 Julia Stuhlträger1*†
Julia Stuhlträger1*† Eberhard von Borell1†
Eberhard von Borell1† Jan Langbein2†
Jan Langbein2† Christian Nawroth2†
Christian Nawroth2† Maria Vilain Rørvang3†
Maria Vilain Rørvang3† Camille M. C. Raoult1,4†
Camille M. C. Raoult1,4†- 1Institute of Agricultural and Nutritional Sciences, Faculty of Natural Science III, Martin-Luther-University Halle-Wittenberg, Halle (Saale), Germany
- 2Research Institute for Farm Animal Biology, Dummerstorf, Germany
- 3Dept. Biosystems and Technology, Swedish University of Agricultural Sciences, Lomma, Sweden
- 4Department of Agroecology and Environment, ISARA, Lyon, France
Understanding the impact of light on, and how it is perceived by, farm animals is crucial for the design of appropriate, high-welfare housing and management conditions. By considering the animal’s visual needs and adaptive capabilities, future housing and management can allow them to better express their natural behavior. In the past, animal environments have been designed primarily to optimize production output and adjusted to human needs. This emphasis has changed toward a more animal-based focus. However, especially in farmed ungulates, there is still limited knowledge about the impact of light parameters on their physiology, performance, and behavior. This poses the risk of not appropriately assessing the importance of these abilities when the animals interact with their physical environment. Here, we provide an overview of the current state of research on the impact of light parameters on farmed ungulates and their preferences for light settings in the most common farmed ungulate species: pigs, sheep, goats, cattle and horses. We focus on three specific light parameters: photoperiod, illumination, and color. Secondly, we identify gaps of knowledge and emphasize their implications for animal welfare and potential improvement of current animal husbandry environments. There was considerable variation in the number of studies by species and light parameters. A large focus of studies looked at illumination in pigs and color perception in horses. Research on cattle, sheep and goats, seems to be underrepresented in the literature. From a human perspective, we tend to overestimate the importance of color perception and preference, whereas contrast discrimination in combination with illumination intensity and quality seems to be more relevant for ungulate orientation and interaction with their environment. Aside from the importance of other senses and their interaction with vision, we conclude that illumination and photoperiodicity seem to be most relevant for securing the welfare of farm ungulates. These aspects should therefore be given more consideration in indoor housing improvements. Future research emphasis should be given to preference testing studies, as they provide insights into the animals’ motivation for specific light conditions that may further improve their welfare, but also health and performance.
1 Introduction
Animals’ senses are shaped by socio-ecological factors and play a crucial role in an animal's interactions with its environment. Animals’ perceptual abilities are adapted to the biology of the animal and therefore are well adjusted to their natural environment. Prey mammals, for example, often have eyes located laterally on either side of the head offering a wide field of vision to be able to scan their surroundings for predators (Piggins and Phillips, 1996; Harman et al., 1999). Artificial environments, for instance farm animal housing systems, however, differ from natural environments and may not fit the animals’ perceptual adaptations. Therefore, modern housing systems can be a source of stress for the animal, for example through obstructed visibility, or suboptimal light conditions, which in turn can affect their health parameters (e.g., increased body lesions) and natural behavioral patterns (e.g., increased development of behavioral disorders or increase in aggression) (Thornton, 2010; Butterworth, 2013). This emphasizes the importance of having detailed knowledge on, and understanding of, the perception and importance of light periods, considering their duration, intensity, and color to allow natural rhythmicity and species-specific interactions with their environment and avoid exposing farmed animals to suboptimal conditions. In this review, we therefore included the aspects of light intensity (illumination) and color, but also photoperiodicity.
In many farm animals (e.g., cattle, sheep, goats, horses), vision plays a key role when animals collect information about their surrounding environment (Baldwin, 1981; Piggins and Phillips, 1996), during individual recognition (e.g., Briefer et al., 2012; Coulon et al., 2007; Hagen and Broom, 2003; Kendrick et al., 1995), and foraging decisions (Bazely and Ensor, 1989). Vision is one of the most studied senses in domestic mammals [unlike other senses such as olfaction (Nielsen et al., 2015)] - probably due to an anthropomorphic bias, as vision is the dominant sensory modality for humans. Research on visual abilities has, however, been conducted with an imbalance of studies across species. Furthermore, these lines of studies are typically done within species, and comparison of knowledge about ungulate farm animals’ visual abilities have not been attempted. Yet, it may yield a basis for elucidating implications for their welfare under commercial settings.
The first aim of this literature review is to provide an overview of the state of research conducted on pig, sheep, goat, cattle, and horse visual abilities and the role of light, as well as their species-specific preferences, by focusing on three distinct light parameters that affect vision: photoperiod, illumination, and color. The focus on these specific parameters is chosen as they can easily be artificially altered in farm animal housing systems (Figure 1). Secondly, the review aims to identify gaps in knowledge and thirdly to discuss their implications on the animals’ welfare in the current and future housing systems. Within this, we additionally aim to provide directions for future research that may contribute to our understanding of ungulate farm animals’ visual perception and how future housing systems and management routines could be better adapted to form high-welfare systems in the future.
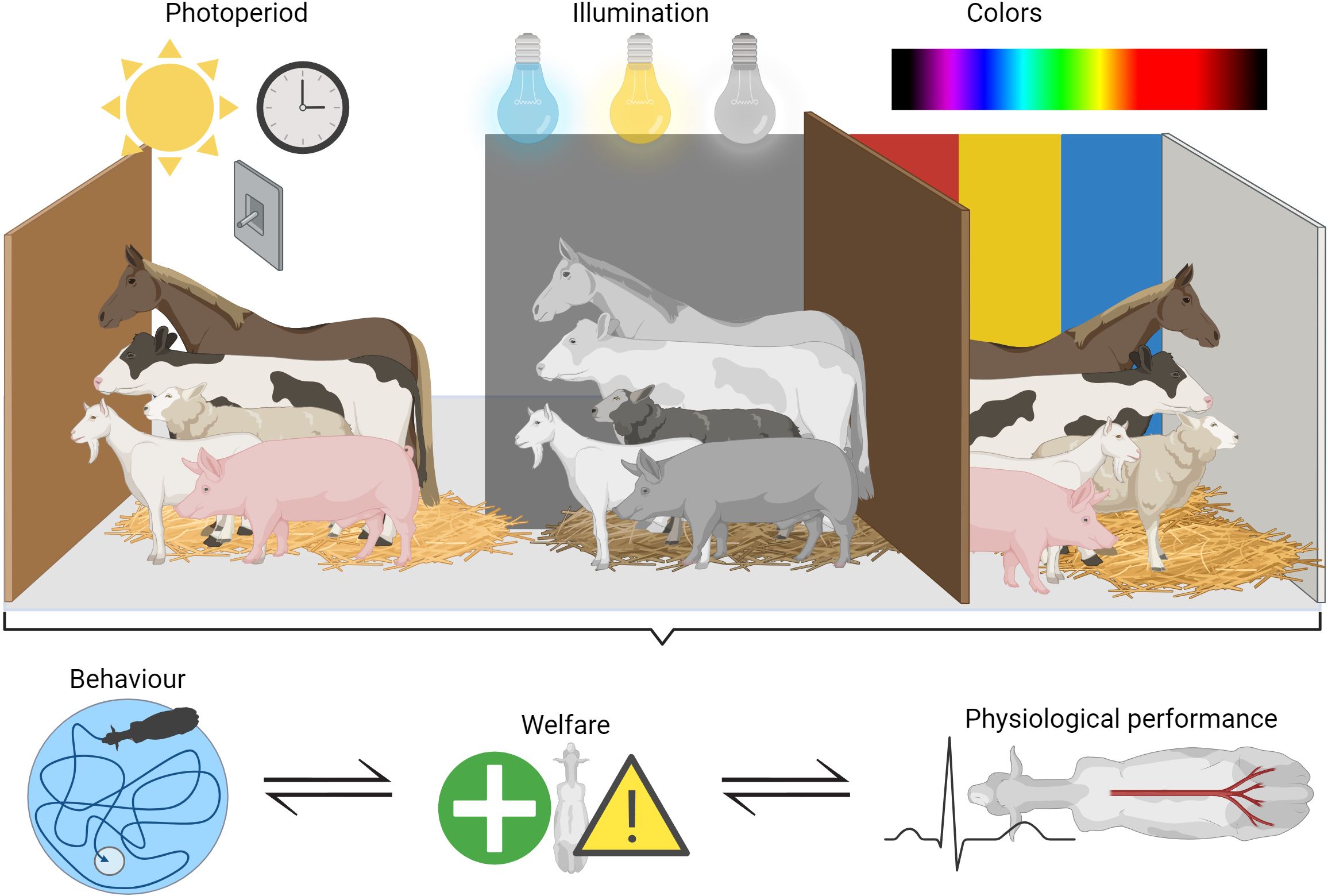
Figure 1. Schematic overview of how the considered light parameters: photoperiod, illumination and color, may affect the considered five production ungulate species: pigs, sheep, goats, cattle and horses. The photoperiod refers to the relative length the animal is exposed to light and dark measured for a 24h period, and is linked to the circadian rhythm of the animal. Illumination is a measure of the incident light an animal (or object) is exposed to. Color refers to colors in the environment of the considered species, which depends heavily on the color vision abilities of each species. All three topics may influence the behavior, welfare and physiology of the considered species, which is illustrated by the collective bracket pointing toward the bottom factors. Drawn by M. V. Rørvang using Biorender.com (publication license obtained) in combination with Power Point.
2 Visual abilities of domestic ungulate species
Domestic ungulates (in particular herbivorous ones, e.g., sheep, goats, cattle, horses) are well adapted to life in open habitats, such as grasslands. The eyes of ungulates are prominent and laterally positioned on either side of the head (Blackshaw et al., 1983; Phillips, 1993). This arrangement offers a wide monocular field of vision, estimated to be 290 – 313° in sheep (Piggins and Phillips, 1996; Kendrick, 2008); 320 – 340° in goats (Piggins and Phillips, 1996); around 330° in cattle (Dimberton, 1999); and 315 – 335° in horses (Timney and Macuda, 2001) with a narrow blind zone at the rear. The monocular field of pigs, whose eyes are less laterally positioned, is still estimated to be around 310° in (Prince, 1977). The large monocular field of vision allows for scanning large horizontal areas and has likely been beneficial for predator detection. In addition to the large monocular field of vision, ungulates also have a small binocular field of vision [35–50° for pigs (Dalmau et al., 2009); 40 – 60° for sheep (Kendrick, 2008); 20 – 60° for goats (Walls, 1942; Hughes and Whitteridge, 1973); 25 – 50° for cattle (Blackshaw et al., 1983); and 55 – 65° for horses (Timney and Macuda, 2001)], which offers a limited stereoscopic vision (i.e., the ability of the brain to combine visual information from both eyes to create a clear three-dimensional image) right in front of them. This means that these species have difficulties perceiving depth and three-dimensional structures (Grandin, 1980; Blackshaw et al., 1983), except for horses who possess true stereopsis (Timney and Keil, 1999) albeit within a limited visual field. Although cattle, sheep and goats cannot assess distance accurately, they have high motion detection (Shrader et al., 2008; Adamczyk et al., 2015). Horses, too, are sensitive to motion, especially when detected in the peripheral visual field. Moreover, most ungulates have astigmatism (i.e., blurred vision both up close and at a distance) and no accommodation (i.e., the ability of the eye to automatically adjust to objects at different distances), and thus a relatively poor visual acuity (Johnson, 1901; Piggins and Phillips, 1996; Adamczyk et al., 2015).
Visual input is important for orientation in space, recognition of flock members and identification of dangers (in combination with hearing) (Blackshaw et al., 1983; Adamczyk et al., 2015). For instance, cattle use vision for foraging and social recognition (Cummins and Myers, 1991; Nawroth et al., 2019), while sheep, goats, and horses also utilize sight to navigate their environment (Culda and Stermin, 2019; Rørvang et al., 2020) and communicate with conspecifics. Pigs are known to have a poor ability to distinguish details and shapes (Zonderland et al., 2008) and do not primarily rely on their visual abilities.
With regard to vision in ungulates per se, these species have two major types of photoreceptors: rods, which is the primary component of the retina and function under low light intensities; and cones, which function in bright light and provide a better visual acuity (i.e., the ability to detect and discriminate details of an object from a given distance) as well as color sensitivity (Greeff, 1894; Braekevelt, 1983). Both photoreceptors have been identified in the retina of horses, cattle, sheep and pigs (Zurn, 1902). The rod-to-cone ratio is 30-40:1 for sheep (Braekevelt, 1983), 20:1 for horses (Wouters and De Moor, 1979), 8:1 on average for pigs (ranging from 3:1 centrally to 16:1 peripherally) (Chandler et al., 1999) and 2-6:1 for cattle (Gilbert and Arave, 1986). Ungulates’ visual system is thus adapted to both photopic (i.e., daylight light intensity vision) and scotopic (i.e., low light vision) conditions (Graf, 1976; Tanida et al., 1991; Ollivier et al., 2004; Hanggi and Ingersoll, 2009; Murphy et al., 2009). In particular, sheep, goats and horses have horizontally elongated slit-shaped pupils (i.e., the pupils can rotate to remain horizontal and allow a greater range of light control), which allow them a clear and sharp horizontal panoramic view all around them while foraging (Banks et al., 2015), as well as a tapetum lucidum, i.e., a light-reflective tissue in the back of the retina (also present in cows). This tissue allows for greater absorption of the light entering the eye (Land, 2006), resulting in good achromatic vision (i.e., low light to night vision), which is thought to aid in the detection of predators (Jacobs et al., 1998; Ollivier et al., 2004; Roth et al., 2008). Pigs have circular (to sightly horizontally oval) pupils and no tapetum (Vestre, 1984; Banks et al., 2015). Besides their ability to see at different light levels, ungulates are also able to perceive colors. Color vision in ungulates is associated with the presence of two spectrally distinct cone types, one sensitive to short wavelengths (440 – 451 nm; S-cones) and the other to middle to long wavelengths (552 – 557 nm; M/L-cones) (Neitz and Jacobs, 1989; Jacobs et al., 1998; Lomas et al., 1998; Carroll et al., 2001). The presence of these cone types provides the basis for dichromate color vision and enables these animals to perceive blue and yellow to green spectra (Jacobs et al., 1998). Dichromatic vision has advantages for motion detection over trichromatic vision (Kendrick, 2008), and is therefore vital for prey animals.
