
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Anim. Sci. , 18 February 2025
Sec. Animal Nutrition
Volume 6 - 2025 | https://doi.org/10.3389/fanim.2025.1540495
 Márcia S. V. Salles1*
Márcia S. V. Salles1* Fábio J. F. Figueiroa2
Fábio J. F. Figueiroa2 Carla M. M. Bittar3
Carla M. M. Bittar3 Viviani Gomes4Raquel S. Marques4
Viviani Gomes4Raquel S. Marques4 Julia A. G da Silveira5Elias J. Facury Filho5José Esler de Freitas6
Julia A. G da Silveira5Elias J. Facury Filho5José Esler de Freitas6 Arlindo Saran Netto2
Arlindo Saran Netto2Introduction: The management and nutrition of young calves are important for the production system due to their great impact on future milk production potential. This study was to evaluate the performance and health of calves in the suckling phase, with supplementation of selenium, iron, and vitamin E while undergoing immunological challenge with Anaplasma marginale.
Methods: An experiment was conducted on an experimental farm using 42 newborn Holstein male calves, aged up to 60 days, in a completely randomized design, and they were assigned to the following three treatments: C (control milk replacer, N = 14); SeVitE (milk replacer supplemented with 0.6 mg organic selenium/kg + 100 IU vitamin E, N = 14); SeVitEFe (milk replacer supplemented with 0.6 mg organic selenium/kg + 100 IU vitamin E + 200 mg Fe chelate/kg, N = 14).
Results: Selenium, iron, and vitamin E intakes were higher in supplemented animals. The mean blood count of Anaplasma marginale was lower in supplemented calves compared to controls and increased with time after inoculation. Increased serum selenium was observed in supplemented calves, without vitamin E and iron changes. Plasma lactate concentration was lower in supplemented animals and those with SeVitEFe had lower urea concentration at 60 days compared to control. Treatments did not influence hematological parameters, while some only changed with the age of animals. Plasma glutathione peroxidase concentrations were higher in supplemented animals, with the interaction between treatment and time, where higher concentrations were observed at 40 days of experiment for supplemented calves compared to control. Treatments did not influence the performance of animals. Animals with SeVitEFe showed a lower incidence of diarrhea in the first thirty days of the experiment.
Discussion: Supplementation of selenium, iron, and vitamin E showed effectiveness in improving the animals’ oxidative metabolism, altering biochemical and hematological parameters, and reducing the pathogens of Anaplasma marginale, and the incidence of diarrhea, but did not enhance the animals’ performance.
The suckling phase, which encompasses the period from birth to weaning, is critical for calves, and mortality and morbidity rates can be high, compromising the efficiency of the dairy cattle production system.
At birth, calves have developed but still immature immune systems. The synepitheliochorial placenta of cattle does not allow the passage of immunoglobulins and other serum proteins (Reis et al., 2007). Soon after birth, calves are challenged by the contaminated environment with harmful viruses and bacteria, as well as by the climate and other external factors (Reis et al., 2007). At birth, they acquire maternal immunoglobulins through the consumption of colostrum, a process called passive immunity transfer, and thus can respond to infections in the first weeks of life (Uyama et al., 2022).
Care and nutrition during the early phase are crucial for the growth and future milk production potential of replacement heifers. Certain nutrients, such as selenium, iron, and vitamin E, are essential for the development of the animal’s immune system. Selenium is an essential micronutrient in ruminant nutrition, being part of the glutathione peroxidase enzyme, an important component in the cellular antioxidant system (NRC, 2001). In addition, as it is involved in the arachidonic acid metabolism, it improves the ability of neutrophils to eliminate pathogenic microorganisms when it is supplemented (NRC, 2001). Like selenium, Vitamin E is involved in inter and intracellular antioxidants (NRC, 2001). In recent years, vitamin E has been shown to play an important role against damage caused by free radicals and in improving the immune system response (McDowell, 2000). Iron is a component of the heme group, a component of enzymes participating in the electron transport chain, and is also found in hemoglobin and myoglobin. Its deficiency results in hypochromic microcytic anemia due to the inability to produce hemoglobin, associated with depressive immune responses. As a result, calves become lethargic, and feed intake and weight gain are reduced (NRC, 2001). The current recommendations of these nutrients for dairy calves in milk replacer are 0.3 mg of selenium/kg of dry matter (DM), 85 mg of iron/kg of DM, and 50 IU of vitamin E/kg of DM for animals between 35 to 125 kg of live weight.
According to the U.S. Department of Agriculture (2015), around 1.9 million calves were lost due to non-predator causes in 2015 in the United States; the main causes being respiratory problems (26.9%), followed by losses related to birth problems (17.8%) and digestive problems (15.4%). The percentages of sick calves during the suckling phase in Brazil are on average 43% for diarrhea, 21% for pneumonia, and 7% for parasitic sadness (Azevedo et al., 2024). In an evaluation carried out in 1,188 rural properties in Brazil, the survival rate of calves up to one year of age was 86.47%, with a still worrying mortality rate of 13.26% (IDEAGRI, 2021). When evaluating the main causes of calf mortality, among the 11,000 deaths that occurred in 69,000 calves evaluated, 21% were due to diarrhea, 20.43% were due to tick fever, and 14.20% were due to pneumonia (IDEAGRI, 2019). Bovine tick fever is composed of two diseases caused by protozoa called babesiosis (caused by Babesia Bigemina and Babesia Bovis) and anaplasmosis (caused by Anaplasma Marginale), which are transmitted by ticks and flies, has and have a major impact on livestock in several countries, as it compromises animal productivity (Ferreira et al., 2022). The prevalence of A. marginale, and Babesia spp. in Latin America was reported as 48.9% and 39.8% (95% CI: 24.6–57.2%), respectively. The prevalence of A. marginale was higher due to the high prevalence of this agent (67.1%) in Mexico (Ferreira et al., 2022).
Newborn calves, adequately fed with colostrum, supplemented with selenium, iron and vitamin E throughout the suckling phase, can have an improved health status in this critical phase, since these nutrients support immune system activity, reduce the incidence of diseases, and consequently improve performance. This study aims to verify the effects of supplementation with selenium, iron, and vitamin E, on the incidence of diarrhea, oxidative metabolism, blood metabolites, hematological parameters, and performance of calves under immunological challenge with Anaplasma marginale in the suckling phase.
The project was approved by the Animal Use Ethics Committee of the Animal Science Institute, with protocol number CEUA/IZ 268-18. We confirm that this study was carried out in compliance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines and all methods were performed following the relevant guidelines and regulations.
The experiment was carried out on the experimental farm of the Animal Science Institute of Ribeirão Preto/SP using 42 male Holstein calves (initial weight 38.33 ± 4.09 kg) for 60 days. Calves were acquired from a commercial farm with 2 to 6 days of age. At the farm, calves were properly fed with the provision of 4 liters of high-quality colostrum (Brix > 22%) within four hours after birth and then another meal of 2 liters from the second milking. Calves received transition milk from the second milking (6 L per day) on the second and third day of age. After the fourth day of life, they received 6 liters of milk a day until they were transported to the farm where the experiment was carried out.
On the day of arrival at the experimental farm, calves remained for 12 hours in a collective pen, with wood shavings bedding and heating with infrared light lamp, and received 2 liters of oral hydration (Calf Care®, De Laval) per animal via bottle. After this period, animals were allocated to individual tropical hutches, with sand bedding, and individual access to starter and drinking water. Supplementation began one day after calves arrived at the experimental farm, considered the first day of the experiment, and all received treatments for 60 days.
The design was completely randomized, with calves allocated to the following treatments: C (control milk replacer, N = 14); SeVitE (milk replacer supplemented with 0.6 mg organic selenium/kg + 100 IU vitamin E, N = 14); SeVitEFe (milk replacer supplemented with 0.6 mg organic selenium/kg + 100 IU vitamin E + 200 mg Fe chelate/kg, N = 14). Each calf was considered an experimental unit.
Supplementation was administered through capsules previously manipulated for each treatment and added to the milk replacer individually at the time of its daily preparation in the bucket with the respective bottle for each calf. The sources of supplemented nutrients were amino acid selenium (YES - Mineral Selenium, 5,000 mg/kg), ferrous glycine hydrate chelate (BASF - Iron Glycinate, 22%), and alpha-tocopherol acetate (LUTAVIT E 50, BASF, 500,000 IU/kg). The amount for capsules was calculated to ensure that the animals received at least 0.6 mg of Se; 200 mg of Fe and 100 IU of vitamin E (149 mg) per kg of dry matter, considering the amount of the respective nutrients from the intake of calf starter and milk replacer according to the dietary program adopted.
Calves were fed for 60 days, receiving 6 liters of milk replacer/day until 30 days from the beginning of the experiment and 4 liters of milk/day, from 31 to 60 days of the experiment. The commercial milk replacer (Sprayfo® Violeta, Trouw Nutrition) was diluted in drinking water preheated to 39°C to reach a dry matter concentration of 14%. The grounded calf starter was offered ad libitum, always in small amounts, being gradually increased according to the animal’s needs. The milk replacer and calf starter composition are shown in Table 1.
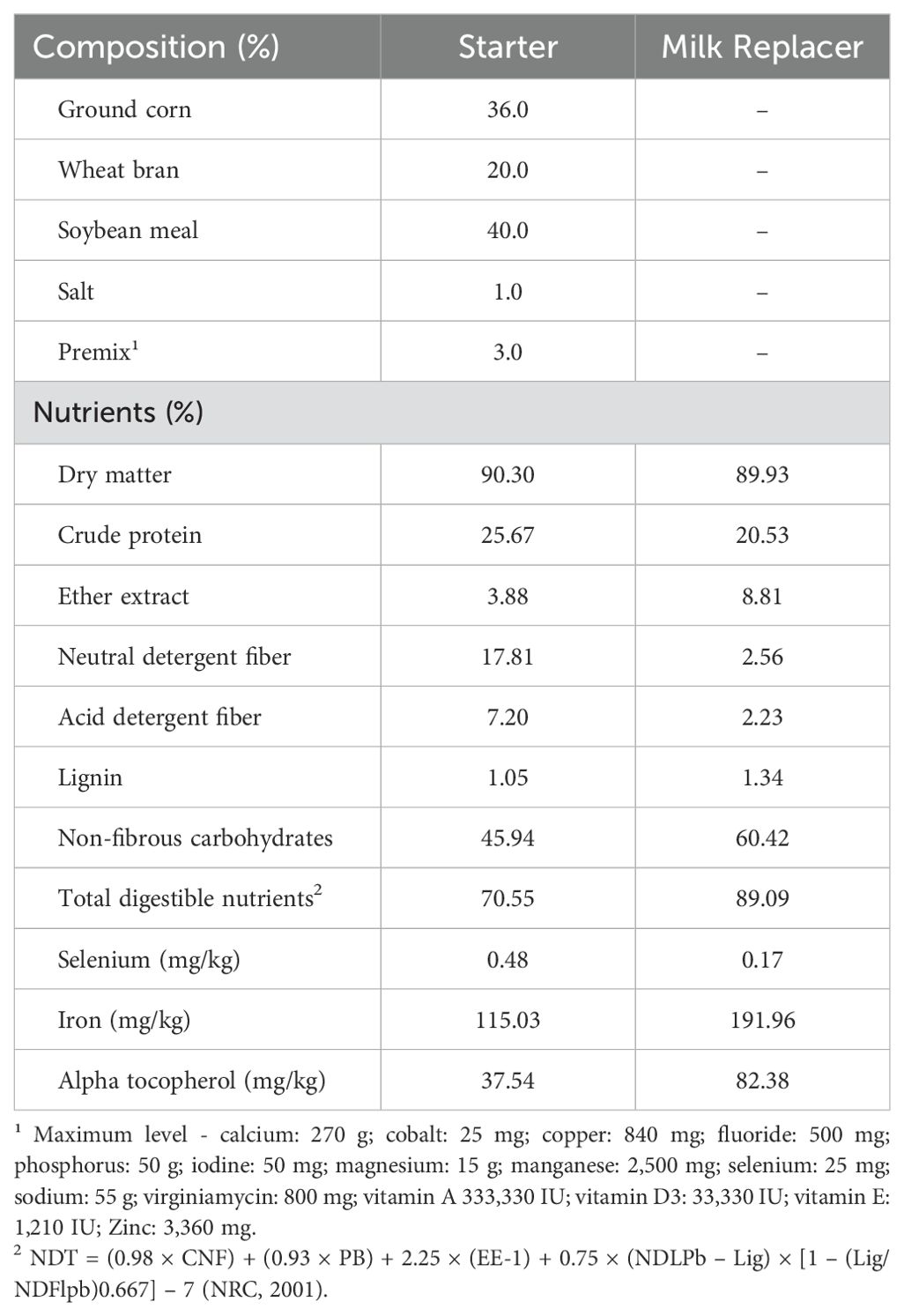
Table 1. Starter ingredients and chemical composition of the ground starter and milk replacer were offered to the calves during the experiment.
