 Claudiney de Jesus Couto1
Claudiney de Jesus Couto1 Bruna Cardoso Braga2*Raphael dos Santos Gomes3Wagner Pessanha Tamy4Severino Delmar Junqueira Villela1
Bruna Cardoso Braga2*Raphael dos Santos Gomes3Wagner Pessanha Tamy4Severino Delmar Junqueira Villela1 Fernando de Paula Leonel1,5*
Fernando de Paula Leonel1,5*- 1Department of Animal Science, Federal University of Jequitinhonha and Mucuri Valleys, Diamantina, MG, Brazil
- 2Department of Animal Science, Federal University of Goiás, Goiânia, GO, Brazil
- 3Federal Institute of Education, Science and Technology of Rondonia, campus Colorado do Oeste, Colorado do Oeste, RO, Brazil
- 4Department of Animal Science and Sustainable Agro-Socio-Environmental Development, Faculty of Veterinary Medicine, Fluminense Federal University, Niterói, RJ, Brazil
- 5Department of Animal Science, Federal University of São João del-Rei, São João del-Rei, MG, Brazil
Introduction: Since the low phosphorus content is one of the reasons for the low degradation of fiber in roughage such as pasture and by-products, fiber transit kinetics can be an important tool to evaluate differences in digestion depending on phosphorus supplementation with different sources. The objective was to compare the potential of different sources of phosphorus in promoting improvement in fiber digestibility parameters.
Methods: Feeds low in phosphorus were chosen to compose the basal diet. Five crossbred heifers with an average initial weight of 150 ± 8.2 kg were used, randomly distributed in a 5x5 Latin square. The treatments were: negative control (NC) without supplementary phosphorus; dicalcium phosphate from igneous rock with sulfuric acidulation, neutralization with limestone, and the show physical form of microgranules (ISLG); Dicalcium phosphate from igneous rock with sulfuric acidulation, neutralization with hydrated lime, and the physical form of powder (ISLhP); Dicalcium phosphate from sedimentary rock with hydrochloric acidulation, neutralization with limestone, and the micro-granulated show physical form (SeCLG); Dicalcium phosphate from the monocalcium phosphate and hydrated lime reaction adjusted with phosphoric acid, and the powder physical form (MLhPaP). The parameters of the kinetics transit of particles with use of the chromium marker, fixed to the roughage cell wall and subsequent recovery of the marker in the heifer feces, were determined.
Results and discussion: The ISLG resulted in a higher (p<0.05) passage rate of particles from the solid to the liquid phase in the rumen-reticulum. There was no difference (p>0.05) between the diets for the time that indigestible particles spent from ruminal escape to their first appearance in feces. The longest retention time (p<0.05) of solid particles in the rumen-reticulum was detected in animals that received NC diet. In conclusion, heifers that do not receive supplemental phosphorus exhibited poorer fiber processing and dry matter intake (DMI).
1 Introduction
Phosphorus (P) is the chemical element whose biological functions are well established. Among the numerous functions attributed to phosphorus, the main ones are the formation of bone structure, participation in the formation of cell membranes, and the use and transfer of energy in the form of ATP (Lehninger, 1994) making it a critical component in almost all metabolic pathways (Suttle, 2010; McDowell, 2003). Reduced food intake is the earliest effect of phosphorus deficiency. Deficiencies of this element have also been associated with reduced growth, anorexia, decreased milk production, and reproductive disorders (Preston et al., 1977; Butterworth, 1985; Underwood, 1996). According to Durand and Kawashima (1980), low dietary P levels can reduce rumen microbial growth, resulting in lower microbial protein production, lower diet digestibility and lower energy supply. The reduction in dry matter intake is also a consequence of the decreased degradability of fiber by rumen microorganisms and the lower metabolic activity of cells, which negatively influence the satiety center (Minson, 1990).
The efficiency of the diet provided to ruminant animals influences both economic viability and environmental sustainability. This is due to its presence in excessive and eutrophic quantities in water bodies, due to the disposal of untreated or unsatisfactorily treated sanitary effluents, causing environmental damage (Sasabuchi et al., 2023). In addition to balanced formulation and animal-diet interactions, diet efficiency also depends on the quality of the ingredients used. In other words, the choice of a particular ingredient or nutrient source depends, among other factors, on the availability of the nutrient for animal metabolism. This is a principle that applies to all sources of nutrients in the diet. However, decision-making is not a simple process. Therefore, in addition to financial cost, the availability of the nutrient in the source options must be considered.
P is essential to metabolism, with mandatory inclusion in diets. This mineral can be obtained from different sources (Satter et al., 2005). Therefore, the choice of source must be based on efficiency criteria, which in this context, may be the one that has the phosphorus best used by the microorganisms in the rumen. Assessing the comparative efficiency of phosphorus sources is not a simple task, as it depends on the characteristics of the source itself (such as origin and processing), its interaction with other nutrients in the diet, the animal category, and productivity. Furthermore, phosphorus can be stored in the body’s reserves and mobilized when necessary. The storage and mobilization process is a complicating factor in comparison tests between sources. However, there are some ways to carry out scientific experiments with the necessary accuracy to compare different phosphorus sources; for example, assays that evaluate ruminal events highly influenced by phosphorus availability.
