
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Anim. Sci., 27 June 2022
Sec. Animal Physiology and Management
Volume 3 - 2022 | https://doi.org/10.3389/fanim.2022.871433
This article is part of the Research TopicBiology Meets Technology: Aquatic Animals in Novel and New Aquaculture Production SystemsView all 6 articles
 Carlo C. Lazado1†
Carlo C. Lazado1† Kevin T. Stiller2†
Kevin T. Stiller2† Khurram Shahzad2Britt Kristin M. Reiten2Yuriy Marchenko2Jascha Gerwins2Filip Strand Radonjic2Bernhard Eckel2Arne Berge3
Khurram Shahzad2Britt Kristin M. Reiten2Yuriy Marchenko2Jascha Gerwins2Filip Strand Radonjic2Bernhard Eckel2Arne Berge3 Åsa Maria Espmark2*
Åsa Maria Espmark2*Closed-containment (CCS) systems offer several advantages in controlling the production environment for Atlantic salmon (Salmo salar) aquaculture, especially at sea, where fish are more exposed to challenging environmental conditions. Here, we report the health and welfare of Atlantic salmon in FishGLOBE V5, a 3500 m3 semi-closed containment system. A group of 200,000 post-smolts was followed from a recirculating aquaculture system (RAS) facility, then for three and a half months in FishGLOBE V5, and three months after release in net pens. Fish samples were collected at four time points during the production period, while water quality was evaluated when the fish were in FishGLOBE V5. The water quality in FishGLOBE V5 was within the recommended range for salmon post-smolts. The mortality rate was 1.4% in FishGLOBE V5 and 4.9% in net pens. There was an increase in the prevalence of eye, snout and dorsal fin damages before the fish left the FishGLOBE V5 phase. The plasma level of magnesium was slightly elevated during this phase as well, indicating osmotic imbalance and stress. Histological evaluation of skin and gills showed sporadic cases of non-specific pathologies. In particular, the transitory stay in FishGLOBE V5 partially improved skin health, but not gill health. Gene expression profiling of some biomarkers showed that rearing fish in FishGLOBE V5 could influence the expression of genes involved in stress response, mucus production, and epithelial integrity. This study documented that rearing salmon for a certain period in FishGLOBE V5 during production affected different health and welfare indicators. These changes should be taken into consideration for the improvement of FishGLOBE V5 as a viable technology for post-smolt production at sea.
In 2019, Norway exported 1.4 million tons of farmed Atlantic salmon and the government has the vision to significantly increase the production in the years to come in a sustainable manner (Ministry of Trade, I.a.F., 2021). This ambitious production target is dependent on how the industry battles major challenges including sea lice (Lepeophtheirus salmonis) and escapees from the farms, both of which have serious economic and societal consequences. In recent years, many salmon aquaculture stakeholders have argued that the increased production must be achieved through the use of additional sustainable production technologies as alternatives to the traditional net pens that are mainly used today. New production technologies must be proven to support optimal conditions for fish health and welfare, and environmental functionality including stringent protection from lice and possibilities for escapees. Floating semi-closed containment systems (S-CCS) are an example of such sustainable systems, as they are proven to be efficient against lice and escapees (Nilsen et al., 2017; Nilsen et al., 2020). Floating S-CCS are aquaculture production systems in which a physical barrier keeps the lice out of the system and prevents the fish from escaping. Water is pumped into the system through pipes, with the inlet located below the level where sea lice normally thrive. Sea lice are normally found in the surface waters, between 1-6 m (Nelson et al., 2018; Fernandez-Gonzalez et al., 2022). However, they may show diel cycles and have been found at 10-17 m during daytime (Nelson et al., 2018). The vertical distribution of lice is also highly dependent on water current and may vary a lot (Asplin et al., 2014). So far, these SCC systems have been used mainly to produce post-smolt up to one kilogram and have thus far served as a stopover before the fish are transferred to net pens, where they are reared until harvest. This means of rearing allows the fish to stay away from the open sea for an extended period and, thus, the chances of sea lice infestation and escapees are lessened. Despite having a significant potential to address biological challenges, the semi-closed systems entail high investment costs and are today regulated similarly to traditional net pens—that is, with license costs and limitations in the allowed maximum biomass. The high costs and regulatory restrictions mean that many producers and farmers of the systems hesitate to proceed and adopt these technologies because profitability is often not assured. To increase the likelihood of improving the regulatory conditions for the semi-closed systems and to further develop them into more sustainable and feasible alternatives, there is a need for scientific documentation of fish performance, health, and welfare in these systems, in addition to holistic research on environmental functionality.
Because the semi-closed systems are relatively new production technologies, the scientific peer-reviewed papers documenting their environmental functionality and biological performance are scarce. Most of the available literature describes and documents the engineering aspects, particularly the hydrodynamic properties and construction (Gorle et al., 2018; Klebert et al., 2018; Kristiansen et al., 2018; Chu et al., 2020; Su et al., 2021). Further, there are only a handful of publications on production capacity (Nilsen et al., 2017) and health, welfare, and performance (Rud et al., 2017; Balseiro et al., 2018; Nilsen et al., 2019). Recently, it was reported that post-smolts from two types of semi-closed facilities had better growth, lower FCR (feed conversion ratio), and lower levels of cortisol compared to fish that had been raised in open net pens (Øvrebø, 2020). In general, these publications describe promising performance, health, and welfare in S-CCS compared to the traditional open net pens. However, most systems are still in a pilot/prototype stage, and further documentation of environmental functionality and biological performance is needed, as the systems are still developing. The scientific challenges with the prototypes are that most of them exist in one version, thus, without replicates, and reliable controls or reference cages. It is also important to note that various S-CCS have different system requirements, which are major contributory factors to how the fish behave and cope when reared in these artificial environments. Some of the technological developments being carried out recently include water and waste treatment, which play an important role in ensuring that the conditions of the rearing environment are optimal. It is being explored how to develop systems and processes at lower costs, as one of the major drawbacks for S-CCS and other closed systems are that the investment costs are much higher than those for traditional cages (Flimlin et al., 2008; Calabrese et al., 2017; Moe, 2021). Additionally, the Norwegian Food Safety Authority is requiring welfare documentation from all new technologies; hence, technology suppliers are obliged to document the welfare conditions of fish in these new environments. For the most optimal welfare monitoring and evaluation, the indicators for monitoring may be system- and procedure-specific (Noble et al., 2018).
One of the S-CCSs available at present is the FishGLOBE V5 (Figure 1A). It is a 3500 m3 S-CCS where water is pumped in through pipes from approximately 14 m deep. The facility is located in Lysefjorden, Rogaland County, Western Norway and is situated on a pier approximately 100 m from land. Each of the three pumps has a capacity of approximately 1000 L/s. The water retention time is 20 min. The system water is oxygenated and maintains dissolved oxygen at 90% saturation. Sludge is collected in the storage tanks. The production of fish starts with the delivery of smolt from the well boat and they are stocked through a hatch in the roof of the system. Feed is provided through perforated pipes that distribute feed from the storage compartment centred 1.5 m above the water level and horizontally to each side of the tank, thus securing even distribution of feed (Figure 1B). Fish appetite is observed with a submerged video camera or with a video camera placed at the filter. From the video, feed spill is evaluated, and feeding may be adjusted according to the fish appetite. When the fish reach the post-smolt size of over 800 g, they are transferred to the well boat via a pipe at the bottom of the globe.
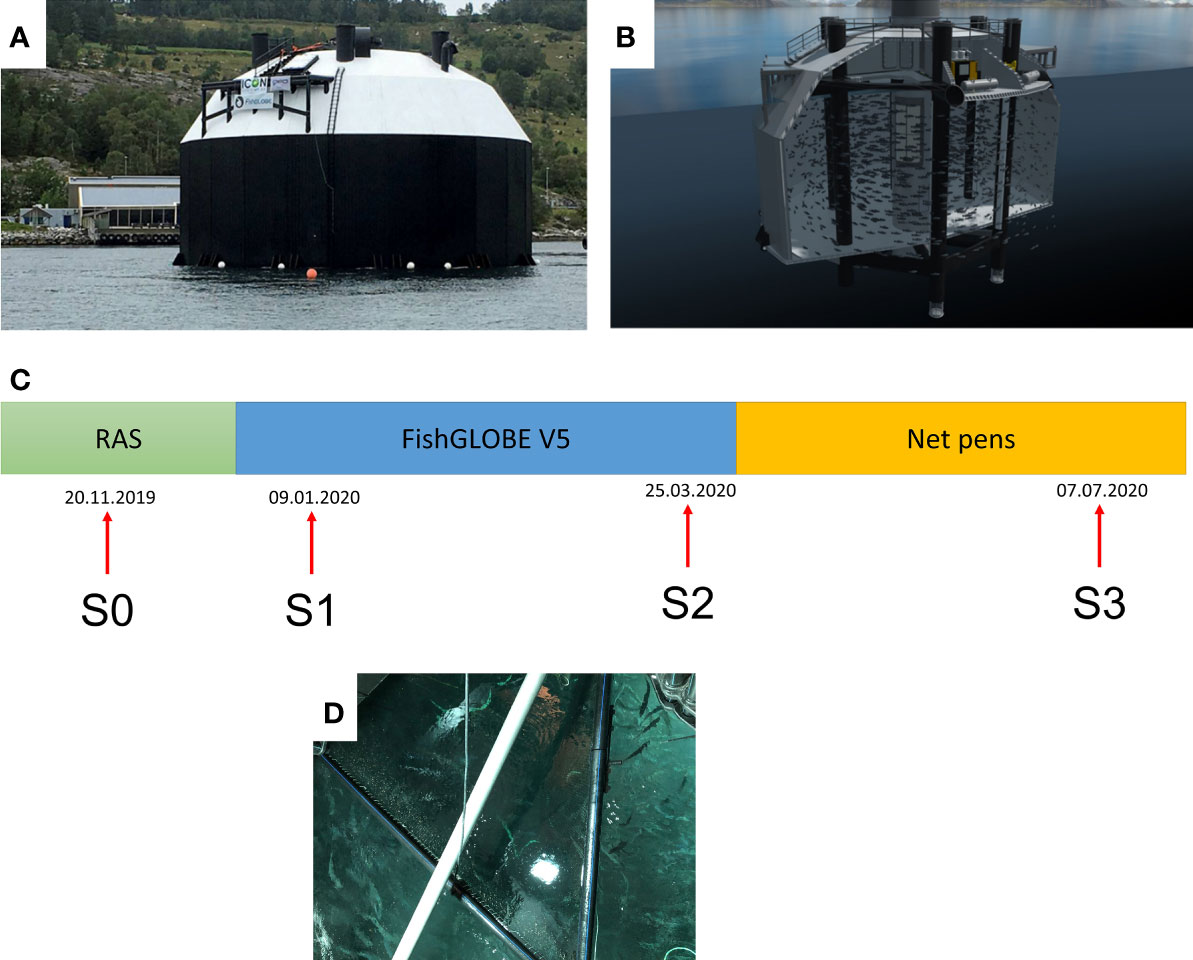
Figure 1 FishGLOBE V5 and summary of monitoring programme employed in the current study. (A) Exterior photo of a FishGLOBE V5 system and (B) diagrammatic representation of the interior, showing the upper deck and the compartment for fish rearing. (C) The monitoring programme of the group of salmon followed 3 main phases – Phase I was the RAS phase, followed by Phase II, where fish were transferred to FishGLOBE V5 and reared for 3 months, and then eventually moved to net pens. There were 4 main sampling points (S), which happened when fish were in the FishGLOBE V5 system. Dates are given as day.month.year. (D) A photo illustrating how fish were taken from the FishGLOBE V5 during sampling. (A) Photo: ©FishGLOBE; (B) Illustration: ©FishGLOBE; (C) Photo: Kevin Stiller
The main aim of this study was to document the fish performance, health, and welfare in FishGLOBE V5 to support the further development of this semi-closed system. Extensive commercial use of FishGLOBE and other semi closed systems demand the security of fish health and welfare, in addition to their functionalities such as being free from sea lice and escapees. A monitoring plan was developed in which a group of production fish was followed during a production cycle from the pre-smolt RAS tanks on land to the FishGLOBE V5 tank in the fjord, and to the first months after transfer to net pens.
