 David Taillis1
David Taillis1 Aleksandra Burdziej1
Aleksandra Burdziej1 Charlyne Bautista1
Charlyne Bautista1 Ferdinand W. M. Haag2
Ferdinand W. M. Haag2 Michael Lebert2,3*
Michael Lebert2,3* Stéphanie Cluzet1*
Stéphanie Cluzet1*- 1Univ. Bordeaux, Bordeaux INP, INRAE, OENO, UMR 1366, ISVV, Villenave d’Ornon, France
- 2Space Cargo Unlimited SAS, Bordeaux, France
- 3Friedrich-Alexander-Universität, Department Biology, Cell Biology, Erlangen, Germany
The control of plant diseases represents a significant challenge in agriculture, particularly with the current emphasis on reducing pesticide use, necessitating alternative approaches. Among them, resistant plant breeding programs offer a promising alternative. Considering the stressful conditions in space, space conditions emerge as an innovative method for advancing crop improvement. This study explored the unique potential of the International Space Station (ISS) environment to induce beneficial modifications in grapevine, resulting in plants with reduced susceptibility to diseases. Canes of two Vitis vinifera L. cultivars, Cabernet Sauvignon (CS) and Merlot (Me), have undergone a 10-month space journey on-board the ISS, while others stayed on Earth (Ground Control, GC). We conducted a 2-year analysis of the response to downy mildew (Plasmopara viticola) of plants developed from ISS and GC canes, called Mother plants. The disease development of progenies produced from Merlot Mother plants was also evaluated. A higher proportion of individuals with low-susceptibility was noted in the ISS Mother group compared to the controls for both cultivars. This trend was also observed for the Merlot progenies. Finally, 11% of Merlot individuals (Mother and progenies, and for the 2 years of study) were distributed in the low susceptibility class. This study suggests that grapevine canes exposure to ISS conditions resulted in an enhanced percentage of plants showing reduction of susceptibility to downy mildew.
1 Introduction
Today, agriculture faces significant difficulties, whether it concerns climate change (Malhi et al., 2021) or the increased concerns about environmental and health issues associated with chemical pesticides (Rani et al., 2021). The grapevine’s perennial nature makes it one of the most affected plants by both challenges. Various pathogens continuously threaten grapes production in terms of quantity and quality. The highly destructive disease, downy mildew, caused by the oomycete Plasmopara viticola, stands out as a significant challenge (Gessler et al., 2011; Bove et al., 2020). Since viticulture is significant in several countries’ economies, there is a growing imperative to explore sustainable alternatives for managing downy mildew. In this study, we chose to focus on the two following cultivars, Vitis vinifera cv. Cabernet Sauvignon and Merlot, being in the top 5 out of the world’s 10,000 known grapevine varieties (CS and Me cover an area of 4% and 3% of the world’s vineyards (341,000 and 266,000 ha in 2015), respectively (OIV, 2017)). These two varieties are widely popular and well-established red wine varietals but are vulnerable to several diseases, mainly downy mildew. Fungicides are regularly applied to avoid quantitative and qualitative losses due to this disease (Rani et al., 2021). However, the intensive use of such chemicals causes human health risks and a negative environmental impact, and thus, it is becoming more and more restrictive.
Cultural practices, resistant grapevine varieties, plant defense stimulators, biopesticides, and biological control agents present promising avenues for sustainable disease management (Pertot et al., 2017). Considering resistant grapevines, most of the breeding programs have been developed to confer protection against downy mildew and powdery mildew, resulting in the availability of new varieties offering durable resistance to these two diseases but often with susceptibility to other major diseases, such as Black rot and gray mold (Foria et al., 2019). Moreover, wine from resistant cultivars sometimes does not reach expected quality standards (Pedneault and Provost, 2016; Bavaresco, 2019; Bavaresco and Squeri, 2022).
Those breeding techniques take several decades to produce a useable grapevine variety (Töpfer and Trapp, 2022). Other ways to obtain resistant plants are to use techniques such as genome editing (Wan et al., 2020) or cisgenesis-generated resistant plants. However, technical issues and legal regulations hamper these approaches (Pirrello et al., 2023). In addition, environmental and chemical stressors can drive the evolution of plants by inducing genetic and epigenetic modifications that could modify plant characteristics. Thus, anatomical and physiological changes could be induced, such as vascular modifications and metabolism changes, in particular at the level of specialized metabolites (Salem-Fnayou et al., 2011; Ferrandino et al., 2023). These adaptive responses enable plants to thrive in diverse and often harsh environments, highlighting the dynamic interplay between environmental stress, genetic variation, and epigenetic regulation in the evolutionary trajectory of plant species. Such environmental conditions can shape the selection and allow the obtaining of plants with traits of agronomical interest (Ahloowalia and Maluszynski, 2001). Evolutionary engineering consists of an alternative approach to conventional modern techniques, such as genetic or metabolic engineering, that mimics the natural evolution process to develop or obtain desired traits, as for instance: enhancement of tolerance of organisms to stress, increase of cell performance, improvement of the production of compounds of interest (for review, see Prasad et al., 2020).
Because of weightlessness and ionizing radiation, space is one of the harshest environments for biological materials. Since the 80s, space crop breeding programs have been carried out on the basis that such a stressful environment, never experienced on Earth for a terrestrial plant, is associated with genetic and phenotypic shifts and then could lead to accelerated evolution (Mohanta et al., 2021). Indeed, space radiation and microgravity profoundly affect plant cells, influencing their fundamental biological processes (Prasad et al., 2020). Space radiation, comprising ionizing particles from sun events and cosmic rays, can impact DNA integrity. Radiations can induce DNA double-stranded break that lead to, if not repaired properly, chromosomal aberrations, stable and inheritable genetic alternations, and epigenetic modifications (Belli and Tabocchini, 2020; Furukawa et al., 2020; Mohanta et al., 2021; De Micco et al., 2023). Microgravity or zero gravity has been shown to impact gene expression and metabolism, thus cellular functions and physiological development (Manzano et al., 2022; De Micco et al., 2023). For example, studies conducted on micro or altered gravity highlight an affected expression of genes involved in mechanisms like cell wall remodeling, photosynthesis, and defense reactions, such as oxidative stress, in Arabidopsis thaliana (Kimbrough et al., 2004; Manzano et al., 2022).
In this study, space, a unique environment that combines many stressors never experienced by organisms on Earth, was used as a potential tool for obtaining grapevines with modified traits, such as plants with varying susceptibility levels to downy mildew. In this experiment, grapevine canes, the organs used for vegetative propagation, were exposed to 10 months of storage inside the International Space Station (ISS). After their return to Earth, leaves of plants developed from these grapevine canes were subjected to P. viticola inoculation.
2 Materials and methods
2.1 Plant conditions
2.1.1 Plant material
Vitis vinifera L. cv. Cabernet Sauvignon (CS, clone 0169) and cv. Merlot (Me, clone 0349) canes from Mercier Nursery (La Lande de Fronsac, France) were cut in 2019 and kept in a controlled environment at 4°C ± 2°C, in the dark and high humidity (75%–80%) until their use. One week before the launch, canes were cut into 4 cm-long sections containing one bud (Figure 1A. They were surface sterilized as follows: 5 min à 54°C (hot water treatment), 1 min in ethanol:H2O (70:30, v/v), water rinsed 2 times, 5 min in hypochlorite sodium:H2O (50:50, v/v) in the presence of Tween 20 (0.1%) and water rinsed 3 times. Then, canes were placed by pair (buds upside down) in each alveolus of specific containers. These containers, designed especially for this experimentation, consisted of plastic boxes with 16 alveoli each (Figures 1B, C). A cap with holes recovered by a membrane in ePTFE closed each alveolus. This membrane allowed gas exchange during the cane’s storage. Each cane was meticulously drilled with two holes, allowing for a secure fixation during travel as the caps presented two inward peaks (Figures 1C, D). A total of 10 containers were prepared for ISS storage: 5 containers for Merlot (160 canes), 5 for Cabernet Sauvignon (160 canes), and 5 containers for the Ground Control condition (GC, Earth): 2.5 containers for Merlot (80 canes) and 2.5 for Cabernet Sauvignon (80 canes). An exploded drawing of a container is presented in Figure 1E.