It is extremely important that the phosphorus meets the requirements of the animal and rumen microorganisms, as its deficiency in the diet can impair digestibility and transit kinetics, which reduces dry matter consumption and, consequently, the overall supply of nutrients (Van Soest, 1994). Thus, an understanding of the availability of phosphorus directly (radioactive tracers) or indirectly (physical, chemical, or biological digestion parameters) is essential for the development of strategies that can increase its efficiency of use. Since the low phosphorus content is one of the reasons for the low degradation of fiber in roughage such as pasture and by-products, fiber transit kinetics can be an important tool to evaluate differences in digestion depending on phosphorus supplementation with different sources.
Therefore, our objective was to compare the potential of different phosphate sources to promote improvements in fiber digestibility. For this we used the technique of evaluation of fiber transit kinetic parameters.
2 Materials and methods
Use of experimental animal statement: i) All animal experimental procedures were approved by the Committee on Animal Use and Care at the Minas Gerais Agricultural Research Company (CEUA EPAMIG01/2019). ii) All experiment procedures were also performed according to relevant guidelines and regulations (ARRIVE guidelines).
Five ruminally fistulated crossbred heifers (Holstein x Gyr) at six months old were used in a 5 × 5 Latin square and randomly distributed into 5 treatments and periods (each 21-d period, being 7-d for adaptation to treatment and 14 for sampling). The animals had a body weight (BW) of 150 ± 8.2 kg (mean ± SD) at the beginning of the experiment and 214 ± 31.9 kg at the end. The heifers were in individual pens, consuming a balanced standard diet to meet the requirements to gain 0.5 kg/day (ingredients: mature grass hay, potato starch, soybean oil, urea, and mineral supplement – Table 1).
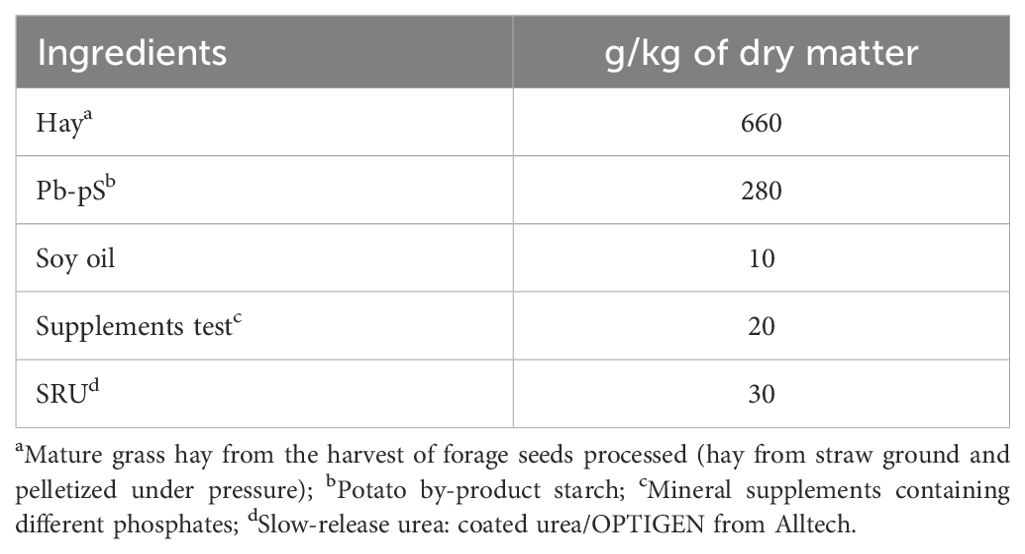
Table 1. Diet formulation.
These diet ingredients were chosen because they are low in phosphorus, this allowed for a greater challenge to the phosphorus sources under study. The mature grass hay offered to heifers was ground and pressed to form briquettes.
The phosphorus sources under study are characterized by differences in the raw rocks, in the acidification processes of these rocks, in the phosphoric acid neutralization bases and in the form of presentation of the product (Powder or Micro-granulated), in addition to the negative control.
Thus, the treatments under study were: 1. negative control (NC) without supplementary phosphorus; 2. dicalcium phosphate from the igneous rock with sulfuric acidulation, neutralization with limestone, and the physical form of microgranules (ISLG); 3. Dicalcium phosphate from the igneous rock with sulfuric acidulation, neutralization with hydrated lime, and powder physical form (ISLhP); 4. Dicalcium phosphate from the sedimentary rock with hydrochloric acidulation, neutralization with limestone, and the micro-granulated physical form (SeCLG); 5. Dicalcium phosphate from the monocalcium phosphate and hydrated lime reaction adjusted with phosphoric acid, and powder physical form (MLhPaP). The phosphorus sources were provided to heifers in premixes (in the same amount: 100 g of P/kg) and included in the total mixed ration (TMR), which had a final concentration of 2 g of P/kg of DM. There was one premix without phosphorus (negative control).
The voluntary dry matter intake was obtained by the difference between the weight of offered feed and the weight of leftovers.