A group of post-smolt from Salmobreed strain (N=200,000, average weight 233 gram) was followed from a RAS facility (S0) to the FishGLOBE V5 system (S1 and S2; Lysefjord) and further on to an on-growing net pen (S3). All three production systems are situated in Rogaland County, in the southwest part of Norway. The documentation and monitoring were carried out during an 8-month period from November 2019 to July 2020 (Figure 1C).
The fish were in 850 m3 tanks with a density of 49 kg/m3 before leaving the RAS facility. There were no reported health or welfare issues when the fish were in the facility. The fish were smoltified by photoperiod manipulation, and smolt status was evaluated using morphology and chloride levels (124 mg/l) at day five in the sea water challenge test which was conducted one week before transfer. FCR before transfer was 0.78. The fish without anaesthesia were transported from the RAS facility (S0) to FishGLOBE V5 with a well boat. The whole operation lasted for three hours. Stocking density during transport was 20.7 kg/m3, O2 = 100.4 ± 2.2%, pH = 7.7± 0.0, and CO2 = 3.2 ± 0.1 mg/l (data given as mean ± SD). Once the well boat arrived at FishGLOBE V5, the smolts were delivered through a hatch in the roof of the FishGLOBE V5 tank. A continuous 24-h photoperiod was maintained both at the RAS facility and in the FishGLOBE V5.
After approximately 3.5 months inside the FishGLOBE V5 (density at the termination of this phase was 75 kg/m3) the post-smolts were transferred through a well boat, which lasted for 3 hours, to the on-growing phase (S3) and placed in one open net pen (160 m in circumference, 20 – 35 m deep). During the S3 cage phase that lasted from April to July the stocking density increased from 9.6 kg/m3 to 22.6 kg/m3.
Several water quality parameters were measured throughout the S0, S1, and S3 phase (Table 1). For S0 and S3 the measurements were provided by the site managers. S1 data were measured by the project team. Due to Covid-19 restrictions, the project team was not allowed to visit the FishGLOBE V5 facility during S2; thus, water quality data from S2 sampling were not available. For S1, measurements inside the FishGLOBE V5 were taken at 3-, 6-, and 9-m depths and at 2.3 m and 4.3 m from the centre wall. The data from the different sampling positions are pooled because no statistical differences were found between different sampled depths or distance from the centre. Parameters including water temperature (°C), pH, conductivity (mS/cm), salinity (ppt), O2 (% saturation), CO2 (mg/l), and alkalinity (mg/l) were measured on-site using dedicated sensors (Table 2). Water samples were sent to AKVALab in Nofima Sunndalsøra for the analysis of TSS (mg/l), turbidity (NTU), TAN (mg/l), nitrite (mg/L), and nitrate (mg/L). In addition to temperature, oxygen and salinity measured at S1 by the project team (Table 1), daily recordings of these parameters were also performed throughout S1 – S2 by site managers (temperature: 8.2 ± 0.8; oxygen: 89.3 ± 1.4; salinity: 31.2 ± 0.5).
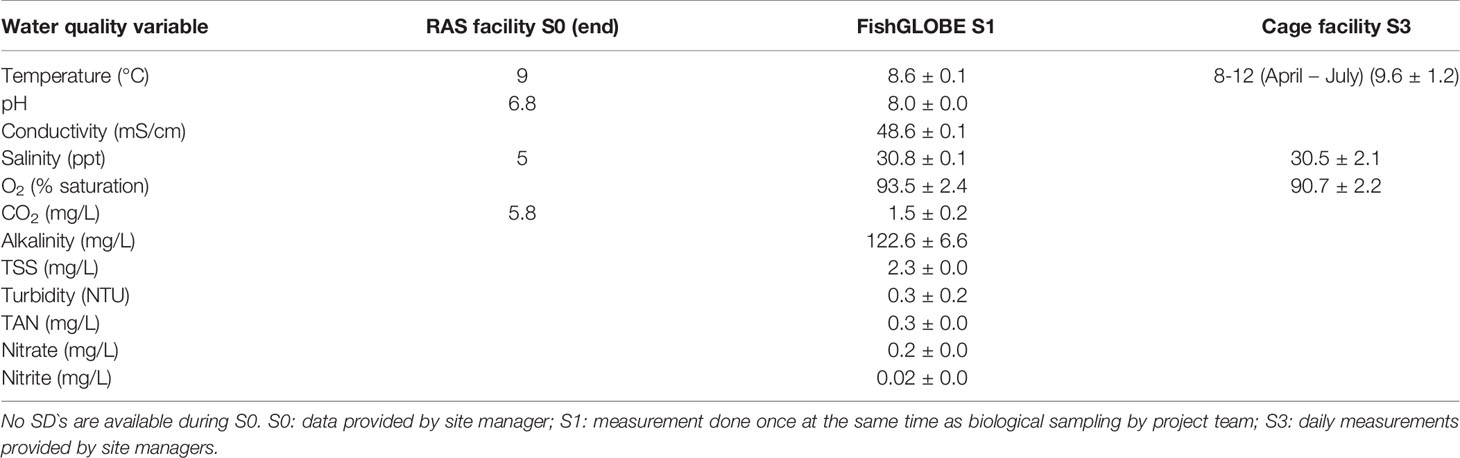
Table 1 Mean ( ± SD) measured water quality variables during end of S0, S1 at 3, 6, and 9 m depth and at 2.3 m and 4.3 m from the centre wall, and S3. Empty cells mean missing data.

Table 2 Description of sensors measuring water quality during S1 sampling.
Four samplings were conducted to document a suite of health and welfare indicators. The first set of samples (S0) was taken at the RAS facility approximately four weeks before transfer to the FishGLOBE V5 system. The second set of samples (S1) was taken after three weeks of acclimation in the FishGLOBE V5, while the third set of samples (S2) was taken two weeks before transfer from FishGLOBE V5 to the net pen. The fourth and final samples (S3) were taken three months after sea transfer in the open net pen (Figure 1C). Fifteen (N=15) individual fish were collected per sampling point.
When fish were collected from FishGLOBE V5 (S1 and S2), a small sampling net (2.5 – 3m per side, approx. 2 m deep) was used (Figure 1D). The net was deployed in FishGLOBE V5 the day before the sampling, and fish that had jumped into the net were hauled out and used as specimen for the sampling. This strategy was employed to lessen the disturbance in the system during sample collection. Sampling from the RAS phase (S0) and open net pen (S3) was done with a commercially available scoop net. In all instances, fish for sampling were humanely euthanised with an overdose of Finquel (250 mg/l). During sampling, fork length and weight were measured. These measurements were used to calculate thermal growth coefficient (TGC), specific growth rate (SGR), and condition factor (CF), using the following formulas:
No more than 5 min after euthanasia, blood was withdrawn from the caudal artery using a heparinised vacutainer and centrifuged at 3000 rpm/min for 10 mins to collect the plasma, which was snap-frozen in dry ice and eventually stored at -20°C until analysis. The external morphological operational welfare indicators (OWI) were assessed according to the scoring scheme described by Noble et al. 2018. Briefly, this scoring system accounts for twelve external parameters (i.e., eye damage, operculum deformity, snot damage, upper jaw deformity, lower jaw deformity, emaciation, vertebral deformity, skin damage, and damage to dorsal, caudal, pectoral and pelvic fins) defining the morphological welfare status of salmon. Each indicator is scored using a 0-to-3 scale, where 0 means in good condition while 3 indicates a severely compromised state.
Gills from the second gill arch and skin right below the dorsal fin were dissected and divided into two. Half of the section was placed in RNAlater® solution (SigmaAldrich, Merck KgaA, Darmstadt, Germany), allowed to penetrate at room temperature overnight, and thereafter transferred to -80°C until RNA isolation. The other half was placed in 10% buffered formalin (Cellstore Pot, CellPath Ltd, UK) for histological evaluation. The fish was opened from the ventral region to expose the abdominal cavity. The liver and heart were weighed for hepatic and cardio somatic index, respectively (100*organ weight/ungutted body weight). The kidney was evaluated for nephrocalcinosis based on a 0-to-4 scoring scheme, where 0 means no sign while 4 indicates a severe case. Cumulative mortality during the FishGLOBE V5 and net pen phases was calculated based on the data provided by the producers.
Plasmatic levels of cortisol were analyzed with ELISA using a commercially available kit (Demeditic Diagnostics GmbH, Kiel, Germany) according to manufacturer instructions, while glucose and magnesium levels were analyzed by the Pentra Clinical Chemistry Analyzer (Pentra C400, HORIBA ABX SAS, Montpellier, France).
The decalcified gills and skin samples were embedded in paraffin following a series of ethanol dehydration (75% → 100%), xylene (3X) clearing, and, finally, paraffin (2X) infiltration in a benchtop histoprocessor paraffin (Leica TP1020, Germany), cut into a 5-μm section using a rotatory microtome (Leica RM2165, Germany) and stained with Periodic Acid Schiff-Alcian Blue (AB-PAS) in an automated stainer (ST5010, Germany). Images were digitalised in an Aperio CS2 digital slide scanner (USA).
Quantitative histological evaluation of the gills was performed following a previously described method (Lazado et al., 2021). Briefly, eight fields in the whole gill arch section were randomly selected, with each field containing forty lamellae. Key branchial histopathological changes were identified including lamellar clubbing, epithelial lifting, hyperplasia, hypertrophy, lamellar fusion, aneurysm, oedema, hyperemia, and necrosis. A lamella that did not show any sign of damage or lesion was defined as “healthy”. The frequency of the lesion was expressed relative to the total number of evaluated lamellae.
