo et al., 2019; see latter reference for an exceptional longevity of 24 h for the Trinitarian cocoa-clone CCN–51), pollen harvested by insects after full dehiscence (09h15) should be viable at the time of maximum stigma/style receptivity (12h15).
2.3.2 Stratum preferences
The bees’ preferences to forage at a certain vegetational stratum provides indirect evidence for whether or not they may visit shaded flower patches. We searched the scientific literature for plant species visited by the meliponine species selected as candidates for targeted cocoa pollination. We found a total of 65 studies, providing either palynological data (analysis of pollen from storage pots, geopropolis deposited in nests, or corbicula pollen loads), or observations of bees at flowers (focal plant studies, bee samplings) (Supplementary Material 2). Information on the respective stratum of each plant species was obtained from the online databases REFLORA (https://floradobrasil.jbrj.gov.br) and Plants of the World Online (https://powo.science.kew.org). For each bee species, we calculated the proportion of plants (relative to total of plants recorded for a bee species) in each of the following strata: trees, palms, lianas, shrubs, subshrubs, and herbs. When more than one stratum type was indicated for a given plant, we attributed the respective fraction to each stratum (for instance, if a plant could grow either as [i] subshrub, [ii] shrub, or [iii] liana, we added a value of 1/3 to each of the given strata). Cases in which more than four different strata were provided by the online databases (mostly when plants had been identified only at the genus or family level in palynological studies) were excluded from the analysis.
For some bee species, we found records for less than 10 plant species. Whenever possible, we pooled the data of these species at genus level (Leurotrigona, Nannotrigona, Nogueirapis, Scaura, Trigonisca) to achieve at least 10 plant species for the analysis. In the case of Trigonisca, many studies did not identify the bees to species level. However, since all species of this genus are very similar in size (Albuquerque and Camargo, 2007), we assumed similar foraging preferences.
2.3.3 Nesting behavior
Stingless bees exhibit a wide range of nesting preferences, including associations with ant and termite nests both above and below the ground. Mostly, however, they built their nests within hollow spaces in trees, branches, rock faces, or even human-built structures. Some meliponine species display multiple nesting habits, building their nests opportunistically either below ground (soil-nesting) or in available cavities above ground (cavity-nesting) (Grüter, 2020). Categorizing bee species based on their nesting habit is important, given that some species (e.g., cavity nesters) can be easily managed in purpose-built hive boxes for rearing stingless bees (Cortopassi-Laurino et al., 2006; Venturieri et al., 2012). We classified nest sites of the selected bee species following the classification proposed by Giannini et al. (2020): cavity-nesting (arboreal, natural cavities), soil-nesting (subterranean, natural cavities), or multiple nesting (when species are reported to build their nests in more than one of the considered categories).
3 Results
Among the 449 species of currently known neotropical Meliponini, 188 species are native to the Brazilian Amazon region (Selection criterion 1: natural occurrence in the Brazilian Amazon). Out of these, 52 species have an intertegular distance ITD ≤ 1.4 mm (Selection criterion 2: body size) (Table 1).
3.1 Body size
The intertegular distance (ITD) of the 52 selected species range from 0.5 mm (Leurotrigona gracilis) to 1.4 mm (Frieseomelitta doederleini). Almost half of the species (23 spp.; 44.2%) have an ITD < 1.0 mm (Figure 2; Table 1). Concerning body length (BL), species vary from BL = 2.2 mm (Trigonisca extrema) to 6.0 mm (Scaura latitarsis). Minute species (ITD < 1.0 mm, BL ≈ 2–4 mm) are mostly represented by the genera Leurotrigona (3 spp.), Trigonisca (10 spp.), and Dolichotrigona (7 spp.).
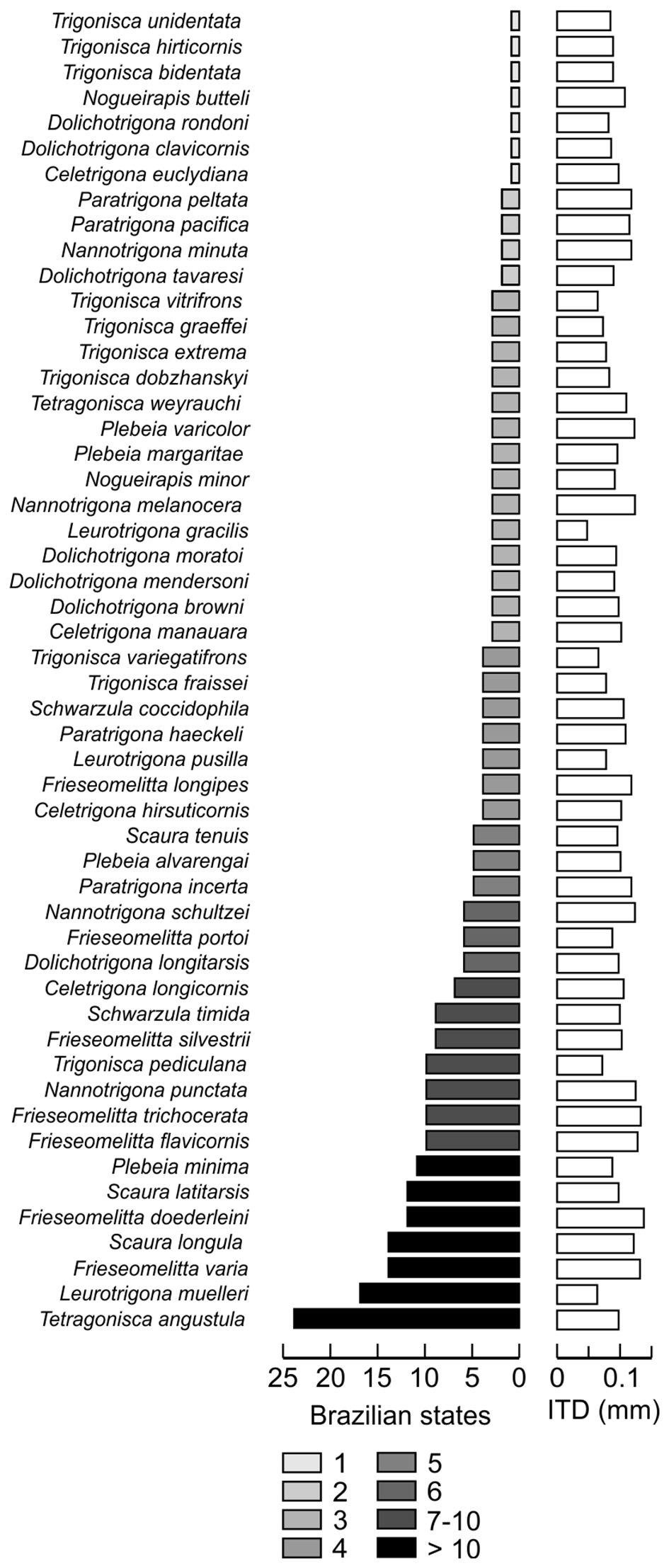
Figure 2 Small stingless bee species native to the Brazilian Amazon. Given are species names, the number of Brazilian states in which each species occurs and their respective intertegular distance (ITD).
3.2 Geographic distribution of species
To get an insight into the geographic distribution of the 52 Meliponini selected as candidates for targeted cocoa pollination, we evaluated the number of Latin-American states in which they have been documented. For the Brazilian territory, the highest numbers the selected species occur in the states of Amazonas (37 spp.), Pará (35 spp.), Rondônia (33 spp.), and Acre (22 spp.) (Figure 1B). Elevated species richness was also found in Mato Grosso (28 spp.) and Maranhão (15 spp.) (Figure 1B), both states with transitions between the Amazon Rainforest and the Cerrado ecoregion (tropical savanna). In neighboring countries, high species numbers are associated with Amazon Rainforest biomes, particularly the Peruvian Amazon jungle plain (Loreto: 21 spp.; Madre de Dios: 18 spp.; San Martin: 16 spp.), the Southwestern Amazon (Beni: 9 spp.) and the adjacent Yungas Forest in Bolivia (La Paz: 10 spp.), as well as the Amazon region in Colombia (7 spp.) (Figure 1B).
Stingless bees native to the Brazilian Amazon also occur in regions beyond the center of origin of T. cacao. Importantly, 10 of the classified species have been documented in the state of Bahia (Figure 1B), which is currently the second largest cocoa producer of Brazil (109,748 tons of cocoa beans in 2022, 40% of Brazilian production) and the largest in terms of area occupied by cocoa plantations (410,185 ha) (for comparison, Pará: 145,995 tons; 152.837 ha; 53% of Brazilian production) (IBGE, 2023).
Although some of the meliponine species identified in our study show a wide geographic distribution, with documented occurrence in more than 10 Brazilian states (7 spp.; 13.5%), most are restricted to one (7 spp.; 13.5%), two (4 spp.; 7.7%), three (14 spp.; 26.9%), or four (7 spp.; 13.5%) states (Figure 2; Supplementary Material 1). The species with the largest distribution is Tetragonisca angustula, ranging from southern Brazil, northern Paraguay and Argentina to Belize and southern Mexico (Figure 3).

Figure 3 Examples of small stingless bee species native to the Brazilian Amazon. Given are geographic positions (orange-filled circles) and the states or provinces of documented occurrences (yellow-shaded areas). Mean intertegular distance (ITD) and body length (BL) of the respective species are indicated as documented in the literature (see Table 1). Grey-shaded background, present-day cocoa distribution; blue-shaded area, presumed origin of Theobroma cacao; based on Zarrillo et al. (2018). Images of the respective species © Entomological Collection “Prof. J.M.F. Camargo,” FFCLRP/USP.
3.3 Timing of foraging
The light intensity necessary for foraging decreased exponentially with increasing body size (nonlinear regression, exponential decay: R²adj = 0.971). The smallest species for which we found information on light intensity thresholds was Trigonisca pipioli (ITD = 0.8 mm), who initiates foraging, on average, at 72 lux. Bigger species, Tetragonisca angustula (ITD = 1.0 mm) and Frieseomelitta doederleini (ITD = 1.4 mm) have documented light intensity thresholds of 30 and 10 lux, respectively (Figure 4A). In open environments or at canopy level, the illuminance necessary for foraging by these three Meliponini are reached already 30 minutes before sunrise. In the understory, however, light intensities are only 1% to 10% of those at canopy level (Figure 4B). In these shady environments, light levels necessary for foraging by T. pipioli are attained only at sunrise, whereas F. doederleini may already be active 30 minutes earlier (Figure 4B). Importantly, at the time of full anther dehiscence of coca flowers (3.8 hours after sunrise, hours) or during the period of maximum receptivity of the stigmata (6.8 hours) (Figure 5B), light levels in the understory are around 5,000 lux (Figure 4B) and, thus, far above activity threshold of small bee species (Figure 4A).
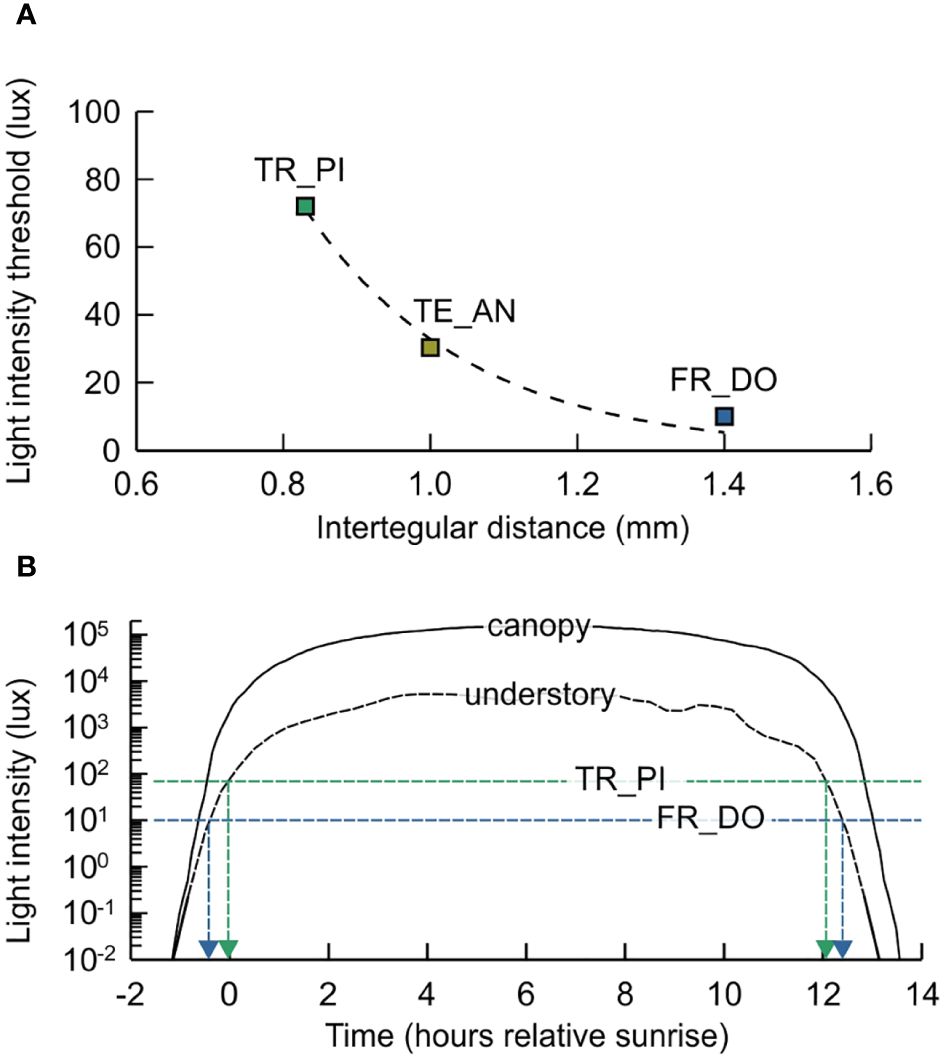
Figure 4 Association between foraging activity of small stingless bees and light intensity. (A) The illuminance level necessary for flight (Light intensity threshold) decreases exponentially with bee size (Intertegular distance) (dashed line, exponential decay regression). (B) Variations in light intensity over a day at canopy level and in the understory of tropical forests (based on MH, unpublished; Yoda, 1974; Becek and Salim, 2019; Matuso et al., 2021). Dashed arrow lines indicate the onset and end of foraging activity of small stingless bees based on their light intensity thresholds (see A). Species are abbreviated as follows: FR_DO, Frieseomelitta doederleini; TE_AN, Tetragonisca angustula; TR_PI, Trigonisca pipioli..