The intake of milk replacer and calf starter was daily monitored, recording the volume of the liquid diet consumed and weighing the calf starter offered and the following day’s leftovers.
Water was offered ad libitum to calves throughout the experimental period and had selenium concentration of 0.008 mg/L and iron concentration of 0.010 mg/L.
An immunological challenge was performed on all calves on day 40 of the experiment and inoculation was carried out by applying 1 mL via intramuscularly puncture of low-virulence isolates of Anaplasma marginale (1x107 red blood cells infected, registered in GenBank under codes EU676176 (CEPA UFMG1). Blood samples inoculated with the pathogen (CEPA UFMG1) were kept in liquid nitrogen with dimethyl sulfoxide cryoprotectant until the date of inoculation. On the respective days of inoculation, the inocula preserved in liquid nitrogen in multiple Eppendorf tubes were thawed, and 25 microliters of the inoculum were mixed with 1 mL of the refrigerated inoculation solution in a sterile environment. The final solution was then placed in Eppendorf tubes inside a thermal box with ice and immediately transported to the calf pen, located 30 meters from the laboratory, for inoculation. The calves were always inoculated in the morning after milk management. Blood samples were collected from the jugular vein before inoculation and 20 days after to quantify the activity of the glutathione peroxidase enzyme (GPx), total antioxidant capacity (TAC), and reduced glutathione (GSH).
From the date of the immunological challenge, daily monitoring of rectal temperature was performed at 7:30 AM and 4:30 PM, as well as observation of the eye score and hematocrit measurements.
To certify the success of inoculation and its effectiveness in the study, blood smears were prepared on days 46, 53, and 60 of the experiment for Anaplasma marginale quantification, and blood samples were collected 20 days after inoculation (60 days of the experiment) to perform indirect fluorescent antibody test (IFAT) and PCR for Anaplasma marginale. As the period of clinical manifestation of the disease generally occurs on average 20 to 38 days after the calf has contact with the pathogen, blood was collected on a fixed date of 20 days after inoculation (60 days of experiment) to maintain equal inoculation days among animals. Clinical signs of the disease were considered when animals had rectal temperatures above 40°C and hematocrit below 60% of the basal value of each calf. To obtain the basal hematocrit level of each calf, a blood sample was collected before inoculation and the red cell count was read. No animal showed clinical signs of the disease during the experiment.
Blood samples were collected from calves by jugular puncture on the first day, 40 and 60 days of the experiment, two hours after offering the milk replacer, for analysis of selenium, iron, blood count, ferritin, and the following biochemical parameters: ß-hydroxybutyrate, glucose, total protein, urea, creatinine, creatinine kinase, lactate dehydrogenase (LDH), lactate, albumin, and IgG. For vitamin E analysis, blood samples were collected from animals on the first day and at 60 days of the experiment.
For selenium, iron, ferritin, and vitamin E measurements, blood samples were collected in vacutainer tubes without anticoagulant and centrifuged at 3,000 RPM to obtain serum, stored frozen in plastic tubes. The tubes used for vitamin E analysis were protected with aluminum foil and the serum was stored in amber plastic tubes.
The stool consistency score (0 = normal, well-formed stools; 1 = abnormal, but not yet showing diarrhea, little pasty stools; 2 = moderate diarrhea, pasty stools; 3 = severe diarrhea; very pasty or liquid stools) was daily assessed. Fecal scores 2 and 3 were considered cases of diarrhea. The incidence of diarrhea, presented as a percentage, was calculated according to the following equation:
Calves with a score of ≥ 2, received 4 L of daily oral electrolytes, divided into two meals, two hours after offering a milk replacer and a lactobacilli-based probiotic, until the consistency of the feces reduced the score. Intravenous electrolytes were administered, when necessary until body rehydration occurred. Calves were medicated with parenteral antimicrobials, a combination of trimethoprim and sulfamethoxazole, and non-steroidal anti-inflammatory drugs with the active ingredient flunixin meglumine, as advised by the veterinarian.
Health status and presence of clinical cases of other diseases were daily observed, and daily environmental temperature and relative humidity data were recorded.
All calves were weighed on a digital scale, and measurements of height at the withers, heart girth, hip width, and body length were recorded at the beginning of the experiment, and every 15 days until the end of 60 days. Height measurements were performed using an appropriate ruler, fixed to a right-angled support, measuring the vertical distance between the ground and the withers. Heart girth, hip length, and width measurements were carried out using a measuring tape. Body length was measured by measuring the distance from the scapula to the beginning of the tail insertion. Hip width was measured by measuring the distance between ischia.
Ground calf starter and milk replacer samples were collected weekly and frozen to prepare a monthly pool for chemical analysis, carried out at the Laboratory of Bromatology - FMVZ/USP. Samples were ground in a mill with a 1 mm sieve for the analysis of dry matter (DM, method 930.15), crude protein (CP, method 984.13), ether extract (EE, method 920.39), mineral matter (MM, method 942.05), acid detergent lignin (method 973.18), according to AOAC (1996). Neutral detergent fiber (NDF) and acid detergent fiber (ADF) analyses were also carried out, according to Van Soest et al. (1991) and Goering and Van Soest (1970), respectively.
To calculate non-fibrous carbohydrates (NFC) and total digestible nutrients (TDN), the following NRC (2001) formulas were used:
Water samples were stored in 300-mL plastic bottles and frozen at –6°C for later analysis of selenium and iron (SMEWW, 2017). Selenium and iron analysis of calf starter, milk replacer, water, and mineral supplement were carried out in the Laboratory of Minerals of the Faculty of Animal Science and Food Engineering at USP/Pirassununga. The samples underwent wet digestion with nitric-perchloric acid using an adapted fluorimetric method (Olson et al., 1975), and iron concentration was analyzed using the colorimetric method (Fe liquiform, Labtest kit). Analysis of alpha-tocopherol in the calf starter, milk replacer, and vitamin supplement were carried out at the CBO Laboratory (Valinhos, SP) using the methodology with HPLC (Isocratic, fluorescence detector) and reverse-phase column separation technique, detection at 326 nm emission and 294 nm excitation and quantification by external standard (Horwitz and Larimer, 2005).
Concentrations of GPx, GSH, TAC, and IgG were determined at the FMVZ/USP laboratory. Vacuum-packed total blood samples were collected in two 8.5-mL tubes containing lytic heparin as an anticoagulant for GPx and TAC analyses. Samples were centrifuged at 3,000 rpm for 10 min at 4°C with slow deceleration, with plasma and buffy coat being collected. After the first centrifugation, an aliquot of plasma was removed, stored in amber Eppendorf tubes, and kept frozen at -80 °C. The total antioxidant capacity (TAC) was determined using a commercial kit (RANSEL® Laboratories, Randox, Crumlin, UK) according to the protocol described by the manufacturer. Centrifugation was repeated two more times with phosphate buffered saline (PBS 10%) until the supernatant was completely clear. The red blood cell mash obtained was packaged in amber microtubes and frozen in a freezer at -80°C for subsequent GPx analysis. Serum GPx activity was determined on a Randox automatic biochemical analyzer (model RX Daytona) using Randox® commercial kits (RANSOD and RANSEL) to determine the values and corrected by the hemoglobin concentration measured by spectrophotometry.
To determine GSH, whole blood collected in vacuum tubes containing EDTA was used. In a test tube, 200 μl of whole blood was hemolyzed with 1,800 μl of distilled water, then, 3 mL of precipitating solution (3.34 g of metaphosphoric acid, 0.44 g of EDTA Na2H20, 60 g of NaCl and distilled water q.s.p 200 ml) were added to the hemolysate. The solution remained at rest for 5 minutes and was then centrifuged for 5 minutes at 3,000 rpm. The intermediate content was collected, stored in amber Eppendorf tubes, and kept in a freezer at -80°C until analysis. To determine the GSH concentration, a 200-μl aliquot of the supernatant was added to 800 μl of Na2HPO4 solution (300 mmol/l). After homogenization, 100 μl of 0.05% DTNB (2-dinitrobenzoic acid) solution was added to the system, with reading being performed at 412 nm within 30 seconds after the addition of DTNB. From the standard curve values, the straight-line equation was calculated and the absorbance values of analyzed samples were converted into mg/dL.
The blood smear technique to obtain the number of Anaplasma marginale was performed with 5 µL of blood and using the GIEMSA dye. Blood smears were taken from all calves at 46, 53, and 60 days of the experiment. There was a problem with the storage of these smear slides, and it was decided to keep only slides in excellent readable condition. Due to this fact, the number of slides used to describe the results were 15 (C=4, SeVitE =4, SeVitEFe=7).
For blood collected for PCR analysis, the tube containing EDTA as an anticoagulant and frozen at -20°C was used. For the indirect fluorescent antibody test (IFAT), blood was collected in a tube without anticoagulant, centrifuged and the serum was frozen at -20°C until sent for analysis. Both analyses were carried out at the Laboratory of Veterinary Protozoology/VET/UFMG. The presence of anti-A. marginale antibodies were detected using IFAT (Silveira, 2012) and anti-bovine monoclonal antibody (AbD Serotec) conjugated to fluorescein isothiocyanate (FITC). The antigens used were produced by the laboratory (IICA, 1987) and the strains utilized in this study were A. marginale strain UFMG1 (Bastos et al., 2010). For each reaction, positive and negative control serum was used (Bastos et al., 2010). Samples were considered positive if they showed fluorescence at dilution of 1:40.
Molecular analysis was conducted (Silveira et al., 2016). DNA was extracted from 300 µl of whole blood using a Wizard Genomic DNA Purification Kit (Promega, Madison, WI, USA) following the manufacturer’s instructions. The nested PCR was performed in two stages: conventional PCR followed by nPCR. An aliquot of the first amplified PCR product was used for a second PCR reaction with a second set of primers. These primers were chosen to amplify the target sequence of the first PCR reaction, thereby increasing sensitivity. The following sets of primers were used to detect the A.marginale msp4 gene. First reaction (872 bp): MSP45: 5GGG-AGCTCCTATGAATTACAGAGAATTGTTTAC3 and MSP43: 5CCG-ATCC TTAGCTGAACAGGAATCTTGC3 (de la Fuente et al., 2008); second reaction (294 bp): AnapF 5_CGCCAGCAAACTTTTCCAAA3 and AnapR 5ATATGGG GACACAGGCAAAT3. Twice-distilled water was used as the negative control (noDNA). As a positive control for the amplification of the msp4 gene fragment, DNA extracted from 300 µl of whole blood from a calf experimentally infected with the UFMG1 strain of A. marginale was used (Bastos et al., 2010).
