 Froylán A. Rodríguez-Soriano1*†
Froylán A. Rodríguez-Soriano1*† Carlos López-Coello2†
Carlos López-Coello2† Ernesto Ávila-González3†
Ernesto Ávila-González3† José Arce-Menocal4†
José Arce-Menocal4† Vitor Barbosa Fascina5Silvestre Chárraga-Aguilar6
Vitor Barbosa Fascina5Silvestre Chárraga-Aguilar6- 1Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, Ciudad de México, Mexico
- 2Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, Departamento de Medicina y Zootecnia de Aves, Ciudad de México, Mexico
- 3Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, Centro de Enseñanza, Investigación y Extensión en Producción Avícola (CEIEPAv), Ciudad de México, Mexico
- 4Facultad de Medicina Veterinaria y Zootecnia, Departamento de Producción Avícola, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico
- 5DSM Nutritional Products LATAM, São Paulo, Brazil
- 6DSM Nutritional Products México, El Salto, Jalisco, Mexico
Introduction: This study aimed to evaluate the effects of a novel sfericase protease—an endopeptidase from the serine protease family, subtilisin subfamily A (MEROPS ID S08.113)—combined with phytase and xylanase on broiler performance, gut morphology, litter quality, and ileal digestibility.
Methods: A total of 1,800 Cobb 500 male chickens were randomly allocated into 36 pens with four dietary treatments and nine replicates per treatment (50 birds per replicate) until 42 days of age. The treatments included: (1) a positive control (PC) following adapted Cobb nutritional recommendations; (2) a negative control (NC) with crude protein and amino acid reductions (0.500% crude protein, 0.041% digestible lysine, 0.040% digestible methionine + cysteine, 0.049% digestible threonine, and 0.032% digestible arginine) compared to PC; (3) NC supplemented with 10,000 new feed protease units (NFP)/kg feed; and (4) NC supplemented with 30,000 NFP/kg feed. Diets, provided in mash form, consisted of corn, soybean meal, canola meal, distiller’s dried grains with solubles, meat and bone meal, corn gluten meal, and soybean oil.
Results: At 42 days of age, broilers fed the NC diet had significantly lower body weight than those in the PC and protease-supplemented groups (p < 0.05). Feed conversion ratio (FCR) was poorest in the NC group, significantly differing from PC and NC + 10,000 NFP/kg (p < 0.05). Gut morphology analysis revealed significant differences in villus length and number, crypt depth, and surface area among treatments (p < 0.05), with the NC exhibiting the lowest surface area per μm². Energy digestibility was lowest in the PC group, whereas protease-supplemented groups (NC + 10,000 and 30,000 NFP/kg) had the highest digestibility values (p < 0.05). Improvements in energy and nitrogen digestibility correlated significantly with body weight, FCR, and gut morphology (p < 0.05).
Discussion: These findings demonstrate that the inclusion of sfericase protease, in combination with phytase and xylanase, positively influences broiler performance, gut morphology, and nutrient digestibility. Optimizing enzyme inclusion based on diet composition and ingredient quality provides practical benefits in commercial broiler production.
1 Introduction
Commercial feed enzymes were first introduced in 1984 to improve the nutritional quality of barley-based rations (Bedford and Partridge, 2001). In the last 15 years, the use of enzymes increased in intensive animal farm production because of their proven effects in reducing antinutritional factors in feeds and improving feed efficiency, digestibility, and bioavailability of nutrients. This has resulted in significant economic and environmental impacts (Dida, 2016). Recently, there has been a growing interest in the effect of enzymes on gastrointestinal functionality, specifically, by reducing substrates for putrefactive microorganisms and increasing substrates for beneficial fermentative organisms, especially with the removal of antibiotics as growth promoters (Cowieson and Kluenter, 2019). In poultry production the use of several types of enzymes, such as phytases, carbohydrases and proteases in feed is common and is based on the diet composition.
Globally phytases are the most widely used feed enzymes and are present in approximately 90% of poultry diets. The use of phytase in feed increases P utilization by hydrolyzing phytate and influences feed conversion, weight gain, egg production, egg traits, mineral availability, and the digestibility of amino acids and energy (Mohamed et al., 2018). Currently, non-starch polysaccharide enzymes in poultry are used worldwide, with a penetration of approximately 70% in poultry diets (Aftab and Bedford, 2018). Xylanases degrade complex non-starch polysaccharides and increase the availability of nutrients, such as starch, protein, and oil, by decreasing digesta viscosity and consequently increasing nutrient absorption (Zhou et al., 2009; Saleh et al., 2018; Raza et al., 2019).
Proteases are also frequently used in the poultry industry and reduce the need for amino acids and energy by improving protein hydrolysis in the presence of anti-nutritional factors such as lectins or trypsin inhibitors (Huo et al., 1993; Ghazi et al., 2002; Cowieson and Roos, 2014). Moreover, proteases may improve ingredient quality by reducing ingredient variability (Cowieson et al., 2016). Since proteases are added to feed to enhance protein hydrolysis and improve nitrogen utilization, there is potential to reduce dietary protein, which can subsequently lower nitrogen levels in manure as observed by Ndazigaruye et al. (2019), when low protein diets where used. Ammonia (NH3) is the primary gas produced in poultry houses as a result of the chemical decomposition of uric acid (the main form of nitrogen excretion in birds) by bacteria present in the litter. The rate of ammonia volatilization depends on factors such as litter pH, humidity, ventilation rate, air velocity, manure nitrogen content and temperature. High ammonia levels have been reported to adversely affect productivity in poultry (>25 ppm) (Swelum et al., 2021). Potentially, if protease supplementation improves nitrogen digestibility, it could lead to reduce uric acid levels in manure, thereby decreasing ammonia emission, as the emissions are directly related to manure nitrogen content. Considering the complex structure of poultry diets and their substrates, proteases are added to poultry diets as part of enzyme mixtures. Since the introduction of the first commercial proteases in the 1990s, their use has grown significantly due to their proven effects (Angel et al., 2011; Cowieson et al., 2019). Extensive research has focused on developing new protease molecules with greater and faster efficiency in protein hydrolysis, enhanced degradation of antinutritional factors, improved stability, and the ability to act on a wide range of raw materials.
After a screening process, a sfericase protease, which is an endopeptidase from the serine protease subtilisin subfamily (MEROPS ID S08.113), was found to be safe and suitable for use in animal production (Cupi et al., 2022). The screening process started with 4,000,000 possible candidates that were narrowed down by exposing the proteases to different tests, including biochemical characterization of feed-relevant parameters (pH, substrate activity, and thermostability), in vitro screening, in vivo digestibility screening, and in vivo extensive performance studies (Cupi et al., 2022). Considering the characteristics of this novel protease observed in this screening process, which are high stability in feed production conditions, a suitable pH profile and a broad specificity that could provide nutritional advantages in practical diets, the objective of this study was to evaluate the efficacy of this newly identified sfericase protease used in combination with a phytase and xylanase, with their respective matrices, in broilers. The combination’s effects on performance, gut health, litter quality, and ileal digestibility were evaluated.
2 Materials and methods
2.1 Facilities and care of experimental animals
The experiment was conducted at Integración y Desarrollo Agropecuario SA de CV experimental poultry farm, Tarimbaro Michoacán, Mexico (19° 48’ N, 101° 10’ W). The location is 1,860 m above sea level with an average temperature of 17.7°C (min, –2.4°C; max, 37.5°C) and has an annual rainfall of 609 mm. Experiments were conducted in accordance with the Official Mexican Norm Guide for Animal welfare; NOM-033SAG/ZOO-2014.
2.2 Experimental design and animal management
One-day-old Cobb 500 male broiler chickens (n = 1,800) were allocated to 36 experimental pens in a completely randomized design with four treatments and nine replicates of 50 chickens each. The birds were housed in an open-sided house with a natural environment and thermal insulation. Each pen (1.8 × 2.5 m) was equipped with two manual 1-L drinkers, a plastic feeding tray, and a mini hopper until seven days of age. From 8–42 days of age, two small feeders and an automatic bell-shaped drinker were used, and the birds were provided feed ad libitum. The diets were provided in mash form and were based on corn, soybean meal, canola meal, distiller’s dried grains with solubles, meat and bone meal, corn gluten meal, and soybean oil. The temperature and lighting programs were based on the Cobb 500 Broiler Management Guide (Cobb-Vantress, 2018). The trial was conducted up to 42 days of age utilizing four feeding phases: Pre-starter (0–7 days), Starter (8–21 days), Grower (22–35 days), and Finisher (36–42 days). For this study, the Cobb nutritional recommendations (Cobb-Vantress, 2022) were adjusted to align with Intensive Commercial Broiler Production Systems (ICBPS) scenarios. The feeding phases were specifically adjusted to optimize productive performance under conditions typically observed on ICBPS. These adaptations represent a common practice in the global poultry industry, where producers tailor guide recommendations to address specific factors, including environmental conditions, management practices, infrastructure, and feed input costs.
The experimental design was as follows:
A positive control (PC) consisted of a diet formulated to meet 100% of an adaptation of Cobb nutritional recommendations (Cobb-Vantress, 2022), specifically aligned with ICBPS scenarios.
A negative control (NC) consisted of a diet with a reduction in crude protein and amino acids compared to the positive control (Table 1). The criteria for reducing crude protein and aminoacids were based on manufacturer recommendations, which consider the inclusion levels of ingredients and the improvements in standardized ileal digestibility observed during in vivo studies (DSM, 2021).