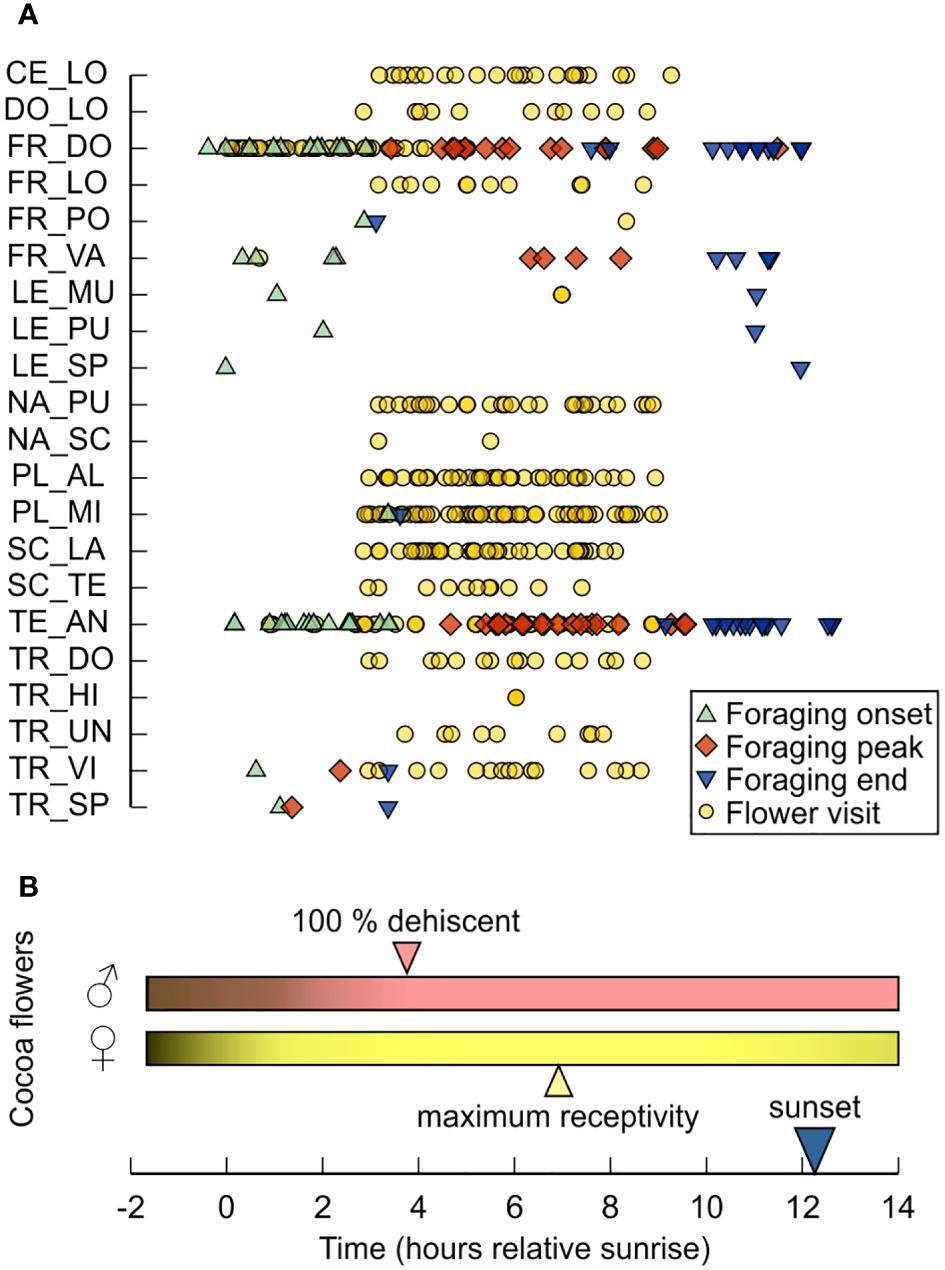
Figure 5 Foraging activity of small stingless bee species native to the Brazilian Amazon. (A) Records of foraging activity of bees in scientific literature, documented either as flower visits (circles) or as the onset (up-poiting triangle), peak (diamond), and end (down-pointing triangle) of colony flight activity; each symbol represents a single literature record (for details and references see Table 1; Supplementary Material 2). (B) Reproductive phenology of cocoa flowers. Given are the times of full dehiscence of the anthers (100% dehiscent) and the maximum receptivity of the stigmata (based on Young et al., 1987). Species are abbreviated as follows: CE_LO, Celetrigona longicornis; DO_LO, Dolichotrigona longitarsis; FR_DO, Frieseomelitta doederleini; FR_LO, Frieseomelitta longipes; FR_PO, Frieseomelitta portoi; FR_VA, Frieseomelitta varia; LE_MU, Leurotrigona muelleri; LE_PU, Leurotrigona pusilla; LE_SP, Leurotrigona sp.; NA_PU, Nannotrigona punctata; NA_SC, Nannotrigona schultzei; PL_AL, Plebeia alvarengai; PL_MI, Plebeia minima; SC_LA, Scaura latitarsis; SC_TE, Scaura tenuis; TE_AN, Tetragonisca angustula; TR_DO, Trigonisca dobzhanskyi; TR_HI, Trigonisca hirticornis; TR_UN, Trigonisca unidentata; TR_VI, Trigonisca vitrifrons; TR_SP, Trigonisca sp.
Concerning foraging activity, we found information on 21 out of the 52 classified Meliponini in 21 studies (Table 1). For 10 species we obtained data on onset and end of colony foraging activity in addition to the timing of flower visits (Figure 5A; Table 1; Supplementary Material 2). Flower visits occurred between sunrise and almost 10 hours after sunrise (earliest observation = 0.02 hours; latest observation = 9.3 hours; average = 5.3 hours) (Figure 5A). Colonies of stingless bee species, for which we found more than a single information on flight activity at the nest entrance, initiated foraging, on average on hour after sunrise, had an activity peak 6 to 7 hours after sunrise, and stopped foraging at sunset (F. doederleini: average foraging onset, FO = 1.6 hours; average foraging peak, FP = 6.5 hours; average foraging end, FE = 10.8 hours, N = 18; F. varia: FO = 1.4 hours, FP = 7.1 hours, FE = 10.9 hours, N = 4; T. angustula: FO = 1.7 hours, FP = 6.9 hours, FE = 11.1 hours, N = 21) (Figure 5A).
3.4 Stratum preferences
For the majority of Meliponini classified in our study, we found little information on the variety of plants visited, given that most species had been observed only in studies focusing on a specific pant/crop (Figure 6A; Table 1; Supplementary Material 2). The known food source spectrum broadened considerably in species for which palynological studies were available (F. flavicornis, F. longipes, F. portoi, F. silvestrii, F. trichocerata, F. varia, Nannotrigona minuta, Nogueirapis butteli, Plebeia minima, Scaura latitarsis, S. tenuis, T. angustula) (Table 1). Floral resource richness was significantly associated with the number of studies available (sigmoidal nonlinear regression: R² adj = 0.78; P < 0.0001) (Figure 6A), which highlights the need for additional investigations to get a more complete picture of the niche breath a given bee species.
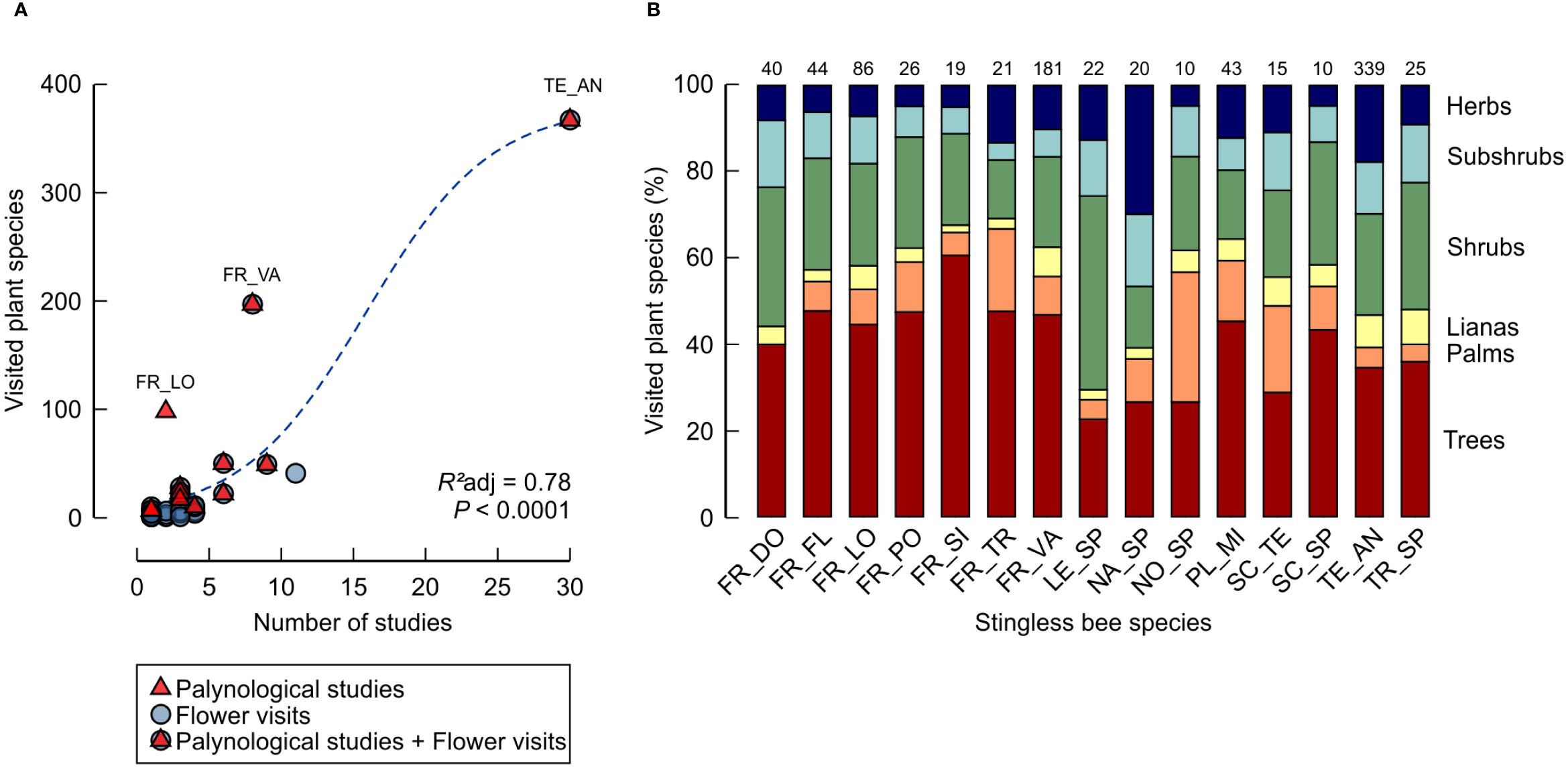
Figure 6 Floral resources used by small stingless bee species native to the Brazilian Amazon. (A) Each symbol represents a meliponine species with some record of floral resource use, including palynological studies (up-pointing triangle) and observations at flowers (circles). The number of plant species documented for a given bee species increases with the number of studies, in which this species has been investigated (dashed line, sigmoidal regression). (B) Stacked bar graph showing the proportion of floral resources in each vegetational stratum (Trees, Palms, Lianas, Shrubs, Subshrubs, Herbs). Numbers above bars indicate the number of evaluable plants (species or genera attributed to not more than 4 different strata, based on the online databases REFLORA and Plants of the World Online). Species are abbreviated as follows: FR_DO, Frieseomelitta doederleini; FR_FL, Frieseomelitta flavicornis; FR_LO, Frieseomelitta longipes; FR_PO, Frieseomelitta portoi; FR_SI, Frieseomelitta silvestrii; FR_TR, Frieseomelitta trichocerata; FR_VA, Frieseomelitta varia; LE_SP, Leurotrigona sp.; NA_SP, Nannotrigona sp.; NO_SP, Nogueirapis sp.; PL_MI, Plebeia minima; SC_TE, Scaura tenuis; SC_SP, Scaura sp.; TE_AN, Tetragonisca angustula; TR_SP, Trigonisca sp. (for details and references see Table 1; Supplementary Material 2).
