
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Anim. Sci., 05 February 2024
Sec. Animal Nutrition
Volume 5 - 2024 | https://doi.org/10.3389/fanim.2024.1308514
 Nienke de Groot1,2*
Nienke de Groot1,2* Fernando Fariñas3Carolina G. Cabrera-Gómez2
Fernando Fariñas3Carolina G. Cabrera-Gómez2 Francisco J. Pallares4
Francisco J. Pallares4 Guillermo Ramis2
Guillermo Ramis2Weaning is a stressful period in a piglet’s life, when many morphological and functional changes occur in the gastrointestinal tract. Examples are an alteration of the barrier function and an increase in production of cytokines, suggesting the immune system of the GIT is activated after weaning. The goal of this study was to evaluate the effect of a commercially available feed additive based on short and medium chain fatty acids on supporting intestinal health and reduce inflammation in the intestine of post-weaning piglets. Seventy piglets were divided over 2 treatments, a control diet and a blend of short and medium chain fatty acids, including butyrate. On day 0, 15, 30 and 45 after weaning blood samples and intestinal tissue samples were collected from sacrificed piglets. Blood and tissue samples were analysed for cytokine gene expression, and tissue samples from jejunum and ileum were analysed on morphology and gene expression of tight junctions. Results show that the use of a blend of short and medium chain fatty acids, including butyrates, improved intestinal morphology, measured by villous length and v/c ratio, reduced expression of tight junction proteins and influenced cytokine production in the intestine, implicating that the use of organic acids as feed additives potentially can reduce damage caused by weaning to the intestinal barrier and due to the higher absorption capacity of the villi potentially improve nutrient absorption.
The intestinal structure is essential for health and performance of all animals. The epithelial lining in the intestine has an important role in integrity and body defense locally (Eckmann, 1995; Pitman and Blumberg, 2000). It creates a physical barrier, preventing toxic components and pathogens from entering the intestinal mucosa and systemic circulation. At the same time, gastrointestinal tract (GIT) is responsible for the uptake of nutrients. The nutritional ingredients can interact with the microbiota and with the host mucosa (Niewold, 2015).
Weaning is a stressful period which in nature occurs between 10 to 12 weeks of age in a gradual matter, around the time the GIT matures. In professional pig farming weaning occurs between three to four weeks of age (Worobec et al., 1999), and it can cause many morphological and functional changes in the GIT (Moeser et al., 2007b; Wijtten et al., 2011). Villi modifications (Wiese et al., 2003) including a decrease in villous height, occur within a minimum of approximately three days after weaning (van Beers-Schreurs et al., 1998; Vente-Spreeuwenberg et al., 2003; Wang et al., 2011). Villous atrophy is mainly caused by increased programmed cell death and decreased rate of cell renewal (van der Peet-Schwering et al., 2007). Furthermore a high cytokine production has been reported, suggesting the activation of the GIT immune system immediately after weaning (Pié et al., 2004; de Groot et al., 2021).
Another negative effect of weaning is a failure of the intestinal epithelial barrier characterized by increased permeability, on which the age of weaning has a negative effect (Smith et al., 2010). Epithelial cells lining the gastrointestinal tract represent the main barrier between the lumen and outside. The quality of this barrier is primarily regulated by tight junctions (TJs), a combination of different proteins interlocking the epithelial cells strongly to each other (Turner, 2009; Marchiando et al., 2010a). Tight junctions regulate the permeability of the intestinal barrier and are composed of intracellular and apical intercellular membrane proteins e.g., zonulin, occludin, and claudins (Turner, 2009).
In the past, high animal productivity and efficiency were obtained using antimicrobial strategies such as the use of antibiotic growth promoters (AGPs), pharmaceutical levels of zinc oxide (ZnO) or a high use of preventive antibiotics. Health concerns have driven worldwide regulatory restrictions, promoting the limited use of antimicrobials. To support optimal animal health, productivity and efficiency without using antimicrobials, nutrition has an important role beyond solely supplying nutrients and energy. Acidifiers or products based on organic acids (OA) can be an alternative of choice to replace antibiotics in pig diets after weaning. A large number of studies have shown that OA, both short chain fatty acids (SCFA) and medium chain fatty acids (MCFA) are beneficial in enhancing growth performance and nutrient digestibility, as well as modulating intestinal microbiota in pigs (Zentek et al., 2011). This study aimed to analyse the effect of a blend of organic acids on intestinal morphology and intestinal cytokine production in weaned piglets.
This study was performed at a high health farm with 72 piglets (Large White) weaned at 22 ± 3 days of age with an average body weight (BW) of 5.51 ± 1.22 kg. A completely randomized design was used after weaning where the piglets were randomly allotted to 8 pens (0.61m×1.22m), 9 animals per pen, and pens were divided over two treatments. Pigs had access to one drinker (ad lib) and one feeder with 4 spaces per pen.
A two-phase pelleted experimental diet was used (Table 1), without additional additives and pharmaceutical levels of zinc oxide. Dietary treatments consisted of a control diet and a treatment diet (control (cont) diet + 0.2% Presan®-FX (PFX)), a feed additive based on a blend of short chain fatty acids (sorbic, benzoic and butyric acid) and medium chain fatty acids (capric C8, caprylic C10 and lauric acid C12). The feed additive used in the present experiment was provided by Selko® B.V., The Netherlands and the commercially advised inclusion rate was followed.
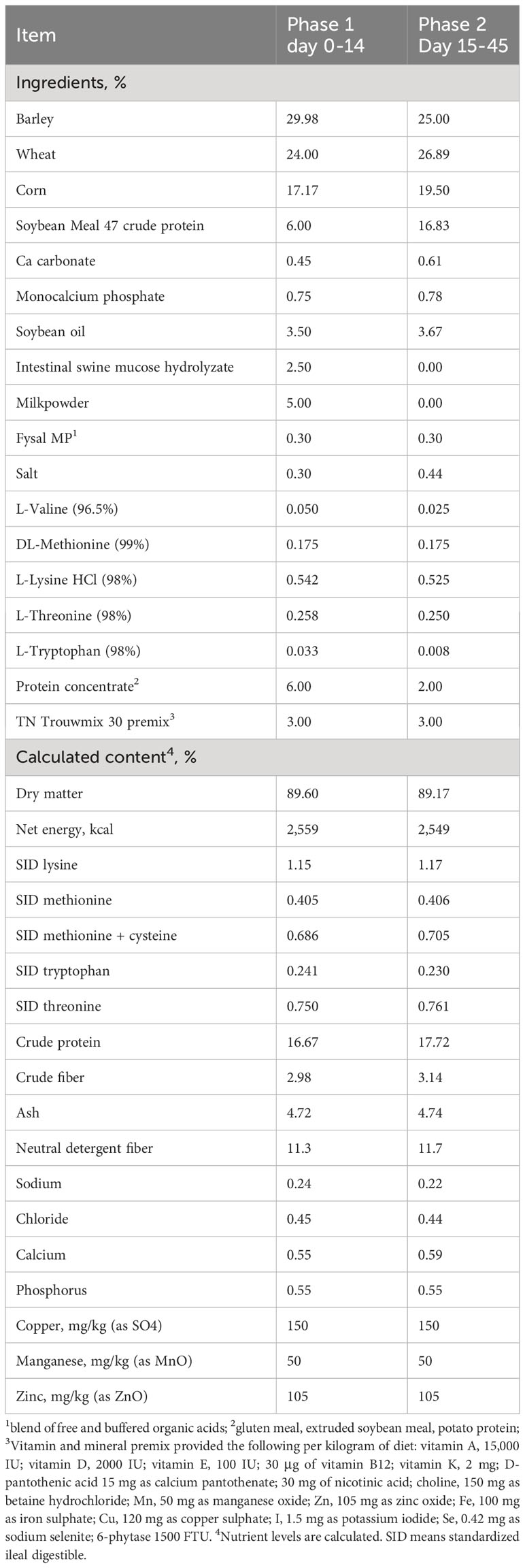
Table 1 Composition of the experimental diets.
Individual BW was recorded at 18 hr after weaning (day 0), days 15, 30, and 45, and average daily gain was calculated. At day 0, 10 control animals were sacrificed to obtain blood from the vena jugular and intestinal tissue samples. Blood samples were collected into Ethylenediaminetetraacetic (EDTA) tubes. Subsequently, piglets were randomly selected for euthanasia, by intravenous injection with an overdose of tiobarbital IV, at day 15 (cont n=10, PFX n=10), day 30 (cont n=7, PFX n=11) and day 45 (cont n=9, PFX n=13) with the same procedure as day 0. Peripheral blood mononuclear cells (PBMCs) were isolated by ficoll Histopaque gradient and preserved in RNAlater (Life Technologies, USA) at -80°C after 24 hours of refrigeration at 8°C. Serum and plasma were isolated from whole blood by centrifugation (251 relative centrifugal force, 10 min at room temperature) and preserved at -80°C up to analysis. Tissue samples were obtained from the ileum, jejunum and colon by 3 cm-long gut sections, and divided in two subsamples, one (20 mg) was preserved in RNAlater (Life Technologies, USA) and one was fixed in 10% buffered formaldehyde.
Ten randomly selected well-oriented intact villi and crypts were measured per piglet and tissue. The villous height (tip to villous-crypt junction) and crypt depth (from villous-crypt junction to the base of villous) were analysed using the ZEISS Efficient Navigation software (Carl Zeiss, Oberkochen, Germany).
Gene expression for cytokines Interleukin (IL)-1α, IL-1β, IL-6, IL-8, IL-10, IL-12p35, IL-12p40, Tumor Necrosis Factor (TNF)-α, Interferon (IFN)-α, IFN-γ and Transforming Growth Factor (TGF)-β was analysed using relative quantification, with primers previously described in the literature (Table 2). Intestinal gene expression of tight junction proteins was studied by doing relative quantification for calprotectin, occludin, zonulin 1 and claudin 1 (Table 2). Total RNA was isolated from tissues by means of Micro RNeasy kit (Qiagen, USA) and DNAc was synthetized using the Geneamp RNA PCR Core Kit (Life Technology, USA). In addition to RNA extracted from tissues as mentioned above, total RNA was isolated from PBMCs. The samples were normalized using the average Ct for glyceraldehyde-3-phosphatedehydrogenase (GAPDH), cyclophilin and β-Actin (Table 2). Data were expressed as fold change, normalized to the lowest value (which was assigned a value 1).