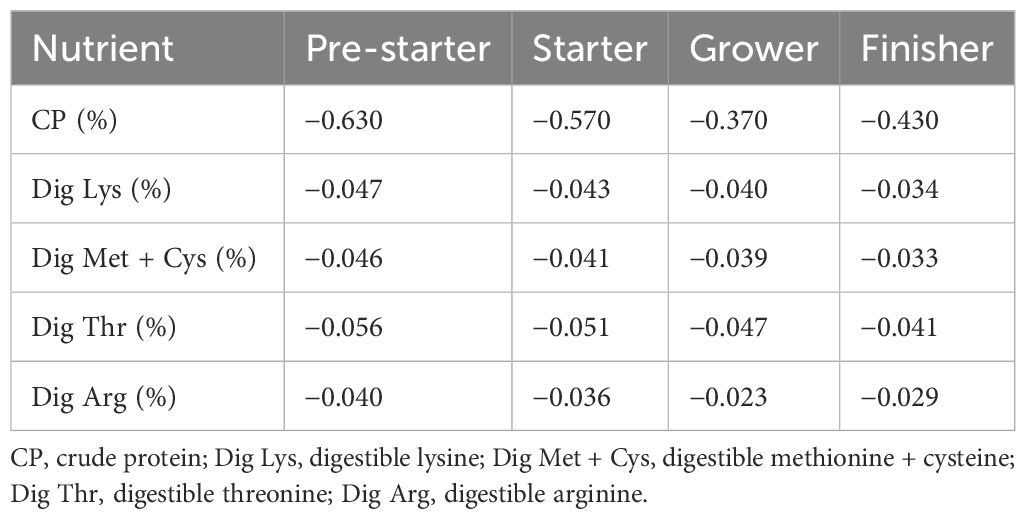
Table 1. Reduction of crude protein and digestible amino acids in the negative control compared to the positive control.
Additional treatments included the NC diet supplemented with sfericase protease at a concentration of 10,000 New Feed Protease units (NFP)/kg feed, and another treatment in which 30,000 NFP/kg feed was added to the NC diet. NFP measures the enzyme amount required to hydrolyze 1 mmol of para-nitroaniline (pNA) from 1 M 128 substrate Suc-Ala-Ala-Pro-Phe-pNA (Cupi et al., 2022).
The used test protease was granulated and contained 600,000 NFP per gram (Proact 360, DSM Nutritional Products Ltd). The additions of 10,000 and 30,000 NFP/kg feed were made at the expense of silica (Sipernat D17, Evonik Industries). All diets included 1,000 phytase units/kg feed (HiPhos GT 20000, DSM Nutritional Products Ltd) and 200 fungal xylanase units/kg feed (Ronozyme WX, DSM Nutritional Products Ltd), with its matrix values for all feeding phases (Table 2). The positive and negative control diets are shown in Tables 3 and 4. Table 5 presents the protease enzyme activity (NFP) analyzed in the finished feeds of treatments supplemented with protease.
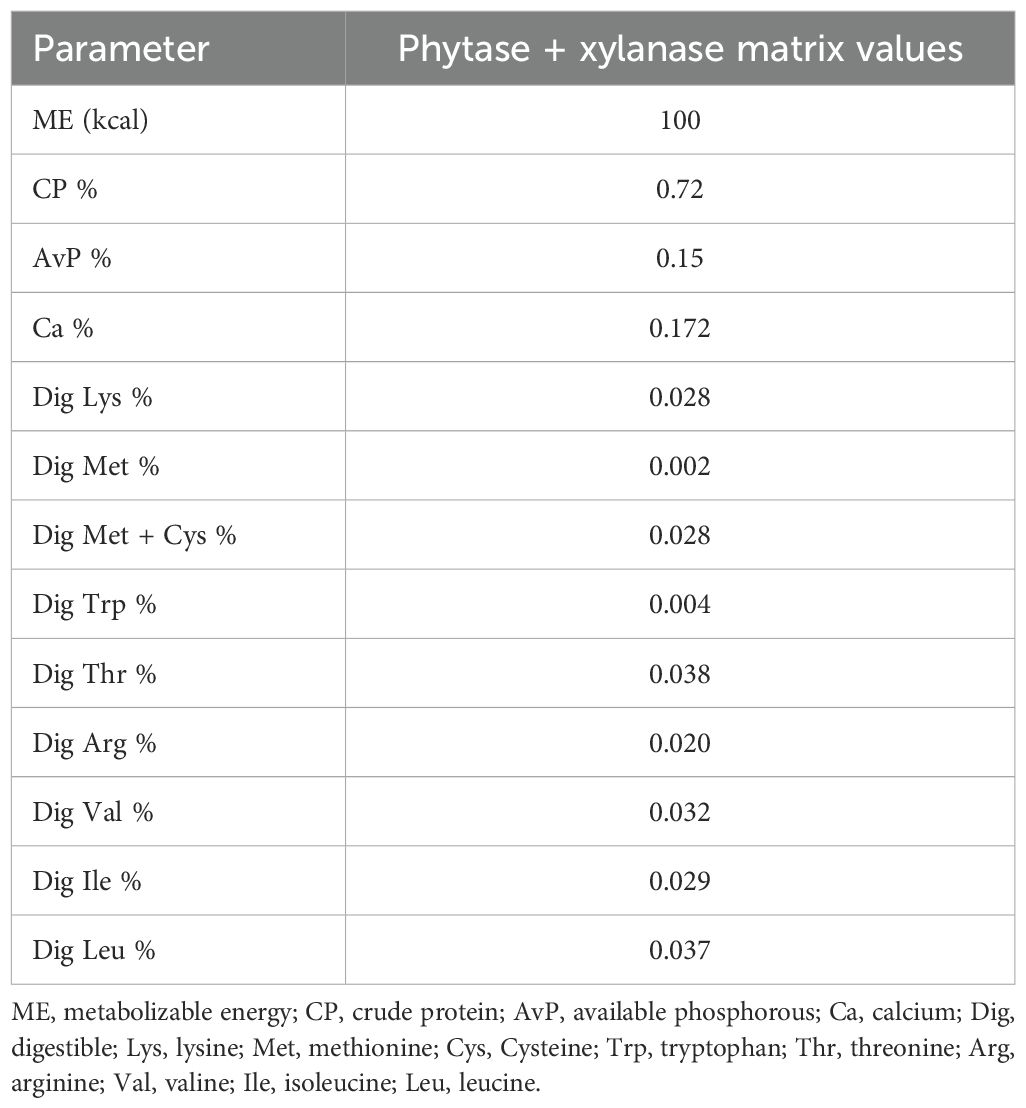
Table 2. Nutritional matrix values for nutrient contribution of phytase and xylanase combination.
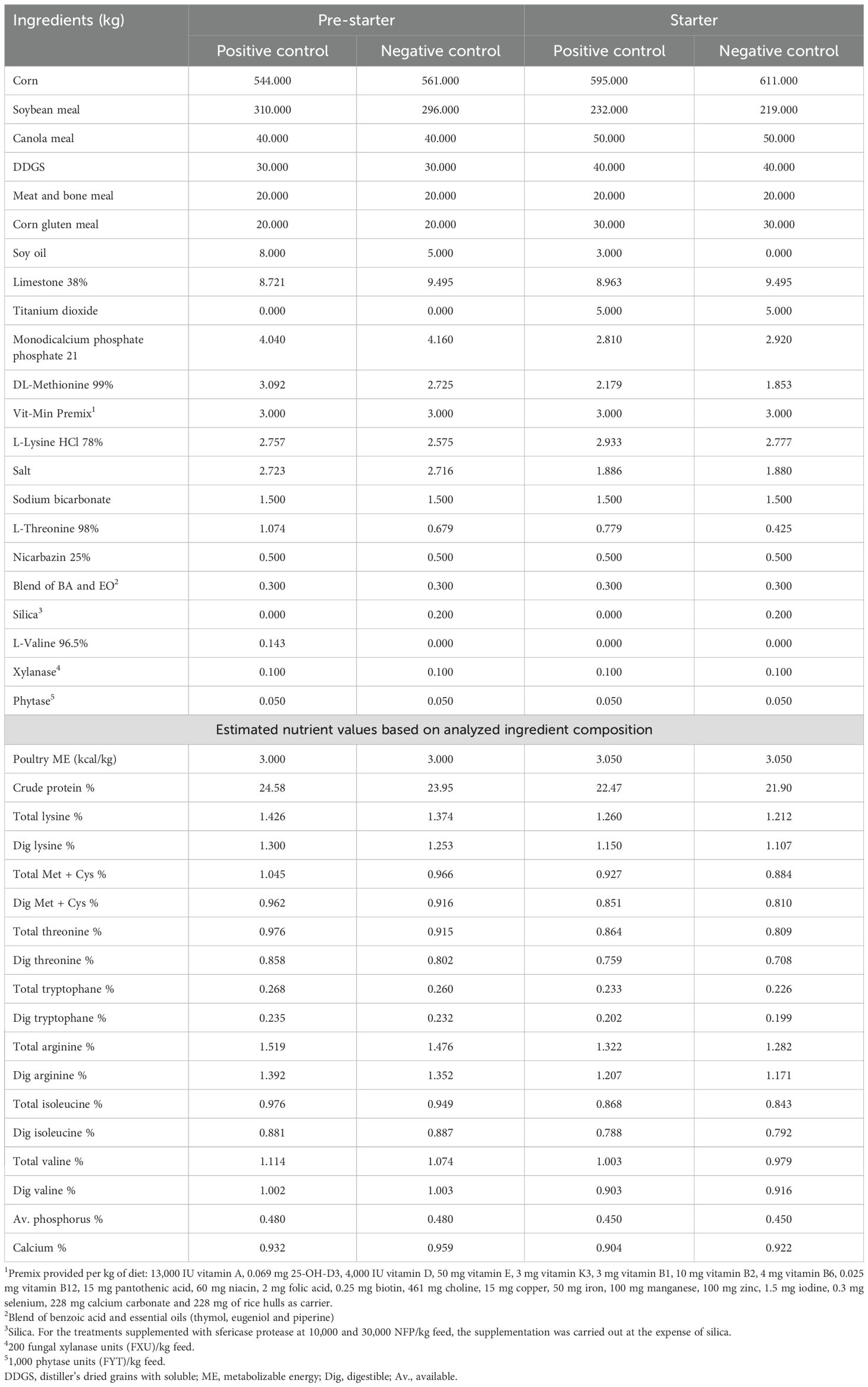
Table 3. Composition of the positive and negative control for the pre-starter and starter diets.