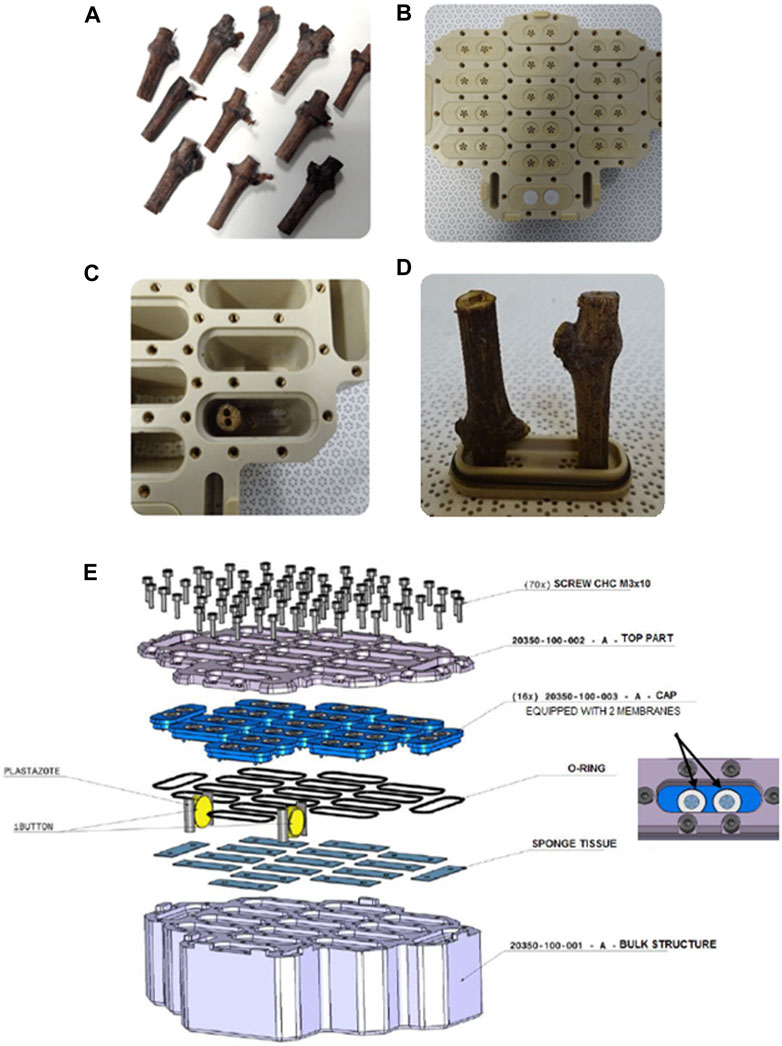
Figure 1. Grapevine cane material and container used (A) Grapevine canes sections of 4 cm in length with one bud. (B) Detailed view of a canes container. (C) Alveolus in which a cane has been placed, (D) Close-up of mounted canes and (E) Exploded drawing of flight hardware.
2.1.2 ISS and Ground Control storage conditions
A total of 10 containers (160 canes of Merlot and 160 canes of CS) were transported aboard a space vessel to the International Space Station (ISS), initiating a 10-month mission spanning from 3 March 2020, to 12 January 2021. In the ISS, the containers were stored inside the Columbus Laboratory at specific temperature and humidity conditions (4°C ± 2°C, 75%–80%), in addition to darkness. Active and passive radiation detectors monitored the radiation environment inside the Columbus Laboratory at fixed locations (DOSTEL-2). The recorded parameters included the absorbed dose rate (D) and the dose equivalent rate (H), measuring 361 µGy/day and 775 µSv/day, respectively. These values amounted to 113.1 mGy and 242.5 mSv throughout the space mission (Table 1). Comprehensive information about all measurements is available in Supplementary Figure S1. A total of 5 containers (80 canes of Merlot and 80 canes of CS) were kept on Earth and stored under conditions similar to those of ISS, such as temperature, humidity, and darkness in a fridge of the ISVV building (Villenave d’Ornon, France).

Table 1. Radiation measurement (dose rate (D) and dose equivalent rate (H)) realized in the Columbus laboratory of the ISS during the mission flight.
2.1.3 Growing condition of plants from canes stored in ISS and on earth
At the end of the storage period, all canes (ISS and Earth stored canes), except 6 canes for each cultivar and storage place condition (planned to be used for further microscopy analyses), were transplanted in pots filled with Substrat 5 perlite, Ref. 446 (Klasmann-Deilmann, France) and placed in greenhouse under the following controlled conditions: 25/20°C day/night air temperature, 75% relative humidity and a 16 h photoperiod (350 μmol/m2/s). A phenotypic monitoring (plant growth and phenological development) was carried out and no difference between ISS and GC plants was observed (Supplementary Figure S2). In October 2021 (10 months after the cane plantation date), the plants developed from canes originally stored in ISS or Earth (Ground Control, GC), called Mother plants, were placed under a tunnel. The use of tunnels was chosen in order to be as close as possible to the natural environment conditions since temperature, light, and humidity cannot be controlled and only protection towards rain was present. Such conditions prevented the development of natural downy mildew, allowing us to test the plants towards this disease in laboratory in a controlled way (e.g., strain, sporangia number). Mother plants were continuously fertilized with a solution of Peters Professional Soluble Plant Food 20–20–20 General Purpose (Scotts company) at 1 g/L. Each plant constituted a specific individual.
2.1.4 Production of progenies and growing condition of these progenies
In January 2022 and January 2023, canes were harvested from each Mother plant. Canes of 10 cm in length with one bud were generated and transplanted in pots filled with Substrat 5 perlite, Ref. 446 (Klasmann-Deilmann, France). Then, they were placed in a greenhouse under controlled conditions: 25/20°C day/night air temperature, 75% relative humidity, and a 16 h photoperiod (350 μmol/m2/s). When these foliar cuttings reached the 6-leaf stage, they were used for downy mildew experiments.
2.2 Downy mildew material and bioassays
The bioassays of downy mildew susceptibility level in laboratory conditions were conducted with the P. viticola isolate ANN-01 collected in 2015 in a vineyard of V. vinifera L. cv. Ugni-blanc in Charente (France). Plasmopara viticola was maintained as previously described (Corio-Costet et al., 2011).
We collected the fourth leaf below the apex to analyze the susceptibility level of Merlot (Me) and Cabernet Sauvignon (CS) plants to P. viticola. Leaves from Mother plants and their respective progenies (only for Merlot) developed from canes stored in ISS conditions (ISS) or Earth (Ground Control, GC) were considered. At least three foliar disks (25 mm wide) were produced from each leaf and placed on the abaxial side upwards into Petri dishes containing humidified Whatman paper. Each plant was tested at least twice, generally three times, for most plants during a growing season. Sporangia from 7 days old P. viticola infected leaves were collected and suspended in distilled water, and the concentration of sporangia was adjusted to obtain a suspension at 5,000 sporangia/mL. Each disk was inoculated with 3 drops of 15 μL of the sporangia suspension and kept in the dark at room temperature one night before vacuuming the drops. Then, Petri dishes were incubated at 25°C with a photoperiod of 16/8 h day/night (Corio-Costet et al., 2011). After 6–7 days of incubation, the estimation of the sporulation density was performed by visual scoring. Three groups were created based on the level of sporulation on leaves, using a descriptor-like scale adapted from the OIV descriptor 452–1 as follows (Supplementary Figure S3): low-susceptibility (0%–35%), moderate-susceptibility (35.1%–60%), and high-susceptibility (60.1%–100%) (Bove et al., 2020; Štambuk et al., 2021).