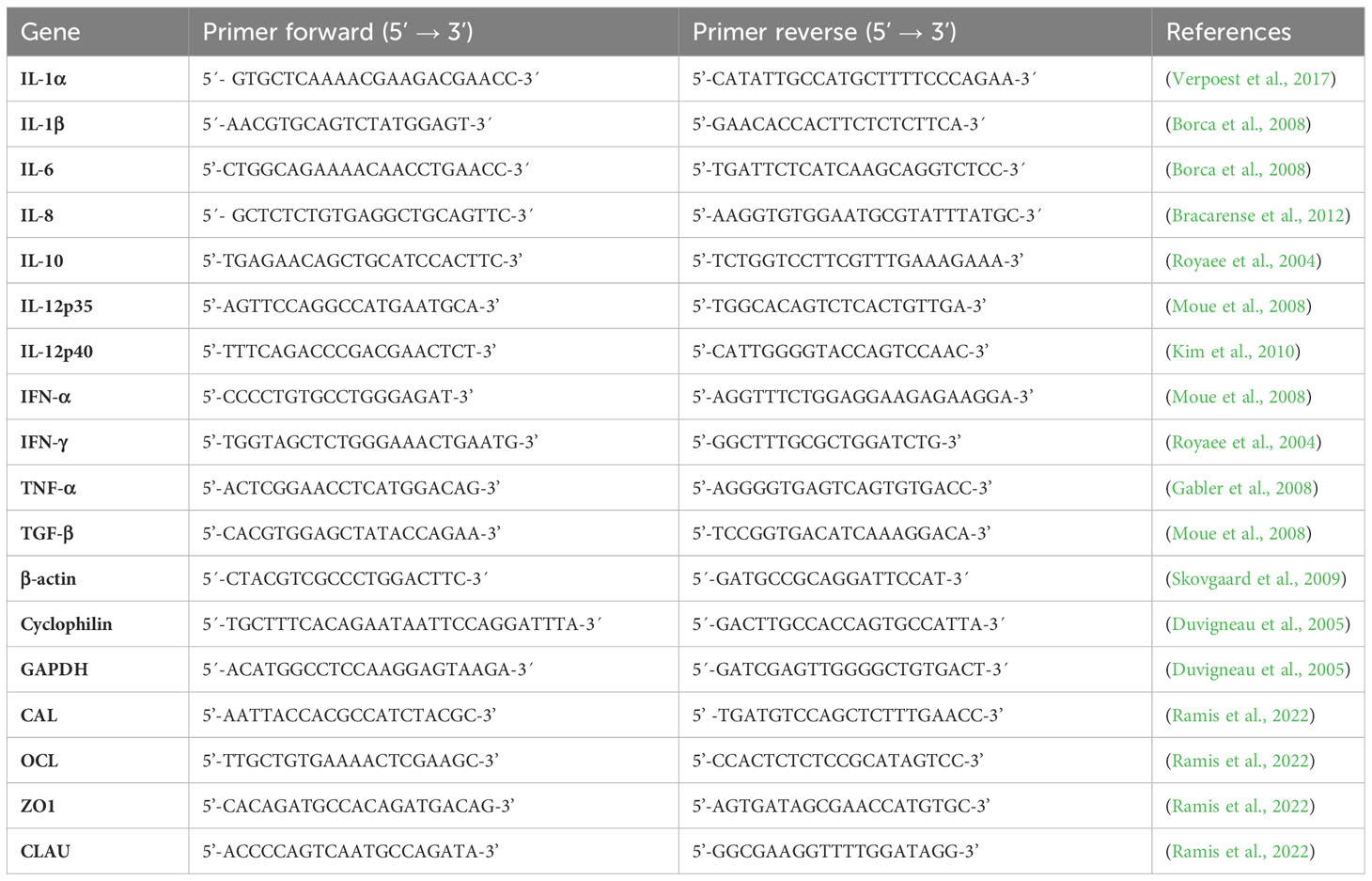
Table 2 Primers of the cytokines IFN-α, IFN-γ, IL-1α, IL-1β, IL-6, IL-8, IL-10, IL-12p35, IL-12p40, TNF-α and TGF-β and primers of glyceraldehyde-3-phosphatedehydrogenase (GAPDH), cyclophilin and β-actin and primers for tight junctions occludin (OCL), zonulin 1 (ZO1), claudin (CLAU) and Calprotectin (CAL).
Data were analysed using the MIXED procedure of SAS (version 9.4, SAS Institute; Cary, USA). The model included the fixed effects of treatment. Animal was the experimental unit for body weight and blood and tissue analyses. Statistical significance and tendency were considered at P < 0.05 and 0.05 < P < 0.10, respectively.
Principal component analysis (PCA, with multidimensional scaling and orthogonal transformation for rotation) and partial least squares regression (PLS) were used to analyse multivariates. The PCA was applied to a dataset including the variables tight junction protein mRNA gene expression and mRNA gene expression of 11 cytokines in jejunum and ileum for 3 different timepoints with time as nesting effects. The PLS analysis was set to assess: (1) variance of tight junction protein mRNA gene expression in jejunum and ileum explained by time and mRNA gene expression of 11 cytokines in jejunum and ileum.
PFX treatment increased BW of piglets on day 30 and 45 after weaning (P < 0.05; Table 3). Piglets had an average feed intake of 18.52 ± 2.3 kg per pen. Two animals died over the experiment, showing no digestive or respiratory related lesions at necropsy.
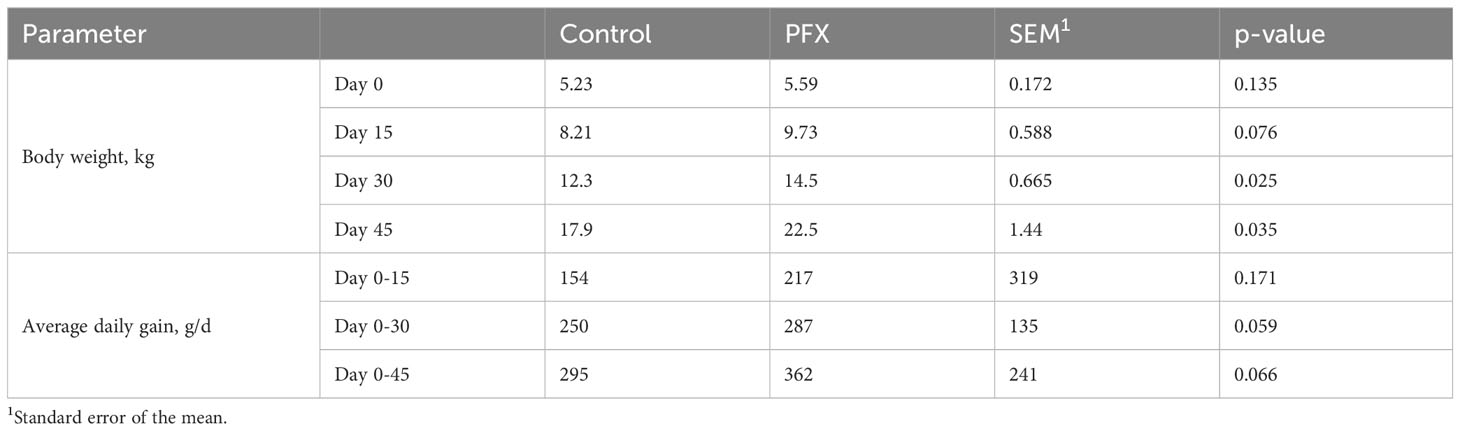
Table 3 Effect of treatment PFX on zootechnical performance parameters at days 15, 30, and 45 after weaning.
The length of villi and V/C ratio in ileal tissue was higher on day 15 for PFX compared to control (P < 0.05). On day 30 in jejunal tissue the villous length tended (P < 0.10) to be higher in PFX compared to control. In ileal tissue the both the villous length as well as the V/C ratio tended to be higher in PFX treatment (P < 0.10). On day 45 a higher villous length in ileal tissue (P < 0.05) was observed for PFX compared to control (Figure 1).