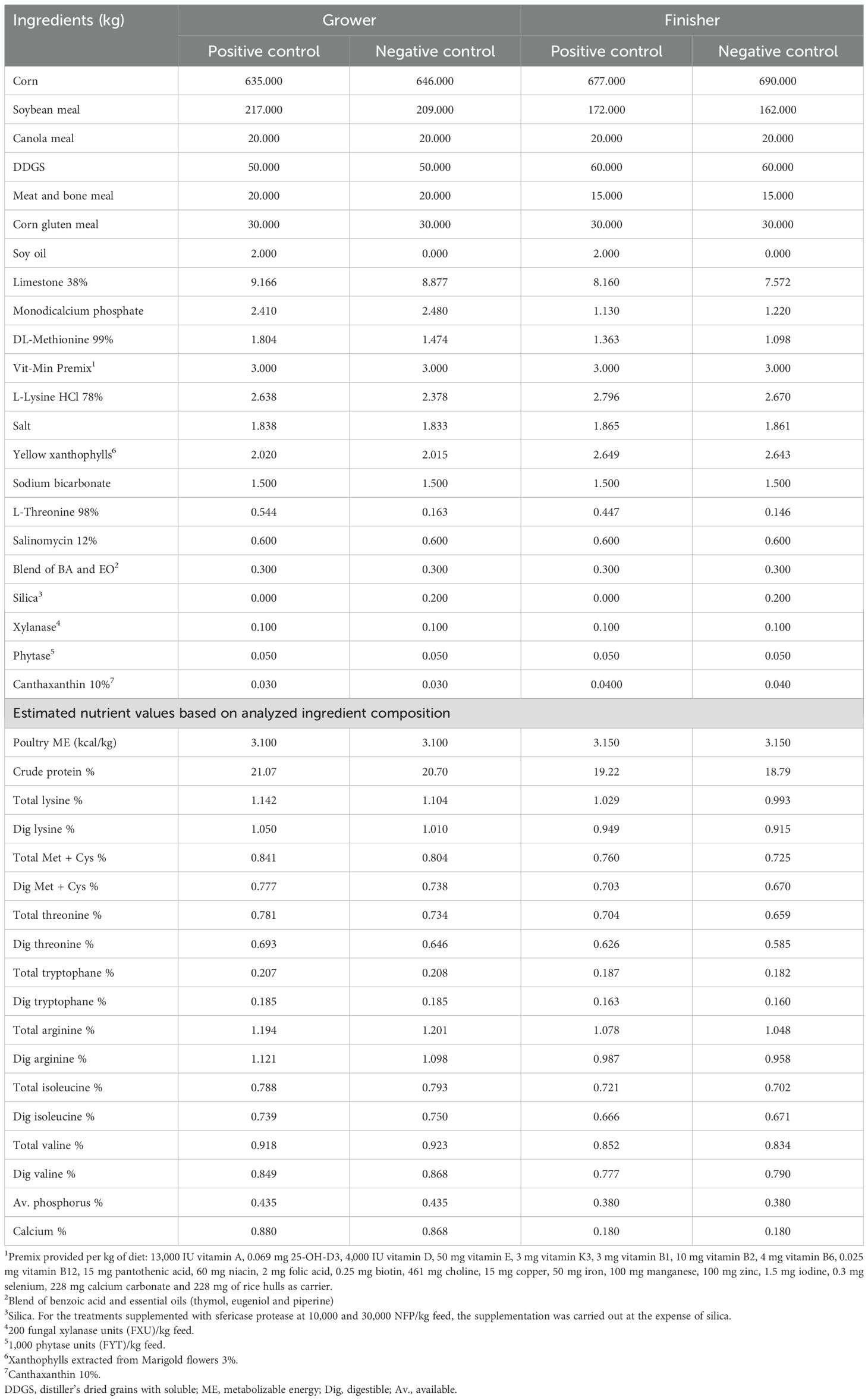
Table 4. Composition of the positive and negative control for the grower and finisher diets.
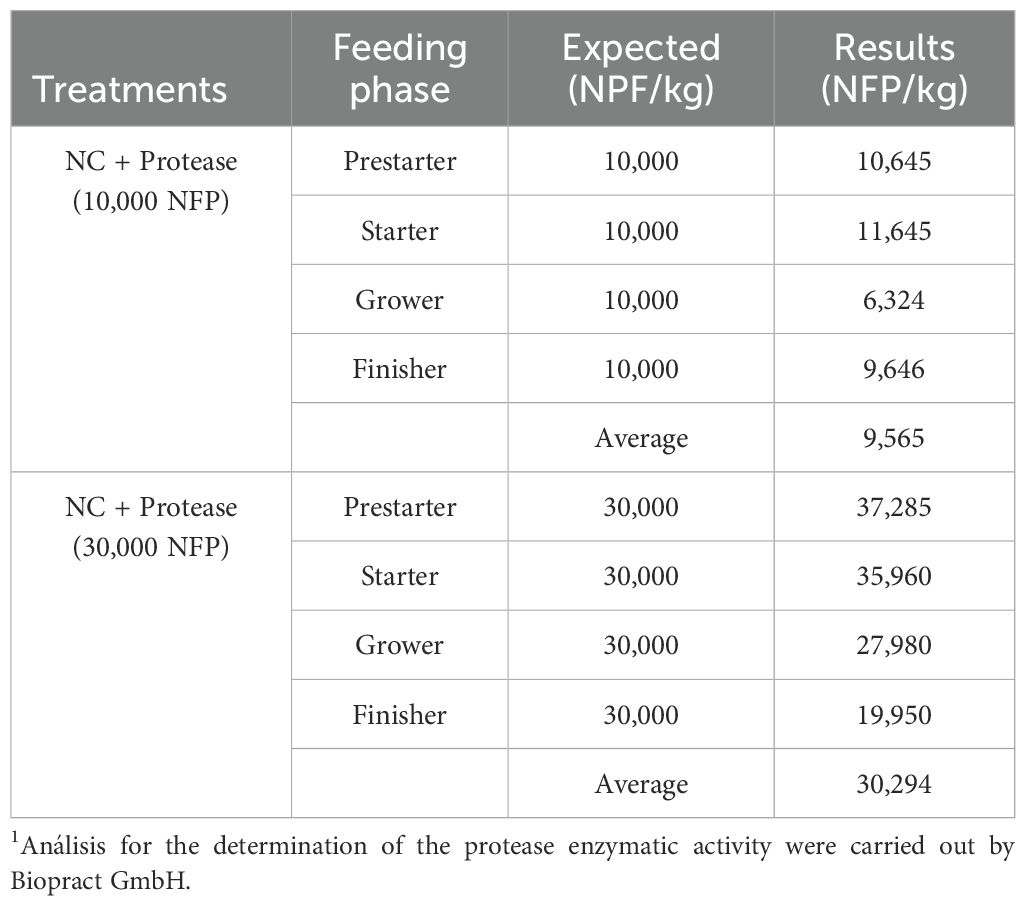
Table 5. Recovery analysis of protease enzyme activity in treatments with Sfericase protease addition.
Prior to formulation, representative samples of corn, soybean meal, canola meal, distiller’s dried grains with solubles, meat and bone meal and corn gluten meal were analyzed to determine their composition. The analysis included dry matter, crude protein, ether extract, crude fiber, ash, starch, acid detergent fiber, neutral detergent fiber, phosphorus, phytic phosphorus, and amino acids (Met, Cys, Met+Cys, Lys, Thr, Trp, Arg, Ile, Leu and Val), using near infrared spectroscopy (AMINONIR, Evonik Industries). Aminoacids coefficients of digestibility where according AMINODAT 6.0 (Evonik Industries).
Representative feed samples of approximately 1 kg were collected from each phase for both PC and NC to undergo proximal chemical analysis. The analyses were conducted as follows: crude protein (CP) was determined using the combustion method (AOAC International, 2012), moisture content was assessed by oven drying at 105°C (AOAC International, 1990a), crude fiber was analyzed using standard procedures (AOAC International, 1990b), and ash content was measured by incineration at 550°C (AOAC International, 1990c). The results obtained are presented in Table 6.

Table 6. Proximal chemical analysis results across different feeding phases for the NC and PC diets.