2.3 Statistical analyses
Statistical analyses were conducted using R software version 4.3.1 and RStudio version 2023.06.2 + 561. To assess the significant differences in means of sporulation level between ISS and GC plants, according to the normality of the data, a Welch t-test was used, and this was done for each year (2022 and 2023) and for each cultivar (Cabernet Sauvignon and Merlot). Moreover, we compared each GC and ISS individual to the total population of individuals (all ISS and GC plants) to identify individuals that differed from the mean of the total population using a Mann-Whitney U test as the distribution of data was not normal. We then classified individuals according to their mean sporulation level in three classes (0%–35%, 35.1%–60%, and 60.1%–100%) and the significance of the difference. The analysis was carried out for each cultivar and year, for Mother plants and progenies separately or with all the 2 years’ data combined. The percentage of individuals belonging to a given category allows for a comparison and discrimination between plants originating from canes stored in GC or ISS conditions.
3 Results
We analyzed the level of susceptibility to downy mildew on the leaves of CS and Me plants that have been stored in ISS or not (GC) for Mother’s plants and their progenies during the years 2022 and 2023. As it was impossible to test them simultaneously due to space and staff time, we performed our assays with plant batches. In 2022, we tested 49 Me ISS, 54 CS ISS, 30 Me GC, and 36 CS GC for the Mother plants and 40 Me ISS and 20 Me GC for progenies, meaning 229 different plants. As repetitions were done, at least 2 were done for Mother plants and 4 or 5 for progenies, meaning a total of 338 assays for Mother plants and 240 for progenies plants; thus, 578 plants were tested. As we did not perform the experimentation at a similar time for all plants (batches), we had to consider the potential occurrence of experimental bias due to, for instance, changing environmental conditions under a tunnel. Thus, to smooth out the variability of the values of downy mildew development among the different batches and years, we chose to apply a correction factor. To calculate this correction factor, a reference value for GC plants was required. Based on our data and the literature, the reference value of sporulation mean of GC plants was set at 50%. Indeed, as example, for Merlot, several authors reported a disease severity around 50% (Calonnec et al., 2013; Bove et al., 2020). To proceed, the sporulation mean of GC plants was calculated for each batch and we determined the correction factor by dividing the reference value (50%) to this value. Then, using this determined correction factor, we transformed the sporulation values of all plants (GC and ISS).
3.1 Sporulation means
Firstly, we considered the mean of downy mildew sporulation, expressed in percentage, for all ISS Mother plants compared to all GC Mother plants for Me and CS in 2022 and 2023. Each year’s value was the mean of two or three bioassays. Based on these 2 years of analysis, we noticed a different profile depending on the year considered. For 2022, no significant differences can be put in evidence between the mean of sporulation of ISS plants and one of GC plants, both for CS and Me. However, in 2023, the levels of P. viticola sporulation on Mother CS and Me ISS plants were significantly lower, with 42% and 41% of mean sporulation, respectively, compared to the GC plants at 50% (Table 2).
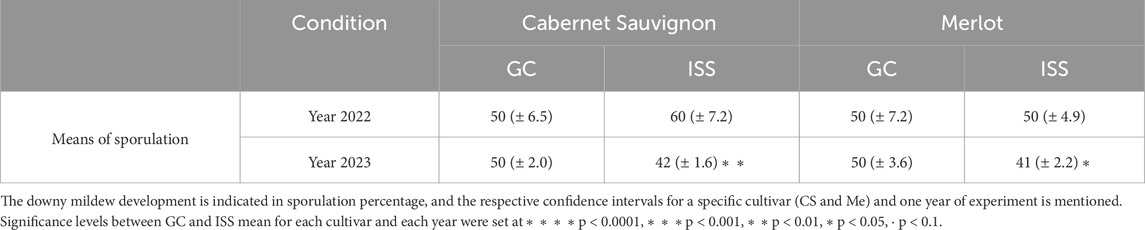
Table 2. Mean of P. viticola sporulation of all ISS (International Space Station) Mother plants compared to all GC (Ground Control) Mother plants.
Then, we considered the mean of downy mildew sporulation for Merlot ISS progenies plants compared to Merlot GC progenies plants for 2022 and 2023. Only Merlot progenies individuals were studied as the experimentation conditions (greenhouse place, personal staff dedicated to these experimentations) did not allow us to evaluate CS plants. We noted a similar profile in both years of experimentation, observing a reduction in downy mildew sporulation on ISS progeny plants, as in 2023 for the Mother plants. Indeed, the levels of downy mildew sporulation on progenies ISS plants were significantly lower with 39.7% and 42% of mean sporulation, respectively, in 2022 and 2023, in comparison to the one of GC plants at 50% (Table 3).

Table 3. Mean of P. viticola sporulation of Merlot ISS (International Space Station) progenies plants compared to Merlot GC (Ground Control) progenies plants.
3.2 Frequency distribution of Merlot and Cabernet Sauvignon Mother individuals
For each Mother individual, according to its respective mean of sporulation level, we classified this plant into one of the three following classes adapted from the OIV descriptor 452–1: low-susceptibility (0%–35%), moderate-susceptibility (35.1%–60%), and high-susceptibility (60.1%–100%). This simple descriptor-like scale that divides each individual inside one of these three classes allowed us to inform about the frequency of distribution of GC and ISS Mother individuals for 2022 and 2023 and the two varieties (CS and Me).
To illustrate the distribution of Merlot and CS plants according to their downy mildew sporulation levels, we made a density plots analysis (Figure 2). We noted, regardless of the year (2022 and 2023), the cultivar (CS and Me), and the condition (ISS and GC) considered, a density distribution that tended to be center around medium values of sporulation, between or relatively close to the two red lines that represent threshold of low and high susceptibility, respectively 35% and 60% (Figure 2). In particular, in 2023, the density distribution of Merlot and CS plants, for ISS and GC conditions, exhibited a Gaussian-like curve shape. This results was confirmed by a frequency analysis in which a high proportion of individuals falled into the class of moderately susceptible plants with between half and two-thirds of frequency distribution (Supplementary Table S1). The other two classes (low and high susceptible plants) had similar proportions of individuals with frequency distribution varying between 7% and 30%. However, differences between ISS and GC conditions were observed.
