 Adrián Alejandro Corrales-Hernández1
Adrián Alejandro Corrales-Hernández1 Patricia Roldán-Santiago2*
Patricia Roldán-Santiago2* Herlinda Bonilla-Jaime3
Herlinda Bonilla-Jaime3 Luis Alberto De la Cruz-Cruz4Ofelia Limón-Morales3
Luis Alberto De la Cruz-Cruz4Ofelia Limón-Morales3 Héctor Orozco-Gregorio5*Raym Pineda-Reyes6
Héctor Orozco-Gregorio5*Raym Pineda-Reyes6- 1Programa de Doctorado en Ciencias de la Producción y de la Salud Animal, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de Mexico, Mexico City, Mexico
- 2Departamento de Reproducción, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de Mexico, Avenida Universidad, Mexico City, Mexico
- 3Departamento de Biología de la Reproducción, Universidad Autónoma Metropolitana, Unidad Iztapalapa, Mexico City, Mexico
- 4Departamento de Producción Agrícola y Animal, Universidad Autónoma Metropolitana, Unidad Xochimilco, Mexico City, Mexico
- 5Ingeniería en Producción Animal, Universidad Politécnica de Francisco I. Madero, Tepatepec, Mexico
- 6Facultad de Agronomía y Veterinaria, Universidad Autónoma de San Luis Potosí, San Luis Potosí, Mexico
Several authors have noted that a sow’s parity has a significant impact not only on her own productivity but also on the performance of her piglets. Analyzing all the factors related to sows and their neonates at each birth event is an effective tool for improving management and reproductive performance on production units. Multiparous sows exhibit cortisol levels up to 38% higher than those of primiparous sows at peripartum, but the latter have prolactin levels 54-70% lower that reduce colostrum production. In addition, the weaning-to-estrous period is 3 days longer in primiparous sows, and their farrowing intervals up to 8 days longer. Farrowing performance is significantly affected, as litter size tends to increase in multiparous sows, thus extending duration. Piglets are also influenced by parity: those born to multiparous sows have higher weights, up to 2.21 kg more in total litter weight (up to 200 g/piglet). Their thermoregulation capacity is better as they show temperatures up to 1.6°C higher, greater overall vitality, and higher growth rates than the neonates of primiparous sows, which also suffer higher mortality rates. Given the obvious performance differences among sows of different parity and their offspring, swine breeders must determine the precise effects of this factor. This review details the key differences related to parity in the events surrounding farrowing in sows and the early lives of their offspring.
1 Introduction
Primiparous sows (PS) have characteristics distinct from other dams on production units (Patterson and Foxcroft, 2019). These emerge when their productive performance is compared to that of multiparous sows (MS) (Ordaz-Ochoa et al., 2013). Among endocrinological changes, PS tend to have lower cortisol levels, though various factors can affect concentrations of this hormone (Hales et al., 2016). It is well known that certain environmental and novel phenomena in the first farrowing experience can intensely affect PS as, for example, alertness tends to decrease in later parities (Jarvis et al., 2001). Prolactin, a hormone linked to colostrum and milk production (Craig et al., 2019), is greatly affected by parity (Quesnel et al., 2013) as MS have higher levels and, hence, produce significantly more colostrum and milk (Yun et al., 2014).
The close relation between reproductive parameters and parity must also be considered. Galíndez and Pulido (2014) hold that farrowing intervals tend to decrease with higher parity, while weaning-to-estrous intervals are typically longer in primiparous sows (Ordaz-Ochoa et al., 2013). Performance at farrowing is clearly influenced by parity, with litter size, the number of liveborn piglets, birthweight, farrowing duration, and birth interval all increasing with higher parity. Age is also a main factor that differentiates the reproductive parameters of sows. Primiparous dams are still growing, so they have high energy requirements as they reach physiological maturity in terms of body weight, body tissue composition, structural solidity, and productive development, compounded by the physiological demands of gestation and subsequent lactation (Menegat and Tokach, 2021). Multiparous sows, in contrast, have larger energy reserves, so they are better adapted to these conditions.
Sow parity also affects piglet performance. Neonates of 1st-parity sows may be as much as 8% lighter than their those of MS (Ruediger and Schulze, 2012; Roldán-Santiago et al., 2019a), while initial body temperatures of newborns can vary by almost 1°C (Roldán-Santiago et al., 2019a; Schild et al., 2020). These factors affect piglet vitality –higher in the litters of MS (Roldán-Santiago et al., 2019a)– and the ability of neonates from PS to adapt to the extrauterine environment, since they present 16% less colostrum intake, lower daily weight gain (DWG) (Rincón-Gainza et al., 2014; Zotti et al., 2017), and higher mortality (Schodl et al., 2019). In light of the foregoing, ensuring the productive longevity of sows in swine production demands attending to the differences between primiparous and multiparous dams (Roongsitthichai and Olanratmanee, 2021) by evaluating distinct management strategies based on parity (Muns et al., 2015). The objective of the present review is to identify and analyze the most important differences between PS and MS in relation to the effect of parity during the peripartum period and its impact on neonate performance.
2 The farrowing process
2.1 The endocrinology of parturition associated with sow parity
As outlined above, at the end of gestation, just before partus, sows experience a cascade of hormonal changes that are responsible for activating various mechanisms before and during delivery, and in preparation for the subsequent phase of lactation (Devillers et al., 2004; Quesnel and Farmer, 2019; Wahner and Fisher, 2005; Walls et al., 2022). The increase in the secretion of the hormone cortisol by fetuses initiates the endocrinal changes that trigger parturition as higher levels in the sow redirect endometrial PGF2α to the uterine vein and, from there, to the bloodstream. Cortisol is also known to be a biomarker of stress (Walls et al., 2022). Several authors have pointed out that hormonal variations occur due to the effect of the sow’s parity (Table 1) (Farmer et al., 1995; Yin et al., 2016; Ison et al., 2018). Hales et al.’s (2016) comparison of 1st- and 2nd-parity dams reported that salivary cortisol levels were associated with parity since, in general, the 2nd-parity group had higher levels than the primiparous dams at the onset of farrowing and 24 h later (45 and 43 nmol/l, respectively, vs. 28 and 30). These results are consistent with those in Ison et al. (2018), who found that salivary cortisol concentrations varied on the day of delivery depending on sow parity, with PS having lower levels than MS (3.4 and 4.6 ng/ml, respectively, P < 0.05). In this regard, Hales et al. (2016) suggest that the lower amount of cortisol that PS produce may be associated with smaller litter sizes compared to MS. Moreover, the larger litters produced by the latter imply a longer farrowing duration that extends the period when cortisol levels remain high. Finally, the impact of stressors may be distinct for PS since they are unfamiliar with the farrowing environment and many of the handling procedures involved (Roelofs et al., 2019). In a related finding, levels of both alertness and absolute cortisol decreased in sows with previous farrowing experience (Jarvis et al., 2001).

Table 1. Effects of parity on the physiological and performance variables of sows.
Progesterone is the hormone responsible for preventing contractions and maintaining pregnancy, but after luteolysis it decreases abruptly and its remnants are transformed into estrogens. This eliminates the myometrial block and allows contractions to begin under the influence of estrogens (Walls et al., 2022). Miller et al. (2004) fed pregnant sows diets with two energy levels (maintenance energy at 460 kJ DE/kg BW0.75 vs. double that amount) from day 100 of gestation to farrowing in sows of parities 1, 2 and 3. Progesterone concentrations were assessed at farrowing –obtaining values of 8.4 ± 0.88, 7.8 ± 0.65, and 9.0 ± 0.93 ng/mL, respectively (P>0.05)– and then at 6 hours postpartum with results of 5.8 ± 0.67, 5.3 ± 0.53, and 7.0 ± 0.72 ng/mL, respectively (P>0.05). There were no significant differences due to parity in the sows evaluated, a finding similar to that in the study by Quesnel et al. (2013), who administered a formulated lactation diet (9.11 MJ NE kg-1, 17.8% crude protein, 0.8% lysine, and 4.4% crude fiber) to sows of parities 1-5 during the week prior to farrowing. They recorded an abrupt decrease in progesterone between the day before farrowing and the day after in all study groups (10.46 ± 0.49, 6.6 ± 0.35, and 1.41 ± 0.12 ng mL-1 on day -1, day 0, and day 1, respectively), though no significant differences appeared due to parity.
Prolactin has been linked to both nest-forming behavior prior to farrowing and stimulation of udder development in preparation for lactation (Farmer, 2022; Walls et al., 2022). Yun et al. (2013) studied serum prolactin levels in PS and MS in the peripartum and postpartum periods (days -3, -2, -1, +1, +2, +4, and +7) under distinct housing and nesting material conditions (cage+1 sawdust bucket, pen+1 sawdust bucket, and pen+2 sawdust buckets with other materials). Regardless of the housing type, prolactin concentrations were affected by parity: the authors found that the PS had 18 pg/ml on the day before farrowing and peaked at ~20 pg/ml on day 1 postpartum, whereas the MS had readings of 37 pg/ml on the day before farrowing and 43 pg/ml on day 1 postpartum (P < 0.01). Clearly, the latter maintained almost double the prolactin level at all measuring intervals, from the day before farrowing to day 7 postpartum.
Higher concentrations of this hormone may positively affect oxytocin levels prior to delivery and during postpartum. Other authors have found that oxytocin plays an important role in regulating prolactin secretion in various species (Samson et al., 1986; Bar-Pelled et al., 1995). During farrowing, this hormone stimulates myometrial contractions for fetus expulsion and stimulates milk letdown (Farmer, 2022; Walls et al., 2022). In this regard, Yun et al. (2014) studied the influence of housing (crate vs. pen) and nesting materials (sawdust, shredded newspaper, branches, chopped straw, sisal rope) in sows of parities 1, 2, and 3-4 on oxytocin and prolactin levels prior to farrowing. Regardless of the housing conditions, they found that the PS had an average prolactin level of 14.8 ng/ml, 2nd-parity sows 18.6 ng/ml, and 3rd- and 4th-parity sows 22.5 ng/ml (P=0.007). Results for oxytocin, irrespective of housing, were 18.3 pg/ml for the PS, 20.3 pg/ml for the 2nd-parity sows, and 23.9 pg/ml for the 3rd- and 4th-parity mothers (P=0.078). The authors concluded that, regardless of the housing environment, parity affected both oxytocin and prolactin levels and confirmed that the former increase with parity while claiming that MS tend to have higher oxytocin concentrations.
Quesnel et al. (2013) also described the effect of parity on prolactin, comparing sows from parities 1-5 over three days, beginning on the day prior to farrowing. Levels increased on that day, reaching 35, 40, 52.5, 61.5, and 58.5 ng/mL-1, respectively. This hormone reached its maximum concentration on the day of farrowing (38, 70, 103, 80, and 127 ng/mL-1, respectively), but decreased in all groups the following day (36, 52.5, 69.5, 60.5, and 104 ng/mL-1, respectively). These results demonstrate that marked differences occurred in the prolactin levels of the five study groups on all three days (P<0.001). These findings show that older sows had higher prolactin concentrations in all three phases: peripartum, parturition, and postpartum. Higher parity was also related to adult sows with better udder development, a more mature hormonal axis, and previous farrowing experience. In contrast, the younger sows required more energy resources to trigger colostrogenesis and, subsequently, lactogenesis, because they were still growing (Menegat and Tokach, 2021). Fully adult sows exhibited greater mammary development and had larger energy reserves for milk production, so they were better adapted for initiating and maintaining lactation (Nuntapaitoon et al., 2019; Tospitakkul et al., 2019). But parity also influenced litter size (Santos et al., 2015), which in turn affected the duration of parturition. Larger litters may stimulate more active suckling by piglets and, since the neonates of MS tend to exhibit greater vitality (Roldán-Santiago et al., 2019a), could lead to more effective stimulation of the udder, thus promoting higher prolactin and oxytocin levels and facilitating milk production.
PS and older sows (those with parities >6) have a higher risk of experiencing poor farrowing, which can compromise their health and increase the danger of postpartum dysgalactia syndrome (Walls et al., 2022). In sows with this condition, cortisol levels increase from 36 h prepartum to 36 h postpartum (Kaiser et al., 2018), possibly an early indication of inflammation or stress, while their prolactin concentrations are lower (Maes et al., 2010). Primiparous sows with large litters that require human intervention at farrowing have a higher risk of developing this syndrome (Gerjets et al., 2011; Bardehle et al., 2012). In relation to this, differences in hormone levels due to parity can affect the performance of sows. Unlike progesterone, which is not influenced by the number of births, prolactin shows marked differences that affect colostrum and milk production in PS. Handlers must be aware of this disadvantage in cases of PS with large litters, as they must consume more food due to the greater wear and tear they suffer (Kim and Easter, 2001).