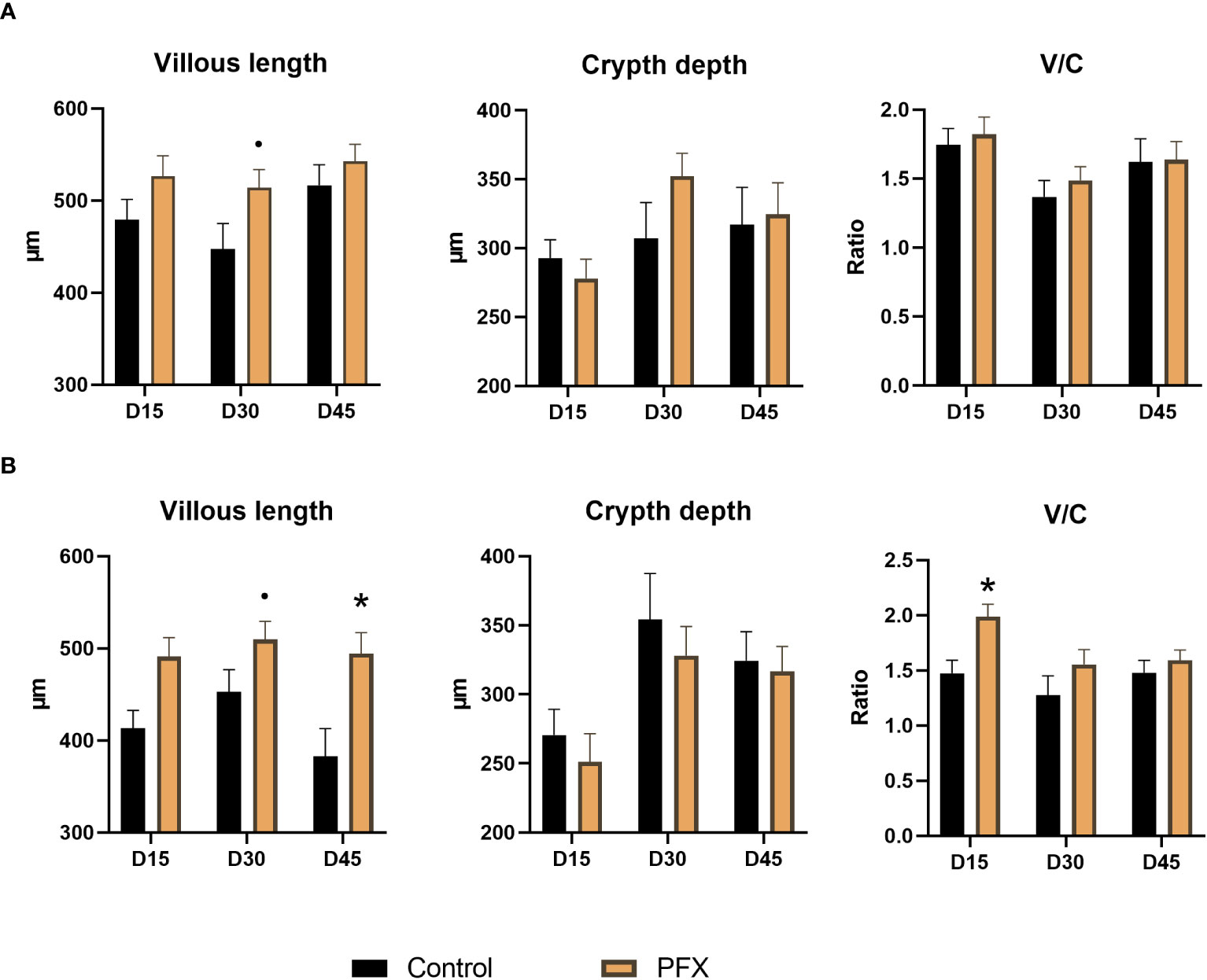
Figure 1 Effect of treatment PFX on intestinal histomorphometry in jejunum (A) and ileum (B) measurements in tissue of piglets at days 15, 30, and 45 after weaning, V/C means the ratio between villous length and crypt depth. Bars represent group mean, and whiskers depict standard deviation from the mean. (*) denotes significant difference (P < 0.05) and (·) denotes tendency (P < 0.10) of piglets receiving PFX compared to control.
On day 15 and 30, the gene expression of occludin in jejunal and ileal tissue was lower in PFX compared to control, as well as a lower gene expression of claudin in the ileum (P < 0.05). On day 45, a lower gene expression of occludin (P < 0.05) in ileal tissue for PFX compared to control was observed (Figure 2).
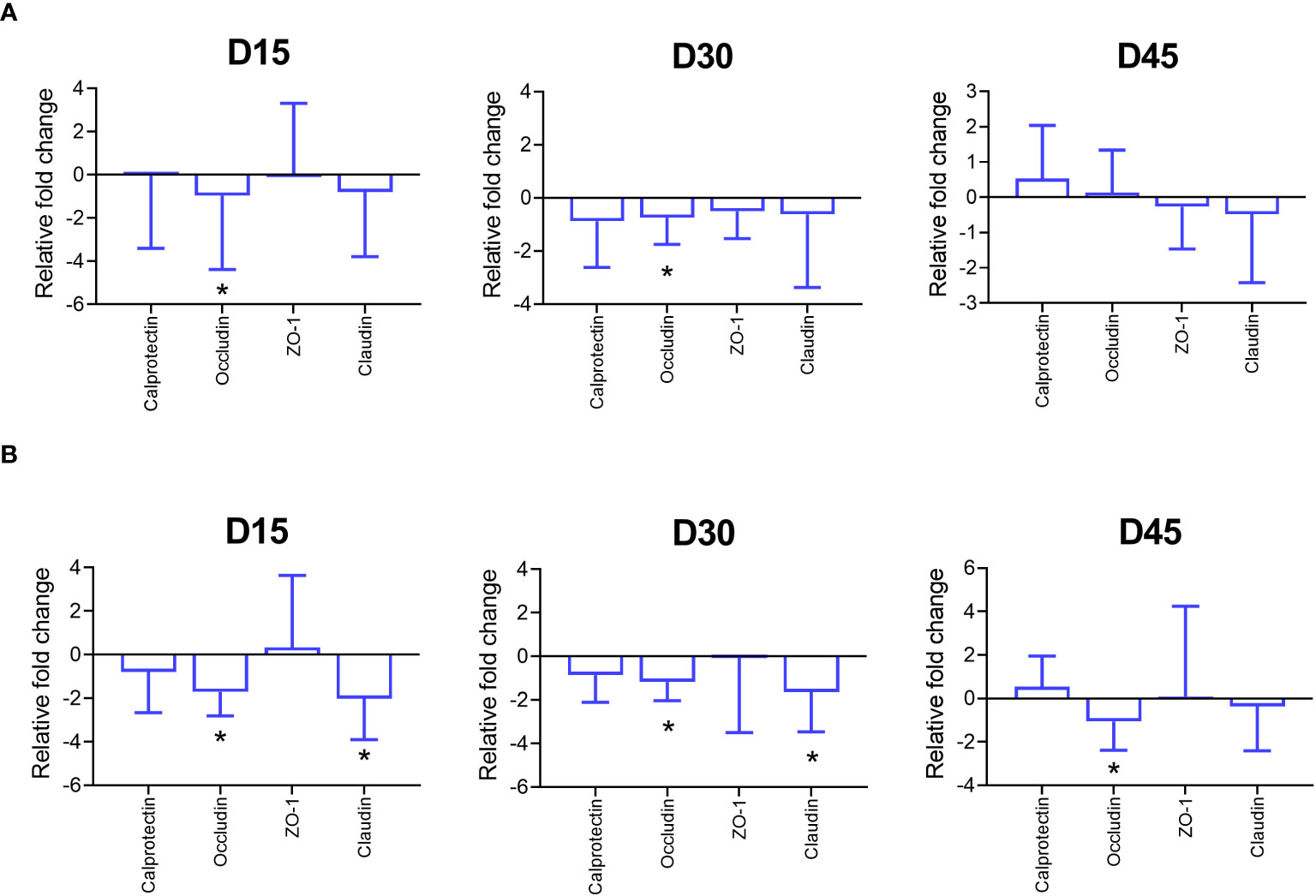
Figure 2 Effects of treatment PFX on tight junction protein expression (calprotectin, Occludin, Zonulin 1 (ZO-1) and Claudin) in tissues of piglets at days 15, 30, and 45 after weaning. Gene expression data is reported as fold change from PFX samples using the control as reference. Bars represent relative difference in upregulation or downregulation of gene expression of tight junction proteins; whiskers depict standard deviation from the mean. (*) denotes significant difference (P < 0.05) in jejunum (A) and ileum (B) of piglets receiving PFX compared to control.
On day 15 in jejunum, a higher gene expression was observed for IFN-α and TGF-β, with IFN-γ and IL-10 being lower in gene expression in PFX compared to control (P < 0.05). The expression of IL-1α and IL-8 tended to be higher for PFX compared to control (P < 0.01). In ileum tissue on day 15, expression of IL-1α, IL-6, IL-12p40 and TGF-β was increased and IFN-γ expression was lower in PFX compared to control (P < 0.05). In colon tissue, expression of IL-1α and TGF-β tended to be higher in PFX compared to control (P < 0.10). In PBMC on day 15, a significant higher expression of IFN-α, IL-12p35 and IL-12p40 was shown in PFX compared to control (P < 0.05). Furthermore, a tendency for higher expression were shown Il-6 in PFX treatment compared to control (P < 0.10; Figure 3).