Total phosphorus was determined by Method 965.17 of AOAC (2012), total DM (method 976.03; AOAC, 1990), ash (method 942.05; AOAC, 1990), total nitrogen (method 981.10; AOAC, 2012), fat (method 996.06; AOAC, 2010) and lignin (method 973.18; AOAC, 1997). The CP content was calculated by multiplying the percentage of N by 6.25. Neutral detergent fiber (aNDF) was determined according to Mertens (2002), without the addition of sodium sulfide and with the addition of thermostable alpha-amylase. The aNDF content was corrected for protein and ash for all samples (Licitra et al., 1996) (Table 2).
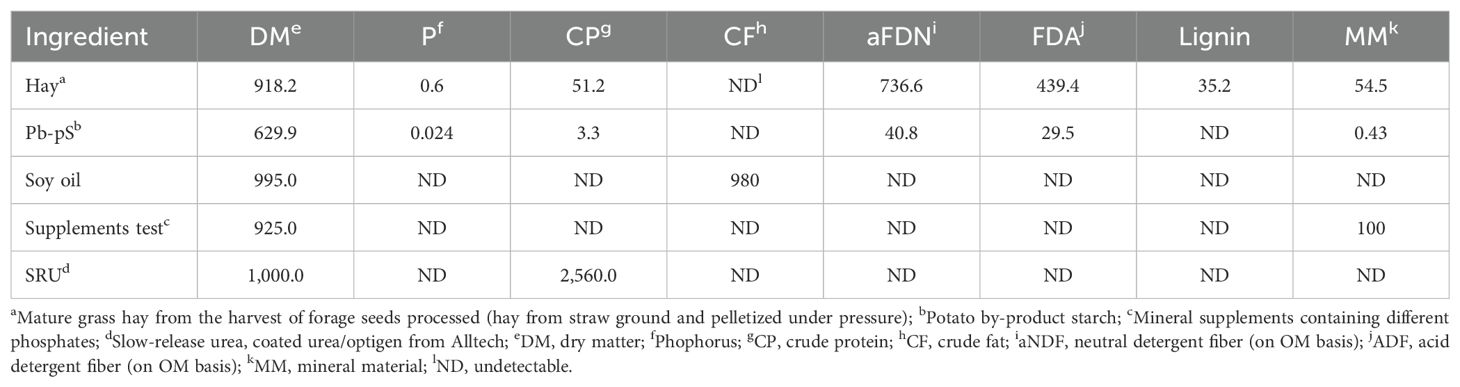
Table 2. Proximate composition (g/kg of DM) of the ingredients.
2.1 Determination of particle transit kinetics parameters
The particle transit kinetic parameters were estimated by employing the chromium-mordant technique on the fiber from the mature grass hay, according to Udén et al. (1980). The samples were dried in a forced-air oven at 55 ± 5 °C for 72 h. Subsequently, this material was boiled with water and neutral detergent for 1 h. The proportion of ingredients used was 100 g of dry sample to 100 mL of detergent and 1 L of water. Then, the material was filtered through a cotton fabric bag and washed in running water until the water was clear to remove the soluble components. After the filtering process, the fiber was returned to the forced-air oven at 55 ± 5 °C, where it remained for 72 h. At the end of the washing and drying process, the fiber was placed in a suitable container and immersed in a solution of potassium dichromate (K2Cr2O7, 2H2O) at the ratio of 13% of chromium relative to the fiber weight. The container with the fiber was completely covered with aluminum foil and oven-dried at 105°C for 24 h. Afterward, a second wash was performed to remove excess potassium dichromate. After this second wash, the material was immersed in a commercial ascorbic acid solution at the proportion of half the fiber weight. The immersion remained at rest for 1 h until it reached an intense green color. Then, a third wash was performed until the water was completely clear. After washing, the fiber was dried in a forced-air oven at 55 ± 5 °C for 72 h.
The labeled fiber (chromium mordant fiber) was inserted directly in the ruminal fistula, at 200 grams per animal. Feces were collected, from rectum, at times zero (immediately after administering the chromium mordant fiber), 1, 2, 4, 6, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 56, 64, 72, 80, 88, 96, 120, 132, 144, and 196 hours, to estimate the parameters of particle passage kinetics (Vieira et al., 1997). After the collections at the indicated times, samples were dried in a forced-ventilation oven at 55 ± 5°C for 72 hours, processed in a mill with 1 mm mesh sieve and stored for later analysis of the chromium content, according to the method proposed by Williams et al. (1962).
Approximately 200 mg of feces respective to each incubation time, animal, and period were diluted in a nitric-perchloric acid solution to remove the organic matter and for subsequent reading of the chromium concentration by atomic absorption spectrometry, following the methodology described by Kimura and Miller (1957).
2.2 Models used to predict particle transit kinetics parameters
Chromium (marker) concentration profiles in feces were described using the generalized bicompartmental model suggested by Matis et al. (1989):
;
to :
The parameters of the model above provide estimates that explain the dynamics of the rate of passage or transit of fibrous particles through the gastrointestinal tract of ruminants in general, where is the concentration of a marker in the feces at a given time t; is the concentration of the marker in the first compartment at t = 0; escape of eligible particles from the rumen-reticulum to the rest of the gastrointestinal tract; ; transfer of particles from the raft pool to the pool of escaping particles dispersed in the liquid phase of the rumen; order of time dependence; denotes the order of time dependency, ∀ 1, 2, …, N and transit time (time equivalent to the marker leaving the reticule-omasal orifice until the first appearance in the feces).