For the skin, a semi-quantitative evaluation was carried out in three randomly selected regions of around 500 μm in the distance per area. Microscopic skin health status was scored by blind evaluation using two key criteria: the general appearance of the epidermis and the quality of the epithelial surface. A three-point scale skin health scoring system was employed (Sveen et al., 2019).
Total RNA was isolated from the skin using the Quick-RNA™ Microprep Kit (Zymo Research, USA) according to the manufacturer’s protocol. Complementary DNA (cDNA) was synthesised using the Taqman Reverse Transcription Kit (Applied Biosystems, USA) in a 20 μL reaction mixture containing 9.6 μL of 500 ng template RNA, 2 μL of 10X RT Buffer, 1.4 μL of 25 mM MgCl2, 4 μL of 10 mM dNTP mix, 1 μL of Rnase Inhibitor, 1 μL of MultiScribe™ Reverse Transcriptase, and 1 μL of Random Hexamers. Thermocycling was carried out in a Veriti™ 96-Well Thermal Cycler (Applied Biosystems, USA) following this protocol: 25°C for 10 min, 37°C for 30 min, and 95°C for 5 min.
The transcript levels of selected genes with involvement in epithelial integrity, stress response, mucin, and antibacterial defence were quantified by real-time quantitative polymerase chain reaction (RT-qPCR) in the QuantStudio™ 5 Real-Time PCR System (Applied Biosystems, USA). The reaction mixture contained 5 μL of PowerUp™ SYBR™ Green Master Mix (Applied Biosystems, USA), 0.5 μL 10 μM of each forward/reverse primer (Invitrogen, USA), and 4 μL of 1:10 cDNA. The cycling param were as follows: pre-incubation at 95°C for 20 sec, amplification with 40 cycles at 95°C for 1 sec and 60°C for 20 sec, and a dissociation stage of 95°C for 1 sec, 60°C for 20 sec, and 95°C for 1 sec. A five-step standard curve of two-fold dilution series was prepared from pooled cDNA to determine the amplification efficiencies. Gene expression was normalised by the geometric mean of three reference genes (Elongation factor alpha-1, Acidic ribosomal protein, and β-actin), as described previously (Nagasawa et al., 2012). Primer sequences of the genes used in the study are provided in Supplementary Table 1.
The production and physiological data were expressed as mean ± SD and analyzed by one-way ANOVA when data were normally distributed. When normal distribution was not the case, as for the physiological stress data, Kruskal-Wallis test followed by Conover-Iman post hoc test was performed. Statistical significance was defined when P < 0.05.
A Shapiro–Wilk test was used to evaluate the normal distribution and a Brown–Forsyth test to check for equal variance of the gene expression and histological data set. One-way ANOVA was then employed to investigate significant differences over time. In addition, the Holm–Sidak test was used to identify pairwise differences. All statistical tests were performed using SigmaPlot 14.0 Statistical Software (Systat Software Inc., London, UK).
Data from external welfare score were not statistically treated because the data are given as proportions. These data are therefore presented as descriptive data (Kolarevic et al., 2021).
During the time that the fish were in the FishGLOBE V5 (S1 + S2), the cumulated mortality was 1.4%, while it was 4.9% during the net pen phase (S3) (Figure 2).
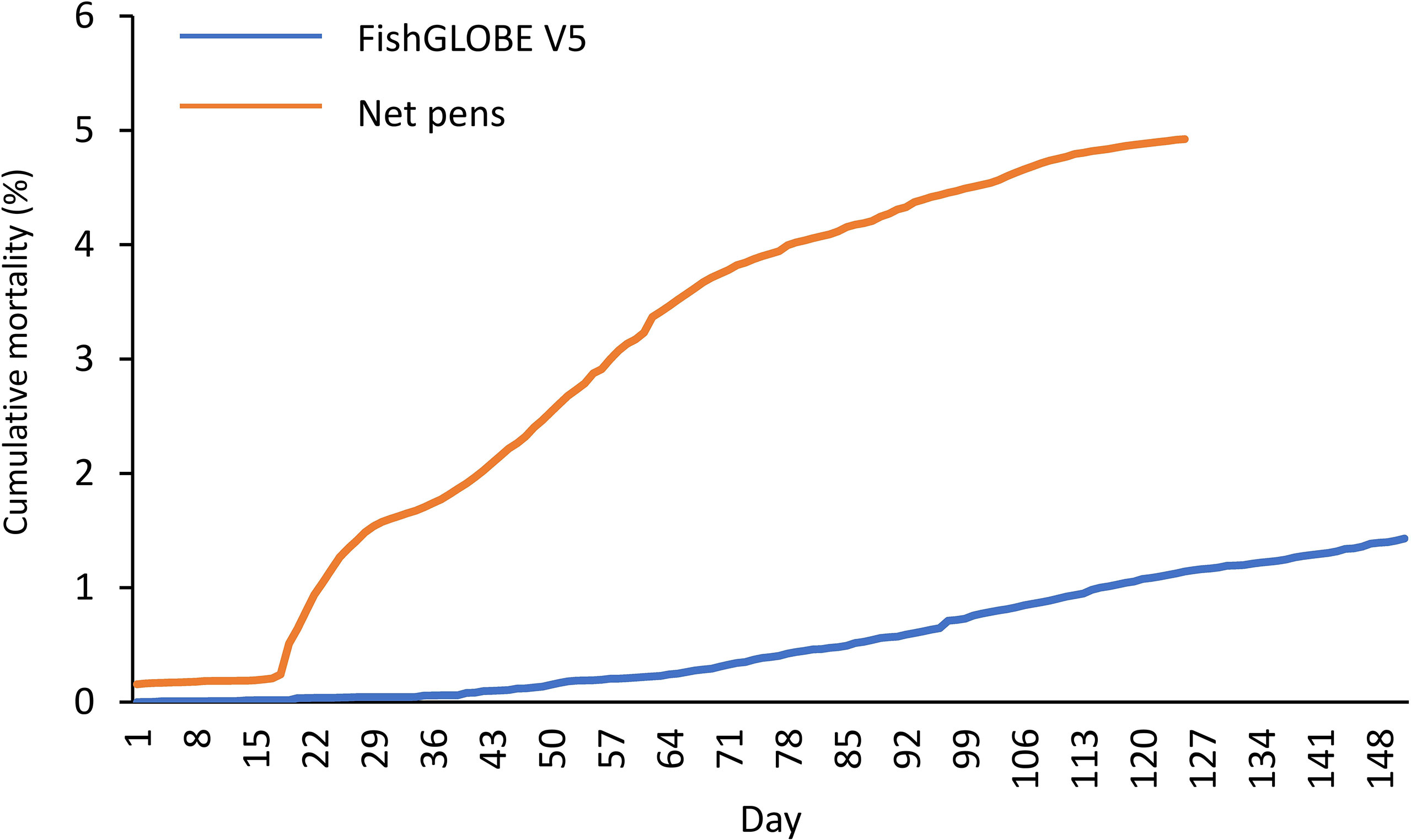
Figure 2 Fish mortality. Cumulative mortality (%) in FishGLOBE V5 (blue line) and three months after release to net pens (orange line). Registrations of dead fish continued after the S3 sampling was carried out in July.
From the S0 sampling at the RAS facility (November 2019) to S3, three months after sea transfer to the net pens (July 2020), the fish grew from 238 g to approximately 1.4 kg (Figure 3A). Expressed as SGR (Figure 3B) and TGC (Figure 3C), the best growth period was when the fish were inside the FishGLOBE V5 (S1 – S2). Between 9 January 2020 (S1) and 25 March 2020 (S2), SGR was 1.3 while TGC was 4.0 (Figures 3B, C). For the entire period, from the initial sampling to the last sampling in the open net pen phase (S0 – S3), SGR and TGC were 0.7 and 2.5, respectively.
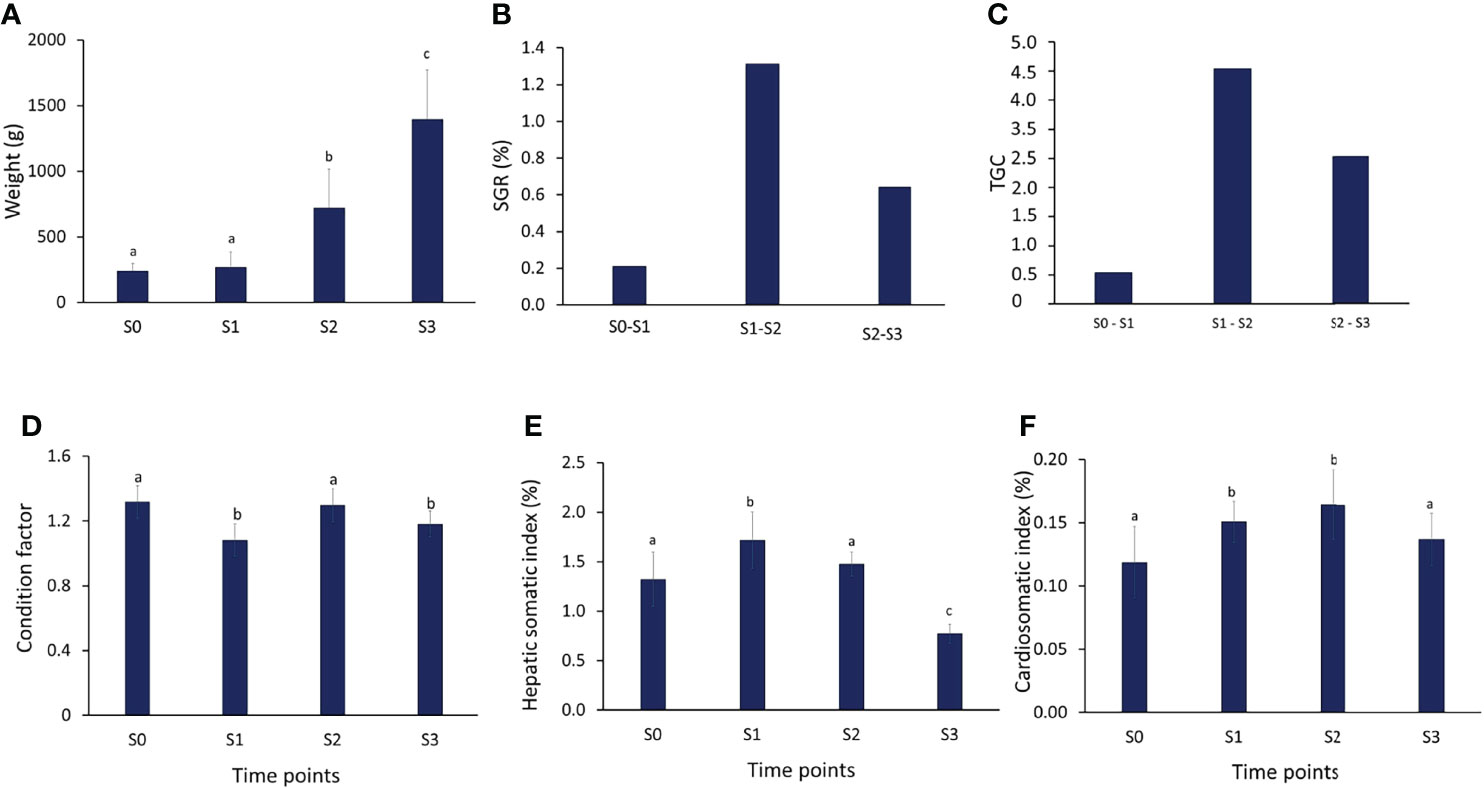
Figure 3 Performance indicators. (A) weight (g), (B) specific growth rate (SGR,%), (C) thermal growth coefficient (TGC), (D) condition factor, (E) hepatic somatic index (%) and (F) cardio somatic index of the representative fish (N=15) during the 3 phases of production. Values are provided as mean ± SD (A, D–F). Different letters indicate significant different at P < 0.05, analyzed by ANOVA followed by a Tukey post-hoc test
The condition factor (CF) is given in Figure 3D. The CF was significantly lower both at the first three weeks inside the FishGLOBE V5 (S1) and at S3 (three months after sea transfer) (P < 0.001). CF between S0 and S2 did not significantly differ.