2.2 Reproductive parameters of sows
The main parameters used to determine reproductive capacity in sows include gestation length, weaning interval, and the return of estrus. Gestation length spans the days from the first insemination during estrus to the farrowing date (Sasaki and Koketsu, 2007; Rydhmer et al., 2008), while the weaning-to-estrus interval is the number of days a sow requires to present signs of heat after weaning (Sasaki and Koketsu, 2007). Significant differences in these reproductive parameters associated with sow parity have been reported. According to Smith et al. (2013), who induced farrowing in nulliparous and primiparous sows on days 113, 114, and 116 of gestation, gilts typically had shorter gestation periods than 2nd-parity sows, as a higher proportion gave birth spontaneously on days 113 (9 vs. 5%) and 114 (23 vs. 9.5%), though this pattern was reversed on day 116 (75 vs. 87%). These results may suggest that the effects of stress during gestation are more pronounced in nulliparous sows, possibly due to constraints on behavior during gestation and because farrowing is a novel event for them (Estienne and Harper, 2010). Yang et al. (2019) found significant differences (P <0.05) in the duration of gestation by parity, as they presented the following results: PS, 115.4 ± 1.3 d; middle-aged sows (parities 2-5), 115.5 ± 1.2 d; and old sows (parities 6-9), 116.7 ± 0.9 d. They concluded that the higher the parity, the lower the impact of stress on reproductive performance.
In an effort to develop a scale to score the ease of farrowing, Mainau et al. (2010) assessed PS and MS (parities 2-7) in relation to duration, postural changes, activity levels, and the presentation of the piglets at expulsion and after birth. They found that parity had no effect on the duration or ease of farrowing, as average times of 221.87 min and inter-piglet intervals of 16.96 min were recorded. This indicates that PS will not necessarily have more difficult farrowings than MS, but contradicts the findings of Wülbers-Mindermann et al. (2002) and Cronin et al. (1993), who reported that parity significantly affects farrowing duration. Here, it is important to consider that PS tend to have smaller litters (Santos et al., 2015), so this may reduce farrowing duration. Although prolonged farrowings may reflect difficult births, various factors can increase duration, such as body condition, housing, gestation length, and the sow’s age, genetics, and parity (Oliviero et al., 2010; Adi et al., 2022).
It is generally accepted that the weaning-estrus interval should be in a range of 5.5-7.5 days (Knox and Zas, 2001; Bolado et al., 2011; Pérez-Sánchez et al., 2015; Yatabe et al., 2019). The return of estrus is influenced not only by parity (Malavé et al., 2007), but also by feed intake, the intensity of lactation, and the consumption of body reserves, key factors that are more intense in 1st- and 2nd-parity sows (Ordaz-Ochoa et al., 2013). The former must distribute their energies and the nutrients they consume among their requirements for growth, reproduction, gestation, and lactation, but MS no longer need to invest energy in growth (Malavé et al., 2007; Leite et al., 2011). Primiparous sows exhibit longer anestrus, but this tendency decreases with higher parity (Pérez-Sánchez et al., 2015). Santos et al. (2015) evaluated the parity of sows selected for the integrity of locomotion, udder morphology, and body condition (3-4 on a scale of 5) with the supplementation of coconut oil to their piglets. Their comparison of the sows’ parity (1-7) revealed that the 1st- and 2nd-parity dams took longer to re-enter the estrous cycle than the 3rd-parity sows (7-10 vs. 5.6 d). Moeller et al. (2004) reported similar results when comparing 1st-4th-parity sows from six different genetic lines, as they found that this interval was shorter in the sows of parities 2 and 3 (8.2-10 vs. 8.5-11.4 d) than in the PS (11.2-15.9 d). Mendoza and Ortega (2009) observed a similar effect, as they reported higher average weaning-to-estrus intervals for PS (12.2 d) and a decrease with higher parity (7.0, 6.1, 6.1, 5.4 and 5.3 d, respectively, for parities 2-6).
In their work, Ordaz-Ochoa et al. (2013) determined that PS had longer weaning-to-estrus interval (8.8 d) than 1st-to-5th-parity dams, and that the latter had the shortest interval (6.7, 6.4, 6.2, and 5.8 d, respectively). Smith et al. (2013), however, reported that the percentage of sows that entered estrus on day 7 post-weaning, and the probability of a subsequent pregnancy, was not affected in primiparous vs. 2nd-parity dams. Their findings concur with Rincón-Gainza et al.’s (2014), comparison of 1st- and 2nd-parity sows, which found no significant variation in this interval due to parity. They also showed that if the loss of weight and backfat during lactation is not severe, the return to estrus post-weaning will not be delayed. Other authors also affirm that the late presentation of post-weaning estrus in PS is associated with significant weight loss and deficient nutrition during lactation (Karvelienė et al., 2008; Han et al., 2020). One such finding is that PS whose backfat thickness is below 18 mm at weaning will take longer to present estrus and will have lower conception rates (Murillo-Galán et al., 2007; Škorjanc et al., 2008).Therefore, backfat thickness is considered a main factor that affect the reproductive performance of sows, especially in relation to litter size (Tummaruk et al., 2007). In this regard, Roongsitthichai and Olanratmanee (2021) evaluated fetal mortality in PS in relation to backfat during insemination and farrowing. Their results showed that the number of stillborn piglets appeared to be proportional to backfat thickness at farrowing, as their study revealed a higher number of stillbirths (9.1%) in sows with more backfat (>21 mm) compared to those with less (≤15 mm, 0.2% stillbirths). They hypothesized that the birth canals of sows with thicker backfat may have more layers of adipose tissue that could form a physical impediment to the expulsion of their piglets.
The length of gestation and the weaning-to-return to estrus interval are highly important parameters in swine production units, as they contribute to determining the productivity of sows. In the case of PS, these times are modified by the fact that they are experiencing their first parity, a condition that may be deemed disadvantageous and require special handling, since inadequate management and the failure to monitor backfat thickness could lead to an increase in the number of stillbirths and in the time required to return to estrus, two factors that reduce the productivity of sows.
2.3 Sows’ behavior at farrowing
Regarding sows’ behavior during farrowing, studies show that both multiparous and primiparous mothers housed in farrowing crates show constant changes in posture. They remain standing or sitting for varying periods of time, but in the 6 hours prior to farrowing they perform the greatest number of postural changes (Mainau et al., 2009; Girardie et al., 2023). Sows in farrowing crates show behavioral patterns similar to those of nest-building (Lammers and De Lange, 1986; Yun et al., 2013; Yin et al., 2016; Rosvold et al., 2019). On the day after farrowing, however, they remain lying laterally 89% of the time (Hötzel et al., 2004; Girardie et al., 2023). Mainau et al. (2010) found that PS had more postural changes (lying, sitting, standing) than MS (106.8 ± 9.8 vs. 96.7 ± 5.5 movements) on the day before farrowing, but 120.6 ± 8 vs. 95.1 ± 6.9 on the day of farrowing. In contrast, Mainau et al. (2009) reported that the proportion of time that sows remained standing, sitting, and lying down, and the frequency of their postural changes, were not affected by parity. This latter finding is consistent with the study by Jarvis et al. (2001), who followed gilts through their first and second gestations. They found no effect of parity on the time spent in each posture but added that there was a tendency for the number of postural changes to decrease with higher parity.
Jarvis et al. (1997) also described behavioral changes related to the postures of gilts 24 h prior to farrowing. They observed that at 16 h before parturition, the sows remained standing for longer periods, and that this reached its maximum level 8 h before farrowing. The lateral lying posture decreased around 16 h prior to farrowing and was at its lowest point 8 h before birthing began. Both the ventral lying and sitting postures varied depending on the time relative to the onset of farrowing. Those authors also mentioned that, regardless of the environment, gilts increased the proportion of time they remained standing mainly during the period that coincided with the performance of nest-building behavior, a conduct that is altered of no material for nest construction is provided. In contrast, the sows housed in cages increased the proportion of time spent sitting (Plush et al., 2021).
Cronin et al. (1993) observed increased nest-building behaviors 24 h prior to farrowing –with notable increases at 12 h pre-farrowing– that included pawing, rooting, and nosing at the floor and cage bars. In that study, the sows generally remained lying laterally throughout farrowing, though some –clearly nervous– dams tended to stand frequently (Randall, 1972; Jarvis et al., 1997). Jarvis et al. (2001) monitored sows during their first and second gestations to compare different types of housing in relation to behavior and plasma cortisol levels. Although they found no significant differences in parity related to the time sows spent in different postures, they did report decreased alertness in the 2nd-parity sows. which they attributed to habituation to the farrowing crates. One indication of this was a pattern of lower absolute cortisol levels. Therefore, previous experience of farrowing in a crate seemed to reduce physiological stress during the second farrowing. It is necessary, however, to consider factors that may influence cortisol levels, such as other housing conditions (Hales et al., 2016) and the changes that occur with higher parity, as these can improve reproductive parameters like litter size (Santos et al., 2015) while also prolonging farrowing (Ju et al., 2022), which affects cortisol levels. In this vein, Thorsen et al. (2017) evaluated sows of parities 1-6 (grouped as 1-2, 3-4, and >4), but found no effect (P=0.614) on the number of postural changes post-delivery.
Some studies of swine suggest that tail-wagging is a common indicator of positive emotions in sows (e.g., Reimert et al., 2013). Yang et al. (2019), however, indicated that tail-wagging at farrowing is a significant indicator of pain. Their experiment determined a negative correlation independent of parity number, with values of r = –0.441 for PS, r = –0.428 for sows from parities 2-5, and r = –0.832 for parities 6-9. Those results suggest that the frequency of tail-wagging increases around the moment of the expulsion of the first piglet but decreases considerably after the last one is expelled. The sows with 6-9 parities exhibited more tail movements before expelling the first piglet (24 with an average of 7.8 ± 6.7 per piglet) followed by the PS (16 with an average of 5.3 ± 4.3), and those from parities 2-5 (9 with an average of 5.6 ± 2.3). These figures indicate that sows with very high parity, along with PS, experience greater pain than dams with 2-5 parities.
It is essential to provide sows with an adequate environment to prevent disruptions in the farrowing process. This is especially important for gilts, whose first experience with farrowing should not be negative. One of the differences between gilts and MS is that the former show greater reactivity because farrowing is a novel event. The number of postural changes and tail movements may reflect discomfort, so all sows must be manipulated gently before farrowing, and handlers must take special care with gilts by, for example, providing materials that allow them to engage in nest-building behavior whenever possible.
2.4 Between-farrowing interval
The farrowing interval (FI) is a critical reproductive parameter, as it represents the number of days that elapse from one farrowing to the next and, hence, the quality of farm management. It spans the lactation, weaning-service interval, and days of gestation (Galíndez and Pulido, 2014; Wu et al., 2019). Studies related to parity suggest that primiparous sows may have longer FIs than multiparous ones (Tantasuparuk et al., 2000; Pandey et al., 2010). Galíndez and Pulido’s (2014) cross-sectional study compared sows with parities 1-5 and ≥6 in two periods. They found that FI tended to decrease as the sow enters full adulthood but does not present a constant tendency (153, 150, 148, 148, 148, and 145 days, respectively, for parities 1-5 and ≥6). This coincides with the report by Koketsu (2005) on his cross-sectional research into the characterization of reproductive performance according to sow parity. Koketsu found longer FIs in sows from 1st-to-2nd parity (152 days) compared to those in the ≥6th-parity group (144 days). However, not all authors agree. Van Dijk et al. (2005), for example, evaluated the length of the expulsion stage and birth interval in piglets from sows of different breeds, reaching the conclusion that FI was not affected by parity.
Returning to the evidence for reduced FI with higher parity, this relation has been attributed to the subsequent development that sows achieve due to a better distribution of nutrients, especially since PS show greater mobilization of fat reserves during lactation (Ek-Mex et al., 2014; Rempel et al., 2022). As mentioned above, primiparous sows must distribute their energy and nutrients among their requirements for growth, reproduction, gestation, and lactation, while multiparous sows no longer invest energy in growth (Malavé et al., 2007; Leite et al., 2011). In addition, PS tend to have longer weaning-to-estrus intervals because they are more susceptible to reproductive failures (e.g., anestrus, repeated estrus, abortion). This can be explained by their immature endocrine system, since they are young and still growing (Ortíz et al., 2004; Rempel et al., 2022). Another factor involved is low feed intake during lactation, as this decreases the secretion of gonadotropins (Koketsu et al., 1996; Butler, 2005), which restricts follicular growth in their ovaries, consequently increasing FI. We cannot ignore findings which suggest that reproductive performance improves as parity increases, apparently peaking between parities 2-5, before declining (Klimas et al., 2020) in older sows that may have decreased ovulation, lower fertilization rates, and higher more stillbirths (Koketsu et al., 2017).
Because PS are young animals that are still growing, they require different handling to achieve their maximum productive potential, especially in terms of nutrition. The feed provided must be optimal so they can maintain high productivity while they continue to grow. This will help prevent the kinds of reproductive failures –caused by the wear and tear that lactation can cause through consumption of body reserves– that occur much less frequently in MS.