2.3 Productive performance
Body weight, weight gain, and feed intake were measured weekly. Feed conversion was adjusted for mortality, and the index was calculated per week. Mortality was recorded on daily, including the cause of death.
2.4 Ileal digestibility
Ileal digestibility was measured using titanium dioxide as an indigestible marker. On day 22, three birds per replicate were humanely euthanized in compliance with NOM-033SAG/ZOO-2014 to collect ileal digesta samples from the section between Meckel’s diverticulum and 2 cm before the ileocecal junction. Samples from each replicate were mixed to obtain one sample per replicate, immediately frozen on dry ice, and maintained frozen at −20°C until lyophilization prior to analysis. The ileal digesta samples were analyzed for dry matter (lyophilized), titanium (Myers et al., 2004), nitrogen (AOAC International, 2012), and energy (by adiabatic bomb calorimetry). Three 500g feed samples per treatment of the starter feed were collected and analyzed for titanium dioxide, dry matter, nitrogen, and energy. The average of the feed results per treatment was used to calculate digestibility.
The ileal digestibility of nitrogen and energy was obtained using the digestibility index described by Kong and Adeola (2014), as follows:
In which and are the concentrations of the index compound (TiO2) in the feed and feces, respectively, and and are the concentrations of the components (energy or nitrogen) in the feed and digesta, respectively.
2.5 Evaluation of gut morphology
Gut morphology was evaluated as described by Serrano (Serrano Gamboa, 2024). At 22 days of age, a 1.5-cm section of the ascending duodenal was obtained from one bird per replicate and preserved in 10% formaldehyde with sterile green dye for subsequent image analysis using an Optisum Industrial Digital Camera Model 9.0 MP 1/2.4. The measurements were performed using Motic Image 2.0. Duodenum gut morphology was evaluated by measuring villi length (µm), villi width (µm), Lieberkühn crypt depth (µm), number of villi on a surface of 1,000,000 µm2, and the ratio between villi length and crypt depth. Additionally, the nutrient contact zone surface area was calculated as follows:
2.6 Litter dry matter, pH and total ammoniacal nitrogen and environmental ammonia
Litter samples were collected at 41 days of age. Samples were collected by taking five 100 g samples per pen from five different areas: four samples from the corners and one from the center. The five samples per pen were thoroughly mixed into a composite sample and preserved with dry ice during transportation for further analysis. For determining dry matter, 100 g per sample were dried at 55°C for 48 h and the weight was determined using a precision scale as described by Brauer-Vigoderis et al. (2014).
The pH was determined by placing 10 g of each sample in beakers with 100 mL of distilled water. After shaking and allowing to stand for 30 min, the pH value was obtained using a pH-meter. Total nitrogen was analyzed according to AOAC, 2001.11 (AOAC International, 2012), and ammoniacal nitrogen was analyzed using distillation with MgO according to AOAC 1980 (modified) (AOAC International, 1980). At 41 days of age, ammonia levels were measured in each pen using a previously calibrated GasAlert Extreme device (NH3 0–400 ppm) (BW Technologies, Honeywell, Schaumburg, IL, USA). The device was placed directly on the litter, 1 meter away from the bell-shaped drinker in each pen.
2.7 Statistical analysis
To evaluate differences among treatments, results were analyzed using analysis of variance (ANOVA) based on General Linear Model (GLM) procedure. When statistically significant differences were detected (p < 0.05), Tukey’s test was applied at a significance of α = 0.05. Weekly mortality, ileal digestibility for energy and nitrogen, litter dry matter, and litter nitrogen percentages were transformed using the arcsine function to ensure normality.
A multivariate correlation analyses were performed to evaluate the linear relationships between the analyzed variables, including body weight, adjusted feed conversion, villus length, and digestibility metrics. Pearson’s correlation coefficient was used to assess linear relationships, with significant correlations identified at p < 0.05.
Additional linear regression analyses were conducted to evaluate the influence of ileal digestibility of energy and nitrogen on body weight and adjusted feed conversion. The analysis included data from the negative control (NC) and treatments supplemented with 10,000 and 30,000 NFP/kg of feed to assess the effect of improved digestibility on performance. The models’ coefficients of determination (R²) were used to assess the strength of these relationships, with significance determined at p < 0.05.
All statistical analyses were performed using JMP® software version 18.1.
3 Results
3.1 Productive performance
Table 7 shows the effects of the diets on broilers performance at 42 days of age, with or without the addition of two sfericase protease concentrations. Performance results indicated a significant effect (p < 0.05) on body weight at 42 days. The negative control group showed lower body weight compared to the positive control and the groups supplemented with sfericase protease (10,000 or 30,000 NFP units/kg feed). The highest body weight was observed in the positive control (p < 0.05). The negative control exhibited the worst adjusted feed conversion rate, which was significantly different from that of the positive control and the group supplemented with 10,000 NFP units/kg feed (p < 0.05). No significant differences were observed between the groups supplemented with 10,000 and 30,000 NFP units/kg feed. Feed intake and mortality did not differ among treatments (p > 0.05).

Table 7. Effects of treatments on broiler performance at 42 days of age.
3.2 Ileal digestibility
Table 8 shows results of ileal digestibility for energy and nitrogen at 22 days of age, measured by using titanium dioxide as indigestible biomarker. The positive control showed the lowest energy digestibility among the treatments (71.0%), while the higher digestibility value was observed in treatment supplemented with protease at 10,000 (77.2%) (p < 0.05). No significant differences were observed in nitrogen digestibility among treatments (p = 0.20). Additionally, it was notably that the energy and nitrogen digestibility percentages followed the same trend.
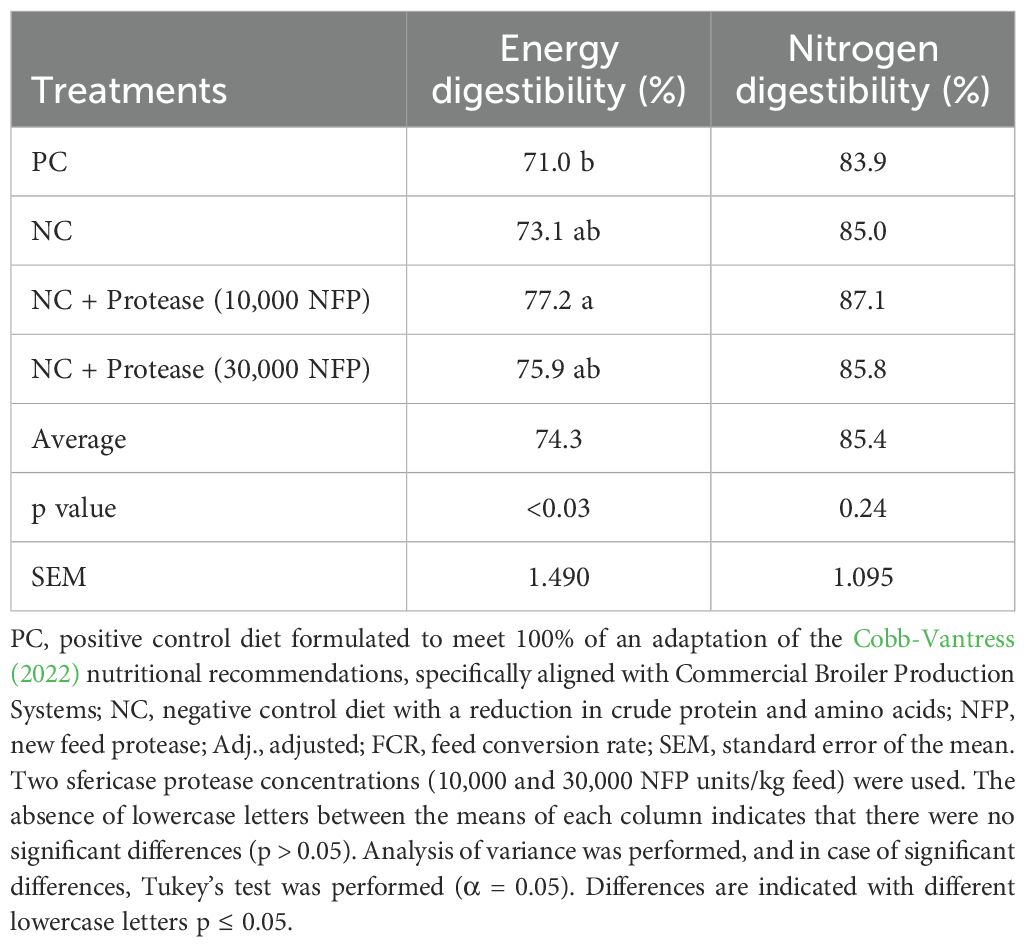
Table 8. Effects of treatments on energy and nitrogen digestibility of broilers at 22 days of age.
3.3 Evaluation of gut morphology
Table 9 presents results of the intestinal villi morphology at 22 days. Significant differences in villus length (p < 0.05) were observed, with the negative control showing the shortest villi length. No significant differences were found between the positive control and the diets supplemented with protease. No differences were observed in villus width (p > 0.05). The Lieberkühn crypt was significantly different among treatments (p < 0.05). The positive control exhibited the shallowest crypt depth, while no differences were observed between the negative control and the diets supplemented with protease. Villi number (1,000,000 µm²) was significantly different among treatments (p < 0.05). The positive control showed the highest number of villi, while the addition of 30,000 NFP/kg feed resulted in the lowest count. The villus surface area per μm2 revealed significant differences among treatments (p < 0.05). The positive control was significantly different from the negative control and the group supplemented with of 10,000 NFP/kg feed; however, no differences were observed between the positive control and the group supplemented with 30,000 NFP/kg feed.