The Meliponini with the highest documented food spectrum were T. angustula (367 spp.), F. varia (197 spp.) F. longipes (98 spp.) (Table 1; Supplementary Material 2). In most meliponine species/genera, for which at least 10 plant resources are specified in the literature, trees were the preferred stratum, accounting for between 22.7 to 60.5% of visited plants (average = 39.9 ± 10.4%) (Figure 6B). However, putative understory strata combined (herbs, subshrubs, shrubs, and lianas) amounted for between 33.3 and 72.7% of visited plants (average = 49.6 ± 11.4%) (Figure 6B).
3.5 Nesting behavior
We found information on the nesting behavior of 38 out of the 52 classified Meliponini in the scientific literature (9 studies). Of these, 21 species build their nests in tree cavities, two nest in the soil, and 15 utilize multiple sites. The nesting habits of 14 species have remained unknown to this date (Table 1).
4 Discussion
The clumped occurrence of ancestral Theobroma cacao, scattered over ample areas of tropical rain forest, asks for medium- to long-range pollen transfer to guarantee allele exchange between patches (Young and Severson, 1994). Thus, population maintenance based on a pure midge-based pollination, whose movement range is usually less than 10 meters (Chumacero de Schawe et al., 2018), appears a rather fruitless endeavor. Although T. cacao might be on an evolutionary transition to pollination by small dipterans (Young, 1985a), there is circumstantial evidence that hints at bees as potential pollinators of cocoa in its wild-type form. (1) The reproductive period of flowers is predominantly diurnal, with progressive dehiscence of the anthers in the early morning (full dehiscence after 09h00) and maximum receptivity of the stigmata around noon (Young et al., 1987; Montoya and Cruzatty, 2014; Jaramillo et al., 2024; Figure 5B). Although midges can be found in cocoa plantations from dawn till dusk (Young, 1985a; Frimpong et al., 2009), their activity on flowers is more common in the early morning and late afternoon (Young, 1985a). Small stingless bees, by contrast, initiate foraging at sunrise. Their peak activity around noon (Figure 5) coincides with the timing of maximum stigma/style receptivity and the concentration peak of volatiles of the floral bouquet, which contains several compounds known to attract bees (Erickson et al., 1987). (2) Flowering of T. cacao peaks between the late dry season and the mid-rainy season yet declines considerably late in the rainy season (Young, 1983; Frimpong et al., 2009 – for precipitation data see Dawoe et al., 2018). Midge populations in cocoa plantations, if not adequately managed, decrease through the dry season, which causes a certain asynchrony between the abundance of these insects and cocoa flowering (Young, 1983; Frimpong et al., 2009). Stingless bees, by contrast, are active all year round, since the maintenance of their perennial colonies requires constant food provisioning (Grüter, 2020). Colony foraging, however, is strongly influenced by the availability of floral resources in the environment (Maia-Silva et al., 2015; Campbell et al., 2019; Freitas et al., 2023). In the Amazon region, the number of flowering plants and, consequently, the food collection by stingless bee colonies decline during the rainy season (Frankie et al., 1974; Roubik, 1982; Veiga et al., 2013; Campbell et al., 2019). Thus, higher bee activity during the dry season and at the beginning of the rainy season concurs with an increase in flowering activity of T. cacao.
4.1 Small stingless bees as potential cocoa pollinators
In the present study, we highlight 52 small stingless bee species native to the Brazilian Amazon region as potential cocoa pollinators. Despite their reduced body size (intertegular distance, ITD = 0.5–1.4 mm; body length = 2.2–6.0 mm), the foraging area of these Meliponini may range between a couple hundred meters to almost one kilometer around their colonies (Grüter and Hayes, 2022). As far as is known, stingless bees collect floral resources during the day (Teixeira and Campos, 2005; Streinzer et al., 2016; Figure 5A). Although bigger species may initiate foraging at nautical twilight levels, at an illuminance below 2 lux, the eye-size and associated visual physiology of small species restrain their activity to higher light levels (Streinzer et al., 2016; Aguiar et al., 2023). It has been argued that the strongly reduced light regime owing to shading in cocoa plantations, particularly in agroforests, limits the visitation of T. cacao flowers by bees (Young, 1985b). However, foraging of small Meliponini may occur at an illuminance corresponding dawn (around 80 lux; Figure 4B). Thus, at least from the viewpoint of visual physiology, they should well be capable to visit cocoa flowers after full dehiscence of the anthers and during maximum receptivity of the stigmata even under heavy shade cover (100% dehiscence at 3.8 hours after sunrise: approx. 111,000 lux at canopy level; approx. 5,000 lux in the understory; maximum receptivity at 6.8 hours after sunrise: approx. 151,000 lux at canopy level; approx. 5,000 lux in the understory; Figure 4B). The bees’ potential to forage under reduced light regimes is corroborated by the finding that, despite a certain preference for trees, between 20 and 60% of the naturally exploited food sources are understory vegetation, including herbs, subshrubs, shrubs, and lianas (Figure 6B). Depending on the height above ground, at which these plants bloom, and the trees’ crown area, the light levels may be less than 1% of the illuminance at canopy level (Yoda, 1974; MacDougall and Kellmann, 1992; Matuso et al., 2021). Thus, food collection at understory vegetation, particularly at herbs and subshrubs in forest environments, relies on the capacity to forage under low light conditions. Although some meliponine species prefer sunlit food patches (Biesmeijer et al., 1999), this is an issue of thermal physiology, associated with body coloration rather than size (Pereboom and Biesmeijer, 2003; Ramos et al., 2024). In any case, bees that are capable of foraging in the sun without overheating may take advantage of flowers located on thin branches in the crown, which is common in ancestral forms of T. cacao in the wild (Young et al., 1987). Bees that preferentially forage at shaded patches to avoid overheating, by contrast, may visit flowers on lower branches even under densely shaded conditions as is the case in cocoa agroforests (Young et al., 1987).
Whether or not stingless bees are, de facto, pollinators of cocoa flowers, has remained an open debate. Early studies showed a low efficiency of Tetragonisca angustula as cocoa pollinator under natural conditions (0.8% of visited cocoa flowers that had not been knocked off during the visit produced fruit; control: 0.2% of flowers sheltered from floral visitors produced fruit; Young, 1981) and postulated that foragers of this meliponine species are mere pollen thieves (Young, 1981, 1985). More recently, however, it has been documented that foragers of T. angustula pass through the staminode fence and enter in contact with the stigmatic surfaces of cocoa flowers (Jaramillo et al., 2024), which gives reason to believe that at least some Meliponini may not only visit but also pollinate T. cacao. In contrast to midges, stingless bees have specialized body structures, such as the corbiculae, which allow them to effectively collect and transport large amounts of pollen, therewith facilitating pollination (Michener, 2007; Martins et al., 2014; Engel and Rasmussen, 2020). Given that social bee foragers do not collect floral resources for their own sustenance, but to provide nutrients for the colony, they typically return to previously known food sources, where they visit numerous flowers (Soria, 1975; Young, 1981). The extended duration of these visits for food collection, coupled with the adherence of pollen grains to their bodies when in contact with pistils, make bees key agents in systems depending on cross-pollination as they move between flowers (Westerkamp, 1996).
Although effective pollen deposition on the female parts of cocoa flowers by small stingless bees cannot be ruled out, it has not been observed so far. Howsoever, pollen grains littered by bees while handling the anthers may sediment in the petal hoods, thereby facilitating eventual adhesion to midges or other minute arthropods crawling through the flowers and, thus, pollen transfer to the stigma (Young et al., 1984; Young, 1985a). In addition to promoting self-pollination (thrip-mediated self-pollination of Ranunculus secleratus and Potentilla rivalis: Baker and Cruden, 1991; ant-mediated self-pollination of Blandfordia grandiflora: Ramsey, 1995), the coincidental deposition by crawling flower visitors has been shown to increase the seed set of self-incompatible plants, yet only in association with flower visits by bees (for Conospermum undulatum: Delnevo et al., 2020). Given the high variety and abundance of minute arthropods in cocoa flowers (Bigger, 2012; Toledo-Hernández et al., 2017), joint pollination involving different species or functional groups appears a promising topic for future research.
4.2 Opportunities and challenges for targeted cocoa pollination by stingless bees
A major drawback concerning the importance of stingless bees for commercial pollination of T. cacao is their reduced attraction to the flowers, which may be the result of artificial selection and cloning for agricultural purposes (Young and Severson, 1994). Thus, Meliponini must be “convinced” to visit the flowers of cultivars in cocoa plantations and agroforests. A possibility to enhance floral visits by social bees to crop flowers is through targeted pollination (Farina et al., 2023a). Prior to introducing managed colonies into crops, they are fed with sugar syrup scented with the odor of the target flowers, or a synthetic odor mixture that mimics the bouquet (Farina et al., 2020). During repeated exposure to the scented food, bees learn to associate the odor with reward and establish specific olfactory memories (Farina et al., 2005). The olfactory information gained inside the nest biases the foraging decisions of individuals in the field (Arenas et al., 2007, 2008), therewith enhancing the colonies’ visitation rate to target crop flowers (Farina et al., 2020, 2022, 2023b). Importantly for the relatively nectarless cocoa flowers (Young et al., 1984), the success of this protocol for managed pollination has been documented also for crops whose flowers offer little or no nectar reward like pear (Farina et al., 2022). Feeding colonies with scented syrup, presumably, activates mainly nectar foragers (Farina et al., 2023a). However, some foragers may switch to pollen collection, particularly if the “promised” nectar source has a lower productivity than expected by the bees based on their experience in the nest (Arenas and Kohlmaier, 2019).
These studies, all using honey bees (Apis mellifera) as model organism, highlight three conditions that must be met to guarantee the success of targeted crop pollination. (1) The bee species in question needs to be manageable. (2) The bees must show a certain level of olfactory associative learning. (3) The bees must show the capacity to transfer scent information learnt inside the nest to the foraging context. Luckily, stingless bees comply with the first two of these requirements, while the third has remained largely understudied so far. (1) Unlike with A. mellifera, there is no standard hive model for stingless bees. Traditionally, many beekeepers, who manage stingless bees, just leave the colony inside the original tree trunk (Crane, 1999; Quezada-Euán, 2018), which limits the possibilities of manipulation (Contrera et al., 2011). To improve the practicability of meliponiculture, several types of hives have been developed over the past fifty years (Nogueira-Neto, 1970; Crane, 1999; Contrera et al., 2011), varying in in size and format to optimize honey extraction and colony division of a particular species (Oliveira, R. C. et al., 2013; Barbieri et al., 2019). For the Meliponini classified in our study as potential cocoa pollinators, we found references for the nesting behavior of 38 species, 26 of which species build their nests, at least facultatively, in tree cavities (Table 1), which facilitates their transfer to rational hives. Although it is possible to keep soil nesting species in nest-boxes (Imperatriz-Fonseca, 1973, 1978; Ribeiro, 2008), new colonies are usually established in underground cavities (Imperatriz-Fonseca, 1990). Thus, attracting these species to trap-nests, a common practice by beekeepers to obtain new colonies (Oliveira, R. C. et al., 2013), is, at the very least, difficult. For only few of the Meliponini highlighted in our study, successful establishment of colonies under managed conditions has been documented so far (Frieseomelitta varia, F. longipes, F. silvestrii, F. trichocerata, Leurotrigona muelleri, Nannotrigona melanocera, Plebeia minima, Scaura latitarsis, S. longula, S. tenuis, Tetragonisca angustula: Nogueira-Neto, 1970; Mateus et al., 2013; Oliveira, F. F. et al., 2013; Barbieri et al., 2019; Leão et al., 2024). However, species usually thrive well when kept in nest-boxes of adequate size and in appropriate thermal conditions (CMS, MH, VLIF, personal observations). (2) Similarly to honey bees, stingless bees show the capacity to establish long lasting olfactory memories when trained on a specific scent (Aguiar et al., 2023). Particularly interesting for protocols of targeted crop pollination is the possibility to induce these olfactory memories through feeding the colonies with scented sucrose solution inside the nest (Mc Cabe and Farina, 2009, 2010). To this date, unfortunately, only two meliponine species have been investigated concerning the acquisition of olfactory information through in-hive feeding, one of these, however, a species classified by our study as potential cocoa pollinator (T. angustula: Mc Cabe and Farina, 2010). (3) It is most likely that foragers use learnt scent-reward contingencies for their foraging decisions. At least when trained to artificially scented food at outdoor feeders, or when exposed to a particular odor in the laboratory, foragers show a preference for this scent in the field (Mc Cabe and Farina, 2010; Roselino and Hrncir, 2012). Crucial, in any case, for a potential targeted cocoa pollination is the fact that stingless bees, similarly to honey bees, may switch from nectar to pollen foraging in case the nectar source becomes unavailable (shown in Plebeia tobagoensis: Hofstede and Sommeijer, 2006).
An important requirement for the success of any forms of directed pollination by stingless bees is an appropriate habitat quality that permits the maintenance of managed colonies within or near crops and, moreover, allows for gene exchange with natural populations. Small Meliponini, in particular, are highly vulnerable to deforestation (Mayes et al., 2019), since their reduced foraging range limits the access to food when resources become scarce or disconnected (Araújo et al., 2004; Greenleaf et al., 2007; Hrncir, 2022). Thus, at landscape-level, protection of continuous and interconnected forest areas next to crop plantations is essential to ensure the survival of bee populations (Campbell et al., 2018). To increase the biodiversity of small bees, the conservation of natural habitats and the sustainable management of agricultural landscapes are fundamental for providing sufficient resources for natural pollinators (Ulyshen et al., 2023). These resources include suitable nesting habitats (e.g., tree cavities and soil substrates) as well as plants that bees may use as additional pollen and nectar sources (Kremen et al., 2007; Ricketts et al., 2008).