The hepatic somatic index (HSI) was highest at S1 and lowest at S3 (Figure 3E). HSI at S1 was significantly higher (P < 0.001) compared to all time points. The cardio somatic index (CSI) increased from S0 to S2 (Figure 3F). CSI in S1 and S2 was significantly higher than in S0 and S3 (P < 0.001).
Weekly sea lice count of 20 fish every week showed no lice in the period between S1 and S2 for the present population. Additionally, no fish escaped from the system during this period.
None of the sampled fish were visually scored for nephrocalcinocis in the FishGLOBE V5 or the time thereafter (S1, S2, or S3; Table 3). However, in the RAS phase (S0, Table 3), 20% of the sampled fish were visually scored with nephrocalcinocis level 1, and 2% were scored with level 2.
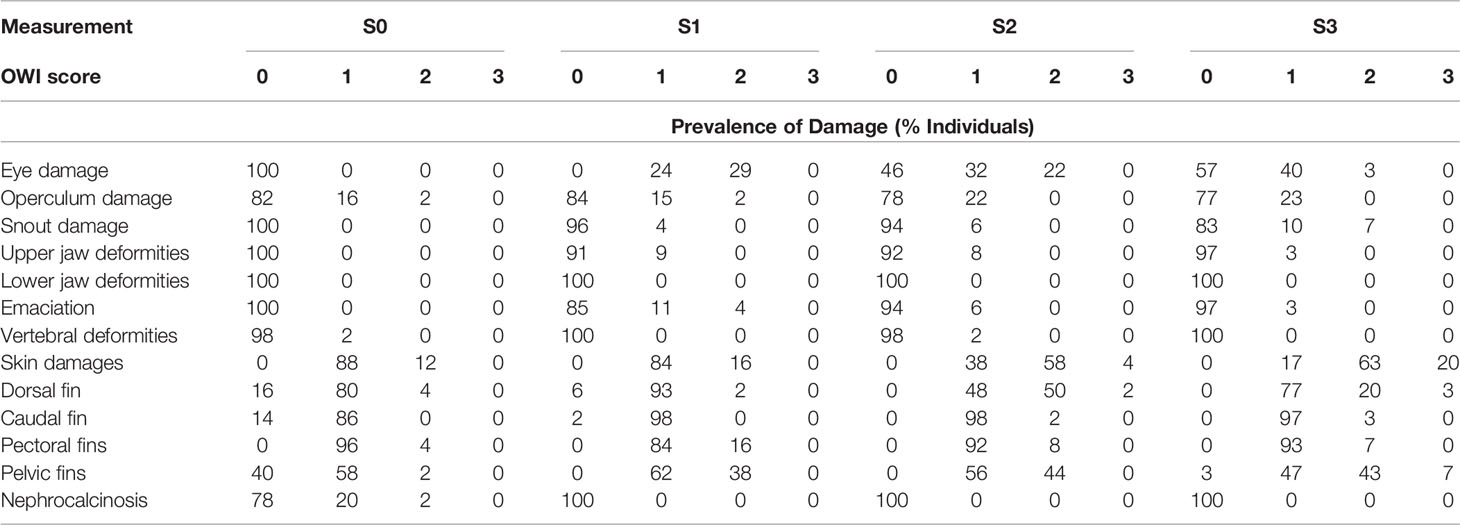
Table 3 Prevalence of damage, deformities, emaciation and nephrocalcinosis (% Individuals), n = 30) of evaluated external morphological operational welfare indicators [OWI, scored from 0 (lowest) to 3 (highest) prevalence] recorded at 3 different time points (T0 = RAS facility, T1 and T2 = FishGLOBE V5, T3 = net pen facility).
There was a high prevalence of eye damage (43 – 54%) in the FishGLOBE V5 and in the net pen (S1 – S3, Table 3), though the proportion declined with time. There was also a moderate prevalence of operculum damages (17 – 23%); however, no apparent trend during the production cycle. The proportion of snout injuries increased, around 17% of snout damages were observed after sea transfer compared to 6% from the first period in FishGLOBE V5. Cases of emaciation increased following transfer to FishGLOBE V5 (S1), but thereafter showed decreasing prevalence. Jaw and vertebral deformities were low in prevalence.
A majority of the sampled fish had slight damages (1 on the scale) in the pectoral fins similar at all sampling points. Fish with more serious (>2) pectoral fin injuries were minimal. Caudal fin damage was accounted for in up to 98% of the sampled population and increased with time. A considerable number of fish had severe pelvic fin damage, and up to 44% of the fish had a damage score of 2, particularly during the period S1 to S3 (Table 3). The proportion of fish with pelvic fin injuries with a severity score of 3 increased when the fish were transferred to the net pen. As for the dorsal fin, the severity of the injuries increased as the fish became bigger, and there were several fish with a severity score of 3 at S2 and S3, respectively.
Plasma cortisol showed large individual variations at S0, S1, and S2, and was significantly higher during the last part of the FishGLOBE V5 phase compared to the first phase (S1: 287.8 ± 203.8 nmol/L; S2: 426.2 ± 262.4 nmol/l, P < 0.001) (Figure 4A). The cortisol levels of fish sampled in the net pen phase (S3: 42.0 ± 29.9 nmol/L) was significantly lower compared to all previous phases (P < 0.0001).
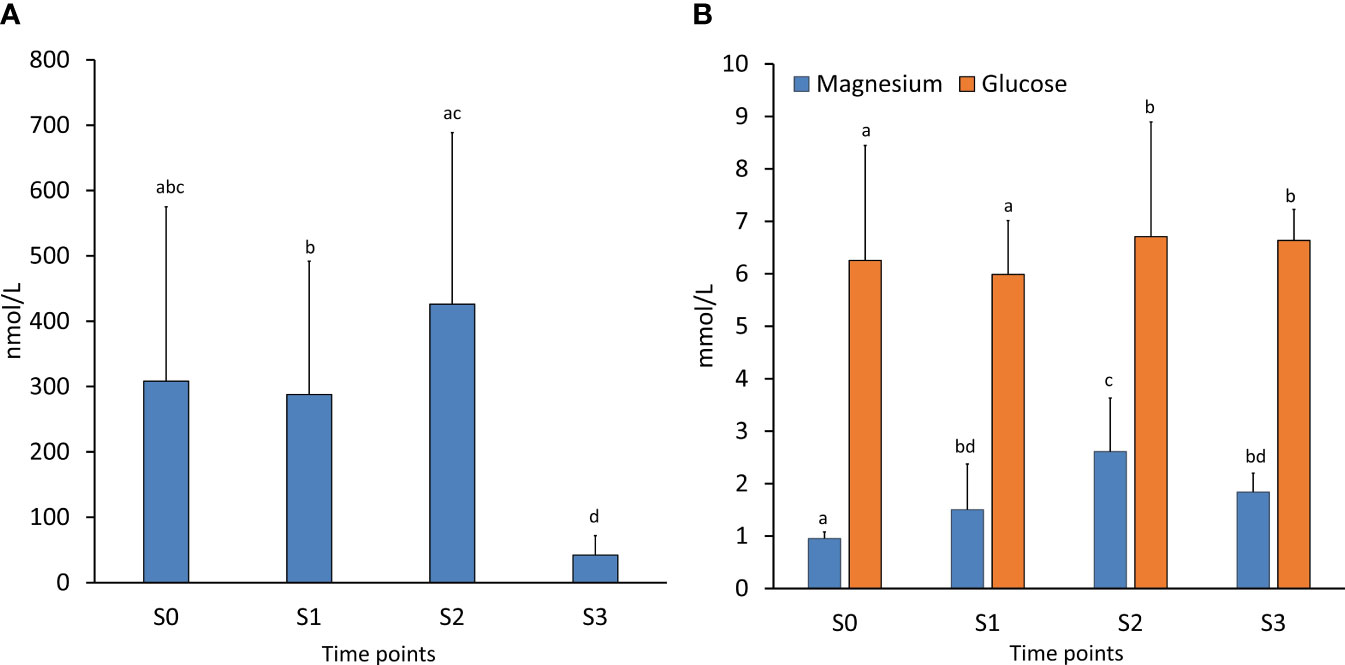
Figure 4 Plasma stress indicators. The levels of (A) cortisol (nmol/l) and (B) magnesium and glucose (mmol/l) were analyzed in the representative fish (N=15) during the 3 phases of production. Values are provided as mean ± SD Different letters indicate a significant difference at P < 0.05, analyzed by Kruskal-Wallis test followed by Conover-Iman post hoc test.
The four sampling points differed regarding plasma glucose levels (P = 0.01), where average values were a little above 6 mmol/L (Figure 4B). Both S0 and S1 sampling points were significantly lower compared to S2 and S3. Plasma magnesium levels increased from S0 (1.0 ± 0.1 mmol/l) to S2 (2.6 ± 7.0 mmol/l), where the level in S2 was significantly different from all sampling points (Figure 4B). The magnesium level between S1 (1.5 ± 0.9 mmol/L) and S2 was significantly different, and both timepoints differed significantly with S0 (P < 0.0001). Mg for S3 was 1.8 ± 0.4 mmol/L.
Histology was focused on two mucosal organs: gills (Figure 5) and skin (Figure 6). Almost 60% of the evaluated filaments in all sampling points were regarded as “healthy” (Figure 5A). Epithelial lifting, lamellar clubbing, hypertrophy, and hyperplasia were the most frequent pathologies (Figures 5A–E) and accounted for 30% of the assessed lesions. There were no statistical differences between S0 – S3 for the different gill conditions. The lowest percentage of filaments with healthy status relative to the total amount of evaluated filaments was registered at S1, immediately after the transfer from the RAS facility to FishGLOBE V5. A higher proportion of gills with the condition “lifting” was also documented at this time point.