2.5 Farrowing duration
Parturition is defined as a three-stage process: the first stage refers to the ripening of the cervix and the initial myometrial contractions to position the fetus in the birth canal; the second is characterized by the onset of strong abdominal contractions, rupture of the allantochorionic sac, and expulsion of the fetuses; the third involves expulsion of the residual fetal membranes (Walls et al., 2022). For productive purposes, the parameter farrowing duration begins in stage 2 with the expulsion of the first fetus, and ends with that of the last piglet (Zaremba et al., 2015). Average duration can vary from 156 to 262 min, but if it lasts more than 300 min it is considered prolonged (Oliviero et al., 2010; Liu et al., 2021), a condition that can increase the mortality rate of piglets (Fraser et al., 1997). Björkman et al. (2017) reported that farrowing duration correlated positively with the number of stillbirths and parity (P = 0.009). In their study, sows with 0 stillbirths had a farrowing duration of 307 ± 141 min, whereas those with 1 stillbirth had an average duration of 437 ± 275 min. In the case of sows with 2 stillbirths, this was 487 ± 219 min; with 3, 429 ± 229 min; with 4, 445 ± 221 min; and with 5, 696 ± 548 min. Farrowing duration can, however, be influenced by various factors, including breed, the age of the sow, gestation length, litter size, and housing and body conditions (van Rens and van der Lende, 2004; Van Dijk et al., 2005; Oliviero et al., 2010).
On the topic of farrowing duration, distinct average times for the between-birth interval of individual piglets have been established by different authors. This interval can be classified as very short –fewer than 5 min–; long, over 20 min (van Rens and van der Lende, 2004; Van Dijk et al., 2005), as 12-18 min (Alonso-Spilsbury et al., 2004), or as 15-28 minutes (van Rens and van der Lende, 2004). In this regard, van Rens and van der Lende (2004) reported that the first piglets expelled have longer birth intervals, while expulsion of the second and final fetuses may take longer in PS (2-28 and 2-150 minutes, respectively).
To study the effect of farrowing duration on the expulsion of the placenta, Björkman et al. (2017) compared sows from the 2nd-to-5th farrowings. They found significant differences (P=0.008) for the effect of parity, as they recorded the following times: 2nd farrowing, 273 ± 147 min; 3rd, 366 ± 183 min; 4th, 377 ± 219 min; and 5th, 481 ± 278 min. Tummaruk and Sang-Gassanee (2013), in turn, suggested that PS have a shorter average farrowing duration (141.2 ± 18.6 min) than sows of parities 2 to 4 (199.2 ± 17.1 min, P=0.025), but that this does not differ significantly from that of sows of parities 5-7 (179.9 ± 16.2 min, P=0.123). They recognized that their results were affected by the litter size in each group of sows. In this sense, some authors claim that high parities result in longer farrowing (Farmer and Robert, 2003), though others do not suggest any significant effect (Van Dijk et al., 2005; Oliviero et al., 2009; Mainau et al., 2010). van Rens and van der Lende (2004) performed a study to evaluate farrowing duration, body weight, inter-expulsion interval, birth order of the piglets, and delay in expelling the placenta in Large White × Meishan F2 PS. They reported an average duration of 130 ± 89 min with between-piglet intervals of 14.9 ± 9.8 min. Oliviero et al. (2010) and Nam and Sukon (2020) reported that parity has no statistically significant effect on farrowing duration.
In contrast, Yang et al.’s (2019) evaluation of the reproductive performance of sows of different parities found a shorter average between-piglet interval in older sows (22.2 min, parities 6-9) than middle-aged (29.5 min, parities 2-5), and young sows (25.9 min, primiparous) The study by Hales et al. (2015), however, which evaluated the effect of confinement of sows from parities 1-7 on farrowing duration, found that the birth interval was shorter in sows of parities 1-2 than in those of parity 3 or greater, irrespective of housing conditions (20 vs. 24 min. p < 0.09). Van Dijk et al. (2005), meanwhile, analyzed the length of the expulsion stage and the birth interval in piglets from sows of different breeds. They reported that the between-birth interval was not affected by parity, but did show an effect of breed.
In a study designed to determine the effect of parity on (i) the total number of piglets born; (ii) the number of liveborn piglets; and (iii) the number of stillbirths, Ju et al. (2022) analyzed 32,231 farrowings from 8,420 sows of parities 1-10. They reported that PS had a shorter average farrowing duration (212.65 ± 61.90) than the MS (243.01 ± 67.68, 247.23 ± 67.23, 250.23 ± 65.84, 251.10 ± 67.23, 252.01 ± 68.35, and 260.40 ± 68.02 min for parities 2-7, respectively, p < 0.05), and added that the average duration of parturition was 4.2 h, though it increased with litter size. They concluded that both litter size and parity influence the duration of parturition. Although as these studies show, the length of farrowing varies amply, verifying the range of the between-piglet interval and the total length of farrowing, as well as considering all the factors involved –including environmental and housing conditions and behavior– can help ensure that timely care is provided when complications arise during farrowing in primiparous sows.
2.6 Litter size
In pigs, the factor of litter size refers to the total number of piglets at birth, including both liveborn neonates and stillborns (Casellas et al., 2004; Rutherford et al., 2013). This parameter can be categorized in four main groups: total piglets born (TB); liveborn piglets (BA); mummified fetuses (MF); and stillbirths (SB) (Roongsitthichai and Olanratmanee, 2021). Genetic improvements in modern swine production have focused on increasing litter size (Rutherford et al., 2013), so today some hyperprolific sows can produce up to 22 piglets per litter (Kobek-Kjeldager et al., 2023). Litter size is now a key indicator of reproductive performance in sows (Roongsitthichai and Olanratmanee, 2021), though larger litters can generate problems such as longer farrowing duration, placental retention, greater weight variation in piglets, low birthweights, and more stillbirths (Björkman et al., 2017). As mentioned previously, Hales et al. (2015) evaluated the effect of confinement on sows from parity 1-7 in relation to the progress of farrowing, finding an effect of parity on piglet mortality, as the sows of parities 1-2 had lower indices than older sows, irrespective of housing conditions (4.27 ± 0.77 vs. 10.9 ± 70.86, P < 0.001). These results agree with those of García-González et al. (2011) in their assessment of the effects of parity on piglet mortality. Their results were 9.22 TB and 8.3 BA for parity 1; 9.43 TB and 8.97 BA for parity 2; and 10.65 TB and 10.05 BA for parity 4. Tummaruk and Sang-Gassanee (2013) also compared sows of different parities. They determined that PS had fewer TB (9.6 ± 0.6) than MS (11.3 ± 0.7 TB for parities 2-4, 11.6 ± 0.6 TB for parities 5-7). Their conclusion was that litter size increases with parity. Some authors, however, do not agree. For example, Yang et al. (2019), who evaluated the reproductive performance of sows of different parities, observed numerically larger litters in 1st-parity sows (16.8 ± 1.8) than in MS (15.5 ± 3.1 for parities 2-5, 12.4 ± 3.2 for parities 6-9), though the effect they calculated did not reach the level of significance (P= 0.113). They further found that the BA piglets tended to be smaller in 1st- and 2nd-parity sows (Quesnel et al., 2008; Tummaruk and Sang-Gassanee, 2013). Regarding stillborn piglets, García-González et al. (2011) found that there is a greater tendency to present stillbirths in sows from parities 1 and 6 (0.92 and 0.94 SB, respectively) compared to mothers from the parities 2-5 (0.47, 0.60, 0.64, 0.61, respectively, P<0.05).
Rincón-Gainza et al.’s (2014) work assessed the reproductive performance of purebred Yorkshire sows and Landrace × Yorkshire crossbreds in four parity groups: 1, 2, 3, and ≥4. Although they did not find significant differences in BA, MF, or SB with respect to litter size (P>0.05), regardless of breed, they did document an effect of parity on the birthweight of the litters (P<0.05), which increased with higher parity. In their study, the litters from 3rd-parity sows were heavier than those of the PS (16.19 ± 0.56, 17.4 ± 0.65, 17.95 ± 0.65, and 18.4 ± 0.64 kg for sows with parities 1-4, respectively). In their research, Santos et al. (2015) evaluated the effects of parity and dietary supplementation with coconut oil. Results showed that sows with higher parity had larger litters (13.62 ± 2.09, 10.25 ± 4.74, 12.25 ± 5.51, 15.0 ± 3.75, 15.0 ± 6.8, 13.5 ± 6.96, and 14.70 ± 2.72 piglets per litter for parities 1-7, respectively), but that the average birthweight of the piglets decreased as their number per litter increased (1.29 ± 0.2, 1.63 ± 0.74, 1.53 ± 0.68, 1.38 ± 0.34, 1.26 ± 0.58, 1.30 ± 0.65, and 1.17 ± 0.16 kg for the sows of parities 1-7, respectively). In addition, they determined that this effect could be exacerbated in hyperprolific sows due to intrauterine growth retardation, which has negative effects on growth performance due to dysfunctions in intestinal morphology (Tang and Xiong, 2022). In this regard, they suggested that the increase in litter size between primiparous and 1st-parity sows could be associated with higher ovulation rates, greater uterine capacity, and the age of the sows (Suriyasomboon et al., 2006; Foxcroft, 2007).
Roongsitthichai and Olanratmanee (2021), who studied the backfat thickness of nulliparous sows during service and farrowing, related fetal mortality to this factor in both nulliparous and primiparous sows. In light of their findings, they recommended that special attention be paid to backfat thickness in these females at the time of mating or service, since sows with thickness ≤12.0 mm produced more MF of non-infectious origin (13.8 ± 4.5 MF). Their experiment generated values of 10.5 ± 2.6 for TB, 9.5 ± 2.7 for BA, 4.0 ± 0.7% for SB, and 4.1 ± 0.9% for MF.
As this research suggests, sows’ productivity is strongly influenced by parity, with PS having lower reproductive performance, including shorter farrowing times and fewer liveborn piglets than sows from parities 2-5 (Koketsu et al., 2017). A second conclusion is that reproductive performance improves with higher parity, reaching its peak between parities 2 and 5, when it begins to decline (Klimas et al., 2020). Reproductive failures, health disorders, and physical defects are among the main reasons for culling sows (Bondoc and Isubol, 2022). According to the data reported to date, the litters of PS are typically smaller than those of MS, so it is essential to manage their feeding carefully before, during, and after gestation to prevent problems such as SB and MF neonates and ensure their productive longevity for subsequent parturitions.
2.7 Colostrum production
Colostrum is the first lacteal secretion from the sow’s udder after farrowing, but it begins to be produced once its components are synthesized and immunoglobulins are transferred into the lacteal secretion. Constituent elements like lactose begin to form approximately 4 days before farrowing (Hartmann et al., 1984). Colostrum secretion is constant during the first 2-4 h after the onset of farrowing, but decreases in frequency until 12-36 h postpartum (Fraser and Rushen, 1992; Quesnel et al., 2012) as production transits toward normal milk (Theil et al., 2014). Ferrari et al. (2014) explored the factors that influence colostrum production by sows and intake by piglets and evaluated colostrum quality in PS and MS (parities 2-6) and the effect of colostrum ingestion on the performance of their piglets. They found that 1st-parity sows produced less colostrum than MS (3.0 ± 0.13 vs. 3.5 ± 0.12 kg, P=0.0058). Devillers et al. (2007) studied the differences in colostrum production of sows in three parity groups: 1, 2-3, and 4-6. They also observed that PS had lower production (3,435 ± 184 g). They suggest that sows from parities 4 to 6 produce greater quantities of colostrum (3616 ± 288 g) than PS, while those from parities 2 and 3 produce more (4,278 ± 288 g) than those from parities 4 to 6 (P= 0.059). Nuntapaitoon et al. (2019) studied colostrum production at 1 and 6 h postpartum in sows of parities 1, 2, 3, and 4-7. They concurred that PS produce less than sows of parities 2 and 3 (4,532 ± 225, 5,372 ± 331, and 5,505 ± 331 g, respectively, P < 0.05), but that no differences appeared in the comparison of the sows of parities 4-7 (4936 ± 214 g, P = 0.198). Their results concur with those in Tospitakkul et al. (2019), who compared PS to MS (parity 1 vs. parities 2-7) by estimating colostrum production per ingestion by individual piglets. Their comparison showed that PS had lower production than MS (2,772 ± 199.1 vs. 3,667 ± 180.8 g, respectively; P<0.001).
Beyer et al. (2007) observed the same effect in their work with sows of parities 1, 2, and 4, where they measured milk production by weighing the litter before and after suckling, and evaluated colostrum quality during three intervals: 1-6, 7-12, and 13-24 h. These authors reported that the PS produced 15% less energy and milk than the 2nd-parity mothers, whereas those of parity 4 produced 6% more milk and 7-10% more protein and lactose (P < 0.005). While colostrum yield can be influenced by certain characteristics of the litter –like vitality at birth– it is not highly-determined by litter size or the intensity of suckling but, rather, predominantly by factors related to the sow (Quesnel and Farmer, 2019), such as endocrine status; specifically, the relative levels of prolactin and progesterone during the prepartum period (Foisnet et al., 2010), as Miller et al. (2004) and Quesnel et al. (2013) have stated. Parity per se does not appear to affect progesterone levels but does affect prolactin concentrations in the peripartum phase (Quesnel et al., 2013; Yun et al., 2014). This is significant because of this hormone’s role in colostrum production (Quesnel et al., 2013).