The turnover or mean retention time in the rumen-reticulum (MRT) was estimated based on biological interpretations, in which both ascending and descending phases of the excretion profile of markers in feces influence the retention of particles in the rumen-reticulum (Vieira et al., 2008b). The MRT is expressed in hours and was estimated according to the equation by Matis et al. (1989):
The adjusted model parameters for passage kinetics were estimated with the NLIN procedure of SAS (SAS, 2000) (SAS System Inc., Cary, NC, USA, version 9.0). Both Newton and Marquardt algorithms were used. Initially, the preferred form or algorithm was Newton’s, due to its good performance in terms of convergence, however, whenever the correlations between parameter estimates were high, we opted for Marquardt’s algorithm.
The selection of the best version for the order of time dependence () and consequently the best model to explain the passage rate, was evaluated by computing the Akaike information criterion (Akaike, 1974; Burnham and Anderson, 2004). The was calculated by the sum of the error squares , number of estimated parameters including residual variance (), and sample size () for all different versions of , . The differences between the values of (), the probability of likelihood (), and the evidence ratio () were obtained according to Vieira et al. (2012)
The parameter estimates of the passage rate profiles were adjusted according to robust regression procedures (Beaton and Tukey, 1974), to reduce the effect of discrepant observations (outliers) and eliminate subjectivity during the assessment of these points.
To verify the effects of treatments on the analyzed variables, and estimate the parameters of the regression curves, when significant, the PROC MIXED procedure was adopted in the SAS statistical program (version 9.0). The statistical model adopted was as follows:
Where is the observed response, is the overall mean, is the row (heifer) effect, is the column (periods) effect, is the treatment effect and is the experimental error.
3 Results
The highest (p<0.05) transfer rate of particles from the mat or raft (digesta that floats in the dorsal part of the rumen) to the pool of escaping particles dispersed in the liquid phase of the rumen (λ) was detected in animals that received the diet containing dicalcium phosphate from igneous rock with sulfuric acidulation, neutralization with limestone, and presented in microgranule form (ISLG) (Table 3). Fiber from the ISLG diet also showed greater (p<0.05) escape of eligible particles from the rumen-reticulum to the rest of the gastrointestinal tract (k) than the NC diet (without supplemental phosphorus); however, it did not differ (p>0.05) of the other diets, but the k of NC did not differ (p<0.05) from those of SeCLG, ISLhP, and MLhPaP (Table 3).
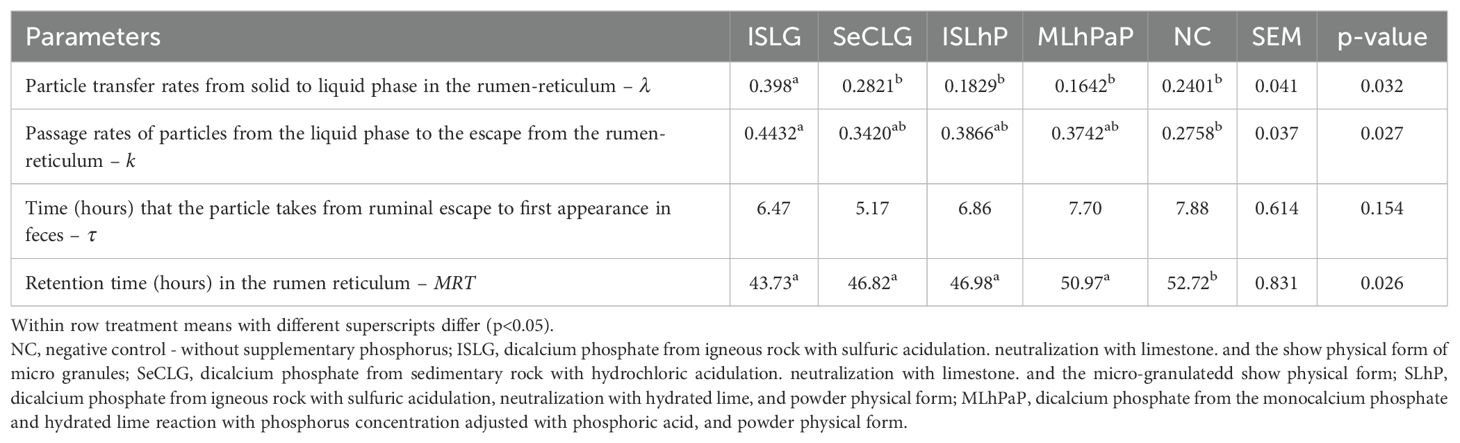
Table 3. Fiber transit kinetic parameters in heifers that received diets with different sources of phosphate.
The transit time of indigestible particles leaving the rumen-reticulum until their appearance in the feces (τ) did not differ (p>0.05) according to the phosphorus sources (Table 3). This parameter has little effect on dry matter intake since the events take place in the digestive compartments after the rumen-reticulum. The longest retention time (p<0.05) of solid particles in the rumen reticulum was detected in animals that received the diet without supplemental phosphorus (Table 3).
