
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Amphib. Reptile Sci., 06 March 2024
Sec. Behavior and Ecology
Volume 2 - 2024 | https://doi.org/10.3389/famrs.2024.1347541
 Deanna H. Olson1*
Deanna H. Olson1* Evan H. Campbell Grant2
Evan H. Campbell Grant2 Molly Bletz2,3
Molly Bletz2,3 Jonah Piovia-Scott4
Jonah Piovia-Scott4 David Lesbarrères5
David Lesbarrères5 Jacob L. Kerby6Michael J. Adams7Maria Florencia Breitman8
Jacob L. Kerby6Michael J. Adams7Maria Florencia Breitman8 Michelle R. Christman9
Michelle R. Christman9 María J. Forzán10
María J. Forzán10 Matthew J. Gray11Aubree J. Hill12
Matthew J. Gray11Aubree J. Hill12 Michelle S. Koo13Olga Milenkaya14Eria A. Rebollar15
Michelle S. Koo13Olga Milenkaya14Eria A. Rebollar15 Louise A. Rollins-Smith16Megan Serr17
Louise A. Rollins-Smith16Megan Serr17 Alexander Shepack18Leonard Shirose19Laura Sprague20
Alexander Shepack18Leonard Shirose19Laura Sprague20 Jenifer B. Walke21Alexa R. Warwick22
Jenifer B. Walke21Alexa R. Warwick22 Brittany A. Mosher23
Brittany A. Mosher23Western palearctic salamander susceptibility to the skin disease caused by the amphibian chytrid fungus Batrachochytrium salamandrivorans (Bsal) was recognized in 2014, eliciting concerns for a potential novel wave of amphibian declines following the B. dendrobatidis (Bd) chytridiomycosis global pandemic. Although Bsal had not been detected in North America, initial experimental trials supported the heightened susceptibility of caudate amphibians to Bsal chytridiomycosis, recognizing the critical threat this pathogen poses to the North American salamander biodiversity hotspot. Here, we take stock of 10 years of research, collaboration, engagement, and outreach by the North American Bsal Task Force. We summarize main knowledge and conservation actions to both forestall and respond to Bsal invasion into North America. We address the questions: what have we learned; what are current challenges; and are we ready for a more effective reaction to Bsal’s eventual detection? We expect that the many contributions to preemptive planning accrued over the past decade will pay dividends in amphibian conservation effectiveness and can inform future responses to other novel wildlife diseases and extreme threats.
Pervasive global amphibian population declines were recognized by 1990 (Blaustein and Wake, 1990), with subsequent research focusing on main contributing factors (Stuart et al., 2004; Wake and Vredenburg, 2008). After three decades, these declines continue to ‘top the charts’ as one of humanity’s leading global biodiversity concerns (Finn et al., 2023; Luedtke et al., 2023; IUCN Red List, 2023). Infectious disease, along with climate change and habitat loss, has been a leading driver of declines across all continents where amphibians are present (Luedtke et al., 2023). In North American amphibian populations, disease continues to contribute to amphibian declines (Grant et al., 2016a). Herein, we address strategic preparedness for the likely emergence in North America of the disease chytridiomycosis caused by the fungal pathogen Batrachochytrium salamandrivorans (Bsal). The disease Bsal-chytridiomycosis was described over a decade ago (Martel et al., 2013) and predicted to affect palearctic salamanders (Martel et al., 2014), leading to a call-to-arms to protect the North American salamander biodiversity hotspot (Gray et al., 2015).
Although several pathogens have been linked to species- or site-specific amphibian losses (e.g., Ranavirus: Lesbarrères et al., 2012; Perkinsea: Isidoro-Ayza et al., 2017; Isidoro-Ayza et al., 2019), the globalization of the disease chytridiomycosis caused by the aquatic fungal pathogen B. dendrobatidis (Bd) has received the most attention. Unexplained population declines in relatively undisturbed habitats were initially perplexing (Blaustein and Wake, 1990) and several context-specific hypotheses were investigated (review: Alford and Richards, 1999; e.g., acidification: Sadinski and Dunson, 1992; agricultural chemicals: Bishop, 1992; Carey and Bryant, 1995; UV-B radiation: Blaustein et al., 1997). Disease was on the radar, as amphibian mortality from bacterial and viral pathogens was known, but the sentiment at the time was well articulated by Alford and Richards (1999): “Little is known about the diseases of wild amphibians.” In that same year, Bd was described from amphibian skin infections occurring in captive and wild frogs from the Americas and Australia (Longcore et al., 1999).
With research on the origins and effects of Bd in naive systems occurring years after its initial effects were being wrought (e.g., Skerratt et al., 2007; Lips et al., 2008; Carvalho et al., 2017; O’Hanlon et al., 2018), reactive mitigations were required during the height of the Bd panzootic (e.g., Bosch et al., 2001; Central America ex situ captive breeding: Gagliardo et al., 2009; Australia Bd Threat Abatement Plan: Commonwealth of Australia, 2016). Today’s Bd conservation strategies include: metapopulation-scale species reintroduction and management (e.g., Chiricahua Leopard Frog (Lithobates chiricahuensis): U.S. Fish & Wildlife Service, 2007); ex-situ species-rescue measures (AmphibianArk, 2023); and implementation of hygiene protocols to reduce inadvertent human-mediated Bd transmission (Gray et al., 2017; Julian et al., 2020; Olson et al., 2021a; Bletz et al., 2023a; Bletz et al., 2023b).
With this historical context in mind, the more recent emergence of fatal amphibian chytridiomycosis caused by the novel amphibian chytrid fungus Bsal in Europe (Martel et al., 2013; Stegen et al., 2017) with potential palearctic reach (Martel et al., 2014) resulted in an immediate proactive call for strategic efforts in North America (Gray et al., 2015), and a more recent alert echoing concerns for neotropical amphibians (García-Rodríguez et al., 2022). Heightened concerns for the world centers of salamander biodiversity represented by Appalachia and the Pacific Northwest were supported by initial Bsal risk models and maps (Yap et al., 2015; Richgels et al., 2016) that integrated early knowledge of infection risk by host amphibian taxon (Martel et al., 2014) with North American host ranges, environmental conditions, and proximity to international ports and pet stores (as the pet trade was identified as a likely route of introduction to middle European populations: Martel et al., 2013; Martel et al., 2014; see also Cunningham et al., 2017; Nguyen et al., 2017; Fitzpatrick et al., 2018; Yuan et al., 2018). This early work helped inform US (U.S. Fish & Wildlife Service, 2016), Canadian (CBSA, 2018), and pet trade (Pet Product News, 2015; PIJAC Canada, 2016) guidance and policy to reduce imports of suspected Bsal-carrier salamander species (Martel et al., 2014). The North American Bsal Task Force (hereafter: Bsal Task Force; https://www.salamanderfungus.org/) was initiated in June 2015, convening a diverse set of amphibian disease specialists, ecologists, and conservation biologists at a workshop aimed to frame the decisions surrounding an expected Bsal introduction to naive host populations in North America (Grant et al., 2016b; Grant et al., 2017). Multiple working groups were formed to advance knowledge and develop mitigation and management actions (Grant et al., 2016b).
To date, Bsal has not been detected in North America. In the intervening years since the pathogen’s discovery (Martel et al., 2013), proactive research and management advances have been developed by conservation and disease specialists from North American governments, institutions, and organizations including the Bsal Task Force and its partners. Herein we review the state of knowledge and response to the Bsal threat to North American amphibians, with a focus on actions by the Bsal Task Force. We summarize the governance structure of the Bsal Task Force, review the major accomplishments and knowledge advances over the last 10 years by thematic Task Force working groups and others, describe current challenges to maintaining Bsal responsiveness, and discuss how we might collectively better prepare for a potential introduction, which remains a real risk (Grear et al., 2021; Luedtke et al., 2023). Insights from this reflection focus a new lens on emerging priorities and can inform future strategic planning for novel wildlife threats.