The main components of colostrum are proteins, including the immunoglobulins IgG, IgM, and IgA. IgG is the bioactive component with the highest concentration in the first hours postpartum (Rooke and Bland, 2002; Wu et al., 2010), but decreases abruptly after 24 h (Herpin et al., 2005; Vallet et al., 2013). In PS, IgG values at the onset of farrowing are lower than in MS (Cabrera et al., 2012). Piglets from large litters born to PS are at a disadvantage in this regard, since their mothers produce less colostrum (Nuntapaitoon et al., 2019). In relation to this, Cabrera et al. (2012) explored the association among birthweight, birth order, and IgG levels by measuring IgG concentrations in sows of parities 1-8 at the onset of farrowing. They found that parity had a significant effect (P <0.001) on IgG concentrations in the colostrum, as PS produced up to 5% less than MS from farrowings 2-8. They also observed that this amount decreased more rapidly in the first 24 h. On the quality of colostrum, other authors have reported that there are no significant differences due to the effect of parity (Craig et al., 2019).
With respect to the management of the litters of PS, it is important to take into account that they produce less colostrum with lower IgG content than MS, especially as litter size increases. In these cases, using nurse sows may be a good option to ensure that the neonates receive sufficient food. Multiparous sows from parities 2 to 4 could be the best candidates for this service since they have higher colostrum production than dams of other parities. This so-called “cross-fostering” strategy may prove advantageous once the piglets have consumed the colostrum from their own mothers (Piñeiro et al., 2019). Farmer (2022) recommends employing this practice after day 3 postpartum to ensure adequate mammary development in PS.
3 Performance of neonate piglets
3.1 Birthweight
The birthweight of piglets is another important parameter because it is associated with neonate survival and growth rates (Vázquez-Gómez et al., 2020). We cannot, however, ignore the genetic advancements in swine production that have fomented greater litter size, nor the variation in birthweight within litters, especially the higher number of low-weight piglets (Beaulieu et al., 2010). Low birthweight neonates have less muscle mass, lower energy reserves, and a higher risk of death. Moreover, they take longer to reach the udder and suckle, a condition that affects their subsequent performance and survival (Le Dividich, 1999; Vodolazska et al., 2023).
Weight also influences the efficiency of thermoregulation in piglets, so it is considered a key indicator of potential survival (Muns et al., 2013). Milligan et al. (2002a) evaluated weight variation in neonates and its relation to mortality in sows of parities 1, 2, 3-5, and ≥6 by grouping them as “low-” and “high-weight” offspring. The average low weights were 1.31, 1.24, 1.14, and 1.12 kg, respectively, while the corresponding mean high weights were 1.77, 1.75, 1.64, and 1.64 kg. Results showed that the low-weight piglets had higher mortality rates, ranging from 6.9 to 18.6% above those of the heavier neonates. Meanwhile, Bierhals et al.’s (2012) study of the adaptation and performance of piglets from PS, classified piglets weighing 1-1.2 kg as “low-weight”, and those in the range of 1.4-1.6 kg as “medium weight”, while van Rens and van der Lende (2004) found average weights of 1.236 kg for the neonates of PS. There is no doubt that parity affects the birthweight of piglets (Table 2) but other factors must be considered, such as the sow’s prolificacy, since individual piglets born from high-prolificacy dams can weigh up to 43% less than those in litters from low-prolificacy mothers (Moreira et al., 2020).
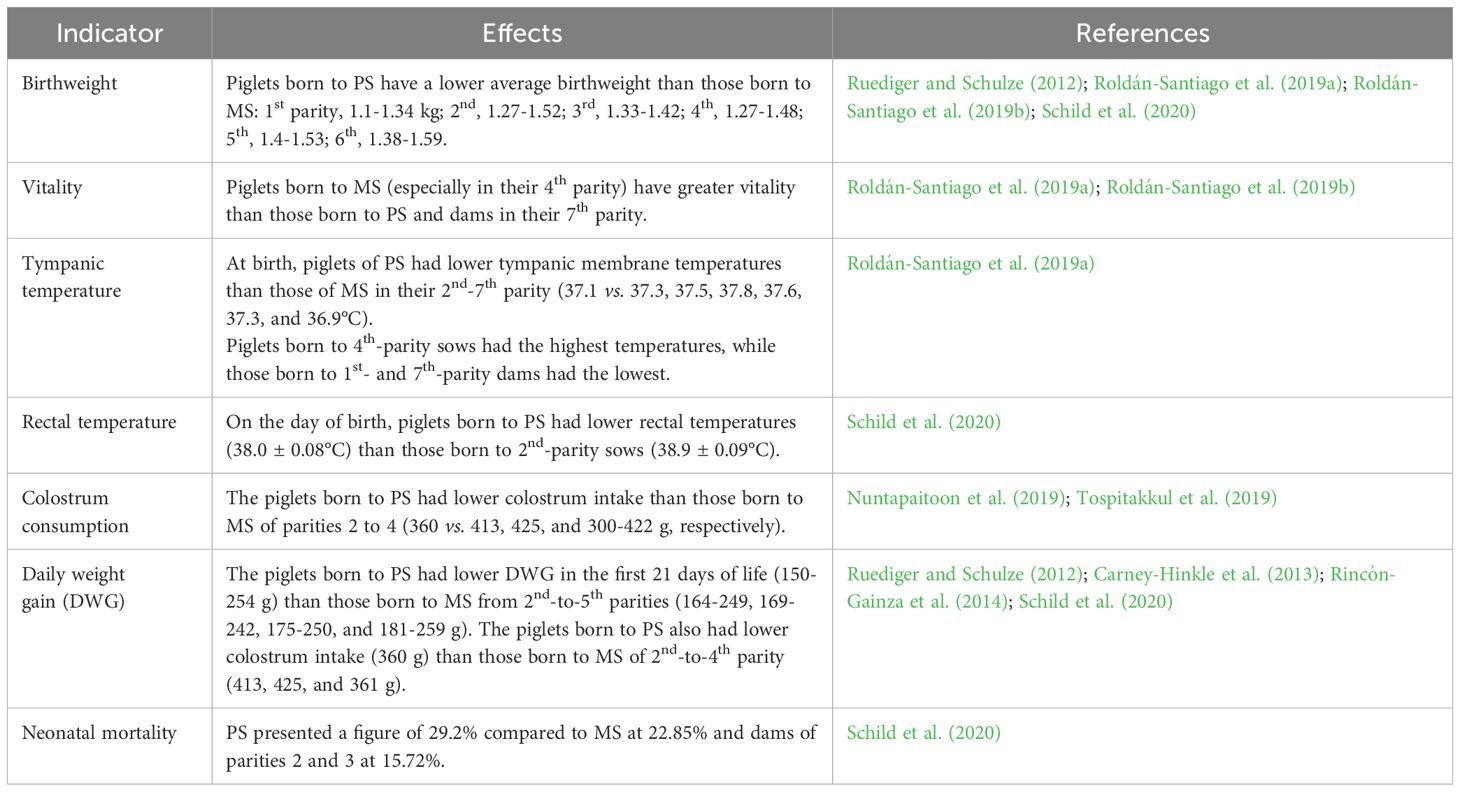
Table 2. Main differences in piglets due to the effect of sow parity.
Extensive research has demonstrated that parity impacts productive performance, particularly in terms of litter size, birthweight, and BA (Ordaz-Ochoa et al., 2013), and that combining parity with large litters affects birthweight (Kitkha et al., 2017). Carney-Hinkle et al. (2013) analyzed the effect of sow parity on breeding performance by comparing dams of parities 1 and 4. They found differences in piglet weight at birth (1.32 vs. 1.56 kg) and litter weight gain (25.7 vs. 30.0 kg on day 7; P < 0.03; 44.4 vs. 51.5 kg on day 14; P < 0.06) due to parity. According to Santos et al. (2015), piglets from large litters typically have lower individual weights, motor performance capacity, body mass, and feed intake capacity. They attributed this to overcrowding in the uterus and competition for space and resources among fetuses due to their large number (Vanden Hole et al., 2018).
Rincón-Gainza et al. (2014) evaluated the reproductive performance of purebred Yorkshire sows and Landrace × Yorkshire crossbreeds, classifying them into parities 1, 2, 3, and ≥4. They observed that the birthweight of the litters of the 1st-parity sows was 16.19 kg, lower than in for the 3rd- and 4th-parity mothers (17.95 and 18.4 kg, respectively), but similar to that of the litters born to the 2nd-parity dams (17.4 kg) (P <0.05). They added that the weight of the piglets born to PS on day 10 was lower (3.19 kg, P <0.01) than that of the 2nd- and >4th-parity sows (3.40 and 3.56 kg, respectively), possibly due to the influence of the mothers’ milk production. Although sow parity effects birthweight, it does not explain all the weight variation among piglets in the same litter, an important parameter that may reflect the influence of the sow’s body condition at the end of gestation on birthweights within the litter (Quesnel et al., 2008). Conversely, González-Hernández et al. (2002) evaluated the effects of two parameters –parity and seasonality– on breeding performance in a cross-sectional study with data from 339 farrowings of sows with parities 1-6. Their findings included that litter size is influenced by parity, as the smallest litters were produced by PS (9.52, 9.59, 10.22, 10.69, 10.96, and 10.17 for parities 1-6 respectively, P<0.05). They added that very large litters resulted in markedly low individual piglet weights due to the number of fetuses that occupied space in the uterus. It is important to mention that birthweight is also affected by the sow’s body condition at the end of gestation and her degree of prolificacy (Moreira et al., 2020).
In the work by Muns et al. (2015), who compared the individual weights of piglets from PS to those of neonates from MS (parities 2-7), we see significant differences due to the effect of parity, as the piglets from the former had the lowest weights (1.28 vs. 1.35 kg, P= 0.002). The lower weight of those piglets constitutes a disadvantage for their survival that must be mitigated by providing an adequate environment with ideal temperature conditions to minimize energy expenditures by the piglets in thermoregulation. Likewise, breeders must ensure that the piglets consume sufficient feed to attain their optimal DWG. This may entail using nurse sows if the PS do not produce enough milk.
3.2 Piglet vitality
The parameter of vitality is based on the piglet’s vigor, strength, and ability to reach the teat and suckle; that is, their capacity to adapt to extrauterine life. High vitality, then, increases the likelihood of survival (Muns et al., 2013; Tucker et al., 2021). Neonate vitality is influenced, in part, by the dam’s parity since, as we have seen, this affects litter size (Santos et al., 2015) and weight (Rincón-Gainza et al., 2014), farrowing duration (Ju et al., 2022), and the birthweights of individual piglets (Roldán-Santiago et al., 2019a). Farrowing duration, litter size, birth interval, and position in the birth order are other factors that strongly influence vitality, a condition that is known to be associated with the risk of suffering neonatal asphyxia (Herpin et al., 1996). Studies have demonstrated a relation between low neonate vitality and intrapartum hypoxia, which can damage the central nervous system of piglets (Trujillo-Ortega et al., 2007). In this regard, the lack of previous maternal experience in PS must be given special consideration since it may affect nursing behavior and the nourishment their offspring receive, thus placing their survival at risk (Li et al., 2023). Several studies have analyzed physiological, behavioral, and metabolic variables to assess piglet vitality after birth (Herpin et al., 1996; Casellas et al., 2004; Muns et al., 2013; Roldán-Santiago et al., 2019a; Schodl et al., 2019).
Roldán-Santiago et al. (2019b) assessed the effect of parity on the vitality and physiological parameters of piglets born to sows of parities 1-7, reporting that this factor definitely impacts the vitality of neonates from sows that have eutocic farrowings. They discovered that piglets from PS showed greater alterations in vitality (P < 0.05) because they required more time to reach the teat (54.5 min), 9.9% showed severe meconium staining, and 11.8% had broken cords. The piglets from the 4th-parity sows had the highest vitality (23.9 min to take the teat; 0.9% severe meconium staining; 1.8% broken cords, P < 0.05). These notable differences in vitality may well be related to the age of the dam since young sows that are still growing have narrower cervixes (Llamas-López et al., 2019), so the weight of the piglets could be an additional factor that affects the birth interval. In later work, Roldán-Santiago et al. (2019a) examined the relation among birthweight, vitality, and temperature in piglets born to sows of parities 1, 2-5, and 6-7. Findings showed that the newborns from PS and MS had distinct weights. They concluded that a relation between birthweight and vitality exists since they found that low-vitality piglets from PS weighed less (1393.9 ± 11.86 g, P < 0.05) than those with medium (1472.4 ± 16.78 g) and high vitality (1590.9 ± 12.13 g). The average difference between the low- and high-vitality piglets was almost 200 g, indicative of an influence of birthweight on neonate vitality that has to do with the sow’s parity. This suggests that the factor of prolificacy must also be taken into account due to its impact on within-litter weight variation (Foisnet et al., 2010) and, therefore, vitality.