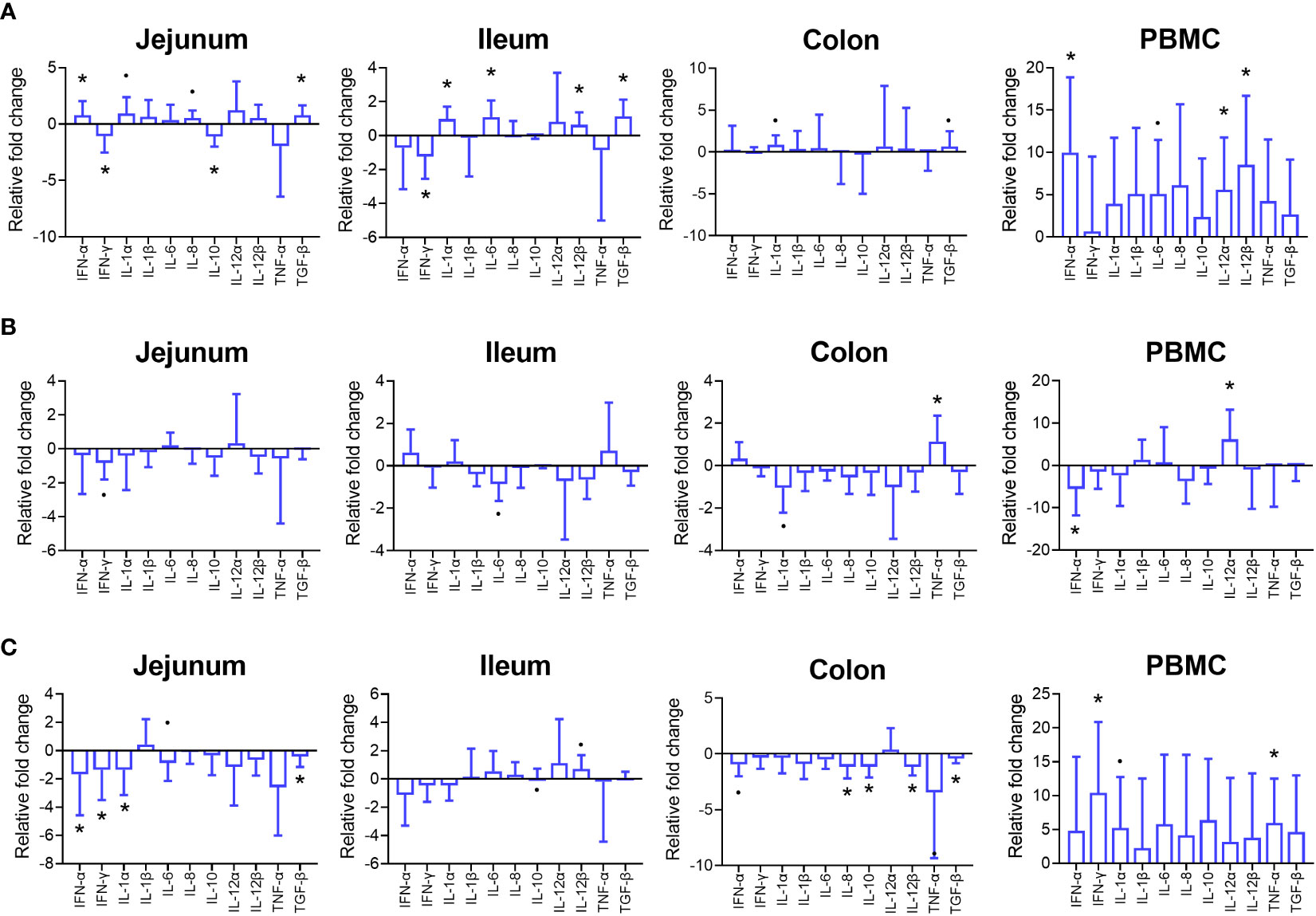
Figure 3 Effects of treatment PFX on cytokine expression in intestinal tissue and PBMC of piglets at days 15, 30, and 45 after weaning. Gene expression data is reported as fold change from PFX samples using the control as reference. Bars represent relative difference in upregulation or downregulation of gene expression cytokines; whiskers depict standard deviation from the mean. (*) denotes significant difference (P < 0.05) and (·) denotes tendency (P < 0.10) on day 15 (A), day 30 (B) and day 45 (C) of piglets receiving PFX compared to control.
On day 30, a tendency was observed for a lower expression (P < 0.10) expression of IFN-γ in the jejunum of piglets receiving PFX. In the ileum PFX tended to lower the expression of IL-6 (P < 0.10). In colon tissue the expression of TNF-α was higher in PFX treatment compared to control (P < 0.05), while IL-1α gene expression tended to be lower (P < 0.10). In PBMC, a lower expression for IFN-α and a higher expression for IL-12p35 was observed for PFX compared to control (P < 0.05).
On day 45, lower IFN-α, IFN-γ, IL-1α and TGF-β expression in the jejunum (P < 0.05) was observed, and a tendency for a higher expression of IL-6 (P < 0.10). In the ileum IL-10 and Il-12p40 expression tended to be higher (P < 0.10). In colon tissue a lower IL-8, IL-10, IL-12p40 and TGF-β expression (P < 0.05) and a tendency (P < 0.10) for a lower IFN-α and TNF-α expression for PFX compared to control was observed on day 45. Furthermore, in PBMC, IFN-γ and TNF-α expression was higher (P < 0.05) in PFX compared to control, with a tendency for a higher expression of IL-1α.
Principle component analysis (PCA) of gene expression of tight junction proteins and cytokines in jejunum and ileum tissue revealed a total variance of 70.6% explained by 2 factors of 36.6% and 34.0%, respectively (Figure 4). Based on the plot of scores and eigen vectors, Factor 1 (x axis), consisting of occludin, zonulin-1 and claudin in jejunum tissue and occludin in ileum, grouped together with IFN-γ, IL-12p35, IL-12p40, TNF-α, IL-10 and IL-6 in jejunum tissue. Within Principal Component Factor 2 (y axis), tight junction protein abundance of zonulin-1, claudin and occludin in ileum tissue, and zonulin-1 and occludin in jejunum tissue appear to be more central in the scatter plot. Cytokines IL-1α, IL-1β and IL-8 in jejunum and IL-1α and IL-1β in ileum tissue are more extreme, explaining more of the variance.
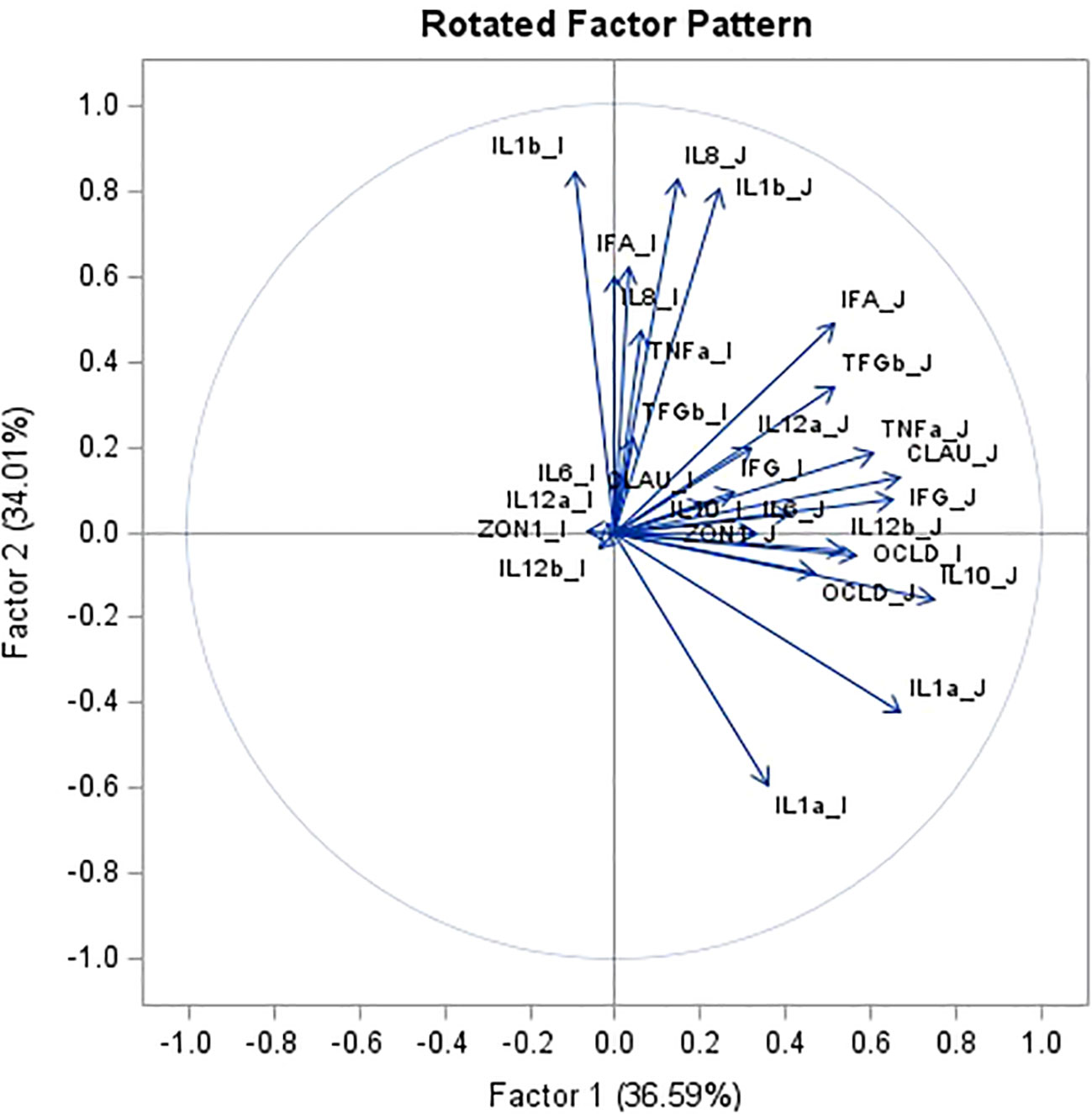
Figure 4 Principal component analysis for tight junction protein1 gene expression and cytokine2 gene expression in jejunum (J) and ileum (I) tissue. Data combines piglets at day 1 after weaning (n = 9), at 15 days after weaning (n = 10), at 30 days after weaning (n = 18) and at 45 days after weaning (n = 21) for both control as treatment group. 1mRNA expression (log2) of tight junction protein occludin (OCLD), claudin (CLAU) and zonulin-1 (ZON1). 2mRNA expression (log2) of 11 cytokines: interleukin IL-1α (IL1a), IL-1β (IL1b), IL-6 (IL6), IL-8 (IL8), IL-10 (IL10), IL-12p35 (IL12a), IL-12p40 (IL12b), tumor necrosis factor alpha (TNFa), interferon alpha and gamma (IFNa and IFNg), transforming growth factor beta (TGFb).
The most opposing components in Factor 1 (x axis) are IL-10 in jejunum, grouped with occludin, claudin, IL-12p40, IFN-γ and TNF-α in jejunum, contrasted against IL-6, IL-12p35, IL-12p40 and zonulin-1 in the ileum. In Factor 2 (y axis), IL-1α in jejunum and ileum grouped with occludin in jejunum and ileum and IL-10 in jejunum, contrasted against a group consisting of IL-8 in jejunum and ileum, IL-1β jejunum and ileum and IFN-α in ileum.
Partial Least Square analysis of expression of tight junction proteins and cytokine gene expression in jejunum and ileum tissue conducted reveals that occludin expression in jejunum and ileum accounts for the highest percentage of the variability in the 6 responses model (Figure 5). Tight junction protein expression is explained primarily by time, and by IL-10 in jejunum (x axis as Factor 1) and by IL-6 and IFN-α in jejunum (y axis as Factor 2), which are then organized as the scores in 4 concentric circles (clouds; Figure 5).