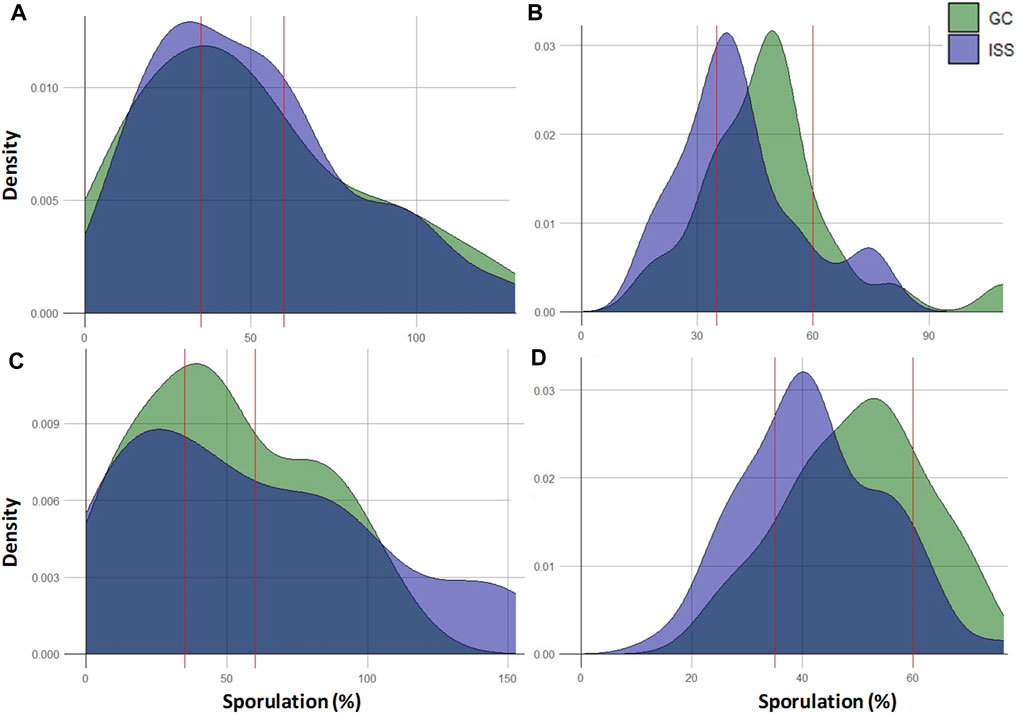
Figure 2. Density plots of Merlot and CS Mother plants according to the observed level of downy mildew sporulation. (A) Merlot 2022; (B) Merlot 2023; (C) CS 2022 and (D) CS 2023. Plants from the ISS condition are represented in light purple and plants from the GC condition are represented in green. The dark purple color indicates areas where the two conditions overlap. Red lines on the x-axis illustrates the threshold of low to moderate susceptibility (35%) and moderate to high susceptibility (60%).
Considering Merlot cultivar in 2022, based on the density plot, a similar density curve shape with a major peak was noticed for both ISS and GC conditions, although the ISS peak was present at a slightly lower sporulation percentage (next to the left side of the 35% red line) and higher compared to the GC peak (Figure 2A). However, the GC density was slightly higher than the one of ISS at very low and very high sporulation percentage. It was confirmed by the frequency analysis (Supplementary Table S1). Specifically, we noted a similar frequency distribution for ISS and GC plants categorized as highly-susceptible, and a tendency to have fewer individuals of ISS compared to GC. In 2023, both conditions had again a similar shape but with steeper density curves. However, we noted that ISS density curve was shifted to the left compared to the GC curve, peaking at lower sporulation percentages (ISS at 35% and GC at 50%) indicating a tendency to have more ISS plants with a low susceptibility level to P. viticola compared to Control (Figure 2B). Indeed, a higher number of ISS individuals were classified in the low-susceptible class (30% of ISS plants in comparison to 17% of GC) (Supplementary Table S1). Concerning the highly-susceptible class, as in 2022, a similar frequency distribution between ISS and GC was observed.
Regarding Cabernet Sauvignon plants and the year 2022, the density distribution at the lowest sporulation percentages was similar between ISS and GC conditions (Figure 2C). A higher density of GC plants was observed around the low sporulation threshold (35%) and at moderate and high sporulation percentage compared to the ISS plants density. At high sporulation percentages, the ISS density was higher than that of GC. However, frequency analysis highlighted that there was slightly more individuals classified as low-susceptible under ISS conditions compared to GC (Supplementary Table S1).
In 2023, similarly to Merlot, density curves displayed for CS were steeper and with higher peaks than in 2022. ISS and GC density curves were very similar but ISS one was shifted to the left, peaking at lower sporulation percentages (40% compared to GC that peaked at 55%). These observations indicated a tendency towards a higher proportion of ISS plants that displayed lower susceptibility to P. viticola compared to Control plants (Figure 2D). These observations were supported by the frequency data: 26% of ISS plants were classified as low-susceptible compared to 11% of GC plants, while only 7% of ISS plants were classified into the highly-susceptible class compared to 17% of GC plants (Supplementary Table S1). A graphical representation of the distribution of each CS and Merlot Mother plant according to the average sporulation percentages obtained over the 2 years of experimentation is provided in Supplementary Figures S4, 5, respectively.
With all these results, it came out that, whatever the grapevine variety, plants of the ISS condition showed a tendency towards being classified as low susceptible, despite slightly pronounced results in 2022.
3.3 Frequency distribution of Merlot progenies individuals
For each Merlot progeny individual, according to its mean sporulation level, we classified it into one of the three following classes: low-susceptibility (0%–35%), moderate-susceptibility (35.1%–60%), and high-susceptibility (60.1%–100%), as done for the Mother plants.
The density plots for both 2022 and 2023 exhibited a similar trend (Figure 3). However, the peak for the ISS condition was shifted to the left compared to the GC condition, highlighting a higher proportion of ISS plants with a low downy mildew susceptibility. For example, in 2022, the density curve of the GC and ISS progenies peaked around 45% and 35% sporulation, respectively. Moreover, the ISS progenies density curve showed a higher density at very low sporulation percentages, while the GC condition was more widely distributed at sporulation levels above 50%. As for Mother plants, a large overlap at moderate development percentages was noted for GC and ISS both conditions.
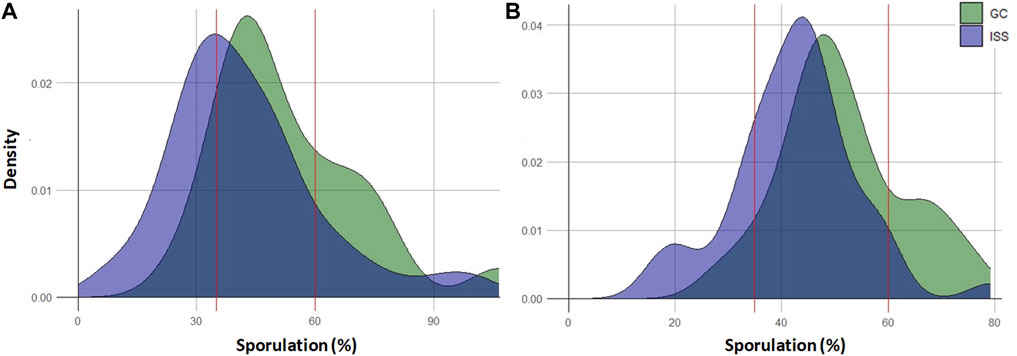
Figure 3. Density plots of Merlot progenies 2022 (A) and 2023 (B) according to the observed level of downy mildew sporulation. Plants from the ISS condition are represented in light purple and plants from the GC condition are represented in green. The dark purple color indicates areas where the two conditions overlap. Red lines on the x-axis illustrates the threshold of low to moderate susceptibility (35%) and moderate to high susceptibility (60%).
Frequency analysis revealed that the largest number of ISS and GC individuals (approximately two-thirds of them) were classified into the moderately-susceptible group (Supplementary Table S2). This analysis confirmed a distinct distribution of progenies in the low and high-susceptible classes based on the condition considered (ISS or GC) (Supplementary Table S2). More GC progeny plants were categorized as highly-susceptible (15% and 17% in 2022 and 2023, respectively) compared to 5 and 2% for ISS plants. The inverse frequency distribution was observed in the low-susceptibility class with more ISS individuals (23% and 13% of ISS compared to 5% and 4% of GC in 2022 and 2023, respectively). These results confirmed that progenies of individuals originated from canes placed in the ISS environment were more likely to exhibit a lower level of susceptibility towards downy mildew.