In light of the findings reviewed to this point, it is clear that the farrowing process requires close supervision and care, especially when it involves primiparous sows. It is important to emphasize that piglets born to PS tend to have lower vitality and may take longer to reach the teat, which delays their first feeding. Therefore, it is crucial to ensure that they receive colostrum quickly to prevent depletion of their energy reserves.
3.3 Thermoregulation in neonates
Thermoregulation is the physiological mechanism that allows animals to maintain their body temperature. The efficiency of thermoregulation in piglets has been related to birthweight, vitality, and colostrum intake (Herpin et al., 2002; Nuntapaitoon et al., 2019; Roldán-Santiago et al., 2019a), three factors that, as we have seen, are influenced by parity (Muns et al., 2015; Nuntapaitoon et al., 2019). It is well known that newborn piglets have no reserves of brown fat, an energy resource used in thermoregulation (Berthon et al., 1994; Hou et al., 2017). Moreover, after expulsion, the skin of neonates is moist with residues of their mother’s placental fluids, so if the farrowing environment is cold their body temperature may drop. This is especially dangerous for small neonates because they have a greater surface:volume ratio than larger ones that leaves them more susceptible to cold (Kammersgaard et al., 2011; Pedersen et al., 2016). We must recall, as well, that piglets born to 1st-parity sows tend to weigh less than those from MS (Roldán-Santiago et al., 2019a) and that in the first hours of life their temperature may be compromised by such factors as their lack of hair and humidity levels, which can exacerbate heat loss and accentuate the consequences of their limited thermoregulatory ability at birth. This means that maintaining the body temperature of neonates depends almost exclusively on shivering thermogenesis, but this action has low efficiency and entails high energy costs (Berthon et al., 1994; Herpin et al., 2002; Farmer and Edwards, 2022). For this reason, piglets tend to adopt behaviors such as huddling and taking certain postures to maintain their temperature. Colostrum ingestion is especially important in the thermoregulation of neonate piglets (Rooke and Bland, 2002; Inoue and Tsukahara, 2021). Here, once again, the sow’s parity plays a role since it influences prolactin levels, and as noted above, the prolactin-to-progesterone ratio is closely related to colostrum production (Quesnel et al., 2013; Yun et al., 2014). Finally, ensuring effective thermoregulation in piglets is vital during the first 2 days postpartum, for only then do they begin to develop a reasonably effective thermoregulation capacity (Berthon et al., 1994).
Low-weight piglets –with low vitality and poor thermoregulation capacity that increases heat loss– are at a significant disadvantage in competition for the udder (Kirkden et al., 2013; Vodolazska et al., 2023). Roldán-Santiago et al. (2019a) measured the surface temperature of piglets based on readings from the lacrimal caruncle using infrared thermography to evaluate the relation of birthweight and vitality in piglets born to sows of parities 1, 2-5, and 6-7. Results showed that the piglets of higher-parity sows had a better thermoregulation capacity than those born to young ones (up to 1.6°C higher temperatures), while better vitality at birth also meant higher temperatures in the neonates of sows of the same parity (P < 0.05). They further reported that the temperature of piglets from PS varied according to their vitality at the moment they took the mother’s teat. Thus, low-vitality piglets from PS had temperatures up to 2.8°C lower at the time of first contact with the teat, and surface temperatures 0.7°C lower than high-vitality piglets at 24 h postpartum. Finally, they found that a piglets’ thermoregulatory capacity was linked to the sows’ parity, as neonates from higher-parity mother showed better thermoregulation, regardless of vitality. This highlights the significance of parity on the temperature of neonate piglets.
Schild et al. (2020) studied sows from their 1st to 2nd parities to evaluate the characteristics of hyperprolific dams. Among other parameters, they measured differences in the rectal temperatures of their neonates. The piglets from the PS had lower readings than those from the 2nd-parity sows (38.0 ± 0.08 vs. 38.9 ± 0.09°C, P<0.001), thus, it is important to account for (i) the variability in weight within litters, which increases in high-prolificacy sows (Moreira et al., 2020) and (ii) temperature variation. Studies have shown that rectal temperature at 24 h is associated with the amount of colostrum consumed (200 g at 37.5°C, 400 g at 38°C) (Tuchscherer et al., 2000; Devillers et al., 2011). Colostrum ingestion by neonates of PS has been reported in a range of 25-60 g less than that consumed by the offspring of MS (Ferrari et al., 2014; Nuntapaitoon et al., 2019). In a study designed to analyze the effects of distinct management routines during farrowing on suckling latency, DWG, piglet mortality, and heat loss, Vasdal et al. (2011) gathered data on the weight and rectal temperatures of piglets from sows of parities 1-2, 3-4, and 4-7. They found that piglets from the sows of parities 1-2 presented the largest drops in rectal temperature at 2 h, a result they attributed to the poor development of the sows’ udders, which produced less colostrum than those of the dams of higher parity. This finding is also related to birthweight, as the heavier piglets had higher surface temperatures in the first 30 and 45 min postpartum, and, therefore, suffered less heat loss due to postnatal hypothermia (Caldara et al., 2014; Vodolazska et al., 2023).
As the studies summarized above show, newborn piglets have poor thermoregulation, regardless of the mother’s parity. However, neonates born to PS show some significant differences compared to those from MS. Here, lower birthweight and reduced feed intake are associated with potential disadvantages for survival, which means that those piglets require greater supervision and special handling during and after farrowing, especially post-expulsion drying and monitoring of feed intake. Providing an environment with a comfortable temperature that is suitable for neonates is also essential, especially for all piglets born to PS and low-weight neonates from MS.
3.4 Growth rate
Growth rate is another key indicator of piglet performance. It is largely driven by feed intake (Magowan et al., 2007), while colostrum ingestion is strongly influenced by vitality (Uddin et al., 2022). Piglet vitality at birth is a factor that correlates positively with growth rates and indices of neonate survival to weaning (Muns et al., 2013). Roldán-Santiago et al. (2019b) evaluated the effect of parity on the vitality and physiological parameters of piglets born to sows of parities 1-7. Their results describe that parity influences the time piglets require to reach the teat and begin to ingest colostrum. They also found that the piglets born to sows in their 1st- and 7th-parity groups took longer to take the teat than those from the sows in the other groups (54.5, 36.6, 28.0, 23.9, 29.5, 34.8, and 48.8 min for parities 1-7 respectively, P < 0.05). It appeared that their low vitality prevented those piglets from feeding efficiently and impacted their initial growth rate. The probability of surviving to weaning is also greater in piglets with higher average birthweights. On this topic, Milligan et al. (2002b) and Carney-Hinkle et al. (2013) suggest that the capacity for growth of piglets born to PS is affected by poor health due to the low level of immunoprotection they acquire through their dams’ colostrum (Cabrera et al., 2012).
Ruediger and Schulze (2012) evaluated administering azaperone at the end of farrowing on piglet performance. Comparing sows of parities 1-6, they determined that birthweight affected the neonates’ DWG during lactation. Considering only the effect of parity on the control group, they reported the following figures for the DWG of the piglets born to the sows in parities 1-6: mean 179 g (range: 151-249); 209 g (164-249 g); 206 g (169-242); 215 g (175-250); 218 g (181-259); and 205 g (154-246) (P < 0.001). Zotti et al. (2017) observed a similar effect in their work with piglets from sows of parities 1-5 where they sought to determine the effect of parity on birthweight and postnatal development during 3 periods: days 0-21, 21-59, and 59-168. They found that piglets from PS presented up to 10% less DWG than those from the other parities during lactation, so they concluded that birthweight and parity affect postnatal development mainly during early extrauterine life. It is important to recall that high prolificacy increases within-litter body weight variation, which may result in low-birthweight piglets exhibiting lower growth rates (López-Vergé et al., 2018). This is explained, in part, by the reduced size of the intestines, shorter villus height, and the villus:crypt ratio (Che et al., 2016), though weaning-induced stress must also be considered. In addition, separation from the mother affects growth rates, and additional factors, such as the mixing of litters, may also have an influence by causing distress and behavioral disorders (Jensen, 2002) that have consequences for piglets’ endocrine and immune systems (Merlot et al., 2004).
Ferrari et al.’s (2014) comparative analysis of the effect of colostrum consumption in piglets born to PS vs. MS (parities 4-6) revealed that the neonates of the former consumed less colostrum than those in the MS group (271.9 ± 7.7 vs. 297.6 ± 7.7 mg/ml; P < 0.003). Their results agree with those of Nuntapaitoon et al. (2019), who compared the intake of piglets born to sows of parities 1, 2, 3, and 4-7 (362 ± 11, 413 ± 13, 425 ± 13, and 422 ± 9 g, respectively, P<0.001). They recommended a consumption level of at least 400 g of colostrum in the first 24 h of life to improve survival potential and prevent effects on growth rates, since piglets that consume more than this amount show higher growth rates during lactation. To prevent the development of large weight differences between piglets from PS and MS, it is necessary to ensure that those from the former consume sufficient colostrum after birth. This is a crucial management issue because those piglets are disadvantaged in two ways: first, they take at least 21 more min to reach the mother’s teat; second, their DWG is 30 g lower. To reduce the differences in productive performance between these two groups of piglets, greater attention must be paid to the care and management of neonates.
4 Conclusions
Having reviewed and analyzed extensive information on multiple factors associated with the sows’ parity and its impact during the farrowing process and on neonate performance, we can conclude that certain hormonal differences –especially prolactin levels– are related to parity during labor since values in primiparous sows may be as much as 50% below those found in multiparous sows. One consequence of this is lower colostrum and milk production that may oblige breeders to consider the use of nurse sows when primiparous sows have large litters to ensure adequate feed intake, promote normal growth, and correct thermoregulation. In line with the findings reported in various studies, we can also affirm that primiparous sows do not necessarily experience difficult births but do take a little longer to return to estrus, due primarily to energy depletion during lactation and/or dietary deficiencies that have a more pronounced effect on young sows who are still developing.
Primiparous sows are also more susceptible to the physical environment due to the novel and unfamiliar experience of farrowing, which may maintain them in a constant state of alert. This is another aspect that must be considered when moving primiparous sows prior to farrowing in order to avoid adding stressors that could affect their performance. To prevent negative effects on the productivity of these sows –which is naturally lower because they are still growing– it is essential to consider that both their own performance and that of their offspring will be lower than those of multiparous sows and their litters. In addition, primiparous sows may have longer weaning-to-estrus intervals, smaller and lighter litters, lower DWG by their piglets, and higher FI.
The parity of sows, therefore, directly impacts neonates and this is particularly evident in primiparous sows whose piglets tend to have lower birthweights than those of multiparous mothers. This difference subsequently influences their overall vitality, feeding ability, thermoregulation capacity, and ultimately, mortality rates. Only by understanding these significant differences due to parity will breeders be able to intervene effectively in the areas where primiparous sows present disadvantages.
Author's note
AC-H is in receipt of SECIHTI fellowship 795362. This work was submitted in partial fulfillment of the requirements of the Ph.D. degree program for AC-H at the Universidad Nacional Autónoma de México. PR-S, HB-J, LC-C, OL-M, HO-G are members of SNI, SECIHTI.
Author contributions
AC-H: Investigation, Methodology, Writing – review & editing, Conceptualization, Writing – original draft. PR-S: Conceptualization, Data curation, Project administration, Writing – review & editing. HB-J: Conceptualization, Project administration, Supervision, Writing – review & editing. LC-C: Validation, Visualization, Writing – review & editing. OL-M: Conceptualization, Supervision, Validation, Visualization, Writing – review & editing. HO-G: Investigation, Methodology, Validation, Visualization, Writing – review & editing. RP-R: Conceptualization, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adi Y. K., Boonprakob R., Kirkwood R. N., Tummaruk P. (2022). Factors associated with farrowing duration in hyperprolific sows in a free farrowing system under tropical conditions. Animals 12, 2943. doi: 10.3390/ani12212943
Alonso-Spilsbury M. a., Mota-Rojas D., Martı́nez-Burnes J., Arch E., Mayagoitia A. L., Ramı́rez-Necoechea R., et al. (2004). Use of oxytocin in penned sows and its effect on fetal intra-partum asphyxia. Anim. Reprod. Sci. 84, 157–167. doi: 10.1016/j.anireprosci.2003.11.002
Bardehle D., Preissler R., Lehmann J., Looft H., Kemper N. (2012). Analysis of fertility and performance parameters in piglet production considering partus induction, birth assistance and Mastitis-Metritis-Agalactia (MMA). Züchtungskunde 84, 293–306.
