 Rémi Millot*
Rémi Millot* Camille Debattice
Camille Debattice Viviana Ligorini
Viviana Ligorini Jérémy BracconiNicolas GattaccecaAmbre CrescioniJean-Baptiste Ronchi-PerfettiAlexandre VelaRomain BastienAntoine Aiello
Jérémy BracconiNicolas GattaccecaAmbre CrescioniJean-Baptiste Ronchi-PerfettiAlexandre VelaRomain BastienAntoine Aiello Jean-José Filippi*
Jean-José Filippi*- Unité d'Appui et de Recherche (UAR) 3514 STELLA MARE–University of Corsica / CNRS, Lieu-dit U Casone, lido de la Marana, Biguglia, France
The Mediterranean spider crab, Maja squinado (Herbst, 1788), is one of the largest species of spider crab and holds significant interest for commercial and stock enhancement purposes. Thus, it is essential to identify optimal rearing conditions to attempt mass production, as several factors affect the growth and survival of this species during the early juvenile phase in aquaculture. In this study, we evaluated the effect of stocking density on the growth parameters and survival rate of M. squinado reared in an experimental hatchery as part of a spider crab stock enhancement feasibility project. Groups of spider crabs were reared at two initial densities (1.6 and 4.6 individuals·L−1). After 29 days of culture (i.e., from megalopae to C3 juveniles), early juveniles reared at the higher density showed a significantly higher weight gain ratio and specific growth rate as well as a significantly higher Fulton’s condition index specifically for C3 juveniles, reflecting a better general well-being. In contrast, the survival at the higher density was negatively affected compared to the lower rearing density. This study improves baseline knowledge about M. squinado rearing to help develop commercial applications and establish management strategies and restoration programs for endangered M. squinado populations going forward.
1 Introduction
In the Mediterranean Sea, the major biological research topics are sustainability of resources and conservation of biodiversity. The Mediterranean Sea is a small, oligotrophic, semi-enclosed sea representing 0.82% of the world’s ocean (Karydis, 2021). Despite its small volume, it is a biodiversity hotspot and includes 4% to 18% of the world’s marine diversity (Coll et al., 2010). Benthic communities are among the most diverse in the Mediterranean Sea, however, they are highly sensitive to natural and anthropogenic perturbations (de Madron et al., 2011). Currently, combined pressures from overfishing, invasive species, and climate change are rapidly altering the biodiversity of Mediterranean marine communities (Boudouresque et al., 2017; de Madron et al., 2011).
The Mediterranean spider crab Maja squinado (Herbst, 1788) (Crustacea: Decapoda: Majidae), is one of the largest spider crabs and is a commercially important European crab species. This crustacean is restricted to the Mediterranean Sea (Sotelo et al., 2008) and is found from the subtidal area to about 50 m depth (Sotelo et al., 2009). M. squinado is captured by small-scale fisheries, especially off the Mediterranean coasts of Croatia, France and Spain (http://www.sealifebase.org/report/FAO), where the species is of notable local gastronomic interest. In addition to being a relevant economic resource, M. squinado possesses biological characteristics, such as high fecundity and rapid larval development (Durán et al., 2012), that are favorable for its potential aquacultural rearing.
Environmental degradation and poor fisheries management are the main causes of the decline and even decimation of certain fisheries (Born et al., 2004). In the last decades of the 20th century, M. squinado became a target species in fisheries of the Mediterranean Sea, leading to overexploitation (Abad, 2003). In some areas, this crab species has almost disappeared, such as off the Balearic Islands, the island of Majorca, and along the Catalan coast (Rotllant et al., 2010). Because of its high ecological value and diminished population size, M. squinado is considered endangered and was included in the Action Plan for the Mediterranean (UNEP, 1996). The Bern Convention lists M. squinado as a protected species (Appendix III) (Bern Convention, 1979), and the Barcelona Convention identifies it as an exploitation-regulated species (Annex III) (Barcelona Convention, 1995). In certain areas, such as coastal Corsica, the recreational fishing of M. squinado is prohibited (Prefectoral decree no. 2014177-001; Mirmand, 2014). An absence of scientific monitoring in the Mediterranean Sea has led to a lack of knowledge needed to assess stock status. Consequently, the International Union for Conservation of Nature (IUCN) has no official conservation status for this species.
Restocking (also referred to as stock enhancement) is one option to help depleted stocks recover, with an emphasis on developing methods for mass-production. The Balearic Islands government has developed a program for the recovery of wild populations through the release of hatchery-reared juveniles M. squinado (Durán et al., 2012). Moreover, other programs aimed at enhancing stocks of severely depleted crustacean species have been implemented, such as the common European lobster (Homarus gammarus) in Europe (Williams, 2007), the mud crab (Scylla paramamosain) in Asia (Ut et al., 2007), and the blue crab (Callinectes sapidus) in North America (Zohar et al., 2008).
A substantial amount of prior zootechnical knowledge is essential to successfully produce a large number of individuals, for both market demands and stock restoration purposes. However, in the case of M. squinado, information on the rearing of this species is scarce. In contrast, numerous examples of such studies exist for the European spider crab, Maja brachydactyla (Castejón et al., 2015, 2019; Domingues et al., 2012; Guerao et al., 2012; Rotllant et al., 2015; Simeó et al., 2015). Until morphological and genetic analyses were conducted, M. brachydactyla and M. squinado were considered as the same taxon (M. squinado). However, recent studies distinguished M. brachydactyla as the Atlantic species and M. squinado as the Mediterranean species (Guerao et al., 2011; Neumann, 1998; Sotelo et al., 2008).
The few existing studies on M. squinado focus on the larval rearing, growth and reproductive process under intensive culture conditions (Durán et al., 2012, 2013; Guerao et al., 2008; Guerao and Rotllant, 2010). M. squinado, like most brachyuran crustaceans, exhibits a complex biphasic life cycle characterized by a pelagic larval phase followed by a benthic juvenile-adult phase. Ovigerous females carry fertilized eggs, from which larvae emerge initially as planktonic zoeae. These larvae undergo two successive zoeal stages (zoea 1 and zoea 2) in the plankton before metamorphosing into a semi-benthic megalopal stage. Subsequently, the megalopae settle and molt into the first juvenile crab instar, referred to as the C1 stage. Growth proceeds through additional juvenile instars (C2, C3, etc.), each marked by successive molts accompanied by incremental increases in size (Durán et al., 2012; Guerao and Rotllant, 2010).
It is imperative for restocking purposes to determine the optimal environmental conditions and nutritional requirements as possible limiting factors for attaining elevated survival and growth in the juvenile phase of M. squinado. Growth and survival rates in crab aquaculture are affected by abiotic and biotic factors, such as food availability (Alaminos and Domingues, 2008), holding space (Swiney et al., 2013) and stocking density (Daly et al., 2009). Cannibalism during crab rearing is the main response to effect of these factors, which affects the productivity and profitability of aquaculture rearing. The occurrence of cannibalism in aquaculture has been recognized as a major problem since the mid-1970s (Hecht and Appelbaum, 1988) and is well-documented for crustacean species, for example, the swimming crab Portunus pelagicus (Marshall et al., 2005) and M. brachydactyla (Domingues and Alaminos, 2008). Several methods exist for mitigating antagonistic interactions among crabs, including artificial habitats and shelters, substrate and high-quality diets, grading size by classes or separating individuals by compartments in order to reduce cannibalism among crabs and achieve higher productivity (Castejón et al., 2019; Daly et al., 2009; Fernandez, 1999; Mann et al., 2007; Swiney et al., 2013; Ut et al., 2007; Zmora et al., 2005).