These above descripted characteristics of the visual system, that partly vary among domesticated ungulates, suggest that they perceive their environment differently, which implies a need for species-specific adaptations when ungulates are housed in artificial farm environments. The following sections will refer to three selected light parameters that influence such indoor farm animal environments, namely photoperiod (i.e. light duration), illumination (i.e. light intensity) and color (i.e. colors and colored light). For this narrative review, we identified peer-reviewed studies published, or in press and available online, with no specific date limit but until November 11th 2024, by searching the Web of Science (http://www.webofknowledge.com) using the key word combination ‘(animal AND visual perception)’, as well as replaced within this combination the key word ‘animal’ by the species, i.e. ‘pig’ OR ‘swine’, OR ‘sheep’, OR ‘goat’, OR ‘cattle’ OR ‘cow’ OR ‘calf’, OR ‘horse’. We used additional searches consisting of the following key word combinations: ‘(illumination AND ‘species’)’, ‘(color vision AND ‘species’)’, ‘(light color AND ‘species’)’, and ‘(colored light AND ‘species’). Only studies that concerned the targeted farmed ungulate species (i.e. cattle, sheep, goat, pig and horse) and foci (sorted according to perception, physiological performance, and behavior) were considered. Review studies’ cited references were checked for additional potential missed studies. In addition, we checked for grey literature on the web search engine and various databases. A table with selected relevant studies is available as Supplementary Material. After reviewing the state of research for each species and for each parameter, we provide the main implications for housing and welfare and future directions.
2.1 Photoperiod
Photoperiod is linked to the circadian rhythm of the animal and is defined as the relative length the animal is exposed to light and dark measured for a 24h period (American Heritage Dictionary of the English Language, 2016). This light-dark-ratio is important to maintain the animal’s welfare because it plays a crucial role in biological functions, such as hormonal secretions and the maintenance of homeostasis during the daily cycles of light and darkness (Hastings et al., 2003). As reviewed by Ungerfeld and Bielli (2012), seasonal variations in photoperiod length substantially influence reproductive function and behavior in mammals, even under artificial indoor housing conditions. Here we examine the known importance of photoperiodicity on the production, reproduction, behavior and welfare of domestic pigs, sheep and goats, cattle and horses.
2.1.1 Pigs
Photoperiodicity appears to play an important role for pigs, particularly in relation to food intake (Weiler et al., 1996), sexual maturity (Claus and Weiler, 1985; Andersson, 2000), and resting behavior (Baldwin and Start, 1985; Ni et al., 2017). In Europe, it is legally required to provide pigs under artificial housing with lighting (at least 40 lux) for at least 8 consecutive hours a day (Council of the European Union, 2008), although these precise recommendations are based on little scientific evidence. A study on nursing sows and their litters found that a photoperiod of 16h compared to less than 1h per day led to greater weight gain in the suckling piglets (Stevenson et al., 1983). This contrasts the findings of McGlone et al. (1988), who found that a change in the photoperiod from 1 to 16h did not affect weight gain in suckling piglets, but a photoperiod of only 1h combined with temperatures of around 30°C caused heat stress and a greater lactation weight loss in the sows (McGlone et al., 1988). Similarly, Zeng et al. (2023) revealed that an elongated daily photoperiod from 8h to 10h did not affect piglets’ growth parameters (i.e. average daily feed intake and weight gain), blood serum parameters (e.g., immunoglobin A or G), and excretion or drinking behavior. However, lying behavior in piglets was 15.48% higher when they were exposed to 10h light at 5-20 lux compared to piglets exposed only to 8h light per day at 5-20 lux (Zeng et al., 2023).
In growing pigs a photoperiod of 14h and 16h compared to 8h (even at relatively low illumination levels of 40 lux) improved growth parameters (i.e. daily weight gain), ham nutritional quality, and resting time, as well as reduced abnormal behaviors (Martelli et al., 2005, 2015). Fredriksen et al. (2006) found that a reduction of the photoperiod from 13h to 11h and 8h, under medium illumination levels of 60 lux, lead to an increase of aggressive behaviors from around 25% to 45%, respectively (in entire male pigs from weaning to slaughter). In contrast, an increase of the photoperiod from 14h to 18h at higher illuminances (440 lux) did neither reduce nor increase aggressive behaviors in those pigs (Fredriksen et al., 2006). Recently, Wallgren et al. (2024) reported that day length influences tail biting events in growing pigs. During months with longer light periods (April – September), pigs showed fewer tail injuries than during months with shorter light periods (October – March).
Even though the above-mentioned studies lack some degree of comparability as they used different illumination levels, the results indicate that pigs exposed to longer lighting periods attained a higher degree of welfare. Nevertheless, several studies showed that pigs prefer darkness for sleep (Baldwin and Start, 1985; Taylor et al., 2006; Götz et al., 2022).
2.1.2 Sheep and goats
Photoperiodicity is highly relevant for small ruminants of the temperate zone, as they are seasonal and short-day breeders (Chemineau et al., 1992; Thiéry et al., 2002; Duarte et al., 2010); thus, their reproductive behavior and their lactation performance are heavily influenced by daylight intensity and the light-dark-ratio. Therefore, in artificial husbandry environments, a change of the natural light-dark-ratio may affect the reproduction cycle of these animals. Despite this knowledge, there are no official husbandry regulations for the design of the photoperiod in indoor-housed goats and sheep.
In an experimental setup, Delgadillo et al. (2002, 2004) showed that it is possible to break the seasonal anestrus in goats under artificial light conditions by shortening the day length, i.e. these female goats can be mated with males exposed to artificially long days for 2.5 months for breeding against the natural day length. Studies from practice recommend a minimum of 16h light per day (with a light intensity of 200 lux at eye level) for a minimum of 45 days to extend the breeding season (Dalton, 2017). However, goats exposed to a long daily photoperiod (20h light) often showed pseudo- pregnancies (Garcia-Hernandez et al., 2007). Interestingly, exposure of bucks to a long photoperiod in winter can be beneficial to reestablish estrus cycles in an ovulatory doe (Rivas-Muñoz et al., 2010).
There are a few studies that focus on the relationship between photoperiod, physiology and performance. For both sheep (Pollott and Gootwine, 2004; Morrissey et al., 2008) and goats (Garcia-Hernandez et al., 2007; Véliz et al., 2009; Russo et al., 2013), an elongated photoperiod positively affects milk yield. While studies on sheep revealed varying optimal light periods, studies on goats showed that an elongation of the light period up to 16h or more resulted in an increase in milk yield up to a 20% (Véliz et al., 2009; Garcia-Hernandez et al., 2007), with the greatest response occurring in late-lactating animals (Russo et al., 2013). However, the increase in milk yield could also be attributed as a secondary effect of increased feed intake due to the extended photoperiod (Bocquier et al., 1990). Whether exposure to an artificial long-day photoperiod, with or without nutritional supplement, increases milk production in subtropical goats kept under extensive grazing conditions was investigated in a study by Flores et al. (2015). They found a higher mean milk yield in goats exposed to a long-day photoperiod (16 h) compared to goats exposed to a natural photoperiod (11 h). This difference in the mean milk yield was greater in goats that were given a nutritional supplement. Changes in the photoperiod has been shown to impact feed intake and ingestive behavior of sheep studied from October to April, in Belgium (Fischer et al., 1997). Light appears to be an important factor in regulating the feeding behavior of sheep (Schanbacher and Crouse, 1981). In sheep, it was found that a 12h photoperiod compared to 8h and 16h, respectively, increased daily dry matter intake and weight gain (Casamassima et al., 1990). A study on goat kids born in autumn in the subtropics found that artificial long days promote a high body weight during early growth phase (Flores et al., 2015, 2018).
In addition to milk yield, changes in milk content due to prolonged photoperiod have also been reported. Goats exposed to a long day photoperiod showed a decrease in milk fat, milk protein, milk solids, but a slightly higher milk lactose percentage, when compared to animals kept under normal photoperiod (Russo et al., 2013). Similarly, milk protein was found to be reduced when ewes were subjected to 15.5h of light compared to 8.5h (Bocquier et al., 1990) and a reduction in milk fat was reported for sheep under a 16:8 light-dark-cycle. All these results provide strong support for the use of alterations in the photoperiod as an effective, non-invasive method to enhance milk production in sheep and goats. Nevertheless, the potential effects of these long light periods on sheep and goats’ welfare have not yet been investigated.
In small ruminants, most, but not all, activity is concentrated in the light phase. Daily motor activity is lowest for goats during winter months (Piccione et al., 2008), although other factors associated with season should be considered. Rumination is also affected by the light-dark-cycle – when light periods dominate over the day, rumination is increased during light phases, while no such differences have been found when dark periods dominate the cycle (Oshiro et al., 1996). To our knowledge, no studies have yet been conducted on other behavioral changes, e.g., altered social behavior or increased aggression behavior as a result of the altered light-dark-cycle. Such studies are necessary before recommending changes in the housing systems of these animals.
2.1.3 Cattle
Cattle, like other bovines, are diurnal animals. Cattle probably distinguish both rate of change and actual photoperiod (Phillips, 2002). After Dahl et al. (2000) reviewed the photoperiodic effects on dairy cattle, Dahl and Petitclerc (2003) proposed an optimal photoperiod for adult dairy cows of 16 to 18h light per day. Several studies of the late 20th century already pointed out the positive effect of light on cow behaviors, reproduction [although cattle are not seasonal breeders; reviewed by Dahl et al. (2000)], milk production and hormone levels (Dahl et al., 2012). In two early studies, Peters et al. (1978, 1981) have demonstrated that supplementing cows with artificial light up to 16h of light (i.e. long day simulation) increases milk yield by around 7%. Similar results were also found in later studies (e.g., Bilodeau et al., 1989; Evans and Hacker, 1989; Miller et al., 1999; Reksen et al., 1999; Hjalmarsson et al., 2014; Lim et al., 2021), which seem to show the advantages of long light periods for the physiological performance of dairy cows. Similarly, Osborne et al. (2007) and de Almeida et al. (2015) found that supplementing 8 weeks old dairy calves up to 18 to 20h light (i.e. long-day photoperiods) stimulated their feed consumption and weight gain compared with calves with short-day photoperiods (i.e. 10 to 12h light). However, while long light periods probably have an indirect positive effect on performance (e.g., by acting on feed and water intake, rest and rumination), potential negative effects on animal welfare have not yet been investigated.
Studies on behavioral responses to changes in the light-dark-ratio revealed that the provision of supplementary light for up to 18h per day resulted in reduced agonistic behaviors in calves and further decreased their activity levels (Weiguo and Phillips, 1991). In the same study, it was also shown that calves with free choice to spend either time in an illuminated or a darkened area, spent more time in the illuminated area, which additionally led to an increase in their lying time. Similarly, supplementary light (mean intensity of 481 lux) reduced daily activity levels and increased lying time in dairy cows (Phillips and Schofield, 1989). More recently, it was shown that daily activity levels and lying time of dairy cows seem to be unaffected by a change in photoperiod (Andersson, 2012; Bernhart et al., 2018). However, it was shown that a reduction of the photoperiod from 18h and 16h to 13h led to a shift in the activity levels and lying behavior during the evening (activity peak was 3h earlier with 13h photoperiod) (Bernhart et al., 2018). In another study, it was shown that the provision of additional light during the night (11, 33 and 74 lux) did not affect dairy cows’ activity level during this period (Hjalmarsson et al., 2014). Nevertheless, milking frequencies increased when the provided light had an illuminance of 33 lux compared to 11 and 74 lux (Hjalmarsson et al., 2014). The findings of some of the studies discussed here suggest that manipulating the light-dark-ratio might influence cattle behavior to some extent. Overall, these insights contribute to our understanding of how light management can be a valuable factor in optimizing the welfare and productivity of dairy animals.
2.1.4 Horses
Under natural conditions, horses are diurnal animals. Therefore, it seems plausible that their higher activity levels occur during light periods (Berger et al., 1999). In domesticated horses, it has been shown that this diurnal pattern persists (Mayes and Duncan, 1986; Piccione et al., 2005; Bertolucci et al., 2008). For example, feeding behavior in horses is observed during light periods, throughout the day, but mostly directly after dawn and before dusk (Mayes and Duncan, 1986). In another study, it was suggested that the highest activity levels occur during springtime, when the natural photoperiod is around 13h per day, while the lowest activity patterns occurred during winter, when the photoperiod is less than 13h per day (Bertolucci et al., 2008). Surprisingly, in autumn when the photoperiod is comparable to that in spring, the activity levels remain more or less comparable. The authors suggested that an increase of the photoperiod in spring in combination with an increase in temperatures may be the cause of these increased activity levels (Bertolucci et al., 2008). Later in 2010, however, a study found that horses have endogenous circadian regulation of muscle function, which means that although horses are affected by external factors such as human activity, they are still influenced endogenously by their natural, internal 24-hours clock (Martin et al., 2010). As a result of this, horse training which follows the light hours of the day or even mimics the diurnal rhythm of horses, have been recommended (Murphy, 2019). It may even be possible to manipulate diurnal rhythms or seasonality in horses by use of, for instance, blue light, to stimulate estrus in anestrus mares (Murphy et al., 2014). Murphy et al. (2014) showed that the reproductive period of mares could be advanced using low-level blue light administered to just one eye from head-worn light mask and that this method was as effective as keeping mares indoors under artificial lighting. These studies collectively show that, although more research on the topic is needed, the photoperiod affects horses, and more research should focus on how circadian rhythms influence physiological and behavioral processes which should be considered when scheduling feeding, training, and resting times in order to optimize welfare.