The North American Bsal Task Force is a multi-tiered volunteer partnership (Figure 1) with representatives from the United States, Canada, and Mexico. Eight thematic working groups (WGs: Figure 1, bottom) serve as the foundation: response & control, decision science, surveillance & monitoring, research, diagnostics, data management, healthy trade, and outreach & communication. WG participation is intended to be inclusive, with representation of university and government scientists, natural resource managers, pet trade representatives, state and federal resource managers, and students (Bsal Task Force, 2023a). Overall, many more than 100 people have had significant input to Bsal Task Force directions over its first decade, and in numerous ways this group has been a model of collaborative conservation planning, as it has interwoven key sectors of the conservation community. Accomplishments of the Bsal Task Force with its partners are included in the sections below, ordered by the Task Force organizational structure (Figure 1). Summaries of advances include how the Task Force has addressed its guiding principles and main objectives (Table 1) and contributed to the growing Bsal scientific literature (Table 2).
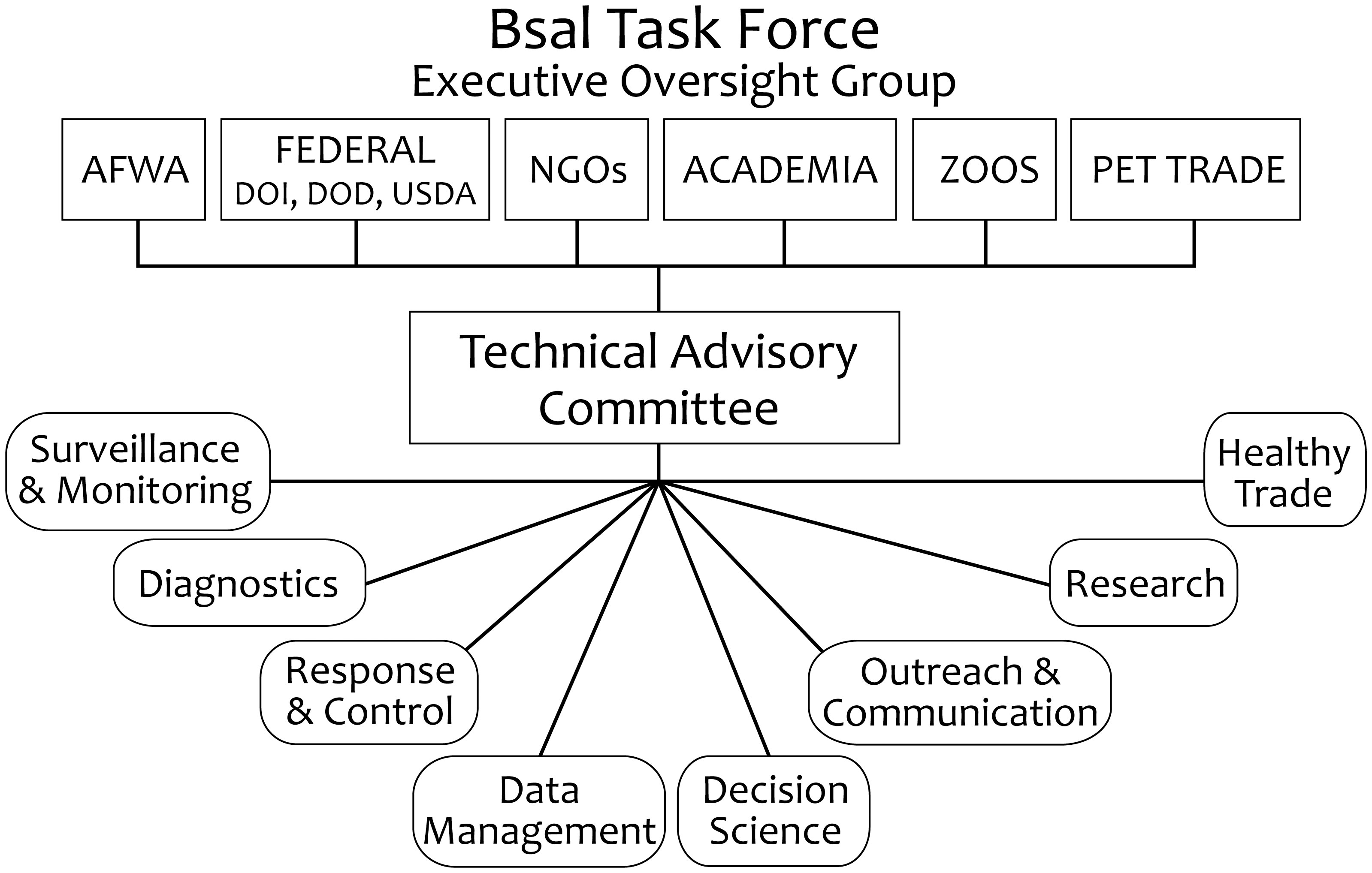
Figure 1 Organizational structure of the North American Bsal (Batrachochytrium salamandrivorans) Task Force, a collaborative group proactively addressing Bsal invasion in advance of arrival to North America.
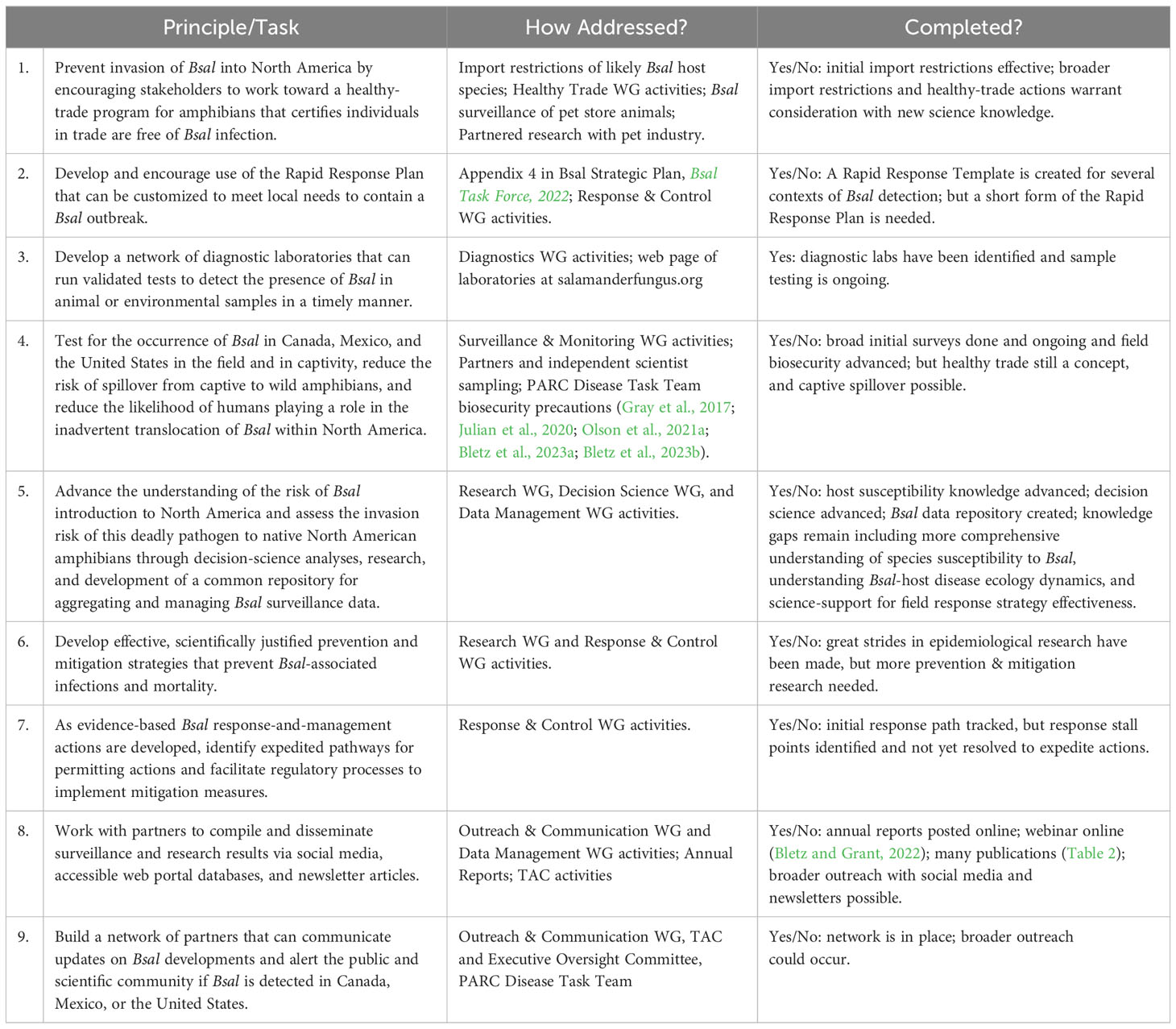
Table 1 Guiding principles and main tasks of the North American Bsal Task Force (Bsal = Batrachochytrium salamandrivorans; Bsal Task Force, 2022).