Since 2019, a Mediterranean crustaceans restocking research program conducted at the Stella Mare Research Unit, Corsica, France, has focused particularly on M. squinado. The program aims to increase the production of juvenile crabs to eventually supply sufficient quantities for ecological restoration experiments around the island. To support these restoration efforts, this study assessed the effect of stocking density on the growth parameters and survival of M. squinado post-settlement juveniles (megalopae to C1–C3 juveniles) reared under controlled conditions to assess the potential of this species for restocking programs.
2 Material and methods
2.1 Broodstock and larval rearing
Four adult ovigerous females were captured in the northwestern Mediterranean Sea, along the Bastia Coast (Corsica, France) by a local fisherman in May 2023 using a trammel net. Particular attention was paid to avoiding shock or stress to the females after their capture. Within less than 3 h of capture, the females were transported alive inside an insulated container—at approximately 18°C to avoid thermal shock—to the Stella Mare Marine Research Centre of the University of Corsica–CNRS (Biguglia, France). At the Stella Mare hatchery, the ovigerous females’ anatomical integrity was verified and females were weighed using a Ranger 7000 precision balance (Ohaus, Florham Park). The females weighed 1.46 ± 0.24 kg. They were brushed, cleaned, thoroughly rinsed with seawater and transferred a 200 L semi-square flat-bottom fiberglass tank (i.e., 0.5m depth x 0.8m width x 0.6m length) connected to a recirculated aquaculture system (RAS) that provided 0.1 μm filtered seawater at 19°C with photoperiod maintained at 12L:12D with Luxina T8 LED 21 W lamps. Each day, broodstock were fed ad libitum a mixed diet of frozen squid, mussels, fish, and shrimp. The active, newly hatched zoea of one female were distributed equally among four white 850 L cylindrical–conical tanks (water temperature of 19°C), for a density around 70 larvae·L−1 until the larvae reached the megalopae instar (i.e., 13 days). Only one female was used because the eggs of the females were not at same developmental phase (indicated by the color of the egg mass; Durán et al., 2012). The water for all larval tanks was sourced from the same treatment system, and the renewal rate of the water was approximately 75% per hour in each tank. The larvae (i.e., zoea 1 and zoea 2) were fed 1st instar Artemia nauplii (Artemia salina) at a density of 4.3 Artemia·mL−1.
2.2 Experimental design
Thirteen-day-old larval M. squinado (i.e., reaching megalopae instar) with an initial carapace length of 2.17 ± 0.15 mm were utilized to initiate the study. Larvae were reared over a 29-day period (up to the 3rd crab instar, C3). To initiate the study, larvae were gently and randomly transferred in suspension using a transparent beaker, in 2000 L square flat-bottom fiberglass tanks (0.5m depth x 2m width x 2m length) connected to a RAS to maintain constant physico-chemical parameters, especially temperature. Particular care was taken to ensure larvae were never directly handled and remained submerged in water throughout the process. Larvae were stocked at two stocking densities: low: 1.6 ind·L−1 (241.8 ind·m−2); and high: 4.6 ind·L−1 (712.5 ind·m−2), corresponding to 3114 and 9176 megalopa per 2 m3 tank, respectively. Density treatments were conducted in triplicate.
Because Pastor et al. (2011) (Maja squinado) and Zmora et al. (2005) (Callinectes sapidus) demonstrated the role of shelters in promoting settlement and reducing cannibalism in crabs because of the increased surface area, this informed the decision to install in each tank a shelter/artificial habitat. The shelters/artificial habitats consisted of 20 collector cups (Bys Ostréiculture, Meze) (corresponding to one black rod of 47 black cups) attached to each other with ropes in the following arrangement: 5 horizontally and 4 vertically. Each shelter took up the entire space of the tank, going from the bottom to the surface. Aeration bubblers were positioned to avoid areas of low turbulence and homogenize conditions in the tanks (Figure 1).
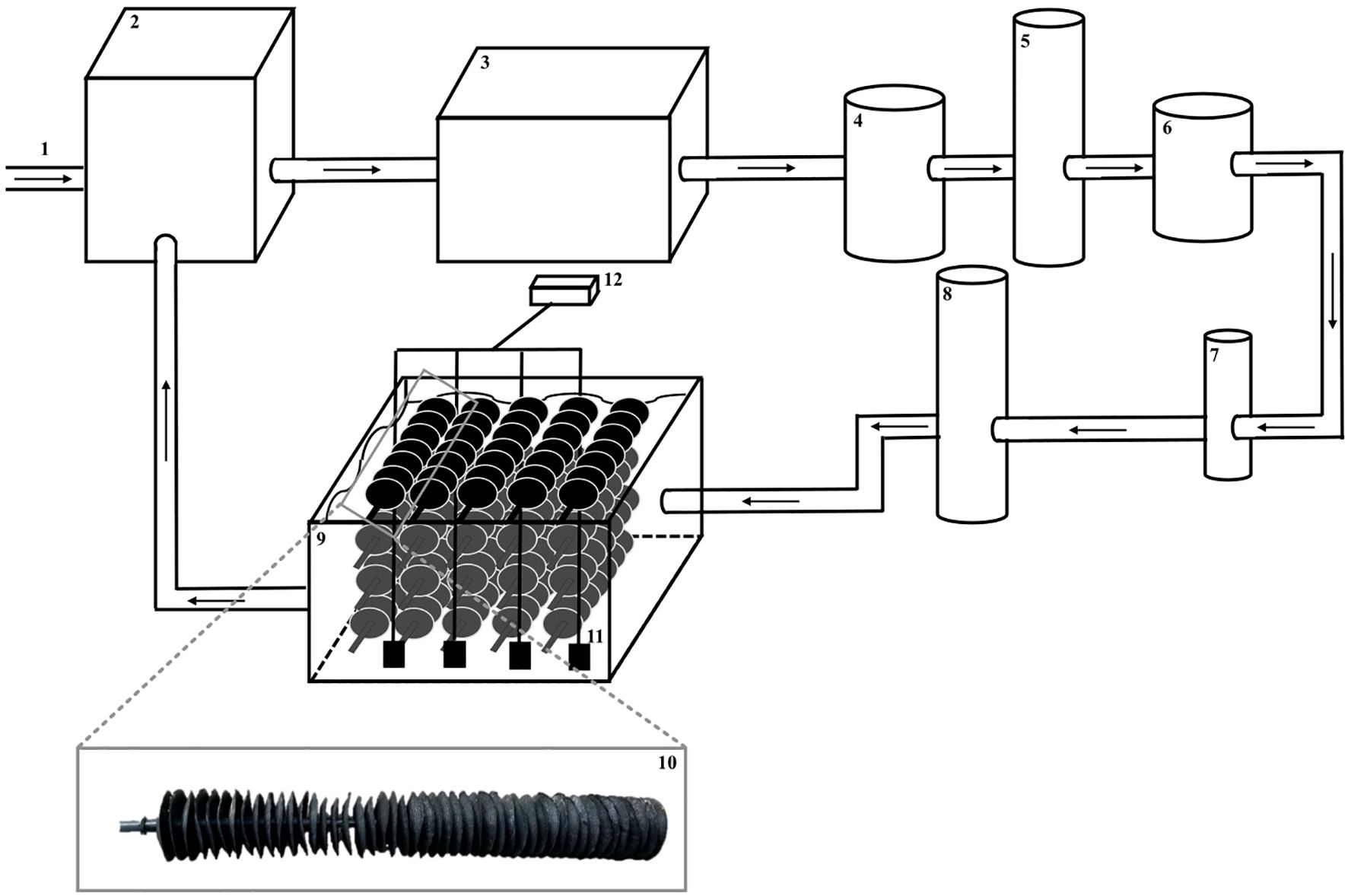
Figure 1. Schematic representation of the rearing system: (1) inflow for water renewal, (2) buffer tank, (3) pump, (4) mechanical filter, (5) ultraviolet sterilizer, (6) biological filter, (7) heat pump, (8) degassing column, (9) rearing tank (2 m3) with artificial habitat/shelter which composed of 20 collector cups (10) detail of the structure of each collector cup (Bys Ostréiculture, Meze), (11) aeration bubblers, (12) aeration system.