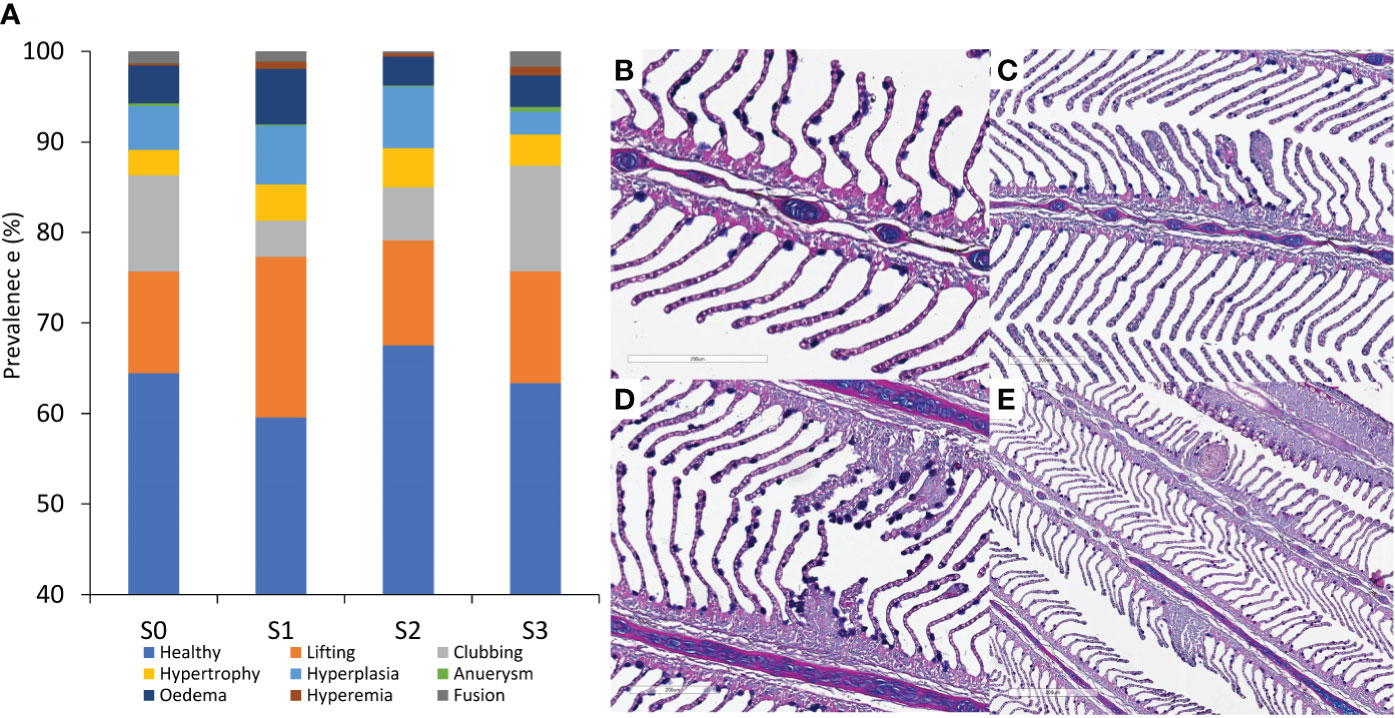
Figure 5 Gill health status. (A) Quantitative histological evaluation was performed in a group of fish from the four sampling points, where common gill pathologies were accounted for and expressed in relation to the number of evaluated healthy lamella. Representative histological photos of (B) healthy gill filaments with defined structure and fair distribution of mucus cells, and some of the pathologies identified, including (C) clubbing, (D) hyperplasia at the base of the filament, with an increased number of mucus cells, and (E) aneurysm and advanced stage of hyperplasia. The data represent the evaluation from 15 individual fish (N=15) per sampling point.

Figure 6 Skin health status. (A) Histological evaluation of the skin was carried out by identifying the status of two key criteria, general appearance and surface quality, using a 0-to-3 scoring scheme. Different letters indicate a significant difference at P < 0.05. The data represent the mean ± SD of 15 individual fish (N=15) per sampling point. Representative histological photos of (B) healthy skin, with defined epidermal and dermal layers, and (C) comprised skin, where the surface is rough and missing a significant portion of the epidermis.
Skin histological condition was assessed by two criteria: 1) general appearance and 2) skin surface quality (Figure 6A). Though there was considerable variability in the scores for the general appearance of the skin, no significant temporal differences were observed (P = 0.091). Nonetheless, the average scores were above 1.5, which was midway in the 0-to-3 scoring scheme used. In addition, the average scores for S1 and S3 were almost at the same level. In most of the fish evaluated, the skin surface was rough and had a missing epidermis (Figure 7C). The average scores for S0, S1, and S3 were above 2. There was a significant change in the status of the skin surface quality from S1 (right after transfer to FishGLOBE V5) to S2 (right before fish were taken out of the FishGLOBE V5), with a significantly lower score found in the latter (P = 0.008).
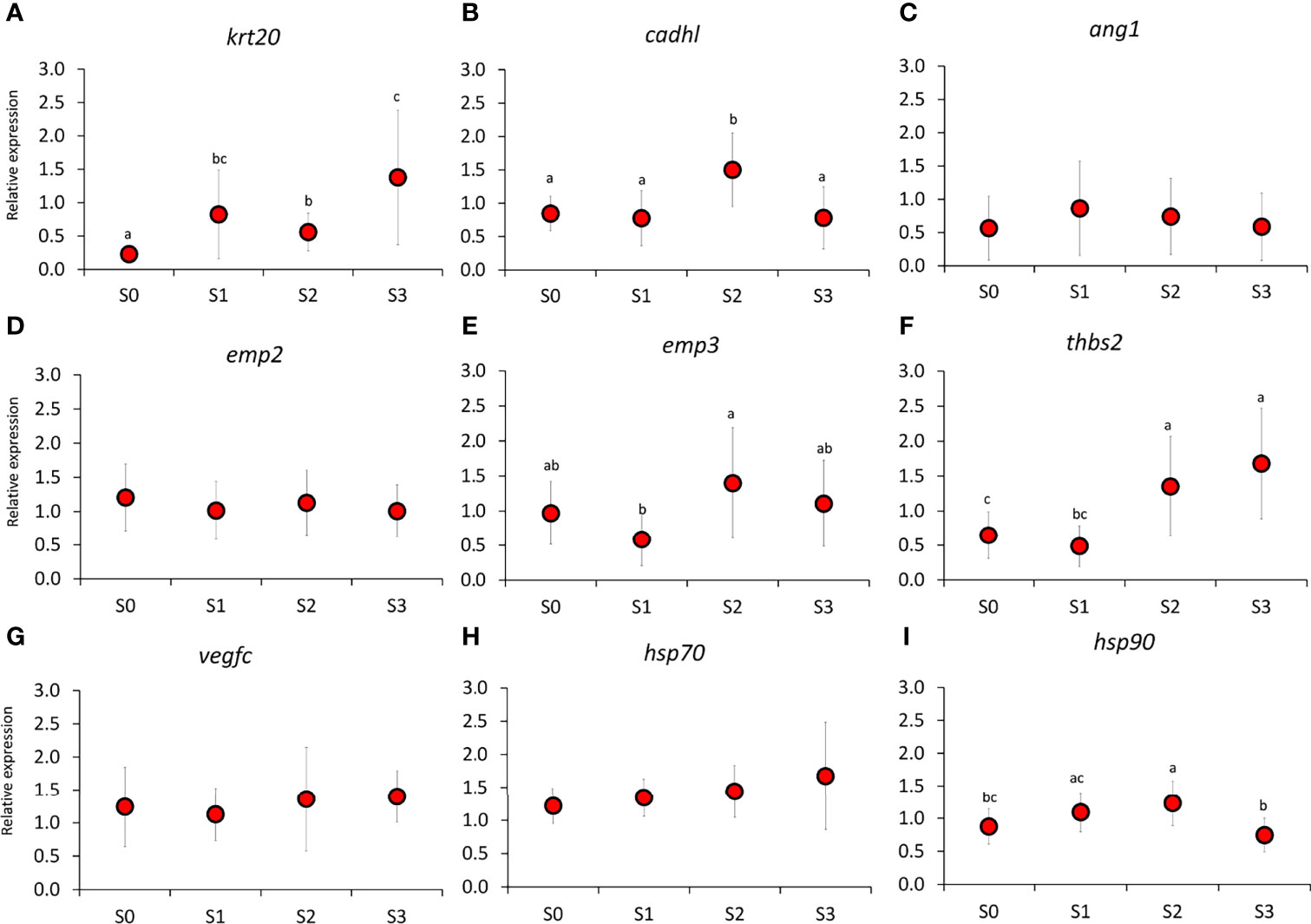
Figure 7 Expression of key genes important for (A–G) epithelial integrity and (H, I) stress response in the skin. Different letters indicate a significant difference at < 0.05, analyzed by ANOVA followed by a Holm–Sidak post-hoc test. Values are presented as mean ± SD of 15 individual fish (N=15) per sampling point.
To further elucidate the changes in the skin, we performed expression analysis of selected genes with key roles in tissue structure and integrity, stress, mucus biochemistry, and immunity (Figures 7, 8). From the genes important for tissue structure and integrity, keratin, type I cytoskeletal 20 (krt20, P = <0.001), B-cadherin-like (cadhl, P = <0.001), epithelial membrane protein 3 (emp3, P = 0.009), and thrombospondin-2 (thbs2, P = <0.001) demonstrated significant temporal changes (Figures 7B, E, F). Krt20 and thbs2 showed an increasing tendency during the production cycle while the expression of cadhl and emp3 was significantly elevated at S2 compared to other time points. In addition, there was a significant difference in the expression of cadhl, emp3, and thbs2 between S1 and S2. For stress genes, only heat shock protein 90 (hsp90, P = <0.001) showed significant changes (Figure 7I), where the level at the later part of FishGLOBE V5 phase was significantly higher compared to S0 and S3, but not with S1. There were significant temporal changes in the expression of all mucin genes in the skin (Figures 8A–C). The expression patterns among these genes showed high similarity with each other, that is, a significant downregulation after transfer to FishGLOBE V5 and elevation at the end of production in the system (S2), specifically with mucin 5b (muc5b, P = 0.001) and mucin 2 (muc2, P = 0.002). From the selected antibacterial defence genes, only defensin (def, P = 0.004) showed significant changes during production, with a decreasing tendency observed from S0 to S2. The expression of def from S1 to S3 did not significantly vary.