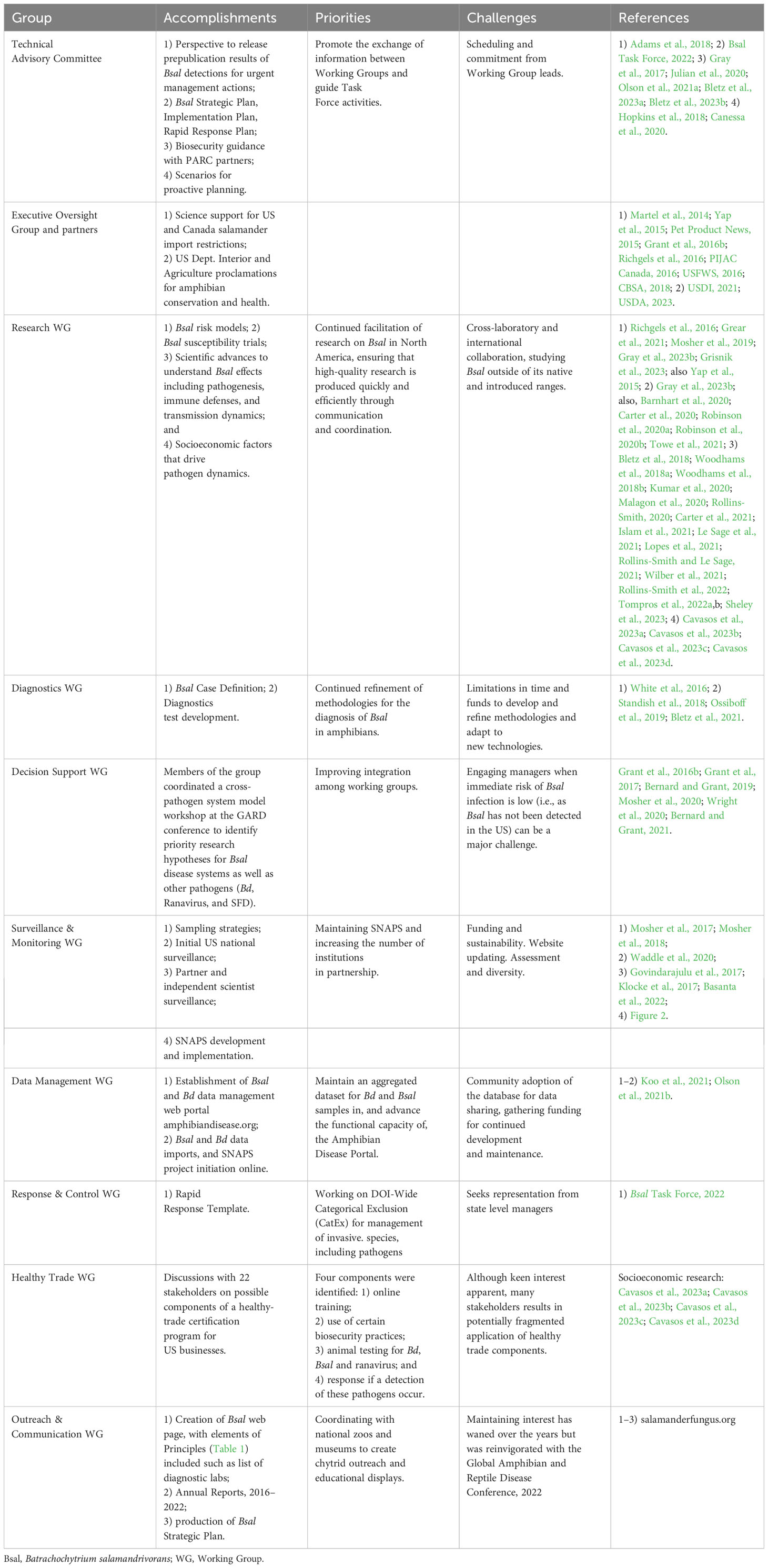
Table 2 Accomplishments of the North American Bsal Task Force members and partners.

Figure 2 Composition and actions of an emergency Incident Command Structure for initial responses if Bsal (Batrachochytrium salamandrivorans) were to be detected in North America (see Hopkins et al., 2018).
The TAC was established to strategically guide the direction of the Bsal Task Force and working group activities, and to improve communication and collaborations among members and partners (Figure 1). With representatives from each working group, it serves a facilitation and oversight function that has resulted in the development of the interdisciplinary aims of the North American Bsal Strategic Plan (Bsal Task Force, 2022), including its adaptive components—the Bsal Implementation Plan and Rapid Response Template. TAC group members have: 1) aided broad participation in WG product development (e.g., Grant et al., 2016b; Grant et al., 2017); 2) contributed to scenario-building exercises to understand how to coordinate for a Bsal die-off response, identifying both an Incident Command Structure (Figure 2) to guide site-specific responses and stall points across potentially complex land- and species-management authorities (e.g., Hopkins et al., 2018; Canessa et al., 2020); and 3) provided conservation perspectives to external science, management, and public groups (e.g., Adams et al., 2018). The TAC had representation at the 2022 Global Amphibian and Reptile Disease conference (Olson et al., 2022; Gray et al., 2023a) and did public outreach during International Amphibian Week kick-off activities at the Smithsonian Institute Museum of Natural History, Washington, D.C. (PARC, 2023b). Cross-membership between the TAC and the Partners in Amphibian and Reptile Conservation (PARC) Disease Task Team, a group addressing multiple herpetofaunal diseases, has resulted in complementary outreach and product development. Examples include development of the Herpetofaunal Disease Alert System (Gray et al., 2018) for early detection of and response to disease-caused die-offs; development of disease biosecurity measures (Gray et al., 2017; Julian et al., 2020; Olson et al., 2021a; Bletz et al., 2023a; Bletz et al., 2023b; webinar: Haman and Henry, 2022); and Bsal outreach by co-production of a “Bsal Basics” webinar (Bletz and Grant, 2022), a Bsal factsheet (PARC, 2022), and a video addressing herpetofaunal disease biosecurity inclusive of Bsal (PARC, 2023a).

Figure 3 Example North American amphibian species identified as susceptible to Batrachochytridium salamandrivorans infection through laboratory trials (Carter et al., 2020; Towe et al., 2021; Gray et al., 2023b): (A) red salamander, Pseudotriton ruber (©T. Pierson); (B) rough-skinned newt, Taricha granulosa with circular, necrotic lesions (©T. Amacker); (C) frosted flatwoods salamander, Ambystoma cingulatum (©M. Mandica); (D) Cuban tree frog, Osteopilus septentrionalis (this species is invasive to the US; ©T. Pierson); (E) eastern newt, Notophthalmus viridescens (©T. Amacker); (F) eastern spadefoot, Scaphiopus holbrookii (©T. Pierson); (G) ensatina, Ensatina eschscholtzii (©I. Parr); (H) green salamander, Aneides aeneus (©T. Pierson); (I) Blue Ridge two-lined salamander, Eurycea wilderae (©K. Stohlgren).