A graphical representation of the distribution of each Merlot progeny plant according to the average sporulation percentages obtained over the 2 years of experimentation is provided in Supplementary Figure S6.
As one of the main goals of this project was to benefit the space environment as a tool for obtaining grapevines with modified traits such as reducted susceptiblity to downy mildew, we considered the percentage of individuals based on the number of specific individuals that presented a mean of sporulation level significantly different from the total population (all individuals) through the 2 years of experimentation and considering Mothers and their respective progenies. Firstly, we noted that 41% of GC individuals and 62% of ISS were always distributed in the same class, regardless of the year of the experimentation and if we consider the Mother plant or one of its progenies. We also noted that a higher percentage of plants fell in the moderate-susceptibility class and less in the two other classes (Table 4). Then, our results showed a higher percentage of ISS Merlot individuals said low-susceptible compared to GC, with 11% and 0%, respectively (Table 4). Conversely, a lower percentage of ISS individuals than GC was noted in the highly-susceptible class (0% and 6%, respectively). Space conditions likely resulted in an enhanced percentage of plants showing a reduced susceptibility to downy mildew. Secondly, we considered all data of sporulation mean percentage for each Merlot Mother plant and its progenies over the 2 years of experimentation to obtain a representative value for each individual, regardless of the generation and the year. For that, a graphical representation of the distribution of each individual plant is provided in Figure 4. We observed that the sporulation percentages were in the range of 27% and 70%, with a mean of the total population of 44%. The majority of the plants (81% and 82% for GC and ISS, respectively) presented an moderate mean of sporulation. Considering GC plants, we noted one highly-susceptible plant (representing 6% of all GC plants) that was significantly different from the total population and one that is low-susceptible. No individual can be classified as highly-susceptible for the ISS condition even though four plants presented a sporulation mean higher than 55%. On the contrary, six plants (18% of all ISS plants) were significantly considered as low-susceptible.
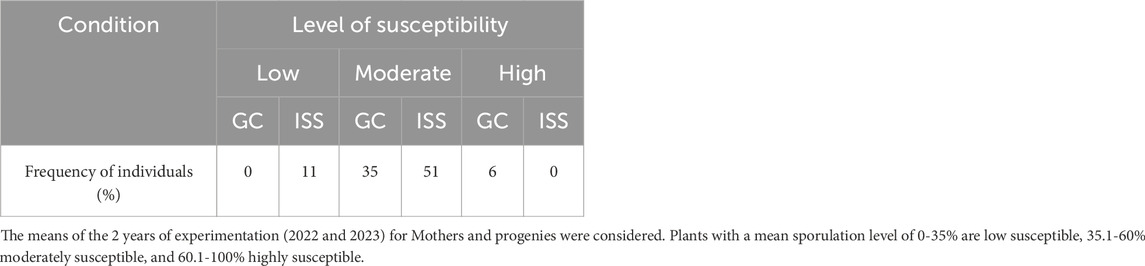
Table 4. Percentage of GC (Ground Control) and ISS (International Space Station) Merlot individuals classified as Low-Susceptibility, Moderate-Susceptibility, or High-Susceptibility) during the 2 years of experimentation (2022 and 2023).
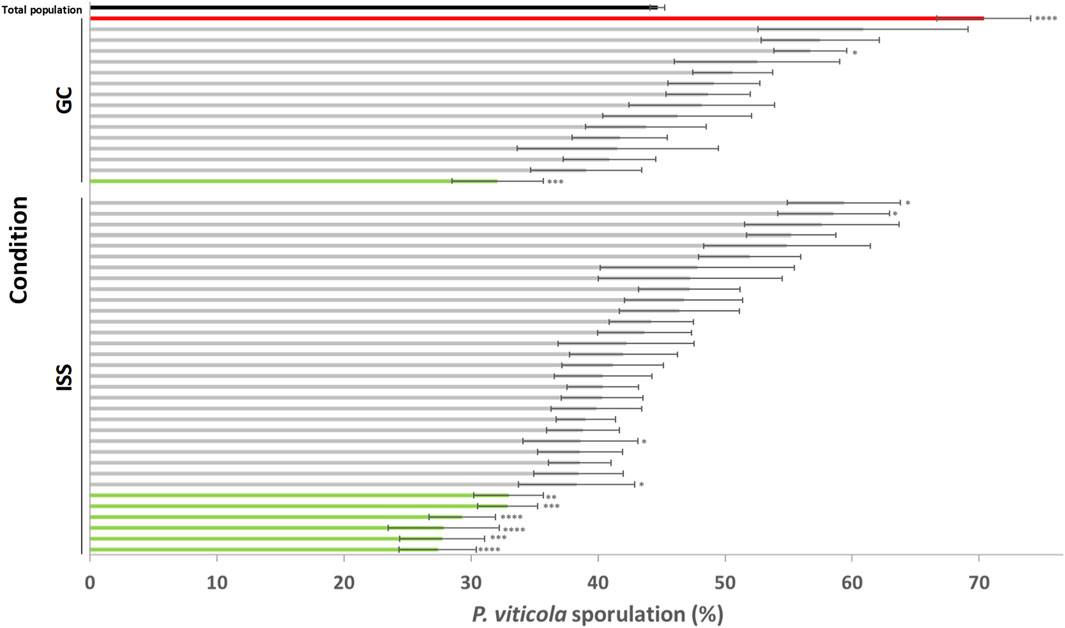
Figure 4. Sporulation percentage for each GC and ISS Merlot individual. The sporulation percentage is given for Ground Control (GC) plants and for those originating from canes stored in space conditions (ISS). It represented the mean of the data obtained from a Mother and its progenies for each individual over the 2 years of the experiment (2022 and 2023). The mean of the total population corresponding to all plants tested over the 2 years of experiment (2022 and 2023) Mother and progenies is represented by a black bar. Colors are assigned depending on the level of susceptibility against Plasmopara viticola as follows: red corresponds to highly susceptible plants (60.1%–100%), grey corresponds to moderately susceptible plants (35.1%–60%), and green corresponds to low susceptible plants (0%–35%). Significant difference levels of each individual compared to the total population were set at ****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.05.
4 Discussion
This study investigated the ISS environment’s influence on grapevine material during cane storage with the idea of taking advantage of evolutionary engineering, which could result in grapevines having higher resistance to various stresses. For that, dormant grapevine canes bearing one bud were exposed to a 10-month storage inside the International Space Station (ISS). A prolonged storage period was chosen to enhance the probability of triggering modifications of the phenotype of plants. In this study, we investigated the downy mildew susceptibility level of leaves of developed plants from ISS-stored canes.
Here, we chose to work with grapevine canes since they represent the basic material for vegetative propagation, the most prevalent technique in viticulture. A particular advantage of using canes (cuttings) is that with this method, genetically identical plants and clones of the parent vine were obtained. Thus, it allows the maintenance of desirable traits of the Mother grapevine of interest as these traits are more likely to be fixed across generations thanks to the vegetative propagation (Kashtwari et al., 2022). On the contrary, seeds will lead to distinct grapevine varieties that result from crossings of two parent varieties, so there is a risk of losing some desirable traits. Nevertheless, as seeds, canes can stay alive for several months without any specific maintenance. It is an excellent advantage as, in the ISS, no astronaut intervention is required. They must be maintained in the dark, near 4°C, and at a high humidity (70%–80%).
In our study, two grapevine cultivars, Cabernet Sauvignon (CS) and Merlot (Me), chosed as they are among the most cultivated across the world, were mainly distributed in the moderate- or high-susceptiblility classes (with a sporulation density above 35%). This observation is in accordance with the literature that mentioned that these two cultivars are susceptible to downy mildew (Calonnec et al., 2013; Bove et al., 2019).