During the experiment, water temperature, pH, nitrite-nitrogen, nitrate-nitrogen and un-ionized ammonia nitrogen concentration, dissolved oxygen (DO) as percent saturation, and total gas pressure were continuously monitored using Oxyguard Handy Polaris 2 and TGP probes (Oxyguard International, Farum, Denmark). Particular attention was paid to ensure that those parameters remained within a physiologically optimal range (pH: 8.05 ± 0.01; DO: 95.27 ± 0.42%; un-ionized ammonia nitrogen: <0.001 mg.L−1, nitrite-nitrogen: <0.001 mg.L−1 and nitrate-nitrogen: <0.001 mg.L−1). For the megalopa-juvenile transition period, temperature and salinity were maintained at 22.84 ± 0.16°C and 37.6 ± 0.2 ppt respectively and total dissolved gas pressure in water was maintained in equilibrium with the ambient atmospheric pressure. Tanks were illuminated from above with Luxina T8 LED 21 W lamps, with an intensity of 400 lx at the water surface and a 12L:12D photoperiod regime controlled by a timer.
The megalopa larvae (3rd larval stage), C1, C2, and C3 juveniles crabs were all fed ad libitum with live prey: 2nd instar Artemia salina metanauplii (INVE, Belgium) enriched with Easy DHA (docosahexaenoic acid) Selco, containing a mix of omega-3 fatty acids and vitamins (INVE Aquaculture, Utah). However, for C2 and C3 juveniles, frozen mysid and frozen Artemia HUFA super shrimp (Ocean Nutrition, Belgium) were added to their diet.
2.3 Data collection
In each tank, once all individuals reached the next developmental stage, they were counted in order to obtain the number of individuals dead or alive. This count was realized at 23, 34 and 42 DAH, corresponding to the times where all individuals reached the next stage (13 DAH: beginning or megalopae; 23 DAH: megalopae molts to C1 juveniles; 34 DAH: C1 juvenile molts to C2 juveniles; 42 DAH: C2 juvenile molts to C3 juveniles; i.e., corresponding to 29 days of experiment). During cleaning procedure (at each of the three-molting transition between the stages investigated, i.e. megalopa to C1 juvenile, C1 to C2, C2 to C3), a bucket filled with seawater was placed next to the tank in which DO was monitored. The artificial habitats/shelters were removed from the tanks and the individuals attached were dislodged (using gentle water flow), then counted before being placed in the bucket. Siphoning was then employed to recover the waste feed and individuals remaining in the bottom of the tank. At the end of each of the three cleaning procedures (one procedure for each molting transition), different mesh sieves were applied in function of stage (1000 µm: megalopae to C1 juveniles; 2000 µm from C1 to C2 and C2 to C3 juveniles). The individuals retained by the sieve were counted and placed in the bucket. Then, the artificial habitats/shelters were rearranged in the same configuration as initially, and the individuals were released into their respective tanks.
In parallel to counting, larvae from each treatment were randomly sampled for biometric measurements (n = 20 per replicate). Measurements were taken with a Stemi 2000-C stereomicroscope (Zeiss, Oberkochen) micrometric eyepiece and rounded to the nearest mm. Carapace width was measured as the greatest distance across the carapace (measured between the tips of the 3rd branchial spines) and carapace length as the distance between the rostral margin (without rostral spines) and the posterior margin of the carapace (without intestinal spines), as reported by Guerao and Rotllant (2010). Wet weight was determined with a TA 164i precision balance accurate to 0.0001 g (VWR, Radnor).
2.4 Calculations and statistical analysis
The following parameters were used to evaluate the growth and performance of the experimental animals (Guerao and Rotllant, 2009; Solanki et al., 2023):
where Oi was the number of megalopae larvae at the start of the experiment, Os was the number of individuals that successfully molted to each stage and O1 was the number of individuals in the previous OS stage. Wf represents the final wet weight (mg), Wi represents the initial wet weight (mg), CLf denote the final carapace length (mm) and CLi denote the initial carapace length (mm).
All growth parameters were analyzed for normality and homogeneity of variances by examining residual values using Shapiro–Wilk’s test and Levene’s test, respectively. The non-normal and heterogeneous data were transformed with arcsine to stabilize variation (Snedecor and Cochran, 1989). One-way ANOVA was performed to check for significant differences among treatments, followed with Cohen’s d to measure the effect size. The evolution of cumulative survival rates over the duration of the experiment (survival time) for each stocking density treatments were analyzed using the Kaplan–Meier survival analysis with a Weibull distribution of errors using the R package ‘survival’ (Therneau and Grambsch, 2000; Therneau, 2015). The Mantel–Cox test results were represented by Kaplan–Meier curves produced using the package ‘survminer’ (Kassambara et al., 2021). The Mantel–Cox test or log-rank test is a non-parametric and hypothesis-based test for comparing the distribution of survival curves of at least two samples and describes the probability per time unit of the event occurring as a function of basic probability. The Proportional Hazards Assumption was tested by Schoenfeld residual test, in order to assess whether the Mantel Cox model adequately described the data. Data were expressed as mean ± standard error (SE). All statistical analyses were performed using R software v.4.2.0 (R Core Team, 2022), considering P < 0.05 as the level of significance.
3 Results
3.1 Growth parameters and survival after a 29-day rearing period
The growth parameters recorded after the 29-day rearing period (the entire experiment from megalopa to C3 juveniles) are presented in Table 1. At the end of the 29-day experiment, the highest specific growth rate and weight gain ratio were observed in individuals reared at the high stocking density treatment (ANOVA, F = 57.13, P < 0.05, d = 4.32; F = 54.19, P < 0.05, d = 4.21, respectively), with a specific growth rate of 5.33 ± 0.03 and a weight gain ratio of 368.90 ± 4.34%, compared to 5.07 ± 0.01 and 334.56 ± 1.70% for individuals reared at the low stocking density treatment. In contrast, there were no significant differences between density treatments concerning final mean weight and length, average weight and length gain, condition factor (K), and yield (i.e., biomass). The final survival rate was similar for both density treatments but the survival time (survival rate over time) was significantly different. Indeed, the log-rank (Mantel–Cox) test for Kaplan–Meier survival curves revealed a significant difference in survival time (log-rank: χ2 = 55.35, P < 0.001; Schoenfeld residual test, χ2 = 1.94, P > 0.05; Figure 2).