In Brazil, cocoa is mostly cultivated in agroforests (Cuenca and Nazario, 2004), which are a valuable resort for managed stingless bee colonies. Agroforestry is a land management approach that involves the incorporation of trees and shrubs into agricultural and rural landscapes, aiming at enhancing productivity, profitability, diversity, and overall ecosystem sustainability (Kang and Akinnifesi, 2000; Nair, 2007, 2011; Jose, 2009; François et al., 2023). Brazilian agroforests may play an important role in stingless bee conservation due to their high floral and structural diversity (Jose, 2012), which increases the availability of both potential food sources and nesting options (Jha and Vandermeer, 2010; Delgado et al., 2022) and improves the microclimatic conditions for foraging and development (Tscharntke et al., 2011; Betrup et al., 2019). Moreover, agroforests contribute to biodiversity conservation through enhancing landscape connectivity between habitat remnants of natural habitat (Moguel and Toledo, 1999; Betrup et al., 2019). The consequently reduced pressure on biodiversity promotes functional services, including pollination (Toledo-Hernández et al., 2021) and pest control (Andow, 1991; Tscharntke et al., 2011; Ferreira, D. F. et al., 2023), which, eventually, lead to increases in crop yields (Betrup et al., 2019; Sabino et al., 2022).
4.3 The way forward
In Brazil, cocoa cultivation plays a vital role in family farming, providing income and subsistence for smallholder farmers (Cuenca and Nazario, 2004). Enhancing cocoa production through targeted crop pollination, therefore, not only increases yields, but also improves the livelihood of rural communities (Donald, 2004). Although our study highlights the potential of small Meliponini as pollinators of T. cacao, it reveals considerable gaps-of-knowledge. So far, we know only little concerning the nesting behavior and foraging activity of most neotropical species, particularly the smallest ones that, owing to size, might be most suitable for cocoa pollination (Table 1). We know even less about the Meliponini of Africa and Southeast Asia, which currently share more than 80% of the global cocoa production (Suri and Basu, 2022). Intriguingly, a survey of potential pollinators of T. cacao in Ghana reported visits of cocoa flowers by Liotrigona bouyssoui (as L. parvula) (Frimpong et al., 2009), a minute stingless bee species with a body length (BL) of less than 4 mm (Grüter, 2020). Moreover, pollen of T. cacao was detected in the honey of small Indonesian Meliponini, Tetragonula laeviceps (BL = 4.0 mm) and Heterotrogona itama (BL = 6.1 mm) (Anggadhania et al., 2020; Jayadi and Susandarini, 2020; for body size see Purwanto et al., 2022). These findings highlight the global potential of targeted cocoa pollination by stingless bees. Yet, studies are urgent to determine the physiological and behavioral requirements for targeted crop pollination, such as the capacity of olfactory associative learning and the capability to transfer scent information learnt inside the nest to the foraging context. From our list of potential cocoa pollinators, scent learning-studies have only been performed on Tetragonisca angustula (Mc Cabe and Farina, 2010). Thus, important next steps should comprise detailed studies particularly on foraging behavior and olfactory learning to validate the potential of the meliponine species from our list as cocoa pollinators. In sequence, managed colonies of the most promising species should be introduced into cocoa plantations, particularly in agroforests, to quantify the effect of their pollination services for crop yields. Studies investigating the efficiency of such directed pollination by stingless bees, so far, documented a positive effect on the yield of several economically important crops, including acai (Muto et al., 2020), apple (Viana et al., 2014), macadamia (Heard, 1999), and watermelon (Layek et al., 2021). Moreover, significant contributions of pollination by Meliponini to the fruit set in avocado (Can-Alonzo et al., 2005), coconut (Heard, 1999), and coffee (Klein et al., 2003) point to the potential of this social bee group for directed crop pollination. At local level, the incorporation of managed stingless bee colonies into family farms and agroforests may provide an efficient strategy to secure crop productivity, smallholder farmers’ income and biodiversity conservation.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Author contributions
CM: Conceptualization, Methodology, Project administration, Writing – original draft, Writing – review & editing. MH: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. TG: Writing – review & editing. MT: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. VI: Conceptualization, Writing – review & editing, Supervision, Funding acquisition.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by grants of the National Council for Scientific and Technological Development (CNPq); CMS (grants 444384/2018-9 and 382076/2023-0); MH (grant 311564/2022-4); VLIF (grants 444384/2018-9 and 315957/2023-9).
Acknowledgments
We would like to thank Eduardo Almeida for providing images of stingless bees deposited in the Entomological Collection “Prof. J.M.F. Camargo,” FFCLRP/USP, which were used to illustrate the species listed in the figures. Sincere thanks also to Christopher Brown, who shared his detailed dataset on stingless bees collected in Rondônia, and two referees, whose valuable comments and suggestions helped to improve the quality of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
VI-F declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frbee.2024.1357811/full#supplementary-material
References
Absy M. L., Camargo J. M. F., Kerr W. E., Miranda I. P. A. (1984). Espécies de plantas visitadas por Meliponinae (Hymenoptera; Apoidea), para coleta de pólen na região do Médio Amazonas. Rev. Bras. Biol. 44, 227–237.
Aguiar C. M. L. (2003). Utilização de recursos florais por abelhas (Hymenoptera, Apoidea) em uma área de Caatinga (Itatim, Bahia, Brasil). Ver. Bras. Zoo. 20, 457–467. doi: 10.1590/S0101-81752003000300015
Aguiar C. M. L., Martins C. F., Moura A. C. (1995). Recursos florais utilizados por abelhas (Hymenoptera, Apoidea) em áreas de caatinga (São João do Cariri, Pb). Rev. Nord. Biol. 10, 101–117. doi: 10.1590/S0101-81752003000300015
Aguiar C. M. L., Santos G. M. M., Martins C. F., Presley S. J. (2013). Trophic niche breadth and niche overlap in a guild of flower-visiting bees in a Brazilian dry forest. Apidologie 44, 153–162. doi: 10.1007/s13592-012-0167-4
Aguiar J.M.R.B.V., Silva R. C., Hrncir M. (2023). Ecological drivers of bee cognition: insights from stingless bees. Behav. Ecol. Sociobiol. 77, 128. doi: 10.1007/s00265-023-03406-7
Albuquerque P. M. C. D., Camargo J. M. F. (2007). Espécies novas de Trigonisca Moure (Hymenoptera, Apidae, Apinae). Rev. Bras. Entomol. 51, 160–175. doi: 10.1590/S0085-56262007000200005
Aleixo K. P., Faria L. B., Garófalo C. A., Imperatriz Fonseca V. L., Silva C. I. (2013). Pollen collected and foraging activities of Frieseomelitta varia (Lepeletier) (Hymenoptera: Apidae) in an urban landscape. Sociobiology 60, 266–276. doi: 10.13102/sociobiology.v60i3.266-276
Alvarez L. J. (2015). Diversidad de las abejas nativas de la tribu Meliponini (Hymenoptera, Apidae) en Argentina (La Plata: Universidad Nacional de la Plata).
Andow D. A. (1991). Vegetational diversity and arthropod population response. Annu. Rev. Entomol. 36, 561–586. doi: 10.1146/annurev.en.36.010191.003021
Anggadhania L., Wahyuni N., Rizqiani K. D. (2020). Melissopalinological characteristic of stingless bee (Trigona/Tetragonula) honey in Lombok, West Nusa Tenggara. Environ. Earth Sci. 457, 12062. doi: 10.1088/1755-1315/457/1/012062
Araújo E. D., Costa M., Chaud-Netto J., Fowler H. G. (2004). Body size and flight distance in stingless bees (Hymenoptera: Meliponini): inference of flight range and possible ecological implications. Braz. J. Biol. 64, 563–568. doi: 10.1590/S1519-69842004000400003
Arenas A., Kohlmaier M. G. (2019). Nectar source profitability influences individual foraging preferences for pollen and pollen-foraging activity of honeybee colonies. Behav. Ecol. Sociobiol. 73, 1–10. doi: 10.1007/s00265-019-2644-5
Arenas A., Fernández V. M., Farina W. M. (2007). Floral odor learning within the hive affects honeybees’ foraging decisions. Naturwissenschaften 94, 218–222. doi: 10.1007/s00114-006-0176-0
Arenas A., Fernández V. M., Farina W. M. (2008). Floral scents experienced within the colony affect long-term foraging preferences in honeybees. Apidologie 39, 714–722. doi: 10.1051/apido:2008053
Armengot L., Barbieri P., Andres C., Milz J., Schneider M. (2016). Cacao agroforestry systems have higher return on labor compared to full-sun monocultures. Agron. Sustain. Dev. 36, 70. doi: 10.1007/s13593-016-0406-6
Ascher J. S., Pickering J. (2020) Discover life bee species guide and world checklist (Hymenoptera: Apoidea: Anthophila). Available at: http://www.discoverlife.org/mp/20q?guide=Apoidea_species (Accessed January 10, 2024).
Baker J. D., Cruden R. W. (1991). Thrips-mediated self-pollination of two facultatively xenogamous wetland species. Botany 78, 959–963. doi: 10.1002/j.1537-2197.1991.tb14499.x
Barbieri C., Pinheiro G. L., Drago P. M., Francoy T. M. (2019). A scientific note on a stingless bee hive model for ecological and behavioral studies and for environmental education. Sociobiology 66, 186–189. doi: 10.13102/sociobiology.v66i1.3401
Barrett M., O’Donnell S. (2023). Individual reflectance of solar radiation confers a thermoregulatory benefit to dimorphic males bees (Centris pallida) using distinct microclimates. PloS One 18, e0271250. doi: 10.1371/journal.pone.0271250
Barth O. M. (2006). Palynological analysis of geopropolis samples obtained from six species of Meliponinae in the Campus of the Universidade de Ribeirão Preto, USP, Brazil. Apiacta 1, 1–14.
Becek K., Salim K. A. (2019). Air temperature and light intensity in a tropical rainforest of Brunei Darussalam: Time series recorded in 2017. Earth Syst. Sci. Data Discuss. doi: 10.5194/essd-2019-35. preprint.
Beg M. S., Ahmad S., Jan K., Bashir K. (2017). Status, supply chain and processing of cocoa-A review. Trends Food Sci. Technol. 66, 108–116. doi: 10.1016/j.tifs.2017.06.007
Bentley J. W., Boa E., Stonehouse J. (2004). Neighbor trees: shade, intercropping, and cacao in Ecuador. Hum. Ecol. 32, 241–270. doi: 10.1023/B:HUEC.0000019759.46526.4d
Betrup G., Hopwood J., Adamson N. L., Vaughan M. (2019). Temperate agroforestry systems and insect pollinators: a review. Forests 10, 981. doi: 10.3390/f10110981
Bezerra L. A., Campbell A. J., Brito T. F., Menezes C., Maués M. M. (2020). Pollen loads of flower visitors toaçaí palm (Euterpe oleracea) and implications for management of pollination services. Neotrop. Entomol. 49, 482–490. doi: 10.1007/s13744-020-00790-x
Biesmeijer J. C., Slaa E. J. (2006). The structure of eusocial bee assemblages in Brazil. Apidologie 37, 240–258. doi: 10.1051/apido:2006014
Biesmeijer J. C., Richter J. A. P., Smeets M. J. P., Sommeijer M. J. (1999). Niche differentiation in nectar collecting stingless bees: the influence of morphology, floral choice and interference competition. Ecol. Entomol. 24, 380–388. doi: 10.1046/j.1365-2311.1999.00220.x
Biesmeijer J. C., Slaa E. J., Siqueira M. C., Viana B. F., Kleinert A., Imperatriz-Fonseca V. L. (2005). Connectance of Brazilian social bee – food plant networks is influenced by habitat, but not by latitude, altitude or network size. Biota Neotrop. 5, 1–9. doi: 10.1590/S1676-06032005000100010
Bigger M. (2012). A geographical distribution list of insects and mites associated with cocoa, derived from literature published before 2012 (INCOCOA Virtual Library). Available at: http://www.incocoa.org/incoped/documents.php (Accessed March 12, 2024).
Blaser-Hart W. J., Hart S. P., Oppong J., Kyereh D., Yeboah E., Six J. (2021). The effectiveness of cocoa agroforests depends on shade-tree canopy height. Agric. Ecosyst. Environ. 322, 107676. doi: 10.1016/j.agee.2021.107676
Borges L. A., Sobrinho M. S., Lopes A. V. (2009). Phenology, pollination, and breeding system of the threatened tree Caesalpinia eChinata Lam. (Fabaceae), and a review of studies on the reproductive biology in the genus. Flora: Morphol. Distrib. Funct. Ecol. Plants. 204, 111–130. doi: 10.1016/j.flora.2008.01.003
Boscolo D., Rodrigues B. N., Ferreira P. A., Lopes L. E., Tonetti V. R., Santos I. C. R., et al. (2023). Atlantic flower–invertebrate interactions: A data set of occurrence and frequency of floral visits. Ecology 104, e3900. doi: 10.1002/ecy.3900
Braga J. A., Sales É. O., Neto J. S., Conde M. M., Barth O. M., Lorenzon M. C. (2012). Floral sources to Tetragonisca angustula (Hymenoptera: Apidae) and their pollen morphology in a Southeastern Brazilian Atlantic Forest. Ver. Biol. Trop. 60, 1491–1501.