Our study highlight a tendency to obtain a grapevine plants with a low susceptibility level to downy mildew after a space storage of the canes inside the ISS. Those results were observed for both cultivars. However, we noted discrepancies between both years of analysis in the sporulation means and frequencies of plants falling into the different categories. These discrepancies could result in a bias generated by our classification choice. Indeed, by looking in more detail at the values, we noted that the frequency distribution could be more comparable between years if we include plants from a larger sporulation threshold. Indeed, if instead of considering as highly-susceptible, plants with a mean of sporulation above 60.1%, we considered plants with a mean sporulation level above 55%, for instance, in 2023 the value of CS ISS highly-susceptible individuals will be 18% (instead of 7), wich is closer to the one observed in 2022. In addition, we noted a higher variability in the sporulation results and a lower amount of data in 2022 compared to 2023 that could lead to underestimating significantly different plants from the general population. Moreover, it is also possible that the environmental conditions encountered by the Mother plants, which were grown under a tunnel and not in a greenhouse, modified the susceptibility level of the plants to a further downy mildew infection. Such variations of the susceptibility level were already reported for experiments done with powdery mildew leaf disk assays performed on V. vinifera subsp. sylvestris seedlings for several years (Lukšić et al., 2022). Nevertheless, in our study, such heterogeneity was not noted for all individuals, and it did not hinder us from identifying a higher proportion of plants with a significant susceptibility reduction in the ISS condition compared to the GC condition, particularly in the case of Merlot. Leaf disk assays are recognized as an accurate method for performing the first screening of plants to evaluate their susceptibility level towards downy mildew (Calonnec et al., 2013). Nevertheless, such bioassay’s estimated level of plant susceptibility was sometimes not linked to what was further observed in a vineyard (Vezzulli et al., 2018). In our study, the Mother plants were placed under a tunnel, and such environmental conditions were similar to field conditions. Therefore, our results could accurately reflect the level of disease susceptibility during a vineyard assay. However, such vineyard assays must be done to confirm the results, especially due to a diversity of P. viticola isolates.
Concerning Progenies downy mildew susceptibility, unlike what was observed with Mother plants, no discrepancies between the data obtained in 2022 and 2023 have been noted. It could result from the fact that, during these two consecutive years, the progenies have grown under controlled conditions in a greenhouse, thus reducing the variability of plant responses to downy mildew.
A key finding of our study consist on the obtention of plants lineages, from the Mothers who have directly undergone space storage to their progenies, that show a significantly reduced susceptibility towards P. viticola. In this study, with regards to Merlot, space environment conditions (ISS) lead to 11% of individuals with a significant low-susceptibility to downy mildew, identified based on bioassays performed on Mother plants and their progenies over 2 years of experimentation. As a comparison, when a classical methodology for grapevine selection is performed, between 1% and 10% of clones with characteristics of interest are usually obtained (Gonçalves et al., 2019). Such classical clonal selection is time-consuming, taking approximately 15–20 years, and requires a genetic variability of the starting population (Rühl et al., 2004; Gonçalves et al., 2019; Farkas et al., 2023). Our results highlighted that using space environment could be a time-saving method that did not necessitate genetic variability of the starting population to obtain individuals with characteristics of interest in a proportion similar to or even higher than a classical selection methodology.
However, levels of disease severity observed in the less susceptible plants in our study is relatively high compared to what can be observed in studies on resistant grapevines, which could even be close to zero sporulation in the case of Vitis riparia and associated to the possession of resistance genes (Poltronieri et al., 2020; Štambuk et al., 2021). From our point of view, the effect we observed should not be described as a resistance phenomenon but as a reduced susceptibility towards downy mildew. Nonetheless, our results are promising since we demonstrated a marked effect. To that extent, it could be interesting to repeat this type of experiment, particularly by extending the space-storage time.
Space combines stresses, radiations and microgravity, for any terrestrial organism. In our case, this harsh environment can have resulted in a higher percentage of ISS plants with a significantly reduced susceptibility to P. viticola than GC plants. Ionizing radiations inside the ISS during the storage of the canes corresponding to radiations present in Low Earth Orbit (LEO) were 100 times higher than those measured on Earth (Restier-Verlet et al., 2021) by comparison to the natural absorbed radiation on Earth’s surface at 2.4 m/year. Thus, such radiations can induce genetic modifications, such as DNA mutations, in ISS canes. Indeed, a previous study reported that a short spaceflight of 6 days causes DNA mutations in rice seeds, resulting in modified phenotypes of plants that have developed from them. Some of these modified traits were transmitted to the next generations (Yu et al., 2007).
It has already been reported that exposing organisms to space environment conditions could result in the apparition of new strains of bacteria, yeasts, plants, or other organisms with agricultural or other industrial relevance (Horneck et al., 2010; Huang et al., 2015). He et al. (2006) mentioned that experiments done in space on seeds of more than 1,000 varieties of crops, fruits, or even forest trees resulted in plants developed from space-exposed seeds with favorable physiological and genetic variations.
Among several control plants, we occasionally observed reduction of the level of susceptibility that was similar to that observed in the low-susceptible ISS-stored plants, but this reduction did not persist from 1 year to the next nor from generation to the next. It could therefore be attributed to the natural variability of phenotypes. On the other hand, a significant percentage of ISS Mother plants and their progenies showed a low susceptibility level to downy mildew, indicating that the plant’s capacity to reduce disease severity is maintained through years and generations. Among the various possible hypotheses, this maintained downy mildew susceptibility reduction could be attributed to epigenetic modifications that would be induced by the conditions aboard the ISS. Indeed, it was mentioned that for an organism, facing a stress could lead to long-term behavioural effects, parlty through changes in the epigenome. Furthermore, some authors reported that a process involving epigenetic mechanisms, know as priming, can be set up. If occurred, by such mechanism, the plant will partially memorized the stress and could initiate a faster and/or more intense response to a subsequent stress (Berger et al., 2023). This priming effect could be transmitted to progenies through the transmission of epigenetic imprints which seems to operate more effectively in the case of agamous reproduction (Gallusci et al., 2023). Indeed, it was mentioned that spaceflight conditions induced epigenetic changes in A. thaliana seedlings with DNA methylation modifications (Xu et al., 2018). Some imprints of these DNA methylation modifications were observed in the offspring, accompanied by phenotypic modifications and changes in gene expression of various cellular metabolic pathways (Xu et al., 2021). Moreover, it has been shown that a tolerant cultivar exhibits a lower upregulation of the percentage of 5-methylcytosine (5-mC) at the early stage following P. viticola infection compared to a susceptible cultivar and a modified regulation of gene expression, suggesting that epigenetic regulation plays a role in disease susceptibility (Azevedo et al., 2022). In the future, genomic and methylome analyses could be conducted to specify potential genetic or epigenetic modifications occurring in ISS plants during the storage.