Table 1. Zootechnical performance parameters of Maja squinado juveniles, reared under two stocking densities (low: 1.6 ind·L−1 and high: 4.6 ind·L−1) on the whole experiment (from Megalopa to C3, i.e., 29 days). Differences were tested using one-way ANOVA.
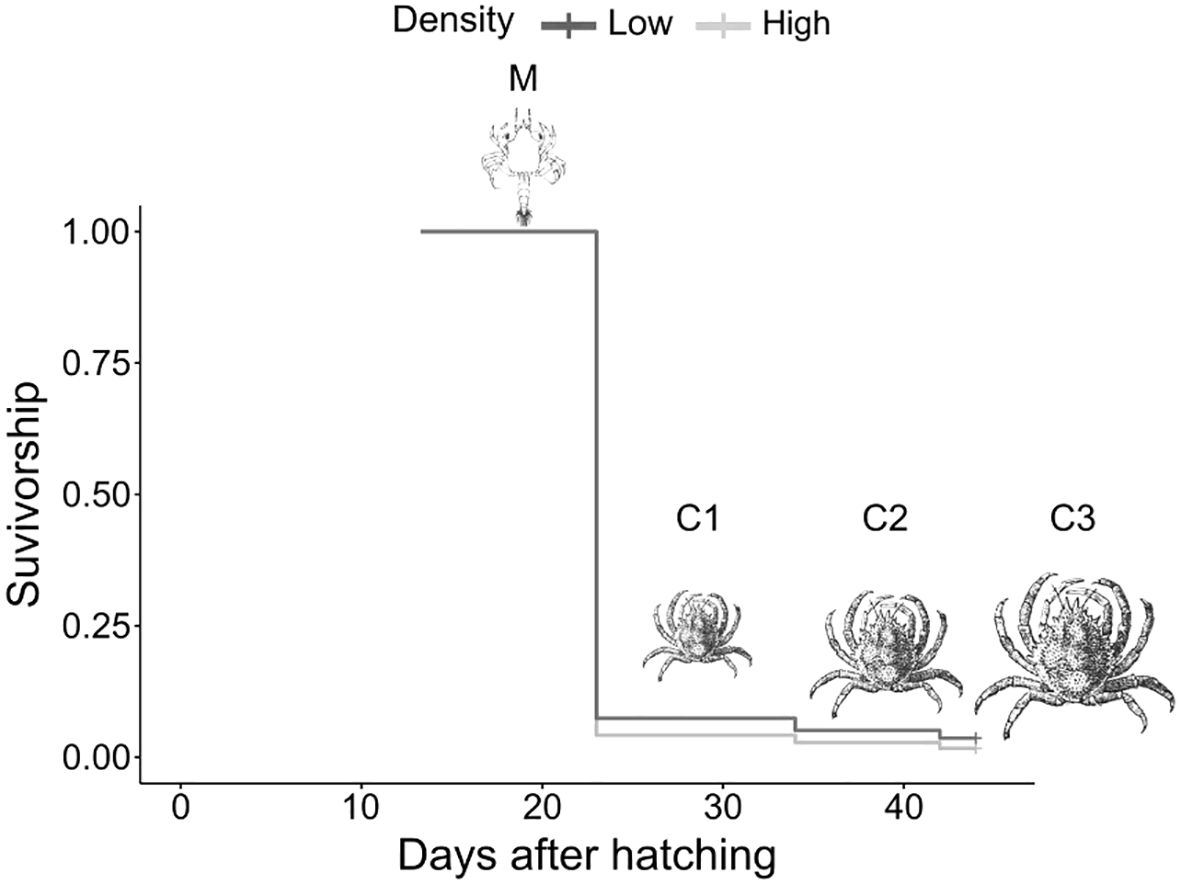
Figure 2. Maja squinado development survival rate over time per stocking density treatment, determined using Kaplan-Meier survival analysis.
3.2 Growth parameters and survival through molts
Table 2 presents the various growth parameters observed through molts (developmental stages) across the experimental period. No significant differences were detected between stocking density treatments for each molt concerning all production parameters measured except for Fulton’s condition index. The C3 juveniles reared at the high stocking density had a significantly higher Fulton’s condition index (0.015 ± 0.4 x 10−3) than C3 juveniles reared at the low stocking density (0.014 ± 0.3 x 10−3), but with a small effect size (Table 2) (ANOVA, F = 4.726, P < 0.05, d = 0.394). Regarding growth parameters related to carapace length, parameters were similar for each both stocking density treatment at each molt, while parameters related to weight tended to be higher at the last stage of the experiment (C3 juveniles) for the high stocking density treatment compared with the low stocking density treatment (Table 2).
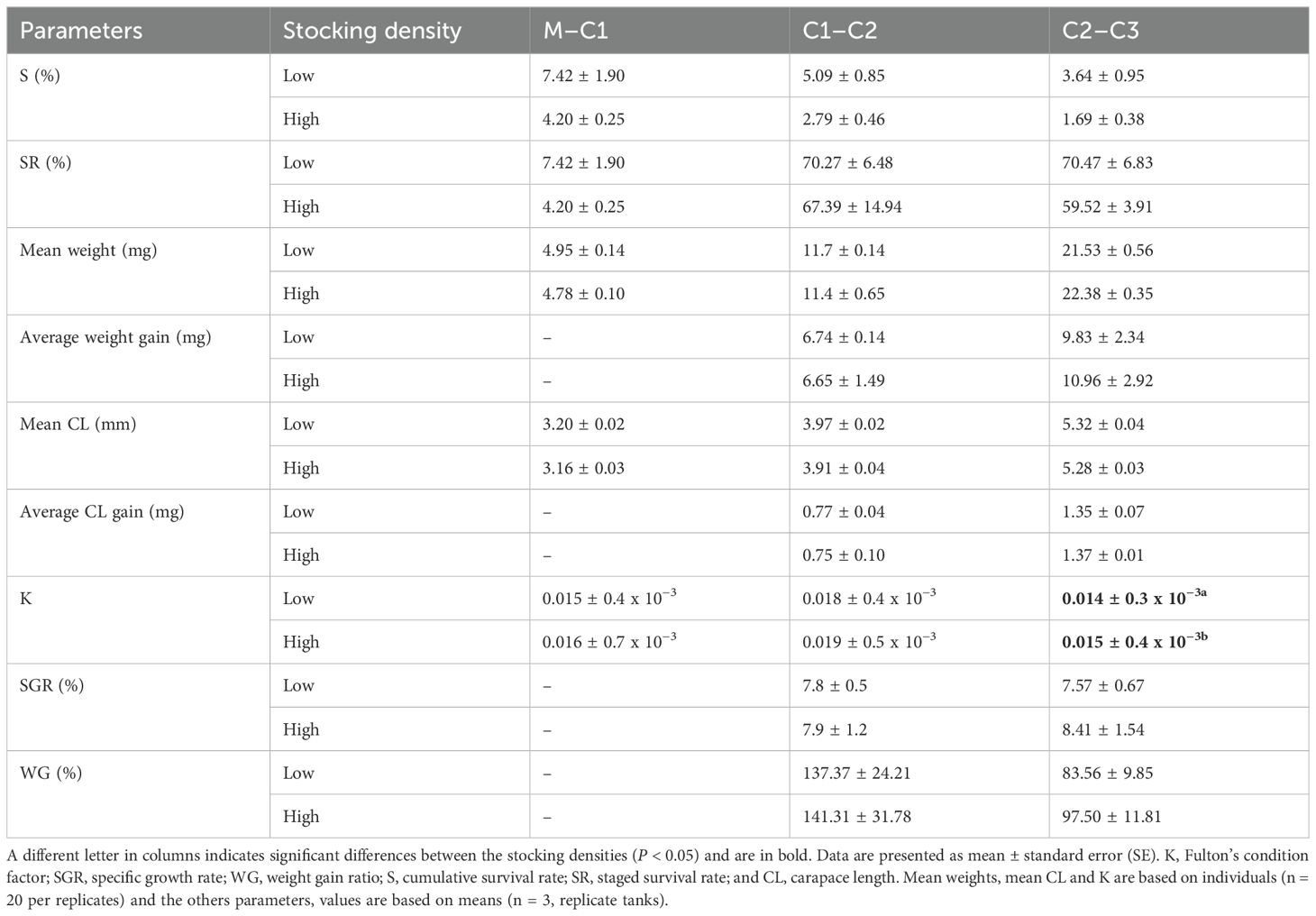
Table 2. Summary of zootechnical performance parameters of Maja squinado juveniles, reared under two stocking densities (low: 1.6 ind·L−1 and high: 4.6 ind·L−1) during verifications or molts (i.e., stages) throughout the experimental period. Differences were tested using on one-way ANOVA.