Branco S. M. J., Silva D. V., Lopes U. V., Corrêa R. X. (2018). Characterization of the sexual self- and cross-compatibility in genotypes of cacao. Am. J. Plant Sci. 9, 1794–1806. doi: 10.4236/ajps.2018.99131
Brown J. C., Oliveira M. L. (2014). The impact of agricultural colonization and deforestation on stingless bee (Apidae: Meliponini) composition and richness in Rondônia, Brazil. Apidologie 45, 172–188. doi: 10.1007/s13592-013-0236-3
Bueno F. G. B., Kendall L., Alves D. A., Tamara M. L., Heard T., Latty T., et al. (2023). Stingless bee floral visitation in the global tropics and subtropics. Glob. Ecol. Conserv. 43, e02454. doi: 10.1016/j.gecco.2023.e02454
Camargo J. M. F., Moure J. S. (1994). Meliponini neotropicais: os gêneros Paratrigona Schwarz 1938 e Aparatrigona Moure 1951 (Hymenoptera, Apidae). Arquivos Zool. 32, 33–109. doi: 10.11606/issn.2176-7793.v32i2p33-109
Camargo J. M. F., Pedro S. R. M. (2002). Uma espécie nova de Schwarzula da Amazônia (Hymenoptera, Apidae, Meliponini). Iheringia 92, 101–112. doi: 10.1590/S0073-47212002000300011
Camargo J. M. F., Pedro S. R. M. (2005). Meliponini neotropicais: o gênero Dolichotrigona Moure (Hymenoptera, Apidae, Apinae). Rev. Bras. Entomol. 49, 69–92. doi: 10.1590/S0085-56262005000100008
Camargo J. M. F., Pedro S. R. M. (2009). Neotropical Meliponini: the genus Celetrigona Moure (Hymenoptera: Apidae, Apinae). Zootaxa 2155, 37–54. doi: 10.11646/zootaxa.2155.1
Camargo J. M. F., Pedro S. R. M., Melo G. A. R. (2023).Meliponini lepeletier 1836. In: Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region - Online Version. Available at: http://www.moure.cria.org.br/catalogue (Accessed August 12, 2023).
Campbell A. J., Carvalheiro L. G., Maués M. M., Jaffé R., Giannini T. C., Freitas M. A. B., et al. (2018). Anthropogenic disturbance of tropical forests threatens pollination services to açaí palm in the Amazon River delta. J. Appl. Ecol. 55, 1725–1736. doi: 10.1111/1365-2664.13086
Campbell A. J., Gomes R. L. C., da Silva K. C., Contrera F. A. L. (2019). Temporal variation in homing ability of the neotropical stingless bee Scaptotrigona aff. postica (Hymenoptera: Apidae: Meliponini). Apidologie 50, 720–732. doi: 10.1007/s13592-019-00682-z
Can-Alonzo C., Quezada-Euán J. J. G., Xiu-Ancona P., Moo-Valle H., Valdovinos-Nunez G. R., Medina-Peralta S. (2005). Pollination of “criollo” avocados (Persea americana) and the behaviour of associated bees in subtropical mexico. J. Apicult. Res. 44, 3–8. doi: 10.1080/00218839.2005.11101138
Carneiro L. T., Martins C. F. (2012). Africanized honey bees pollinate and preempt the pollen of Spondias mombin (Anacardiaceae) flowers. Apidologie 43, 474–486. doi: 10.1007/s13592-011-0116-7
Carvalho A. M. C., Bego L. R. (1997). Exploitation of available resources by bee fauna (Apoidea-Hymenoptera) in the Reserva Ecológica do Panga, Uberlândia, state of Minas Gerais, Brazil. Rev. Bras. Entomol. 41, 101–107.
Castro M. S. (1994). Composição, fenologia e visita às flores pelas espécies de Apidae em um ecossistema de caatinga (Nova Casa Nova, Bahia 9°26’S/41°50’W). Master thesis. Universidade de São Paulo, São Paulo.
Castro M. S. (2001). Comunidade de abelhas (Hymenoptera: Apoidea) de uma área de caatingas arbórea entre os inselbergs de Milagres (12°53’S; 39°51’W), Bahia. Doctoral thesis. Universidade de São Paulo, São Paulo.
Chumacero de Schawe C., Kessler M., Hensen I., Tscharntke T. (2018). Abundance and diversity of flower visitors on wild and cultivated cacao (Theobroma cacao L.) in Bolivia. Agrofor. Syst. 92, 117–125. doi: 10.1007/s10457-016-0019-8
Claus G., Vanhove W., Van Damme P., Smagghe G. (2018). “Challenges in cocoa pollination: The case of Côte d’Ivoire,” in Pollination in Plants. Ed. Mokwala P. W. (IntechOpen, London), 39–58. doi: 10.5772/intechopen.75361
Clough Y., Faust H., Tscharntke T. (2009). Cacao boom and bust: sustainability of agroforests and opportunities for biodiversity conservation. Conserv. Lett. 2, 197–205. doi: 10.1111/j.1755-263X.2009.00072.x
Contrera F. A. L., Menezes C., Venturieri G. C. (2011). New horizons on stingless beekeeping (Apidae, Meliponini). Rev. Bras. Zootecnia 40, 48–51.
Cornejo O. E., Yee M. C., Dominguez V., Andrews M., Sockell A., Strandberg E., et al. (2018). Population genomic analyses of the chocolate tree, Theobroma cacao L., provide insights into its domestication process. Commun. Biol. 1, 1–12. doi: 10.1038/s42003-018-0168-6
Cortopassi-Laurino M., Imperatriz-Fonseca V. L., Roubik D. W., Dollin A., Heard T., Aguilar I. B., et al. (2006). Global meliponiculture: challenges and opportunities. Apidologie 37, 275–292. doi: 10.1051/apido:2006027
Costa N. B. L., Silva M. C. J., Souza E. A., Brasil D. F., Guimarães-Brasil M. O. (2016). “Influência dos fatores climáticos na atividade de voo da abelha Frieseomelitta doederleini (Friese 1900) (Apidae: Meliponinae) em Marcelino Vieira, Rio Grande do Norte, Brasil,” in Anais do I. Congresso Internacional da Diversidade do Semiárido (Instituto Nacional do Semiárido, Campina Grande).
Crane E. (1999). The World History of Beekeeping and Honey Hunting (New York: Routledge). doi: 10.4324/9780203819937
Cruz R. M., Martins C. F. (2015). Pollinators of Richardia grandiflora (Rubiaceae): an important ruderal species for bees. Neotrop. Entomol. 44, 21–29. doi: 10.1007/s13744-014-0252-7
Cuenca M. A. G., Nazario C. C. (2004). Importância Econômica e Evolução da Cultura do Cacau no Brasil e na Região dos Tabuleiros Costeiros da Bahia entre 1990 e 2002 (Aracaju: Empresa Brasileira de Pesquisa Agropecuária).
Dawoe E. K., Barnes V. R., Oppong S. K. (2018). Spatio-temporal dynamics of gross rainfall partitioning and nutrient fluxes in shaded-cocoa (Theobroma cocoa) systems in a tropical semi-deciduous forest. Agroforest. Syst. 92, 397–413. doi: 10.1007/s10457-017-0108-3
de Bruijn L. L. M., Sommeijer M. J. (1997). Colony foraging in different species of stingless bees (Apidae, Meliponinae) and the regulation of individual nectar foraging. Insect. Soc 44, 35–47. doi: 10.1007/s000400050028
Delgado C., Fine P. V. A., Atías G. (2022). Plant species used by Melipona eburnea bees in seasonally-flooded agroforestry systems in the Peruvian Amazon. Livest. Res. Rural. Dev. 34, 104.
Delnevo N., van Etten E. J., Clemente N., Fogu L., Pavarani E., Byrne M., et al. (2020). Pollen adaptation to ant pollination: a case study from the Proteaceae. Ann. Bot. 126, 377–386. doi: 10.1093/aob/mcaa058
Donald P. F. (2004). Biodiversity impacts of some agricultural commodity production systems. Conserv. Biol. 18, 17–37. doi: 10.1111/j.1523-1739.2004.01803.x
Dorneles L. L., Zillikens A., Steiner J., Padilha M. T. S. (2013). Biologia da polinização de Euterpe edulis Martius (Arecaceae) e associação com abelhas sociais (Apidae: Apini) em sistema agroflorestal na Ilha de Santa Catarina. Iheringia Ser. Bot. 6, 47–57.
Engel M. S., Rasmussen C. (2020). “Corbiculate bees,” in Encyclopedia of Social Insects. Ed. Starr C. (Springer, Cham). doi: 10.1007/978-3-319-90306-4_30-1
Engel M. S., Rasmussen C., Ayala R., Oliveira F. F. (2023). Stingless bee classification and biology (Hymenoptera, Apidae): a review, with an updated key to genera and subgenera. Zookeys 1172, 239–312. doi: 10.3897/zookeys.1172.104944
Erickson B. J., Young A. M., Strand M. A., Erickson E. H. (1987). Pollination biology of Theobroma and Herrania (Sterculiaceae)—II. Analyses of floral oils. Insect Sci. Applic. 8, 301–310. doi: 10.1017/S1742758400002575
Erickson E. H., Young A. M., Erickson B. J. (1988). Pollen collection by honeybees (Hymenoptera: Apidae) in a Costa Rican cacao (Theobroma cacao) plantation. J. Apicult. Res. 27, 190–196. doi: 10.1080/00218839.1988.11100801
Falcão M. A., Paraluppi N. D., Clement C. R. (2002). Fenologia e produtividade do jambo (Syzygium malaccensis) na Amazônia Central. Acta Amazon. 32, 3–8. doi: 10.1590/1809-43922002321008
Falque M., Vincent A., Vaissiere B. E., Eskes A. B. (1995). Effect of pollination intensity on fruit and seed set in cacao (Theobroma cacao L.). Sex Plant Reprod. 8, 354–360. doi: 10.1007/BF00243203
FAOSTAT (2020). Food and Agriculture Organization of the United Nations: Data Production and Trade. Available at: http://www.fao.org/faostat/en/#data (Accessed July 26, 2023).
Farina W. M., Grüter C., Díaz P. C. (2005). Social learning of floral odours inside the honeybee hive. Proc. R. Soc B. 272, 1923–1928. doi: 10.1098/rspb.2005.3172
Farina W. M., Arenas A., Díaz P. C., Susic Martin C., Estravis-Barcala M. C. (2020). Learning of a mimic odor within honey bee hives improves pollination service efficiency in a commercial crop. Curr. Biol. 30, 1–7. doi: 10.1016/j.cub.2020.08.018
Farina W. M., Arenas A., Díaz P. C., Susic Martin C., Corriale M. J. (2022). In-hive learning of specific mimic odours as a tool to enhance honey bee foraging and pollination activities in pear and apple crops. Sci. Rep. 12, 20510. doi: 10.1038/s41598-022-22985-5
Farina W. M., Arenas A., Estravis-Barcala M. C., Palottini F. (2023a). Targeted crop pollination by training honey bees: advances and perspectives. Front. Bee Sci. 1. doi: 10.3389/frbee.2023.1253157
Farina W. M., Palottini F., Estravis-Barcala M. C., Arenas A., Balbuena M. S., González A. (2023b). Conditioning honeybees to a specific mimic odor increases foraging activity on a self-compatible almond variety. Apidologie 54, 40. doi: 10.1007/s13592-023-01019-7
Ferreira M. G., Absy M. L., Rezende A. C. C. (2023). Pollen collected and trophic interactions between stingless bees of the genera Melipona, Frieseomelitta and Plebeia (Apidae: Meliponini) raised in Central Amazon. J. Apicult. Res. 62, 692–704. doi: 10.1080/00218839.2021.1898837
Ferreira D. F., Jarrett C., Wandji A. C., Atagana P. J., Rebelo H., Maas B., et al. (2023). Birds and bats enhance yields in Afrotropical cacao agroforests only under high tree-level shade cover. Agric. Ecosyst. Environ. 345, 108325. doi: 10.1016/j.agee.2022.108325
François M., Pontes M. C. G., Lima A. S., Mariano-Neto E. (2023). Impacts of cacao agroforestry systems on climate change, soil conservation, and water resources: a review. Water Policy 25, 564–581. doi: 10.2166/wp.2023.164
Frankie G. W., Baker H. G., Opler P. A. (1974). Comparative phenological studies of trees in tropical wet and dry forests in the lowlands of Costa Rica. J. Ecol. 62, 881–891. doi: 10.2307/2258961
Franzen M., Mulder M. B. (2007). Ecological, economic and social perspectives on cocoa production worldwide. Biodivers. Conserv. 16, 3835–3849. doi: 10.1007/s10531-007-9183-5
Freitas P. V. D. X., Faquinello P., Arnold E., Ferro D. A. C., Ferro R. A. C., Lacerda M. L. G., et al. (2023). Flight radius and climatic conditions affect the external activity of stingless bee Melipona rufiventris (Lepeletier 1836). Braz. J. Biol. 83, e275645. doi: 10.1590/1519-6984.275645
Frimpong E. A., Gordon I., Kwapong P. K., Gemmill-Herren B. (2009). Dynamics of cocoa pollination: tools and applications for surveying and monitoring cocoa pollinators. Int. J. Trop. Insect Sci. 29, 62–69. doi: 10.1017/S1742758409990117
Frimpong-Anin K., Adjaloo M. K., Kwapong P. K., Oduro W. (2014). Structure and stability of cocoa flowers and their response to pollination. J. Bot. 2014, 513623. doi: 10.1155/2014/513623
Giannini T. C., Costa W. F., Borges R. C., Miranda L., Costa C. P. W., Saraiva A. M., et al. (2020). Climate change in the Eastern Amazon: crop-pollinator and occurrence-restricted bees are potentially more affected. Reg. Environ. Change 20, 9. doi: 10.1007/s10113-020-01611-y
Glendinning D. R. (1972). Natural pollination of cocoa. New Phytol. 71, 719–729. doi: 10.1111/j.1469-8137.1972.tb01284.x
Gouw M. S., Gimenes M. (2013). Differences of the daily flight activity rhythm in two Neotropical stingless bees (Hymenoptera, Apidae). Sociobiology 60, 183–189. doi: 10.13102/sociobiology.v60i2.183-189
Greenleaf S. S., Williams N. M., Winfree R., Kremen C. (2007). Bee foraging ranges and their relationship to body size. Oecologia 153, 589–596. doi: 10.1007/s00442-007-0752-9
Gribel R., Queiroz A. L., Assis M. G., Oliveira F. F., Queiroz M. L., Palácio C. (2008). Polinização e Manejo dos Polinizadores do Cupuaçu (Theobroma grandiflorum) (Manaus: Instituto Nacional de Pesquisas da Amazônia).