2.1.5 Implications for housing and welfare
Housing designs and management routines for farmed ungulates should consider photoperiodicity to allow the animals to exhibit their natural daily rhythms for various activities and rest. Photoperiodicity and illumination intensity seem to be strongly inter-correlated and should therefore both be taken into account when studying effects on behavior, welfare and performance.
For pigs: Research on photoperiod is underrepresented in the literature. For suckling pigs, we only found two peer-reviewed papers, both from the 1980th. While in the paper of McGlone et al. (1988), a photoperiod of 16h did not positively affect piglets weight gain, the opposite was found by Stevenson et al. (1983). This emphasizes the need for more research on suckling piglets including farrowing sows. For weaned piglets and growing/finishing pigs, an elongated photoperiod seems to be more effective. Lying behavior increases already at a photoperiod of 10h, while aggressive behaviors decrease with a photoperiod of 13h. A photoperiod of more than 14h leads to greater weight gain and a greater ham nutrition rate (Martelli et al., 2005, 2015). Even though the studies used varying photoperiods, one can notice that a photoperiod of 8h as requested by the Council of the European Union (2008) does not seem satisfactory for pig’s growth parameters and behavioral performances. This emphasizes the need for a well-regulated light-dark-ratio, with at least more than 10h of light, to fulfill the pigs’ needs according to their age, and physiological as well as reproductive stage. However, more research under controlled conditions is necessary to draw conclusive assumptions on the impact of photoperiod on welfare and productivity metrics.
For sheep and goats: Seasonal breeding sheep and goats are short-day breeders. Photoperiodicity, whether natural or artificial, should therefore enable undisturbed seasonal reproductive behavior. Other than reproductive function and productivity, there seems to be a lack of scientific publications (and, consequently, of potential recommendations) on the impact of varying light-dark-cycles on the performance of specific behaviors such as socio-positive play and comfort behaviors that are explicitly relevant for their welfare.
For cattle: Cattle seem to be less affected in their daily activity patterns by variations in light regimen. Nevertheless, long light periods of up to 16 hours support milk production in dairy cows, while long-day photoperiod increases feed consumption and weight gain in dairy calves. It however remains unclear if artificially raising milk production causes concerns for the welfare of the cows, which should thus be a future research focus.
For horses: Housing facilities that allow daylight to enter the building and enable horses to follow their natural circadian rhythm would be beneficial. Likewise, horse training and stable activities should follow the natural light conditions to enable horses to synchronize their equine circadian rhythm to being active. Activities during dark hours should be minimized to safeguard horse welfare and optimize training outcomes, but the exact impact of equine activities during dark hours remains unexplored and should be a future focal point.
2.2 Illumination
Illumination is a measure of the incident light an animal (or object) is exposed to (American Heritage Dictionary of the English Language, 2016). Recent studies focused on illumination levels (also called illuminances or light intensities; indicated in lux) or on light color temperatures (indicated in kelvin; Table 1). The level of illuminance (lux) per se may not accurately reflect the actual light intensity perceived by the eye of an animal, as this depends on surface characteristics (such as floors) from which light is reflected before it enters the eye of an individual. As emphasized by Werner et al. (2018), measuring the actual light density (candela per m2) seems to be essential for contrast vision assessment in farm buildings, which is relevant for undisturbed movement, and to distinguish installations such as feeders and barriers. In this section, we review the recommendations and knowledge of the effect of illumination on indoor-housed farmed ungulates.

Table 1. Relevant lighting and vision parameters, definitions and units [adapted from Werner et al. (2018)].
2.2.1 Pigs
As mentioned above, it is recommended that pigs under artificial light receive at least 40 lux (Council of the European Union, 2008; Hirt et al., 2020) and 5 to 20 foot-candles (i.e. 50 to 215 lux) in the USA (National Pork Board, 2018) over an 8h light period per day. Illumination levels (intensity) can affect pigs and influence their body condition and behavior. For example, miniature pigs that were kept continuously under 2500 lux for over 1 month developed retinal damage and showed reduced pupillary reflex (Dureau et al., 1996). Similarly, it was found recently that pigs kept under high light intensities of 968 lux developed conjunctivitis more often than pigs kept in pens with illumination gradients ranging from 71 to 330 lux (Scaillierez et al., 2024). However, variations in light intensity do not affect piglets’ survival and development per se (Mutton, 1987; Zeng et al., 2023).
Zeng et al. (2023) investigated whether different light intensities lead to behavioral changes, and found that abnormal behaviors (e.g., fighting, biting railings, and tail biting) in weaned piglets were twice as high when they were kept under a light intensity of 5 - 20 lux compared to piglets kept under 40 - 50 lux. However, it must be mentioned that the total rate of abnormal behaviors in the group kept under 5 - 20 lux did not exceed 7% (Zeng et al., 2023). A higher illumination rate (40 - 50 lux compared to 5 - 20 lux) also increased resting time in piglets; at least when they were exposed to 8h light per day (Zeng et al., 2023). For growing pigs, it was found that a light intensity of 80 lux reduced aggressive behaviors in comparison to pigs kept under 40 lux (Martelli et al., 2010). In another study, medium levels of illumination (60 lux) provided exclusively as artificial light, resulted in high percentages of aggressive behaviors in entire male pigs (observed from weaning to slaughter) when compared to illuminations of 440 lux (Fredriksen et al., 2006). This result has to be interpreted with caution as the 440 lux were provided by a combination of daylight through the windows and artificial light. Simulation of intense outdoor light conditions of 600 lux in a controlled laboratory setup for growing pigs indicated that a daylight intensity of 600 lux affected behavioral activities (indirectly measured) that are associated with increased skin lesion scores (Zheng et al., 2015). In contrast, independently from light intensity, body core temperature pattern followed the same biphasic diurnal activity rhythm as seen in other behavioral studies with pigs (Zheng et al., 2015). A new study by Scaillierez et al. (2024) found that growing pigs of around 14 weeks of age displayed more negative social interactions when exposed to either low (45 lux) or high (968 lux) light intensities compared to medium (198 lux) light intensities. Light intensities impinged abnormal behaviors only at pigs’ age of around 18 weeks, with fewer abnormal behaviors in the groups kept under 198 lux compared to 45 lux (Scaillierez et al., 2024). Abnormal behaviors were also less frequent in pigs kept in a pen with light gradients (71 – 330 lux). When pigs were around 20 weeks of age, they showed more exploratory behavior when exposed to light intensities of 198 lux compared to pigs that were kept under 45 lux (Scaillierez et al., 2024). The results of Scaillierez et al. (2024) suggest that light intensities influence behavior differently at different ages, but there was no consistency between the observed behaviors and the illumination level. Hence, it remains difficult to make detailed recommendations about adequate lighting of indoor housing systems.
Glatz (2001) reported on sex differences in aggressive behavior of piglets, kept under similar light conditions before and after weaning, due to variations in light color temperatures. While male piglets showed more incidences of fighting under TP/PR light (i.e. Triphosphor (TP) lighting to simulate daylight during the day and Pascal red (PR) lighting during the night) compared to conventional cool-white light during the day only, the opposite was observed in female piglets (Glatz, 2001). Independent of sex, the provision of additional PR light during the night led to poorer body integument conditions (ear, neck and genitals were the body areas that were affected most) in the piglets (Glatz, 2001). It was, however, also shown that the TP/PR illuminated piglets increased their feed intake during the first experimental week. Hence, the study showed that the TP/PR light provision had some advantages but also that the red-light color temperature was disadvantageous in rearing barns. Another study analyzed tail injuries in growing pigs kept either in pens with standard fluorescent tubes with an invisible flickering light of 30 – 40% for 14h daily or non-flickering LED light for 14h daily (Wallgren et al., 2024). The authors found significantly more tail injuries in the first mentioned group (Wallgren et al., 2024). Hence, additional research in this area needs to be conducted to investigate the optimal light color temperatures, which also relate to the respective ontogenetic developmental stages of pigs.
In addition to studies with fixed illumination levels, several studies investigated pigs’ preferences for different illuminances but came to conflicting results. Tanida et al. (1996) found that suckling piglets of 1 week of age actively moved toward bright areas and away from dark areas, while Baldwin and Start (1985) and Andersen et al. (2000) found that weaned pigs preferred dimmed lighting (10 lux or 2.5 lux) compared to bright lighting (110 lux or 90 lux) or complete darkness. In contrast, Taylor et al. (2006) found that weaned piglets only preferred a dim illumination (around 2.4 lux or less) for resting: they spent 78.7% of their resting time in those compartments with the lowest illumination. Nevertheless, the animals used all the provided compartments during this experiment (4, 40 and 400 lux) and used compartments illuminated with 40 and 400 lux significantly more for defecation, while other behaviors such as drinking, eating and playing were not influenced by the illuminance. The fact that, for example, eating was independent of illumination levels concur with other studies which found that foraging in pigs is mainly driven by olfactory and tactile senses (e.g., Nowicki and Klocek, 2012). Götz et al. (2022) investigated the preference of weaned piglets for compartments illuminated with 600 lux vs. dark compartments. They found that illumination preferences change with age and partly depend on the performed behavior of the piglets. At an age of 4 weeks, piglets clearly preferred the lighted compartment for all behaviors. This agrees with findings of Tanida et al. (1996). However, in the Götz et al. (2022) study, when piglets reached 5 weeks of age, they preferred to stay mostly in the dark compartments except during midday where pigs increased the time spent in the lighted compartments eating or being active (Götz et al., 2022). Both the studies of Taylor et al. (2006) and Götz et al. (2022) indicate that pigs should have access to darkened areas, which can be provided either temporally or spatially. A spatial provision might be more attractive for pigs because this gives them the possibility to freely choose between compartments according to the performed behavior, e.g., leaving the darkened compartment for defecation. Such a spatial separation would also be beneficial for the farmers because it would simplify the cleaning of the barn.
In addition to preferences for illumination levels, pigs seem to show preferences for specific light color temperatures. A recent study revealed that weaned piglets first preferred a color temperature of 3000 K at 4 weeks of age just after weaning, before this preference decreased over time (Götz et al., 2020). Piglets of 6 weeks of age preferred the 3000 K color temperature for resting only while they preferred the 6500 K color temperature for defecation. By the age of 8 weeks, they additionally preferred 6500 K during feeding (Götz et al., 2020). This study therefore shows that pigs are able to discriminate between light color temperatures of 3000 K and 6500 K, and that their preferences for specific color temperatures seem to depend on their age and performed behaviors.
2.2.2 Sheep and goats
Recommendations for specific illuminances for sheep and goat housing are rare. For example, Berge (1997) recommends an illumination level of 75 to 100 lux in sheep barns. Casamassima et al. (1993) found that a light intensity of 100 lux could lead to an increase in the time sheep spent standing, which could be related to the sheep being less relaxed. Very low light intensities of 10 lux were shown to reduce the time spent eating and increase abnormal behaviors. Meanwhile, light intensities of 1000 lux resulted in hyperactive behaviors in lambs (Casamassima et al., 1993). In sheep (and pigs), elimination of shadows seems to also facilitate movement (Grandin, 1979). A German magazine for sheep and goat husbandry recommends providing a window area of 3% of the barn floor area to allow natural light to enter (Mösenbacher-Molterer, 2018). According to the legal requirements in Germany, the minimum light intensity in the animal area is 40 lux. However, for optimum utilization of productivity, the authors recommend raising this value to 200 lux (Mösenbacher-Molterer, 2018). This is in line with the illumination of 220 lux in sheep and goat husbandry systems recommended by the MidWest Plan Service [MidWest Plan Service (1994) cited by Thonney et al. (2020)].
There is a lack of studies on illumination preferences, making it challenging to draw conclusions about the optimal illumination levels for goats or sheep in husbandry environments. In one of the very few studies assessing illumination preferences in small ruminants, sheep were placed in a dark chamber and had the possibility to turn on the lights by interrupting an infrared beam. The results showed that the sheep had a strong motivation to press the beam switch with their muzzles rewarding them with light, which can be interpreted as a preference for light over darkness (Baldwin and Start, 1981). To our knowledge, no studies on perception, behavior or preferences related to light color temperatures and its effect on physiological parameters or welfare have been conducted yet for these species.
2.2.3 Cattle
The visual system of cattle is sensitive to motion and contrasts of light and dark (Grandin, 2007). Calves are able to discriminate different light intensities but to a lesser extent than humans (Phillips and Weiguo, 1991). German regulations have, for instance, specified that calves must be kept at a light intensity of at least 80 lux (TierSchNutztV14, 2021). However, studies on illumination within cattle barns are underrepresented in the scientific literature.
Phillips and Schofield (1989) investigated whether 10h supplementary light with intensities of 0, 101, 191, or 529 lux, provided in addition to a 9h natural daylight, influenced physiological performance of lactating cows. They found that both a light intensity of 101 or 529 lux led to a decrease in milk fat concentration, while feed intake and milk yield were unaffected by light intensities (Phillips and Schofield, 1989). In contrast, Lim et al. (2021) observed an increase in milk fat concentration but not in milk yield in dairy cows illuminated during 16h light with 100 lux vs. a lower or higher light intensity (i.e. 50 and 200 lux). A comparison of LED illumination (short-wave light) and sodium-vapor illumination (long-wave light) on the behavior of dairy cows revealed that the first treatment had several advantages (Bernhart et al., 2018). Cows that were exposed to LED light increased their daily lying time. The lying time again increased with an illuminance of 80 lux, which was especially visible during noon, while activity levels increased when an illuminance of 150 lux was used (Bernhart et al., 2018). Dannenmann et al. (1985) examined the influence of light intensity (8 h/day of artificial light at 2, 20, 100 vs. 130 lux) on the behavior of calves and found that the higher the intensity, the more feeding and social behaviors occurred, while the reverse happened for resting behavior. Lindkvist et al. (2023) later observed that light uniformity may affect locomotion in cows. In particular, medium intensity, non-uniform red light (i.e., an average of 4 lux at a uniformity of 0.1 µmol.m-2.s-1 and a wavelength around 650 nm) affected the speed of pregnant non-lactating cows when navigating through an obstacle course. Also, in loose-housing systems, darkness did not seem to affect cows’ locomotion (Lindkvist et al., 2023).