The Executive Oversight Group (EOG) (Figure 1) is intended to improve the communication and coordination of Bsal Task Force activities with other national-scale wildlife governing authorities working to improve amphibian conservation efforts. This extension of the Bsal Task Force is designed to increase awareness of the threat Bsal poses to North American amphibians. The EOG is not intended as a communication channel for advocacy, but rather to inform wildlife and natural-resource managers and policy makers of relevant science advances that may affect their decision making, as per the North American Model for Wildlife Management (Leopold et al., 2018). At this time, the Executive Oversight Group has not been formalized. The reason for this is complex but in part reflects ongoing efforts to develop a broader national-scale approaches to address non-agricultural wildlife health. This is a larger umbrella topic under which amphibian diseases including Bsal-chytridiomycosis could be addressed along with other emerging infectious diseases of nongame native wildlife such as bat white-nose syndrome caused by the introduced fungal pathogen Pseudogymnoascus destructans. (e.g., Cheng et al., 2021). The overarching need remains for an EOG group to be kept abreast of the Bsal threat and related Bsal Task Force efforts, and in-turn provide feedback on management and policy perspectives to the Task Force. Nevertheless, two US Department Secretaries have developed Proclamations for amphibian conservation, including statements of health and welfare priorities (USDI, 2021; USDA, 2023). In lieu of an active EOG, members of the Bsal TAC have served in consultation roles through communication networks across government and science-management partners. The initial publications of Bsal predictions for North America (Martel et al., 2014; Yap et al., 2015; Grant et al., 2016b; Richgels et al., 2016) were key products in this vein, providing important science support for amphibian-import restrictions (U.S. Fish and Wildlife Service, 2016; CBSA, 2018), and a voluntary pet trade import moratorium (Pet Product News, 2015; PIJAC Canada, 2016).
The Research WG has been the most active subgroup of the Bsal Task Force relative to peer-reviewed science-product development (Table 2). Their robust research portfolio aims to fill information gaps about Bsal epidemiology and to better characterize the potential threat of Bsal chytridiomycosis to North American amphibians. The WG has made significant advances in understanding Bsal host-species susceptibility, pathogenesis, immune defenses, and transmission dynamics in susceptible North American and European salamanders. As new information has developed, novel North American Bsal risk models have been created by participants of both the Research and Decision Science WGs (US: Grear et al., 2021; Gray et al., 2023b), as well as by independent scientists and partners (Mexico: Basanta et al., 2019; Ontario, Canada: Crawshaw et al., 2022; US: Moubarak et al., 2022). Also, socioeconomic research has been initiated to address biosecurity practices in US pet amphibians (see Healthy Trade WG; Cavasos et al., 2023a; Cavasos et al., 2023b; Cavasos et al., 2023c) and the role of visitors in preventing pathogen threats in natural areas (Cavasos et al., 2023d). Specific examples follow.
Bsal susceptibility trials in 35 North American amphibian species (10 families) showed that infection was detectable in 74% of species and deaths were observed in 35% of species (Gray et al., 2023b), building on the earlier studies such as Martel et al. (2014); Friday et al. (2020); Carter et al. (2020), and DiRenzo et al. (2021) that also documented susceptibility in a diversity of North American species (Table 3; Supplementary Materials). These results have supported the contention that many North American amphibian species are at risk for population declines, with the greatest predicted biodiversity losses in the Appalachian Region and along the Pacific Coast, confirming the expectations from the early Bsal risk maps (e.g., Richgels et al., 2016). Susceptible species (Supplementary Materials; Figure 3; Table 3) have included both salamanders (see also Carter et al., 2020) and anurans (see also Towe et al., 2021), including species that commonly occur in the pet trade (e.g., Osteopilus septentrionalis, Ambystoma mexicanum; Gray et al., 2023b). Some of these species were susceptible to Bsal infection and may be carriers, as not all tested species developed lethal disease in laboratory trials. Ongoing studies focus on Bsal-infection and -chytriodiomycosis susceptibility in additional native North American amphibian species as well as other species that occur in the pet trade.
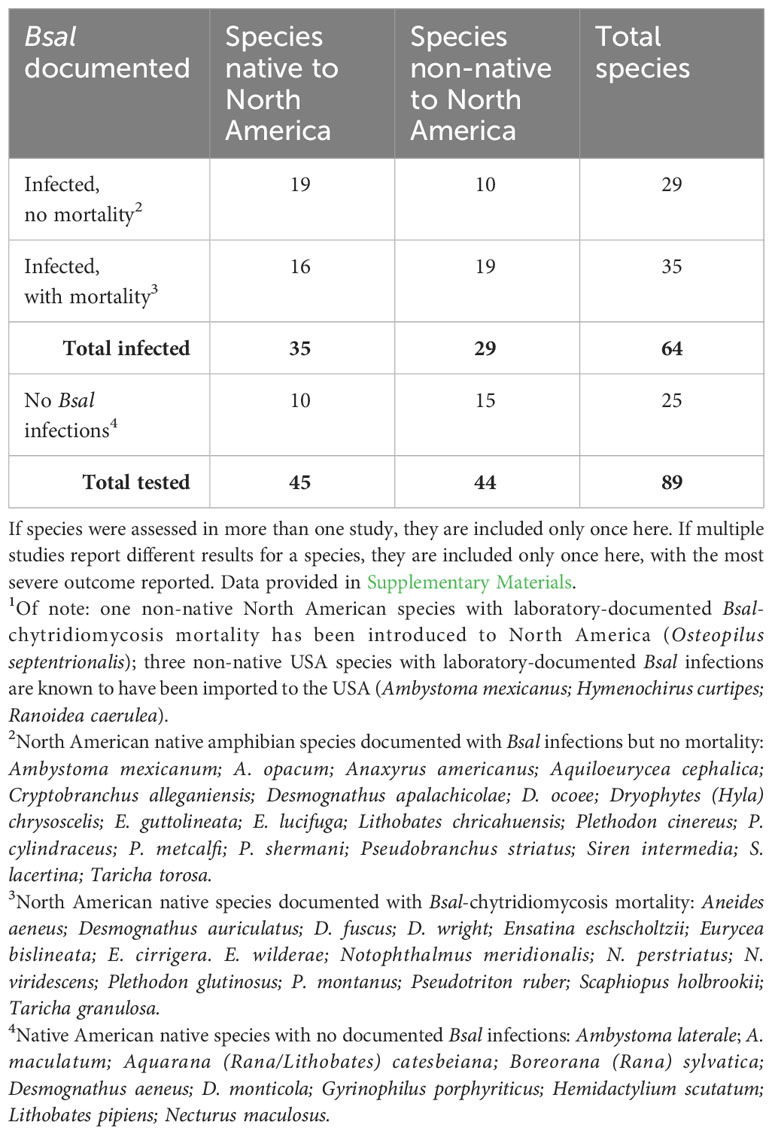
Table 3 Number of native and non-native North American amphibian species that have been laboratory-documented with Batrachochytrium salamandrivorans (Bsal) infections with and without mortality, and with no Bsal infections1.