Groeneveld J. H., Tscharntke T., Moser G., Clough Y. (2010). Experimental evidence for stronger cacao yield limitation by pollination than by plant resources. Perspect. Plant Ecol. Evol. Syst. 12, 183–191. doi: 10.1016/j.ppees.2010.02.005
Grüter C., Hayes L. (2022). Sociality is a key driver of foraging ranges in bees. Curr. Biol. 32, 5390–5397e3. doi: 10.1016/j.cub.2022.10.064
Harland S. C. (1925). Studies in cacao. Part I. The method of pollination. Ann. Appl. Biol. 12, 403–409. doi: 10.1111/j.1744-7348.1925.tb04236.x
Heard T. (1999). The role of stingless bees in crop pollination. Annu. Rev. Entomol 44, 183–206. doi: 10.1146/annurev.ento.44.1.183
Hernandez J. (1965). Insect pollination of cacao (Theobroma cacao L.) in Costa Rica. Doctoral thesis. University of Wisconsin, Madison.
Hofstede F. E., Sommeijer M. J. (2006). Effect of food availability on individual foraging specialisation in the stingless bee Plebeia tobagoensis (Hymenoptera, Meliponini). Apidologie 37, 387–397. doi: 10.1051/apido:2006009
Hrncir M. (2022). Pollinator conservation: Where will bees go in the Anthropocene future? Curr. Biol. 32, R1351–R1353. doi: 10.1016/j.cub.2022.11.024
IBGE - Instituto Brasileiro de Geografia e Estatística (2023). Produção de cacau. Available at: https://www.ibge.gov.br/explica/producao-agropecuaria/cacau/br (Accessed November 22, 2023).
Imperatriz-Fonseca V. L. (1973). Miscellaneous observations on the behaviour of Schwarziana quadripunctata (Hym., Apidae, Meliponinae). Bol. Zool. Biol. Mar. 30, 633–640. doi: 10.11606/issn.2526-3366.bzbm.1973.121366
Imperatriz-Fonseca V. L. (1978). Studies on Paratrigona subnuda (Moure) Hymenoptera, Apidae, Meliponinae - III: queen supersedure. Bol. Zool. 3, 153–162. doi: 10.11606/issn.2526-3358.bolzoo.1978.121658
Imperatriz-Fonseca V. L. (1990). “Swarming activity in Schwarziana quadripunctata (Apidae, Meliponinae),” in Social Insects and the Environment. Eds. Veeresh G. K., Mallik B., Viraktamath C. A. (Oxford & IBH, New Delhi), 744–745.
Imperatriz-Fonseca V. L., Alves D. A. (2020). Stingless bees of Pará: based on the scientific expeditions of João MF Camargo (Belém: Instituto Tecnológico Vale).
Jaramillo M. A., Reyes-Palencia J., Jimenez P. (2024). Floral biology and flower visitors of cocoa (Theobroma Cacao L.) in the upper Magdalena Valley, Colombia. Flora 313, 152480. doi: 10.1016/j.flora.2024.152480
Jayadi L. Z., Susandarini R. (2020). Melissopalynological analysis of honey produced by two species of stingless bees in Lombok Island, Indonesia. Nusantara Biosci. 12, 97–108. doi: 10.13057/nusbiosci/n120203
Jha S., Vandermeer J. H. (2010). Impacts of coffee agroforestry management on tropical bee communities. Biol. Conserv. 143, 1423–1431. doi: 10.1016/j.biocon.2010.03.017
Jose S. (2009). Agroforestry for ecosystem services and environmental benefits: an overview. Agroforest. Syst. 76, 1–10. doi: 10.1007/s10457-009-9229-7
Jose S. (2012). Agroforestry for conserving and enhancing biodiversity. Agroforest. Syst. 85, 1–8. doi: 10.1007/s10457-012-9517-5
Kaminski A. C., Absy M. L. (2006). Bees visitors of three species of Clusia (Clusiaceae) flowers in Central Amazonia. Acta Amazon. 36, 259–264. doi: 10.1590/S0044-59672006000200016
Kang B. T., Akinnifesi F. K. (2000). Agroforestry as alternative land-use production systems for the tropics. Nat. Res. Forum 24, 137–151. doi: 10.1111/j.1477-8947.2000.tb00938.x
Kaufmann T. (1974). Behavioral biology of a cocoa pollinator, Forcipomyia inornatipennis (Diptera: Ceratopogonidae) in Ghana. J. Kansas Entomol. Soc 47, 541–548.
Kaufmann T. (1975a). Ecology and behavior of cocoa pollinating Ceratopogonidae in Ghana, W. Africa. Environ. Entomol. 4, 347–351. doi: 10.1093/ee/4.2.347
Kaufmann T. (1975b). Studies on the ecology and biology of a cocoa pollinator, Forcipomyia squamipennis I. & M. (Diptera, Ceratopogonidae), in Ghana. Bull. Entomol. Res. 65, 263–268. doi: 10.1017/S0007485300005940
Kendall L. K., Rader R., Gagic V., Cariveau D. P., Albrecht M., Baldock K. C. R., et al. (2019). Pollinator size and its consequences: Robust estimates of body size in pollinating insects. Ecol. Evol. 9, 1702–1714. doi: 10.1002/ece3.4835
Klein A. M., Cunningham S. A., Bos M., Steffan-Dewenter I. (2008). Advances in pollination ecology from tropical plantation crops. Ecology 89, 935–943. doi: 10.1890/07-0088.1
Klein A. M., Steffan-Dewenter I., Tscharntke T. (2003). Fruit set of highland coffee increases with the diversity of pollinating bees. Proc. R. Soc Lond. B 270, 955–961. doi: 10.1098/rspb.2002.2306
Kremen C., Williams N. M., Aizen M. A., Gemmill-Herren B., LeBuhn G., Minckley R., et al. (2007). Pollination and other ecosystem services produced by mobile organisms: a conceptual framework for the effects of land-use change. Ecol. Lett. 10, 299–314. doi: 10.1111/j.1461-0248.2007.01018.x
Layek U., Kundu A., Bisui S., Karmakar P. (2021). Impact of managed stingless bee and western honey bee colonies on native pollinators and yield of watermelon: A comparative study. Ann. Agric. Sci. 66, 38–45. doi: 10.1016/j.aoas.2021.02.004
Leão K. L., Campbell A., Veiga J., Menezes C., Contrera F. A. L. (2024). Colony size of amazonian stingless bees and its assessment through intrinsic parameters. J. Apicult. Res. doi: 10.1080/00218839.2024.2327114
Lemos C. Q. (2014). Abelha Plebeia cf. flavocincta como potencial polinizador do cacaueiro (Theobroma cacao L.) no semiárido brasileiro. Master thesis. Universidade Federal do Ceará, Fortaleza.
Lopes A. V., MaChado I. C. (1998). Floral biology and reproductive ecology of Clusia nemorosa (Clusiaceae) in northeastern Brazil. Plant Syst. Evol. 213, 71–90. doi: 10.1007/BF00988909
Lorenzon M. C. A., Matrangolo C. A. R., Schoereder J. H. (2003). Flora visited by the eusocial bees (Hymenoptera, Apidae) in a savanna of the south of Piauí, Brazil. Neotrop. Entomol. 32, 27–36. doi: 10.1590/S1519-566X2003000100004
MacDougall A., Kellmann M. (1992). The understorey light regime and patterns of tree seedlings in tropical riparian forest patches. J. Biogeogr. 19, 667–675. doi: 10.2307/2845708
Maia-Silva C., Hrncir M., Silva C. I., Imperatriz-Fonseca V. L. (2015). Survival strategies of stingless bees (Melipona subnitida) in an unpredictable environment, the Brazilian tropical dry forest. Apidologie 46, 631–643. doi: 10.1007/s13592-015-0354-1
Marques-Souza A. C. (2010). Ocorrência do pólen de Podocarpus sp. (Podocarpaceae) nas coletas de Frieseomelitta varia Lepeletier 1836 (Apidae: Meliponinae) em uma área de Manaus, AM, Brasil. Acta Bot. Bras. 24, 558–566. doi: 10.1590/S0102-33062010000200024
Martínez-Hernández E., Cuadriello-Aguilar J. I., Ramírez-Arriaga E., Medina-Camacho M., Sosa-Nájera M. S., Melchor-Sánchez J. E. (1994). Foraging of Nannotrigona testaceicornis, Trigona (Tetragonisca) angustula, Scaptotrigona mexicana and Plebeia sp. in the Tacaná region, Chiapas, Mexico. Grana 33, 205–217. doi: 10.1080/00173139409429001
Martínez-Hernández E., Cuadriello-Aguilar J. I., Téllez-Valdez O., Ramírez-Arriaga E., Sosa-Nájera M. S., Melchor-Sánchez J. E., et al. (1993). Atlas de las plantas y el polen utilizados por las cinco especies principales de abejas productoras de miel en la región del Tacaná, Chiapas, México (México: Universidad Nacional Autónoma de México).
Martins A. C., Melo G. A., Renner S. S. (2014). The corbiculate bees arose from New World oil-collecting bees: implications for the origin of pollen baskets. Mol. Phylogenet. Evol. 80, 88–94. doi: 10.1016/j.ympev.2014.07.003
Mateus S. (1998). Abundância relativa, fenologia e visita as flores pelos Apoidea do cerrado da Estação Ecológica de Jataí - Luiz Antônio - SP. Master thesis. Universidade de São Paulo, Ribeirão Preto.
Mateus S., Menezes C., Vollet-Neto A. (2013). “Leurotrigona muelleri, a pequena pérola entre as abelhas sem ferrão,” in Stingless Bees Process Honey and Pollen in Cerumen Pots. Eds. Vit P., Roubik D. W. (Universidad de Los Andes, Mérida), 1–8. Available at: http://www.saber.ula.ve/handle/123456789/37000.
Matuso T., Martínez-Ramos M., Bongers F., van der Sande M. T., Poorter L. (2021). Forest structure drives changes in light heterogeneity during tropical secondary forest succession. J. Ecol. 109, 2871–2884. doi: 10.1111/1365-2745.13680
Maués M. M., Couturier G. (2002). Floral biology and reproductive phenology of the camu-camu (Myrciaria dubia (H.B.K.) McVaugh, Myrtaceae) in the state of Pará, Brazil. Braz. J. Bot. 25, 441–448. doi: 10.1590/S0100-84042002012000008
Maués M. M., Sousa L. A., Miyanaga R. (2000). Insetos Polinizadores do Cupuaçuzeiro (Theobroma grandiflorum (Willd. Ex Sprengel) Schum - Sterculiaceae) no Estado do Pará, Brasil (Belém: Embrapa Amazônia Oriental).
Mayes D. M., Bhatta C. P., Shi D., Brown J. C., Smith D. R. (2019). Body size influences stingless bee (Hymenoptera: Apidae) communities across a range of deforestation levels in Rondônia, Brazil. J. Insect Sci. 19, 23. doi: 10.1093/jisesa/iez032
Mc Cabe S. I., Farina W. M. (2009). Odor information transfer in the stingless bee Melipona quadrifasciata: effect of in-hive experiences on classical conditioning of proboscis extension. J. Comp. Physiol. A 195, 113–122. doi: 10.1007/s00359-008-0391-6
Mc Cabe S. I., Farina W. M. (2010). Olfactory learning in the stingless bee Tetragonisca angustula (Hymenoptera, Apidae, Meliponini). J. Comp. Physiol. A 196, 481–490. doi: 10.1007/s00359-010-0536-2
Mendes F. N. (2007). Polinização, sistema reprodutivo e abelhas visitantes de duas espécies simpátricas de Byrsonima (Malpighiaceae) em área de cerrado no Maranhão, Brasil. Master thesis. Universidade Federal do Maranhão, São Luís.
Michener C. D. (2007). The Bees of the World, 2nd edition (Baltimore-MD: The John Hopkins University Press).