Bar-Pelled U., Maltz E., Bruckental I., Folman Y., Kali Y., Gacitua H., et al. (1995). Relationship between frequent milking or suckling in early lactation and milk production of high producing dairy cows. J. Dairy Sci. 78, 2726–2736. doi: 10.3168/jds.S0022-0302(95)76903-X
Beaulieu A.D., Aalhus J.L., Williams N.H., Patience J.F. (2010). Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork. J. Anim. Sci. 88 (8), 2767–2778. doi: 10.2527/jas.2009-2222
Berthon D., Herpin P., Le Dividich J. (1994). Shivering thermogenesis in the neonatal pig. J. Thermal Biol. 19, 413–418. doi: 10.1016/0306-4565(94)90040-X
Beyer M., Jentsch W., Kuhla S., Wittenburg H., Kreienbring F., Scholze H., et al. (2007). Effects of dietary energy intake during gestation and lactation on milk yield and composition of first, second and fourth parity sows. Arch. Anim. Nutr. 61, 452–468. doi: 10.1080/17450390701563433
Bierhals T., Magnabosco D., Ribeiro R., Perin J., Da Cruz R., Bernardi M., et al. (2012). Influence of pig weight classification at cross-fostering on the performance of the primiparous sow and the adopted litter. Livest. Sci. 146, 115–122. doi: 10.1016/j.livsci.2012.02.026
Björkman S., Oliviero C., Rajala-Schultz P., Soede N., Peltoniemi O. (2017). The effect of litter size, parity and farrowing duration on placenta expulsion and retention in sows. Theriogenology 92, 36–44. doi: 10.1016/j.theriogenology.2017.01.003
Bolado P. M., Pereda M. J., Gonzáles H. C., Izquierdo P., Palacio C. D. (2011). Influencia de la paridad de la cerda sobre las características de sus crías en el período predestete. Rev. Producción Anim. 23, 75–81.
Bondoc O., Isubol J. (2022). Reasons for culling in relation to longevity and lifetime efficiency of landrace and large white sows in a nucleus breeding farm in the Philippines. Trop. Anim. Sci. J. 45, 30–36. doi: 10.5398/tasj.2022.45.1.30
Butler W. R. (2005). Inhibition of ovulation in the postpartum cow and the lactating sow. Livest. Prod. Sci. 98, 5–12. doi: 10.1016/j.livprodsci.2005.10.007
Cabrera R. A., Lin X., Campbell J. M., Moeser A. J., Odle J. (2012). Influence of birth order, birth weight, colostrum and serum immunoglobulin G on neonatal piglet survival. J. Anim. Sci. Biotechnol. 3, 1–10. doi: 10.1186/2049-1891-3-42
Caldara F. R., Dos Santos L. S., MaChado S. T., Moi M., de Alencar Nääs I., Foppa L., et al. (2014). Piglets’ surface temperature change at different weights at birth. Asian-Australasian J. Anim. Sci. 27, 431. doi: 10.5713/ajas.2013.13505
Carney-Hinkle E., Tran H., Bundy J., Moreno R., Miller P., Burkey T. (2013). Effect of dam parity on litter performance, transfer of passive immunity, and progeny microbial ecology. J. Anim. Sci. 91, 2885–2893. doi: 10.2527/jas.2011-4874
Casellas J., Rauw W., Piedrafita J., Sánchez A., Arqué M., Noguera J. (2004). Viability of Iberian× Meishan F2 newborn pigs. I. Analysis of physiological and vitality variables. J. Anim. Sci. 82, 1919–1924. doi: 10.2527/2004.8271919x
Che L., Hu L., Liu Y., Yan C., Peng X., Xu Q., et al. (2016). Dietary nucleotides supplementation improves the intestinal development and immune function of neonates with intra-uterine growth restriction in a pig model. PloS One 11, e0157314. doi: 10.1371/journal.pone.0157314
Craig J. R., Dunshea F. R., Cottrell J. J., Wijesiriwardana U. A., Pluske J. R. (2019). Primiparous and multiparous sows have largely similar colostrum and milk composition profiles throughout lactation. Animals 9, 35. doi: 10.3390/ani9020035
Cronin G., Schirmer B., McCallum T., Smith J., Butler K. (1993). The effects of providing sawdust to pre-parturient sows in farrowing crates on sow behaviour, the duration of parturition and the occurrence of intra-partum stillborn piglets. Appl. Anim. Behav. Sci. 36, 301–315. doi: 10.1016/0168-1591(93)90128-C
Devillers N., Farmer C., Le Dividich J., Prunier A. (2007). Variability of colostrum yield and colostrum intake in pigs. Animal 1, 1033–1041. doi: 10.1017/S175173110700016X
Devillers N., Farmer C., Mounier A.-M., Ledividich J., Prunier A. (2004). Hormones, IgG and lactose changes around parturition in plasma, and colostrum or saliva of multiparous sows. Reprod. Nutr. Dev. 44, 381–396. doi: 10.1051/rnd:2004043
Devillers N., Le Dividich J., Prunier A. (2011). Influence of colostrum intake on piglet survival and immunity. Animal 5, 1605–1612. doi: 10.1017/S175173111100067X
Ek-Mex J. E., Segura-Correa J. C., Batista-Garcia L., Alzina-López A. (2014). Factores ambientales que afectan los componentes de producción y productividad durante la vida de las cerdas. Trop. Subtrop. Agroecosystems 17, 447–462.
Estienne M., Harper A. (2010). Type of accommodation during gestation affects growth performance and reproductive characteristics of gilt offspring. J. Anim. Sci. 88, 400–407. doi: 10.2527/jas.2009-2081
Farmer C. (2022). Achieving optimal sow performance, still an ongoing challenge in 2022. Anim. Front. 12, 53–55. doi: 10.1093/af/vfac064
Farmer C., Edwards S. A. (2022). Review: Improving the performance of neonatal piglets. Animal 16, 100350. doi: 10.1016/j.animal.2021.100350
Farmer C., Robert S. (2003). Hormonal, behavioural and performance characteristics of Meishan sows during pregnancy and lactation. Can. J. Anim. Sci. 83, 1–12. doi: 10.4141/A02-034
Farmer C., Robert S., Matte J., Girard C., Martineau G. (1995). Endocrine and peripartum behavioral responses of sows fed high-fiber diets during gestation. Can. J. Anim. Sci. 75, 531–536. doi: 10.4141/cjas95-080
Ferrari C., Sbardella P., Bernardi M., Coutinho M., Vaz I. Jr., Wentz I., et al. (2014). Effect of birth weight and colostrum intake on mortality and performance of piglets after cross-fostering in sows of different parities. Prev. Vet. Med. 114, 259–266. doi: 10.1016/j.prevetmed.2014.02.013
Foisnet A., Farmer C., David C., Quesnel H. (2010). Relationships between colostrum production by primiparous sows and sow physiology around parturition1. J. Anim. Sci. 88, 1672–1683. doi: 10.2527/jas.2009-2562
Foxcroft G. (2007). “Pre-natal programming of variation in post-natal performance-how and when? Adv. Pork Prod. 18, 167–189.
Fraser D., Phillips P., Thompson B. (1997). Farrowing behaviour and stillbirth in two environments: an evaluation of the restraint-stillbirth hypothesis. Appl. Anim. Behav. Sci. 55, 51–66. doi: 10.1016/S0168-1591(97)00007-5
Fraser D., Rushen J. (1992). Colostrum intake by newborn piglets. Can. J. Anim. Sci. 72, 1–13. doi: 10.4141/cjas92-001
Galíndez R., Pulido F. (2014). Efectos genéticos y no genéticos sobre el intervalo entre partos de cerdas Duroc, Hampshire y sus cruces recíprocos. Zootecnia Trop. 32, 139–148.
García-González J. S., Herradora-Lozano M. A., Martínez-Gamba R. G. (2011). Efecto del número de parto de la cerda, la caseta de parición, el tamaño de la camada y el peso al nacer en las principales causas de mortalidad en lechones. Rev. mexicana Cienc. pecuarias 2, 403–414.
Gerjets I., Traulsen I., Reiners K., Kemper N. (2011). Assessing individual sow risk factors for coliform mastitis: A case–control study. Prev. Vet. Med. 100, 248–251. doi: 10.1016/j.prevetmed.2011.04.012
Girardie O., Bonneau M., Billon Y., Bailly J., David I., Canario L. (2023). Analysis of image-based sow activity patterns reveals several associations with piglet survival and early growth. Front. Vet. Sci. 9. doi: 10.3389/fvets.2022.1051284
González-Hernández C., de Armas Rodríguez I., Paz Sieres C., Guevara Viera G., Tamayo Escobar Y. (2002). Influencia del número de partos y la época del año sobre indicadores reproductivos en una unidad porcina. Rev. Producción Anim. 14, 69–72.
Hales J., Moustsen V. A., Devreese A. M., Nielsen M. B. F., Hansen C. F. (2015). Comparable farrowing progress in confined and loose housed hyper-prolific sows. Livest. Sci. 171, 64–72. doi: 10.1016/j.livsci.2014.11.009
Hales J., Moustsen V., Nielsen M., Hansen C. (2016). The effect of temporary confinement of hyperprolific sows in Sow Welfare and Piglet protection pens on sow behaviour and salivary cortisol concentrations. Appl. Anim. Behav. Sci. 183, 19–27. doi: 10.1016/j.applanim.2016.07.008
Han T., Björkman S., Soede N. M., Oliviero C., Peltoniemi O. A. T. (2020). IGF-1 concentration patterns and their relationship with follicle development after weaning in young sows fed different pre-mating diets. Animal 14, 1493–1501. doi: 10.1017/S1751731120000063
Hartmann P. E., Whitely J. L., Willcox D. L. (1984). Lactose in plasma during lactogenesis, established lactation and weaning in sows. J. Physiol. 347, 453–463. doi: 10.1113/jphysiol.1984.sp015075
Herpin P., Damon M., Le Dividich J. (2002). Development of thermoregulation and neonatal survival in pigs. Livest. Prod. Sci. 78, 25–45. doi: 10.1016/S0301-6226(02)00183-5
Herpin P., Le Dividich J., Hulin J. C., Fillaut M., De Marco F., Bertin R. (1996). Effects of the level of asphyxia during delivery on viability at birth and early postnatal vitality of newborn pigs. J. Anim. Sci. 74, 2067–2075. doi: 10.2527/1996.7492067x
Herpin P., Louveau I., Damon M., Le Dividich J. (2005). “Environmental and hormonal regulation of energy metabolism in early development of the pig,” in Biology of growing animals (Amsterdam, Netherlands: Elsevier), 351–374.
Hötzel M. J., Wolf F. M., Dalla Costa O. A. (2004). Behaviour of sows and piglets reared in intensive outdoor or indoor systems. Appl. Anim. Behav. Sci. 86, 27–39. doi: 10.1016/j.applanim.2003.11.014
Hou L., Shi J., Cao L., Xu G., Hu C., Wang C. (2017). Pig has no uncoupling protein 1. Biochem. Biophys. Res. Commun. 487, 795–800. doi: 10.1016/j.bbrc.2017.04.118
Inoue R., Tsukahara T. (2021). Composition and physiological functions of the porcine colostrum. Anim. Sci. J. 92, e13618. doi: 10.1111/asj.13618
Ison S. H., Jarvis S., Hall S. A., Ashworth C. J., Rutherford K. M. (2018). Periparturient behavior and physiology: further insight into the farrowing process for primiparous and multiparous sows. Front. Vet. Sci. 5. doi: 10.3389/fvets.2018.00122
Jarvis S., Lawrence A., McLean K., Deans L., Chirnside J., Calvert S. (1997). The effect of environment on behavioural activity, ACTH,(β-endorphin and cortisol in pre-farrowing gilts. Anim. Sci. 65, 465–472. doi: 10.1017/S1357729800008663
Jarvis S., van der Vegt B. J., Lawrence A. B., McLean K. A., Deans L. A., Chirnside J., et al. (2001). The effect of parity and environmental restriction on behavioural and physiological responses of pre-parturient pigs. Appl. Anim. Behav. Sci. 71, 203–216. doi: 10.1016/S0168-1591(00)00183-0
Jensen P. (2002). “Behaviour of pigs,” in The ethology of domestic animals: an introductory text, 1st ed. Ed. Jensen P. (CABI Publishing, Wallingford, UK), 159–172.
Ju M., Wang X., Li X., Zhang M., Shi L., Hu P., et al. (2022). Effects of litter size and parity on farrowing duration of landrace × Yorkshire sows. Animals 12, 94. doi: 10.3390/ani12010094
Kaiser M., Jacobsen S., Andersen P. H., Bækbo P., Cerón J. J., Dahl J., et al. (2018). Hormonal and metabolic indicators before and after farrowing in sows affected with postpartum dysgalactia syndrome. BMC Vet. Res. 14, 334. doi: 10.1186/s12917-018-1649-z
Kammersgaard T., Pedersen L., Jørgensen E. (2011). Hypothermia in neonatal piglets: interactions and causes of individual differences. J. Anim. Sci. 89, 2073–2085. doi: 10.2527/jas.2010-3022
Karvelienė B., Šernienė L., Riškevičienė V. (2008). Effect of different factors on weaning-to-first-service interval in Lithuanian pig herds. Veterinarija ir zootechnika 41, 64–69.