Recent practical studies with LED bulbs in dairy barns indicate that light color temperatures adapted for specific areas such as the milking parlor (5000 to 6000 Kelvin) may enhance visibility and motivation to visit these specific areas (Werner et al., 2018). Practical recommendations for dairy cows refer to an illumination level of at least 150 lux for all areas of a barn during daylight and less than 10 lux during nighttime. Sudden changes of illumination levels within the barn should be avoided, due to the slow adaptation of the cattle pupil to different light conditions (Ryba and Uhl, 1989). In cattle, the light-dark adaptation and vice versa takes about 4 - 5 times longer than in humans (Bayerische Landesanstalt für Landwirtschaft, 2012). Thus, moving cattle into sudden bright light or dark areas, for example from the barn to the milking parlor and back, could lead to fearful reactions.
2.2.4 Horses
Specific illuminance recommendations for horse housing are rare. Most recommendations on illumination with regard to horses, are recommendations for lighting of riding arenas (100-150 lux for riding) and during sporting events (250-300 lux at the eye-level of the horse) (Svenska Ridsportförbundet, 2020). Some recommendations depend of the number of people rather than the number of horses at an event, with increasing people yielding higher lux (e.g., > 1000 people > 750 lux horizontal) (Riksidrottsförbundet, 2023). In Sweden, there are a set of recommendations for lighting of horse facilities with 100 lux as the general recommendation for indoor horse facilities (Brandskyddsföreningens Service AB, 2023). At the international level, the FEI (i.e., the international federation for equestrian sports) recommends stable and riding conditions with good lighting as a minimum requirement for sporting events that includes horses (Fédération Equestre Internationale, 2020). Overall, bright light, large light variations or uneven light as well as sudden changes in light intensity should be avoided (Svenska Ridsportförbundet, 2020).
Horse eyes are large and contain elongated pupils which can dilate greatly to capture sparse photons at night (Wouters and De Moor, 1979). At the back of the horse’s eyes, the reflecting tapetum lucidum provides a higher chance of the photons to be captured by the photoreceptors (Ollivier et al., 2004). All these features give the horse good visual abilities under low light intensities. Studies on equine scotopic abilities have shown that horses maintain grazing and interacting with conspecifics during nighttime, i.e., under light intensities similar to moon light (Berger, 1986; Mayes and Duncan, 1986). Saslow (1999) further noted that horses are more capable of seeing details on overcast days as opposed to sunny days. Some of the more recent studies show that horses are able to solve two-dimensional discrimination tasks in almost complete darkness, and that horses possess good visual capacity under both natural and artificial light conditions (Hanggi, 2006; Hanggi and Ingersoll, 2009). However, a crucial difference between horses and humans is the ability to adapt to sudden changes in light intensity. While humans transfer from bright light to darker conditions (or vice versa) within a few seconds, for horses the transfer may take considerably longer. The size of the horse’s horizontal pupil is reduced by relatively weak circular muscles (Samuelson, 1999), making it slow to adapt. Horses may thus be more easily blinded by exposure to sudden bright light, in comparison with humans. In the reverse situation (transition from bright light to darker conditions) the adaptation time depends on the initial light intensity to which the horse was exposed. The higher the light intensity, the longer the adaptation period, which can extend for up to 45 min (Wouters and De Moor, 1979; Hanggi and Ingersoll, 2009).
Moreover, Neveux et al. (2023) investigated the effects of different LED lighting conditions on young horses during trailer loading and stationary confinement. They found that homogeneous warm (3000 K and 100% luminous flux) and cold (6500 K) LED light color temperatures during the stationary phase seem to have a calming effect when horses return to their stable, which was not the case under natural light conditions. Additionally, horses loaded faster into the trailer and expressed less stress-related behaviors with a high illuminance level (> 4500 lux) inside the trailer compared to a lower illuminance level (< 3000 lux) (Neveux et al., 2023).
2.2.5 Implications for housing and welfare
Illumination is essential for biological functions, health and visual capabilities of farmed ungulates to recognize and discriminate between conspecifics as well as for technical structures in their housing environment. However, evidence for clear recommendations on essential illumination intensities is still scarce. In comparison to humans, ungulates are better adapted to nocturnal and crepuscular conditions. Adapting light conditions and management can ease handling of farm animals and lower stress. Therefore, not only illumination intensity, but also light color temperature and luminance density for contrast vision assessment should be considered for future housing and welfare improvement.
For pigs: Pigs are able to see at very low illumination intensities of less than 1 lux. Most of the current recommendations are aimed at much higher intensities for orientation (which is more important for human supervision) and daylight conditions of a minimum of 40 to 80 lux for which pigs do not show clear preferences. However, some studies also revealed that pigs kept under illumination levels lower than 40 lux increase their aggressive or abnormal behaviors. On the other hand, pigs seem to avoid very intense illumination and exhibit increased aggression when permanently exposed to these conditions (> 440 lux). This wide range of illumination levels (between 40 and 440 lux) makes it difficult to propose any recommendations for future light settings. Furthermore, most of the literature available was conducted on growing/finishing pigs, less on piglets and none on farrowing sows. This leads to a gap of knowledge regarding whether illumination levels can affect growth parameters and agonistic behaviors across production stages.
If pigs seem to be able to discriminate and develop preferences for certain illumination levels and light color temperatures, these preferences seem to depend on their age and performed behaviors (e.g., piglets of 4 weeks of age preferred dimmed illuminations and warm light colors for all behaviors; piglets of more than 5 weeks of age preferred dimmed illuminations and warm light colors only for resting) and should thus be considered for illumination devices in pig barns. Habits certainly also play a part. As pigs clearly establish functional areas for resting and other activities, illumination intensity and quality could help among other climatic factors to support this motivation for separation of functional areas. Future housing should not only provide them with access to natural light conditions but also allow them to choose between dark/dimmed and illuminated areas to better support their behavioral and functional needs such as resting, exploration, feeding, defecation and social activities at different times of the day. As very intense outdoor illumination levels might be harmful (including sunburn in unpigmented domestic pigs), shades or dimmed areas for resting seem to be essential for their welfare.
For sheep and goats: Studies on illumination requirements for sheep and goats are scarce. One can assume that most small ruminants are housed outdoors and well adapted to these conditions or exposed to natural light conditions when housed indoors, explicitly during the winter periods. Few studies indicate that indoor illumination intensities between 10 and 100 lux seem to be appropriate for sheep while others recommend light intensity around 200 lux for optimal performance. Future studies should systematically investigate if low or medium illumination levels in indoor housing systems can promote sheep and goats’ welfare, and whether they prefer specific light intensities to perform behaviors such as resting, exploration and feeding.
For cattle: Although illumination seems to influence the behavior of cattle, studies on light intensity preference have not been reported so far. Most studies in cattle are concerned with the effect of additional (LED lighting) illuminance (in combination of an increased photoperiod) and/or light color temperatures on behavior, performance and milk yield in dairy cows. Contradictory results were reported with regard to the effect of light intensity on milk fat concentration. Lower illumination levels seem to increase lying time, both in calves and lactating dairy cows, which could positively affect rumination, while illumination levels up to 130 lux seem to increase feeding and social behaviors in calves. Furthermore, it seems that adapted LED light color temperatures for specific crucial areas in the barn (at 150 lux) may enhance visibility and thus increase the motivation to visit these areas. Based on the available literature, it remains difficult to draw definite conclusions on the effect of illumination on cows’ behavior and welfare.
For horses: As for cattle, horses seem to be influenced by illumination, but preferences for distinct illumination levels have not been systematically investigated. Training horses in low light conditions might make it easy for the horse to see its surroundings, but in turn makes it harder for the rider or trainer. Illumination of the area in which a horse is, regardless of the purpose (training/housing/riding) is thus important but the horse’s slow adaptation from light to dark is possibly the most important factor to consider. This may explain why horses are sometimes reluctant to go from lighted areas into dark ones, for instance, the interior of a trailer might be visible to humans in bright daylight, but to a horse, it can appear to be a dark hole. As a result, constant light intensity is preferable for horses to minimize sudden or large changes in illumination.
2.3 Color
The perception of colors is linked to the presence of cones in the eyes and varies among species. For an individual to perceive color, the retina must contain at least two types of photoreceptors, each with peak sensitivity in a different wavelength of light. These receptors are sometimes referred to as red, green, and blue cones, sensitive respectively to long-wavelength light (L), middle-to-long-wavelength light (M/L), and short-wavelength light (S). To date, it appears that horses, pigs, goats, cows, and sheep are lacking red (L) cones and therefore have the photopigment basis for dichromatic color vision (Neitz and Jacobs, 1989; Jacobs et al., 1998). For dichromats, there is a neutral point in the wavelength spectrum so that light may appear as gray (Roth et al., 2007).
It has been shown that domesticated ungulates respond differently to various light color temperatures (see section 2.2). These species are also able to discriminate a variety of colored objects (e.g., Manida et al., 1989; Tanaka et al., 1989a; Tanida et al., 1991). Here, we will review to which extend farmed ungulates discriminate colors. In humans, it has been shown that colors can affect mood (O’Connor, 2011; Elliot, 2015; Yang and Shen, 2022), however, little is known about the effect of color on domesticated ungulates and whether colors and colored light (indicated in nm) can affect physiological parameters, e.g., reduce stress levels.
2.3.1 Pigs
There are various studies on pigs’ color perception that affirm that pigs are able to discriminate colors. Tanida et al. (1991) pointed out that pigs could discriminate only blue among three colors (i.e. vs. green and red) as well as distinguish blue from gray. However, these results must be viewed with caution because the stimuli (i.e. cards) presented to the pigs did not match in brightness, but only in luminosity, and therefore the results might be biased (Tanida et al., 1991). More recent studies indicate that piglets can differentiate between blue, yellow, red, and green, even though pigs are assumed to have dichromatic color vision, i.e. be red-green deficient (Deligeorgis et al., 2006; Klocek et al., 2016). More precisely, these studies showed that piglets prefer to feed and drink from blue troughs (Klocek et al., 2016) and red drinkers (Deligeorgis et al., 2006), respectively. In contrast, the piglets used yellow troughs (Klocek et al., 2016) and green drinkers (Deligeorgis et al., 2006) less frequently. Deligeorgis et al. (2006), but not Klocek et al. (2016), found that the preferences slightly vary with sex; female piglets preferred blue drinkers while males more often used red ones. Foppa et al. (2018) on the other hand, found that female pigs had no preference for environmental enrichment objects of a specific color (red, blue, yellow). In a study by Babicz and Paprocka (2018), it was investigated whether two different pig breeds (Pulawska and Polish Large White) show preferences to interact with objects of certain colors (yellow, blue, red) in a “free-choice behavioral-test”. Surprisingly, the authors found that the Pulawska and Polish Large White preferred blue and yellow objects, respectively, and both breeds interacted least with the red objects. Besides the different preferences, the results clearly show that these animals can distinguish between colors, and that they prefer blue and yellow over red (Babicz and Paprocka, 2018). However, the results on the Polish Large White breed contradict the findings of Klocek et al. (2016), where blue and red were preferred over yellow. One possible explanation for the dissenting preferences might be due to genetic variation between the breeds; Klocek et al. (2016) used Polish Landrace x Polish Large White crossbred piglets. Another possibility could be simply due to differences in their familiar environment before the trial. However, at this stage, it remains only speculative and more detailed investigations should be done focusing on color preferences among various pig breeds.
A recent study of Paggi et al. (2020) investigated suckling piglets’ preference of alternative light colors (i.e. blue, green, yellow and red) in their creep area over LED white light, and whether it affected their behavior. Piglets showed significant preferences only for white light over blue and red light and preferred green lighting among the various colors, even though no color influenced their behavior. No other studies investigating pig preferences for specific light colors were found.
2.3.2 Sheep and goats
Understanding color vision in sheep and goats is crucial for better understanding their behavior in various environmental and housing contexts (Tribe and Gordon, 1949). However, there is still some debate about the extent of color vision in both species. Jacobs et al. (1998) used flicker photometry to assess the spectral properties of cones in sheep and goats and concluded that these animals have the necessary photopigments for dichromatic color vision, similar to most other mammals (Bowmaker, 1998). Studies on color discrimination in sheep have shown that lambs can differentiate between red, green, blue, and gray (Tanaka et al., 1989a) and between various shades of green (Bazely, 1988; Tanaka et al., 1989b). Bazely and Ensor (1989) further showed that sheep could not differentiate green and yellow of the same brightness. Goats, on the other hand, have been found to distinguish between orange, yellow, green, blue, and violet (Buchenauer and Fritsch, 1980), with some preliminary evidence indicating trichromacy (Siemers et al., 1999), whereas Jacobs et al. (1998) reported only two types of cones in goats - the basis for dichromatic color vision.
Moreover, Tribe and Gordon (1949) have even postulated that color vision in sheep has no importance for grazing and feed acceptance. There are reports that sheep become irritated or fearful when exposed to novel colors associated with painted conspecifics or offspring (Alexander and Shillito, 1977), although this response was also seen in response to increased darkness of gray (Alexander and Stevens, 1979). In addition, a study on light colors found that green light (560 nm) can increase feeding time and reduce resting time in lambs compared to white (solar spectrum), red (680 nm), and blue (470 nm) lights (Casamassima et al., 1994).
To the best of our knowledge, further studies on color perception abilities and preferences of goats and sheep are not available and it remains subject of future research if different colors may affect physiological parameters and welfare.