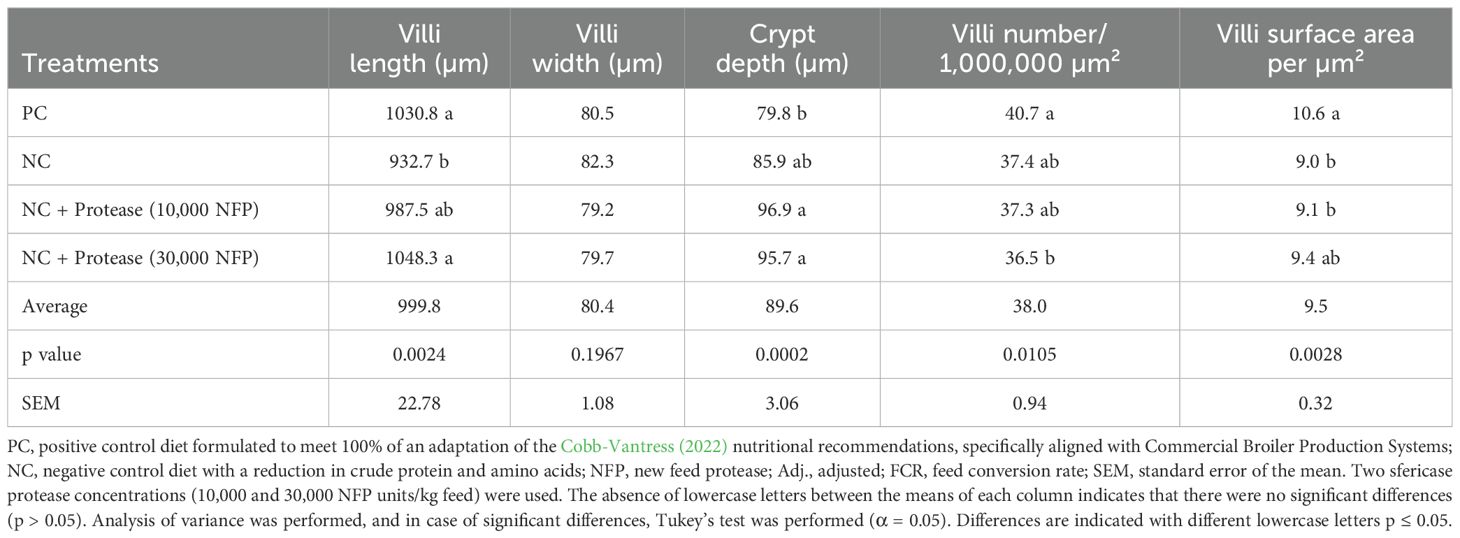
Table 9. Effect of treatments on gut morphology in broilers at 22 days of age.
3.4 Litter dry matter, pH, total nitrogen, ammoniacal nitrogen and environmental ammonia
Litter was kept in an excellent–good condition on day 22 and maintained throughout the experimental period (dry and crumbly). The trial was conducted during season characterized by average temperatures exceeding 19°C and relative humidity below 46%, occurring between March and April. These conditions allowed excellent ventilation and low environmental moisture. The average, maximum and minimum temperatures within the broiler house were 25.2°C, 31.9°C and 19.2°C, respectively, while relative humidity values were 43.1%, 62.3% and 21.8, respectively, throughout the experimental period.
Table 10 shows results for litter dry matter, pH, total nitrogen, ammoniacal nitrogen and environmental ammonia. No significant differences were observed in litter pH, litter nitrogen, or ammoniacal nitrogen (p > 0.05). However, the positive control showed the highest values for litter pH, nitrogen, and ammoniacal nitrogen. Litter dry matter differed among treatments, with the negative control showing a higher value compared to the treatment supplemented with 10,000 NFP/kg feed protease. Environmental ammonia levels did not differ significantly among treatments(p > 0.05).
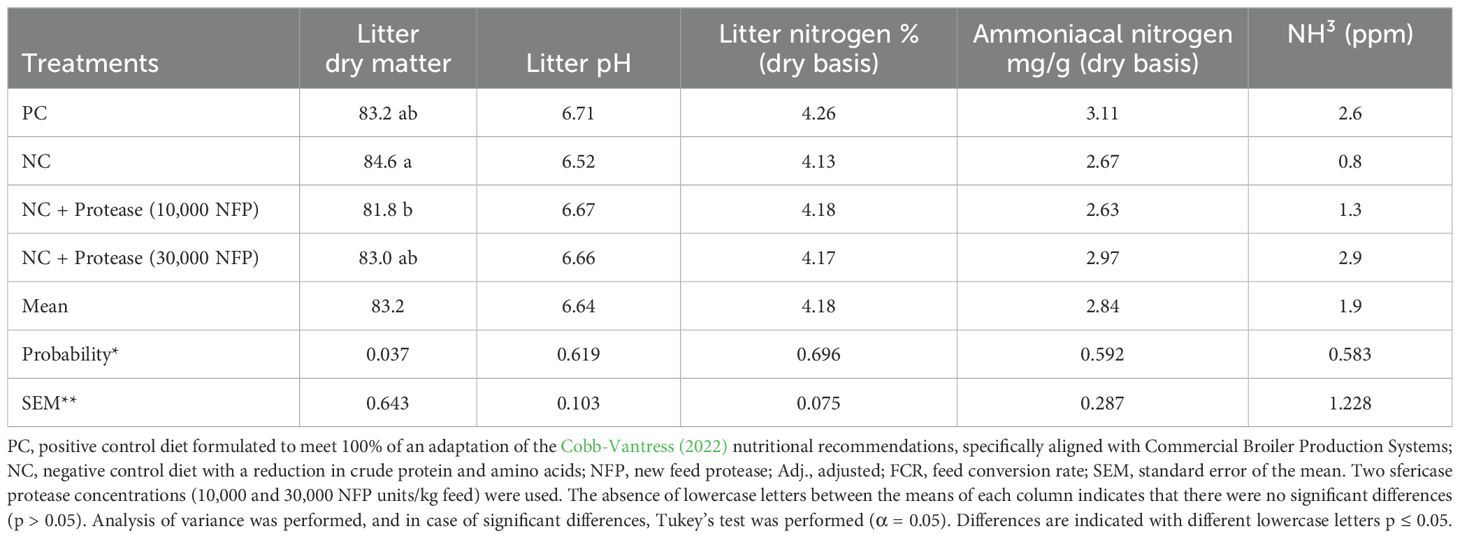
Table 10. Effect of treatments on litter dry matter, pH, total nitrogen, ammoniacal nitrogen and environmental ammonia at 41 days of age.
3.5 Multivariate correlation analysis of the variables
Table 11 presents results of P-values and correlations coefficients, obtained in the multivariate correlation analysis. The multivariate analysis included data from the PC, NC, NC + Protease (10,000 NFP) and NC + Protease (30,000 NFP). The variables analyzed were energy digestibility, nitrogen digestibility, body weight at 42 days, adjusted feed conversion, villi length, villi width, crypt depth, villi number/1,000,000 µm² and villi surface area/µm².
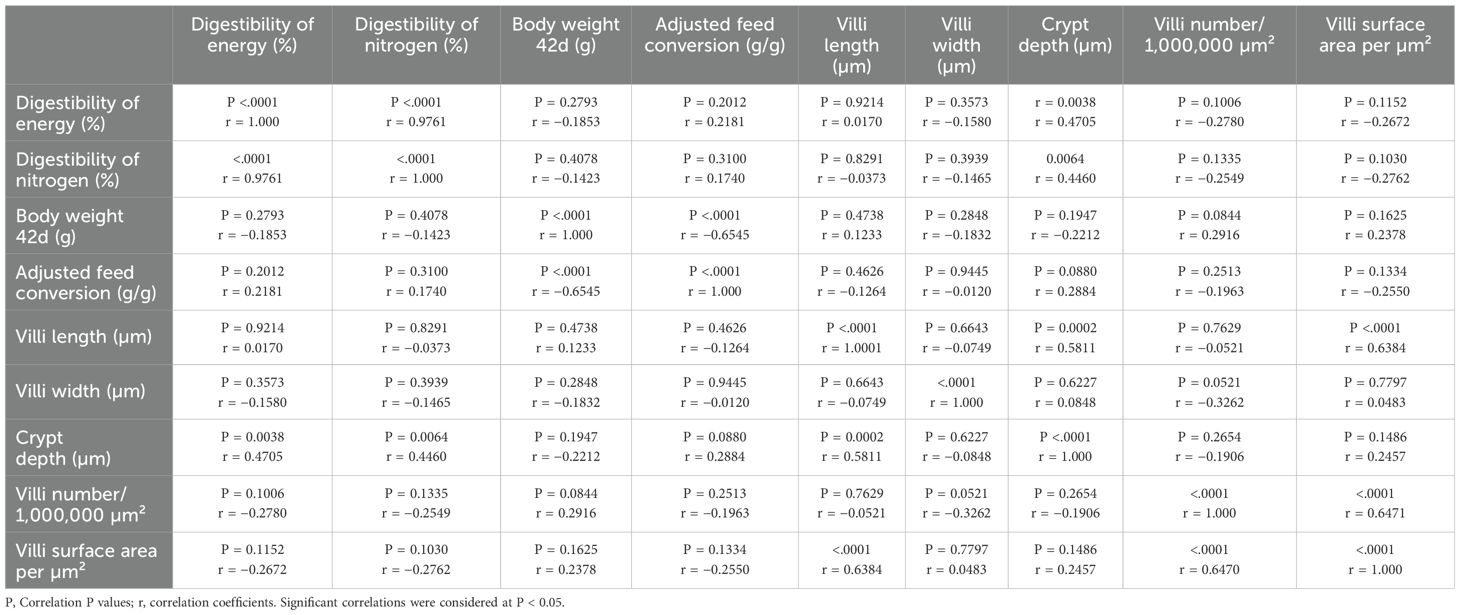
Table 11. Correlation P-values and correlation coefficients obtained in the multivariate analysis for energy digestibility (%), nitrogen digestibility (%), body weight at 42 days (g), adjusted feed conversion (g/g), villi length (µm), villi width (µm), crypt depth (µm), villi number/1,000,000 µm and villi surface area per µm.