The first reaction mixture comprised 1.2 µl dNTPs (2.5 mM), 0.15 µl Taq polymerase (0.05 U), 1.5 µl reaction buffer IB (1×), 0.6 µl of a solution containing the mixed primers (10 mM), and 10.05 µl sterile ultra-pure water. A 1.5 µl aliquot of the DNA template was added to the reaction mixture to give a final volume of 15 µl. The second reaction mixture comprised 2.0 µl dNTPs (2.5 mM), 0.25 µl Taq polymerase (0.05 U), 2.5 µl reaction buffer IB (1x), 1.0 µl of a solution containing the mixed primers (10 mM) and 16.75 µl sterile ultra-pure water. An aliquot (2.5 µl) of the amplicon obtained from the first reaction was added to the reaction mixture to give a final volume of 25 µl. Amplification was carried out under the conditions described. Amplification was performed using an Eppendorf Mastercycler®thermocycler (Eppendorf, São Paulo, SP, Brazil). The protocol used to amplify A. marginale was as follows: 94 °C for 5 min (initial denaturation step); 30 cycles of 92 °C for 1 min (denaturation), 54 °C for 1 min (annealing), 72 °C for 2 min (extension); and a final extension step at 72 °C for 8 min. Following amplification, the reaction mixtures were maintained at 12 °C. PCR amplicons were separated by electrophoresis on 1% agarose gel (40 min, 100 V), stained with gelred (Biotium, São Paulo, SP, Brazil), and visualized under ultraviolet light.
Selenium and iron analysis in serum were carried out in the same laboratory as those in feed and used the same analysis methodologies.
To perform blood cell count, blood samples were collected using vacutainer tubes containing EDTA as an anticoagulant. Immediately after collection, samples were refrigerated and transported to the veterinary laboratory for analysis. Blood samples were analyzed in an automated pochi-100ivdiff® cell counter, which uses impedance and spectrophotometry techniques. The following parameters were analyzed: erythrocyte, hematocrit, and hemoglobin count, mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), leukocyte count, and platelet count.
Serum ferritin was analyzed by two-site enzyme immunoassay (sandwich), the chemiluminescent substrate (Lumi-Phos* 530) was added to the container and the light generated by the reaction was measured with a luminometer.
Analysis of biochemical parameters were conducted at the Metabolism Laboratory at ESALQ/USP. Blood samples were collected with the aid of a vacuum tube containing sodium fluoride to obtain plasma and tubes without anticoagulants to obtain blood serum. Samples were centrifuged at 3,000 rpm for 15 minutes. Specific commercial enzymatic kits from Labtest Diagnóstica S.A. (Lagoa Santa, MG, Brazil) were used to analyze plasma glucose (Ref. 85), serum total protein (Ref. 99-250), plasma urea (Ref. 104), plasma creatinine (Ref. 35), plasma creatinine kinase (Ref. 96), plasma lactate dehydrogenase (Ref. 86), plasma lactate (Ref. 116), and serum albumin (Ref. 19). The concentration of plasma ß hydroxybutyrate (BHB) using the commercial kit RANBUT (Randox Laboratories, Life Sciences Ltd., Crumlin, United Kingdom). All biochemical metabolites were analyzed in an automatic biochemical system (SBA-200; CELM, Barueri, SP, Brazil).
To estimate the concentration of serum proteins, Megabrix® portable manual refractometer (IONLab Barigui, Brazil) was used. Blood samples were centrifuged in a laboratory centrifuge model CEOO1-B1® - Kacil (Recife, Brazil), at a speed of 2,500 rpm for 5 minutes.
The quantity of serum IgG samples from the calves was measured using a sandwich ELISA, according to the procedures previously published (Reber et al., 2008). Briefly, rabbit anti-bovine IgG antibody (capture antibody, B5645; Sigma, St. Louis, MO) diluted 1:400 in sodium carbonate buffer was used to coat Immulon 4HBX plates (Thermo Corp., Milford, MA) at 4–8 °C overnight. After, all plates were washed three times using the wash buffer. For this assay, plasma samples were diluted 1:1 000 000. The samples and a standard dilution of bovine gamma globulin preparation were placed in duplicate wells and incubated for 1 h at 20–26 °C. The plates were washed three times. Bound IgG was detected using a rabbit anti-bovine IgG conjugated to horseradish peroxidase (detection antibody, A5295; Sigma, St. Louis, MO). The detection antibody was diluted 1:1000 and incubated for 30 min. The plates were washed three times. The quantity of bound detection antibody was measured using the substrate 2,20-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS, A-9941; Sigma, St. Louis, MO) containing 20 µL of 30% hydrogen peroxide per 11 mL ABTS (preconditioned at RT for 15 minutes) and incubated 30 min. The color signal was measured using a plate reader with a 405 nm filter. An immunoglobulin standard (commercial bovine gamma globulin, I5506; Sigma, St. Louis, MO) was prepared as a serial dilution with a range of 50–0.39 ng/mL. The intra-test coefficient of variation was 11.0% and the inter-test coefficient was 13.1%.
For serum alpha-tocopherol analysis, samples were saponified to hydrolyze and solubilize fats and proteins and extracted with hexane. Serum alpha-tocopherol analyses were performed using high-performance liquid chromatography with a fluorescence detector (Arnaud et al., 1991).
Results were submitted to statistical analysis according to a completely randomized design using PROC MIXED from SAS 9.4 (SAS, 2013) according to the model below:
Where Y ij = dependent variable; µ = overall average; Di = diet effect (i = 1 to 3); Aj(D)k = random effect of the animal within treatment (k =1 to 3); εijk = assumed random error NID ~ (0,σ2).
Blood variables were analyzed as repeated measures over time, verifying the normality of residuals and homogeneity of variances by PROC UNIVARIATE procedure. The model collected over time according to the previous design considering the physiological stage of calf growth or weeks of collection (time, 0, 40, and 60 days), diets (Control, SeVitE, SeVitEFe), and the interaction between time and treatment using PROC MIXED from SAS, according to the following model:
Where Yijklm is the dependent variable value, μ is the overall mean, Di is the fixed effect of diet l (i = 1–3), Aj(D)k, is the random effect of the animal within of diet, εijk is the residual error. Tm, is the fixed effect of sampling time (m= 1–3), Tm×Di is the fixed interaction effect between sampling time and diet, and ωijklm is the random experimental error. All means were obtained using LSMEANS. The Akaike information criterion (AIC) was used to determine the structured covariance matrix. Degrees of freedom were defined according to Kenward and Roger (1997). All means were estimated by the LSMEANS procedure and compared by the Tukey test, considering 0.05 as the probability of type-I error. Treatment responses were tested and a significance level of 0.05 was adopted.
The dry matter intake of the feeds and total dry matter intake were not different among treatments. However, the intakes of selenium, iron, and vitamin E, were higher in supplemented animals throughout the experimental period compared to control treatment (P< 0.03, Table 2).
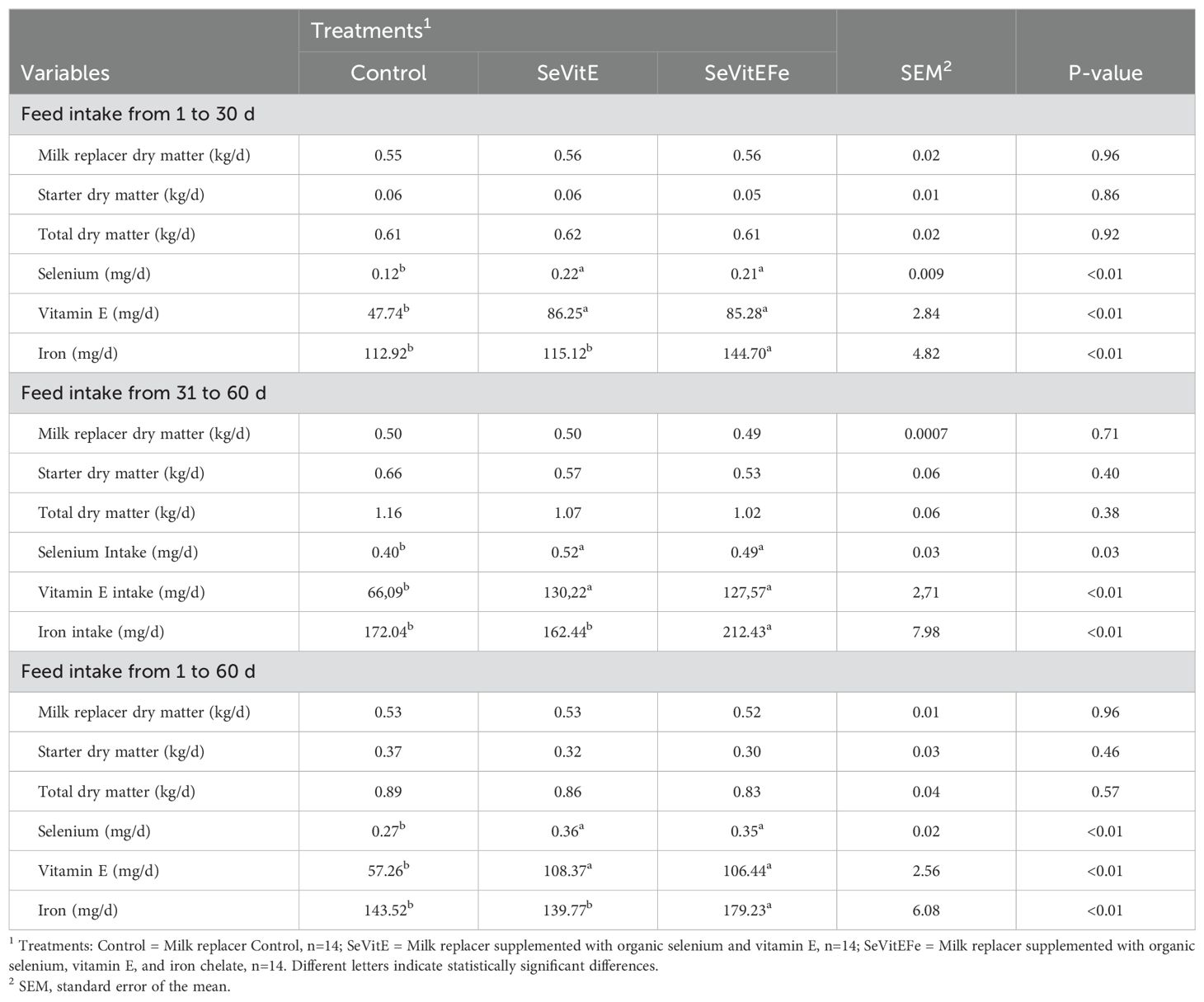
Table 2. Dry matter intake and of micronutrients by calves supplemented with different micronutrients during the experimental period.
Regarding oxidative metabolism, glutathione peroxidase activity was higher in supplemented compared to control calves (P=0.03, Table 3). Interaction between treatment and time was observed (P<0.01, Figure 1), with higher glutathione concentrations in the plasma of supplemented calves after 40 days of experiment compared to control. Reduced glutathione and total plasma antioxidant activity were not different among treatments (P>0.05).
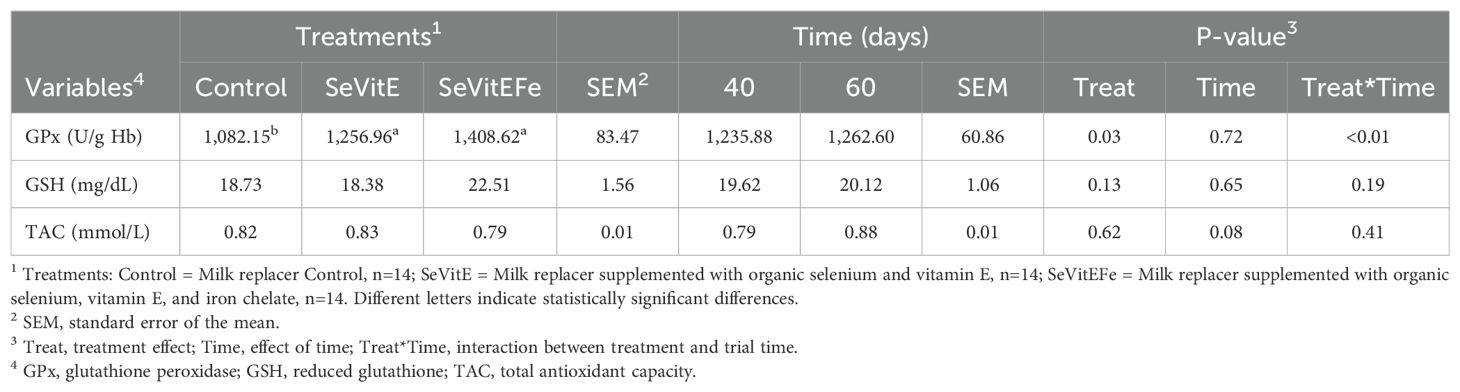
Table 3. Blood oxidative metabolism during immunological challenge of calves supplemented with micronutrients.