2.3.3 Cattle
According to Jacobs et al. (1998), cattle have dichromatic vision with peak photopic sensitivities occurring at 455 nm (blue) and 554 nm (green). This means that cattle, like all other ungulates, see colors (Riol et al., 1989; Phillips and Lomas, 2001). Discrimination experiments suggest that cattle can distinguish short (492 nm blue) and medium (538 nm green) wavelengths from long (610 nm red) wavelengths (Gilbert and Arave, 1986; Riol et al., 1989; Phillips and Lomas, 2001), but the discrimination between blue and green light seems to be difficult (Gilbert and Arave, 1986). However, nothing shows that the intensity of the light sources has been taken into account during these tests. Two recent studies investigated whether Japanese Black cows were able to differentiate between green fresh forages and brown dead forages, and assessed whether this ability was affected by sunlight versus shadow. The results indicated that the cows successfully differentiated between forage types independently of ambient light conditions (Hirata et al., 2019; Hirata and Kusatake, 2020). Manida et al. (1989) conducted an experiment to study the ability of cattle to discriminate color in a grazing management context. They demonstrated that cattle were most willing to stay inside the paddock and therefore away from the electric fence when the tape had the color red, followed by blue, white, orange, yellow, green and violet (Manida et al., 1989). In another study, it was shown that the presence of green light seems to reduce fear and improved maze performances of calves compared to their performance under blue or red light (Lomas, 1994). This positive effect of green light was also seen in calves that were exposed to a fearful stimulus, i.e. a loud noise behind them (Phillips and Lomas, 2001). The result of Lomas (1994) and Phillips and Lomas (2001) indicates that cattle can distinguish between green and blue light, which contrasts with the findings of Gilbert and Arave (1986).
Lindkvist et al. (2021) studied the effects of achromatic and chromatic (i.e. artificial blue, red, or white similar to sunlight) lights on endocrinology, activity, and milk production in dairy cows. They observed that milk yield was not affected whereas plasma melatonin was higher during the night, when the cows were exposed to blue and red dim light compared to white light (Lindkvist et al., 2021). Murphy et al. (2021) observed a melatonin secretion suppression but no milk production effect in dairy cows after administrating a LED short wavelength blue light (468 nm) at high intensity (250 lux) to one eye. Contrasting to these studies, Son et al. (2020) found that supplementary LED blue (470 nm) and yellow (580 nm) light during the night led to a decrease in milk yield in lactating cows compared to white or no light during the night. In addition, blue light supplementation led to higher stress levels in those cows (Son et al., 2020). Elsabagh et al. (2020) also found higher melatonin levels during nighttime in female dairy calves with yellow vs. blue LED (for 2h before the darkness of long-day photoperiod) which might negatively impact their development. They additionally observed higher hay intake, frequency of water intake, rumination time and body weight gain when exposed to the yellow rather than the white LED (Elsabagh et al., 2020).
Lindkvist et al. (2021), also analyzed the relationship between light colors and activity patterns and found that standing during the night was slightly higher (but not significant) when blue light was provided compared to white light. They argued that in a tie-stall system (as used in their experiment), different light regimes did not change activity levels of dairy cows. Whether the same may be true for loose housing systems need to be further investigated. Wilson et al. (2022) found no difference in lactating cows’ resting behavior when briefly exposed to supplemental LED wavelengths [full-spectrum white (380 – 780 nm), yellow-green (564 nm), and blue (483 nm)] in the resting area compared to a dark control treatment.
Generally, literature on cattle color vision abilities, behavioral responses and preferences is scarce and not always controlled for other parameters. Further studies to fulfill this knowledge gap are needed in order to elucidate if color affects cattle welfare, and if so, how color could be used to optimize cattle housing and management on-farm.
2.3.4 Horses
Horses also have dichromatic color vision (Macuda and Timney, 1999; Carroll et al., 2001; Timney and Macuda, 2001). It means that horses have only two distinct spectral receptors in their retina: blue (S) and green (M/L) cones. Because horses are lacking red (L) cones, they likely see colors similarly to red-green colorblind humans (i.e., protanopia) (Rørvang et al., 2020). With only two color receptors (Timney and Macuda, 2001), several combinations of wavelength and luminance will induce equal response ratios in the receptors, so that various colors will appear similar to one another. For horses, as well as for other dichromats, there is a neutral point in the wavelength spectrum so that light may appear as gray (Roth et al., 2007). Although horses probably perceive wavelengths from across the spectrum as colored (apart from that at the neutral point), these colors may be seen as variations of only two hues (blue and green). This means that horses would be unable to use wavelength information to discriminate between colors that lie on the same side of the neutral point similar to the way human protanopes commonly confuse reds and greens (Murphy et al., 2009). Different dichromats may additionally differ in their color vision abilities depending on the spectral tuning of their cone pigments (Carroll et al., 2001) resulting in different achromatic (i.e., grayscale) zones across different dichromatic species.
In behavioral studies, horses have been shown capable of discriminating blue and red from gray but had difficulties with green and yellow (Pick et al., 1994; Macuda and Timney, 1999). Grzimek (1952) pointed out that horses can discriminate yellow best from gray, followed by green and blue, while they had difficulties discriminating red. Similarly, Hall et al. (2006) found that the colors orange, yellow, and blue can be clearly discriminated from gray, but not from red. However, in the same study, it was also found that horses had difficulties to identify yellow and blue when compared to green (Hall et al., 2006). Smith and Goldman (1999), on the other hand, found that horses can discriminate red, yellow, green as well as blue from gray, and that only single individuals may have difficulties identifying green and yellow.
The ability of horses to distinguish among various colors was also tested in relation to flooring by Hall and Cassaday (2006). The results reveal that white-, blue-, and yellow-colored mats on black floors caused a great number of adverse reactions, while green, red, brown, and gray caused less adverse reactions (Hall and Cassaday, 2006). The level of the adverse reactions diminished in most cases once no negative consequences had occurred, i.e. after acclimatization (Hall and Cassaday, 2006). A study by Burla et al. (2016) further found a general calming effect (on activity and heart rate) of color illumination (15 min at 240 lux), regardless of the (625 nm red, 571 nm yellow, 534 nm green, and 470 nm blue) color of the light.
The results of the above-mentioned studies reveal that there is a lack of clarity about which colors horses can discriminate, and which factors influence this ability. It is well-known that certain breeds and colors of horses can have implications for hearing ability (Magdesian et al., 2009) and visual ability (Andersson et al., 2013). Hence, it is possible that such factors also influence color vision abilities and preferences of individual horses. However, instead of studies that focus on simple discrimination tasks, studies investigating how horses may react to different colors in different situations may be of greater interest. Such studies are needed in order to evaluate if colors might benefit horse welfare if used in stable design and horse management.
2.3.5 Implications for housing and welfare
Although pigs, sheep and goats, cattle and horses have dichromatic color vision, evidence suggests that they do not all see nor discriminate colors the same way, having potentially different achromatic (grayscale) zones. In addition, further highly controlled studies are needed to identify to which extent ruminants and horses (in particular) discriminate colors and whether this ability is relevant for housing enrichment and handling. Research should, for example, focus on physiological and behavioral reactions to colored features and colored lights.
For pigs: Pigs seem to be able to discriminate between colors but the studies on preferences for any particular color associated with feeders, troughs etc. give no clear results. Compared to humans and birds one can assume that they do not see red as humans see it, as they have problems discriminating red from gray. Their preference for resources seems to be more dependent on their excellent senses of smell, sound and touch rather than vision. Therefore, environmental enrichment with coloration to enhance visibility or colored materials to initiate exploration such as colored “toys” may not fulfill their purpose in pigs.
For sheep and goats: As for all farmed ungulates, sheep and goats’ visibility and discrimination of objects, conspecifics, handlers and predators do not seem to depend much on their sense of color vision. One can assume that they are, similar to other species, able to discriminate and develop preferences between color intensities, such as pastures with dark or bright green grass. However, we do not know yet if it is slightly different wavelengths that makes them choose one pasture or another.
For cattle: Literature on cattle color vision and discrimination abilities is scarce. If cattle seem to be able to see and discriminate between short (blue) to medium (green) wavelengths and long (red) wavelengths, further studies are needed to better identify to which extent cattle rely on gray nuances to distinguish between colors. There is evidence that green light has a calming effect on calves (Lomas, 1994). Whether this effect remains in later life stages needs to be investigated. Nevertheless, the use of green light to improve the environment for cattle in specific situations, e.g., during transportation or in slaughterhouses, where the animals may experience fearful and stressful situations, may contribute positively to their welfare. Studies on performance rates of dairy cows and calves, i.e. daily milk yield, melatonin levels and body weight gain, come to conflicting results, and therefore need further investigations. Further studies should focus on fulfilling this knowledge gap before conducting research on color effect and colored features on cattle welfare.
For horses: Although it is still not clear exactly how horses see colors, current knowledge suggests that some colors are visible to horses whereas others are not. Further research on horses’ behavioral reactions to colored features within their home environment or colored light is needed to fully map horses’ reaction to colors and their impact on horse welfare. It is, however, clear that colors impact horses especially when the horse can see contrasts, such as a yellow, white or blue mat on a dark surface. Such information is useful in the design of high welfare housing systems but also in the daily management and training of horses. In some situation it may be beneficial to have clear contrasts in order for the horse to see the object (e.g., a jump in horse riding), whereas in other situations using colors with less contrast would be beneficial in order to lower adverse reactions (e.g., flooring, inside of a trailer or features in the home pen).
3 Conclusions
In conclusion, the welfare and productivity of farmed ungulates can be influenced by photoperiodicity, lighting, and color perception. For pigs, longer photoperiods (≥10 hours) improve growth rates and reduce aggression, although further research is needed to establish optimal lighting conditions, especially for younger pigs. Sheep and goats, as seasonal breeders, require photoperiods that align with their natural reproductive cycles, with potential benefits in milk production, though the full impact on welfare remains unclear. Cattle benefit from extended light periods expressed in increased milk yield and feed intake, but more research is needed to understand the long-term welfare implications of such lighting practices. Horses, with their circadian rhythms closely tied to their behavior and performance, benefit from routines aligned with natural light cycles, suggesting that minimizing nighttime activities can enhance welfare and training outcomes.
In terms of lighting, species-specific needs are crucial for safeguarding welfare and production, with pigs needing moderate light levels to avoid aggression and stress, and cattle benefitting from tailored lighting systems that support feeding and social behaviors. Sheep and goats also require adequate illumination, with studies suggesting lighting between 75-100 lux to minimize stress. Horses, with unique visual needs, require gradual transitions in light intensity, particularly for tasks like trailer loading, highlighting the importance of lighting systems that reduce stress and support natural behaviors across species.
Colored light, while still an underexplored area of research, appears to influence the behavior and welfare of farmed ungulates. Pigs can distinguish between colors, but their preferences are often driven by other more salient sensory cues such as olfaction and touch than by color. Sheep and goats can differentiate between some colors, though the significance of this in their behavior remains uncertain. Cattle show a preference for green light, which may have a calming effect, and horses can distinguish blue and yellow, with varying reactions to color depending on the context. Although these findings suggest that color can affect behavior and thus welfare, more research is needed to fully understand how colors influence physiological responses and how they may be utilized.
Ultimately, a central aspect to enhancing the welfare and productivity of ungulates lies in understanding and tailoring environmental factors such as different light parameters to meet the species-specific needs of pigs, sheep, goats, cattle, and horses. Future research should continue to explore these areas to optimize housing systems and management practices, ensuring the well-being of farmed ungulates.
Author contributions
JS: Conceptualization, Project administration, Writing – original draft, Writing – review & editing. EvB: Conceptualization, Writing – original draft, Writing – review & editing. JL: Writing – original draft, Writing – review & editing. CN: Writing – original draft, Writing – review & editing. MR: Visualization, Writing – original draft, Writing – review & editing. CR: Conceptualization, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
We thank the Martin-Luther-University Halle-Wittenberg (Germany) for financial support for publishing this manuscript with an Open Access license.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2024.1433181/full#supplementary-material
Supplementary Table 1 | Selected studies on photoperiod, illumination, and color showing different study foci (sorted according to perception, physiological performance, and behavior) and setups.
References
Adamczyk K., Górecka-Bruzda A., Nowicki J., Gumułka M., Molik E., Schwarz T., et al. (2015). Perception of environment in farm animals - A review. Ann. Anim. Sci. 15, 565–589. doi: 10.1515/AOAS-2015-0031
Alexander G., Shillito E. E. (1977). Importance of visual clues from various body regions in maternal recognition of the young in Merino sheep (Ovis aries). Appl. Anim. Ethol. 3, 137–143. doi: 10.1016/0304-3762(77)90022-0
Alexander G., Stevens D. (1979). Discrimination of colours and grey shades by Merino ewes: tests using coloured lambs. Appl. Anim. Ethol. 5, 215–231. doi: 10.1016/0304-3762(79)90057-9
Andersen I. L., Færevik G., Bøe K. E., Janczak A. M., Bakken M. (2000). Effects of diazepam on the behaviour of weaned pigs in three putative models of anxiety. Appl. Anim. Behav. Sci. 68, 121–130. doi: 10.1016/S0168-1591(00)00098-8
Andersson H. (2000). Photoperiodism in pigs. (Uppsala: Swedish University of Agricultural Sciences).
Andersson L. (2012). The effects of two light programs on sleep in dairy cattle. (Uppsala: SLU, Dept. of Animal Nutrition and Management).