4 Discussion
Stocking density is a crucial factor influencing the growth performance of aquacultured animals (Cuvin-Aralar et al., 2009; del Mar Gil et al., 2019; Yuan et al., 2018; Zhang et al., 2024). The effects of high stocking density on the growth and survival of crustaceans have been well documented, generally showing a negative correlation between growth and stocking density (González et al., 2010; Li et al., 2007; Naranjo-Páramo et al., 2004). Nevertheless, despite these numerous studies focused on crustaceans, few reports have documented the effect of stocking density on the growth and survival of M. squinado. Only two studies on the effect of stocking density were available on this species before the present study. The first study, by del Mar Gil et al. (2019), constitutes a microscale investigation, using one or two individuals per 1.5 L cylinder (i.e., 0.67 and 1.33 ind·L−1) from C1 to C5 juveniles, while the second study, by Pastor et al. (2011), provided a macroscale analysis on 2,352 3,794 5,500 6,721 and 9,740 individuals reared in 1,300 L tanks (1.96, 3.6, 4.58, 5.6 and 8.12 ind·L−1) from megalopae to C5 in a brief communication lacking any statistical analysis. Because of this scarcity of M. squinado rearing studies, our experiment represents a preliminary contribution, aiming to identify key trends in growth and survival for improved juvenile culture methods of this species towards future stock enhancement efforts.
The high stocking density treatment in this study positively impacted the wet weight gain, the specific growth rate, and Fulton’s CI (specifically for C3 juveniles) of early juveniles of M. squinado reared at two initial densities (1.6 and 4.6 individuals·L−1), but negatively impacted the survival time (survival rate over time). However, no differences were observed regarding the final survival rate at the end of the experiment. Our results contrast with those of del Mar Gil et al. (2019), who reported higher survival time with two M. squinado juveniles per cylinder (a small experimental unit in their study) than with one, but no significant difference concerning the growth parameters, except for C3 juveniles. These discrepancies may be due to different initial densities and rearing configurations: del Mar Gil et al. (2019) studied the stocking density effect between C1 and C5, whereas our study started at the megalopa stage. Moreover, in their study, rearing was conducted in 10 cm diameter PVC cylinders (1.5 L, 78.54 cm2) with 1 mm mesh bottoms. Thus, the density applied in their study was much lower than in our experimental design, particularly in terms of surface area: 0.01 ind.m−² (low density) and 0.03 ind.m−2 (high density), and the individuals were fed fresh food (i.e., mussel flesh) and dry pellets. Thus, the reasons for the difference between our results and those of del Mar Gil et al. (2019) might be multifactorial. According to Dong et al. (2018), the relationship between rearing density, growth performance, and survival must be carefully assessed in relation to the specific culture management system, as the effect of stocking density can vary considerably. In general, high stocking densities will increase social interactions for limited resources (e.g., food, territory, shelter or any other crucial factor for survival) as demonstrated by previous studies (Damodaran et al., 2018; Savolainen et al., 2004). These changes of social interaction may lead to two possibilities: 1) Triggering a rising energy demand for activities and lead to the reduction in the available energy for growth such as demonstrated for various crustaceans (González et al., 2010; Li et al., 2007; Naranjo-Páramo et al., 2004; Zhang et al., 2024). 2) Improving nutritional needs through the supply of additional nitrogen, vitamins, and other essential substances linked to cannibalism and lead to the increase of the energy available for growth, as demonstrated on Paralithodes camtschaticus (red king crab) (Daly et al., 2009). While water quality deterioration can also be a factor (Sookying et al., 2011), it was likely negligible in our study due to maintained high water quality. Hence, intraspecific cannibalism emerged as the principal explanation for our observations, perhaps which led to lower survival time but higher weight gain ratio and specific growth of individuals reared at the high stocking density, as well as a higher Fulton’s condition index for C3 juveniles, reflecting better overall well-being. This similar behavior has been demonstrated in various spider crab studies (M. brachydactyla; Durán et al., 2012) and, specifically, in M. squinado (Pastor et al., 2011; del Mar Gil et al., 2019).
In the present study, M. squinado exhibited a typical mortality pattern regardless of stocking density with high levels of survival after the critical settlement phase, a well-known pattern for this species (del Mar Gil et al., 2019; Pastor et al., 2011). During the settlement period (megalopa to first juvenile crab), M. squinado populations markedly decreased, having a survival rate between 7.42 ± 1.90% (low density) and 4.20 ± 0.25% (high density), closely matching the mass-culture outcomes reported by Pastor et al. (2011). The specific causes of M. squinado mortalities at settlement remain uncertain but appear multifactorial. They may stem from factors not directly related to stocking density. With regard to handing stress, Iglesias et al. (2002) suggested that early juvenile M. brachydactyla are very delicate, as a high handling stress-related mortality was observed and their observation appears to be even more critical for M. squinado. In the case of individual culture of M. squinado, survival during the settlement period did not exceed 20% (Guerao and Rotllant, 2010) and was lower than that of M. brachydactyla (about 48%) under similar experimental conditions (Guerao and Rotllant, 2009). Hence, M. squinado larvae may be more sensitive to handling stress than M. brachydactyla. In the present study, high mortality may be explained by the individuals being manipulated at each molt in the rearing system. In terms of larval nutrition, Guerao and Rotllant (2009) suggested that high mortality in M. brachydactyla rearing may stem from accumulated nutritional deficiencies during larval development. This problem is confirmed in other species, such as the mud crab Scylla serrata, and it is accentuated in the case of other species with several zoeal stages (Hamasaki et al., 2002). Additionally, regarding behavioral aspects, Urcera et al. (1993) proposed that an inability to adapt led to significant mortality at the change from predatory pelagic behavior to benthic behavior of M. brachydactyla larvae. The high mortality of early developmental stages of M. squinado recorded in this study also identified the settlement phase as a critical period for development. Finally, cannibalism is often cited as the main factor causing large losses in mass culture of early stage crabs, possibly linked to several factors such as lack of space and proper diet induced by high stocking density (Borisov et al., 2007; del Mar Gil et al., 2019; Pastor et al., 2011). Inter-cohort cannibalism can function as a density-dependent self-regulatory mechanism (Moksnes et al., 1997). In the present study, cannibalism was observed in both density treatments, although it was greater at the higher stocking density treatment. Mortality decreased for subsequent molts, from C1 to C3 juvenile stages, similar to the findings of Pastor et al. (2011) on the same species, who reported less cannibalism in these later stages. Typically, newly molted crabs with soft exoskeletons are most vulnerable to attacks by conspecifics (Elgar and Crespi, 1992; Marshall et al., 2005), a phenomenon also seen been in M. brachydactyla (Domingues and Alaminos, 2008) and Paralithodes camtschaticus (Borisov et al., 2007). Although individual rearing could minimize this issue (Kovatcheva et al., 2006), such a system is not practical for commercial aquaculture operations that require high-density conditions to maximize production (Daly et al., 2009).