Significant correlations (p < 0.05) were as follows:
Energy digestibility with nitrogen digestibility (r = 0.9761) and crypt depth (r = 0.4705).
Nitrogen digestibility with crypt depth (r = 0.4460).
Body weight with adjusted feed conversion (r = -0.6545)
Villi length with crypt depth (r = 0.5811) and villi surface area/µm² (r = 0.6384).
Villi number/1,000,000 µm² with villi surface area/µm² (r = 0.6470).
3.6 Relationship between digestibility metrics and performance variables
The detailed regression results are presented in Table 12. which summarizes the variability explained (R²) and statistical significance of the models. Models were considered significant if p < 0.05.
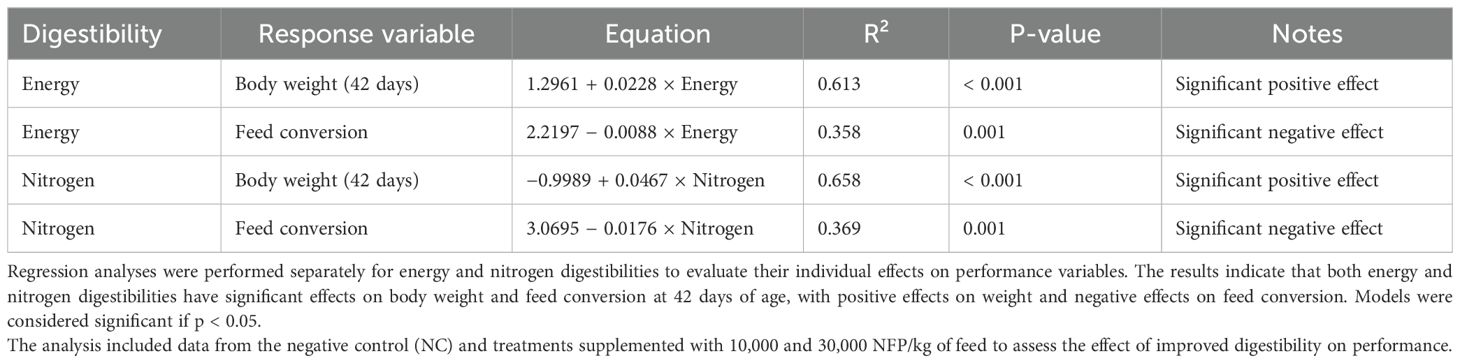
Table 12. Effects of digestibility metrics on broiler performance at 42 days of age.
For body weight at 42 days, the model with digestible energy explained 61.3% of the variability (R² = 0.613, p < 0.001), indicating that 1%increase in digestible energy corresponded to a 0.0228 kg increase in body weight. Similarly, the model using digestible nitrogen explained 65.8% of the variability (R² = 0.658, p < 0.001), with 1% increase in digestible nitrogen leading to a 0.0467 kg increase in body weight.
For adjusted feed conversion, the model with digestible energy explained 30.3% of the variability (R² = 0.303, p = 0.003), where a 1% increase in digestible energy resulted in a reduction of 0.0088 units. In comparison, the model with digestible nitrogen explained 34.4% of the variability (R² = 0.344, p = 0.001), with a 1% increase in digestible nitrogen corresponding to a reduction of 0.0185 units in adjusted feed conversion.
These results suggest that nitrogen digestibility has a slightly stronger influence on both performance variables compared to energy digestibility.
4 Discussion
4.1 Performance parameters
In the present study, differences were observed in broiler body weight and feed conversion at 42 days of age (p < 0.05). The use of sfericase protease resulted in a positive effect on performance as evidenced by the lower efficiency exhibited in the treatment with reduced amino acid and crude protein content without protease supplementation. The response obtained for sfericase protease was observed in the presence of a phytase and xylanase in the diets, with their matrix value considered. The composition of diets and nutrient concentrations play key roles in feed enzyme responses. Thanabalan et al. (2021) and Doskovic̈ et al. (2013) used multienzyme complexes that included carbohydrases, phytases, and proteases and produced variable results. These discrepancies may be attributable to confounding effects resulting from the presence of several enzyme activities. Moreover, these trials did not consider the separate effects of the phytase-carbohydrase and the protease, specifically the effects of proteases in each ingredient of the diet. Therefore, a partitioning of the value for the phytase-carbohydrase and protease combination should be considered. Several studies have reported beneficial effects of monocomponent proteases on feed efficiency (Freitas et al., 2011; Vieira et al., 2013; Ding et al., 2016), body weight gain, and feed intake (Angel et al., 2011). Lee et al. (2023) evaluated the effect of the sfericase protease using similar protease matrix values than in the current experiment and found no differences in BW, BWG, FI, or FCR at 35 days of age. In Lee et al. (2023) experiment, supplementation with exogenous sfericase protease numerically improved these parameters in the same manner as in the present experiment, in which differences were observed; nevertheless, the diet composition was different. In this experiment, ingredients such as canola meal, meat and bone meal, and corn gluten meal were used to calculate the protease matrix value, considering the level of inclusion of each (DSM, 2021). In another trial, Walk et al. (2019) did not find differences in the use of different proteases; nevertheless, in that case, the protease was added to a nutrient-adequate diet and its matrix value according to ingredient inclusion was not considered. As mentioned by Walk et al. (2019), the lack of effects could be associated with the nutrient adequacy of the diet and the digestibility of the adequate diet.
4.2 Ileal digestibility
In previous studies, Cowieson and Roos (2014) showed that the effect on increasing the apparent ileal digestibility on poultry diets of a first-generation monocomponent protease was +4.5% on average for the most critical amino acids (lysine, cysteine, methionine, and threonine) and was dependent on the inherent amino acid digestibility of the control diet. In the current study, as with performance parameters, energy digestibility showed differences among treatments (p < 0.05), even though no differences were observed for nitrogen digestibility (p > 0.05). In a previous study, Lee et al. (2023) reported significant improvements in the apparent total digestibility of protein, arginine, threonine, and glycine with the addition of sfericase protease when compared with that of the negative control. An important consideration regarding the efficiency of enzyme use is the amount of substrate required to improve digestibility, and proteases are known to differ in their efficacy due to inherent characteristics such as stability, activity at specific pH levels, and substrate specificity.
4.3 Gut morphology
The gastrointestinal system digests and absorbs ingested nutrients and excretes waste products from digestion. Most nutrients are ingested as chemical structures that are too complex to absorb. In the gastrointestinal tract, many of these substances are solubilized and further degraded by enzymes to simple molecules absorbed by the mucosal epithelium (Hornbuckle et al., 2008). The morphology of the villi is associated with the absorption of nutrients (Yamauchi et al., 1996; Awad et al., 2009; Zulkifli et al., 2009; Rysman et al., 2023); long villi provide a large absorption area, which is expected to result in improved broiler performance (Laudadio et al., 2012), whereas deeper crypts and shorter villi result in smaller absorption areas, which are expected to result in poor performance (Awad et al., 2009). The use of monocomponent enzymes or combinations of multi-enzymes has been shown to have some effect on gut morphology by increasing villus length (Ayoola et al., 2015; Kim et al., 2021; Lee et al., 2023; Vasanthakumari et al., 2023). The proposed mechanisms underlying the effects of enzymes on gut morphology include a reduction in digesta viscosity, which may stimulate cell division and tissue renewal of intestinal cells, this process can result in shorter villus height or deeper crypt depth, additionally enzymes have shown to increase the production of short fatty acids (Iji et al., 2001; Lee et al., 2017; Kim et al., 2021). Previous studies have shown beneficial effects on gut morphology with the use of proteases (Tajudeen et al., 2022; Lee et al., 2023).
4.4 Nitrogen environmental impact
In the current study, no differences were observed in total or ammoniacal nitrogen or pH. Interestingly, higher values for total and ammoniacal nitrogen were recorded in the positive control. This suggest that the positive control may not have had an excess of nitrogen, leading to significant higher nitrogen output to the litter via feces. Ammonia is a product of the microbial decomposition of uric acid excreted by poultry. Numerous factors affect ammonia emissions from broiler litter, including temperature, humidity, air exchange rate, pH, moisture content, nitrogen content, and litter type (Liu et al., 2006). The present study was conducted in an open house, during season characterized by average temperatures exceeding 19°C and relative humidity below 46%, which allowed for adequate ventilation. These conditions may have favored the control of ammonia production. Additionally, no differences in pH were observed among treatments, with values ranging between 6.5 and 6.71. Reece et al. (1979) demonstrated that ammonia release from the litter was negligible when litter pH was below 7. In the current study, the pH remained threshold. As previously mentioned, the ammonia concentrations measured on day 41 were very low, which could be attributed to the consistent lower levels of the main factors involved in ammonia production through the entire period. Ammonia concentration measured in all treatments were below the maximum recommendations for poultry, which are less than 10 ppm and should not exceed 25 ppm (Ross-Aviagen, 2015; Cobb-Vantress, 2018; Bist et al., 2023).