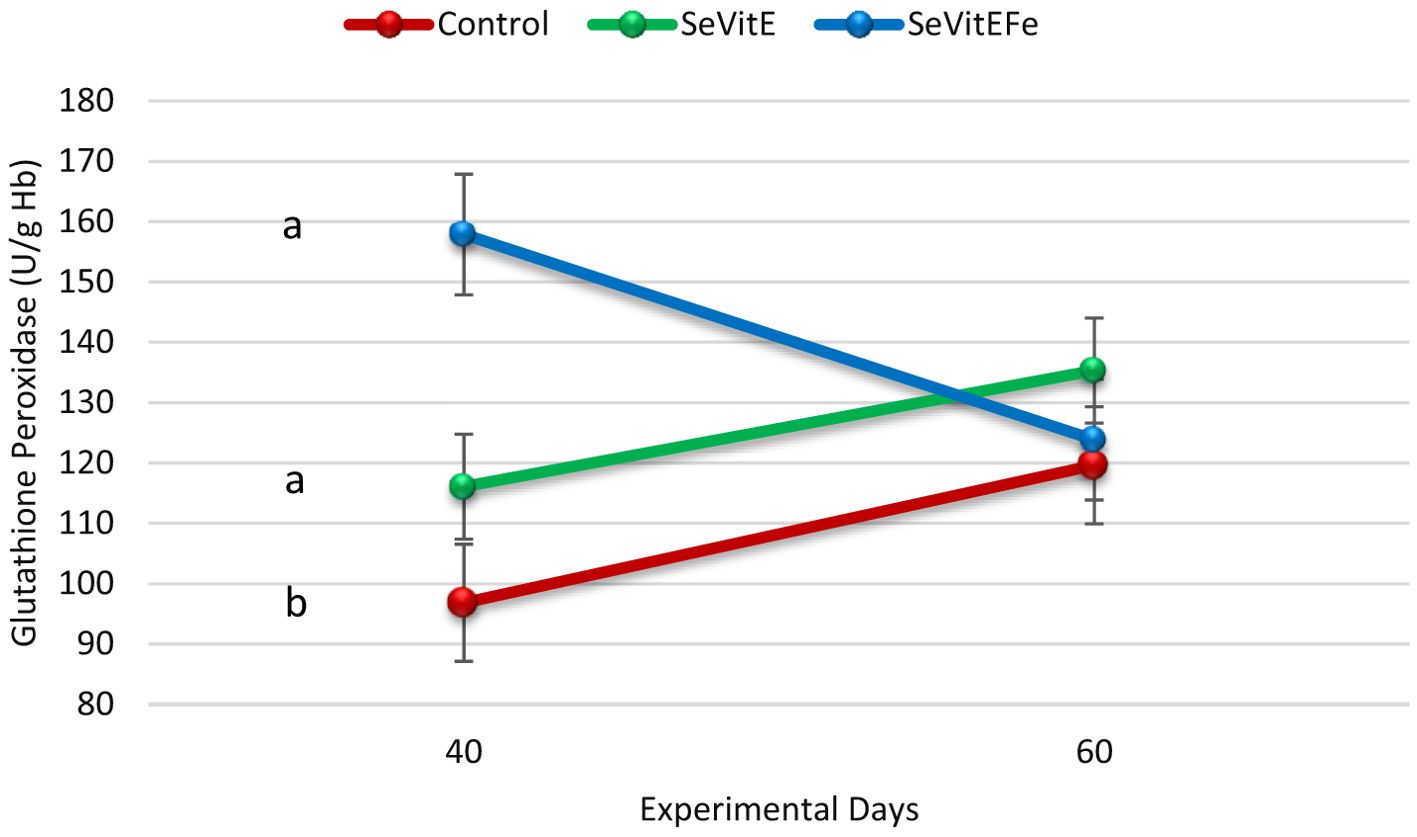
Figure 1. Glutathione peroxidase concentration in the blood plasma of calves in the different treatments (Control = Milk replacer Control, N=14; SeVitE = Milk replacer supplemented with organic selenium and vitamin E, N=14; SeVitEFe = Milk replacer supplemented with organic selenium, vitamin E, and Iron chelate, N=14) on the days of experiment (Different letters indicate statistically significant differences, treatment and time interaction effect, P<0.01, SEM = 6.62).
The mean numbers/mL of Anaplasma marginale were lower in supplemented calves compared to controls (P = 0.0408, SEM = 17.02, Figure 2) and increased with time after inoculation (P = 0.0408, SEM = 17.39, Figure 3).
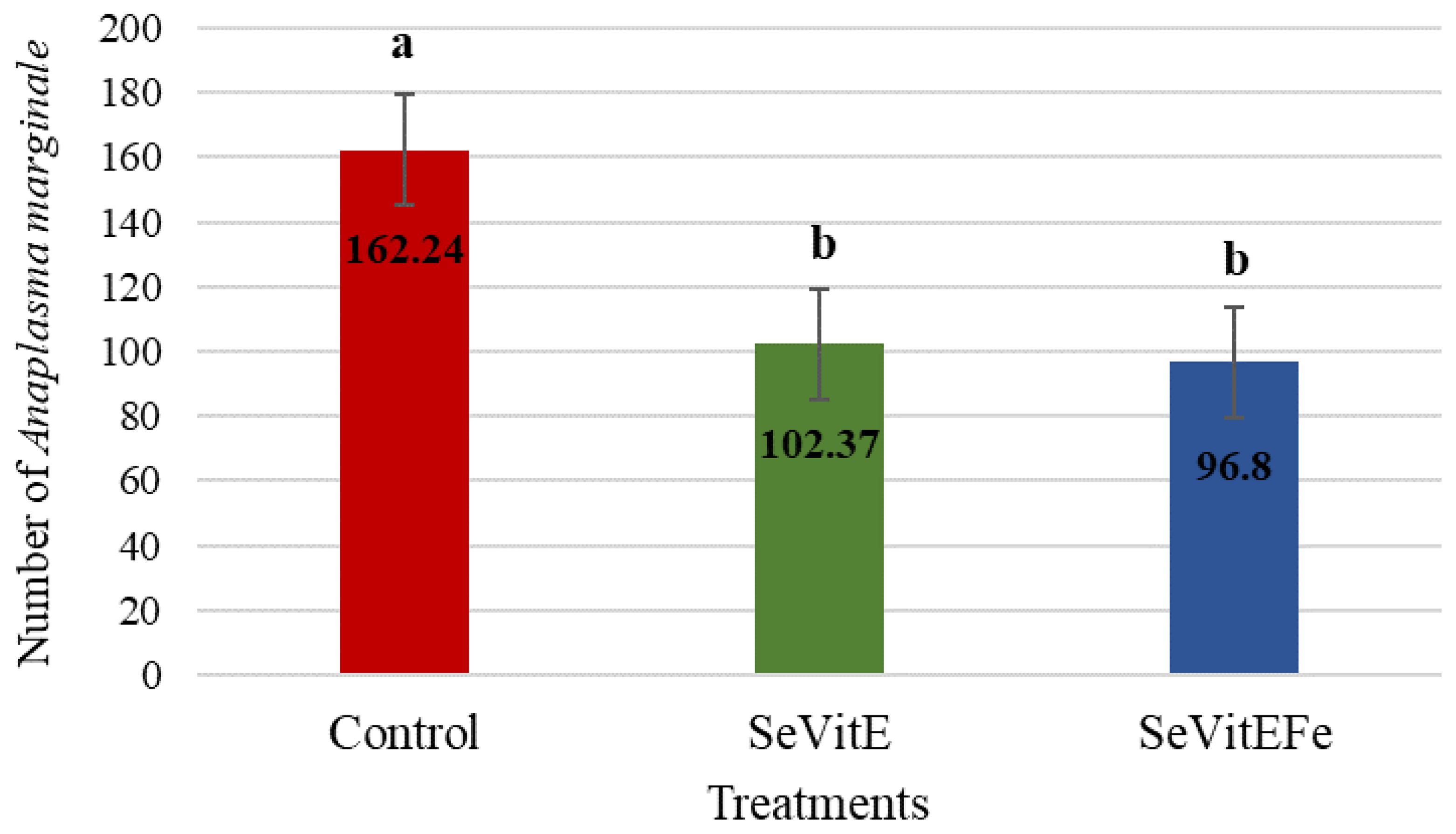
Figure 2. Average number of Anaplasma marginale in treatments (Control = Milk replacer Control; SeVitE = Milk replacer supplemented with organic selenium and vitamin E; SeVitEFe = Milk replacer supplemented with organic selenium, vitamin E, and Iron chelate) at 46, 53, 60 days of experiment (Different letters indicate statistically significant differences, P= 0.0408, SEM = 17.02).
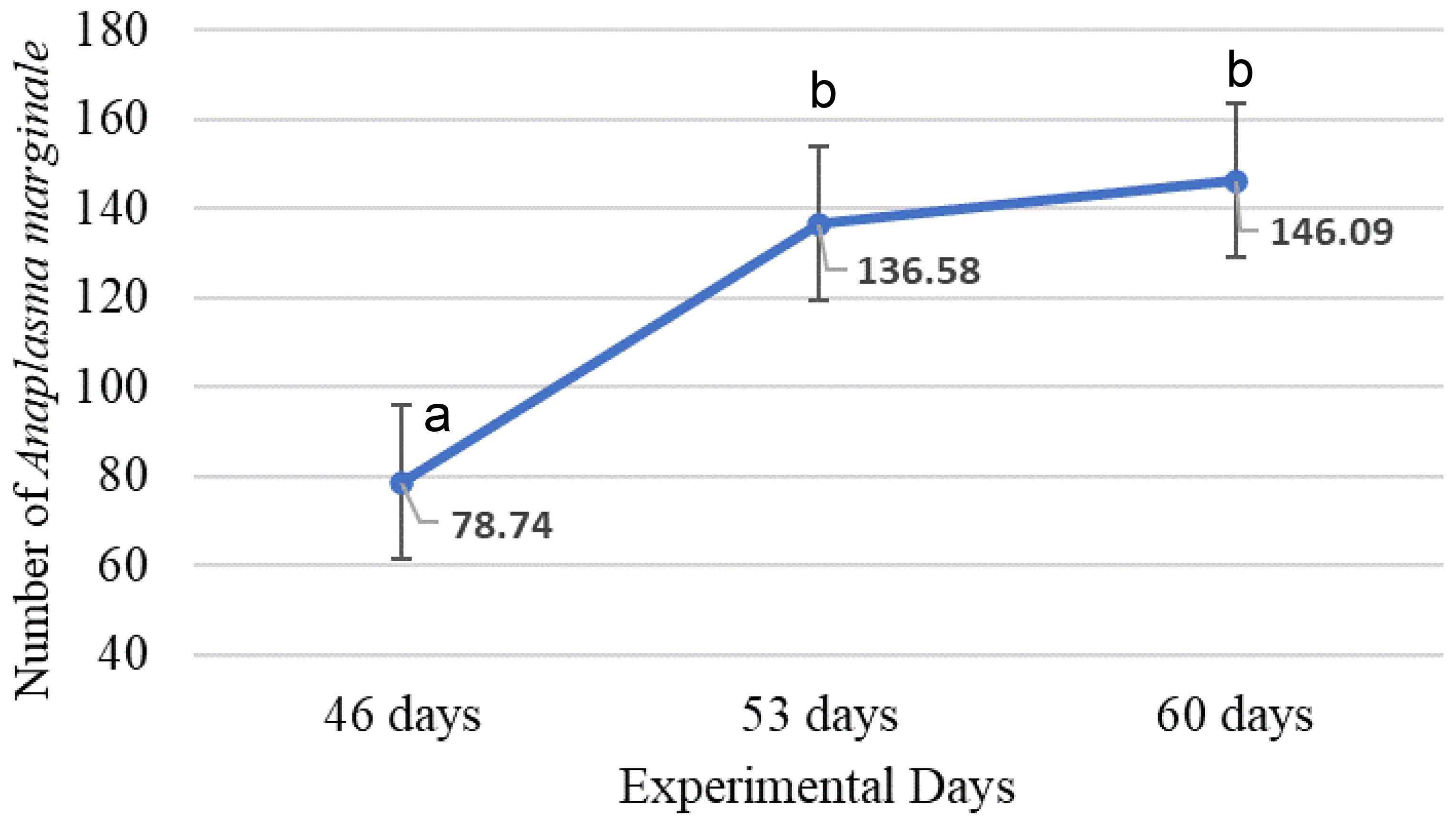
Figure 3. Mean number of Anaplasma marginale at 46, 53 and 60 days of experiment (Different letters indicate statistically significant differences, P = 0.0353, SEM = 17.39).