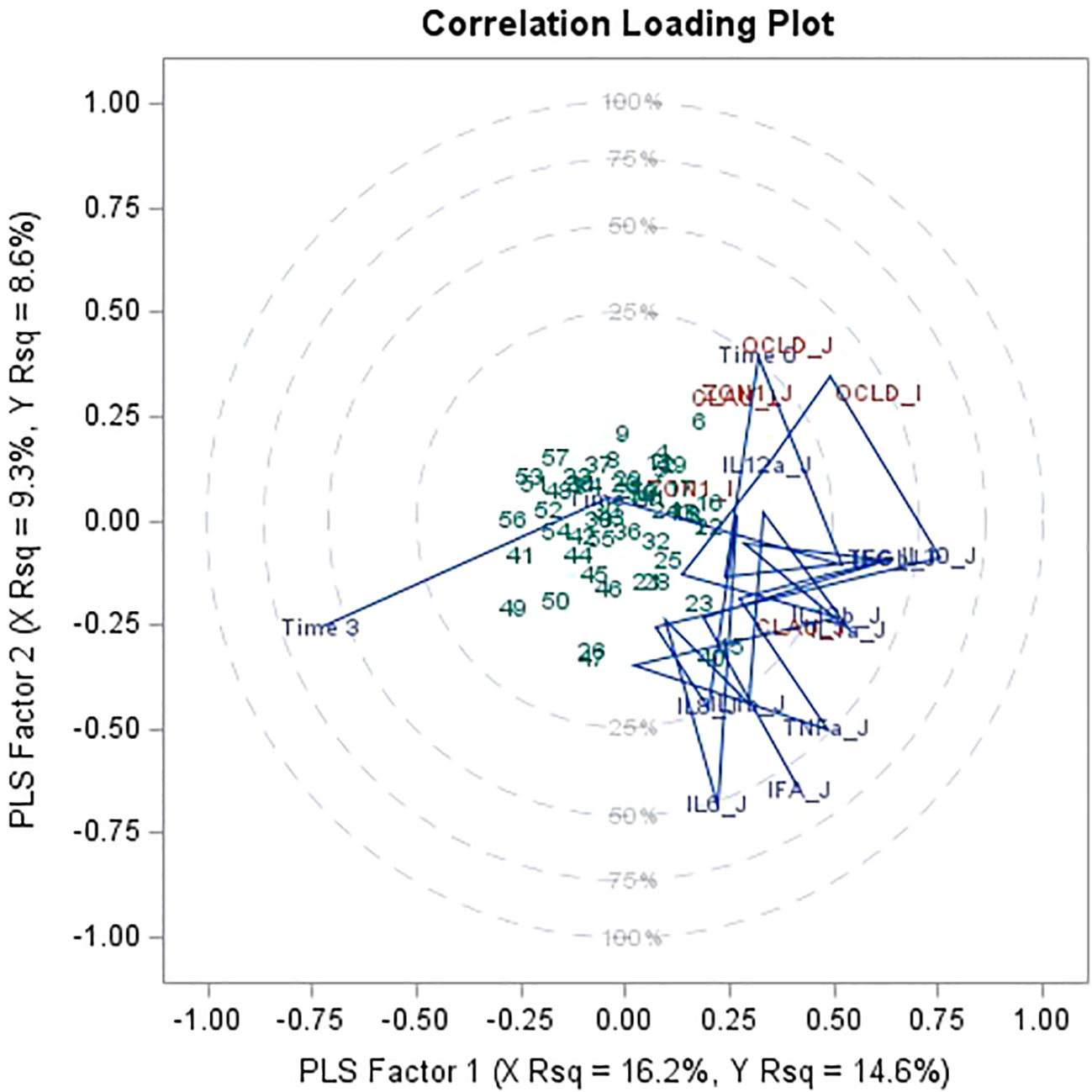
Figure 5 Partial least square regression (PLS) and two extracted factors for variance of tight junction protein1 gene expression in jejunum and ileum explained by time (time 0 = day 1 after weaning, time 1 = day 15 after weaning, time 2 = day 30 after weaning, time 3 = day 45 after weaning), mRNA expression of 11 immune biomarkers2 from jejunum and ileum. Data combines piglets at day 1 after weaning (n = 9), at 15 days after weaning (n = 20), at 30 days after weaning (n = 18) and at 45 days after weaning (n = 21). 1mRNA expression (log2) of tight junction proteins occludin (OCLD), claudin (CLAU) and zonulin-1 (ZON1). 2mRNA expression (log2) of 11 cytokines: interleukin IL-1α (IL1a), IL-1β (IL1b), IL-6 (IL6), IL-8 (IL8), IL-10 (IL10), IL-12p35 (IL12a), IL-12p40 (IL12b), tumor necrosis factor alpha (TNFa), interferon alpha and gamma (IFNa and IFNg), transforming growth factor beta (TGFb) for jejunum and ileum.
Immune parameter responses also partly explain tight junction gene expression variance and appear clustered by time, tissue, or the combination thereof. More specifically, IL-12p35 in jejunum clusters with occludin, zonulin-1 in jejunum and occludin and claudin in ileum, together with time 0 (1-day after weaning). IL-1α and IL-12p40 in jejunum cluster with claudin in jejunum.
Weaning is a stressful period in a piglet’s life which has been reported to impair the intestinal architecture and function, leading to gut-associated disorders and diarrhea (Wijtten et al., 2011; Hu et al., 2013; McLamb et al., 2013). Low feed intake immediately after weaning could be attributed as the main responsibility for gut morphological alterations such as villous atrophy (Wiese et al., 2003), resulting in a lower absorption of nutrients and reduced energy available for growth.
In the current study, the use of a blend of short and medium chain fatty acids, including butyrate, increased body weights of piglets on day 30 and 45 after weaning, and improved intestinal morphology, measured by villous length and v/c ratio, on day 15, 30 and 45. The villous height has a important effect on the intestinal structure, and may be reduced to 75% compared to pre-weaning structures within 24 h of weaning at 21 days of age (Hampson, 1986). Damage to the intestinal integrity of morphology has been reported after weaning, where the most serious damage to the intestinal barrier occurred from day 3 to 5 after weaning (Mei and Xu, 2005; Xiao et al., 2014). The intestinal mucosa is able to repair itself rapidly after damage (Sangild et al., 2000), with Hu et al. (2013) suggesting that villous height and crypt depth returned to pre-weaning values on day 14 after weaning. Studies can show an increase in cell proliferation to repair the damaged intestinal epithelium and others show a decrease in cell proliferation due to changes in cell expressions (Steeb et al., 1995; Yang et al., 2013). These divergent results may be due to the differences in the time of sample collection. Due to the lack of pre-weaning data on intestinal morphology in current study, it is not possible to show the effects of weaning on these parameters. However, data demonstrates that the use of the acid blend increased villous length in jejunum and ileum, along with an improved v/c ratio in the ileum, implying a potential increased nutrient absorption capacity of the GIT. It needs be taken in account that weaning is known to change the intestinal morphology, caused by the changes in feed intake pattern and a decrease in intake after weaning (Pluske et al., 1997; Moeser et al. 2007a), effects that are not studied separately in current study. The potential increased villous length could be caused by an increase in feed intake, however further research is needed to prove this hypothesis.
Several nutritional strategies have been adopted to improve gut health and maximize the production of weaned pigs (Lallès et al., 2004; Domeneghini et al., 2006; Liu et al., 2018) with different aims. One of them is to improve nutrient digestion and absorption, as well as optimizing intestinal integrity of weaned pigs and potentially alleviate the detrimental effects of weaning on GIT barrier integrity (Modina et al., 2019). The use of organic acids, which are broadly distributed in nature as elements of plant or animal tissue, is one of those nutritional strategies. Organic acids are also produced by the microbial fermentation of carbohydrates, predominantly in the large intestine of pigs. If used as animal feed supplements, with the right doses, they can contribute to improved growth performance and reduced colonization of pathogens in the intestine (Partanen and Mroz, 1999). Important to mention is that free acids are easily absorbed early on in the gastrointestinal tract, therefor certain protection mechanisms are used such as encapsulation or coatings to assure the target point of release is reached. The product used in this study contains butyric acids in the form of salts by binding them to sodium and calcium. Another example is the use of alkyl esters in MCFAs, which can reach lower parts of the gastrointestinal tract than free MCFAs or triglycerides because they need a different pH or a different enzyme activity to be broken down (Hedemann et al., 2003; Pereira et al., 2023).
In commercial practice, piglets are weaned early, which means that their intestine has limited digestive and absorption capacities, mainly caused by insufficient production of hydrochloric acid, pancreatic enzymes and therefore the sudden changes in the consistency and intake of feed is a major challenge (Cranwell, 1985). Therefore organic acids are frequently used after weaning in piglets (Suiryanrayna and Ramana, 2015). Besides the effect in the feed, there is direct or indirect action of organic acids on the mucosa of the GIT, mostly on the mucosa of the large intestine (Sakata, 1987; Sakata et al., 1995). Similar to current study, Long et al. (2018) observed an increase in villous height and the ratio of villous height to crypt depth in the jejunum and ileum in piglets fed PFX after weaning. Increased villous height in butyrate-supplemented weaned pigs have been observed in previous studies (Piva et al., 2002; Kotunia et al., 2004; Shen et al., 2009; Tonel et al., 2010). Mroz et al. (2000) reported that pigs fed with sodium butyrate showed proliferation of ileal microvilli cells as well as increases in the length of the microvilli, resulting in higher villous height, which is consistent with the effect in the present research.
Another example of a nutritional strategy is the use of other types of organic acids, being MCFAs, as they can improve growth performance in weaned piglets (Zentek et al., 2011) and have broad-spectrum antibacterial effects (Marounek et al., 2012). Previous studies have shown that a blend of MCFA and SCFA can be utilized by enterocytes as energy sources and can attenuate the negative effects of weaning on villous length and crypt depth in pigs (Lee et al., 2007), as well as improve growth performance (Zentek et al., 2011; Han et al., 2018; Li et al., 2018; Long et al., 2018). The effect of using a mixture of acids (as the blend used in this study) is generally better than that of a single acid due to synergism and their dissociation properties at different locations within the digestive tract (Huang et al., 2015). Fumaric acid, lactic acid, capric acid and caprylic acids didn’t prove to have an effect on the morphometry of jejunum in piglets (Ferrara et al., 2017). In the same study an increasing number of potential effector cells (principally CD3+ cytotoxic T lymphocytes) was found. Similar analyses were not performed in current study.