Misiewicz T. M., Kraichak E., Rasmussen C. (2014). Distance and habitat drive fine scale stingless bee (Hymenoptera: Apidae) community turnover across naturally heterogeneous forests in the western Amazon. Sociobiology 61, 407–414. doi: 10.13102/sociobiology.v61i4.407-414
Moguel P., Toledo V. M. (1999). Biodiversity conservation in traditional coffee systems of Mexico. Conserv. Biol. 13, 1–21. doi: 10.1046/j.1523-1739.1999.97153.x
Montoya M. A. M., Cruzatty L. C. G. (2014). Eficiencia Reproductiva y Receptividad Estigmática en Clones de Cacao (Theobroma cacao L.) en la Zona Central del Litoral Ecuatoriano (Quito: Ministerio de Agricultura, Ganadería, Acuacultura y Pesca). Available at: https://fliphtml5.com/ijia/fukf/basic (Accessed July 02, 2024).
Morán C. Q., Schettini G. N., Ochoa D. F., González A. B., Cantos I. S., Jazayeri S. M. (2021). Selection of self-compatible progenies of high productivity cocoa in the Ecuadorian coast. Cienc. Tecnol. 14, 11–21. doi: 10.18779/cyt.v14i1.448
Moreno E., Vit P., Aguilar I., Barth O. M. (2023). Melissopalynology of Coffea arabica honey produced by the stingless bee Tetragonisca angustula (Latreille 1811) from Alajuela, Costa Rica. AIMS Agric. Food 8, 804–831. doi: 10.3934/agrfood.2023043
Moure J. S. (1988). Uma nova espécie de Frieseomelitta do oeste da Amazônia (Hymenoptera, Apoidea). Acta Biologica Paranaense 17, 141–145.
Muto N. A., Leite R. O. S., Pereira D. S., Rogez H. L., Venturieri G. C. (2020). Impact of the introduction of stingless bee colonies (Scaptotrigona aff. postica) on the productivity of acai (Euterpe oleracea). Rev. Verde 15, 265–273. doi: 10.18378/rvads.v15i3.8404
Nair P. K. R. (2007). The coming of age of agroforestry. J. Sci. Food Agric. 87, 613–1619. doi: 10.1002/jsfa.2897
Nair P. K. R. (2011). Agroforestry systems and environmental quality: introduction. J. Environ. Qual. 40, 784–790. doi: 10.2134/jeq2011.0076
Nascimento E. S., Santos R. H. S. (2021). Visitantes florais do abricoteiro (Mammea americana L., Calophyllaceae) em Belém, PA. Doctoral thesis. Universidade Federal Rural da Amazônia, Belém.
Nery L. S., Takata J. T., Camargo B. B., Chaves A. M., Ferreira P. A., Boscolo D. (2018). Bee diversity responses to forest and open areas in heterogeneous Atlantic Forest. Sociobiology 65, 686–695. doi: 10.13102/sociobiology.v65i4.3472
Neto W. V. D. C., Maues M. M. (2008). Manutenção de uma Criação de Plebeia minima e Tetragonisca angustula (Apidae, Meliponini) para a Polinização do Cupuaçuzeiro (Theobroma grandiflorum) (Belém: Empresa Brasileira de Pesquisa Agropecuária Amazônia Oriental).
Neves E. L. (2001). Composição e visita às flores pelas abelhas eusociais (Hymenoptera: Apidae: Apinae) nas dunas interiores da margem esquerda do Médio Rio São Francisco, Barra, Bahia. Master thesis. Universidade Federal da Bahia, Salvador.
Nevill A. M., Teixeira L. V., Marques M. D., Waterhouse J. M. (2004). Using covariance to unravel the effects of metorological factors and daily and seasonal rhythms. Biol. Rhythm. Res. 35, 159–169. doi: 10.1080/09291010412331313313
Nogueira D. S. (2016). Sistemática de Scaura Schwarz 1938 (Hymenoptera: Apidae, Meliponini), com notas biológicas. Master thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Nogueira D. S., Oliveira F. F., Oliveira M. L. (2017). The real taxonomic identity of Trigona latitarsis Friese 1900, with notes on type specimens (Hymenoptera, Apidae). Zookeys 2, 113–130. doi: 10.3897/zookeys.713.11653
Nogueira D. S., Ribeiro C. F., Oliveira M. L. (2020). Redescription of Nogueirapis Moure 1953 with two new species from the Amazon Forest (Hymenoptera: Apidae: Meliponini). Zootaxa 4859, 138–150. doi: 10.11646/zootaxa.4859.1.6
Nogueira D. S., Rocha E. E. M., Félix J. A., Andrade M. A. P., Cortopassi-Laurino M., Alves R. M. O., et al. (2023). Notes on the biology of Scaura (Hymenoptera: Apidae: Meliponini). J. Apicult. Res. 62, 47–63. doi: 10.1080/00218839.2021.1940743
Nogueira-Neto P. (1970). A Criação de Abelhas Indígenas Sem Ferrão, 2nd edn (São Paulo: Editora Chácaras e Quintais).
Novais J. S., Absy M. L. (2013). Palynological examination of the pollen pots of native stingless bees from the Lower Amazon region in Pará, Brazil. Palynology 37, 218–230. doi: 10.1080/01916122.2013.787127
Novais J. S., Absy M. L. (2015). Melissopalynological records of honeys from Tetragonisca angustula (Latreille 1811) in the Lower Amazon, Brazil: pollen spectra and concentration. J. Apicult. Res. 54, 11–29. doi: 10.1080/00218839.2015.1041294
Novais J. S., Absy M. L., Santos F. A. R. (2013). Pollen grains in honeys produced by Tetragonisca angustula (Latreille 1811) (Hymenoptera: Apidae) in tropical semi-arid areas of north-eastern Brazil. Arthropod Plant Interact. 7, 619–632. doi: 10.1007/s11829-013-9276-x
Novais J. S., Absy M. L., Santos F. A. R. (2014). Pollen types collected by Tetragonisca angustula (Hymenoptera: Apidae) in dry vegetation in Northeastern Brazil. Eur. J. Entomol. 111, 25–34. doi: 10.14411/eje.2014.004
Novais J. S., Absy M. L., Santos F. A. R. (2015). Comparative pollen spectra of Tetragonisca angustula (Apidae, Meliponini) from the Lower Amazon (N Brazil) and caatinga (NE Brazil). Apidologie 46, 417–431. doi: 10.1007/s13592-014-0332-z
Obregón D., Rodríguez C. Á., Chamorro F. J., Nates-Parra G. (2013). “Botanical origin of pot-honey from Tetragonisca angustula Latreille in Colombia,” in Pot-Honey: A Legacy of Stingless Bees. Eds. Vit P., Pedro S. R. M., Roubik D. (Springer, New York), 337–346.
Oliveira F. F., Richers B. T. T., Silva J. R., Farias R. C., Matos T. A. L. (2013). Guia Ilustrado das Abelhas “Sem Ferrão” das Reservas Amanã e Mamirauá, Amazonas, Brasil (Hymenoptera, Apidae, Meliponini) (Tefé: Instituto de Desenvolvimento Sustentável Mamirauá).
Oliveira M. M. (2018). Insetos visitantes florais associados a um guaranazal (Paullinia cupana var. sorbilis (Mart.) Ducke), com notas sobre sua polinização. Doctoral thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Oliveira R. C., Menezes C., Soares A. E. E., Imperatriz-Fonseca V. L. (2013). Trap-nests for stingless bees (Hymenoptera, Meliponini). Apidologie 44, 29–37. doi: 10.1007/s13592-012-0152-y
Paz F. S., Pinto C. E., Brito R. M., Imperatriz-Fonseca V. L., Giannini T. C. (2021). Edible fruit plant species in the Amazon Forest rely mostly on bees and beetles as pollinators. J. Econ. Entomol. 114, 710–722. doi: 10.1093/jee/toaa284
Pedro S. R. M. (2014). The stingless bee fauna in Brazil (Hymenoptera: Apidae). Sociobiology 61, 348–354. doi: 10.13102/sociobiology.v61i4.348-354
Pedro S. R. M., Camargo J. M. F. (2009). Neotropical Meliponini: the genus Leurotrigona Moure two new species (Hymenoptera: Apidae, Apinae). Zootaxa 1983, 23–44. doi: 10.11646/zootaxa.1983.1
Pedro S. R. M. (1992). Sobre as abelhas (Hymenoptera – apoidea) em um ecossistema de cerrado (Cajuru, néctar estado de são paulo): Composição, fenologia e visita às flores (Ribeirão Preto: Universidade de São Paulo).
Pereboom J. J. M., Biesmeijer J. C. (2003). Thermal constraints for stingless bee foragers: the importance of body size and coloration. Oecologia 137, 42–50. doi: 10.1007/s00442-003-1324-2
Pimentel A. D. A. (2020). Recursos polínicos coletados por Melipona Illiger 1806 e Frieseomelitta Ihering 1912 (Apidae-Apinae-Meliponini) em matas de igapós do Rio Negro, Amazonas, Brasil. Master thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Purwanto H., Soesilohadi R. C. H., Trianto M. (2022). Stingless bees from meliponiculture in South Kalimantan, Indonesia. Biodiversitas 23, 1254–1266. doi: 10.13057/biodiv/d230309
Quezada-Euán J. J. G. (2018). Stingless Bees of Mexico–The Biology, Management and Conservation of an Ancient Heritage (Cham: Springer). doi: 10.1007/978-3-319-77785-6
Ramalho M. (2004). Stingless bees and mass flowering trees in the canopy of Atlantic Forest: a tight relationship. Acta Bot. Bras. 18, 37–47. doi: 10.1590/S0102-33062004000100005
Ramírez-Arriaga E., Pacheco-Palomo K. G., Moguel-Ordoñez Y. B., Moreno R. Z. G., Godínez-García L. M. (2018). “Angiosperm resources for stingless bees (Apidae, Meliponini): a pot-pollen melittopalynological study in the Gulf of Mexico,” in Pot-Pollen in Stingless Bee Melittology. Eds. Vit P., Pedro S. R. M., Roubik D. (Springer, Cham), 111–130.
Ramos D. L., Menezes C., Bustamante M. M. C., Pires C. S. S. (2024). Effect of ultraviolet and green radiation and temperature on flight activity and foraging of three tropical stingless bees. J. Apicult. Res., 1–14. doi: 10.1080/00218839.2023.2293589
Ramsey M. (1995). Ant pollination of the perennial herb Blandfordia grandiflora (Liliaceae). Oikos 74, 265–272. doi: 10.2307/3545656
Rasmussen C., Delgado C. (2019). Abejas Sin Aguijón (Apidae: Meliponini) en Loreto, Perú (Iquitos: Instituto de Investigaciones de la Amazonia Peruana).
Rasmussen C., Gonzalez V. H. (2009). Abejas sin aguijón del Cerro Escalera, San Martín, Perú (Hymenoptera: Apidae: Meliponini). Sist. Agroeco. Mod. Biomatematic. 2, 26–32.
Rasmussen C., Gonzalez V. H. (2017). The neotropical stingless bee genus Nannotrigona Cockerell (Hymenoptera: Apidae: Meliponini): An illustrated key, notes on the types, and designation of lectotypes. Zootaxa 4299, 191–220. doi: 10.11646/zootaxa.4299.2.2
Raudales D. P. (2022). Comunidades de abejas (Hymenoptera: Apidae) en ambientes contrastantes de um paisaje de la Chiquitanía, Santa Cruz, Bolívia. [master’s tesis]. Centro Agronómico Tropical de Investigación y Enseñanza, Turrialba.
Rech A. R., Absy M. L. (2011a). Pollen sources used by species of Meliponini (Hymenoptera: Apidae) along the Rio Negro channel in Amazonas, Brazil. Grana 50, 150–161. doi: 10.1080/00173134.2011.579621
Rech A. R., Absy M. L. (2011b). Pollen storages in nests of bees of the genera Partamona, Scaura and Trigona (Hymenoptera, Apidae). Rev. Bras. Entomol. 55, 361–372. doi: 10.1590/S0085-56262011005000026
Ribeiro M. F. (2008). Manejo de uruçu do chão (Melipona quinquefasciata) no interior do Ceará e Pernambuco (Mensagem Doce). Available at: https://www.apacame.org.br/mensagemdoce/95/artigo.htm (Accessed April 12, 2024).
Ribeiro M. F., Rodrigues F., Lima C. B. S., Braga J. R. (2012). “Atividade externa da abelha branca (Frieseomelitta doederleini) em período seco e chuvoso em Petrolina, PE,” in Anais do VII. Congresso Nordestino de Produção Animal (Sociedade Nordestina de Produção Animal, Maceió).
Ricketts T. H., Regetz J., Steffan-Dewenter I., Cunningham S. A., Kremen C., Bogdanski A., et al. (2008). Landscape effects on crop pollination services: are there general patterns? Ecol. Lett. 11, 499–515. doi: 10.1111/j.1461-0248.2008.01157.x
Roselino A. C., Hrncir M. (2012). Repeated unrewarded scent exposure influences the food choice of stingless bee foragers, Melipona scutellaris. Anim. Behav. 83, 755–762. doi: 10.1016/j.anbehav.2011.12.025
Roubik D. W. (1982). Seasonality in colony food storage, brood production and adult survivorship: studies of Melipona in tropical forest (Hymenoptera: Apidae). J. Kansas Entomol. Soc 55, 789–800.