The IFAT analyses in the serum of the calves were all reactive for serology and the PCR were all positive for Anaplasma marginale.
Increased selenium concentration in the serum of supplemented calves (P<0.01) compared to control was observed (Table 4). However, vitamin E (P=0.57) and iron (P=0.59) concentrations in the serum were not different among calves. Plasma lactate dehydrogenase (LDH) was lower in calves supplemented with SeViEFe (P=0.01) and lactate was lower in supplemented animals (P=0.03) compared to control.
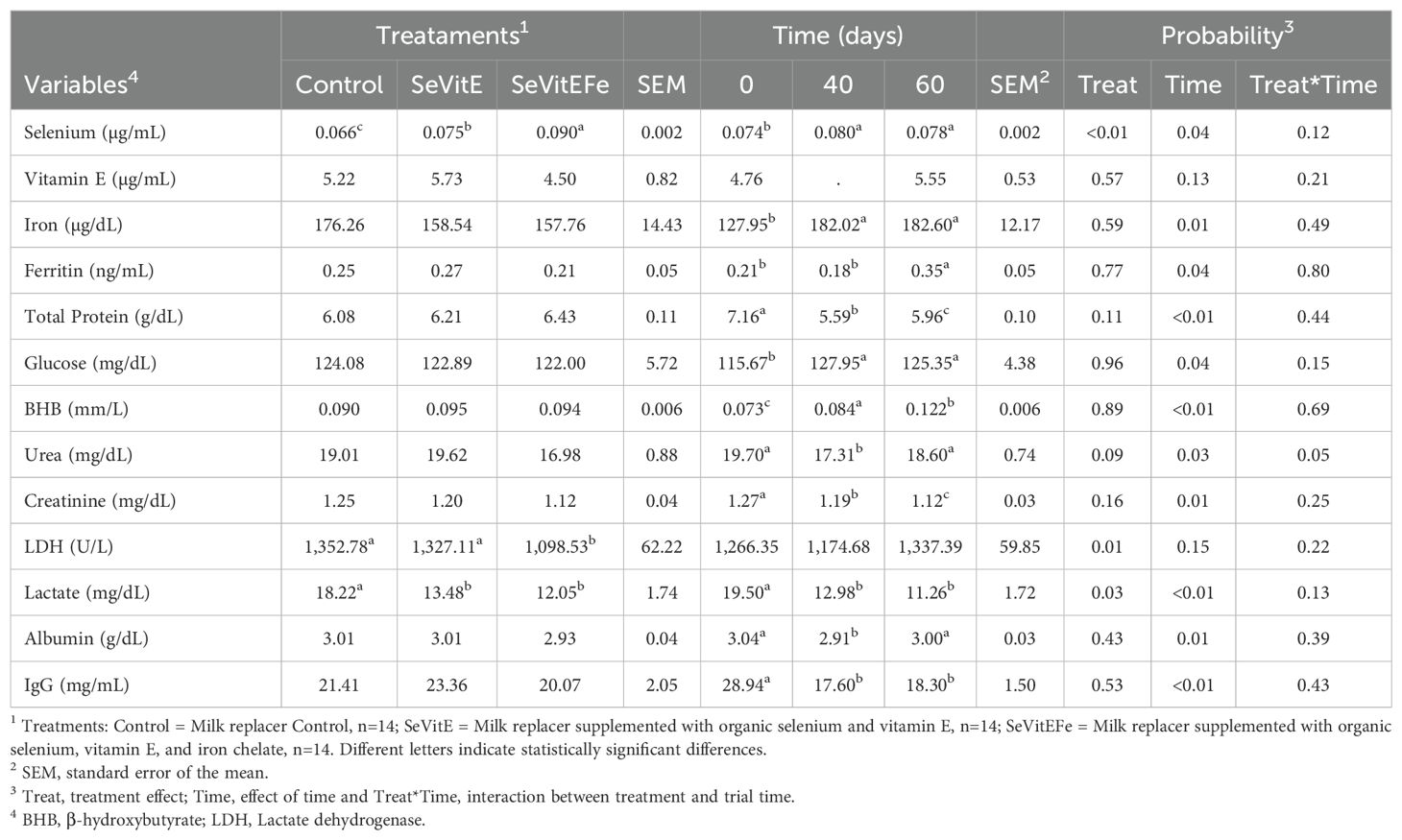
Table 4. Blood metabolites of calves in the different treatments and during the experimental period.
Blood selenium, vitamin E, glucose, and beta-hydroxybutyrate concentrations increased during the experiment (P<0.05, Table 4). Ferritin was higher at 60 days compared to previous periods (P=0.04). Total protein decreased from the beginning of the experiment until 40 days, then increased, but at lower values compared to the initial value (P<0.01). There was an interaction between treatment and experimental time on plasma urea concentrations (P=0.05, Figure 4), and it was observed that the values among treatments were very close at the beginning of the experiment and 40 days of supplementation, but there was a difference at 60 days when animals supplemented with SeVitEFe showed a lower concentration compared to control animals. Plasma creatinine, lactate, albumin, and IgG concentrations decreased during the experimental period (P<0.01).
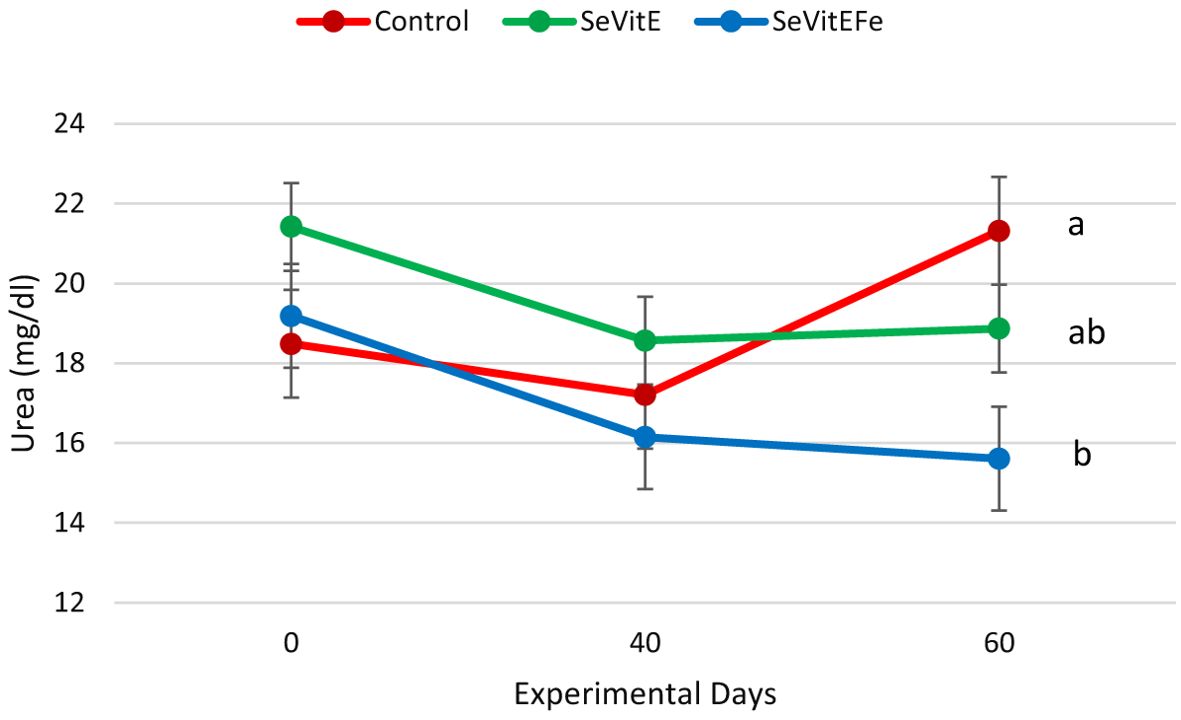
Figure 4. Urea concentration in the blood serum of calves in the different treatments (Control = Milk replacer Control, N=14; SeVitE = Milk replacer supplemented with organic selenium and vitamin E, N=14; SeVitEFe = Milk replacer supplemented with organic selenium, vitamin E, and Iron chelate, N=14) on the days of the experiment (Different letters indicate statistically significant differences, interaction effect of treatment and time, P=0.05, SEM = 0.67).
The other plasma concentrations of metabolites were not affected by supplementation. None of the other blood metabolite variables had a significant interaction between treatment and time.
Mean corpuscular hemoglobin (P=0.05) and its concentration (P=0.04) decreased from the beginning to 40 days of supplementation and increased at 60 days to values similar to the initial ones. The percentage of eosinophils (P<0.01) increased from the beginning to 40 days of supplementation and decreased at 60 days. Platelet count decreased during the experiment (P<0.01, Table 5).
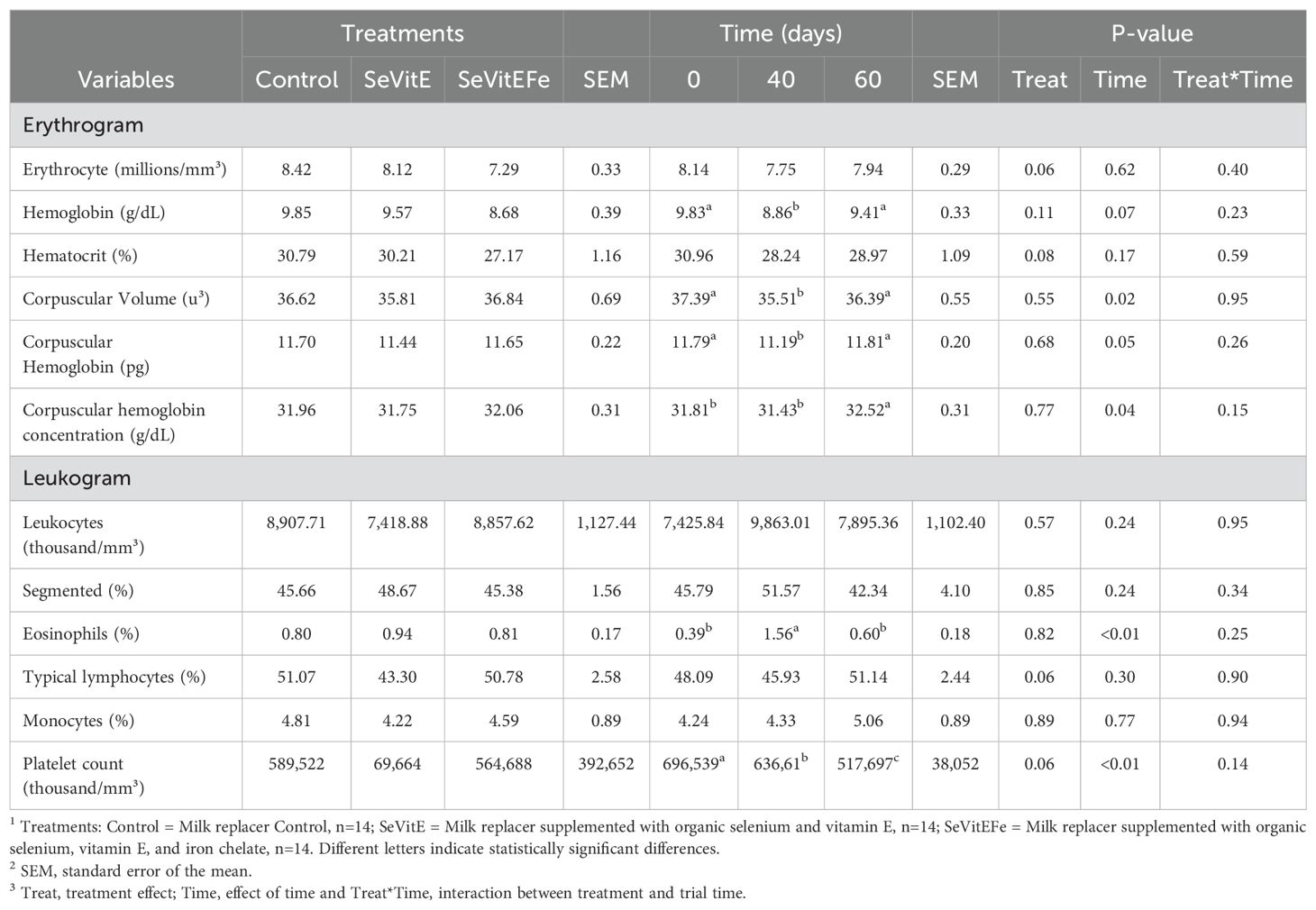
Table 5. Blood count of calves in different treatments and during the experimental period.