Similar to intestinal morphology results in current study, improvements in intestinal morphology were observed in piglets after a gastric infusion of SCFA (acetic, propionic and butyric acids). Coinciding with this reported intestinal morphology improvement, the SCFA infusion also increased the relative mRNA expression of claudin-1 in the jejunum, and occludin and claudin-1 in the duodenum and ileum (Diao et al., 2019). The positive effect on TJ protein expression was demonstrated by in vitro studies on Intestinal Porcine Epithelial Cell line-J2 (IPEC-J2) cultured in increasing doses of butyrate (Ma et al., 2012), reconfirmed in vivo with a general upregulation of occludin after butyrate administration, mainly in the small intestine. The same authors, however, observed that butyrate caused a down-regulation of claudin-1 in the small intestine of weaned pigs (Grilli et al., 2016). SCFA and especially butyrate and propionate are known to regulate cellular function and regulate the immune response by acting as histone deacetylase (HDAC) inhibitor (Zou et al., 2021). This has been shown by reducing cytokine production and NO after treatment with a HDAC inhibitor (Chang et al., 2014), similar results have been found in this study. Furthermore, the current study demonstrated a downregulation of gene expression of claudin 1 and occludin in the ileum and jejunum of piglets receiving a blend of SCFA and MCFA, including butyrate, while an improved intestinal morphology is observed at the same tissue locations. Some differences could be explained by a direct and indirect effect of organic acids on the expression of tight junctions and the gut barrier integrity. For example caprylic (C8), capric (C10), and lauric (C12) acids have been investigated as enhancers of drug absorption via the TJ pathway (Lindmark et al., 1995; Suzuki, 2020). This mechanism requires further study. In Caco-2 cells, C10 and C12 enhance TJ permeability by activating protein kinase C (PKC) and MLCK. The C10 effect also involves phospholipase C activation and ZO-1 redistribution. In current study no analyses were done in colon tissue, nor was the design of the study adequate to analyse effect of individual type of acids, therefore further study is necessary.
The GIT is acting as a large and important barrier between external environment and the organism. The permeability of the intestinal barrier is regulated by tight junctions (TJ), structures that are composed of membrane proteins such as Zona occludens (ZO), occludin and claudins (Edelblum and Turner, 2009; Groschwitz and Hogan, 2009; Turner, 2009; Marchiando et al., 2010b). These membrane proteins regulate the permeability of the epithelium through the “gate function”, selecting the epithelial ions and modulating pore size. Wang et al. (2016) found the mRNA abundances of occludin as well as occludin and ZO-1 protein in the jejunum and ileum decreased 3 and 5 days after weaning, indicating an impairment of the intestinal barrier. Similarly, Hu et al. (2013) and Xiao et al. (2014), demonstrated that TJ protein levels were decreased on day 3 and/or day 7 after weaning. Another example of disruption of intestinal cell tight junction proteins is the stress of heat and oxidative damage, resulting in increased permeability to luminal endotoxins (Zuhl et al., 2014). Wang et al. (2016) found lower abundances of different jejunal TJ proteins of weaned piglets than those in suckling piglets at 21 days of age.
The cytokine expression and its effects on the TJ barrier can result in immune activation and tissue inflammation, and subsequently important in the initiation and/or development of several intestinal and systemic diseases (Turner, 2009). Other growth factors might play a role in protection and maintenance of TJ integrity (Capaldo and Nusrat, 2009), such as interferon-γ (Bruewer et al., 2005), TNF-α (Ma et al., 2005; Schulzke et al., 2006; Mankertz et al., 2009), IL-1β (Dinarello, 1994; Al-Sadi and Ma, 2007; Al-Sadi et al., 2009), IL-6 (Suzuki et al, 2011). Its expression can impair or disrupt the TJ regulation, thereby increasing permeability, while IL-10 opposes the cellular functions induced by TNF-a and IFN-γ (Kucharzik et al., 2001; Sun et al., 2008), suggesting that IL-10 has a role in the protection of the intestinal barrier, similarly for TGF-ß (Howe et al., 2005). Interestingly, the current study demonstrated in piglets fed with PFX a downregulation of IFN-γ in jejunum tissue, during day 15, 30 and 45, a cytokine that is known to impair or disrupt the TJ regulation. This higher level of IFN-γ is coinciding with a lower level of IL-10 in jejunum on day 15, showing their interrelation. Furthermore TGF-ß expression has been increased in jejunum and ileum tissue on day 15 and 30, a cytokine with protective or promotive effects on intestinal barrier function. Interestingly, at the same time at day 15 and 30 a lower expression of occludin and claudin-1 is observed at the same timepoints in jejunum and ileum tissue.
Correlation analysis in current study demonstrated a clustering of tight junction gene expression in jejunum tissue with cytokines expression IFN-γ, IL-12p35, IL-12p40, TNF-α, IL-10 and IL-6 in jejunum tissue, while expression in ileum tissue clustered with IL-1α and IL-1β in jejunum and ileum tissue. IL-10, an anti-inflammatory cytokine and mentioned as a cytokine that opposes the cellular functions induced by TNF-a and IFN-γ seems one of the most contrasting cytokines and explaining most of the variation in the regression analyses. A possible explanation of the lower expression of tight junction genes in PFX could be explained by the effect of IL-10 in reducing the inflammation by opposing TNF-a and IFN-γ, thereby causing less damage to the tight junction proteins and lower gene expression of these proteins. However, this hypothesis needs more investigation. A clear limitation in this study is the lack of functional analysis before weaning to compare before and after, and also a shorter timeframe in post-weaning analysis. By day 15 the intestine could have recovered from the damage and differences can be more difficult to notice.
In post-weaning piglets, using a feed additive based on different types of organic acids can improve intestinal morphology, measured by villous length and v/c ratio, reduced expression of tight junction proteins and influenced cytokine production in the intestine, implicating these components can potentially reduce damage caused by weaning to the intestinal barrier and due to the higher absorption capacity of the villi potentially improve nutrient absorption. Further research is needed to confirm these findings.
The original contributions presented in the study are included in the article/supplementary files, further inquiries can be directed to the corresponding author/s.
The animal studies were approved by Bioethical Committee of the University of Murcia (CEEA-OH 465/2018). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
NdG: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. GR: Conceptualization, Supervision, Validation, Writing – review & editing. FF: Conceptualization, Investigation, Methodology, Writing – review & editing. CC-G: Methodology, Writing – review & editing. FP: Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Author NG was employed by the company Trouw Nutrition.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Al-Sadi R., Boivin M., Ma T. (2009). Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Bioscience 14 (7), 2765–2778. doi: 10.2741/3413
Al-Sadi R. M., Ma T. Y. (2007). IL-1β Causes an increase in intestinal epithelial tight junction permeability. J. Immunol. 178 (7), 4641–4649. doi: 10.4049/jimmunol.178.7.4641
Borca M. V., Gudmundsdottir I., Fernández-Sainz I. J., Holinka L. G., Risatti G. R., et al. (2008). Patterns of cellular gene expression in swine macrophages infected with highly virulent classical swine fever virus strain Brescia. Virus Res. 138 (1–2), 89–96. doi: 10.1016/j.virusres.2008.08.009
Bracarense A.-P. F. L., Lucioli J., Grenier B., Drociunas Pacheco G., Moll W.-D., Schatzmayr G., et al. (2012). Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 107 (12), 1776–1786. doi: 10.1017/S0007114511004946
Bruewer M., et al. (2005). Interferon-γ induces internalization of epithelial tight junction proteins via a macropinocytosis-like process. FASEB J. 19 (8), 923–933. doi: 10.1096/fj.04-3260com
Capaldo C. T., Nusrat A. (2009). “Cytokine regulation of tight junctions,” in Biochimica et biophysica acta - biomembranes 1788 (4), 864–871. doi: 10.1016/j.bbamem.2008.08.027
Chang P. V., Hao L., Offermanns S., Medzhitov R. (2014). The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. 111 (6), 2247–2252. doi: 10.1073/pnas.1322269111
Cranwell P. D. (1985). The development of acid and pepsin (EC 3. 4. 23. 1) secretory capacity in the pig; the effects of age and weaning: 1. Studies in anaesthetized pigs. British J. Nutrit. 54 (1), 305-320. doi: 10.1079/BJN19850113
de Groot N., Fariñas F., Cabrera-Gómez C. G., Pallares F. J., Ramis G. (2021). Weaning causes a prolonged but transient change in immune gene expression in the intestine of piglets. J. Anim. Sci. 99 (4). doi: 10.1093/jas/skab065
Diao H., Jiao A. R., Yu B., Mao B., Chen D. W. (2019). Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 14 (1), 4. doi: 10.1186/s12263-019-0626-x
Dinarello C. A. (1994). The interleukin-1 family: 10 years of discovery. FASEB journal : Off. Publ. Fed. Am. Societies Exp. Biol. 8 (15), 1314–1325.