In summary, the apparent contradiction of reduced survival time yet improved specific growth parameters under high-density rearing conditions may be linked to increased foraging behavior and nutritional benefits derived from agonistic interactions. Asynchronous molting leads to size variability, potentially intensifying predation on smaller individuals and stimulating overall growth (Daly et al., 2009). Conspecific cannibalism, might supply additional nitrogen, vitamins, and other essential nutrients, even under conditions of excess food (del Mar Gil et al., 2019). However, the relatively brief duration of this study may explain the limited impact on growth parameters and the small effect size value observed regarding Fulton’s Condition Index (CI) as well as shelter/artificial habitats effect by reducing cannibalism as reported by Daly et al. (2009). Future research should be conducted over longer periods as well as to assess the shelter effect in order to enhance statistical analyses. In addition, video monitoring could be employed to quantify instances of cannibalism. Ultimately, a tradeoff between survival rates and production must be determined (Daly et al., 2009). From a hatchery standpoint, a lower survival rate may be acceptable if it maximizes yield (Zmora et al., 2005; Ut et al., 2007). Nonetheless, future experiments should be carried out to propose solutions to cannibalism, such as feed formulation adapted to nutritional requirements or the use of shelters or artificial habitats, which have already proved useful in increasing the survival rate of juveniles by reducing cannibalism in other crustacean species (Moksnes et al., 1997; Mann et al., 2007).
5 Conclusions
Under the rearing conditions examined in this study, a higher stocking density of early developmental stage of M. squinado positively affected growth parameters including weight gain ratio, specific growth rate and Fulton’s condition index, despite lower survival time. One of the major causes of the results of this study might be due to agonistic behavior (i.e., conspecific cannibalism). Ultimately, successfully rearing Mediterranean spider crabs holds considerable promise not only for commercial aquaculture development but, more importantly, for restoring depleted natural populations. Despite limitations, this work offers valuable preliminary insights into improving early stage rearing protocols for the under-researched species M. squinado, providing a crucial step toward establishing reliable production methods for both commercial and ecological restoration purposes.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements. The study concerns M. squinado, a crustacean species, which according to French legislation in force does not require ethical approval.
Author contributions
RM: Data curation, Investigation, Visualization, Writing – original draft, Writing – review & editing. CD: Conceptualization, Data curation, Methodology, Visualization, Writing – original draft, Writing – review & editing. JB: Investigation, Resources, Writing – review & editing. NG: Investigation, Resources, Writing – review & editing. AC: Investigation, Resources, Writing – review & editing. J-BR-P: Resources, Writing – review & editing. AV: Writing – review & editing. RB: Project administration, Writing – review & editing. AA: Funding acquisition, Project administration, Supervision, Writing – review & editing. J-JF: Conceptualization, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. VL: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was part of the DHAVID research program (Développement Halieutique: Valorisation, Innovation et Déploiement) and was financially supported by the French CPER (Contrat de Plan État-Région).
Acknowledgments
The authors would like to thank the Corsican fisherman for providing berried M. squinado females.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abad R. (2003). Actas de las I Jornadas sobre Reservas Marinas y I Reunión de la Red Iberoamericana de Reservas Marinas (RIRM). Ministerio de Agricultura, Pesca y Alimentación. Secretaría Gen. Técnica, 47–54.
Alaminos J., Domingues P. (2008). Effects of different natural or prepared diets on growth and survival of juvenile spider crabs, Maja brachydactyla (Balss 1922). Aquac. Int. 16, 417–425. doi: 10.1007/s10499-007-9154-6
Barcelona Convention (1995). Annex II and Annex III, Convention for the Protection of The Mediterranean Sea Against Pollution. Available online at: http://eunis.eea.europa.eu/references/1818/species (Accessed September 2, 2024).
Borisov R. R., Epelbaum A. B., Kryakhova N. V., Tertitskaya A. G., Kovatcheva N. P. (2007). Cannibalistic behavior in red king crabs reared under artificial conditions. Rus. J. Mar. Biol. 33, 227–231. doi: 10.1134/S1063074007040049
Born A. F., Immink A. J., Bartley D. M. (2004). “Marine and coastal stocking: global status and information needs,” in Marine ranching, vol. 429. (FAO Fisheries Technical Paper, Rome), 1–19.
Boudouresque C. F., Blanfuné A., Fernandez C., Lejeusne C., Pérez T., Ruitton S., et al. (2017). Marine biodiversity-warming vs. biological invasions and overfishing in the Mediterranean Sea: Take care,’one train can hide another’. MOJ Ecol. Environ. Sci. 2, 1–13. doi: 10.15406/mojes.2017.02.00031
Castejón D., Rotllant G., Giménez L., Torres G., Guerao G. (2015). The effects of temperature and salinity on the survival, growth and duration of the larval development of the common spider crab Maja brachydactyla (Balss 1922) (Brachyura: Majidae). J. Shellf. Res. 34, 1073–1083. doi: 10.2983/035.034.0334
Castejón D., Rotllant G., Guerao G. (2019). Factors influencing successful settlement and metamorphosis of the common spider crab Maja brachydactyla Balss 1922 (Brachyura: Majidae): Impact of larval density, adult exudates and different substrates. Aquaculture 501, 374–381. doi: 10.1016/j.aquaculture.2018.11.053
Coll M., Piroddi C., Steenbeek J., Kaschner K., Ben Rais Lasram F., Aguzzi J., et al. (2010). The biodiversity of the Mediterranean sea: estimates, patterns, and threats. PloS One 5, e11842. doi: 10.1371/journal.pone.0011842
Convention on the Conservation of European Wildlife and Natural Habitats (Bern Convention) (1979). Annex II and III. Bern/Berne, 19. Available online at: http://conventions.coe.int/Treaty/FR/Treaties/Html/104-2.htmhttp://conventions.coe.int/Treaty/FR/Treaties/Html/104-3.htm.