Animals supplemented with SeVitEFe had a lower incidence of diarrhea (P = 0.03) compared to the other treatments in the first 30 days of supplementation (Table 6), with no change in stool consistency scores during the experiment.
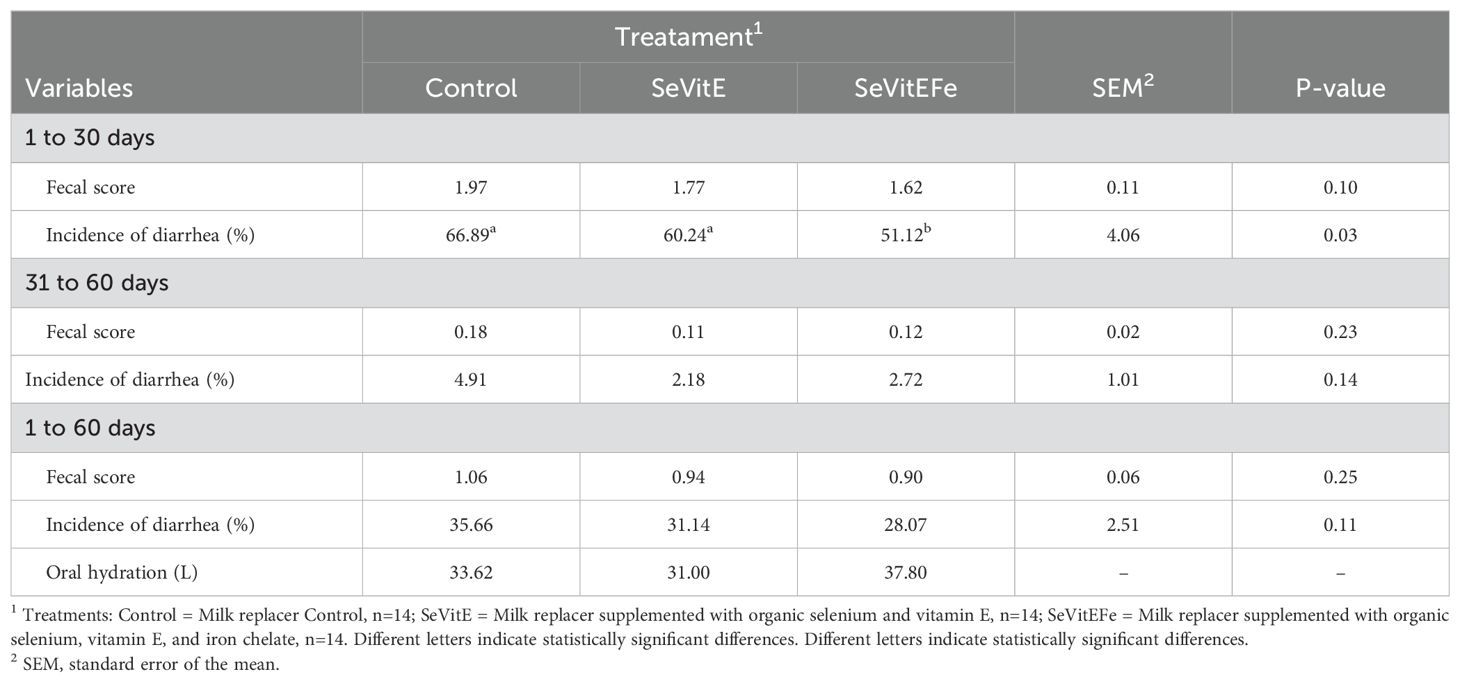
Table 6. Average fecal score and incidence of diarrhea in calves in different treatments during the first 30 days, from 31 to 60 days, and throughout the experimental period.
Performance and feed efficiency variables were not different among treatments (Table 7).
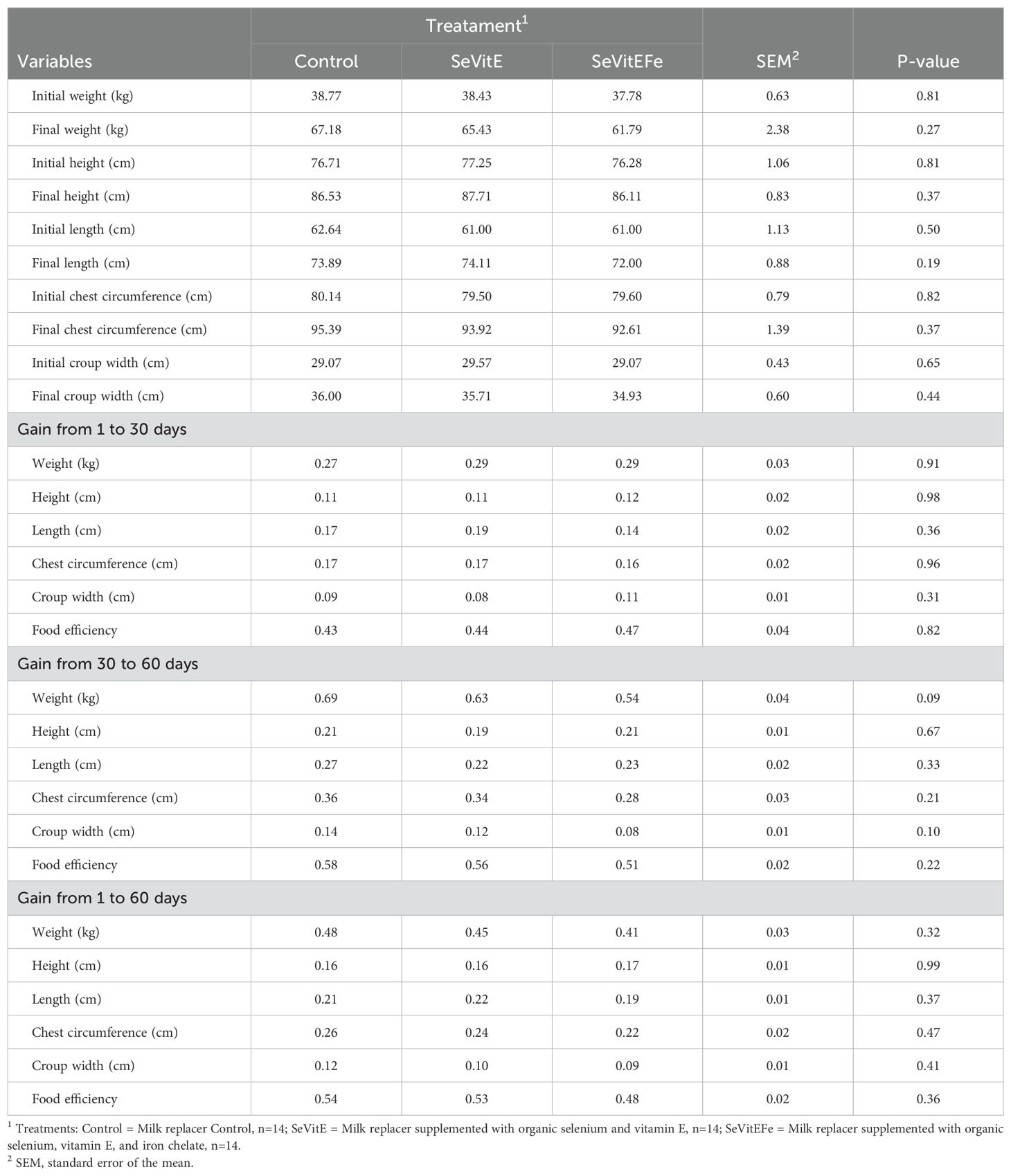
Table 7. Performance of calves in different treatments and during the experimental period.
Selenium, iron, and vitamin E intake were increased in supplemented animals throughout the experimental period and changes in the concentrations of blood metabolites of animals were observed, indicating their supportive role against pathogens. Selenium is a cofactor necessary for the function of the glutathione peroxidase enzyme (Goff, 2018), which plays a unique role in the battle against oxidative stress. The increased selenium intake in supplemented calves resulted in higher plasma selenium and glutathione peroxidase concentrations. The glutathione concentration in supplemented calves before the immunological challenge was on average 19.26 U/g Hb (20%) for SeVitE and 61.06 U/g Hb (63%) for SeVitEFe, more than in control animals. Glutathione is an important antioxidant that can prevent cellular damage caused by various oxidative stressors (Wangsanut and Pongpom, 2022), a can for instance protects hemoglobin from oxidative degradation and inhibits inflammation (Brigelius-Flohé and Flohé, 2020). We can infer that with a greater amount of glutathione in the blood of SeVitEFe calves, there was better systemic support to protect from the pathogen Anaplasma marginale, since SeVitE and SeVitEFe supplemented calves had a lower blood pathogen count. There was an increase in the blood pathogen count after inoculation for all calves since they received the same amount of Anaplasma marginale during inoculation. The initial corpuscles of this pathogen, when inoculated into cattle, adhere to erythrocytes and multiply and invade other erythrocytes. Supplemented calves presented the lowest blood count of pathogens indicating that the immune system of these animals was better able to cope with the infection. Additionally, calves supplemented with SeVitEFe performed more effectively to bacterial challenges, with a lower incidence of diarrhea than animals maintained on control diets, proving the immunostimulant effect of the micronutrients. In the literature, there are no reports to date of a specific interaction between iron metabolism with selenium and vitamin E (Ensley, 2020; NASEM, 2021). There may be an interaction between these nutrients during supplementation, particularly when the physiological state differs from deficiency or marginal supplementation. Further research is needed to explore this interaction in greater depth.
Supplementation with vitamin E, iron, and selenium increased the selenium concentration in the serum of animals, with no effect on Fe and Vitamin E. There are few studies reporting the effect of selenium and vitamin E supplementation on calves during the suckling phase. There are studies in literature assessing Se and vitamin E supplementation in peripartum cows and the concentration of these nutrients in the colostrum and blood of calves in the first days of life, that is, newborns (Abdelrahman and Kincaid, 1995; Gunter et al., 2003; Jaaf et al., 2020; Kamada et al., 2007; LeBlanc et al., 2002; Ranches et al., 2017). Calves that were supplemented with 0.80 mg of organic Se/day during the suckling phase, presented an increase in the selenium concentration in the serum (0.068 µg/ml) compared to control animals (0.051 µg/ml) and increased phagocytic activity of macrophages (Salles et al., 2014). Selenium glyceride was offered orally on the second day of life to calves at doses of 0.5 or 1 mg of Se/kg of body weight and blood parameters were monitored until the fourteenth day of life (Żarczyńska et al., 2021). An increase in serum Se from 0.063 to 0.184 µg/mL at the dose of 0.5 mg/kg BW, and of 0.200 µg/mL with 1 mg/kg was observed on the second day. However, the blood glutathione peroxidase concentration only increased after ten days of supplementation, with calves receiving a dose of 0.5 mg/kg BW having values of 180 U/gHb and those receiving 1 mg Se/kg having 220 U/gHb, compared to control animals (130 U/gHb) (Żarczyńska et al., 2021). It was observed that with an increase in the selenium supply, there is also an increase in the amount of glutathione peroxidase. These authors worked with higher supplementation doses, compared to the present study, and reported that it was effective in improving Se status, and that the biochemical parameters monitored did not indicate a negative effect on the functioning of liver and kidneys (Żarczyńska et al., 2021).