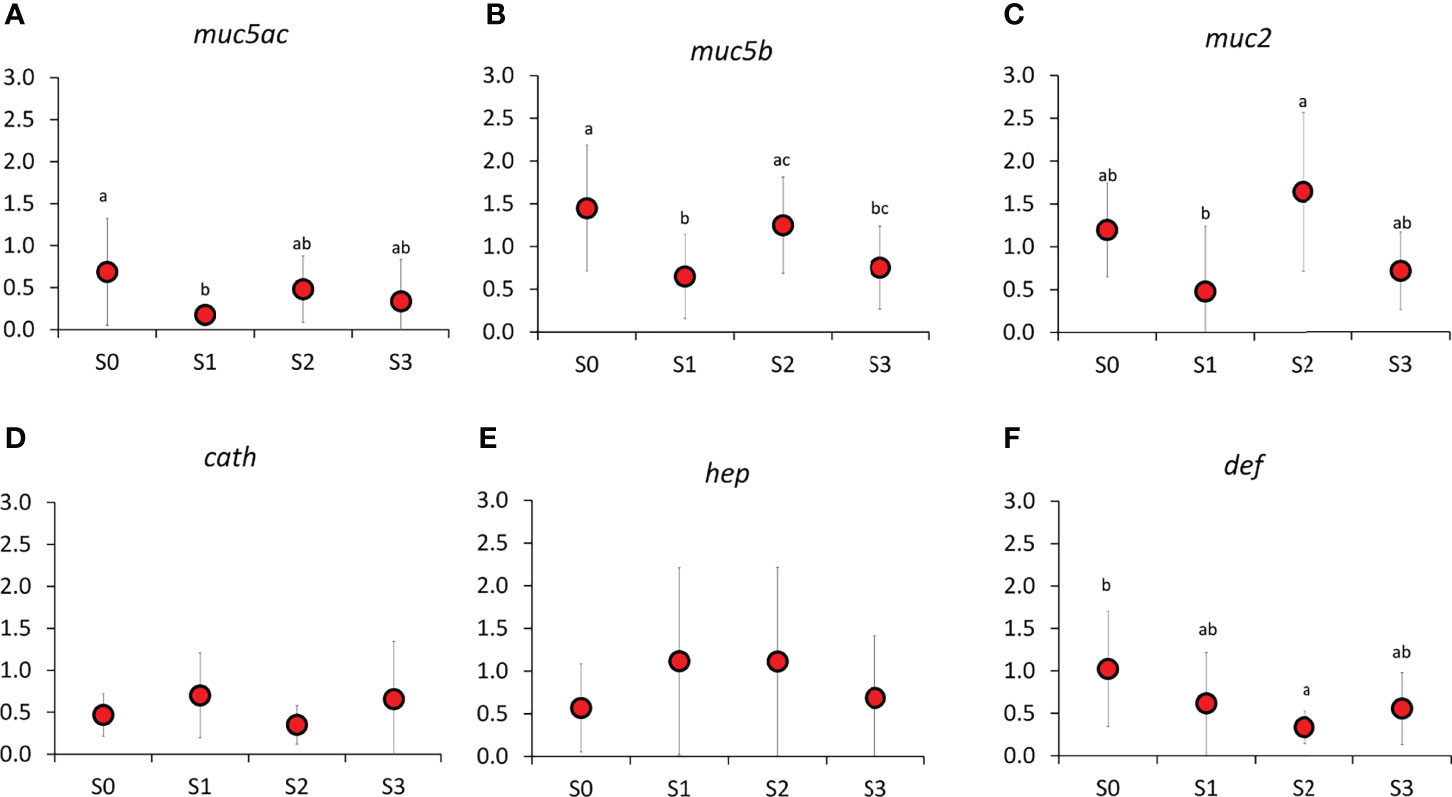
Figure 8 Expression of key genes important for (A–C) mucin and (E, F) antibacterial defence in the skin. Different letters indicate a significant difference at P < 0.05, analyzed by ANOVA followed by a Holm–Sidak post-hoc test. Values are presented as mean ± SD of 15 individual fish (N=15) per sampling point.
The study documented the health and welfare of a group of Atlantic salmon from a land-based RAS farm until three months after transfer to a net pen at sea, where, in between these productions, fish were reared in FishGLOBE V5, a novel semi-closed containment system. The use of semi-closed systems in the industry is still at a pilot stage, and most of the systems available still exist as one unit (N = 1) with no reference cage nearby. This has been considered a major challenge in several studies documenting the biological consequences of rearing fish in S-CCS prototypes (Balseiro et al., 2018; Karlsen et al., 2018), since we at present lack the possibility of controlled experiments with reliable replicates and controls. Also, most farmers are prohibited by regulations to place a reference cage so close to the semi-closed system. We are aware of these limitations, but still, the present study is needed to further develop the systems that are believed to be of importance for future salmon aquaculture since they have shown to be very promising regarding lice and escapees. One alternative in future study would be to replicate the study with new populations in the same system, but this also entails a certain limitation since populations differ in so many ways. This may require advanced mathematical models, so that monitoring data can be compared.
The water quality inside the FishGLOBE V5 was within the recommended levels for salmon post-smolts. Optimum hydrodynamics is important for even distribution of good water quality in all areas of the system to secure fish health and welfare. In addition, optimal hydrodynamics helps the self-cleaning of the system. Though not provided in this paper, the hydrodynamics inside the FishGLOBE system were monitored and found to be sufficient in ensuring that the parameters are within the limits supporting the biological requirements of post-smolts (Espmark et al., 2020).
The mortality inside the FishGLOBE V5 was low, at 1.4%, but increased to 4.9% during the first three months following transfer to net pens (S3). The mortality reported by the production staff was mainly attributed to weaker fish that did not grow. The weight from S0 to S3 was steadily increasing, with the best growth, in terms of SGR and TGC, during the 3.5 months that the fish stayed inside the FishGLOBE V5. The growth in FishGLOBE V5, in terms of SGR and TGC, is also acceptable with reference to commercial growth tables and reference studies (Espmark et al., 2017; Ytrestøyl et al., 2020). The drop in SGR and TGC from S2 to S3 was likely not associated with a drop in condition, but rather that the growth inside the FishGLOBE V5 was much better than during three months after transfer to cage, since SGR and TGC in the cages were not low compared to the mentioned references. Decreased growth right after transfer to new systems is expected, but in this case, the transfer from S0 to S1 did not cause a drop in growth, while transfer from FishGLOBE V5 to cage showed the expected drop. Also, the condition factor (CF) indicates that the fish at all sampling points were in good condition. The drop in CF from S0 to S1 may be due to sea water transfer (Alne et al., 2011). The second drop in CF from S2 to S3 may also be due to transfer. However, it is worth mentioning that the condition factor during S3 was approximately 1.2 and not low compared to benchmark studies (e.g., Ytrestøyl et al., 2020).
The cardio somatic index was higher inside FishGLOBE V5 (S1 – S2) compared to both phases before (S0) and after (S3). Larger hearts may be a consequence of training (Ytrestøyl et al., 2020) and can be seen as an indicator of good condition in fish. However, the benefit of increased CSI is not conclusive (Timmerhaus et al., 2021). Also, the hepatic somatic index showed differences between the phases, with significantly larger HSI during S1, and significantly lower HSI during S3. Smaller HSI during the sea phase (S3) was likely a direct effect of the fact that the fish was bigger in size. Also, larger HSI during the first period of the FishGLOBE V5 phase (S1) in relation to S2 and S3 can be fish-size-related. We would like to emphasise that the HSI was calculated from un-gutted fish. HSI is also considered to be an indicator of feeding status, where low feeding rate may result in smaller livers (Pandit and Gupta, 2019). Since in this study we did not systematically control and determine feed intake, we do not have reliable data to verify the difference in feed intake between the sampling points.
The general external welfare status was not sub-optimal and that rearing fish in FishGLOBE V5 did not have substantial negative consequences for the fish. Nonetheless, some indicators presented concerns for future consideration. Over 50% of the evaluated fish during the FishGLOBE V5 phase, regardless of whether it was during the early or late period of rearing, exhibited varying degrees of eye damages. Cataracts have many different causal factors, such as genetic conditions, nutritional deficiency, and water temperature (Hargis, 1991; Ersdal et al., 2001). Cataracts in fish are often considered irreversible, but osmotic cataracts have been reported to be reversible (Hargis, 1991). We could not establish the cause of the increased cases of cataracts, however, the moderate osmotic imbalance after sea transfer (S0 to S1) could be linked to the reported eye lesions. Around 25% of the evaluated fish demonstrated opercular shortening and allowing the fish to stay in FishGLOBE V5 prior to the open net pen did not aggravate the condition. Operculum deformities seem to have genetic causes, in addition to problems that occurred in early larval stages (Galeotti et al., 2000). This deformity can be an influencing factor for infection because gills are more exposed to the environment in malformed operculum (Lazado et al., 2020). This reflects some non-specific pathologies inferred from gill histology. The fish in this study showed increased snout damage from 6% to 17% from S1 to S3. This may originate from transport out of the FishGLOBE V5 and exemplify the issue of increased mechanical damage during transport in salmon (Sveen et al., 2020).
Fin erosion or damage is considered an important visible measurement of the welfare of the animals (Noble et al., 2007). These damages can be caused by several factors during production including handling, pumping, water velocity, density, pathogenic infection, and/or aggression (Timmerhaus et al., 2021). Salmon commonly exhibit fin damage at farms (Noble et al., 2007; Noble et al., 2018), and, usually, “mild” forms of damage (small area affected) are not potentially harmful at first, though when fish are crowded or stressed, and if water quality deteriorates, the condition rapidly increases in severity and causes serious damage to the affected animals. Except for dorsal fin damage, the FishGLOBE V5 phase did not worsen the cases of fin damage. The prevalence of dorsal fin damage with a score of 2 increased at the end of the FishGLOBE V5 phase but a considerable recovery was noted during the net pen phase.
Skin damage by visual gross scoring did not correspond well with the histology results, and the methods of evaluation could account for this difference. The gross scoring of skin evaluates the overall appearance of the skin surface while histological evaluation covers only a small portion. Nonetheless, both offer insights into the skin health status. The increased cases of higher scores (>2) for skin damage in S2 and S3 could be attributed to multiple factors, though very likely they were caused by the increased density (from S1 to S2) and handling and transport-related stress during transfer from FishGLOBE V5 to the net pen. Mechanical-related skin wounds are often an issue in salmon reared in high densities, and could be predisposing factors for infections (e.g. winter ulcers) (Sveen et al., 2020). In the future, it would be interesting to follow, at several time points during the FishGLOBE production, the skin gross appearance to establish whether rearing in this system has a pronounced impact on skin quality and the development of the damage. Nonetheless, most of the skin damage accounted for minor scale loss and haemorrhaging.
Nephrocalcinosis has been identified as a growing concern in salmon farming, especially in the last few years and it appears to be as common in flow-through as in RAS systems (Klykken et al., 2021). Around 20% of the fish evaluated during the RAS phase exhibited mild nephrocalcinosis (scores 1 and 2). Though there is an ongoing debate over the main causes of such a condition, water quality, such as the increased use of seawater during the hatchery stage, and CO2 in RAS, have been implicated as potential causal factors (Fivelstad et al., 2018). It is important to highlight that the CO2 levels were very low both during the RAS and FishGLOBE V5 phase and far below the threshold that has been shown to influence fish welfare (Skov, 2019). It is interesting to highlight that not a single case of nephrocalcinosis was identified during the FishGLOBE V5 and net pen phases. Despite the limited sampled fish, this suggests the possibility that the conditions in FishGLOBE V5 did not trigger nephrocalcinosis, or it could be that the condition in the system allowed the fish to recover. This supports the recent survey that mild nephrocalcinosis appears to be reversible after transfer to sea (Klykken et al., 2021).