Cuvin-Aralar M. L. A., Lazartigue A. G., Aralar E. V. (2009). Cage culture of the Pacific white shrimp Litopenaeus vannamei (Boone 1931) at different stocking densities in a shallow eutrophic lake. Aquac. Res. 40, 181–187. doi: 10.1111/j.1365-2109.2008.02081.x
Daly B., Swingle J. S., Eckert G. L. (2009). Effects of diet, stocking density, and substrate on survival and growth of hatchery-cultured red king crab (Paralithodes camtschaticus) juveniles in Alaska, USA. Aquaculture 293, 68–73. doi: 10.1016/j.aquaculture.2009.04.010
Damodaran D., Mohammed Koya K., Mojjada S. K., Lalaji C. D., Dash G., Vase V. K., et al. (2018). Optimization of the stocking parameters for mud spiny lobster Panulirus polyphagus (Herbst 1793) capture-based aquaculture in tropical open sea floating net cages. Aquacult. Res. 49, 1080–1086. doi: 10.1111/are.13557
del Mar Gil M., Pastor E., Durán J. (2019). Survival and growth of hatchery-reared Mediterranean spider crab juveniles, Maja squinado, under different rearing conditions. Aquaculture 498, 37–43. doi: 10.1016/j.aquaculture.2018.08.001
de Madron X. D., Guieu C., Sempéré R., Conan P., Cossa D., D’ortenzio F., et al. (2011). Marine ecosystems’ responses to climatic and anthropogenic forcings in the Mediterranean. Prog. Oceanogr. 91, 97–166. doi: 10.1016/j.pocean.2011.02.003
Domingues P., Alaminos J. (2008). Efectos de la densidad de cultivo y de elementos de refugio en el crecimiento y supervivencia de juveniles de centollo, Maja brachydactyla (Balss 1922). Rev. biol. marina y oceanogr. 43, 121–127. doi: 10.4067/S0718-19572008000100012
Domingues P., Alaminos J., García-Garrido S., Hachero-Cruzado I., Rosas C. (2012). Growth and survival of juvenile spider crabs, Maja brachydactyla (Balss 1922), fed with fresh or frozen mussels. Aquac. Res. 43, 167–177. doi: 10.1111/j.1365-2109.2011.02813.x
Dong J., Zhao Y. Y., Yu Y. H., Sun N., Li Y. D., Wei H., et al. (2018). Effect of stocking density on growth performance, digestive enzyme activities, and nonspecific immune parameters of Palaemonetes sinensis. Fish shellf. Immunol. 73, 37–41. doi: 10.1016/j.fsi.2017.12.006
Durán J., Palmer M., Pastor E. (2013). Growing reared spider crabs (Maja squinado) to sexual maturity: the first empirical data and a predictive growth model. Aquaculture 408, 78–87. doi: 10.1016/j.aquaculture.2013.05.031
Durán J., Pastor E., Grau A., Valencia J. M. (2012). First results of embryonic development, spawning and larval rearing of the Mediterranean spider crab Maja squinado (Herbst) under laboratory conditions, a candidate species for a restocking program. Aquac. Res. 43, 1777–1786. doi: 10.1111/j.1365-2109.2011.02983.x
Elgar M. A., Crespi B. J. (1992). Cannibalism: Ecology and Evolution among diverse Taxa (USA: Oxford University Press).
Fernandez M. (1999). Cannibalism in Dungeness crab Cancer magister: effects of predator-prey size ratio, density, and habitat type. Mar. Ecol. Prog. Ser. 182, 221–230. doi: 10.3354/meps182221
González R., Celada J. D., González A., García V., Carral J. M., Sáez-Royuela M. (2010). Stocking density for the intensive rearing of juvenile crayfish, Pacifastacus leniusculus (Astacidae), using Artemia nauplii to supplement a dry diet from the onset of exogenous feeding. Aquac. Int. 18, 371–378. doi: 10.1007/s10499-009-9250-x
Guerao G., Andree K., Froglia C., Simeo C., Rotllant G., Simeó C. (2011). Identification of European species of Maja (Decapoda: Brachyura: Majidae): RFLP analyses of COI mtDNA and morphological considerations. Sci. Mar. 75, 129–134. doi: 10.3989/scimar.2011.75n1129
Guerao G., Pastor E., Martin J., Andres M., Estevez A., Grau A., et al. (2008). The larval development of Maja squinado and M. brachydactyla (Decapoda, Brachyura, Majidae) described from plankton collected and laboratory-reared material. J. Nat. Hist. 42, 2257–2276. doi: 10.1080/00222930802254755
Guerao G., Rotllant G. (2009). Survival and growth of post-settlement juveniles of the spider crab Maja brachydactyla (Brachyura: Majoidea) reared under individual culture system. Aquaculture 289, 181–184. doi: 10.1016/j.aquaculture.2008.12.033
Guerao G., Rotllant G. (2010). Development and growth of the early juveniles of the spider crab Maja squinado (Brachyura: Majoidea) in an individual culture system. Aquaculture 307, 105–110. doi: 10.1016/j.aquaculture.2010.06.006
Guerao G., Simeó C. G., Anger K., Urzúa Á., Rotllant G. (2012). Nutritional vulnerability of early zoea larvae of the crab Maja brachydactyla (Brachyura, Majidae). Aquat. Biol. 16, 253–264. doi: 10.3354/ab00457
Hamasaki K., Suprayudi M. A., Takeuchi T. (2002). Mass mortality during metamorphosis to megalops in the seed production of mud crab Scylla serrata (Crustacea, Decapoda, Portunidae). Fish. Sci. 68, 1226–1232. doi: 10.1046/j.1444-2906.2002.00559.x
Hecht T., Appelbaum S. (1988). Observations on intraspecific aggression and coeval sibling cannibalism by larval and juvenile Claias gariepinus (Clariidae: Pisces) under controlled conditions. J. zool. 214, 21–44. doi: 10.1111/j.1469-7998.1988.tb04984.x
Iglesias J., Sánchez F. J., Moxica C., Fuentes L., Otero J. J., Pérez J. L. (2002). Datos preliminares sobre el cultivo de larvas y juveniles de centolla Maja squinado Herbst 1788 en el Centro Oceanográfico de Vigo del Instituto Español de Oceanografía. Bol. Inst. Esp. Oceanogr. 18, 25–30.
Karydis M. (2021). Biodiversity Changes in the Mediterranean Marine Environment: The role of species Invasions and Climate Change. Adv. Mar. Biol. 1, 1–45. doi: 10.52305/AVHN5735
Kassambara A., Kosinski M., Biecek P. (2021). survminer: drawing survival curves using ‘ggplot2’Available online at: https://CRAN.R-project.org/package=survminer (Accessed July 1, 2019).
Kovatcheva N., Epelbaum A., Kalinin A., Borisov R., Lebedev R. (2006). Early life history stages of the red king crab Paralithodes camtschaticus (Tilesius 1815): biology and culture. VNIRO Publishing, Moscow, p 116.
Li X., Dong S., Lei Y., Li Y. (2007). The effect of stocking density of Chinese mitten crab Eriocheir sinensis on rice and crab seed yields in rice-crab culture systems. Aquaculture 273, 487–493. doi: 10.1016/j.aquaculture.2007.10.028
Mann D. L., Asakawa T., Kelly B., Lindsay T., Paterson B. (2007). Stocking density and artificial habitat influence stock structure and yield from intensive nursery systems for mud crabs Scylla serrata (Forsskål 1775). Aquac. Res. 38, 1580–1587. doi: 10.1111/j.1365-2109.2006.01626.x
Marshall S., Warburton K., Paterson B., Mann D. (2005). Cannibalism in juvenile blue-swimmer crabs Portunus pelagicus (Linnaeus 1766): effects of body size, moult stage and refuge availability. Appl. Anim. Behav. Sci. 90, 65–82. doi: 10.1016/j.applanim.2004.07.007
Mirmand C. (2014). Arrêté n°2014177-001 portant interdiction de pêche de certaines espèces marines aux pêcheurs de loisir dans les eaux territoriales autor de las Corse, June 26, 2014 (République Française: Direction Interrégionale de la Mer Méditerranée Service Rêglementation Contrôle).