4.5 Multivariate analysis and correlations
In the present experiment, multivariate analysis was employed to assess responses of multiple variables simultaneously, enabling a deeper understanding of their interrelationships. Significant correlations were observed between energy digestibility and nitrogen digestibility (r = 0.9761), as well as between these metrics and crypt depth (r = 0.4705 and r = 0.4460, respectively). Additionally, villi length was positively correlated with crypt depth (r = 0.5811) and villi surface area (r = 0.6384), supporting the hypothesis that improvements in digestibility metrics are associated with positive changes in intestinal morphology. Regression analysis further demonstrated that nitrogen digestibility had a slightly stronger influence on body weight (R² = 0.658) and feed conversion (R² = 0.344) compared to energy digestibility (R² = 0.613 and R² = 0.303, respectively). A 1% increase in nitrogen digestibility corresponded to an increase of 0.0467 kg in body weight and a reduction of 0.0185 units in feed conversion.
4.6 Study limitations and future directions
This study had a few limitations. Poultry diets used worldwide exhibit significant variability in composition, quality and nutritional profiles. It is essential to consider these differences, including diet compositions (substrate), ingredient quality, potential improvement on digestibility, and partitioning nutrient values for each enzyme used. Further, more extensive research is required to assess the effects of this sfericase protease under diverse production conditions, across different diet types, and in combination with other enzymes, including detailed evaluations of digestibility of individual amino acids.
Additionally, further studies are necessary to elucidate the mechanism by which proteases could exert their beneficial effects on gut morphology, overall health, nutrient absorption processes, and the microbiome. Regarding environmental ammonia emissions, it is crucial to acknowledge the numerous factors influencing these emissions. Measuring blood biomarkers related to nitrogen metabolism, such as uric acid and NH3, could serve as more sensitive indicators of nitrogen metabolism and environmental ammonia emissions.
5 Conclusion
The objective of this study was to evaluate the efficacy of a newly identified sfericase protease, used in combination with phytase and xylanase, in broiler diets. The results showed that this protease enhanced broiler performance, improved ileal nutrient digestibility, and positively influenced gut morphology. Furthermore, litter quality parameters remained unaffected, indicating no adverse environmental impacts. These findings confirm the efficacy of this enzyme in improving broiler productivity and optimizing nutrient utilization.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The experiment was carried out in accordance with Official Mexican Norm (NOM-033-SAG/ZOO-2014) guidelines for animal welfare, and experimental protocols were approved by the Institutional Animal Care and Use Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
FR-S: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. CL-C: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. EA-G: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JA-M: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. SC: Conceptualization, Investigation, Methodology, Supervision, Validation, Writing – review & editing. VF: Supervision, Writing – review & editing, Conceptualization, Funding acquisition, Investigation, Methodology.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by DSM Nutritional Products AG, Wurmisweg 576, CH-4303, Kaiseraugst, Switzerland, and was conducted at the Integración y Desarrollo Agropecuario SA de CV Research Center. Av. Paseo Altozano, 1015 -T2 -1101, 58260; Morelia, Michoacán.
Acknowledgments
We thank DSM Nutritional Products AG, Integración y Desarrollo Agropecuario SA de CV, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias – CENID Fisiología, and the persons involved for their support, including materials, installations, laboratory analysis, and monitoring on the experimental farm. We would like to thank Editage (www.editage.com) for English language editing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2025.1453735/full#supplementary-material
References
Aftab U., Bedford M. R. (2018). The uses of NSP enzymes in poultry nutrition: myths and realities. J. World Poult. Sci. 74, 277–286. doi: 10.1017/S0043933918000272
Angel C. R., Saylor W., Vieira S. L., Ward N. (2011). Effects of a monocomponent protease on performance and protein utilization in 7 to 22 day-old broiler chickens. Poultr. Sci. 90, 2281–2286. doi: 10.3382/ps.2011-01482
AOAC International. (1980). Official methods of analysis of the Association of Analytical Chemists (13th ed.) (Washington, D.C., United States: Association of Official Analytical Chemists).
AOAC International. (2012). Official mehod 2001.11. Protein (crude) in animal feed, forage (plant tissue), grain, and oilseed (Gaithersburg, MD, United States: AOAC International).
AOAC International. (1990a). “Moisture in animal feed – oven drying method,” in Official methods of analysis of AOAC International (Gaithersburg, MD, United States: AOAC International). Official Method 934.01.
AOAC International. (1990b). “Crude fiber in animal feed – weende method,” in Official methods of analysis of AOAC International (Gaithersburg, MD, United States: AOAC International). Official Method 978.10.
AOAC International. (1990c). “Ash in animal feed – muffle furnace method,” in Official methods of analysis of AOAC International (Gaithersburg, MD, United States: AOAC International). Official Method 942.05.
Awad W. A., Ghareeb K., Abdel-Raheem S., Böhm J. (2009). Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poultr. Sci. 88, 49–56. doi: 10.3382/ps.2008-00244
Ayoola A. A., Malheiros R. D., Grimes J. L., Ferket P. R. (2015). Effect of dietary exogenous enzyme supplementation on enteric mucosal morphological development and adherent mucin thickness in Turkeys. Front. Vet. Sci. 2. doi: 10.3389/fvets.2015.00045
Bedford M. R., Partridge G. G. (2001). “Preface,” in Enzymes in farm animal nutrition. Eds. Bedford M. R., Partridge G. G. (Wallingford, United Kingdom: CAB International). doi: 10.1046/j.1439-0396.2001.0335b.x
Bist R. B., Subedi S., Chai L., Yang X. (2023). Ammonia emissions, impacts, and mitigation strategies for poultry production: a critical review. J. Environ. Manage 328, 116919. doi: 10.1016/j.jenvman.2022.116919
Brauer-Vigoderis R., Ferreira-Tinôco I., Pandorfi H., Bastos-Cordeiro M., de-Souza-Júnior J. P., de-Carvalho-Guimarães M. C. (2014). Effect of heating systems in litter quality in broiler facilities in winter conditions. Dyna 81, 36–40. doi: 10.15446/dyna.v81n185.35762
Cobb-Vantress (2018). Cobb broiler management guide. Available online at: https://www.cobb-vantress.com/assets/Cobb-Files/045bdc8f45/Broiler-Guide-2021-min.pdf (Accessed December 20, 2024).
Cobb-Vantress (2022). Cobb500 broiler performance & Nutrition supplement. Available online at: https://www.cobbgenetics.com/assets/Cobb-Files/2022-Cobb500-Broiler-Performance-Nutrition-Supplement.pdf (Accessed December 20, 2024).
Cowieson A. J., Kluenter A. M. (2019). Contribution of exogenous enzymes to potentiate the removal of antibiotic growth promoters in poultry production. Anim. Feed. Sci. Technol. 250, 81–92. doi: 10.1016/j.anifeedsci.2018.04.026
Cowieson A. J., Lu H., Ajuwon K. M., Knap I., Adeola O. (2016). Interactive effects of dietary protein source and exogenous protease on growth performance, immune competence and jejunal health of broiler chickens. Anim. Prod Sci. 57, 252–261. doi: 10.1071/AN15523
Cowieson A. J., Roos F. F. (2014). Bioefficacy of a mono-component protease in the diets of pigs and poultry: a meta-analysis of effect on ileal amino acid digestibility. J. Appl. Anim. Nutr. 2, e13. doi: 10.1017/jan.2014.5
Cowieson A. J., Toghyani M., Kheravii S. K., Wu S. B., Romero L. F., Choct M. (2019). A mono-component microbial protease improves performance, net energy, and digestibility of amino acids and starch, and upregulates jejunal expression of genes responsible for peptide transport in broilers fed corn/wheat-based diets supplemented with xylanase and phytase. Poultr. Sci. 98, 1321–1332. doi: 10.3382/ps/pey456
Cupi D., Thorsen M., Elvig-Jørgensen S. G., Wulf-Andersen L., Berti-Sorbara J. O., Cowieson A. J., et al. (2022). Efficacy and safety profile of a subtilisin protease produced by fermentation in bacillus licheniformis to be used as a feed additive. Heliyon 8, e10030. doi: 10.1016/j.heliyon.2022.e10030
Dida M. F. (2016). Review paper on enzyme supplementation in poultry ration. Int. J. Bioorganic Chem. 1, 1–7. doi: 10.11648/j.ijbc.20160101.11
Ding X. M., Li D. D., Li Z. R., Wang J. P., Zeng Q. F., Bai S. P., et al. (2016). Effects of dietary crude protein levels and exogenous protease on performance, nutrient digestibility, trypsin activity and intestinal morphology in broilers. Livest. Sci. 193, 26–31. doi: 10.1016/j.livsci.2016.09.002
Doskovic̈ V., Bogosavljevic̈-Boskovic̈ S., Pavlovski Z., Miloševic̈ B., Škrbï Z., Rakonjac S., et al. (2013). Enzymes in broiler diets with special reference to protease. J. World’s Poult. Sci. 69, 343–360. doi: 10.1017/S004393391300034
DSM. (2021). DSM proteases matrix calculator. (2021) Available online at: https://proteases.dsm.com/login (Accessed January 10, 2025).