The heifers that received the supplemental phosphorus sources ISLG and SeCLG exhibited the highest (p<0.05) dry matter intakes (kg/heifer/day, and kg/100 kg of body weight). There was no difference (p>0.05) for these variables between heifers that did not receive supplemental phosphorus and those that received ISLhP and MLhPaP (Table 4).
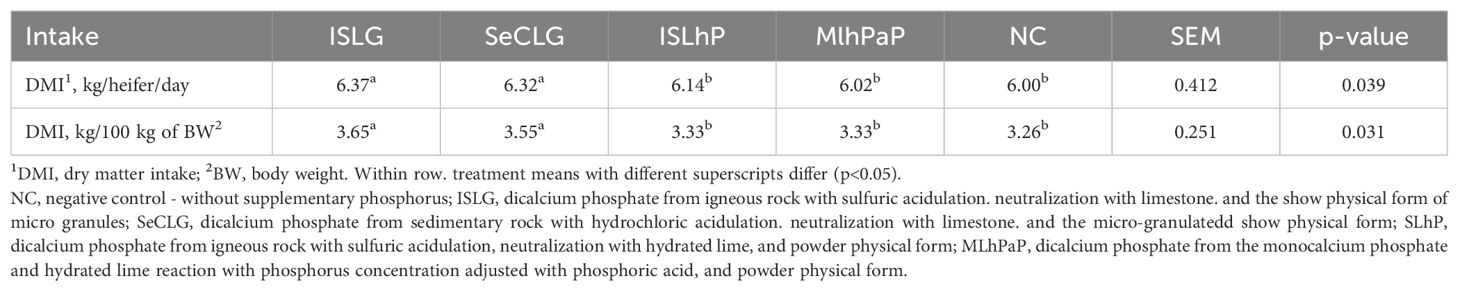
Table 4. Dry matter intake by heifers fed diets containing phosphate from different sources.
4 Discussion
It is important to provide a general summary of the phosphates manufacturing process: 1. An acid attack is carried out on the phosphate rock (apatite), with the product being the phosphoric acid and calcium sulfate; 2. Neutralize this phosphoric acid with a basic source; 3. The final solid product is processed in the following ways: a) grinding into powder form or b) going through a granulation process so that the phosphate is presented in micro-granulated form. Therefore, some phosphates differ in terms of the origin of the rock (igneous or sedimentary), the form of acidification (sulfuric or hydrochloric), and the neutralizing base.
The higher transfer rate of particles from the mat to the pool of escaping particles dispersed in the liquid phase of the rumen (λ), in diet with ISLG, allows us to infer that there was better processing of the fibrous mass by the animal (rumination) or by ruminal microflora or both effects. This indicates that the indigestible fraction has the possibility of leaving the rumen more quickly, which is desirable when the objective is to increase the animal’s dry matter intake and consequently, increase the overall supply of nutrients (Vieira et al., 2020). Greater escape of particles from the rumen compartment may be necessary when animals are fed a low-digestibility diet (Dufreneix et al., 2019).
The greater availability of phosphorus from ISLG for the ruminal microbiota, in relation to diets containing other phosphates sources, may have been the factor that provided the improvement in λ values. Phosphorus is an essential mineral for microbial growth, required by ruminal microorganisms to digest cellulose (Yoshihara et al., 2013; Suttle, 2010; Wu et al., 2000). Therefore, more available phosphorus in the diet improves microbial growth, enhancing cellulose degradation and, consequently, increasing the particle transfer rate (Suttle, 2010; Van Soest, 1994). In this study, forage was the basal diet of the heifers; thus, the predominant ruminal microbial population was cellulolytic bacteria (Butyrivibrio fibrisolvens, Fibrobacter succinogenes, Ruminococcus albus, Ruminococcus flavefaciens), which have a high phosphorus requirement. Preston and Pfander (1964) reported that these microorganisms have higher phosphorus requirements than the host animal itself. Durand and Komisarczuk (1988) state that ruminal available phosphorus should be at least 5 g/kg OMD (organic matter digestible) to optimize fiber degradation.
It is noteworthy that regardless of the treatment, the fiber of mature grass hay showed high transfer rates of particles from the solid to the liquid phase, in the rumen-reticulum (λ), when comparing this parameter with that of other roughage feeds. The high transfer rate (λ) observed in mature grass hay is due to its smaller particle size in relation to silages. This smaller particle size is consequence of the grinding to enable the briquetting process that was done to prepare the grass hay into pressed briquettes. Roughages with smaller particles tend to be transferred from the solid to the liquid phase in the rumen-reticulum (λ) more quickly. This occurs because the particles have their density increased through the comminution process, resulting from rumination and the action of ruminal microorganisms (Vieira et al., 2020; Rohem Júnior et al., 2020). Smaller particles require less rumination and are more quickly acted upon by rumen microorganisms, due to the greater surface exposure for microbial attack (Regadas Filho et al., 2014; Vieira et al., 2008b). This microbial action increases the digestion rate of potentially digestible cell walls (Processi et al., 2016; Allen, 2014; Durand and Kawashima, 1980). Rumination time is considerably influenced by the physical characteristics of the diet, such as particle size (Van Soest, 1994).