Calves are deficient in selenium at birth, and supplementation at this phase of life can contribute to the development of their immune system (NASEM, 2021) and/or prepare the defense system to act against pathogens. In the present study, supplemented calves ingested on average of 0.09 and 0.08 mg/kg more selenium and obtained an increase in serum level of 0.009 and 0.024 µg/mL and an increase in glutathione peroxidase of 17.48 and 32.65 U/g Hb, respectively for SeVitE and SeVitEFe to control. It was demonstrated that even with the increase in glutathione peroxidase in SeVitE in relation to control calves, there was no effective action against Anaplasma pathogens and in the incidence of diarrhea. However, when calves were supplemented together with iron (SeVitEFe), they showed a 1.86-fold increase in glutathione concentration in relation to SeVitE and had an effective reduction in the number of Anaplasma pathogens. Furthermore, combined supplementation of micronutrients resulted in a 10% reduction in the incidence of diarrhea in the first 30 days of life, a phase in which the disease appears most frequently.
The purpose of selenium supplementation above requirements recommended by NRC (2001) and maintained in Nasem (2021), would be to increase the concentration of the nutrient in the blood of calves, and consequently increase and maintain higher glutathione peroxidase concentrations throughout the entire suckling period. So, if the calf faces any stress, pathogen, or disease, the enzyme would be available to act immediately. The increase in the blood glutathione peroxidase concentration depends on the increase in serum selenium (Thompson et al., 1981), and this increase in amount and time depends on the dose of supplemented selenium. In the study in which high doses were offered to calves (1 mg Se/kg BW resulting in 0.200 µg Se/mL in the blood) an increase in the concentration of the enzyme was observed 10 days after starting supplementation (Żarczyńska et al., 2021). On the other hand, with doses of 0.113 mg Se/kg of dry matter, resulting in 0.030 µg Se/mL, it took almost two months to observe an increase in the concentration of the enzyme in the calves’ blood (Thompson et al., 1981).
Regarding vitamin E and iron supplementation, although their intake increased, 107.40 vs 57.26 mg/day of vitamin E for supplemented and control animals, and 179.23 vs 141.64 mg/day of Fe for SeVitEFe and non-supplemented calves; values remained unchanged in the calves’ blood (5.11 vs 5.22 µg/mL of vitamin E for supplemented and control animals, and 157.76 vs 167.40 µg/dL of Fe for SeVitEFe and non-supplemented animals). When Jersey calves were orally supplemented with high DL-alpha-tocopherol acetate doses (two grams daily) from birth to 45 days of age, their serum vitamin E concentrations increased over time, rising from 1.55 to 4.38 μg/mL (Reis et al., 2007). These authors observed that calves, between birth and the first twenty days of life, presented very low serum vitamin E concentrations, which suggests the need for supplementation of this vitamin in the newborn’s diet (Reis et al., 2007). In the current study, even offering double (108.37 and 106.44 mg/day for SeVitE and SeVitEFe) the daily vitamin E recommendation (44.7 mg/day), which are lower than the values used in the study with Jerseys calves (Reis et al., 2007), it was not sufficient to change the concentration of the nutrient in the blood of supplemented calves; however, vitamin E supplementation improves the ability to kill neutrophils and increases macrophage function in cows (Politis et al., 1995; Xiao et al, 2021). The biochemical actions of Se and vitamin E complement the body’s physiological mechanism. Vitamin E increases Se retention and prevents the auto-oxidation of lipids within cell membranes, preventing the formation of peroxides; however, only GPx can destroy the peroxides already formed (Lippman et al., 2009; Pak et al., 2002).
A study was carried out to understand the need for iron supplementation for the development of suckling calves and reported that blood iron concentrations in newborn calves before colostrum feeding were equal to those in their mothers (166 µg/dL) and decreased up to two months after birth to values of 141.52 µg/dL (Atyabi et al., 2006). In the present study, there were no differences in iron concentration among treatments, but iron values increased in serum over the experimental period (127.95, 182.02, and 182.60 µg/dL, for initial value, 40 and 60 days, respectively) due to iron supplementation since the iron concentration in the blood of calves tends to fall shortly after birth (Getty et al., 1968). Iron is a microelement and the main component of hemoglobin. Newborn mammals exhibit physiologically low hemoglobin values, and it is known that milk is deficient in iron and can lead a nursing animal to iron deficiency relatively easily (Getty et al., 1968). One of the ways in which the nutrient is stored is in the form of ferritin and hemosiderin proteins (McDowell, 2000), with ferritin being stored in the liver and spleen, and when required by the body, it is mobilized (Grotto, 2010). There are still few studies reporting the influence of chelated minerals, especially iron, on ruminant production. Unlike what occurs with monogastric, in which there is emphasis and recommendation on supplementation, there is little discussion about iron supplementation for ruminants because deficiency in adult animals is rare. The new recommendations of nutritional needs for dairy cattle indicate that iron deficiency in calves is associated with low immune response (NASEM, 2021), however, there is no greater approach to supplementation and/or increased supply of the mineral in the first phase of suckling.
It was expected that calves supplemented with iron would show an increase in erythrogram and ferritin, but this result was not observed in the current study. However, the general erythrogram parameters of calves indicate that they were not in an anemic state, since there was no decrease in plasma hemoglobin and other parameters such as mean corpuscular volume and mean corpuscular hemoglobin remained within normal values. Fe supplementation together with the other micronutrients contributed to improving oxidative metabolism, changing some biochemical parameters and the immune system, with reduced the number of Anaplasma marginale pathogens and the incidence of diarrhea compared to calves supplemented only with Se and VitE.
Among blood metabolites, animals supplemented with SeVitEFe had lower urea, lactate, and LDH values. Urea was lower in SeVitEFe animals at 60 days of experimentation. Urea is one of the main indicators of the animal’s protein metabolism, and its change in the blood is faster than albumin when there is a protein deficit in the animal’s diet (Lima et al., 2012). Blood lactate is related to rumen lactate, which is produced in the rumen when animals begin to ingest solid feed, and the rumen fermentation process becomes more active. The dry matter intake of the animals did not differ between treatments, although the SeVitEFe treatment was numerically slightly lower, but not to the point of perhaps influencing the levels of urea, LDH and lactate. This reduction may be more related to the defense metabolism of the animals against pathogens, which was more efficient in this treatment. Urea, LDH and lactate values are close to those found by other authors evaluating the metabolite in calves during the suckling phase (Feitosa et al., 2007; Lima et al., 2012).
As there was no difference among treatments for feed intake, another approach in addition to nutritional to explain these biochemical and hematological changes in the blood is the fact that control and SeVitE calves presented a higher incidence of diarrhea. Biochemical parameters were evaluated in neonatal calves with diarrhea, and it was reported higher urea and creatinine concentrations in animals with diarrhea compared to healthy ones, probably because of dehydration (Akyüz and Kükürt, 2021; Kozat et al., 2021).
One of the indices to determine the diagnosis of metabolic diseases in ruminants is the total activity of lactate dehydrogenase and its isoenzymes, and there is an increase in LDH activity when clinical symptoms of the disease are present (Klein et al., 2020). In sick calves, a slight increase in LDH and a significant increase in LDH1 activity were detected (Klein et al., 2020). In the present study, it was observed that calves in the control and SeVitE groups had higher LDH concentrations and higher incidences of diarrhea when compared to those in the SeVitEFe group.
It is already known in practice and in the literature that calves at this early stage of life are likely of having higher incidence of diseases and are also submitted to the critical stress period of weaning management. Therefore, immunonutrient supplementation would be a strategy to assist in the metabolism of calves at this early stage of life. The results show that glutathione peroxidase for supplemented calves was higher before inoculation with Anaplasma marginale (CEPA UFMG1), showing higher values of the enzyme in SeVitEFe calves. This made the oxidative metabolism of these animals to be more active to combat pathogens during immunological challenge and combat diarrhea-causing pathogens. This is a very important response for the production system, because the costs associated with treating diarrhea at this stage of life are high. In the present study, each case of diarrhea costs around US$ 22.00 (antibiotic, antipyretic, probiotic and hydration), and many calves had recurrence. This decrease in the incidence of diarrhea in SeVitEFe supplemented calves reduces costs on medicines and labor and can positively impact the adulthood productivity. The occurrence of diarrhea and other neonatal parameters such as time of birth, birth weight, and ease of birth are associated with detrimental effects on subsequent production and reproductive performance (Aghakeshmiri et al., 2017).
Supplementation of the micronutrients selenium, iron and vitamin E together showed effectiveness in improving the animals’ oxidative metabolism, altering biochemical and hematological parameters, reducing the blood count of the Anaplasma marginale and the incidence of diarrhea, but did not enhance performance. Although this study demonstrated the importance of immunonutrient supplementation for calves at this early stage of life, additional studies should be conducted for nutritional recommendations for these supplements.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by Animal Use Ethics Committee of the Animal Science Institute, with protocol number CEUA/IZ 268-18. The study was conducted in accordance with the local legislation and institutional requirements.
MS: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. FF: Data curation, Investigation, Writing – review & editing. CB: Investigation, Methodology, Writing – review & editing. VG: Conceptualization, Methodology, Writing – review & editing. RM: Methodology, Writing – review & editing. JS: Conceptualization, Investigation, Writing – review & editing. EF: Conceptualization, Methodology, Writing – review & editing. JF: Formal analysis, Writing – review & editing. AS: Conceptualization, Investigation, Methodology, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. We are grateful to the São Paulo Research Foundation (FAPESP 2017/04165-5, 2019/03516-4, and 2020/01116-6) and CNPQ 301990/2022-0 for the financial support of this study.
The authors also would like to acknowledge the support from Animal Science Institute in Ribeirão Preto and the technical help provided by the co-workers.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Żarczyńska K., Sobiech P., Tobolski D., Mee J. F., Illek (2021). Effect of a single, oral administration of selenitetriglycerides, at two dose rates, on blood selenium status and haematological and biochemical parameters in Holstein-Friesian calves. J. Ir. Vet. J. 74, 11. doi: 10.1186/s13620-021-00192-4
Abdelrahman M. M., Kincaid L. (1995). Effect of selenium supplementation of cows on maternal transfer of selenium to fetal and newborn calves. J. Dairy Sci. 78, 625–630. doi: 10.3168/jds.S0022-0302(95)76673-5
Aghakeshmiri F., Azizzadeh M., Farzaneh N., Gorjidooz M. (2017). Effects of neonatal diarrhea and other conditions on subsequent productive and reproductive performance of heifer calves. Vet. Res. Commun. 41, 107–112. doi: 10.1007/s11259-017-9678-9
Akyüz E., Kükürt A. (2021). Evaluation of oxidative stress index and some biochemical parameters in neonatal calves with diarrhea. Acta Sci. Vet. Sci. 3, 58–63. doi: 10.31080/ASVS.2021.03.0188
A.O.A.C. Association of official analytical chemists (1996). Official Methods of Analysis. Sixteenth edition (Washington, D.C: Association of Official Analytical Chemists, Inc.).