Andersson L. S., Wilbe M., Viluma A., Cothran G., Ekesten B., Ewart S., et al. (2013). Equine multiple congenital ocular anomalies and silver coat colour result from the pleiotropic effects of mutant PMEL. PloS One 8, e75639. doi: 10.1371/journal.pone.0075639
Babicz M., Paprocka S. (2018). Analysis of the weaned piglets reaction on the handling material with coloured balls and possibility of exploration and chewing. Ann. Warsaw. Univ. Life Sci. - SGGW. - Anim. Sci. 57, 87–93. doi: 10.22630/AAS.2018.57.2.8
Baldwin B. A. (1981). Shape discrimination in sheep and calves. Anim. Behav. 29, 830–834. doi: 10.1016/S0003-3472(81)80017-6
Baldwin B. A., Start I. B. (1981). Sensory reinforcement and illumination preference in sheep and calves. Proc. R. Soc London. Ser. B. Biol. Sci. 211, 513–526. doi: 10.1098/rspb.1981.0020
Baldwin B. A., Start I. B. (1985). Illumination preferences of pigs. Appl. Anim. Behav. Sci. 14, 233–243. doi: 10.1016/0168-1591(85)90004-8
Banks M. S., Sprague W. W., Schmoll J., Parnell J. A. Q., Love G. D. (2015). Why do animal eyes have pupils of different shapes? Sci. Adv. 1, e1500391. doi: 10.1126/sciadv.1500391
Bayerische Landesanstalt für Landwirtschaft. (2012). Licht und Lichtprogramme in der Rinderhaltung, (Poing: Bayerische Landesanstalt für Landwirtschaft (LfL)).
Bazely D. R. (1988). Foraging behaviour of sheep (Ovis ones L.) grazing on swards of perennial ryegrass (Lolium perenne L.) (Oxford: University of Oxford).
Bazely D. R., Ensor C. V. (1989). Discrimination learning in sheep with cues varying in brightness and hue. Appl. Anim. Behav. Sci. 23, 293–299. doi: 10.1016/0168-1591(89)90098-1
Berge E. (1997). Housing of sheep in cold climate. Livest. Prod. Sci. 49, 139–149. doi: 10.1016/S0301-6226(97)00010-9
Berger J. (1986). Wild horses of the Great Basin: social competition and population size. Wildl. Behav. Ecol. 326. doi: 10.3/JQUERY-UI.JS
Berger A., Scheibe K. M., Eichhorn K., Scheibe A., Streich J. (1999). Diurnal and ultradian rhythms of behaviour in a mare group of Przewalski horse (Equus ferus przewalskii), measured through one year under semi-reserve conditions. Appl. Anim. Behav. Sci. 64, 1–17. doi: 10.1016/S0168-1591(99)00026-X
Bernhart K., Werner D., Greif M., Esselmann C., Schwenzfeier-Hellkamp E., Reiter K. (2018).Einfluss von unterschiedlichen Lichtspektren, Lichtlängen und Beleuchtungsstärken auf das Verhalten von Milchkühen. In: Aktuelle Arbeiten zur artgemäßen Tierhaltung, (KTBL). Available online at: https://cris.fau.de/converis/portal/publication/204755515?lang=de_DE (Accessed January 27, 2023).
Bertolucci C., Giannetto C., Fazio F., Piccione G. (2008). Seasonal variations in daily rhythms of activity in athletic horses. Animal 2, 1055–1060. doi: 10.1017/S1751731108002267
Bilodeau P. P., Petitclerc D., St. Pierre N., Pelletier G., St. Laurent G. J. (1989). Effects of photoperiod and pair-feeding on lactation of cows fed corn or barley grain in total mixed rations. J. Dairy. Sci. 72, 2999–3005. doi: 10.3168/JDS.S0022-0302(89)79452-2
Blackshaw J. K., Allan D. J., McGreevy P. (1983). Notes on some topics in applied animal behaviour (Brisbane: University of Queensland).
Bocquier F., Kann G., Thériez M. (1990). Relationships between secretory patterns of growth hormone, prolactin and body reserves and milk yield in dairy ewes under different photoperiod and feeding conditions. Anim. Sci. 51, 115–125. doi: 10.1017/S0003356100005213
Bowmaker J. K. (1998). Evolution of colour vision in vertebrates. Eye 12, 541–547. doi: 10.1038/eye.1998.143
Braekevelt C. R. (1983). Retinal photoreceptor fine structure in the domestic sheep. Cells Tissues. Organs. 116, 265–275. doi: 10.1159/000145750
Brandskyddsföreningens Service AB (2023). Produktion och form: Förlaget Brandskyddsföreningen. (Stockholm: Förlaget Brandskyddsföreningen).
Briefer E. F., de la Torre M. P., McElligott A. G. (2012). Mother goats do not forget their kids’ calls. Proc. Biol. Sci. 279, 3749–3755. doi: 10.1098/RSPB.2012.0986
Buchenauer D., Fritsch B. (1980). Zum Farbsehvermögen von Hausziegen (Capra hircus L.). Z. Tierpsychol. 53, 225–230. doi: 10.1111/J.1439-0310.1980.TB01051.X
Burla J.-B., Bachmann I., Hillmann E., Westerath H. S. (2016). The effects of coloured LED light on behaviour and physiology in healthy horses. Open Biol. Sci. J. 2, 1–16. doi: 10.2174/2352633501602010001
Butterworth A. (2013). On-farm broiler welfare assessment and associated training. Braz. J. Poult. Sci. 15, 71–78. doi: 10.1590/S1516-635X2013000200001
Carroll J., Murphy C. J., Neitz M., Ver Hoeve J. N., Neitz J. (2001). Photopigment basis for dichromatic color vision in the horse. J. Vis. 1, 2–2. doi: 10.1167/1.2.2
Casamassima D., Cagnetta P., Muscio A. (1990). Effect of photoperiod on the performance of artificially fed lambs. Zootec. e. Nutr. Anim. 16, 255–262.
Casamassima D., Sevi A., Montemurro O. (1993). Effect of light intensity on performance and behaviour of Comisana ewe-lambs intensively reared. Zoot. Nutr. Anim. 19, 251–259.
Casamassima D., Sevi A., Montemurro O. (1994). Effect of colour of light on production and behaviour of lambs for slaughter. Zootec. e. Nutr. Anim. 20.
Chandler M. J., Smith P. J., Samuelson D. (1999). Photoreceptor density of the domestic pig retina. Vet. Ophthalmol. 2, 179–184. doi: 10.1046/j.1463-5224.1999.00077.x
Chemineau P., Malpaux B., Delgadillo J. A., Guérin Y., Ravault J. P., Thimonier J., et al. (1992). Control of sheep and goat reproduction: use of light and melatonin. Anim. Reprod. Sci. 30, 157–184. doi: 10.1016/0378-4320(92)90010-B
Claus R., Weiler U. (1985). Influenceof light and photoperiodicityon pig prolificacy. J. Reprod. Fertil. 33, 185–197.
Coulon M., Deputte B. L., Heyman Y., Delatouce L., Richard C., Baudoin C. (2007). Visual discrimination by heifers (Bos taurus) of their own species. J. Comp. Psychol. 121, 198–204. doi: 10.1037/0735-7036.121.2.198
Council of the European Union (2008). European Union Council Directive. Council Directive 2008/120/EC of 18 December 2008 Laying Down Minimum Standards for the Protection of Pigs. Available online at: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX:32008L0120 (Accessed March 7, 2024).
Culda C. A., Stermin A. N. (2019). Horses’ Senses involvement in food location and selection. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Anim. Sci. Biotechnol. 76, 94–101. doi: 10.15835/buasvmcn-asb:0008.19
Cummins K. A., Myers L. J. (1991). Olfactory and visual cues, individual recognition, and social aggression in lactating dairy cows. J. Dairy. Sci. 74.
Dahl G. E., Buchanan B. A., Tucker H. A. (2000). Photoperiodic effects on dairy cattle: A review. J. Dairy. Sci. 83, 885–893. doi: 10.3168/jds.S0022-0302(00)74952-6
Dahl G. E., Petitclerc D. (2003). Management of photoperiod in the dairy herd for improved production and health. J. Anim. Sci. 81, 11–17. doi: 10.2527/2003.81SUPPL_311X
Dahl G. E., Tao S., Thompson I. M. (2012). Lactation Biology Symposium: Effects of photoperiod on mammary gland development and lactation. J. Anim. Sci. 90, 755–760. doi: 10.2527/jas.2011-4630
Dalmau A., Llonch P., Velarde A. (2009). Professional pig community. Pig. Vis. Manag. Available at: https://www.pig333.com/articles/pig-vision-and-management-handling_981/ (Accessed April 14, 2024).
Dalton I. (2017). Using light to extend the breeding season in goats. Data Driven. Dairy. Decis. Farmers. Available at: https://4d4f.eu/content/using-light-extend-breeding-season-goats (Accessed April 14, 2024).
Dannenmann K., Buchenauer D., Fliegner H. (1985). The behaviour of calves under four levels of lighting. Appl. Anim. Behav. Sci. 13, 243–258. doi: 10.1016/0168-1591(85)90048-6
de Almeida G. L. P., Pandorfi H., Baptista F., Guiselini C., da Cruz V. F., de Almeida G. A. P. (2015). Efficiency of use of supplementary lighting in rearing of dairy calves during milk feeding stage. Rev. Bras. Eng. Agric. e. Ambient. 19, 989–995. doi: 10.1590/1807-1929/AGRIAMBI.V19N10P989-995
Delgadillo J. A., Cortez M. E., Duarte G., Chemineau P., Malpaux B. (2004). Evidence that the photoperiod controls the annual changes in testosterone secretion, testicular and body weight in subtropical male goats. Reprod. Nutr. Dev. 44, 183–193. doi: 10.1051/rnd:2004024
Delgadillo J. A., Flores J. A., Véliz F. G., Hernández H. F., Duarte G., Vielma J., et al. (2002). Induction of sexual activity in lactating anovulatory female goats using male goats treated only with artificially long days. J. Anim. Sci. 80, 2780–2786. doi: 10.2527/2002.80112780X
Deligeorgis S. G., Karalis K., Kanzouros G. (2006). The influence of drinker location and colour on drinking behaviour and water intake of newborn pigs under hot environments. Appl. Anim. Behav. Sci. 96, 233–244. doi: 10.1016/J.APPLANIM.2005.06.006
Dimberton A. (1999). La sécurité de l’éleveur et le bien être des bovins pendant la contention: étude de méthodes adaptées au comportement animal. University of Lyon.
Duarte G., Nava-Hernández M. P., Malpaux B., Delgadillo A., Agraria A., Narro A., et al. (2010). Ovulatory activity of female goats adapted to the subtropics is responsive to photoperiod. Anim. Reprod. Sci. 120, 65–70. doi: 10.1016/J.ANIREPROSCI.2010.04.004
Dureau P., Jeanny J., Clerc B., Dufier J., Courtois Y. (1996). Long term light-induced retinal degeneration in the miniature pig - PubMed. Mol. Vis. 2, 7–14.
Elliot A. J. (2015). Color and psychological functioning: A review of theoretical and empirical work. Front. Psychol. 6. doi: 10.3389/FPSYG.2015.00368/BIBTEX
Elsabagh M., Mon M., Takao Y., Shinoda A., Watanabe T., Kushibiki S., et al. (2020). Exposure to blue LED light before the onset of darkness under a long-day photoperiod alters melatonin secretion, feeding behaviour and growth in female dairy calves. Anim. Sci. J. 91, 1–8. doi: 10.1111/ASJ.13353
Evans N. M., Hacker R. R. (1989). Effect of chronobiological manipulation of lactation in the dairy cow. J. Dairy. Sci. 72, 2921–2927. doi: 10.3168/JDS.S0022-0302(89)79443-1
Fédération Equestre Internationale (2020). MULTI-SPORT GAMES - EQUESTRIAN VENUE REQUIREMENTS/MINIMUM STANDARDS.
Fischer W., Deswysen A. G., Despres L., Dutilleul P., Lobato J. (1997). Ingestive behavior of sheep fed a hay-based diet over a six-month period. Soc Bras. Zootec. Rev. 26, 1032–1038.
Flores M. J., Delgadillo J. A., Flores J. A., Pastor F. J., Duarte G., Vielma J., et al. (2015). Artificial long days increase milk production in subtropical lactating goats managed under extensive grazing conditions. J. Agric. Sci. 153, 335–342. doi: 10.1017/S0021859614000379
Flores M. J., Flores J. A., Duarte G., Vielma J., Delgadillo J. A., Hernández H. (2018). Artificial long-day photoperiod in the subtropics increases body weight in goat kids born in the autumn. Small. Rumin. Res. 169, 181–185. doi: 10.1016/J.SMALLRUMRES.2018.11.013
Foppa L., Caldara F. R., de Moura R., MaChado S. P., Nääs I. A., Garcia R. G., et al. (2018). Pig’s behavioral response in nursery and growth phases to environmental enrichment objects. Spanish. J. Agric. Res. 16, e0507–e0507. doi: 10.5424/SJAR/2018163-12303
Fredriksen B., Nafstad O., Lium B. M., Marka C. H., Dahl E., Choinski J. U. (2006). Artificial light programmes in entire male pig production – effects on androstenone, skatole and animal welfare. Acta Vet. Scand. 48, 1–2. doi: 10.1186/1751-0147-48-S1-P3
Garcia-Hernandez R., Newton G., Horner S., Nuti L. C. (2007). Effect of photoperiod on milk yield and quality, and reproduction in dairy goats. Livest. Sci. 110, 214–220. doi: 10.1016/J.LIVSCI.2006.11.005
Gilbert B. J., Arave C. W. (1986). Ability of cattle to distinguish among different wavelengths of light. J. Dairy. Sci. 69, 825–832. doi: 10.3168/JDS.S0022-0302(86)80472-6
Glatz P. C. (2001). Effect of different lighting sources on behavior and growth of weanling pigs. Asian-Australasian. J. Anim. Sci. 14, 280–287. doi: 10.5713/AJAS.2001.280
Götz S., Raoult C. M. C., Reiter K., Wensch-Dorendorf M., von Borell E. (2022). Lying, feeding and activity preference of weaned piglets for LED-illuminated vs. Dark pen compartments. Animals 12, 1–8. doi: 10.3390/ani12020202
Götz S., Raoult C. M. C., Reiter K., Wensch-Dorendorf M., Werner D., von Borell E. (2020). Influence of different LED light colour temperatures on the preference behaviour of weaned piglets. Agriculture 10, 594. doi: 10.3390/agriculture10120594
Graf R. (1976). Das visuelle Orientierungsvermögen der Schweine in Abhängigkeit von der Beleuchtungsstärke. Available online at: https://library.wur.nl/WebQuery/wurpubs/reports/466800 (Accessed August 3, 2022).