In low-digestibility roughages, the transit time that is computed from the compartment containing the liquid phase until escape through the reticle-omasal orifice is important, as the faster the indigestible particles leave the rumen-reticulum (RR) the lower the negative effect of rumen repletion on the dry matter intake (Allen, 2014; Vieira et al., 2008a). Thus, higher particle transfer rates from the liquid phase to the RR exhaust (k) can result in greater dry matter consumption, consequently greater nutrient input, and greater animal performance (Dixon et al., 2020a, 2018; Dixon and Coates, 2011; Vieira et al., 2008a, b).
The lack of significant difference between the diets for () passage rate is possibly due to the fact that phosphorus has effects on the microorganisms that degrade fiber, that is, it has an effect on events that happen within the rumen compartments. In the rest of the gastrointestinal tract, there is no effective action of microorganisms that have effect on fiber kinetic parameters.
The longer mean particle retention time in the rumen-reticulum (MRT) of animals that did not receive supplemental phosphorus in the diet was probably due to the low amount of phosphorus in the rumen environment. This impaired microbial growth and consequently cellulose degradation (Yoshihara et al., 2013; Suttle, 2010; Wu et al., 2000; Hungate, 1966). Possible reductions in the fibrolytic bacterial population due to the drop in ruminal pH caused by the treatments are unlikely since several studies did not find changes in the pH of the medium as a function of phosphorus source (Coneglian et al., 2010; Barreto et al., 2009; Komisarczuk et al., 1987; Witt and Owens, 1983). There are reports that in conditions of low phosphorus concentration (< 4 mg/L), there is a drop in VFA production, with a consequent increase in pH. However, this is not an effect of the phosphorus source but rather a consequence of the low fermentation of fiber resulting from the poor growth of cellulolytic bacteria (Komisarczuk et al., 1987). In other words, the best parameters of fiber transit kinetic in the diets with ISLG and SeCLG can be attributed to the greater phosphorus availability in these sources.
When there is low phosphorus availability in the diet, ruminants normally mobilize the phosphorus stored in the skeleton and send it to the rumen via saliva or the ruminal wall (Schatz et al., 2023; Dixon et al., 2020a; Underwood and Suttle, 2001). This recycling of phosphorus appears not to have been sufficient in this study, probably because animals in the growth phase were used and this meant that phosphorus was prioritized for bone growth and not for recycling.
The MRT is also related to dry matter intake capacity (Processi et al., 2016). The lower the MRT, the greater the possibility of the animal increasing its dry matter intake per unit of time without the rumen-fill regulation system acting, enabling a greater supply of nutrients to the animal (Araújo et al., 2014).
The higher dry matter intake by heifers receiving supplementation with ISLG and SeCLG supports the results of the ruminal kinetic parameters of fiber (λ and k) in the diets that have included these phosphorus sources. Also, as with the transit kinetics, the high dry matter intake, regardless of the treatment, can be justified by the small particle size of the hay. In order to make the briquetting process viable, the hay was finely ground.
5 Conclusion
Phosphorus supplementation improved fiber transit kinetics, regardless of the phosphate source. ISLG and SeCLG demonstrated the most significant improvements in fiber transit kinetic parameters and dry matter intake among the sources tested. Phosphorus sources that promote better parameters of ruminal fiber transit kinetics could improve animal growth, milk production, and reproductive performance.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal studies were approved by Committee on Animal Use and Care at the Minas Gerais Agricultural Research Company (CEUA EPAMIG01/2019). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
CC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. BB: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. RG: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. WT: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. SV: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. FL: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
The authors thank the Coordination for the Improvement of Higher Education Personnel (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES), for support with scholarship to graduate student.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akaike H. (1974). A new look at statistical model identification. IEEE Trans. Automatic Control 19, 716–723. doi: 10.1109/TAC.1974.1100705
Allen M. S. (2014). Drives and limits to feed intake in ruminants. Anim. Production Sci. 54, 1513–1524. doi: 10.1071/AN14478
Araújo K. G., Villela S. D. J., Leonel F. P., Tamy W. P., Gomes R. S., Abreu M. L. C. (2014). Rumen degradation and passage kinetics of silage made from different corn hybrids. Rev. Bras. Zootecnia 43, 519–523. doi: 10.1590/S1516-35982014001000002
Association of Official Analytical Chemists. (1990). Official Methods of Analysis of the Association of Official Analytical Chemists. 15th ed (Washington, DC: AOAC International). Available at: https://archive.org/details/officialmethodso02horw.
Association of Official Analytical Chemists (1997). Official Methods of Analysis of AOAC International. 16th ed (Arlington, VA: AOAC International).
Association of Official Analytical Chemists. (2010). Official Methods of Analysis of Association of Official Analytical Chemists. 18th ed (Washington, DC: AOAC International).
Association of Official Analytical Chemists (2012). Official methods of analysis of AOAC International. 19th ed (Gaithersburg, MD: AOAC International).