Domeneghini C., Di Giancamillo A., Arrighi S., Bosi G. (2006). Gut-trophic feed additives and their effects upon the gut structure and intestinal metabolism. State of the art in the pig, and perspectives towards humans. Histol. histopathology 21 (3), 273–283. doi: 10.14670/HH-21.273
Duvigneau J. C., Hartl R. T., Groiss S., Gemeiner M. (2005). Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods 306 (1–2), 16–27. doi: 10.1016/j.jim.2005.06.021
Eckmann L. (1995). Intestinal epithelial cells as watchdogs for the natural immune system. Trends Microbiol. 3 (3), 118–120. doi: 10.1016/S0966-842X(00)88894-0
Edelblum K. L., Turner J. R. (2009). The tight junction in inflammatory disease: communication breakdown. Curr. Opin. Pharmacol. 9 (6), 715–720. doi: 10.1016/j.coph.2009.06.022
Ferrara F., Tedin L., Pieper R., Meyer W., Zentek J. (2017). Influence of medium-chain fatty acids and short-chain organic acids on jejunal morphology and intra-epithelial immune cells in weaned piglets. J. Anim. Physiol. Anim. Nutr. 101 (3), 531–540. doi: 10.1111/jpn.12490
Gabler N. K., Spencer J. D., Webel D. W., Spurlock M. E. (2008). n-3 PUFA attenuate lipopolysaccharide-induced down-regulation of toll-like receptor 4 expression in porcine adipose tissue but does not alter the expression of other immune modulators. J. Nutr. Biochem. 19 (1), 8–15. doi: 10.1016/j.jnutbio.2006.11.014
Grilli E., Tugnoli B., Foerster C. J., Piva A. (2016). Butyrate modulates inflammatory cytokines and tight junctions components along the gut of weaned pigs. J. Anim. Sci. 94 (suppl_3), 433–436. doi: 10.2527/jas.2015-9787
Groschwitz K. R., Hogan S. P. (2009). Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 124 (1), 3–20. doi: 10.1016/j.jaci.2009.05.038
Hampson D. J. (1986). Alterations in piglet small intestinal structure at weaning. Res. Veterinary Sci. 40 (1), 32–40. doi: 10.1016/S0034-5288(18)30482-X
Han Y. S., Tang C. H., Zhao Q. Y., Zhan T. F., Zhang K., Han Y. M., et al. (2018). “Effects of dietary supplementation with combinations of organic and medium chain fatty acids as replacements for chlortetracycline on growth performance, serum immunity, and fecal microbiota of weaned piglets,” in Livestock science., vol. 216m 210–218. doi: 10.1016/j.livsci.2018.08.013
Hedemann M. S., Hojsgaard S., Jensen B. B. (2003). Small intestinal morphology and activity of intestinal peptidases in piglets around weaning. J. Anim. Physiol. Anim. Nutr. 87 (1–2), 32–41. doi: 10.1046/j.1439-0396.2003.00405.x
Howe K. L., Reardon C., Wang A., Nazli A., McKay D. M. (2005). Transforming growth factor-β Regulation of epithelial tight junction proteins enhances barrier function and blocks enterohemorrhagic escherichia coli O157:H7-induced increased permeability. Am. J. Pathol. 167 (6), 1587–1597. doi: 10.1016/S0002-9440(10)61243-6
Hu C. H., Xiao K., Luan Z. S., Song J. (2013). Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs1. J. Anim. Sci. 91 (3), 1094–1101. doi: 10.2527/jas.2012-5796
Huang C., Song P., Fan P., Hou C., Thacker P., Ma X. (2015). Dietary sodium butyrate decreases postweaning diarrhea by modulating intestinal permeability and changing the bacterial communities in weaned piglets. J. Nutr. 145 (12), 2774–2780. doi: 10.3945/jn.115.217406
Kim C. J., Kovacs-Nolan J. A., Yang C., Archbold T., Fan M. Z., Mine Y. (2010). l-Tryptophan exhibits therapeutic function in a porcine model of dextran sodium sulfate (DSS)-induced colitis. J. Nutr. Biochem. 21 (6), 468–475. doi: 10.1016/j.jnutbio.2009.01.019
Kotunia A., Woliński J., Laubitz D., Jurkowska M., Romé V., Guilloteau P., et al. (2004). Effect of sodium butyrate on the small intestine development in neonatal piglets fed [correction of feed] by artificial sow. J. Physiol. Pharmacol. 55 Suppl 2, 59–68.
Kucharzik T., Lügering N., Pauels H. G., Domschke W., Stoll R., et al. (2001). IL-4, IL-10 and IL-13 down-regulate monocyte-chemoattracting protein-1 (MCP-1) production in activated intestinal epithelial cells. Clin. Exp. Immunol. 111 (1), 152–157. doi: 10.1046/j.1365-2249.1998.00481.x
Lallès J.-P., Boudry G., Favier C., Le Floc’h N., Luron I., Montagne L., et al. (2004). Gut function and dysfunction in young pigs: physiology. Anim. Res. 53 (4), 301–316. doi: 10.1051/animres:2004018
Lee D. N., Liu S. R., Chen Y. T., Wang R. C., Lin S. Y., Weng C. F., et al. (2007). Effects of diets supplemented with organic acids and nucleotides on growth, immune responses and digestive tract development in weaned pigs. J. Anim. Physiol. Anim. Nutr. 91 (11–12), 508–518. doi: 10.1111/j.1439-0396.2007.00684.x
Li S., Zheng J., Deng K., Chen L., Zhao X. L., Jiang X., et al. (2018). Supplementation with organic acids showing different effects on growth performance, gut morphology, and microbiota of weaned pigs fed with highly or less digestible diets. J. Anim. Sci. 96 (8), 3302–3318. doi: 10.1093/jas/sky197
Lindmark T., Nikkilä T., Artursson P. (1995). Mechanisms of absorption enhancement by medium chain fatty acids in intestinal epithelial Caco-2 cell monolayers. J. Pharmacol. Exp. Ther. 275 (2), 958–964.
Liu Y., Espinosa C. D., Abelilla J. J., Casas G. A., Lagos L. V., Lee S. A., et al. (2018). Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 4 (2), 113–125. doi: 10.1016/j.aninu.2018.01.007
Long S. F., Xu Y. T., Pan L., Wang Q. Q., Wang C. L., Wu J. Y., et al. (2018). Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets. Anim. Feed Sci. Technol. 235 (April 2017), 23–32. doi: 10.1016/j.anifeedsci.2017.08.018
Ma T. Y., Boivin M. A., Ye D., Pedram A., Said H. M. (2005). Mechanism of TNF-α modulation of Caco-2 intestinal epithelial tight junction barrier: role of myosin light-chain kinase protein expression. Am. J. Physiology-Gastrointestinal Liver Physiol. 288 (3), G422–G430. doi: 10.1152/ajpgi.00412.2004
Ma X., Fan P. X., Li L. S., Qiao S. Y., Zhang G. L., Li D. F. (2012). Butyrate promotes the recovering of intestinal wound healing through its positive effect on the tight junctions1. J. Anim. Sci. 90 (suppl_4), 266–268. doi: 10.2527/jas.50965
Mankertz J., Amasheh M., Krug Fromm S. M. A., Amasheh S., Hillenbrand B., et al. (2009). TNFα up-regulates claudin-2 expression in epithelial HT-29/B6 cells via phosphatidylinositol-3-kinase signaling. Cell Tissue Res. 336 (1), 67–77. doi: 10.1007/s00441-009-0751-8
Marchiando A. M., Graham W. V., Turner J. R. (2010a). Epithelial barriers in homeostasis and disease. Annu. Rev. Pathology: Mech. Dis. 5 (1), 119–144. doi: 10.1146/annurev.pathol.4.110807.092135
Marchiando A. M., Shen L., Graham W. V., Weber C. R., Schwarz B. T., Austin J. R., et al. (2010b). Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. J. Cell Biol. 189 (1), 111–126. doi: 10.1083/jcb.200902153
Marounek M., Putthana V., Benada O., Lukešová D. (2012). Antimicrobial activities of medium-chain fatty acids and monoacylglycerols on Cronobacter sakazakii DBM 3157T and Cronobacter malonaticus DBM 3148. Czech J. Food Sci. 30 (6), 573–580. doi: 10.17221/433/2011-CJFS
McLamb B. L., Gibson A. J., Overman E. L., Stahl C., Moeser A. J. (2013). Early weaning stress in pigs impairs innate mucosal immune responses to enterotoxigenic E. coli challenge and exacerbates intestinal injury and clinical disease. PloS One 8 (4), e59838. doi: 10.1371/journal.pone.0059838
Mei J., Xu R.-J. (2005). Transient changes of transforming growth factor-β expression in the small intestine of the pig in association with weaning. Br. J. Nutr. 93 (1), 37–45. doi: 10.1079/bjn20041302
Modina S. C., Polito U., Rossi R., Corino C., Di Giancamillo A. (2019). Nutritional regulation of gut barrier integrity in weaning piglets. Animals 9 (12), 1–15. doi: 10.3390/ani9121045
Moeser A. J., Vander Klok C., Ryan K. A., Wooten J. G., Little D., Cook V. L., et al. (2007b). Stress signaling pathways activated by weaning mediate intestinal dysfunction in the pig. Am. J. Physiol. - Gastrointestinal Liver Physiol. 292 (1), 173–181. doi: 10.1152/ajpgi.00197.2006
Moeser A. J., Ryan K. A., Nighot P. K., Blikslager A. T. (2007a). Gastrointestinal dysfunction induced by early weaning is attenuated by delayed weaning and mast cell blockade in pigs. Am. J. Physiology-Gastrointestinal Liver Physiol. 293 (2), G413–G421. doi: 10.1152/ajpgi.00304.2006
Moue M., Tohno M., Shimazu T., Kido T., Aso H., Saito T., et al. (2008). Toll-like receptor 4 and cytokine expression involved in functional immune response in an originally established porcine intestinal epitheliocyte cell line. Biochim. Biophys. Acta (BBA) - Gen. Subj. 1780 (2), 134–144. doi: 10.1016/j.bbagen.2007.11.006
Mroz Z., Jongbloed A. W., Partanen K. H., Vreman K., Kemme P. A., Kogut J., et al. (2000). The effects of calcium benzoate in diets with or without organic acids on dietary buffering capacity, apparent digestibility, retention of nutrients, and manure characteristics in swine. J. Anim. Sci. 78 (10), 2622. doi: 10.2527/2000.78102622x
Niewold T. (2015). Intestinal Health: Key to Maximise Growth Perform. Livestock (Leiden, The Netherlands: Wageningen Academic), 1–276. doi: 10.3920/978-90-8686-792-9
Partanen K. H., Mroz Z. (1999). Organic acids for performance enhancement in pig diets. Nutr. Res. Rev. 12 (1), 117–145. doi: 10.1079/095442299108728884
Pereira E., Fernandes J.-M., Gonçalves R., Pinheiro A. C., Duarte M. S., Alves M. M., et al. (2023). Evaluating the in vitro digestion of lipids rich in medium-chain fatty acids (MCFAs) using dynamic and static protocols. Food Chem. 406, 135080. doi: 10.1016/j.foodchem.2022.135080
Pié S., Lallès J. P., Blazy F., Laffitte J., Sève B., Oswald I. P., et al. (2004). Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 134 (3), 641–647. doi: 10.1093/jn/134.3.641
Pitman R. S., Blumberg R. S. (2000). First line of defense: the role of the intestinal epithelium as an active component of the mucosal immune system. J. Gastroenterol. 35 (11), 805–814. doi: 10.1007/s005350070017
Piva A., Morlacchini M., Casadei G., Gatta P. P., Biagi G., Prandini A., et al. (2002). Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Ital. J. Anim. Sci. 1 (1), 35–41. doi: 10.4081/ijas.2002.35
Pluske J. R., Hampson D. J., Williams I. H. (1997). Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livestock Production Sci. 51 (1–3), 215–236. doi: 10.1016/S0301-6226(97)00057-2
Ramis G., Pérez-Esteruelas L., Gómez-Cabrera C. G., de Pascual-Monreal C., Gonzalez-Guijarro B., Párraga-Ros E., et al. (2022). Oral and Parenteral Vaccination against Escherichia coli in Piglets Results in Different Responses. Animals 12 (20), 2758. doi: 10.3390/ani12202758
Royaee A. R., Husmann R. J., Dawson H. D., Calzada-Nova G., Schnitzlein W. M., Zuckermann F. A., et al. (2004). Deciphering the involvement of innate immune factors in the development of the host response to PRRSV vaccination. Veterinary Immunol. Immunopathology 102 (3), 199–216. doi: 10.1016/j.vetimm.2004.09.018
Sakata T. (1987). Stimulatory effect of short-chain fatty acids on epithelial cell proliferation in the rat intestine: a possible explanation for trophic effects of fermentable fibre, gut microbes and luminal trophic factors. Br. J. Nutr. 58 (1), 95–103. doi: 10.1079/BJN19870073
Sakata T., Adachi M., Hashida M., Sato N., Kojima T. (1995). Effect of n-butyric acid on epithelial cell proliferation of pig colonic mucosa in short-term culture. Dtsch Tierarztl Wochenschr. 102 (4), 163–164.