The final intention was to propose plants with a reduced susceptibility to downy mildew while maintaining the genetic background of the two international varieties studied here, Merlot and Cabernet Sauvignon. Continuing to grow such cultivars, but with enhanced disease resistance, will allow winegrowers to conduct their vineyard and winemaking process as they are accustomed to. As mentioned, the results presented in this study must be confirmed with field assays. In addition, it has to be investigated that no particular issues are encountered until the finished product is obtained, particularly when considering the wine quality. Such new clones could offer winegrowers to have at disposal a mixture of clones of these famous cultivars in their vineyard and so limiting naturally, for instance, disease development, or contrary increasing resilience of the plant to abiotic factors (Schöb et al., 2015; Newton, 2016; Savary et al., 2019; Delrot et al., 2020; Vezzulli et al., 2022). We hope that the modifications triggered by the ISS environment will be relatively different and increase the genetic diversity within these varieties. To conclude, using space environment could help to improve grapevine plasticity, representing a promising tool for developing grapevines with enhanced traits of interest to answer actual and future agricultural concerns and will contributing to reduce pesticide use.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Author contributions
DT: Data curation, Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing–original draft, Writing–review and editing. AB: Formal Analysis, Investigation, Methodology, Writing–original draft, Writing–review and editing. CB: Formal Analysis, Investigation, Methodology, Writing–original draft, Writing–review and editing. FH: Conceptualization, Project administration, Resources, Writing–original draft, Writing–review and editing. ML: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Visualization, Writing–original draft, Writing–review and editing. SC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by Space Cargo Unlimited SAS, Bordeaux.
Acknowledgments
We are grateful to Anaïs Ferrer for her help in the canes integration and S. Gambier and J. Jolivet for their help in plant production and maintenance.
Conflict of interest
Authors FH and ML were employed by Space Cargo Unlimited SAS.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fspas.2024.1415376/full#supplementary-material
References
Ahloowalia, B. S., and Maluszynski, M. (2001). Induced mutations-A new paradigm in plant breeding. Euphytica 118, 167–173. doi:10.1023/A:1004162323428
Azevedo, V., Daddiego, L., Cardone, M. F., Perrella, G., Sousa, L., Santos, R. B., et al. (2022). Transcriptomic and methylation analysis of susceptible and tolerant grapevine genotypes following Plasmopara viticola infection. Physiol. Plant. 174 (5), e13771. doi:10.1111/ppl.13771
Bavaresco, L. (2019). Impact of grapevine breeding for disease resistance on the global wine industry. Acta Hortic. 1248, 7–14. doi:10.17660/ActaHortic.2019.1248.2
Bavaresco, L., and Squeri, C. (2022). Outlook on disease resistant grapevine varieties. BIO Web Conf. 44, 06001. doi:10.1051/bioconf/20224406001
Belli, M., and Tabocchini, M. A. (2020). Ionizing radiation-induced epigenetic modifications and their relevance to radiation protection. Int. J. Mol. Sci. 21, 5993–6034. doi:10.3390/ijms21175993
Berger, M. M. J., Stammitti, L., Carrillo, N., Blancquaert, E., Rubio, B., Teyssier, E., et al. (2023). Epigenetics: an innovative lever for grapevine breeding in times of climatic changes. Oeno One 57 (2), 265–282. doi:10.20870/oeno-one.2023.57.2.7405
Bove, F., Bavaresco, L., Caffi, T., and Rossi, V. (2019). Assessment of resistance components for improved phenotyping of grapevine varieties resistant to downy mildew. Front. Plant Sci. 10, 1559. doi:10.3389/fpls.2019.01559
Bove, F., Savary, S., Willocquet, L., and Rossi, V. (2020). Simulation of potential epidemics of downy mildew of grapevine in different scenarios of disease conduciveness. Eur. J. Plant Pathol. 158, 599–614. doi:10.1007/s10658-020-02085-8
Calonnec, A., Wiedemann-Merdinoglu, S., Delière, L., Cartolaro, P., Schneider, C., and Delmotte, F. (2013). The reliability of leaf bioassays for predicting disease resistance on fruit: a case study on grapevine resistance to downy and powdery mildew. Plant Pathol. 62, 533–544. doi:10.1111/j.1365-3059.2012.02667.x
Corio-Costet, M. F., Dufour, M. C., Cigna, J., Abadie, P., and Chen, W. J. (2011). Diversity and fitness of Plasmopara viticola isolates resistant to QoI fungicides. Eur. J. Plant Pathol. 129, 315–329. doi:10.1007/s10658-010-9711-0
Delrot, S., Grimplet, J., Carbonell-Bejeranoo, P., Schwandner, A., Bert, P. F., Bavaresco, L., et al. (2020). “Genetic and genomic approaches for adaptation of grapevine to climate change,” in Genomic designing of climate-smart fruit crops. Editor C. Kole (Cham: Springer). doi:10.1007/978-3-319-97946-5_7
De Micco, V., Aronne, G., Caplin, N., Carnero-Diaz, E., Herranz, R., Horemans, N., et al. (2023). Perspectives for plant biology in space and analogue environments. NPJ Microgravity 9, 67. doi:10.1038/s41526-023-00315-x
Farkas, E. A., Jahnke, G., Szőke, B., Deák, T., Oláh, R., Oláh, K., et al. (2023). Clonal selection of autochthonous grape varieties in Badacsony, Hungary. Horticulturae 9, 994. doi:10.3390/horticulturae9090994
Ferrandino, A., Pagliarani, C., and Pérez-Álvarez, E. P. (2023). Secondary metabolites in grapevine: crosstalk of transcriptional, metabolic and hormonal signals controlling stress defence responses in berries and vegetative organs. Front. Plant Sci. 14, 1124298. doi:10.3389/fpls.2023.1124298
Foria, S., Monte, C., Testolin, R., Di Gaspero, G., and Cipriani, G. (2019). Pyramidizing resistance genes in grape: a breeding program for the selection of elite cultivars. Acta Hortic. 1248, 549–554. doi:10.17660/actahortic.2019.1248.73
Furukawa, S., Nagamatsu, A., Nenoi, M., Fujimori, A., Kakinuma, S., Katsube, T., et al. (2020). Space radiation biology for ”Living in space. Biomed. Res. Int. 2020, 1–25. doi:10.1155/2020/4703286
Gallusci, P., Agius, D. R., Moschou, P. N., Dobránszki, J., Kaiserli, E., and Martinelli, F. (2023). Deep inside the epigenetic memories of stressed plants. Trends Plant Sci. 28 (2), 142–153. doi:10.1016/j.tplants.2022.09.004
Gessler, C., Pertot, I., and Perazzolli, M. (2011). Plasmopara viticola: a review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 50, 3–44. doi:10.14601/Phytopathol_Mediterr-9360
Gonçalves, E., Graça, A., and Martins, A. (2019). Grapevine clonal selection in Portugal: a different approach. BIO Web Conf. 12, 01003. doi:10.1051/bioconf/20191201003
He, X., Liu, M., Lu, J., Xue, H., and Pan, Y. (2006). “Space mutation breeding: a brief introduction of screening new floricultural, vegetable and medicinal varieties from earth-grown plants returned from China's satellites/spaceships” in the Floriculture,”. Ornamental and plant biotechnology. Editor J. T. da Silva (London, United Kingdom: Global Science Books), IV. Chapter: 28.