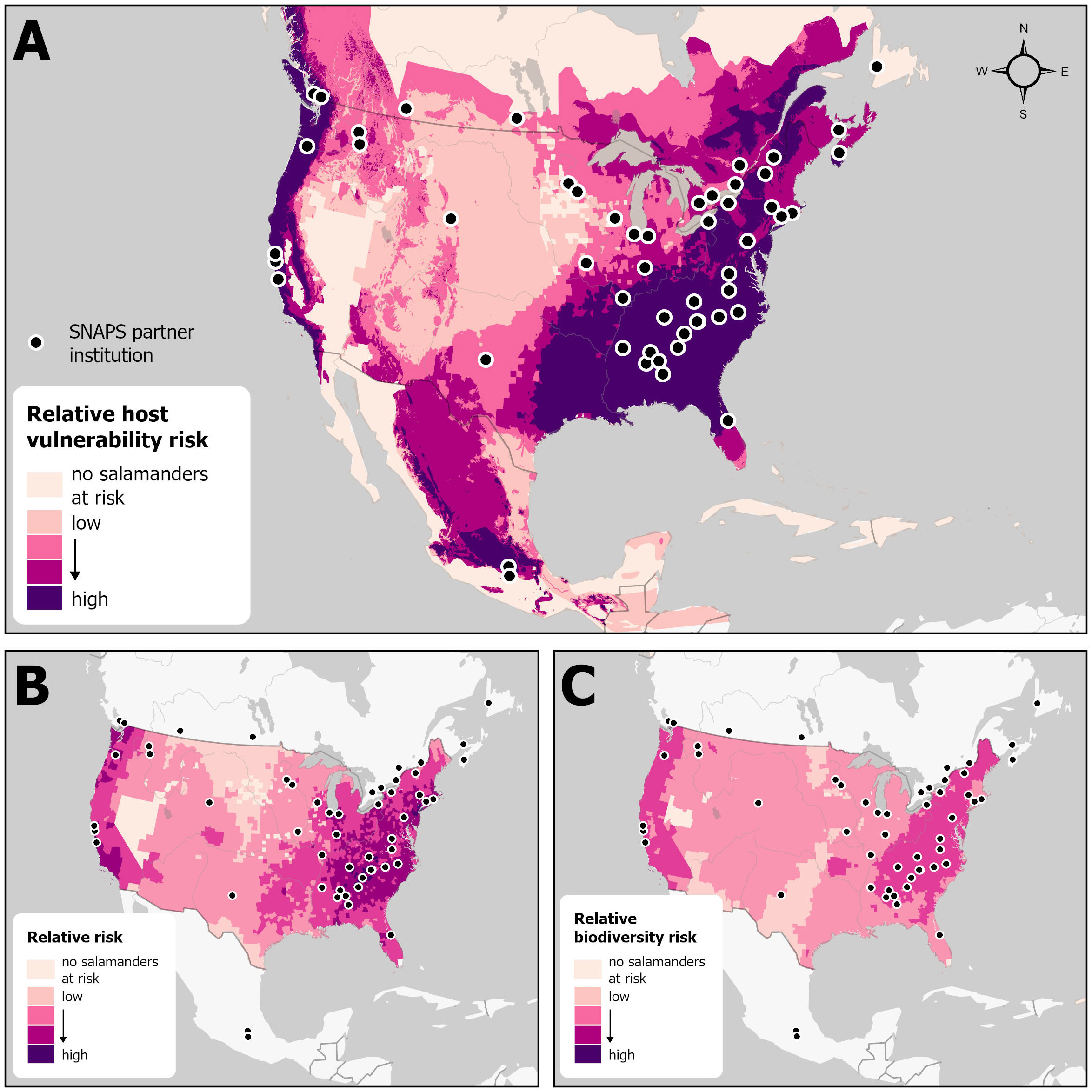
Figure 4 Locations of 54 SNAPS (Student Network for Amphibian Population Surveillance, Surveillance and Monitoring Working Group, North American Bsal Task Force) partnering institutions (2 in Mexico, 10 in Canada, 40 in USA) overlaying maps showing three predictive risk models of relative host susceptibility to Batrachochytrium salamandrivorans (Bsal; darker shades denote higher risk): (A) before implementation of import restrictions (from North America model by Yap et al., 2015); (B) after implementation of import restrictions (only the US was modeled: Grear et al., 2021); (C) with updated Bsal-host susceptibility data (only the US was modeled: Gray et al., 2023b). The lightest shade on the maps denotes that no salamanders are at risk in that geographic area.
Using Bsal-susceptible rough-skinned newts (Taricha granulosa) and eastern newts (Notophthalmus viridescens) as model species, critical information about the mechanisms of Bsal-induced physiological changes and immune defenses have been revealed. For example, to address the question of what may lead to death from Bsal chytridiomycosis, an extensive study of the blood cells, blood chemistry, and histological changes at various stages of disease progression was conducted in the rough-skinned newts. Electrolyte imbalances were associated with clinically diseased newts as compared to non-diseased newts, and as Bsal loads increased, blood potassium levels increased, sodium and chloride ion concentrations decreased, and disruption of the skin led to dehydration and potential disruption of granular glands that produce defensive peptides (Sheley et al., 2023). Studies of the capacity of skin secretions from additional salamander species to inhibit Bsal in vitro showed mixed results, with some species showing effective secretions whereas others had less-effective secretions (Pereira et al., 2018; Smith et al., 2018; Carter et al., 2021; Pereira and Woodley, 2021; Hardman et al., 2023). Although clinically diseased rough-skinned newts showed evidence of a systemic inflammatory response, they were unable to clear the infection prior to death (Sheley et al., 2023). Although Bsal can induce immune responses, the pathogen can also inhibit lymphocyte responses, suggesting local immune suppression (Rollins-Smith et al., 2022).
In a comprehensive study of temperature effects on susceptibility of eastern newts to Bsal (Carter et al., 2021), greater mortality of both eft-stage newts and adult newts was observed at 14°C in comparison with 6°C or 22°C. Because 22°C is approaching the upper limit of Bsal growth, there were no persistent Bsal infections at this temperature except in efts exposed to the highest initial dose. When eastern newts were exposed to relatively high numbers of zoospores (5 x 104) and maintained at a cold temperature (6°C), their production or release of putative defensive peptides from granular glands was reduced in comparison with unexposed controls. The relative abundance of bacteria inhibitory to Bsal was greater at 22°C as compared to the lower temperatures of 14°C and 6°C, suggesting that temperature-mediated shifts in the skin microbiome may contribute to host protection at warmer temperatures in eastern newts. This supports an earlier study of the skin microbiome of fire salamanders (Salamandra salamandra) in Europe in which wild salamanders had anti-Bsal bacteria in their communities but at low abundance, and the experimental increase of these anti-Bsal bacteria by daily additions slowed disease progression (Bletz et al., 2018). More broadly, changing climatic conditions may lead to some decreases in climatic suitability for Bsal in the United States (Grisnik et al., 2023).
Lastly, research has investigated modes of Bsal transmission and utility of some applied management strategies. Transmission between individual amphibian hosts can occur by swimming zoospores or by direct contact of infected individuals with susceptible hosts (Malagon et al., 2020; Tompros et al., 2022a). In eastern newts, all susceptible hosts were infected following one or more contacts with an infected host, resulting in 89% mortality over three months (Malagon et al., 2020). Even one contact for about one second was sufficient to result in transmission (Malagon et al., 2020). Although host-contact rates are density-dependent (Malagon et al., 2020), further analysis suggested that transmission is frequency-dependent (Tompros et al., 2022a). Relative to considerations for applied management actions to reduce Bsal effects on susceptible hosts, introduction of artificial plants to increase habitat complexity reduced host contacts but did not markedly reduce mortality, and transmission remained high (Malagon et al., 2020). Possible management strategies for Bsal include use of plant-derived fungicides (Tompros et al., 2022b) or probiotics that may reduce Bsal transmission or possibly treating Bsal-contaminated habitats or hosts (also see below, Decision Science WG; Bernard and Grant, 2021). The management response to Bsal detection at a field site in Spain 1000 km from the nearest European Bsal outbreak included biosecurity measures, habitat management and disinfection, host removal, and disease surveillance, resulting in temporary Bsal containment (Martel et al., 2020). This response can inform North American approaches, as can additional approaches considered for Bd management (Garner et al., 2016; Mendelson et al., 2019). Additional research guided by management-driven needs identified by the Research, Decision Science (e.g., Bernard and Grant, 2019; Bernard and Grant, 2021), Response and Control, and Healthy Trade WGs will help to advance conservation efficacy in North American jurisdictions.
Despite a decade of progress, much remains to be learned about amphibian immune defenses against Bsal, including innate defenses (e.g., antimicrobial peptides), adaptive immune defenses (lymphocyte-mediated), and the role of the microbiome. Identifying strategies to boost host immunity (e.g., use of probiotics, vaccination) or altered environmental characteristics (e.g., habitat complexity, temperature) to protect vulnerable species is a top priority. Identifying additional microbes that could be used to modify naturally occurring microbial communities in the skin mucus (e.g., a probiotics approach) to protect highly vulnerable species, while challenging, merits exploration. Investigating how host-amphibian community structure influences the impacts of Bsal in a natural setting could inform mitigation efforts if Bsal is accidentally introduced to susceptible North American communities. Because many North American amphibian species are also infected with Bd, determining how a dual infection with both Bd and Bsal would affect amphibian communities is another high-priority challenge. One study of experimental dual infections in eastern newts suggested that simultaneous Bd and Bsal infections were more harmful than single infections (Longo et al., 2019).
The Diagnostics WG includes interdisciplinary experts who regularly examine specimen samples for the presence of amphibian pathogens. An initial critical task for the group was to reach consensus on a Bsal case definition to establish diagnostic criteria for validating a positive Bsal-chytridiomycosis DNA detection and develop criteria for establishing infection and disease presentation (White et al., 2016). This effort aided standardized communication of results and consistent reporting among diagnostic laboratories to provide reliable information to amphibian-disease ecologists and wildlife managers. Although many options exist for laboratory diagnostic methods for Bsal DNA detection in a specimen sample, the ‘gold standard’ relies on molecular methods with quantitative polymerase chain reactions (qPCR) for verification of Bsal (Blooi et al., 2013; Martel et al., 2013). Critical to this designation is the recognition that a positive Bsal-DNA result from one diagnostic laboratory does not necessarily imply an infected population.