Sangild P., Fowden A., Trahair J. (2000). How does the foetal gastrointestinal tract develop in preparation for enteral nutrition after birth? Livestock Production Sci. 66 (2), 141–150. doi: 10.1016/S0301-6226(00)00221-9
Schulzke J.-D., Bojarski C., Zeissig S., Heller F., Gitter A. H., Fromm M. (2006). Disrupted barrier function through epithelial cell apoptosis. Ann. New York Acad. Sci. 1072 (1), 288–299. doi: 10.1196/annals.1326.027
Shen Y. B., Piao X. S., Kim S. W., Wang L., Liu P., Yoon I., et al. (2009). Effects of yeast culture supplementation on growth performance, intestinal health, and immune response of nursery pigs1. J. Anim. Sci. 87 (8), 2614–2624. doi: 10.2527/jas.2008-1512
Skovgaard K., Mortensen S., Boye M., Poulsen K. T., Campbell F. M., Eckersall P. D., et al. (2009). Rapid and widely disseminated acute phase protein response after experimental bacterial infection of pigs. Veterinary Res. 40 (3), 23. doi: 10.1051/vetres/2009006
Smith F., Clark J. E., Overman B. L., Tozel C. C., Huang J. H., Rivier, et al. (2010). Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. - Gastrointestinal Liver Physiol. 298 (3), 352–363. doi: 10.1152/ajpgi.00081.2009
Steeb C. B., Trahair J. F., Read L. C. (1995). Administration of insulin-like growth factor-I (IGF-I) peptides for three days stimulates proliferation of the small intestinal epithelium in rats. Gut 37 (5), 630–638. doi: 10.1136/gut.37.5.630
Suiryanrayna M. V. A. N., Ramana J. V. (2015). A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. J. Anim. Sci. Biotechnol. 6 (1), 1–11. doi: 10.1186/s40104-015-0042-z
Sun X., Yang H., Nose K., Nose S., Haxhija E. Q., Koga H., et al. (2008). Decline in intestinal mucosal IL-10 expression and decreased intestinal barrier function in a mouse model of total parenteral nutrition’. Am. J. Physiology-Gastrointestinal Liver Physiol. 294 (1), G139–G147. doi: 10.1152/ajpgi.00386.2007
Suzuki T. (2020). Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 91, e13357. doi: 10.1111/asj.13357
Suzuki T., Yoshinaga N., Tanabe S. (2011). Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 286 (36), 31263–31271. doi: 10.1074/jbc.M111.238147
Tonel I., Pinho M., Lordelo M. M., Cunha L. F., Garres P., Freire J. P. B., et al. (2010). Effect of butyrate on gut development and intestinal mucosa morphology of piglets. Livestock Sci. 133 (1–3), 222–224. doi: 10.1016/j.livsci.2010.06.069
Turner J. R. (2009). “Intestinal mucosal barrier function in health and disease,” in Nature reviews immunology, vol. 9, 799–809. doi: 10.1038/nri2653
van Beers-Schreurs H. M. G., Nabuurs M. J., Vellenga L., Kalsbeek-van der Valk H. J., Wensing T., Breukink H. J. (1998). Weaning and the weanling diet influence the villous height and crypt depth in the small intestine of pigs and alter the concentrations of short-chain fatty acids in the large intestine and blood. J. Nutr. 128 (6), 947–953. doi: 10.1093/jn/128.6.947
van der Peet-Schwering C. M. C., Jansman A. J., Smidt H., Yoon I. (2007). Effects of yeast culture on performance, gut integrity, and blood cell composition of weanling pigs1,2. J. Anim. Sci. 85 (11), 3099–3109. doi: 10.2527/jas.2007-0110
Vente-Spreeuwenberg M. A. M., Verdonk J. M. A J., Verstegen M. W. A., Beynen A. C. (2003). Villus height and gut development in weaned piglets receiving diets containing either glucose, lactose or starch. Br. J. Nutr. TNO Defence Secur. Saf. 90 (5), 907–913. doi: 10.1079/bjn2003981
Verpoest S., Cay B., Favoreel H., De Regge N. (2017). Age-dependent differences in pseudorabies virus neuropathogenesis and associated cytokine expression. J. Virol. 91 (2), 1–17. doi: 10.1128/jvi.02058-16
Wang D., Piao X. S., Zeng Z. K., Lu T., Zhang Q., Li P. F., et al. (2011). Effects of keratinase on performance, nutrient utilization, intestinal morphology, intestinal ecology and inflammatory response of weaned piglets fed diets with different levels of crude protein. Asian-Australasian J. Anim. Sci. 24 (12), 1718–1728. doi: 10.5713/ajas.2011.11132
Wang J., Zeng L., Tan B., Li G., Huang B., Xiong X., et al. (2016). Developmental changes in intercellular junctions and Kv channels in the intestine of piglets during the suckling and post-weaning periods. J. Anim. Sci. Biotechnol. J. Anim. Sci. Biotechnol. 7 (1), 1–10. doi: 10.1186/s40104-016-0063-2
Wiese F., Simon O., Weyrauch K. D. (2003). Morphology of the small intestine of weaned piglets and a novel method for morphometric evaluation+. Anatomia Histologia Embryologia: J. Veterinary Med. Ser. C 32 (2), 102–109. doi: 10.1046/j.1439-0264.2003.00430.x
Wijtten P. J. A., van der Meulen J., Verstegen M. W. A. (2011). Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 105 (7), 967–981. doi: 10.1017/S0007114510005660
Worobec E., Duncan I. J., Widowski T. (1999). The effects of weaning at 7, 14 and 28 days on piglet behaviour. Appl. Anim. Behav. Sci. 62 (2–3), 173–182. doi: 10.1016/S0168-1591(98)00225-1
Xiao K., Song Z. H., Jiao L. F., Ke Y. L., Hu C. H. (2014). Developmental changes of TGF-β1 and smads signaling pathway in intestinal adaption of weaned pigs’. PloS One 9 (8), 1–7. doi: 10.1371/journal.pone.0104589
Yang H., Xiong X., Yin Y. (2013). “Development and renewal of intestinal villi in pigs,” in Nutritional and physiological functions of amino acids in pigs (Vienna: Springer Vienna), 29–47. doi: 10.1007/978-3-7091-1328-8_3
Zentek J., Buchheit-Renko S., Ferrara F., Vahjen W., Van Kessel A. G., Pieper R., et al. (2011). Nutritional and physiological role of medium-chain triglycerides and medium-chain fatty acids in piglets. Anim. Health Res. Rev. / Conf. Res. Workers Anim. Dis. 12 (1), 83–93. doi: 10.1017/S1466252311000089
Zou F., Qiu Y., Huang Y., Zou H., Cheng X., Niu Q., et al. (2021). Effects of short-chain fatty acids in inhibiting HDAC and activating p38 MAPK are critical for promoting B10 cell generation and function. Cell Death Dis. 12 (6), 582. doi: 10.1038/s41419-021-03880-9
Keywords: piglets, gut health, morphology, post-weaning, cytokines, immune response
Citation: de Groot N, Fariñas F, Cabrera-Gómez CG, Pallares FJ and Ramis G (2024) Blend of organic acids improves gut morphology and affects inflammation response in piglets after weaning. Front. Anim. Sci. 5:1308514. doi: 10.3389/fanim.2024.1308514
Received: 06 October 2023; Accepted: 09 January 2024;
Published: 05 February 2024.
Edited by:
Carla Maris Machado Bittar, University of São Paulo, BrazilReviewed by:
Marcos Elias Duarte, North Carolina State University, United StatesCopyright © 2024 de Groot, Fariñas, Cabrera-Gómez, Pallares and Ramis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nienke de Groot, bmllbmtlZGdyb290QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.