Kim S. W., Easter R. A. (2001). Nutrient mobilization from body tissues as influenced by litter size in lactating sows1. J. Anim. Sci. 79, 2179–2186. doi: 10.2527/2001.7982179x
Kirkden R., Broom D., Andersen I. (2013). Piglet mortality: the impact of induction of farrowing using prostaglandins and oxytocin. Anim. Reprod. Sci. 138, 14–24. doi: 10.1016/j.anireprosci.2013.02.009
Kitkha S., Boonsoongnern A., Ratanavanichrojn N., Jirawattanapong P., Pinyopummin A. (2017). Effects of the higher parity and litter size on piglet birth weight and survival rate of later born piglets. Thai J. Vet. Med. 47, 79–86. doi: 10.56808/2985-1130.2815
Klimas R., Klimienė A., Sobotka W., Kozera W., Matusevičius P. (2020). Effect of parity on reproductive performance sows of different breeds. South Afr. J. Anim. Sci. 50, 434–441. doi: 10.4314/sajas.v50i3.10
Knox R. V., Zas S. L. R. (2001). Factors influencing estrus and ovulation in weaned sows as determined by transrectal ultrasound1. J. Anim. Sci. 79, 2957–2963. doi: 10.2527/2001.79122957x
Kobek-Kjeldager C., Larsen M. L. V., Pedersen L. J. (2023). Changes in piglet and litter characteristics across parities in two highly prolific sow hybrids in an outdoor organic herd. Anim. Sci. J. 94, e13840. doi: 10.1111/asj.13840
Koketsu Y. (2005). Within-farm variability in age structure of breeding-female pigs and reproductive performance on commercial swine breeding farms. Theriogenology 63, 1256–1265. doi: 10.1016/j.theriogenology.2004.04.018
Koketsu Y., Dial G. D., Pettigrew J. E., Marsh W. E., King V. L. (1996). Influence of imposed feed intake patterns during lactation on reproductive performance and on circulating levels of glucose, insulin, and luteinizing hormone in primiparous sows. J. Anim. Sci. 74, 1036–1046. doi: 10.2527/1996.7451036x
Koketsu Y., Tani S., Iida R. (2017). Factors for improving reproductive performance of sows and herd productivity in commercial breeding herds. Porcine Health Manage. 3, 1–10. doi: 10.1186/s40813-016-0049-7
Lammers G., De Lange A. (1986). Pre-and post-farrowing behaviour in primiparous domesticated pigs. Appl. Anim. Behav. Sci. 15, 31–43. doi: 10.1016/0168-1591(86)90020-1
Le Dividich J. (1999). Review: management to reduce variation in pre-and post-weaned pigs. Cranwell PD Australas. Pig Sci. Assoc., 135–155.
Leite C., Lui J., Albuquerque L., Alves D. (2011). Environmental and genetic factors affecting the weaning-estrus interval in sows. Genet. Mol. Res., 2692–2701. doi: 10.4238/2011.November.4.2
Li S., Lu C., Jia M., Zhang Z., Zhang H., Song Z., et al. (2023). Comparison between maternal behaviors of primiparous and multiparous sows. Livest. Sci. 267, 105127. doi: 10.1016/j.livsci.2022.105127
Liu Y., Zhou Q., Theil P. K., Fang Z., Lin Y., Xu S., et al. (2021). The differences in energy metabolism and redox status between sows with short and long farrowing duration. Animal 15, 100355. doi: 10.1016/j.animal.2021.100355
Llamas-López P. J., López-Úbeda R., López G., Antinoja E., García-Vázquez F. A. (2019). A new device for deep cervical artificial insemination in gilts reduces the number of sperm per dose without impairing final reproductive performance. J. Anim. Sci. Biotechnol. 10, 11. doi: 10.1186/s40104-019-0313-1
López-Vergé S., Gasa J., Farré M., Coma J., Bonet J., Solà-Oriol D. (2018). Potential risk factors related to pig body weight variability from birth to slaughter in commercial conditions. Trans. Anim. Sci. 2, 383–395. doi: 10.1093/tas/txy082
Maes D., Papadopoulos G., Cools A., Janssens G. P. J. (2010). Postpartum dysgalactia in sows: pathophysiology and risk factors. Tierarztl Prax Ausg G Grosstiere Nutztiere 38, S15–S20. doi: 10.1055/s-0038-1623058
Magowan E., McCann M. E. E., Beattie V. E., McCracken K. J., Henry W., Smyth S., et al. (2007). Investigation of growth rate variation between commercial pig herds. animal 1, 1219–1226. doi: 10.1017/S1751731107000572
Mainau E., Dalmau A., Ruiz-de-la-Torre J. L., Manteca X. (2009). Validation of an automatic system to detect position changes in puerperal sows. Appl. Anim. Behav. Sci. 121, 96–102. doi: 10.1016/j.applanim.2009.09.005
Mainau E., Dalmau A., Ruiz-de-la-Torre J., Manteca X. (2010). A behavioural scale to measure ease of farrowing in sows. Theriogenology 74, 1279–1287. doi: 10.1016/j.theriogenology.2010.05.034
Malavé T., Alfaro M., Hurtado E. (2007). Efecto del número de partos, tamaño y peso de la camada al destete sobre el intervalo destete-estro en cerdas. Rev. Unellez Ciencia y Tecnología 25, 10–15.
Mendoza U., Ortega R. (2009). Factores genéticos y ambientales que influyen el intervalo destete-servicio en cerdas (Cuba: Instituto de Investigaciones Porcinas).
Menegat M., Tokach M. D. (2021). 263 review of current nutrition knowledge and practices for gilt development. J. Anim. Sci. 99, 105–106. doi: 10.1093/jas/skab054.171
Merlot E., Meunier-Salaün M.-C., Prunier A. (2004). Behavioural, endocrine and immune consequences of mixing in weaned piglets. Appl. Anim. Behav. Sci. 85, 247–257. doi: 10.1016/j.applanim.2003.11.002
Miller H. M., Foxcroft G. R., Aherne F. X. (2004). Increasing feed intake in late gestation does not affect plasma progesterone concentration in the sow. Theriogenology 62, 1618–1626. doi: 10.1016/j.theriogenology.2004.03.002
Milligan B. N., Dewey C. E., de Grau A. F. (2002a). Neonatal-piglet weight variation and its relation to pre-weaning mortality and weight gain on commercial farms. Prev. Vet. Med. 56, 119–127. doi: 10.1016/S0167-5877(02)00157-5
Milligan B. N., Fraser D., Kramer D. L. (2002b). Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 76, 181–191. doi: 10.1016/S0301-6226(02)00012-X
Moeller S., Goodwin R., Johnson R., Mabry J., Baas T., Robison O. (2004). The National Pork Producers Council Maternal Line National Genetic Evaluation Program: A comparison of six maternal genetic lines for female productivity measures over four parities. J. Anim. Sci. 82, 41–53. doi: 10.2527/2004.82141x
Moreira R. H. R., Pérez Palencia J. Y., Moita V. H. C., Caputo L. S. S., Saraiva A., Andretta I., et al. (2020). Variability of piglet birth weights: A systematic review and meta-analysis. J. Anim. Physiol. Anim. Nutr. 104, 657–666. doi: 10.1111/jpn.13264
Muns R., Manteca X., Gasa J. (2015). Effect of different management techniques to enhance colostrum intake on piglets’ growth and mortality. Anim. Welfare 24, 185–192. doi: 10.7120/09627286.24.2.185
Muns R., Manzanilla E. G., Sol C., Manteca X., Gasa J. (2013). Piglet behavior as a measure of vitality and its influence on piglet survival and growth during lactation. J. Anim. Sci. 91, 1838–1843. doi: 10.2527/jas.2012-5501
Murillo-Galán C., Herradora-Lozano M. A., Martínez-Gamba R. (2007). Relación entre la pérdida de grasa dorsal de Cerdas Lactantes con el consumo de alimento, tamaño de la camada, peso de los Lechones al destete y días de Lactancia. Rev. Científica 17, 380–385.
Nam H. N., Sukon P. (2020). Associated factors for farrowing duration in sows with natural parturition in intensive conditions. World 10, 320–324. doi: 10.36380/scil.2020.wv41
Nuntapaitoon M., Muns R., Theil P. K., Tummaruk P. (2019). Factors influencing colostrum consumption by piglets and their relationship with survival and growth in tropical climates. Livest. Sci. 224, 31–39. doi: 10.1016/j.livsci.2019.04.008
Oliviero C., Heinonen M., Valros A., Peltoniemi O. (2010). Environmental and sow-related factors affecting the duration of farrowing. Anim. Reprod. Sci. 119, 85–91. doi: 10.1016/j.anireprosci.2009.12.009
Oliviero C., Kokkonen T., Heinonen M., Sankari S., Peltoniemi O. (2009). Feeding sows with high fibre diet around farrowing and early lactation: impact on intestinal activity, energy balance related parameters and litter performance. Res. Vet. Sci. 86, 314–319. doi: 10.1016/j.rvsc.2008.07.007
Ordaz-Ochoa G., Juárez-Caratachea A., García-Valladares A., Pérez-Sánchez R. E., Ortiz-Rodríguez R. (2013). Efecto del número de parto sobre los principales indicadores reproductivos de las cerdas. Rev. Científica 23, 511–519.
Ortíz R., Ortega R., Becerril J., Consultora O. (2004). Efectos ambientales en cerdas sometidas a lactancias de 12 y 21 días en México: Rasgos del comportamiento reproductivo (Punta Brava, La Habana, Cuba: Instituto de Investigaciones Porcinas).
Pandey A., Singh S., Pandey A. (2010). Studies on genetic and nongenetic factors affecting reproductive traits in landrace desi and their crossbred pigs. J. Ecol. Environ. Sci. 1, 7–10.
Patterson J., Foxcroft G. (2019). Gilt management for fertility and longevity. Animals 9, 434. doi: 10.3390/ani9070434
Pedersen L. J., Larsen M. L. V., Malmkvist J. (2016). The ability of different thermal aids to reduce hypothermia in neonatal piglets1. J. Anim. Sci. 94, 2151–2159. doi: 10.2527/jas.2015-0219
Pérez-Sánchez R. E., Ordaz-Ochoa G. O., Juárez-Caratachea A., García-Valladares A., Ortíz-Rodríguez R. (2015). Efecto del número de parto sobre el consumo de alimento voluntario de las cerdas durante la fase de lactancia y su repercusión en el intervalo destete-estro. Nota técnica. Rev. Científica 25, 145–152.
Piñeiro C., Manso A., Manzanilla E. G., Morales J. (2019). Influence of sows’ parity on performance and humoral immune response of the offspring. Porcine Health Manage. 5, 1. doi: 10.1186/s40813-018-0111-8
Plush K. J., McKenny L. A., Nowland T. L., van Wettere W. H. E. J. (2021). The effect of hessian and straw as nesting materials on sow behaviour and piglet survival and growth to weaning. Animal 15, 100273. doi: 10.1016/j.animal.2021.100273
Quesnel H., Brossard L., Valancogne A., Quiniou N. (2008). Influence of some sow characteristics on within-litter variation of piglet birth weight. Animal 2, 1842–1849. doi: 10.1017/S175173110800308X
Quesnel H., Farmer C. (2019). Review: nutritional and endocrine control of colostrogenesis in swine. Animal 13, 26–34. doi: 10.1017/S1751731118003555
Quesnel H., Farmer C., Devillers N. (2012). Colostrum intake: Influence on piglet performance and factors of variation. Livest. Sci. 146, 105–114. doi: 10.1016/j.livsci.2012.03.010
Quesnel H., Ramaekers P., van Hees H., Farmer C. (2013). Relations between peripartum concentrations of prolactin and progesterone in sows and piglet growth in early lactation. Can. J. Anim. Sci. 93, 109–112. doi: 10.4141/cjas2012-106
Randall G. (1972). Observations on parturition in the sow. I. Factors associated with the delivery of the piglets and their subsequent behaviour. Vet. Rec. 90, 178–182. doi: 10.1136/vr.90.7.178
Reimert I., Bolhuis J. E., Kemp B., Rodenburg T. B. (2013). Indicators of positive and negative emotions and emotional contagion in pigs. Physiol. Behav. 109, 42–50. doi: 10.1016/j.physbeh.2012.11.002
Rempel L. A., Keel B. N., Oliver W. T., Wells J. E., Lents C. A., Nonneman D. J., et al. (2022). Dam parity structure and body condition during lactation influence piglet growth and gilt sexual maturation through pre-finishing. J. Anim. Sci. 100. doi: 10.1093/jas/skac031
Rincón-Gainza C., Colina-Rivero J., Araque-Molina H., Martínez-García G. (2014). Desempeño productivo de cerdas Yorkshire puras y cruzadas en una granja comercial de Venezuela. Rev. Científica 24, 543–552.