Grandin T. (1979). Understanding animal psychology facilitates handling livestock. Vet. Med. Small. Anim. Clin. 74, 697–706.
Grandin T. (1980). Livestock behavior as related to handling facilities design. J. Study. Anim. Probl. 1, 33–52.
Grandin T. (2007). “Behavioural principles of handling cattle and other grazing animals under extensive conditions,” in Livestock handling and transport (CABI, UK), 44–64. doi: 10.1079/9781780643212.0039
Greeff R. (1894). Die retina der Wirbelthiere: Untersuchungen mit der Golgi-Cajal’schen Chromsilbermethode und der Ehrlich’schen Methylenblaufärbung (Wiesbaden: Verlag von J.F. Bergmann).
Grzimek B. (1952). Versuche über das Farbsehen von Pflanzenessern. Z. Tierpsychol. 9, 23–39. doi: 10.1111/J.1439-0310.1952.TB01645.X
Hagen K., Broom D. M. (2003). Cattle discriminate between individual familiar herd members in a learning experiment. Appl. Anim. Behav. Sci. 82, 13–28. doi: 10.1016/S0168-1591(03)00053-4
Hall C. A., Cassaday H. J. (2006). An investigation into the effect of floor colour on the behaviour of the horse. Appl. Anim. Behav. Sci. 99, 301–314. doi: 10.1016/j.applanim.2005.10.018
Hall C. A., Cassaday H. J., Vincent C. J., Derrington A. M. (2006). Cone excitation ratios correlate with color discrimination performance in the horse (Equus caballus). J. Comp. Psychol. 120, 438–448. doi: 10.1037/0735-7036.120.4.438
Hanggi E. B. (2006). Equine cognition and perception: understanding the horse. Divers. Cogn. Evol. Dev. Domest. Pathol. Kyoto. Univ. Press Kyoto., 98–118.
Hanggi E. B., Ingersoll J. F. (2009). Stimulus discrimination by horses under scotopic conditions. Behav. Processes. 82, 45–50. doi: 10.1016/j.beproc.2009.04.009
Harman A. M., Moore S., Hoskins R., Keller P. (1999). Horse vision and an explanation for the visual behaviour originally explained by the ‘ramp retina.’. Equine. Vet. J. 31, 384–390. doi: 10.1111/j.2042-3306.1999.tb03837.x
Hastings M. H., Reddy A. B., Maywood E. S. (2003). A clockwork web: circadian timing in brain and periphery, in health and disease. Nat. Rev. Neurosci. 4, 649–661. doi: 10.1038/nrn1177
Hirata M., Arimoto C., Hattori N., Anzai H. (2019). Can cattle visually discriminate between green and dead forages at a short distance while moving in the field? Anim. Cogn. 22, 707–718. doi: 10.1007/S10071-019-01268-Z/FIGURES/5
Hirata M., Kusatake N. (2020). How cattle discriminate between green and dead forages accessible by head and neck movements by means of senses: reliance on vision varies with the distance to the forages. Anim. Cogn. 23, 405–414. doi: 10.1007/s10071-019-01344-4
Hirt A., Maisack C., Moritz J. (2020). Tierschutzgesetz: Mit TierSchHundeV, TierSchNutztV, TierSchVersV, TierSchTrV, EU-Tiertransport-VO, TierSchlV, EU-Tierschlacht-VO. (Germany: Verlag Franz Vahlen).
Hjalmarsson F., Olsson I., Ferneborg S., Agenäs S., Ternman E., Hjalmarsson F., et al. (2014). Effect of low light intensity at night on cow traffic in automatic milking systems. Anim. Prod. Sci. 54, 1784–1786. doi: 10.1071/AN14215
Hughes A., Whitteridge D. (1973). The receptive fields and topographical organization of goat retinal ganglion cells. Vision Res. 13, 1101–1IN7. doi: 10.1016/0042-6989(73)90147-8
Jacobs G. H., Deegan J. F., Neitz J. (1998). Photopigment basis for dichromatic color vision in cows, goats, and sheep. Vis. Neurosci. 15, 581–584. doi: 10.1017/S0952523898153154
Johnson G. L. (1901). I. Contributions to the comparative anatomy of the mammalian eye, chiefly based on ophthalmoscopic examination. Philos. Trans. R. Soc London. Ser. B. Contain. Pap. Biol. Character. 194, 1–82. doi: 10.1098/rstb.1901.0001
Kendrick K. M. (2008). “Sheep senses, social cognition and capacity for consciousness,” in The Welfare of Sheep (Springer, Dordrecht), 135–157. doi: 10.1007/978-1-4020-8553-6_4
Kendrick K. M., Atkins K., Hinton M. R., Broad K. D., Fabre-Nys C., Keverne B. (1995). Facial and vocal discrimination in sheep. Anim. Behav. 49, 1665–1676. doi: 10.1016/0003-3472(95)90088-8
Klocek C., Nowicki J., Brudzisz B., Pabiańczyk M. (2016). Colour preferences in pigs. Sci. Ann. Polish. Soc Anim. Prod. 12, 123–129. doi: 10.5604/01.3001.0013.5420
Land M. F. (2006). Visual optics: the shapes of pupils. Curr. Biol. 16, R167–R168. doi: 10.1016/j.cub.2006.02.046
Lim D.-H., Kim T.-I., Park S.-M., Ki K.-S., Kim Y. (2021). Effects of photoperiod and light intensity on milk production and milk composition of dairy cows in automatic milking system. J. Anim. Sci. Technol. 63, 626–639. doi: 10.5187/jast.2021.e59
Lindkvist S., Ferneborg S., Ståhlberg K., Bånkestad D., Ekesten B., Agenäs S., et al. (2023). Effect of light intensity, spectrum, and uniformity on the ability of dairy cows to navigate through an obstacle course. J. Dairy. Sci. 106, 7698–7710. doi: 10.3168/jds.2023-23469
Lindkvist S., Ternman E., Ferneborg S., Bånkestad D., Lindqvist J., Ekesten B., et al. (2021). Effects of achromatic and chromatic lights on pupillary response, endocrinology, activity, and milk production in dairy cows. PloS One 16, e0253776. doi: 10.1371/journal.pone.0253776
Lomas C. A. (1994). The effect of supplementary light on the behaviour, physiology and productivity of cattle (Bangor: University of Wales).
Lomas C. A., Piggins D., Phillips C. J.C. (1998). Visual awareness. Appl. Anim. Behav. Sci. 57, 247–257. doi: 10.1016/S0168-1591(98)00100-2
Macuda T., Timney B. (1999). Luminance and chromatic discrimination in the horse (Equus caballus). Behav. Processes. 44, 301–307. doi: 10.1016/S0376-6357(98)00039-4
Magdesian K. G., Williams D. C., Aleman M., LeCouteur R. A., Madigan J. E. (2009). Evaluation of deafness in American Paint Horses by phenotype, brainstem auditory-evoked responses, and endothelin receptor B genotype. J. Am. Vet. Med. Assoc. 235, 1204–1211. doi: 10.2460/javma.235.10.1204
Manida M., Satou M., Kurohiji I. (1989). Behavioral response of cattle to electric fence tapes in different colors. Japanese. J. Zootech. Sci. 60, 236–239.
Martelli G., Boccuzzi R., Grandi M., Mazzone G., Zaghini G., Sardi L., et al (2010). The effects of two different light intensities on the production and behavioural traits of Italian heavy pigs. Berl. Munch. Tierarztl. Wochenschr. 123, 457–462. doi: 10.2376/0005-9366-123-457
Martelli G., Nannoni E., Grandi M., Bonaldo A., Zaghini G., Vitali M., et al. (2015). Growth parameters, behavior, and meat and ham quality of heavy pigs subjected to photoperiods of different duration. J. Anim. Sci. 93, 758–766. doi: 10.2527/JAS.2014-7906
Martelli G., Scalabrin M., Scipioni R., Sardi L. (2005). The effects of the duration of the artificial photoperiod on the growth parameters and behaviour of heavy pigs. Vet. Res. Commun. 29 Suppl 2, 367–369. doi: 10.1007/S11259-005-0367-8
Martin A.-M., Elliott J. A., Duffy P., Blake C. M., Attia S., Katz L. M., et al. (2010). Circadian regulation of locomotor activity and skeletal muscle gene expression in the horse. J. Appl. Physiol. 109, 1328–1336. doi: 10.1152/japplphysiol.01327.2009
Mayes E., Duncan P. (1986). Temporal patterns of feeding behaviour in free-ranging horses. Behaviour 96, 105–129. doi: 10.1163/156853986X00243
McGlone J. J., Stansbury W. F., Tribble L. F., Morrow J. L. (1988). Photoperiod and heat stress influence on lactating sow performance and photoperiod effects on nursery pig performance. J. Anim. Sci. 66, 1915–1919. doi: 10.2527/jas1988.6681915x
Miller A. R. E., Stanisiewski E. P., Erdman R. A., Douglass L. W., Dahl G. E. (1999). Effects of long daily photoperiod and bovine somatotropin (Trobest®) on milk yield in cows. J. Dairy. Sci. 82, 1716–1722. doi: 10.3168/jds.S0022-0302(99)75401-9
Morrissey A. D., Cameron A. W. N., Tilbrook A. J. (2008). Artificial lighting during winter increases milk yield in dairy ewes. J. Dairy. Sci. 91, 4238–4243. doi: 10.3168/JDS.2007-0918
Murphy B. A. (2019). Circadian and circannual regulation in the horse: internal timing in an elite athlete. J. equine. Vet. Sci. 76, 14–24. doi: 10.1016/j.jevs.2019.02.026
Murphy J., Hall C., Arkins S. (2009). What horses and humans see: A comparative review. Int. J. Zool. 2009, 721798. doi: 10.1155/2009/721798
Murphy B. A., Herlihy M. M., Nolan M. B., O’Brien C., Furlong J. G., Butler S. T. (2021). Identification of the blue light intensity administered to one eye required to suppress bovine plasma melatonin and investigation into effects on milk production in grazing dairy cows. J. Dairy. Sci. 104, 12127–12138. doi: 10.3168/jds.2021-20526
Murphy B. A., Walsh C. M., Woodward E. M., Prendergast R. L., Ryle J. P., Fallon L. H., et al. (2014). Blue light from individual light masks directed at a single eye advances the breeding season in mares. Equine. Vet. J. 46, 601–605. doi: 10.1111/evj.12153
Mutton W. J. (1987). The influence of photoperiod and light intensity on the farrowing sow and litter. BSAP. Occas. Publ. 11, 137–138. doi: 10.1017/S0263967X00001907
Nawroth C., Langbein J., Coulon M., Gabor V., Oesterwind S., Benz-Schwarzburg J., et al. (2019). Farm animal cognition-linking behavior, welfare and ethics. Front. Vet. Sci. 6. doi: 10.3389/fvets.2019.00024
Neitz J., Jacobs G. H. (1989). Spectral sensitivity of cones in an ungulate. Vis. Neurosci. 2, 97–100. doi: 10.1017/S0952523800011949
Neveux C., Ferard M., Melac E., Pousset N. (2023). Effects of different LED lighting conditions on young horses during trailer loading and stationary confinement. Appl. Anim. Behav. Sci. 261, 105885. doi: 10.1016/j.applanim.2023.105885
Ni J.-Q., Liu S., Radcliffe J. S., Vonderohe C. (2017). Evaluation and characterisation of Passive Infrared Detectors to monitor pig activities in an environmental research building. Biosyst. Eng. 158, 86–94. doi: 10.1016/j.biosystemseng.2017.03.014
Nielsen B. L., Jezierski T., Bolhuis J. E., Amo L., Rosell F., Oostindjer M., et al. (2015). Olfaction: An overlooked sensory modality in applied ethology and animal welfare. Front. Vet. Sci. 2. doi: 10.3389/FVETS.2015.00069/BIBTEX
Nowicki J., Klocek C. (2012). The effect of aromatized environmental enrichment in pen on social relations and behavioural profile of newly mixed weaners. Ann. Anim. Sci. 12, 403–412. doi: 10.2478/v10220-012-0034-2
O’Connor Z. (2011). Colour psychology and colour therapy: Caveat emptor. Color Res. Appl. 36, 229–234. doi: 10.1002/col.20597
Ollivier F. J., Samuelson D. A., Brooks D. E., Lewis P. A., Kallberg M. E., Komáromy A. M. (2004). Comparative morphology of the tapetum lucidum (among selected species). Vet. Ophthalmol. 7, 11–22. doi: 10.1111/j.1463-5224.2004.00318.x
Osborne V. R., Odongo N. E., Edwards A. M., McBride B. W. (2007). Effects of photoperiod and glucose-supplemented drinking water on the performance of dairy calves. J. Dairy. Sci. 90, 5199–5207. doi: 10.3168/JDS.2007-0402
Oshiro S., Nakamae H., Hirayama T., Furuta K., Hongo F., Hirakawa M., et al. (1996). Effects of duration of photoperiod on the rumination behavior of goats. Small. Rumin. Res. 22, 97–102. doi: 10.1016/S0921-4488(96)00886-3
Paggi A. C., Diesel T. A., de Oliveira P. A. V., Coldebella A., Paiano D., Marcanzoni V. C. B., et al. (2020). Alternative light colors in suckling piglet creep area: Preference and validation tests in swine maternity. Cienc. Rural 50, 1–10. doi: 10.1590/0103-8478CR20190328
Peters R. R., Chapin L. T., Emery R. S., Tucker H. A. (1981). Milk yield, feed intake, prolactin, growth hormone, and glucocorticoid response of cows to supplemented light. J. Dairy. Sci. 64, 1671–1678. doi: 10.3168/JDS.S0022-0302(81)82745-2
Peters R. R., Chapin L. T., Leining K. B., Tucker H. A. (1978). Supplemental lighting stimulates growth and lactation in cattle. Sci. (80-.). 199, 911–912. doi: 10.1126/science.622576
Phillips C. J. C., Lomas C. A. (2001). The perception of color by cattle and its influence on behavior. J. Dairy. Sci. 84, 807–813. doi: 10.3168/jds.S0022-0302(01)74537-7
Phillips C. J. C., Schofield S. A. (1989). The effect of supplementary light on the production and behaviour of dairy cows. Anim. Sci. 48, 293–303. doi: 10.1017/S0003356100040290
Phillips C. J. C., Weiguo L. (1991). Brightness discrimination abilities of calves relative to those of humans. Appl. Anim. Behav. Sci. 31, 25–33. doi: 10.1016/0168-1591(91)90150-V
Piccione G., Caola G., Refinetti R. (2005). Temporal relationships of 21 physiological variables in horse and sheep. Comp. Biochem. Physiol. - A. Mol. Integr. Physiol. 142, 389–396. doi: 10.1016/j.cbpa.2005.07.019
Piccione G., Giannetto C., Casella S., Caola G. (2008). Seasonal change of daily motor activity rhythms in Capra hircus. Can. J. Anim. Sci. 88, 351–355. doi: 10.4141/CJAS07125
Pick D. F., Lovell G., Brown S., Dail D. (1994). Equine color perception revisited. Appl. Anim. Behav. Sci. 42, 61–65. doi: 10.1016/0168-1591(94)90007-8
Piggins D., Phillips C. J. C. (1996). The eye of the domesticated sheep with implications for vision. Anim. Sci. 62, 301–308. doi: 10.1017/S1357729800014612
Pollott G. E., Gootwine E. (2004). Reproductive performance and milk production of assaf sheep in an intensive management system. J. Dairy. Sci. 87, 3690–3703. doi: 10.3168/JDS.S0022-0302(04)73508-0
Prince J. H. (1977). “The eye and vision,” in Dukes physiology of domestic animals. Ithaca: Cornell University Press, 696–712.