Moksnes P. O., Lipcius R. N., Pihl L., Van Montfrans J. (1997). Cannibal–prey dynamics in young juveniles and postlarvae of the blue crab. J. Exp. Mar. Biol. Ecol. 215, 157–187. doi: 10.1016/S0022-0981(97)00052-X
Naranjo-Páramo J., Hernandez-Llamas A., Villarreal H. (2004). Effect of stocking density on growth, survival and yield of juvenile redclaw crayfish Cherax quadricarinatus (Decapoda: Parastacidae) in gravel-lined commercial nursery ponds. Aquaculture 242, 197–206. doi: 10.1016/j.aquaculture.2004.05.017
Neumann V. (1998). A review of the Maja squinado (Crustacea: Decapoda: Brachyura) species-complex with a key to the eastern Atlantic and Mediterranean species of the genus. J. Nat. Hist. 32, 1667–1684. doi: 10.1080/00222939800771191
Pastor E., Durán J., Caimari M. A. (2011). “Experiences in mass production of mediterranean spider crab Maja squinado juveniles for restocking studies,” in Proceedings Aquaculture Europe Congress (European Aquaculture Society, Greece), 843–844.
R Core Team (2022). R: A language and environment for statistical computing. Available online at: https://www.r-project.org/ (Accessed April 20, 2022).
Rotllant G., Alaminos J., Cerezo J., Fernández-Palacios E., Pastor E. (2010). “Cría de centolla (Maja sp.),” in Informe final Junta de cultivos marinos Planes nacionales de cultivos marinos (JACUMAR, Madrid, Spain).
Rotllant G., Simeó C. G., Macià G., Estévez A. (2015). High environmental salinity reduces the reproductive potential of the spider crab Maja brachydactyla (Decapoda, M ajidae). Mar. Eco 36, 496–505. doi: 10.1111/maec.12157
Savolainen R., Ruohonen K., Railo E. (2004). Effect of stocking density on growth, survival and cheliped injuries of stage 2 juvenile signal crayfish Pasifastacus leniusculus Dana. Aquaculture 231, 237–248. doi: 10.1016/j.aquaculture.2003.09.045
Simeó C. G., Andrés M., Estévez A., Rotllant G. (2015). The effect of male absence on the larval production of the spider crab Maja brachydactyla Balss 1922. Aquac. Res. 46, 937–944. doi: 10.1111/are.12250
Snedecor G. W., Cochran W. G. (1989). Statistical methods. 8th edn (Ames: The Iowa State University Press).
Solanki S., Meshram S. J., Dhamagaye H. B., Naik S. D., Shingare P. E., Yadav B. M. (2023). Effect of C/N ratio levels and stocking density of catla spawn (Gibelion catla) on water quality, growth performance, and biofloc nutritional composition in an indoor biofloc system. Aquacult. Res. 2023, 2501653. doi: 10.1155/2023/2501653
Sookying D., Silva F. S. D., Davis D. A., Hanson T. R. (2011). Effects of stocking density on the performance of Pacific white shrimp Litopenaeus vannamei cultured under pond and outdoor tank conditions using a high soybean meal diet. Aquaculture 319, 232–239. doi: 10.1016/j.aquaculture.2011.06.014
Sotelo G., Morán P., Posada D. (2008). Genetic identification of the northeastern Atlantic spiny spider crab as Maja brachydactyla Balss 1922. J. Crust. Biol. 28, 76–81. doi: 10.1651/07-2875R.1
Sotelo G., Morán P., Posada D. (2009). Molecular phylogeny and biogeographic history of the European Maja spider crabs (Decapoda, Majidae). Mol. Phylogenet. Evol. 53, 314–319. doi: 10.1016/j.ympev.2009.05.009
Swiney K. M., Long W. C., Persselin S. L. (2013). The effects of holding space on juvenile red king crab, Paralithodes camtschaticus (Tilesius 1815), growth and survival. Aquac. Res. 44, 1007–1016. doi: 10.1111/j.1365-2109.2012.03105.x
Therneau T. M. (2015). A Package for Survival Analysis in S. Available online at: https://cran.r-project.org/package=survival (Accessed December 17, 2024).
Therneau T. M., Grambsch P. M. (2000). Modeling Survival Data: Extending the Cox Model (New York: Springer).
UNEP (1996). Acta final de la reunión de plenipotenciarios sobre los anexos del protocolo referente a las zonas especialmente protegidas y la diversidad biológica en el Mediterráneo (Spain: UNEP (OCA)/MED IG 10/4), 86.
Urcera M., Arnaiz R., Rua N., Coo A. (1993). Cultivo de la centolla Maja squinado: Influencia de la dieta en el desarrollo larvario. Actas IV Congreso Nacional Acuicult. 269–274.
Ut V. N., Le Vay L., Nghia T. T., Hong Hanh T. T. (2007). Development of nursery culture techniques for the mud crab Scylla paramamosain (Estampador). Aquac. Res. 38, 1563–1568. doi: 10.1111/j.1365-2109.2006.01608.x
Williams K. C. (2007). Nutritional requirements and feeds development for post-larval spiny lobster: a review. Aquaculture 263, 1–14. doi: 10.1016/j.aquaculture.2006.10.019
Yuan Q., Qian J., Ren Y., Zhang T., Li Z., Liu J. (2018). Effects of stocking density and water temperature on survival and growth of the juvenile Chinese mitten crab, Eriocheir sinensis, reared under laboratory conditions. Aquaculture 495, 631–636. doi: 10.1016/j.aquaculture.2018.06.029
Zhang G., Jiang X., Zhou W., Chen W., Levy T., Wu X. (2024). Stocking density affects culture performance and economic profit of adult all-female Chinese mitten crabs (Eriocheir sinensis) reared in earthen ponds. Aquaculture 581, 740352. doi: 10.1016/j.aquaculture.2023.740352
Zmora O., Findiesen A., Stubblefield J., Frenkel V., Zohar Y. (2005). Large-scale juvenile production of the blue crab Callinectes sapidus. Aquaculture 244, 129–139. doi: 10.1016/j.aquaculture.2004.11.012
Keywords: Maja squinado, stocking density, growth performance, crustacean, Mediterranean spider crab
Citation: Millot R, Debattice C, Ligorini V, Bracconi J, Gattacceca N, Crescioni A, Ronchi-Perfetti J-B, Vela A, Bastien R, Aiello A and Filippi J-J (2025) Effect of stocking density on survival and growth of post-settlement juveniles of aquaculture reared Mediterranean spider crab Maja squinado (Herbst, 1788). Front. Aquac. 4:1548270. doi: 10.3389/faquc.2025.1548270
Received: 19 December 2024; Accepted: 31 March 2025;
Published: 23 April 2025.
Edited by:
Charles Weirich, NOAA National Sea Grant Office, United StatesReviewed by:
Diego Castejón Bueno, Agência Regional para o Desenvolvimento da Investigação Tecnologia e Inovação (ARDITI), PortugalGuillermo Guerao, Independent researcher, Barcelona, Spain
Alexandria Walus, University of Florida, United States
Md. Mahmud Al Hasan, Australian National University, Australia
Copyright © 2025 Millot, Debattice, Ligorini, Bracconi, Gattacceca, Crescioni, Ronchi-Perfetti, Vela, Bastien, Aiello and Filippi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rémi Millot, bWlsbG90MDJAaG90bWFpbC5mcg==; Jean-José Filippi, ZmlsaXBwaV9qakB1bml2LWNvcnNlLmZy