Barreto J. C., Branco A. F., Santos G. T., Magalhães V. J. de A., Coneglian S. M., Teixeira S. (2009). Avaliação dos efeitos de fontes de fósforo na dieta sobre parâmetros do meio ruminal e eficiência de síntese microbiana, digestibilidade dos nutrientes e fósforo plasmático em bovinos. Rev. Bras. Zootecnia 38, 760–769. doi: 10.1590/S1516-35982009000400024
Beaton A. E., Tukey J. W. (1974). The fitting of power series, meaning polynomials, illustrated on bandspectroscopic data. Technometrics 16, 147–185. doi: 10.1080/00401706.1974.10489171
Burnham K. P., Anderson D. R. (2004). Multimodel inference: understanding AIC and BIC in model selection. Sociological Methods Res. 33, 261–304. doi: 10.1177/0049124104268644
Butterworth M. H. (1985). “Phosphorus deficiencies and their correction,” in Beef cattle nutrition and tropical pastures. Ed. Butterworth M. H. (Logman, London), 40–75.
Coneglian S. M., Branco A. F., Guimarães K. C., Mano D. S., Barreto J. C., Fávaro V. R. (2010). Replacement of dicalcium phosphate by rock phosphate in cattle diets: nutrients digestibility, plasma parameters, ruminal fermentation and microbial synthesis efficiency. Rev. Bras. Zootec. 39, 815–823. doi: 10.1590/S1516-35982010000400017
Dixon R. M., Anderson S. T., Kidd L. J., Fletcher M. T. (2020a). Management of phosphorus nutrition of beef cattle grazing seasonally dry rangelands: a review. Anim. Production Sci. 60, 863–879. doi: 10.1071/AN19344
Dixon R. M., Coates D. B. (2011). The relationship between the concentration of phosphorus in the diet and in faeces of cattle grazing tropical grass and grass-legume pastures. Adv. Anim. Biosci. 2, 337.
Dixon R. M., Coates D. B., Reid D. J. (2018). The effects of supplementary P in the diet on the concentration of P in faeces. Anim. Production Sci. 58, 2600.
Dufreneix F., Faverdin P., Peyraud J. L. (2019). Influence of particle size and density on mean retention time in the rumen of dairy cows. J. Dairy Sci. 102, 3010–3022. doi: 10.3168/jds.2018-15926
Durand M., Kawashima R. (1980). “Influence of minerals in rumen microbial digestion,” in Digestive Physiology and Metabolism in Ruminants. Eds. Ruckebusch, Thivend (Dordrecht: Springer), 375–383. doi: 10.1007/978-94-011-8067-2_18
Durand M., Komisarczuk S. (1988). Influence of major minerals on rumen microbiota. J. Nutr. 118, 249–260. doi: 10.1093/jn/118.2.249
Kimura F. T., Miller V. L. (1957). Improved determination of chromic oxide in cal feed and feces. J. Agric. Feedstuffs Chem. 5, 216. doi: 10.1021/jf60073a008
Komisarczuk S., Merry R. J., Mc Allan A. B. (1987). Effects of different levels of phosphorus on rumen microbial fermentation and synthesis determined using a continuous culture system. Br. J. Nutr. 57, 279–290. doi: 10.1079/BJN19870033
Licitra G., Hernandez T. M., Van Soest P. J. (1996). Standardization of procedures for nitrogen fractionation of ruminants feeds. Anim. Feed Sci. Technol. 57, 347–358. doi: 10.1016/0377-8401(95)00837-3
Matis J. H., Wehrly T. E., Ellis W. C. (1989). Some generalized stochastic compartment models for digesta flow. Biometrics 45, 703–720. doi: 10.2307/2531678
Mertens D., Allen M., Carmany J., Clegg J., Davidowicz A., Drouches M., et al. (2002). Gravimetric determination of amylase-treated neutral detergent fiber in feed swith refluxing in beaker or crucibles: collaborative study. J. AOAC Int. 85, 1217–1240.
Preston L. R., Jacobson N. L., Wiggers K. D., Wiggers M. H., Jacobson G. N. (1977). Phosporus in ruminant nutrition (Iowa: National Feed Ingredients Association), 43p.