Reksen O., Tverdal A., Landsverk K., Kommisrud E., Bøe K. E., Ropstad E. (1999). Effects of photointensity and photoperiod on milk yield and reproductive performance of Norwegian red cattle. J. Dairy. Sci. 82, 810–816. doi: 10.3168/jds.S0022-0302(99)75300-2
Riksidrottsförbundet (2023). Riksidrottsförbundets måttuppgifter för fritidsanläggningar. Available online at: https://www.rf.se/download/18.5424ab70183abb853551a3d/1665068711761/Anl%C3%A4ggnings-och%20idrottsmilj%C3%B6politiskt%20program.pdf (Accessed November 29, 2024).
Riol J. A., Sanchez J. M., Eguren V. G., Gaudioso V. R. (1989). Colour perception in fighting cattle. Appl. Anim. Behav. Sci. 23, 199–206. doi: 10.1016/0168-1591(89)90110-X
Rivas-Muñoz R., Carrillo E., Rodriguez-Martinez R., Leyva C., Mellado M., Véliz F. G. (2010). Effect of body condition score of does and use of bucks subjected to added artificial light on estrus response of Alpine goats. Trop. Anim. Health Prod. 42, 1285–1289. doi: 10.1007/s11250-010-9563-9
Rørvang M. V., Nielsen B. L., McLean A. N. (2020). Sensory abilities of horses and their importance for equitation science. Front. Vet. Sci. 7. doi: 10.3389/fvets.2020.00633
Roth L. S. V., Balkenius A., Kelber A. (2007). Colour perception in a dichromat. J. Exp. Biol. 210, 2795–2800. doi: 10.1242/jeb.007377
Roth L. S. V., Balkenius A., Kelber A. (2008). The absolute threshold of colour vision in the horse. PloS One 3, e3711. doi: 10.1371/journal.pone.0003711
Russo V. M., Cameron A. W. N., Dunshea F. R., Tilbrook A. J., Leury B. J. (2013). Artificially extending photoperiod improves milk yield in dairy goats and is most effective in late lactation. Small. Rumin. Res. 113, 179–186. doi: 10.1016/j.smallrumres.2013.01.002
Ryba N., Uhl R. (1989). In vitro dark adaptation and preservation of electrical light responses in the retina from bovine eyes. Exp. Brain Res 74, 618–624.
Saslow C. A. (1999). Factors affecting stimulus visibility for horses. Appl. Anim. Behav. Sci. 61, 273–284. doi: 10.1016/S0168-1591(98)00205-6
Scaillierez A. J., van Nieuwamerongen-de Koning S. E., Boumans I., van der Tol P. P. J., Schnabel S. K., Bokkers E. A. M. (2024). Effect of light intensity on behaviour, health and growth of growing-finishing pigs. Animal 18, 101092. doi: 10.1016/j.animal.2024.101092
Schanbacher B. D., Crouse J. D. (1981). Photoperiodic regulation of growth: a photosensitive phase during light-dark cycle. Am. J. Physiol. 4, E1–E5. doi: 10.1152/ajpendo.1981.241.1.E1
Shrader A. M., Brown J. S., Kerley G. I. H., Kotler B. P. (2008). Do free-ranging domestic goats show ‘landscapes of fear’? Patch use in response to habitat features and predator cues. J. Arid. Environ. 72, 1811–1819. doi: 10.1016/j.jaridenv.2008.05.004
Siemers B., Backhaus W., Franz H. (1999). “Colour vision in farm animals: evidence for trichromacy in dwarf goats,” in Proceedings of the 27th neurobiology conference (Göttingen), Georg Thieme Verlag Stuttgart, NY (Georg Thieme Verlag Stuttgart, Göttingen).
Smith S., Goldman L. (1999). Color discrimination in horses. Appl. Anim. Behav. Sci. 62, 13–25. doi: 10.1016/S0168-1591(98)00206-8
Son J., Park J., Kang D., Belal S. A., Cha J., Shim K. (2020). Effects of white, yellow, and blue colored LEDs on milk production, milk composition, and physiological responses in dairy cattle. Anim. Sci. J. 91, 1–9. doi: 10.1111/ASJ.13337
Stevenson J. S., Pollmann D. S., Davis D. L., Murphy J. P. (1983). Influence of Supplemental Light on Sow Performance during and after Lactation. J. Anim. Sci. 56, 1282–1286. doi: 10.2527/jas1983.5661282x
Tanaka T., Asakawa K., Kawahara Y., Tanida H., Yoshimoto T. (1989a). Color discrimination in sheep. Japanese. J. Livest. Manage. 24, 89–94. doi: 10.20652/kachikunokanri.24.3_89
Tanaka T., Sekino M., Tanida H., Yoshimoto T. (1989b). Ability to discriminate between similar colors in sheep. Japanese. J. Zootech. Sci. 60, 880–884. doi: 10.2508/chikusan.60.880
Tanida H., Miura A., Tanaka T., Yoshimoto T. (1996). Behavioral responses of piglets to darkness and shadows. Appl. Anim. Behav. Sci. 49, 173–183. doi: 10.1016/0168-1591(96)01039-8
Tanida H., Senda K., Suzuki S., Tanaka T., Yoshimoto T. (1991). Color discrimination in weanling pigs. J. Anim. Sci. Technol. 62, 1029–1034.
Taylor N., Prescott N., Perry G., Potter M., Le Sueur C., Wathes C. (2006). Preference of growing pigs for illuminance. Appl. Anim. Behav. Sci. 96, 19–31. doi: 10.1016/j.applanim.2005.04.016
Thiéry J. C., Chemineau P., Hernandez X., Migaud M., Malpaux B. (2002). Neuroendocrine interactions and seasonality. Domest. Anim. Endocrinol. 23, 87–100. doi: 10.1016/S0739-7240(02)00148-0
Thonney M. L., Waldron D. F., Zobel G., MacNeil M. D. (2020). “Domestic sheep and goats,” in Guide for the care and use of agricultural animals in research and teaching. (Champaign: American Dairy Science Association), 141–155.
Thornton P. K. (2010). Livestock production: recent trends, future prospects. Philos. Trans. R. Soc B. Biol. Sci. 365, 2853–2867. doi: 10.1098/rstb.2010.0134
TierSchNutztV14 (2021). Verordnung zum Schutz Landwirtsch. Nutztiere und anderer zur Erzeugung Tier. Produkte gehaltener Tiere bei ihrer Haltung (Tierschutz-Nutztierhaltungsverordnung - TierSchNutztV). Available online at: https://www.gesetze-im-internet.de/tierschnutztv/BJNR275800001.html (Accessed April 20, 2024).
Timney B., Keil K. (1999). Local and global stereopsis in the horse. Vision Res. 39, 1861–1867. doi: 10.1016/S0042-6989(98)00276-4
Timney B., Macuda T. (2001). Vision and hearing in horses. J. Am. Vet. Med. Assoc. 218, 1567–1574. doi: 10.2460/javma.2001.218.1567
Tribe D. E., Gordon J. G. (1949). The importance of colour vision to the grazing sheep. J. Agric. Sci. 39, 313–314. doi: 10.1017/S0021859600004597
Ungerfeld R., Bielli A. (2012). “Seasonal and social factors affecting reproduction,” in Livestock reproduction: bovine, swine, and ruminants (Eolss Publishers, Oxford, UK).
Véliz F. G., Mellado M., Carrillo E., Meza-Herrera C. A., Rivas-Muñoz R. (2009). Effects of a long daily photoperiod on milk yield and ovarian activity of saanen goats in northern Mexico. J. Appl. Anim. Res. 36, 287–290. doi: 10.1080/09712119.2009.9707078
Vestre W. A. (1984). Porcine ophthalmology. Vet. Clin. North Am. Large. Anim. Pract. 6, 667–676. doi: 10.1016/S0196-9846(17)30016-2
Wallgren P., Johansson M., Wallgren T., Susic Z., Sigfridson K., Johansson S.-E. (2024). Impact of feed, light and access to manipulable material on tail biting in pigs with intact tails. Acta Vet. Scand. 66, 2. doi: 10.1186/s13028-023-00716-8
Walls G. L. (1942). The vertebrate eye and its adaptive radiation. Bloomsfield Hills: Cranbrook Institute of Science.
Weiguo L., Phillips C. J. C. (1991). The effects of supplementary light on the behaviour and performance of calves. Appl. Anim. Behav. Sci. 30, 27–34. doi: 10.1016/0168-1591(91)90082-9
Weiler U., Claus R., Dehnhard M., Hofäcker S. (1996). Influence of the photoperiod and a light reverse program on metabolically active hormones and food intake in domestic pigs compared with a wild boar. Can. J. Anim. Sci. 76, 531–539. doi: 10.4141/cjas96-080
Werner D., Schneider M., Schwenzfeier-Hellkamp E., Müller M., Reiter K., Pelzer A. (2018). Beleuchtung und Beleuchtungstechnik im Rinderstall (DLG-Merkblatt), 415.
Wilson A. M., Wright T. C., Cant J. P., Osborne V. R. (2022). Preferences of dairy cattle for supplemental light-emitting diode lighting in the resting area. Animals 12, 1894. doi: 10.3390/ani12151894
Wouters L., De Moor A. (1979). Ultrastructure of the pigment epithelium and the photoreceptors in the retina of the horse. Am. J. Vet. Res. 40, 1066–1071.
Yang J., Shen X. (2022). The application of color psychology in community health environment design. J. Environ. Public Health 4, 1–10. doi: 10.1155/2022/7259595
Zeng Y., Long D., Hu B., Wang H., Pu S., Jian Y., et al. (2023). Study on illumination intensity and duration of LED light sources for a weaned piglet house without natural light. Agriculture 13, 2121. doi: 10.3390/agriculture13112121
Zheng J., Schäfer S., Ebschke S., Nawroth C., Von Borell E. (2015). “Effects of light intensity on rhythmicity of core body temperature and body lesion scores in growing pigs,” in Proceedings of the 49th Congress of the International Society for Applied Ethology. 195 (Wageningen Academic).
Zonderland J. J., Cornelissen L., Wolthuis-Fillerup M., Spoolder H. A. M. (2008). Visual acuity of pigs at different light intensities. Appl. Anim. Behav. Sci. 111, 28–37. doi: 10.1016/j.applanim.2007.05.010
Keywords: illuminance, color, photoperiodicity, animal welfare, behavior, husbandry implications
Citation: Stuhlträger J, von Borell E, Langbein J, Nawroth C, Rørvang MV and Raoult CMC (2025) The role of light and vision in farmed ungulates and implications for their welfare. Front. Anim. Sci. 5:1433181. doi: 10.3389/fanim.2024.1433181
Received: 15 May 2024; Accepted: 30 December 2024;
Published: 28 January 2025.
Edited by:
Shelly Druyan, Agricultural Research Organization (ARO), IsraelReviewed by:
Dana L. M. Campbell, Commonwealth Scientific and Industrial Research Organisation (CSIRO), AustraliaDeborah Temple, AWEC Animal Welfare Education Center UAB, Spain
Copyright © 2025 Stuhlträger, von Borell, Langbein, Nawroth, Rørvang and Raoult. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julia Stuhlträger, anVsaWEuc3R1aGx0cmFlZ2VyQGxhbmR3LnVuaS1oYWxsZS5kZQ==
†ORCID: Julia Stuhlträger, orcid.org/0000-0001-9340-3578
Eberhard von Borell, orcid.org/0000-0003-4044-3909
Jan Langbein, orcid.org/0000-0002-1170-5431
Christian Nawroth, orcid.org/0000-0003-4582-4057
Maria Vilain Rørvang, orcid.org/0000-0002-3503-2059
Camille M. C. Raoult, orcid.org/0000-0001-5587-108X