Arnaud J., Fortis I., Blachier S., Kia D., Favier A. (1991). Simultaneous determination of retinol, α-tocopherol and β-carotene in serum by isocratic high-performance liquid chromatography. J. Chromatogr. B Biomed. Appl. 572, 103–116. doi: 10.1016/0378-4347(91)80476-s
Atyabi N., Gharagozloo F., Nassiri S. M. (2006). The necessity of iron supplementation for normal development of commercially reared suckling calves. Comp. Clin. Pathol. 15, 165–168. doi: 10.1007/s00580-006-0624-4
Azevedo R. A., Teixeira A. M., da Silva A. L., Arantes A. N., Sabino Resende A. L. S., et al. (2024) (Uberaba, Minas Gerais), 170. doi: 10.26626/9786556682082.2024B0001
Bastos C. V., Passos L. M., Facury-Filho E. J., Rabelo E. M., de la Fuente J., Ribeiro M. F. (2010). Protection in the absence of exclusion between two Brazilian isolates of Anaplasma marginale in experimentally infected calves. Vet. J. 186, 374–378. doi: 10.1016/j.tvjl.2009.09.013
Brigelius-Flohé R., Flohé L. (2020). Antioxidants & Redox Signaling Vol. 33 (Larchmont, United States: Publishing Mary Ann Liebert). doi: 10.1089/ars.2019.79054982020
de la Fuente J., Estrada-Pena A., Venzal J. M., Kocan K. M., Sonenshine D. E. (2008). Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 13, 6938–6946. doi: 10.2741/3200
Ensley E. (2020). Evaluating mineral status in ruminant livestock. Vet Clinics North America: Food Anim. Pract. 36, 525–546. doi: 10.1016/j.cvfa.2020.08.009
Feitosa F. L. F., Mendes C. N., Peiró J. R., Cadioli F. A., Yanaka R., Bovino F., et al. (2007). Influence of age on values of hepatic enzymatic activity and of urea and creatinine in Holstein calves at birth until 365 days of life. Ciênc. Vet. Tróp. 10, 54–61.
Ferreira G. C. M., Canozzi M. E. A., Peripolli V., Moura G. P., Sánchez J., Martins C. E. N. (2022). Prevalence of bovine Babesia spp., Anaplasma marginale, and their co-infections in Latin America: Systematic review-meta-analysis. Ticks Tick-borne Dis. 13, 1-8. doi: 10.1016/j.ttbdis.2022.10196
Getty S. M., Beck C. C., Brown L. D., Connor G. H., Ellis D. J., Miller E. R. (1968). Effect of iron on hematology and growth of calves. J. Anim. Sci. 27, 712–717. doi: 10.2527/jas1968.273712x
Goering H. K., Van Soest P. J. (1970). “Forage Fiber Analysis (Apparatus Reagents, Procedures and Some Applications),” in Agriculture Handbook (United States Department of Agriculture, Washington DC).
Goff J. P. (2018). Invited review: Mineral absorption mechanisms, mineral interactions that affect acid–base and antioxidant status, and diet considerations to improve mineral status. J. Dairy Sci. 101, 2763–2813. doi: 10.3168/jds.2017-13112
Grotto H. Z. W. (2010). Fisiologia e metabolismo do ferro. Rev. Bras. Hematol. Hemoter. 32, 8–17. doi: 10.1590/S1516-84842010005000050
Gunter S. A., Beck P. A., Phillips J. M. (2003). Effects of supplementary selenium source on the performance and blood measurements in beef cows and their calves. J. Anim. Sci. 81, 856–864. doi: 10.2527/2003.814856x
Horwitz W., Larimer G. W. (2005). Official Methods of Analysis of the Association of Official Analytical Chemists. Eighteenth edition Vol. 1 (Gaithersburg, Maryland 20877-2417, USA: Association of Official Analytical Chemistry International), 45.
IDEAGRI. (2019). Brazilian Milk Index. Available online at: http://web.ideagri.com.br/ideagriweb/iilb/process/IilbBiblioteca?_ga=2.113017159.2146775234.1670158642-1927443523.1670158642 (Accessed September, 27, 2023).
IDEAGRI. (2021). Brazilian Milk Index. Available online at: https://www.ideagri.com.br/siteideagridados/New/IILB/2021_09/boletim_IILB.pdf (Accessed September, 27, 2023).
IICA – Instituto Interamericano de Cooperacíon para la Agricultura (1987). Técnicas para el diagnostico de babesiosis y anaplasmosis bovinas (San José: Instituto Interamericano de Cooperación para la Agricultura), 79.
Jaaf S., Batty B., Krueger A., Estill C. T., Bionaz M. (2020). Selenium biofortified alfalfa hay fed in low quantities improves selenium status and glutathione peroxidase activity in transition dairy cows and their calves. J. Dairy Res. 87, 184–190. doi: 10.1017/S002202992000028X
Kamada H., Nonaka I., Ueda Y., Murai M. (2007). Selenium addition to colostrum increases immunoglobulin G absorption by newborn calves. J. Dairy Sci. 90, 5665–5670. doi: 10.3168/jds.2007-0348
Kenward M. G., Roger J. H. (1997). Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 53, 983–997.
Klein R., Nagy O., Tóthová C., Chovanová F. (2020). Clinical and diagnostic significance of lactate dehydrogenase and its isoenzymes in animals. Vet. Med. Int. 15, 5346483. doi: 10.1155/2020/5346483
Kozat S., Özkan C., Okman E. N. (2021). Evaluation of homocysteine levels in neonatal calves with diarrhea. Kafkas Univ. Vet. Fak. Derg. 27, 271–277. doi: 10.9775/kvfd.2020.24894
LeBlanc S. J., Duffield T. F., Leslie K. E., Bateman K. G., TenHag J., Walton J. S., et al. (2002). The effect of prepartum injection of vitamin E on health in transition dairy cows. J. Dairy Sci. 85, 1416–1426. doi: 10.3168/jds.S0022-0302(02)74209-4
Lima P. O., Cândido M. J. D., Queiroz M. G. R., Ferreira J. M., Modesto E. C., Lima R. N., et al. (2012). Serum parameters of calves receiving different types of liquid diets. Rev. Bras. Saúde Prod. Anim. 13, 529–540. doi: 10.1590/S1519-99402012000200020
Lippman S. M., Klein E. A., Goodman P. J., Lucia M.S., Thompson I.M., Ford L.G., et al. (2009). Effect of selenium and vitamin E on risk of prostate cancer and other cancers: the selenium and vitamin E cancer prevention trial. JAMA 301, 39–51. doi: 10.1001/jama.2008.864
McDowell L. R. (2000). Vitamins in animal and human nutrition. 2st edition (Iowa, nos Estados Unidos: Iowa State University Press).
NASEM, National Academies of Sciences, Engineering, and Medicine (2021). Nutrient Requirements of Dairy Cattle. Eighth Revised Edition Vol. 143 (Washington, DC: THE NATIONAL ACADEMIES PRESS).
NRC - NATIONAL RESEARCH COUNCIL (2001). Nutrient requirements of dairy cattle. sixth edition (Washington: Academy Press).
Olson O. E., Palmer I. S., Cary E. E. (1975). Modification of the official fluorimetric method for selenium in plants. J. Assoc. Off. Agric. Chem. 58, 117–121.
Pak R. W., Lanteri V. J., Scheuch J. R., Sawczuk I. S. (2002). Review of vitamin E and selenium in the prevention of prostate cancer: implications of the selenium and vitamin E chemoprevention trial. Integr. Cancer Ther. 1, 338–344. doi: 10.1177/1534735402238186.37
Politis I., Hidiroglou M., Batra T. R., Gilmore J. A., Gorewit R. C., Scherf H. (1995). Effects of vitamin E on immune function of dairy cows. Am. J. Vet. Res. 56, 179–184. doi: 10.2460/ajvr.1995.56.02.179
Ranches J., Vendramini J. M. B., Arthington J. D. (2017). Effects of selenium biofortification of hayfields on measures of selenium status in cows and calves consuming these forages. J. Anim. Sci. 95, 120–128. doi: 10.2527/jas.2016.0943
Reber A. J., Donovan D. C., Gabbard J., Galland K., Aceves-Avila M., Holbert K. A., et al. (2008). Transfer of maternal colostral leukocytes promotes development of the neonatal immune system Part II. Effects neonatal lympho. Vet Immunol. Immunopathol. 123, 305–313. doi: 10.1016/j.vetimm.2008.02.009
Reis M. C., Costa J. N., Peixoto A. P. C. (2007). Effect of age on oral supplementation with DL-α-tocopherol acetate on serum vitamin E levels and acetate proteinogram. Rev. Bras. Saúde Prod. An. 8, 151.
Salles M. S. V., Zanetti M. A., Roma Junior L. C., Salles F. A., Azzolini A. E. C. S., Soares E. M., et al. (2014). Performance and immune response of suckling calves fed organic selenium. Anim. Feed Sci. Technol. 188, 28–35. doi: 10.1016/j.anifeedsci.2013.11.008
SAS Statistical Analysis Software (2013). Users’ Guide Statistics Version 9.4 (Cary: SAS Institute Inc.).
Silveira J. A. G. (2012). Ocorrência de hemoparasitos e ectoparasitos em veado-catingueiro (Mazama gouazoubira Fischer 1814), veado-campeiro (Ozotocerus bezoarticus Linnaeus 1758) e cervo-do-pantanal (Blastocerus dichotomus Illiger 1815): utilização de métodos parasitológicos e moleculares (Belo Horizonte, Minas Gerais, Brazil: Universidade Federal de Minas Gerais).
Silveira J. A., Silvestre B. T., Bastos C. V., Ribeiro M. F. (2016). Isolation and attempted cultivation of an Anaplasma marginale strain from Brazilian brown brocket deer (Mazama gouazoubira, Fisher 1814) in the tick cell line IDE8. Ticks Tick Borne Dis. 7, 1102–1108. doi: 10.1016/j.ttbdis.2016.09.001
SMEWW. (2017). Standard Method for the Examination of Water and Wastewater. 23 Edition (Washington, D.C: Samples, American Public Health Association).
Thompson K. G., Fraser A. J., Harrop B. M., Kirk J. A., Bullians J., Cordes D. O. (1981). Glutathione peroxidase activity and selenium concentration in bovine blood and liver as indicators of dietary selenium intake. N. Zeal. Vet. J. 29, 3-6. doi: 10.1080/00480169.1981.34776
USDA - U.S. Department of Agriculture. (2015). Death Loss in U.S. Cattle and Calves Due to Predator and Nonpredator Causes. Available online at: https://content.govdelivery.com/accounts/USDAAPHIS/bulletins/1cd6c28 (Accessed September, 27, 2023).
Uyama T., Kelton D., Winder C., Dunn J., Goetz H., LeBlanc S., et al. (2022). Colostrum management practices that improve the transfer of passive immunity in neonatal dairy calves: A scoping review. PloS One 17, 1-17. doi: 10.1371/journal.pone.0269824
Van Soest P. J., Robertson J. B., Lewis B. A. (1991). Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J.Dairy Sci. 74, 3583–3597. doi: 10.3168/jds.S0022-0302(91)78551-2
Wangsanut T., Pongpom M. (2022). The role of the glutathione system in stress adaptation, morphogenesis and virulence of pathogenic fungi. Int. J. Mol. Sci. 23, 10645. doi: 10.3390/ijms231810645
Keywords: antioxidants, performance, immunology, immunonutrients, nutrition, ruminant
Citation: Salles MSV, Figueiroa FJF, Bittar CMM, Gomes V, Marques RS, da Silveira JAG, Facury Filho EJ, de Freitas JE and Saran Netto A (2025) Supplementation with selenium, iron, and vitamin E in calves under immunological challenge. Front. Anim. Sci. 6:1540495. doi: 10.3389/fanim.2025.1540495
Received: 05 December 2024; Accepted: 31 January 2025;
Published: 18 February 2025.
Edited by:
Bianca Castiglioni, National Research Council (CNR), ItalyReviewed by:
Ravikanth Reddy Poonooru, University of Missouri, United StatesCopyright © 2025 Salles, Figueiroa, Bittar, Gomes, Marques, da Silveira, Facury Filho, de Freitas and Saran Netto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Márcia S. V. Salles, bWFyY2lhLnNhbGxlc0BzcC5nb3YuYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.