Preston R. L., Pfander W. H. (1964). P metabolism in lambs fed varying P intakes. J. Nutr. 83, 369–378. doi: 10.1093/jn/83.4.369
Processi E. F., Fontes C. A. A., Vieira R. A. M. (2016). Degradability and kinetics of fibrous particles in cattle fed diets based on corn silage and concentrates with or without addition of lipids. Rev. Bras. Zootecnia (Online) 45, 773–780. doi: 10.1590/s1806-92902016001200007
Regadas Filho J. G. L., Tedeschi L. O., Vieira R. A. M., Rodrigues M. T. (2014). Assessment of the heterogeneous ruminal fiber pool and development of a mathematical approach for predicting the mean retention time of feeds in goats. J. Anim. Sci. 92, 1099–1109. doi: 10.2527/jas.2013-6866
Rohem Júnior N. M., Silva M. C., Abreu M. L. C., Oliveira J. G., Glória L. S., Tedeschi L. O., et al. (2020). The transit of external markers throughout the ruminant digestive tract: 1. The fitting quality of models to marker profiles in feces using an information-theoretic approach. Anim. Feed Sci. Technol. 261, 114407. doi: 10.1016/j.anifeedsci.2020.114407
Sasabuchi I. T. M., Krieger K. S., Nunes R. S., Ferreira A. C., Xavier G. T. M., Urzedo A. L., et al. (2023). Sustentabilidade no uso de fósforo: uma revisão bibliográfica com foco na situação atual do estado de são paulo, brasil. Química Nova 46, 185–198. doi: 10.21577/0100-4042.20170967
Satter L. D., Klopfenstein T. J., Erickson G. E., Powell J. M. (2005). Phosphorus and dairy/beef nutrition. Faculty Papers Publications Anim. Sci. 46, 587–606. doi: 10.2134/agronmonogr46.c19
Schatz T. J., McCosker K. D., Heeb C. (2023). Phosphorus supplementation improves the growth and reproductive performance of female Brahman cattle grazing phosphorus deficient pastures in the Victoria River District, Northern Territory, Australia. Anim. Production Sci. 63, 544–559. doi: 10.1071/AN21565
Suttle N. (2010). “Mineral nutrition of livestock: Fourth edition,” in Mineral Nutrition of Livestock, 4th ed. (CABI) N. Suttle (Ed.). doi: 10.1079/9781845934729.0000
Udén P., Colucci P. E., Van Soest P. J. (1980). Investigation of chromium, cerium and cobalt as markers in digesta. Rate of passage studies. J. Dairy Sci. 31, 625. doi: 10.1002/jsfa.2740310702
Underwood E. J., Suttle N. F. (2001). The mineral nutrition of livestock. 3.ed (Wallingford: CABI Publishing), 614p.
Van Soest P. J. (1994). Nutritional ecology of the ruminant. 2.ed (Ithaca: Constock Publishing Associates), 476p.
Vieira R. A. M., Pereira J. C., Malafaia P. A. M., Queiroz A. C. (1997). The influence of elephant-grass (Pennisetum purpureum Schum., Mineiro variety) growth on the nutrient kinetics in the rumen. Anim. Feed Sci. Technol. 67, 151–161. doi: 10.1016/S0377-8401(96)01130-3
Vieira R. A. M., Rohem N. M., Abreu M. L. C., Da Silva M. C., De Oliveira J. G., Orlindo Tedeschi L., et al. (2020). The transit of external markers throughout the ruminant digestive tract: 2. The estimation of fiber digestibility, ruminoreticular fill, and related biases. Anim. Feed Sci. Technol. 261, 114420. doi: 10.1016/j.anifeedsci.2020.114420
Vieira R. A. M., Tedeschi L. O., Cannas A. (2008a). A generalized compartmental model to estimate the fibre mass in the ruminoreticulum: 1. Estimating parameters of digestion. J. Theor. Biol. 255, 345–356. doi: 10.1016/j.jtbi.2008.08.014
Vieira R. A. M., Tedeschi L. O., Cannas A. (2008b). A generalized compartmental model to estimate the fibre mass in the ruminoreticulum: 2. Integrating digestion and passage. J. Theor. Biol. 255, 357–368. doi: 10.1016/j.jtbi.2008.08.013
Vieira R. A. M., Tedeschi L. O., Cannas A. (2012). A generalized compartmental model to estimate the fibre mass in the ruminoreticulum: 1. Estimating parameters of digestion. J. Theor. Biol. 255, 345–356. doi: 10.1016/j.jtbi.2008.08.014
Williams C. H., David D. J., Lismaa O. (1962). The determination of chromic oxide in faeces samples by atomic absorption spectrometry. J. Agric. Sci. 59, 381–385. doi: 10.1017/S002185960001546X
Witt K. E., Owens F. M. (1983). Phosphorus ruminal availability and effects on digestion. J. Anim. Sci. 56, 930–937. doi: 10.2527/jas1983.564930x
Wu Z., Satter L. D., Sojo R. (2000). Milk production, reproductive performance, and fecal excretion of phosphorus by dairy cows fed three amounts of phosphorus. J. Dairy Sci. 83, 1028–1041. doi: 10.3168/jds.S0022-0302(00)74967-8
Keywords: by-products, ruminal fiber transit kinetics, seed harvest straw, passage rate, phosphorus source
Citation: Couto CdJ, Braga BC, Gomes RdS, Tamy WP, Villela SDJ and Leonel FdP (2025) Evaluation of phosphate sources using ruminal fiber transit kinetics methodology. Front. Anim. Sci. 6:1490835. doi: 10.3389/fanim.2025.1490835
Received: 03 September 2024; Accepted: 06 January 2025;
Published: 30 January 2025.
Edited by:
James Levi Klotz, United States Department of Agriculture, United StatesReviewed by:
Ravikanth Reddy Poonooru, University of Missouri, United StatesBenjamad Khonkhaeng, Rajamangala University of Technology Isan, Thailand
Copyright © 2025 Couto, Braga, Gomes, Tamy, Villela and Leonel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fernando de Paula Leonel, ZmVybmFuZG9sZW9uZWxAdWZzai5lZHUuYnI=; Bruna Cardoso Braga, YnJhZ2EuYnJhZ2FAZGlzY2VudGUudWZnLmJy