Horneck, G., Klaus, D. M., and Mancinelli, R. L. (2010). Space microbiology. Microbiol. Mol. Biol. R. 74, 121–156. doi:10.1128/mmbr.00016-09
Huang, B., Liu, N., Rong, X., Ruan, J., and Huang, Y. (2015). Effects of simulated microgravity and spaceflight on morphological differentiation and secondary metabolism of Streptomyces coelicolor A3(2). Appl. Microbiol. Biotechnol. 99, 4409–4422. doi:10.1007/s00253-015-6386-7
Kashtwari, M., Mansoor, S., Wani, A. A., Najar, M. A., Deshmukh, R. K., Baloch, F. S., et al. (2022). Random mutagenesis in vegetatively propagated crops: opportunities, challenges and genome editing prospects. Mol. Biol. Rep. 49, 5729–5749. doi:10.1007/s11033-021-06650-0
Kimbrough, J. M., Salinas-Mondragon, R., Boss, W. F., Brown, C. S., and Sederoff, H. W. (2004). The fast and transient transcriptional network of gravity and mechanical stimulation in the Arabidopsis root apex. Plant Physiol. 136, 2790–2805. doi:10.1104/pp.104.044594
Lukšić, K., Zdunić, G., Hančević, K., Mihaljević, M. Ž., Mucalo, A., Maul, E., et al. (2022). Identification of powdery mildew resistance in wild grapevine (Vitis vinifera subsp. sylvestris Gmel Hegi) from Croatia and Bosnia and Herzegovina. Sci. Rep. 12 (1), 2128. doi:10.1038/s41598-022-06037-6
Malhi, G. S., Kaur, M., and Kaushik, P. (2021). Impact of climate change on agriculture and its mitigation strategies: a review. Sustainability 13, 1318. doi:10.3390/su13031318
Manzano, A., Carnero-Diaz, E., Herranz, R., and Medina, F. J. (2022). Recent transcriptomic studies to elucidate the plant adaptive response to spaceflight and to simulated space environments. IScience 25, 104687. doi:10.1016/j.isci.2022.104687
Mohanta, T. K., Mishra, A. K., Mohanta, Y. K., and Al-Harrasi, A. (2021). Space breeding: the next-generation crops. Front. Plant Sci. 12, 771985. doi:10.3389/fpls.2021.771985
Newton, A. C. (2016). Exploitation of diversity within crops— the key to disease tolerance? Front. Plant Sci. 7, 665. doi:10.3389/fpls.2016.00665
OIV (2017). Distribution variétale du vignoble dans le monde. Focus OIV 2017. Available at: https://www.oiv.int/public/medias/5861/fr-distribution-vari-tale-du-vignoble-dans-le-monde.pdf.
Pedneault, K., and Provost, C. (2016). Fungus resistant grape varieties as a suitable alternative for organic wine production: benefits, limits, and challenges. Sci. Hortic. 208, 57–77. doi:10.1016/j.scienta.2016.03.016
Pertot, I., Caffi, T., Rossi, V., Mugnai, L., Hoffmann, C., Grando, M. S., et al. (2017). A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 97, 70–84. doi:10.1016/j.cropro.2016.11.025
Pirrello, C., Magon, G., Palumbo, F., Farinati, S., Lucchin, M., Barcaccia, G., et al. (2023). Past, present, and future of genetic strategies to control tolerance to the main fungal and oomycete pathogens of grapevine. J. Exp. Bot. 74, 1309–1330. doi:10.1093/jxb/erac487
Poltronieri, P., Marrazzo, M. T., and Cipriani, G. (2020). “Grapevine: resistance genes, sRNAs and immunity,” in Applied plant biotechnology for improving resistance to biotic stress. Editors P. Poltronieri, and Y. Hong (Academic Press, Elsevier), 151–179. doi:10.1016/B978-0-12-816030-5.00007-0
Prasad, B., Richter, P., Vadakedath, N., Mancinelli, R., Krüger, M., Strauch, S. M., et al. (2020). Exploration of space to achieve scientific breakthroughs. Biotechnol. Adv. 43, 107572. doi:10.1016/j.biotechadv.2020.107572
Rani, L., Thapa, K., Kanojia, N., Sharma, N., Singh, S., Grewal, A. S., et al. (2021). An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 283, 124657. doi:10.1016/j.jclepro.2020.124657
Restier-Verlet, J., El-Nachef, L., Ferlazzo, M. L., Al-Choboq, J., Granzotto, A., Bouchet, A., et al. (2021). Radiation on Earth or in space: what does it change? Int. J. Mol. Sci. 22, 3739. doi:10.3390/ijms22073739
Rühl, E. H., Konrad, H., Lindner, B., and Bleser, E. (2004). Quality criteria and targets for clonal selection in grapevine. Acta Hortic. 652, 29–33. doi:10.17660/ActaHortic.2004.652.1
Salem-Fnayou, A. B., Bouamama, B., Ghorbel, A., and Mliki, A. (2011). Investigations on the leaf anatomy and ultrastructure of grapevine (Vitis vinifera) under heat stress. Microsc. Res. Tech. 74, 756–762. doi:10.1002/jemt.20955
Savary, S., Willocquet, L., Pethybridge, S. J., Esker, P., McRoberts, N., and Nelson, A. (2019). The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 3, 430–439. doi:10.1038/s41559-018-0793-y
Schöb, C., Kerle, S., Karley, A. J., Morcillo, L., Pakeman, R. J., Newton, A. C., et al. (2015). Intraspecific genetic diversity and composition modify species-level diversity-productivity relationships. New Phytol. 205, 720–730. doi:10.1111/nph.13043
Štambuk, P., Šikuten, I., Preiner, D., Nimac, A., Lazarević, B., Marković, Z., et al. (2021). Screening of Croatian native grapevine varieties for susceptibility to Plasmopara viticola using leaf disc bioassay, chlorophyll fluorescence and multispectral imaging. Plants 10 (4), 661. doi:10.3390/plants10040661
Töpfer, R., and Trapp, O. (2022). A cool climate perspective on grapevine breeding: climate change and sustainability are driving forces for changing varieties in a traditional market. Theor. Appl. Genet. 135, 3947–3960. doi:10.1007/s00122-022-04077-0
Vezzulli, S., Gramaje, D., Tello, J., Gambino, G., Bettinelli, P., Schwandner, A., et al. (2022). “Genomic designing for biotic stress resistant grapevine,” in Genomic designing for biotc stress resistant fruit crops. Editor C. Kole (Cham: Springer), 87–255. doi:10.1007/978-3-030-91802-6_4
Vezzulli, S., Vecchione, A., Stefanini, M., and Zulini, L. (2018). Downy mildew resistance evaluation in 28 grapevine hybrids promising for breeding programs in Trentino region (Italy). Eur. J. Plant Pathol. 150 (2), 485–495. doi:10.1007/s10658-017-1298-2
Wan, D. Y., Guo, Y., Cheng, Y., Hu, Y., Xiao, S., Wang, Y., et al. (2020). CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Hortic. Res. 7, 116. doi:10.1038/s41438-020-0339-8
Xu, P., Chen, H., Hu, J., and Cai, W. (2021). Potential evidence for transgenerational epigenetic memory in Arabidopsis thaliana following spaceflight. Commun. Biol. 4 (1), 835. doi:10.1038/s42003-021-02342-4
Xu, P., Chen, H., Jin, J., and Cai, W. (2018). Single-base resolution methylome analysis shows epigenetic changes in Arabidopsis seedlings exposed to microgravity spaceflight conditions on board the SJ-10 recoverable satellite. Npj Microgravity 4 (1), 12. doi:10.1038/s41526-018-0046-z
Keywords: Vitis vinifera, Plasmopara viticola, ISS, space breeding, microgravity, radiations, Cabernet Sauvignon, Merlot
Citation: Taillis D, Burdziej A, Bautista C, Haag FWM, Lebert M and Cluzet S (2024) Grapevines grown from canes having spent 10 months in space: study of their downy mildew susceptibility. Front. Astron. Space Sci. 11:1415376. doi: 10.3389/fspas.2024.1415376
Received: 10 April 2024; Accepted: 17 June 2024;
Published: 23 October 2024.
Edited by:
Ricardo Amils, Autonomous University of Madrid, SpainReviewed by:
José Manuel Martínez Lozano, Autonomous University of Madrid, SpainVicenta De La Fuente, Autonomous University of Madrid, Spain
Copyright © 2024 Taillis, Burdziej, Bautista, Haag, Lebert and Cluzet. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphanie Cluzet, c3RlcGhhbmllLmNsdXpldEB1LWJvcmRlYXV4LmZy; Michael Lebert, bWljaGFlbC5sZWJlcnRAZmF1LmRl