Although qPCR tests for Bsal DNA were established upon recognition of the novel pathogen (Blooi et al., 2013; Standish et al., 2018; Thomas et al., 2018), the use of different protocols among laboratories has raised concerns regarding the consistency of results (e.g., Iwanowicz et al., 2017). In 2020, with the support of Environment and Climate Change Canada, the Diagnostics WG initiated a round-robin test among over 40 diagnostic and research laboratories worldwide (Bsal Task Force, 2023b). The round-robin exercise aimed to test the proficiency of participating laboratories running qPCR tests to detect Bd and Bsal, either individually (single qPCR) or together (duplex qPCR). Blind samples consisting of various concentrations of the chytrid fungi dripped onto cotton swabs (to mimic samples obtained from swabbing amphibians), were shipped to participants, and results were collated and analyzed. A summary of the results, which was shared with all participants maintaining the anonymity of individual values, resulted in a high level of participation (over 95% of laboratories who received samples reported results), and highlighted discrepancies such as false negative and false positive results. Perhaps the most striking inconsistency related to the calculation of DNA copies per sample: calculations varied by up to three orders of magnitude among some laboratories. Although no specific explanation for this inconsistency has been proposed, the results of the round-robin test are currently being analyzed in the hope of detecting factors that may have caused laboratories to report such varying numbers of DNA copy numbers. Results of these ongoing analyses along with the methodology and results of the round-robin are planned for independent publication. In addition, recently, a non-profit organization (Healthy Trade Institute, Inc., HTI) was created to help facilitate incorporation of biosecurity practices in US pet amphibian businesses and reduce the likelihood of chytrid fungi and ranaviruses in trade. The HTI will be recruiting laboratories to run diagnostic samples associated with a Healthy Trade Certification Program. Diagnostic competency of the laboratories will be verified following a similar round-robin approach.
Owing to the high expected consequences of a Bsal-positive detection in North America, uncertain data resulting from sample contamination during either field collection or laboratory procedures, cross-reactivity of other sample components (e.g., Iwanowicz et al., 2017), or reduced sensitivity of the laboratory test conducted, Bsal-positive detections by two independent laboratories have been incorporated into the Bsal Task Force Rapid Response Template (Bsal Task Force, 2022). In Europe, diagnostic recommendations for Bsal detection were developed by Thomas et al. (2018) and similarly included duplicate samples from live animals for qPCR testing as per procedures by Blooi et al. (2013), and post-mortem screening of skin-swab samples and histopathology confirming the presence of the fungus in the skin. The Diagnostics WG continues to focus on the development of new techniques and technologies. For example, the more-recent development of an in situ hybridization protocol to detect and differentiate Bd and Bsal in histologic samples has increased the reliability of histopathologic diagnosis of Bsal infection (Ossiboff et al., 2019). As a service to the Bsal science-and-management community, for those interested in Bsal testing options, the Diagnostics WG maintains a current list of open diagnostic laboratories on the Bsal Task Force website (diagnostics.salamanderfungus.org).
Decision science is an interdisciplinary field with associated tools for framing and solving complex decision problems and is particularly useful for decisions with large uncertainties. For example, where multiple uncertainties exist, decision theory offers tools to quantify and rank the relative importance of knowledge gaps that impede identifying the optimal management strategy to reduce the risk of Bsal. Further, natural-resource managers must often consider multiple social, economic, and ecological objectives in their decisions, and these may induce difficult trade-offs for any given disease-management strategy. To this end, members of the Decision Science WG bring expertise to support management decisions where Bsal is a known or potential risk for native amphibian population declines.
Decision Science WG accomplishments align with its four chief aims: 1) identify critical research that has the highest information value, which will lead to an improved ability to manage Bsal; 2) identify approaches to improve proactive management of Bsal when uncertainty or competing objectives are impediments to action; 3) conduct and update Bsal risk assessments; and 4) work with management agencies to help frame and solve Bsal management problems. Addressing the first aim, Grant et al. (2016b) identified that the most important proactive conservation mitigation would be to reduce Bsal introduction to North America, providing science support to inform key policy decisions such as the restriction of salamander imports to North America. Correspondingly, in 2015 The Pet Advocacy Network in the US recommended a temporary and immediate voluntary trade moratorium on imports of salamander species imported from Asia that were known to carry Bsal until such time as effective testing and treatment regimens could be developed and distributed (Pet Product News, 2015). In Canada, a similar temporary moratorium was supported for a ban on importation of fire belly and paddle-tail newts (Cynops spp. and Pachytriton labiatus, respectively) native to Asia, which were possible vectors of Bsal (PIJAC Canada, 2016). The US enacted an interim rule of the Lacey Act that banned importation of 201 species of salamanders (U.S. Fish and Wildlife Service, 2016). Canada implemented a permit requirement for the importation of all salamanders whether live or dead (CBSA, 2018).
Addressing the second aim, Bernard and Grant (2019) worked closely with managers to identify decision-problem elements common to both Bsal chytridiomycosis and bat white-nose syndrome, which help to understand impediments to making management decisions. Importantly, whereas research uncertainties are often initially identified as key impediments to mounting a management response, Bernard and Grant (2019) found that other challenges existed; addressing the full complexity of the challenging decisions for Bsal management is expected to lead to improved management responses. Indeed, although uncertainty is an important impediment, the working group proposed that existing models and information can be used to make useful predictions for developing and assessing management strategies (Russell et al., 2017).
To address the third aim, Richgels et al. (2016) developed an initial risk assessment that used environmental suitability for Bsal and host-species richness in the US, combined with risk of introduction from the pet trade, to help inform US Fish and Wildlife Service’s decision-making process for evaluating a ruling under the Lacey Act. This assessment had general agreement to that of Yap et al. (2015), conducted independently, which covered a similar geography using a different approach. The Richgels et al. (2016) county-level risk model informed US surveillance efforts for Bsal (Waddle et al., 2020). Subsequently, Grear et al. (2021) combined the ban on importation of amphibians under the Lacey Act with US surveillance data (Waddle et al., 2020) to update a risk model to inform additional surveillance efforts (see next section) and to identify areas of highest importance for the development and implementation of proactive management actions. The recently updated risk-assessment model by the Research WG (Gray et al., 2023b) added specificity to species-risk predictions resulting from knowledge gains from experimental trials and supports main risk geographies in Richgels et al. (2016).
Finally, to address the fourth aim of the WG, consultations with national-resource managers have identified common concerns and perceived impediments to Bsal management. More in-depth work with multiple resource managers (e.g., US Fish and Wildlife Service; US Forest Service; State of New Mexico) has helped to structure their decision-making processes. The WG used a formal process of expert judgment to parameterize a predictive model to help quantitatively evaluate management actions proposed among TAC working groups (Bernard and Grant, 2021).
The decision science working group will continue to work with resource management agencies to frame and help solve their Bsal decision problems, revealing critical uncertainties that can be addressed by other Bsal Task Force working groups (e.g., Research, Surveillance and Monitoring). Development of decision support tools and synthesis of decision challenges will aid in expanding the utility of the group’s decision analysis work.
The mission of the Surveillance & Monitoring WG is to facilitate and coordinate surveillance for Bsal across North America, and the monitoring of Bsal in the event of its detection. Early detection of Bsal would allow for a rapid response to safeguard native fauna, but achieving a broad and robust surveillance network is difficult and expensive because of the human resources needed, including the costs of laboratory testing. No single organizational entity has this capacity. Instead, the emphasis of the Surveillance & Monitoring WG has been to coordinate and encourage sampling for Bsal by diverse partners, with broad spatiotemporal scope in consideration of high-risk species and geographies (e.g., Yap et al., 2015; Richgels et al., 2016; Basanta et al., 2019; Grear et al., 2021; Gray et al., 2023b; Grisnik et al., 2023).
Initial WG efforts toward this goal included a one-time sampling effort conducted by the US Geological Survey Amphibian Research and Monitoring Initiative (ARMI) from 2014 to 2017 (Waddle et al., 2020), including sample sites across the US and resources allocated according to the estimated risk of Bsal occurrence (Richgels et al., 2016). Over 10,000 amphibians (mostly salamanders) were sampled and Bsal was not detected (Waddle et al., 2020; data archived at amphibiandisease.org by the Data Management Working Group, see below). ARMI and the US Fish and Wildlife Service have continued to sample at a very low level in selected areas where resources allow. However, a targeted effort is ongoing in Appalachia, which is one of the projected high-risk areas in the US (Yap et al., 2015; Richgels et al., 2016; Gray et al., 2023b; Grisnik et al., 2023).