Freitas D. M., Vieira S. L., Angel C. R., Favero A., Maiorka A. (2011). Performance and nutrient utilization of broilers fed diets supplemented with a novel mono-component protease. J. Appl. Poult. Res. 20, 322–334. doi: 10.3382/japr.2010-00295
Ghazi S., Rooke J. A., Galbraith H., Bedford M. R. (2002). The potential for the improvement of the nutritive value of soya-bean meal by different proteases in broiler chicks and broiler cockerels. Br. Poult. Sci. 43, 70–77. doi: 10.1080/00071660120109935
Hornbuckle W. E., Simpson K. W., Tennant B. C. (2008). Gastrointestinal function. Clin. Biochem. Dom. Anim. 14, 413–457. doi: 10.1016/B978-0-12-370491-7.00014-3
Huo G. C., Fowler V. R., Inborr J., Bedford M. R. (1993). “The use of enzymes to denature antinutritive factors in soybean,” in Proc. 2nd int. Workshop on antinutritional factors in legume seed (Wageningen, The Netherlands: EAAP), 60.
Iji P. A., Saki A. A., Tivey D. R. (2001). Intestinal development and body growth of broiler chicks on diets supplemented with non-starch polysaccharides. Anim. Feed. Sci. Technol. 89, 175–188. doi: 10.1016/S0377-8401(00)00223-6
Kim M., Ingale S. L., Hosseindoust A., Choi Y., Kim K., Chae B. (2021). Synergistic effect of exogenous multi-enzyme and phytase on growth performance, nutrients digestibility, blood metabolites, intestinal microflora and morphology in broilers fed corn-wheat-soybean meal diets. Anim. Biosci. 34, 1365–1374. doi: 10.5713/ab.20.0663
Kong C., Adeola O. (2014). Evaluation of amino acid and energy utilization in feedstuff for swine and poultry diets. Asian Australas. J. Anim. Sci. 27, 917–925. doi: 10.5713/ajas.2014.r.02
Laudadio V., Passantino L., Perillo A., Lopresti G., Passantino A., Khan R. U., et al. (2012). Productive performance and histological features of intestinal mucosa of broiler chickens fed different dietary protein levels. Poultr. Sci. 91, 265–270. doi: 10.3382/ps.2011-01675
Lee S. A., Apajalahti J., Vienola K., González-Ortiz G., Fontes C. M. G. A., Bedford M. R. (2017). Age and dietary xylanase supplementation affects ileal sugar residues and short chain fatty acid concentration in the ileum and caecum of broiler chickens. Anim. Feed Sci. Technol. 234, 29–42. doi: 10.1016/j.anifeedsci.2017.07.017
Lee J., Oh H., Kim Y., Song D., An J., Chang S., et al. (2023). Effects of exogenous protease on performance, economic evaluation, nutrient digestibility, fecal score, intestinal morphology, blood profile, carcass trait, and meat quality in broilers fed normal diets and diets considered with matrix value. Poultr. Sci. 102, 102565. doi: 10.1016/j.psj.2023.102565
Liu Z., Wang L., Beasley D. B. (2006). A review of emission models of ammonia released from broiler houses. ASAE Annual Meeting (Portland, MI, United States: American Society of Agricultural and Biological Engineers). Paper 064101. doi: 10.13031/2013.21568
Mohamed E. A. E., Mahmoud A., Muhammad A., Mohamed E., Muhammad S., Muhammad A. A., et al. (2018). The uses of microbial phytase as a feed additive in poultry nutrition–a review. Ann. Anim. Sci. 18, 639–658. doi: 10.2478/aoas-2018-0009
Myers W. D., Ludden P. A., Nayigihugu V., Hess B. W. (2004). A procedure for the preparation and quantitative analysis of samples for titanium dioxide. J. Anim. Sci. 82, 179–183. doi: 10.2527/2004.821179x
Ndazigaruye G., Kim D. H., Kang C. W., Kang K. R., Joo Y. J., Lee S. R., et al. (2019). Effects of low-protein diets and exogenous protease on growth performance, carcass traits, intestinal morphology, cecal volatile fatty acids and serum parameters in broilers. Anim 9, 226. doi: 10.3390/ani9050226
Raza A., Bashir S., Tabbasum R. (2019). An update on carbohydrases: growth performance and intestinal health of poultry. Heliyon 5, e01437. doi: 10.1016/j.heliyon.2019.e01437
Reece F. N., Bates B. J., Lot B. (1979). Ammonia control in broiler houses. Poultr. Sci. 58, 754–755. doi: 10.3382/ps.0580754
Ross-Aviagen (2015). Ross broiler pocket guide. Available online at: https://en.aviagen.com/assets/Tech_Center/BB_Resources_Tools/Pocket_Guides/Ross-Broiler-Pocket-Guide-2015-EN.pdf (Accessed December 12, 2024).
Rysman K., Eeckhaut V., Ducatelle R., Goossens E., Van Immerseel F. (2023). Broiler performance correlates with gut morphology and intestinal inflammation under field conditions. Avian Pathol. 52, 232–241. doi: 10.1080/03079457.2023.2201169
Saleh A. A., Ali H., Abdel-Latif M. A., Emam M. A., Ghanem R., El-Hamid H. S. A. (2018). Exogenous dietary enzyme formulations improve growth performance of broiler chickens fed a low-energy diet targeting the intestinal nutrient transporter genes. PloS One 13, e019808. doi: 10.1371/journal.pone.0198085
Serrano Gamboa M. Y. (2024). Efecto de la inclusión de fitasa, xilanasa y xilo-oligosacáridos en dietas con dos niveles de fibra cruda en el pollo de engorda (Morelia, Michoacán, México: Universidad Michoacana de San Nicolás de Hidalgo). Advisors: Fonseca Madrigal J, López Coello C.
Swelum A. A., El-Saadony M. T., Abd-El-Hack M. E., Abo Ghanima M. M., Shukry M., Alhotan R. A., et al. (2021). Ammonia emissions in poultry houses and microbial nitrification as a promising reduction strategy. Sci. Total Environ. 781, 146978. doi: 10.1016/j.scitotenv.2021.146978
Tajudeen H., Hosseindoust A., Ha S. H., Moturi J., Mun J. Y., Lee C. B., et al. (2022). Effects of dietary level of crude protein and supplementation of protease on performance and gut morphology of broiler chickens. Europ. Poult. Sci. 86. doi: 10.3390/agriculture13030703
Thanabalan A., Mohammadigheisar M., Kiarie E. G. (2021). Amino acids and energy digestibility in extruded or roasted full fat soybean fed to broiler chickens without or with multienzyme supplement containing protease, phytase, and fiber degrading enzymes. Poultr. Sci. 100, 101511. doi: 10.1016/j.psj.2021.101511
Vasanthakumari B. L., Gedye K. R., Abdollahi M. R., Di Benedetto M., Sanchez D. G., Wealleans A., et al. (2023). A new monocomponent xylanase improves performance, ileal digestibility of energy and nutrients, intestinal morphology, and intestinal micro-biota in young broilers. J. Appl. Poult. Res. 32, 1. doi: 10.1016/j.japr.2022.100301
Vieira S. L., Miranda D. J. A., Angel C. R., Favero A., Cruz R. F. A., Sorbara J. O. B. (2013). Effects of a monocomponent protease on performance and protein utilization in 1- to 26-day-of-age Turkey poults. J. Appl. Poult. Res. 22, 680–688. doi: 10.3382/japr.2012-00558
Walk C. L., Juntunen K., Paloheimo M., Ledoux D. R. (2019). Evaluation of novel protease enzymes on growth performance and nutrient digestibility of poultry: enzyme dose response. Poultr. Sci. 98, 5525–5532. doi: 10.3382/ps/pez299
Yamauchi K., Kamisoyama H., Isshiki Y. (1996). Effects of fasting and refeeding on structures of the intestinal villi and epithelial cells in White Leghorn hens. Br. Poult. Sci. 37, 909–921. doi: 10.1080/00071669608417922
Zhou Y., Jiang Z., Lv D., Wang T. (2009). Improved energy-utilizing efficiency by enzyme preparation supplement in broiler diets with different metabolizable energy levels. Poult. Sci. 88, 316–322. doi: 10.3382/ps.2008-00231
Keywords: broilers, protease, Sfericase, digestibility, gut morphology, performance, enzymes combination
Citation: Rodríguez-Soriano FA, López-Coello C, Ávila-González E, Arce-Menocal J, Fascina VB and Chárraga-Aguilar S (2025) Sfericase protease, phytase, and xylanase combination improves body weight, feed conversion rate, ileal digestibility, and gut morphology in broilers. Front. Anim. Sci. 6:1453735. doi: 10.3389/fanim.2025.1453735
Received: 23 June 2024; Accepted: 28 January 2025;
Published: 17 February 2025.
Edited by:
Bianca Castiglioni, National Research Council (CNR), ItalyReviewed by:
José Henrique Stringhini, Universidade Federal de Goiás, BrazilGuanchen Liu, Kansas State University, United States
Copyright © 2025 Rodríguez-Soriano, López-Coello, Ávila-González, Arce-Menocal, Fascina and Chárraga-Aguilar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Froylán A. Rodríguez-Soriano, Y3JveWY0QGV4YWx1bW5vLnVuYW0ubXg=
†These authors have contributed equally to this work and share first authorship