Roubik D. W. (2023). Stingless bee (Apidae: Apinae: Meliponini) ecology. Annu. Rev. Entomol. 68, 231–256. doi: 10.1146/annurev-ento-120120-103938
Sabino W., Costa L., Andrade T., Teixeira J., Araújo G., Acosta A. L., et al. (2022). Status and trends of pollination services in Amazon agroforestry systems. Agric. Ecosyst. Environ. 335, 108012. doi: 10.1016/j.agee.2022.108012
Sabino W., Ferreira O. R. P., Antonini Y. (2011). A apifauna do Parque Estadual do Itacolomi, Ouro Preto, Minas Gerais. MG. Biota 3, 19–31.
Santos C. F., Absy M. L. (2010). Polinizadores de Bertholletia excelsa (Lecythidales: Lecythidaceae): interações com abelhas sem ferrão (Apidae: Meliponini) e nicho trófico. Neotrop. Entomol. 39, 854–861. doi: 10.1590/S1519-566X2010000600002
Santos D. L. (2016). Biologia reprodutiva de duas espécies de Palicourea Aubl. (Rubiaceae) em floresta de terra firme na Amazônia Central. Master thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Saravia-Nava A., Niemeyer H. M., Pinto C. F. (2018). Pollen types used by the native stingless bee, Tetragonisca angustula (Latreille), in an Amazon-Chiquitano transitional forest of Bolivia. Neotrop. Entomol. 47, 798–807. doi: 10.1007/s13744-018-0612-9
Shivanna K. R. (2015). “Management of pollination services to enhance crop productivity,” in Plant Biology and Biotechnology. Eds. Bahadur B., Venkat Rajam M., Sahijram L., Krishnamurthy K. (Springer, New Delhi), 697–711.
Silveira M. S. (2006). Fauna de abelhas (Hymenoptera, Apoidea Apiformes) e recursos tróficos utilizados em áreas urbanas e entorno da cidade de João Pessoa. Master thesis. Universidade Federal da Paraiba, João Pessoa.
Soria S. J. (1975). O papel das abelhas sem ferrão (Meliponinae) na polinização do cacaueiro na América Tropical. Rev. Theobroma 5, 12–20.
Souza J. M. T. (2013). Redes de polinização em áreas restauradas de Floresta Atlântica do Sul do Brasil. Doctoral thesis. Universidade Federal do Paraná, Curitiba.
Souza J. V., Souza E. A., Guimarães-Brasil M. O., Brasil F. (2017). “Atividade de voo da abelha sem ferrão Frieseomelitta doederleini (Apidae: Meliponini) em uma área de domínio da Caatinga,” in Educação Ambiental: Biomas, Paisagens e o Saber Ambiental. Ed. Seabra G. (Barlavento, Ituiutaba), 24–36.
Souza R. R., Pimentel A. D. A., Nogueira L. L., Abreu V. H. R., Novais J. S. (2023). Palynoflora exploited by Frieseomelitta longipes (Smith 1854) (Apinae: Meliponini) in protected areas from the Brazilian Amazon basin. J. Apic. Res. 62, 705–720. doi: 10.1080/00218839.2021.1889824
Steiner J., Zillikens A., Kamke R., Feja E. P., Falkenberg D. B. (2010). Bees and melittophilous plants of secondary Atlantic Forest habitats at Santa Catarina Island, Southern Brazil. Oecol. Aust. 14, 16–39. doi: 10.4257/oeco.2010.1401.01
Streinzer M., Huber W., Spaethe J. (2016). Body size limits dim−light foraging activity in stingless bees (Apidae: Meliponini). J. Comp. Physiol. A 202, 643–655. doi: 10.1007/s00359-016-1118-8
Suri T., Basu S. (2022). Heat resistant chocolate development for subtropical and tropical climates: a review. Crit. Rev. Food Sci. Nutr. 62, 5603–5622. doi: 10.1080/10408398.2021.1888690
Talledo B. G., Zambrano A. B., Cruzatty L. G., Gavilanes F. Z. (2019). Morphology, viability, and longevity of pollen of National Type and Trinitarian (CCN-51) clones of cocoa (Theobroma cacao L.) on the Coast of Ecuador. Braz. J. Bot. 42, 441–448. doi: 10.1007/s40415-019-00541-2
Teixeira A. F. R., Oliveira F. F., Viana B. F. (2007). Utilization of floral resources by bees of the genus Frieseomelitta von Ihering (Hymenoptera: Apidae). Neotrop. Entomol. 36, 675–684. doi: 10.1590/S1519-566X2007000500007
Teixeira L. V., Campos F. N. M. (2005). Início da atividade de vôo em abelhas sem ferrão (Hymenoptera, Apidae): influência do tamanho da abelha e da temperatura ambiente. Rev. Bras. Zoociências 7, 195–220.
Toledo-Hernández M., Wanger T. C., Tscharntke T. (2017). Neglected pollinators: Can enhanced pollination services improve cocoa yields. A review? Agric. Ecosyst. Environ. 247, 137–148. doi: 10.1016/j.agee.2017.05.021
Toledo-Hernández M., Tscharntke T., Tjoa A., Anshary A., Cyio B., Wanger T. C. (2020). Hand pollination, not pesticides or fertilizers, increases cocoa yields and farmer income. Agric. Ecosyst. Environ. 304, 107160. doi: 10.1016/j.agee.2020.107160
Toledo-Hernández M., Tscharntke T., Tjoa A., Anshary A., Cyio B., Wanger T. C. (2021). Landscape and farm-level management for conservation of potential pollinators in Indonesian cocoa agroforests. Biol. Conserv. 257, 109106. doi: 10.1016/j.biocon.2021.109106
Toledo-Hernández M., Tscharntke T., Giannini T. C., Solé M., Wanger T. C. (2023). Hand pollination under shade trees triples cocoa yield in Brazil’s agroforests. Agric. Ecosyst. Environ. 355, 108612. doi: 10.1016/j.agee.2023.108612
Tscharntke T., Clough Y., Bhagwat S. A., Buchori D., Faust H., Hertel D., et al. (2011). Multifunctional shade-tree management in tropical agroforestry landscapes – a review. J. Appl. Ecol. 48, 619–629. doi: 10.1111/j.1365-2664.2010.01939.x
Tscharntke T., Ocampo-Ariza C., Vansynghel J., Ivañez-Ballesteros B., Aycart P., Rodriguez L., et al. (2023). Socio-ecological benefits of fine-flavor cacao in its center of origin. Conserv. Lett. 16, e12936. doi: 10.1111/conl.12936
Ulyshen M., Urban-Mead K. R., Dorey J. B., Rivers J. W. (2023). Forests are critically important to global pollinator diversity and enhance pollination in adjacent crops. Biol. Rev. 98, 1118–1141. doi: 10.1111/brv.12947
Vaast P., Somarriba E. (2014). Trade-offs between crop intensification and ecosystem services: the role of agroforestry in cocoa cultivation. Agroforest. Syst. 88, 947–956. doi: 10.1007/s10457-014-9762-x
Vansynghel J., Ocampo-Ariza C., Maas B., Martin E. A., Thomas E., Hanf-Dressler T., et al. (2022). Cacao flower visitation: Low pollen deposition, low fruit set and dominance of herbivores. Ecol. Solutions Evid. 3, e12140. doi: 10.1002/2688-8319.12140
Veiga J. C., Menezes C., Venturieri G. C., Contrera F. A. L. (2013). The bigger, the smaller: relationship between body size and food stores in the stingless bee Melipona flavolineata. Apidologie 44, 324–333. doi: 10.1007/s13592-012-0183-4
Velez-Ruiz R. I., Gonzalez V. H., Engel M. S. (2013). Observations on the urban ecology of the Neotropical stingless bee Tetragonisca angustula (Hymenoptera: Apidae: Meliponini). J. Melittology 15, 1–8. doi: 10.17161/jom.v0i15.4528
Venturieri G. C., Alves D. A., Villas-Bôas J. K., Carvalho C. A. L. D., Menezes C., Vollet-Neto A., et al. (2012). Meliponicultura no Brasil: Situação Atual e Perspectivas Futuras para o Uso na Polinização Agrícola. Polinizadores no Brasil: Contribuição e Perspectivas para a Biodiversidade, Uso Sustentável, Conservação e Serviços Ambientais (São Paulo: Editora da Universidade de São Paulo).
Viana B. F., Coutinho J. G. E., Garibaldi L. A., Gastagnino L. B., Gramacho K. P., Silva F. O. (2014). Stingless bees further improve apple pollination and production. J. Pollinat. Ecol. 14, 261–269. doi: 10.26786/1920-7603(2014)26
Vieira W. M. (2021). Atividade de voo da abelha jataí (Tetragonisca angustula) durante o ano, no Setor de Meliponiciltura da UFRPE no Recife, PE. Bachelor thesis. Universidade Federal Rural de Pernambuco, Recife.
Wanger T. C., Dennig F., Toledo-Hernández M., Tscharntke T., Lambin E. F. (2021). Cocoa pollination, biodiversity-friendly production, and the global market. arXiv:2112.02877[econ.GN]. doi: 10.48550/arXiv.2112.02877
Westerkamp C. (1996). Pollen in bee-flower relations some considerations on melittophily. Bot. Acta 109, 325–332. doi: 10.1111/j.1438-8677.1996.tb00580.x
Winder J. A. (1977). Field observations on Ceratopogonidae and other Diptera: Nematocera associated with cocoa flowers in Brazil. Bull. Entomol. Res. 67, 57–63. doi: 10.1017/S0007485300010890
Wolcott K. A., Stanley E. L., Gutierrez O. A., Wuchty S., Whitlock B. A. (2023). 3D pollination biology using micro-computed tomography and geometric morphometrics in Theobroma cacao. Appl. Plant Sci. 11, e11549. doi: 10.1002/aps3.11549
Wongnaa C. A., Apike I. A., Babu S., Awunyo-Vitor D., Kyei A. B. (2021). The impact of adoption of artificial pollination technology in cocoa production: Evidence from Ghana. J. Agricult. Food Res. 6, 100208. doi: 10.1016/j.jafr.2021.100208
Yoda K. (1974). Three-dimensional distribution in a tropical rain forest of west Malaysia. Japan. J. Ecol. 24, 247–254.
Young A. M. (1981). The ineffectiveness of the stingless bee Trigona jaty (Hymenoptera: Apidae: Meliponinae) as a pollinator of cocoa (Theobroma cacao L.). J. Appl. Ecol. 18, 149–155. doi: 10.2307/2402485
Young A. M. (1982). Effects of shade cover and availability of midge breeding sites on pollinating midge populations and fruit set in two cocoa farms. J. Appl. Ecol. 19, 47–63. doi: 10.2307/2402990
Young A. M. (1983). Seasonal differences in abundance and distribution of cocoa-pollinating midges in relation to flowering and fruit set between shaded and sunny habitats of the La Lola cocoa farm in Costa Rica. J. Appl. Ecol. 20, 801–831. doi: 10.2307/2403127
Young A. M. (1985a). Studies of cecidomyiid midges (Diptera: Cecidomyiidae) as cocoa pollinators (Theobroma cacao L.) in Central America. Proc. Entomol. Soc Wash. 87, 49–79.
Young A. M. (1985b). Pollen-collecting by stingless bees on cacao flowers. Experientia 41, 760–762. doi: 10.1007/BF02012584
Young A. M., Severson D. W. (1994). Comparative analysis of steam distilled floral oils of cacao cultivars (Theobroma cacao L., Sterculiaceae) and attraction of flying insects: Implications for a Theobroma pollination syndrome. J. Chem. Ecol. 20, 2687–2703. doi: 10.1007/BF02036201
Young A. M., Schaller M., Strand M. (1984). Floral nectaries and trichomes in relation to pollination in some species of Theobroma and Terrania (Sterculiaceae). Am. J. Bot. 71, 466–480. doi: 10.1002/j.1537-2197.1984.tb12532.x
Young A. M., Erickson E. H., Strand M. A., Erickson B. J. (1987). Pollination biology of Theobroma and Herrania (Sterculiaceae)—I. Floral Biol. Insect Sci. Applic. 8, 151–164. doi: 10.1017/S1742758400007153
Young A. M., Erickson B. J., Erickson E. H. (1989). Pollination biology of Theobroma and Herrania (Sterculiaceae)—III. Steam-distilled floral oils of Theobroma species as attractants to flying insects in a Costa Rican cocoa plantation. Insect Sci. Appl. 10, 93–98. doi: 10.1017/S1742758400003404
Keywords: Meliponini, Amazon rainforest, bee morphology, cocoa, directed pollination
Citation: Maia-Silva C, Hrncir M, Giannini TC, Toledo-Hernández M and Imperatriz-Fonseca VL (2024) Small Amazonian stingless bees: an opportunity for targeted cocoa pollination. Front. Bee Sci. 2:1357811. doi: 10.3389/frbee.2024.1357811
Received: 18 December 2023; Accepted: 15 July 2024;
Published: 02 August 2024.
Edited by:
Fabrice Requier, Institut de Recherche Pour le Développement (IRD), FranceReviewed by:
Isabelle Merle, Institut de Recherche Pour le Développement (IRD), FranceSandra V. Rojas-Nossa, University of Vigo, Spain
Copyright © 2024 Maia-Silva, Hrncir, Giannini, Toledo-Hernández and Imperatriz-Fonseca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Camila Maia-Silva, Y2FtaWxhbWFpYUBhbHVtbmkudXNwLmJy