Cortisol, the commonly measured primary stress hormone, was elevated in S1 to S2, compared to S3. In particular, the cortisol level at the beginning of the FishGLOBE V5 phase (S1) was lower than at the end period in the system (S2). In conditions that are less controlled, as in the present study, numerous factors might have influenced this profile, which could be difficult to single out. One example is the fact that the fish were sampled in different ways, because different systems required different strategies. While scoop-net was used in tanks and cage, the inserted net was used in FishGLOBE V5 to obtain a better representative sample in this huge system. However, the values were very similar to those of previous studies, demonstrating the cortisol response after stress induction in salmon. The values at S3 were close to the baseline resting cortisol levels in salmon. There were small but significant differences in plasma glucose level among sampling points, where fish sampled during the last part in FishGLOBE V5 (S2) and in sea cage (S3) phase had slightly higher glucose levels compared to the two first stages in RAS (S0) and early phase in FishGLOBE V5 (S1). The glucose ranged from 6 – 6.7 mmol/L and indicated mild stress, which partly indicates that the increased cortisol level was not chronic but an instantaneous response, likely due to handling-related stress during sampling. Despite the calming effect of anaesthesia, it is known that different chemicals, including Finquel, as used in this study can cause an immediate increase in physiological stress indicators such as cortisol. However, the increase is more severe if the fish are repeatedly exposed over time, and it is not expected to be large compared to the increase in cortisol because of handling (Zahl et al., 2012). In this study all fish were treated similarly, and Finquel was used for euthanasia.
Plasma magnesium is normally between 1 – 2 mmol/L. Levels deviating from this often indicate stress and a poor ability to regulate osmoregulation (Iversen and Eliassen, 2009). Here plasma Mg increased to 2.6 mmol/L in S2, indicating a slightly weakened seawater adaptation. The shift in salinity from 5 ppt in S0 (RAS) to 32 ppt in S1 and S2 (FishGLOBE V5) may have caused the significant increase in Mg (Bakke et al., 1991).
The skin and gills are multifunctional mucosal organs that represent the first line of defence in fish (Cabillon and Lazado, 2019). Because of their direct contact with the environment, their responses are often regarded as a reflection of the quality of the environment. Histological evaluation revealed that gill health status did not improve during the stay in FishGLOBE V5. Lifting, clubbing, and hyperplasia were the 3 pathologies that constituted the major deviations in the analyzed gill samples, accounting for 25 – 30% of the recorded cases. On the other hand, we can also state that rearing in FishGLOBE V5 did not worsen these structural alterations. As for the skin, the surface quality was significantly influenced during the early period in FishGLOBE V5. At the beginning of the period in the system, the skin surface exhibited roughness and, in many cases, the epidermal layer was missing. The skin significantly recovered at the latter phase of rearing in FishGLOBE V5, with an average score less than 2. This may indicate that the rearing environment provided a condition for the skin to partially recover. It may be that the elevated score in S1 is due to handling during transfer. Skin health, characterised by lesions and wounds, is a known issue for salmon during transportation from one production unit to another (Noble et al., 2018; Lazado et al., 2020; Sveen et al., 2020).
To further elucidate the observations in the skin, we performed gene expression analysis of key genes for tissue structure and integrity, stress, mucus, and antibacterial defence. The biological function of EMP3 is not fully known, but current evidence linked it to cell-cell interactions, control of cell proliferation, and apoptosis (Taylor and Suter, 1996). Both of the emp genes were significantly influenced by time in the current study, where a significant increase in expression from S1 to S2 was observed. It was earlier reported that these genes have been implicated in the skin development of salmon in S-CCS (Karlsen et al., 2018). Their pronounced regulation at the end of the FishGLOBE V5 phase likely contributed to the observed improvement in surface quality, as they may have facilitated the regeneration process. Two other genes, krt20 and thbs2, have key functions in the extracellular matrix (ECM) of skin (Agah et al., 2004; Rakers et al., 2010; Karlsen et al., 2018) and their increasing expression as the production progressed could be attributed to the changes relative to growth and further development of the skin in seawater (Karlsen et al., 2018). In a study by Sveen et al., (Sveen et al., 2019), ECM proteins were shown have key involvement in the early of wound healing mechanisms in the skin of salmon. The viscous mucus is an impervious gel, which acts as a lubricant, a physical barrier, and a trap for microbes (Linden et al., 2008; Osório et al., 2022). Here, we identified a significant temporal variability in the expression of mucin, the major glycopolymeric component of mucus, in the skin during the production cycle. We also observed that following transfer to FishGLOBE V5, mucin expression decreased, then was elevated again at the end of the rearing phase in the system. The elevated expression of mucins at S2 relative to S1 was perhaps a form of recovery response in the skin provided by the FishGLOBE V5 environment.
In summary, the monitoring programme employed in the current study documented the health and welfare of salmon from a RAS facility until transfer to a net pen. FishGLOBE V5, a S-CCS, was employed as a transition rearing phase between RAS and net pen. The adoption of S-CCS in salmon farming can shorten the time for the fish in on-growing net pens where they are more vulnerable to infection and erratic environmental conditions. Overall, we have shown that allowing salmon to stay for some time in FishGLOBE V5 did not significantly affect fish health, welfare and performance. Water quality in FishGLOBE V5 was within the threshold that supports a good rearing environment for salmon. This was reflected in the desirable growth, moderate mortality, and a potential recovery from nephrocalcinosis. The prevalence of eye, fin, and skin lesions following transfer to FishGLOBE V5 should be considered in the risk assessment with regard to the use of this S-CCS. Skin health may have benefited from the transition phase in FishGLOBE V5 based on the skin quality immediately after transfer compared to the end of the FishGLOBE V5 phase. Indications of stress induction including elevated plasma cortisol and magnesium levels should also be taken into consideration in the further development of this prototype. Future verification trials for FishGLOBE system and similar semi-closed systems should aim for including replicated systems and comparing reference net pens situated in locations with almost similar environmental conditions, including documentation of the temporal variability of different factors.
The raw data supporting the conclusions of this article will be made available by the authors on request and after the approval from all authors.
In accordance with the Norwegian Food Safety Authority, ethical review and approval was not required for this study.
Conceptualisation: AB and ÅE; Planning: ÅE, KTS and AB; Sampling: KTS, KS, BR, YM, JG, FS, and BE; Laboratory analysis: KTS and CL; Data analysis and curation: ÅE, KTS, and CL; Interpretation; ÅE, KTS, and CL; Visualisation and presentation: ÅE, KTS, and CL; Writing: ÅE, KTS, and CL. All authors contributed to the article and approved the submitted version.
The study was funded by Regionale forskningsfond RFF (ref 259057). The project is associated with the Centre of Research based Innovation project CtrlAQUA (RCN project #237856/O30).
AB was affiliated with FishGLOBE AS.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We acknowledge the staff at the AKVA Lab Nofima Sunndalsøra for the analysis of the water quality parameters. We would like to thank the personnel of Fomas - Fish Health and Environment AS (FoMAS - Fiskehelse og Miljø AS) for assistance during sampling, and staff from the RAS and cage facilities for valuable information reported in this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2022.871433/full#supplementary-material
Supplementary Table 1 | List of primers used in the current study.
Øvrebø T. K. (2020). Growth Performance and Welfare of Post-Smolt (Salmo Salar L.) Reared in Semi Closed Containment Systems (S-CCS) – a Comparative Study (Norway: MSc thesis, University of Bergen).
Agah A., Kyriakides T. R., Letrondo N., Björkblom B., Bornstein P. (2004). Thrombospondin 2 Levels are Increased in Aged Mice: Consequences for Cutaneous Wound Healing and Angiogenesis. Mat. Biol. 22, 539–547. doi: 10.1016/j.matbio.2003.09.004
Alne H., Oehme M., Thomassen M., Terjesen B., Rørvik K.-A. (2011). Reduced Growth, Condition Factor and Body Energy Levels in Atlantic Salmon Salmo Salar L. During Their First Spring in the Sea. Aquacul. Res. 42, 248–259. doi: 10.1111/j.1365-2109.2010.02618.x
Asplin L., Johnsen I. A., Sandvik A. D., Albretsen J., Sundfjord V., Aure J., et al. (2014). Dispersion of Salmon Lice in the Hardangerfjord. Mar. Biol. Res. 10, 216–225. doi: 10.1080/17451000.2013.810755
Bakke H., Bjerknes V., Øvreeide A. (1991). Effects of Rapid Changes in Salinity on the Osmoregulation of Postsmolt Atlantic Salmon (Salmo Salar). Aquaculture 96, 375–382. doi: 10.1016/0044-8486(91)90166-5
Balseiro P., Moe Ø., Gamlem I., Shimizu M., Sveier H., Nilsen T. O., et al. (2018). Comparison Between Atlantic Salmon Salmo Salar Post-Smolts Reared in Open Sea Cages and in the Preline Raceway Semi-Closed Containment Aquaculture System. J. Fish. Biol. 93, 567–579. doi: 10.1111/jfb.13659
Cabillon N., Lazado C. C. (2019). Mucosal Barrier Functions of Fish Under Changing Environmental Conditions. Fishes 4, 2. doi: 10.3390/fishes4010002
Calabrese S., Nilsen T. O., Kolarevic J., Ebbesson L. O. E., Pedrosa C., Fivelstad S., et al. (2017). Stocking Density Limits for Post-Smolt Atlantic Salmon (Salmo Salar L.) With Emphasis on Production Performance and Welfare. Aquaculture 468, 363–370. doi: 10.1016/j.aquaculture.2016.10.041
Chu Y. I., Wang C. M., Park J. C., Lader P. F. (2020). Review of Cage and Containment Tank Designs for Offshore Fish Farming. Aquaculture 519, 734928. doi: 10.1016/j.aquaculture.2020.734928
Ersdal C., Midtlyng P. J., Jarp J. (2001). An Epidemiological Study of Cataracts in Seawater Farmed Atlantic Salmon Salmo Salar. Dis. Aquat. Organ 45, 229–236. doi: 10.3354/dao045229
Espmark Å.M., Kolarevic J., Åsgård T., Terjesen B. F. (2017). Tank Size and Fish Management History Matters in Experimental Design. Aquacul. Res. 48, 2876–2894. doi: 10.1111/are.13121
Espmark Å.M.O., Stiller K. T., Shahzad K., Reiten B. K. M., Marchenko Y., Gerwins J., et al. (2020). Nytt S-CCS Konsept for Oppdrett Av Laks-Postsmoltproduksjon i FishGLOBE. Nofima report nr, 35.