More recently, the emphasis of the Surveillance & Monitoring WG has been to coordinate and facilitate Bsal sampling with broad spatiotemporal scope in consideration of high-risk species and locations. To this end, the WG has successfully developed, launched, and maintained the Student Network for Amphibian Pathogen Surveillance program (SNAPS; https://snaps.amphibiandisease.org/), which aims to increase Bsal sampling and awareness, engage diverse partners and citizen scientists, and utilize dispersed in-kind resources. SNAPS is a cross-continental network of students, scientists, and educators committed to the conservation of amphibians against the threat of emerging pathogens, with an initial focus on Bsal (Figure 4). The SNAPS approach is to embed Bsal surveillance into undergraduate college and university curricula where students learn about the amphibian-chytrid disease system, One Health, biosecurity protocols, and data collection, while contributing to a meaningful conservation initiative. Bsal sampling is a straightforward procedure that involves swabbing the skin of amphibians and is an ideal field activity for novices with training and supervision by their instructors. The network facilitates meaningful field and research experiences for students while harnessing their enthusiasm and geographic breadth to surveille for Bsal across the continent. To date, more than 2,300 amphibians have been sampled (see our open-access data portal at https://amphibiandisease.org/projects/?id=284), with a focus on key geographies that have been flagged as high-risk for Bsal introduction (Yap et al., 2015; Basanta et al., 2019), and 600+ students from more than 70 institutions have participated in sampling events across Canada, the US and Mexico (Figure 4).
Recruitment of additional schools to participate in SNAPS is ongoing although obtaining long-term funding and diversifying participation in SNAPS is challenging. Field-based courses and authentic research experiences for underserved, underrepresented students can help close demographic gaps in Science, Technology, Engineering, and Mathematics (STEM) related fields, foster a sense of self-efficacy and belonging, and enhance feelings of comfort in the outdoors (Zavaleta et al., 2020). The WG’s goals include strategically expanding and diversifying the SNAPS network to include students from populations that are underrepresented in STEM, introductory-level students, students with career interests outside the fields of conservation and ecology, and a greater variety and larger number of institution types, including field stations, community colleges, Historically Black Colleges and Universities, Hispanic-Serving Institutions, R2 institutions and Indigenous First Nations, through Diversity, Equity, and Inclusion initiatives described by Ruggs and Hebl (2012).
Early Bsal detection through surveillance of both captive and wild amphibians is also being conducted by partners and independent scientists (e.g., Canada: Govindarajulu et al., 2017; Fieschi-Méric et al., 2023; Mexico: Basanta et al., 2022; US: Klocke et al., 2017; Gluesenkamp et al., 2018; Hill et al., 2021). A recent study funded by the US National Science Foundation identified factors contributing to amphibian pathogen movements through US trade networks (https://www.healthyamphibiantrade.org). As new surveillance projects develop, key considerations include the diagnostic uncertainties stemming from both sampling and laboratory procedures discussed in the previous section, and the need for sufficient sample sizes per relevant unit of study such as species, life-history stage, or site (e.g., Skerratt et al., 2008; Gray et al., 2017). In addition to the mix of coordinated and independent surveillance by sampling wild or captive amphibians, rapid-response networks have been initiated to report amphibian disease die-offs that might be caused by Bsal, including direct communication with state or provincial authorities or Bsal Task Force members, or use of PARC’s Herpetofaunal Disease Alert System (HDAS; Gray et al., 2018: To report, email disease reports toaGVycF9kaXNlYXNlX2FsZXJ0QHBhcmNwbGFjZS5vcmc=for them to forward to state and provincial authorities; include photograph, species affected, location, other relevant episode information, observer’s name). If sufficient information is provided, social media reports can be routed to HDAS. Lastly, Bd detection has been performed on all samples tested for Bsal, thus expanding Bd monitoring efforts in North America.
The focus of the Data Management WG has been to develop an online Bsal and Bd data aggregation portal to accelerate sharing of planned or completed surveillance projects and scientific studies and to facilitate the pace of learning about these pathogens. Several key products have been completed: 1) establishment and a description of the structural underpinnings of the Bsal and Bd Amphibian Disease web portal (amphibiandisease.org), with an important roadmap for the incorporation of the legacy Bd-maps.net dataset (Koo et al., 2021); 2) imports of USGS survey results for Bsal in 2014–2017 (USA: Waddle et al., 2020), results from the Bsal Consortium of Germany (see special edition of Salamandra, Volume 56[3]), and integration of the SNAPS effort as an ongoing project in the web portal (Figure 4); and 3) a global Bd update (Olson et al., 2021b) with transference of the Bd-maps.net database (formerly archived at Imperial College, London, UK: Olson et al., 2013) updated through 2019 to the new web portal, inclusive of database DOI references. Although the Bsal data repository is growing, the utility of our web portal to the world community is supported by recent Bd metadata analyses and risk-assessment publications using exported globally compiled data archived in the Amphibian Disease database that focus conservation-planning attention on more defined geographies and taxa (e.g., Suriname: Rawien and Jairam-Doerga, 2022; West Africa: Ghose et al., 2023).
Features of the Amphibian Disease web portal are multifaceted. Planned and completed projects can be included, increasing research and management efficiencies across taxa and geographies. Portal data include the capacity for field, captive, and museum-specimen amphibian reports, eDNA results from water samples, pathogen strain identification, and inclusion of infection intensity (zoospore load) and disease symptoms. The portal’s species-specific data summary pages can be dynamically linked to external websites (e.g., AmphibiaWeb) so users can see if any amphibian has been tested for chytridiomycosis. Partnering with the global bio-sampling database Geome (geome-db.org) has enhanced research and forecasting abilities by delivering improved validation services, enhancing security, and improving data accessibility through third-party applications and programming tools.
Ongoing and planned work includes maintenance and web programming tasks to expand data visualizations of the Bd and Bsal data such as country- and species-summary charts and custom query interfaces. Collaboration with the Bsal Surveillance and Monitoring WG has provided a central repository for a network of North American colleges and universities engaged in Bsal surveillance so educators can focus on curricula and simply use the AmphibianDisease portal’s features. Updating data and advancing the functional capacity, especially as an educational tool, of the Amphibian Disease Portal is on the horizon. Further, links to other online scientific web portals would help extend the portal’s reach to other audiences in education and conservation. Expanded outreach to surveillance communities to report projects, disease symptoms, strains, and zoospore loads is needed, inclusive of outreach to journals and permitting agencies to help with portal messaging. Many of these plans highlight the ongoing funding needs for development, which the Data Management Working Group seeks through collaborative grants and strategic partnerships.
The aims of the Response & Control WG have changed as both proactive and reactive management responses to Bsal have been addressed in tandem. The Bsal Rapid Response Template created by the initial Response WG (Grant et al., 2016b) was one of the most urgent initial priorities of the Task Force (Table 1). This template was intended to be updated and improved over time. The most recent revision occurred in 2019 and is included as an Appendix in the 2022 North American Bsal Strategic Plan (Bsal Task Force, 2022). The template considers scenarios of Bsal detection in both wild and captive situations and is intended to be customized to different users and for different contexts of Bsal detection to support a more efficient and rapid response. Scenarios include: a mortality event in the wild or in captivity; non-definitive Bsal detection from a laboratory PCR test result (wild or captive); and a definitive Bsal detection in the wild or captivity. Important components of this template include: 1) a Bsal-positive result from laboratory analysis of a skin swab or eDNA sample is not considered a “definitive detection,” and confirmation by a second independent laboratory is needed (see Diagnostics WG section above); 2) collection of tissue and/or whole animals suspected to be infected with Bsal for diagnostic testing can aid accurate, confirmed diagnosis; 3) directions for swab or tissue collection are included in the template; 4) establishment of biosecurity measures to reduce potential human-mediated Bsal transmission; and 5) mandatory notification of the agency or agencies with management jurisdiction of the site (i.e., initiation of an Incident Command Structure, Figure 2; Hopkins et al., 2018), which can trigger national and international coordination. For example, an immediate report by a US State Animal Health Professional to the US National Animal Health Reporting System (NAHRS) can help advise development of the Incident Command Structure (Figure 2) to help coordinate communication and actions. As this unfolds, consultation with members of the Bsal Task Force can expedite planned response actions in the context of the detection. As Bsal is listed as a reportable disease with the World Organization for Animal Health (WOAH, 2023), this reporting step would be followed by the national authority.