Roelofs S., Godding L., de Haan J. R., van der Staay F. J., Nordquist R. E. (2019). Effects of parity and litter size on cortisol measures in commercially housed sows and their offspring. Physiol. Behav. 201, 83–90. doi: 10.1016/j.physbeh.2018.12.014
Roldán-Santiago P., Martínez-Burnes J., López-Mayagoitia A., Ramírez-Necoechea R., Mota-Rojas D. (2019a). Relationship of vitality and weight with the temperature of newborn piglets born to sows of different parity. Livest. Sci. 220, 26–31. doi: 10.1016/j.livsci.2018.12.011
Roldán-Santiago P., Mota-Rojas D., Martínez-Burnes J., Velarde A., Muns R., López-Mayagoitia A. (2019b). Neurophysiological development of newborn pigs: effect of the sow’s parity number in eutocic farrowings. Anim. Prod. Sci. 59, 216–224. doi: 10.1071/AN16641
Rooke J., Bland I. (2002). The acquisition of passive immunity in the new-born piglet. Livest. Prod. Sci. 78, 13–23. doi: 10.1016/S0301-6226(02)00182-3
Roongsitthichai A., Olanratmanee E.-o. (2021). Fetal mortality associated with backfat thickness at first mating and first farrowing of the primiparous sows raised in a commercial herd in Thailand. Trop. Anim. Health Prod. 53, 1–8. doi: 10.1007/s11250-021-02624-3
Rosvold E. M., Newberry R. C., Andersen I. L. (2019). Early mother-young interactions in domestic sows – Nest-building material increases maternal investment. Appl. Anim. Behav. Sci. 219, 104837. doi: 10.1016/j.applanim.2019.104837
Ruediger K., Schulze M. (2012). Post-farrowing stress management in sows by administration of azaperone: Effects on piglets performance. J. Anim. Sci. 90, 2331–2336. doi: 10.2527/jas.2011-4661
Rutherford K., Baxter E., D’eath R., Turner S., Arnott G., Roehe R., et al. (2013). The welfare implications of large litter size in the domestic pig I: biological factors. Anim. Welfare 22, 199–218. doi: 10.7120/09627286.22.2.199
Rydhmer L., Lundeheim N., Canario L. (2008). Genetic correlations between gestation length, piglet survival and early growth. Livest. Sci. 115, 287–293. doi: 10.1016/j.livsci.2007.08.014
Samson W. K., Lumpkin M. D., McCann S. M. (1986). Evidence for a physiological role for oxytocin in the control of prolactin secretion. Endocrinology 119, 554–560.
Santos L. S., Caldara F. R., MaChado S. T., Nääs I. A., Foppa L., Garcia R. G., et al. (2015). Sow’s parity and coconut oil postnatal supplement on piglets performance. Rev. MVZ Córdoba 20, 4513–4521.
Sasaki Y., Koketsu Y. (2007). Variability and repeatability in gestation length related to litter performance in female pigs on commercial farms. Theriogenology 68, 123–127. doi: 10.1016/j.theriogenology.2007.04.021
Schild S.-L. A., Foldager L., Rangstrup-Christensen L., Pedersen L. J. (2020). Characteristics of piglets born by two highly prolific sow hybrids. Front. Vet. Sci. 7. doi: 10.3389/fvets.2020.00355
Schodl K., Revermann R., Winckler C., Fuerst-Waltl B., Leeb C., Willam A., et al. (2019). Assessment of piglet vitality by farmers—Validation of A scoring scheme and estimation of associated genetic parameters. Animals 9, 317. doi: 10.3390/ani9060317
Škorjanc D., Hohler M., Brus M. (2008). Effect of backfat loss during lactation on weaning-to-oestrus interval of sows at gonadotropin application. Arch. Anim. Breed. 51, 560–571. doi: 10.5194/aab-51-560-2008
Smith H., Selby C., Williams A., Ellersieck M., Lamberson W., Safranski T. (2013). Effects of day of farrowing induction and spontaneous versus induced farrowing on sow and suckling piglet performance. J. Swine Health Prod. 21, 195–202.
Suriyasomboon A., Lundeheim N., Kunavongkrit A., Einarsson S. (2006). Effect of temperature and humidity on reproductive performance of crossbred sows in Thailand. Theriogenology 65, 606–628. doi: 10.1016/j.theriogenology.2005.06.005
Tang X., Xiong K. (2022). Intrauterine growth retardation affects intestinal health of suckling piglets via altering intestinal antioxidant capacity, glucose uptake, tight junction, and immune responses. Oxid. Med. Cell Longev. 2022, 2644205. doi: 10.1155/2022/2644205
Tantasuparuk W., Lundeheim N., Dalin A., Kunavongkrit A., Einarsson S. (2000). Reproductive performance of purebred Landrace and Yorkshire sows in Thailand with special reference to seasonal influence and parity number. Theriogenology 54, 481–496. doi: 10.1016/S0093-691X(00)00364-2
Theil P. K., Lauridsen C., Quesnel H. (2014). Neonatal piglet survival: impact of sow nutrition around parturition on fetal glycogen deposition and production and composition of colostrum and transient milk. Animal 8, 1021–1030. doi: 10.1017/S1751731114000950
Thorsen C. K., Schild S.-L. A., Rangstrup-Christensen L., Bilde T., Pedersen L. J. (2017). The effect of farrowing duration on maternal behavior of hyperprolific sows in organic outdoor production. Livest. Sci. 204, 92–97. doi: 10.1016/j.livsci.2017.08.015
Tospitakkul P., Kraomkaew K., Thammasin K., Uttarak P., Nuntapaitoon M., De Rensis F., et al. (2019). Induction of parturition by double administration of prostaglandin F2alpha in sows reduces the variation of gestation length without affecting the colostrum yield and piglet performance. J. Vet. Med. Sci. 81 (9), 18–0725. doi: 10.1292/jvms.18-0725
Trujillo-Ortega M. E., Mota-Rojas D., Olmos-Hernández A., Alonso-Spilsbury M., González M., Orozco H., et al. (2007). A study of piglets born by spontaneous parturition under uncontrolled conditions: could this be a naturalistic model for the study of intrapartum asphyxia? Acta Biomedica-Ateneo Parmense 78, 29.
Tuchscherer M., Puppe B., Tuchscherer A., Tiemann U. (2000). Early identification of neonates at risk: Traits of newborn piglets with respect to survival. Theriogenology 54, 371–388. doi: 10.1016/S0093-691X(00)00355-1
Tucker B. S., Craig J. R., Morrison R. S., Smits R. J., Kirkwood R. N. (2021). Piglet viability: A review of identification and pre-weaning management strategies. Animals 11, 2902. doi: 10.3390/ani11102902
Tummaruk P., Sang-Gassanee K. (2013). Effect of farrowing duration, parity number and the type of anti-inflammatory drug on postparturient disorders in sows: a clinical study. Trop. Anim. Health Prod. 45, 1071–1077. doi: 10.1007/s11250-012-0315-x
Tummaruk P., Tantasuparuk W., Techakumphu M., Kunavongkrit A. (2007). Age, body weight and backfat thickness at first observed oestrus in crossbred Landrace× Yorkshire gilts, seasonal variations and their influence on subsequence reproductive performance. Anim. Reprod. Sci. 99, 167–181. doi: 10.1016/j.anireprosci.2006.05.004
Uddin M. K., Hasan S., Peltoniemi O., Oliviero C. (2022). The effect of piglet vitality, birth order, and blood lactate on the piglet growth performances and preweaning survival. Porcine Health Manage. 8, 52. doi: 10.1186/s40813-022-00299-2
Vallet J., Miles J., Rempel L. (2013). A simple novel measure of passive transfer of maternal immunoglobulin is predictive of preweaning mortality in piglets. Vet. J. 195, 91–97. doi: 10.1016/j.tvjl.2012.06.009
Vanden Hole C., Aerts P., Prims S., Ayuso M., Van Cruchten S., Van Ginneken C. (2018). Does intrauterine crowding affect locomotor development? A comparative study of motor performance, neuromotor maturation and gait variability among piglets that differ in birth weight and vitality. PloS One 13, e0195961. doi: 10.1371/journal.pone.0195961
Van Dijk A., Van Rens B., van der Lende T., Taverne M. (2005). Factors affecting duration of the expulsive stage of parturition and piglet birth intervals in sows with uncomplicated, spontaneous farrowings. Theriogenology 64, 1573–1590. doi: 10.1016/j.theriogenology.2005.03.017
van Rens B. T., van der Lende T. (2004). Parturition in gilts: duration of farrowing, birth intervals and placenta expulsion in relation to maternal, piglet and placental traits. Theriogenology 62, 331–352. doi: 10.1016/j.theriogenology.2003.10.008
Vasdal G., Østensen I., Melišová M., Bozděchová B., Illmann G., Andersen I. L. (2011). Management routines at the time of farrowing—effects on teat success and postnatal piglet mortality from loose housed sows. Livest. Sci. 136, 225–231. doi: 10.1016/j.livsci.2010.09.012
Vázquez-Gómez M., García-Contreras C., Astiz S., Torres-Rovira L., Fernández-Moya E., Olivares Á., et al. (2020). Piglet birthweight and sex affect growth performance and fatty acid composition in fatty pigs. Anim. Prod. Sci. 60, 573–583. doi: 10.1071/AN18254
Vodolazska D., Feyera T., Lauridsen C. (2023). The impact of birth weight, birth order, birth asphyxia, and colostrum intake per se on growth and immunity of the suckling piglets. Sci. Rep. 13, 8057. doi: 10.1038/s41598-023-35277-3
Wahner M., Fisher K. (2005). Current physiological aspects of fetal growth and parturition in the pig. Biotechnol. Anim. Husbandry 21, 135–148.
Walls A., Hatze B., Lomax S., Bathgate R. (2022). Defining “Normal” in pig parturition. Animals 12, 2754. doi: 10.3390/ani12202754
Wu W., Wang X., Wu G., Kim S., Chen F., Wang J. (2010). Differential composition of proteomes in sow colostrum and milk from anterior and posterior mammary glands. J. Anim. Sci. 88, 2657–2664. doi: 10.2527/jas.2010-2972
Wu P., Wang K., Zhou J., Chen D., Yang Q., Yang X., et al. (2019). GWAS on imputed whole-genome resequencing from genotyping-by-sequencing data for farrowing interval of different parities in pigs. Front. Genet. 10. doi: 10.3389/fgene.2019.01012
Wülbers-Mindermann M., Algers B., Berg C., Lundeheim N., Sigvardsson J. (2002). Primiparous and multiparous maternal ability in sows in relation to indoor and outdoor farrowing systems. Livest. Prod. Sci. 73, 285–297. doi: 10.1016/S0301-6226(01)00257-3
Yang K. Y., Jeon J. H., Kwon K. S., Choi H. C., Kim J. B., Lee J. Y. (2019). Effect of different parities on reproductive performance, birth intervals, and tail behavior in sows. J. Anim. Sci. Technol. 61, 147. doi: 10.5187/jast.2019.61.3.147
Yatabe Y., Iida R., Piñeiro C., Koketsu Y. (2019). Recurrence patterns and lifetime performance of parity 1 sows in breeding herds with different weaning-to-first-mating intervals. Porcine Health Manage. 5, 15. doi: 10.1186/s40813-019-0122-0
Yin G., Liu H., Li X., Quan D., Bao J. (2016). Effect of farrowing environment on behaviour and physiology of primiparous sows with 35-day lactation. Int. J. Appl. Res. Vet. Med. 14, 159–169.
Yun J., Swan K.-M., Farmer C., Oliviero C., Peltoniemi O., Valros A. (2014). Prepartum nest-building has an impact on postpartum nursing performance and maternal behaviour in early lactating sows. Appl. Anim. Behav. Sci. 160, 31–37. doi: 10.1016/j.applanim.2014.08.011
Yun J., Swan K.-M., Vienola K., Farmer C., Oliviero C., Peltoniemi O., et al. (2013). Nest-building in sows: Effects of farrowing housing on hormonal modulation of maternal characteristics. Appl. Anim. Behav. Sci. 148, 77–84. doi: 10.1016/j.applanim.2013.07.010
Zaremba W., Udluft T., Bostedt H. (2015). Effects of various procedures for synchronisation of parturition in sows. Tierärztliche Praxis Ausgabe G: Großtiere/Nutztiere 43, 269–277. doi: 10.15653/TPG-150345
Keywords: sow, gilt, primiparous, neonate, farrowing, endocrinology, behavior, vitality
Citation: Corrales-Hernández AA, Roldán-Santiago P, Bonilla-Jaime H, De la Cruz-Cruz LA, Limón-Morales O, Orozco-Gregorio H and Pineda-Reyes R (2025) A review: influence of the sow’s parity on farrowing and neonate performance. Front. Anim. Sci. 6:1568955. doi: 10.3389/fanim.2025.1568955
Received: 31 January 2025; Accepted: 26 March 2025;
Published: 24 April 2025.
Edited by:
Vishal Suthar, Gujarat Biotechnology University, IndiaReviewed by:
Argana Ajay, Indian Veterinary Research Institute (IVRI), IndiaYosua Kristian Adi, Gadjah Mada University, Indonesia
Venkataramanan Ragothaman, Tamil Nadu Veterinary and Animal Sciences University, India
Copyright © 2025 Corrales-Hernández, Roldán-Santiago, Bonilla-Jaime, De la Cruz-Cruz, Limón-Morales, Orozco-Gregorio and Pineda-Reyes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricia Roldán-Santiago, cGF0cmljaWFyb2xkYW5AZm12ei51bmFtLm14; Héctor Orozco-Gregorio, Z29oZWN0b3I3MkB5YWhvby5jb20ubXg=