Fernandez-Gonzalez V., Ulvan E. M., Sanchez-Jerez P., Diserud O. H., Toledo-Guedes K., Casado-Coy N., et al. (2022). Abundance of Sea Lice Larvae in Plankton Samples: Determination of Optimal Sample Sizes. Aquaculture 551, 737919. doi: 10.1016/j.aquaculture.2022.737919
Fivelstad S., Hosfeld C. D., Medhus R. A., Olsen A. B., Kvamme K. (2018). Growth and Nephrocalcinosis for Atlantic Salmon (Salmo Salar L.) Post-Smolt Exposed to Elevated Carbon Dioxide Partial Pressures. Aquaculture 482, 83–89. doi: 10.1016/j.aquaculture.2017.09.012
Flimlin G., Buttner J., Webster D. (2008). Aquaculture Systems for the Northeast (College Park, MD: Northeast Regional Aquaculture Center).
Galeotti M., Beraldo P., De Dominis S., D'angelo L., Ballestrazzi R., Musetti R., et al. (2000). A Preliminary Histological and Ultrastructural Study of Opercular Anomalies in Gilthead Sea Bream Larvae (Sparus Aurata). Fish. Physiol. Biochem. 22, 151–157. doi: 10.1023/A:1007883008076
Gorle J. M. R., Terjesen B. F., Holan A. B., Berge A., Summerfelt S. T. (2018). Qualifying the Design of a Floating Closed-Containment Fish Farm Using Computational Fluid Dynamics. Biosyst. Eng. 175, 63–81. doi: 10.1016/j.biosystemseng.2018.08.012
Hargis W. J. (1991). Disorders of the Eye in Finfish. Annu. Rev. Fish. Dis. 1, 95–117. doi: 10.1016/0959-8030(91)90025-F
Iversen M., Eliassen R. A. (2009). The Effect of AQUI-S® Sedation on Primary, Secary, and Tertiary Stress Responses During Salmon Smolt, Salmo Salar L., Transport and Transfer to Sea. J. World Aquacul. Soc. 40, 216–225. doi: 10.1111/j.1749-7345.2009.00244.x
Karlsen C., Ytteborg E., Timmerhaus G., Høst V., Handeland S., Jørgensen S. M., et al. (2018). Atlantic Salmon Skin Barrier Functions Gradually Enhance After Seawater Transfer. Sci. Rep. 8, 9510. doi: 10.1038/s41598-018-27818-y
Klebert P., Volent Z., Rosten T. (2018). Measurement and Simulation of the Three-Dimensional Flow Pattern and Particle Removal Efficiencies in a Large Floating Closed Sea Cage With Multiple Inlets and Drains. Aquacul. Eng. 80, 11–21. doi: 10.1016/j.aquaeng.2017.11.001
Klykken C., Reed A. K., Dalum A. S., Olsen R. E., Moe M. K., Attramadal K. J. K., et al (2022). Physiological Changes Observed in Farmed Atlantic Salmon (Salmo Salar L.) With Nephrocalcinosis, in Aquaculture 554, 738104. doi: 10.1016/j.aquaculture.2022.738104
Kolarevic J., Calduch-Giner J., Espmark Å.M., Evensen T., Sosa J., Pérez-Sánchez J. (2021). A Novel Miniaturized Biosensor for Monitoring Atlantic Salmon Swimming Activity and Respiratory Frequency. Animals 11, 2403. doi: 10.3390/ani11082403
Kristiansen D., Lader P., Endresen P. C., Aksnes V. (2018). “Numerical and Experimental Study on the Seakeeping Behavior of Floating Closed Rigid Fish Cages,” in Proceeding Paper in International Conference on Offshore Mechanics and Arctic Engineering American Society of Mechanical Engineers, Vol. 51258, p. V006T05A001.
Lazado C. C., Pedersen L.-F., Kirste K. H., Soleng M., Breiland M. W., Timmerhaus G. (2020). Oxidant-Induced Modifications in the Mucosal Transcriptome and Circulating Metabolome of Atlantic Salmon. Aquat. Toxicol. 227, 105625. doi: 10.1016/j.aquatox.2020.105625
Lazado C. C., Sveen L. R., Soleng M., Pedersen L.-F., Timmerhaus G. (2021). Crowding Reshapes the Mucosal But Not the Systemic Response Repertoires of Atlantic Salmon to Peracetic Acid. Aquaculture 531, 735830. doi: 10.1016/j.aquaculture.2020.735830
Linden S. K., Sutton P., Karlsson N. G., Korolik V., Mcguckin M. A. (2008). Mucins in the Mucosal Barrier to Infection. Mucosal Immunol. 1, 183–197. doi: 10.1038/mi.2008.5
Nagasawa K., Lazado C., Fernandes J. M. (2012). "Validation of Endogenous Reference Genes for qPCR Quantification of Muscle Transcripts in Atlantic Cod Subjected to Different Photoperiod Regimes," in. Aquaculture. doi: 10.5772/28739
Nelson E. J., Robinson S. M. C., Feindel N., Sterling A., Byrne A., Pee Ang K. (2018). Horizontal and Vertical Distribution of Sea Lice Larvae (Lepeophtheirus Salmonis) in and Around Salmon Farms in the Bay of Fundy, Canada. J. Fish. Dis. 41, 885–899. doi: 10.1111/jfd.12692
Nilsen A., Hagen Ø., Johnsen C. A., Prytz H., Zhou B., Nielsen K. V., et al. (2019). The Importance of Exercise: Increased Water Velocity Improves Growth of Atlantic Salmon in Closed Cages. Aquaculture 501, 537–546. doi: 10.1016/j.aquaculture.2018.09.057
Nilsen A., Nielsen K. V., Bergheim A. (2020). A Closer Look at Closed Cages: Growth and Mortality Rates During Production of Post-Smolt Atlantic Salmon in Marine Closed Confinement Systems. Aquacul. Eng. 91, 102124. doi: 10.1016/j.aquaeng.2020.102124
Nilsen A., Nielsen K. V., Biering E., Bergheim A. (2017). Effective Protection Against Sea Lice During the Production of Atlantic Salmon in Floating Enclosures. Aquaculture 466, 41–50. doi: 10.1016/j.aquaculture.2016.09.009
Noble C., Gismervik K., Iversen M. H., Kolarevic J., Nilsson J., Stien L. H., et al. (2018). Welfare Indicators for Farmed Atlantic Salmon: Tools for Assessing Fish Welfare (Tromsø, Norway: Lundblad Media AS).
Noble C., Kadri S., Mitchell D. F., Huntingford F. A. (2007). Influence of Feeding Regime on Intraspecific Competition, Fin Damage and Growth in 1+ Atlantic Salmon Parr (Salmo Salar L.) Held in Freshwater Production Cages. Aquacul. Res. 38, 1137–1143. doi: 10.1111/j.1365-2109.2007.01777.x
Osório J., Stiller K. T., Reiten B.-K., Kolarevic J., Johansen L.-H., Afonso F., et al. (2022). Intermittent Administration of Peracetic Acid is a Mild Environmental Stressor That Elicits Mucosal and Systemic Adaptive Responses From Atlantic Salmon Post-Smolts. BMC Zool. 7, 1–17. doi: 10.1186/s40850-021-00100-x
Pandit D. N., Gupta M. M. L. (2019). Hepato-Somatic Index, Gonado Somatic Index and Condition Factor of Anabas Testudineus as Bio-Monitoring Tools of Nickel and Chromium Toxicity International Journal of Innovations in Engineering and Technology 12, 25–8. doi: 10.21172/ijiet.123.05
Rakers S., Gebert M., Uppalapati S., Meyer W., Maderson P., Sell A. F., et al. (2010). ‘Fish Matters’: The Relevance of Fish Skin Biology to Investigative Dermatology. Exp. Dermatol. 19, 313–324. doi: 10.1111/j.1600-0625.2009.01059.x
Rud I., Kolarevic J., Holan A. B., Berget I., Calabrese S., Terjesen B. F. (2017). Deep-Sequencing of the Bacterial Microbiota in Commercial-Scale Recirculating and Semi-Closed Aquaculture Systems for Atlantic Salmon Post-Smolt Production. Aquacul. Eng. 78, 50–62. doi: 10.1016/j.aquaeng.2016.10.003
Skov P. V. (2019). “"8 - CO2 in Aquaculture,",” in Fish Physiology. Eds. Grosell M., Munday P. L., Farrell A. P., Brauner C. J. (Academic Press), 287–321.
Su B., Kelasidi E., Frank K., Haugen J., Føre M., Pedersen M. O. (2021). An Integrated Approach for Monitoring Structural Deformation of Aquaculture Net Cages. Ocean. Eng. 219, 108424. doi: 10.1016/j.oceaneng.2020.108424
Sveen L., Karlsen C., Ytteborg E. (2020). Mechanical Induced Wounds in Fish – a Review on Models and Healing Mechanisms. Rev. Aquacul. 12, 2446–2465. doi: 10.1111/raq.12443
Sveen L. R., Timmerhaus G., Krasnov A., Takle H., Handeland S., Ytteborg E. (2019). Wound Healing in Post-Smolt Atlantic Salmon (Salmo Salar L.). Sci. Rep. 9, 3565. doi: 10.1038/s41598-019-39080-x
Taylor V., Suter U. (1996). Epithelial Membrane Protein-2 and Epithelial Membrane Protein-3: Two Novel Members of the Peripheral Myelin Protein 22 Gene Family. Gene 175, 115–120. doi: 10.1016/0378-1119(96)00134-5
Timmerhaus G., Lazado C. C., Cabillon N. A., Reiten B. K. M., Johansen L.-H. (2021). The Optimum Velocity for Atlantic Salmon Post-Smolts in RAS is a Compromise Between Muscle Growth and Fish Welfare. Aquaculture 532, 736076. doi: 10.1016/j.aquaculture.2020.736076
Ytrestøyl T., Takle H., Kolarevic J., Calabrese S., Timmerhaus G., Rosseland B. O., et al. (2020). Performance and Welfare of Atlantic Salmon, Salmo Salar L. Post-Smolts in Recirculating Aquaculture Systems: Importance of Salinity and Water Velocity. J. World Aquacul. Soc. 51, 373–392. doi: 10.1111/jwas.12682
Keywords: aquaculture, mucosal health, semi-closed technology, stress, fish welfare, production performance
Citation: Lazado CC, Stiller KT, Shahzad K, Reiten BKM, Marchenko Y, Gerwins J, Radonjic FS, Eckel B, Berge A and Espmark ÅM (2022) Health and Welfare of Atlantic Salmon in FishGLOBE V5 – a Novel Closed Containment System at Sea. Front. Anim. Sci. 3:871433. doi: 10.3389/fanim.2022.871433
Received: 08 February 2022; Accepted: 13 May 2022;
Published: 27 June 2022.
Edited by:
Erin E. Connor, University of Delaware, United StatesReviewed by:
Arve Nilsen, Norwegian Veterinary Institute (NVI), NorwayCopyright © 2022 Lazado, Stiller, Shahzad, Reiten, Marchenko, Gerwins, Radonjic, Eckel, Berge and Espmark. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Åsa Maria Espmark, QXNhLkVzcG1hcmtATm9maW1hLm5v
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.