Currently, the Response & Control WG is focused on: 1) creating a more direct version of the Rapid Response Template resolving likely stall points in rapid, effective Bsal responses after initial detection; and 2) exploring tools or systems to make the response template (or its contents) easier to access or use. Initial stall points have been identified in various scenario exercises (e.g., Hopkins et al., 2018; Canessa et al., 2020), and include regulatory authorities and restrictions, legal issues, and permissions or authorizations necessary to implement field or captive response measures. As these scenarios have unfolded, it has become apparent that numerous and different authorities may need to be consulted, and guidance for approved actions followed per authority. One focus of the WG supports the efforts of the US Fish and Wildlife Service in development of a national-scale US Department of Interior Categorical Exclusion (CatEx) for management of invasive species, including pathogens. Acceptance of the CatEx would be instrumental in rapid-response efforts. Ongoing work also includes development of easy-to-follow Incident Command Structures (Figure 2) similar to the US Federal Emergency Management Agency (FEMA) for disaster response or Early Detection Rapid Response (EDRR) templates created for invasive-species control. Lastly, synergies with the Research and Decision Support WGs are planned as new knowledge of amphibian host susceptibility is forthcoming, and procedural dynamics of a rapid response are further defined.
With the growing concern that Bsal could be translocated to North America via trade, the Healthy Trade WG was created in 2020. This WG includes members from the Pet Advocacy Network, US pet amphibian businesses, and academia. Originally called the Clean Trade WG, we renamed it because the intent of the group is not to promote clean/sterile trade, but rather pathogen-free/healthy trade. By pathogen-free, this group is most concerned with Bsal, but there is also plenty of evidence that Bd and ranaviruses are being translocated globally through amphibian trade (Schloegel et al., 2009; Kolby et al., 2014). Most nations do not require animal-health certificates for globally traded amphibians (Grant et al., 2017), despite Bd, Bsal and ranaviruses being listed as notifiable pathogens by the World Organization for Animal Health (e.g., WOAH, 2023). Nevertheless, methods to expedite detection of disease-causing pathogens in trade are advancing (e.g., Brunner, 2020).
A major concern with domestic and international trading of amphibians infected with novel pathogens or variants is spillover to wild populations and native biodiversity loss, which is hypothesized to be how Bsal emerged in Europe (Martel et al., 2014). Pathogens in trade also cause major losses to industry. As such, it is not surprising that most US businesses and consumers support acquiring pathogen-free amphibians (Cavasos et al., 2023a; Cavasos et al., 2023b). In fact, US pet-amphibian consumers are willing to pay up to 75% more for certified pathogen-free amphibians compared to amphibians with unknown health status (Cavasos et al., 2023c). Thus, in 2022, the Healthy Trade Working Group began discussions with 22 stakeholders on possible components of a healthy-trade certification program for US businesses. Four components were identified: online training; use of certain biosecurity practices; animal testing for Bd, Bsal and ranavirus; and response if these pathogens were detected. Subgroups were subsequently created for each of the plan components to draft details, and a follow-up survey is ongoing to receive feedback from US businesses and consumers on the draft plan (https://www.healthyamphibiantrade.org).
Planned efforts of the Healthy Trade WG include working with Pet Advocacy Network, industry, and academic partners to hopefully launch a market-driven US healthy trade certification program for pet amphibians in the near future. Through socioeconomic surveys (Cavasos et al., 2023a; Cavasos et al., 2023b; Cavasos et al., 2023c) and over two years of discussions with industry stakeholders, the Healthy Trade WG has learned that US pet amphibian businesses are supportive of healthy trade practices, especially in comparison to trade bans, which the federal government has used in the past (U.S. Fish and Wildlife Service, 2016).
The Communication & Outreach WG supports Task Force activities and goals through the production and promotion of Bsal resources and materials for scientists, managers, and the general public (Table 2). The WG maintains the Task Force website (salamanderfungus.org), which provides outreach communication materials, including the strategic plan and Rapid Response Template, recent highlights, links to resources provided by other WGs (e.g., diagnostic laboratory information; the Bsal data management system: amphibiandisease.org), and annual reports. Initial communications provided outreach to wildlife managers, herpetofaunal conservation groups, and the public (e.g., Olson, 2015). Bsal Task Force annual reports are co-produced by the Amphibian Foundation (amphibianfoundation.org) to summarize the current status of the Bsal fungus and to provide WG updates on accomplishments and new directions in work portfolios. The WG collaborates with partners to produce and distribute materials including scientific and more general articles, flyers, and social media. An annual pulse of communication activities coincides with International Amphibian Week activities promoted by Partners in Amphibian and Reptile Conservation (PARC, 2023b), which the Bsal Task Force supports through a Twitter [now X] account managed by the WG (twitter.com/salamanderfungi). The PARC Disease Task Team provides complementary outreach materials for Bsal-chytridiomycosis and other diseases (e.g., PARC, 2023a). To increase awareness, the WG works with Bsal Task Force members to coordinate outreach communication at professional conferences, engagement events, and other pertinent venues.
A decade after the Martel et al. (2014) report of broad susceptibility of Western Palearctic salamanders to Bsal chytridiomycosis, many advances have been made to address the science and management of the Bsal threat throughout the world (review: Castro Monzon et al., 2022), and specifically in North America (e.g., Tables 1, 2). Despite this work, challenges remain in ensuring that native North American amphibian populations remain at low risk from Bsal. The following challenges are described further below: translating science into action; adopting best practices to reduce the introduction and spread of Bsal; preparation for a detection of Bsal in a free-ranging population; making precise forecasts in a no-analog system; conducting a continental-scale surveillance program; maintaining capacity within a volunteer-run organization; and identifying the most critical scientific uncertainties to improve management responses.
A first major challenge relates to the ability for large and disparate natural-resource management entities across a continent to translate science into management. An aim of the Bsal Task Force has been to provide information to address the Bsal threat and identify possible conservation and response actions. Management decisions are made by personnel with designated management and policy authorities using this information (e.g., in the US: Hopkins et al., 2018; Olson and Pilliod, 2024). Because of the multiple authorities responsible for amphibian population, habitat, and disease and invasive-species management, implementing management actions is complex, and existing processes may delay an ability to rapidly implement management or policy responses to new scientific findings (the ‘research-implementation gap’; Cook et al., 2013). For example, although Bsal-host susceptibility trials have been ongoing since 2014 and iteratively updating our understanding of both global and North American amphibian species that can be infected by Bsal (e.g., Castro Monzon et al., 2022; Gray et al., 2023b; Figure 3; Table 3; Supplementary Materials), there is no rapid mechanism to include iterative updates relevant to import policy restrictions for amphibian species that might transmit Bsal.
Implementation of biosecurity measures in field settings and in the pet trade are additional gaps for reducing the risk of Bsal introduction and spread in North America. Although field and captive biosecurity practices have been developed to reduce human-mediated Bsal transmission, adherence to recommendations can be improved. In the wild, biosecurity measures for wildlife health are not yet embedded as an everyday part of outdoor activities for researchers, managers, or recreationists—especially where it is highly relevant for sensitive species and habitats (Olson et al., 2021a). For Bsal in particular, there are ongoing efforts to echo the need for higher field-gear disinfectant concentrations for Bsal inactivation (Van Rooij et al., 2017; Bletz et al., 2023a; Bletz et al., 2023b). The Healthy Trade WG aims to increase the development and application of healthy-trade practices. Immediate improvements in actions to prevent potential spillover of pathogens from captive amphibians to the wild is needed, for example by following successful actions by many zoos and aquariums where water and substrate are disinfected before disposal as amphibian housing is cleaned (Pessier and Mendelson, 2017). Although state and federal law enforcement units routinely act to stop proponents of illegal animal trade, this has not been addressed relative to human-mediated Bsal transmission on potentially infected amphibians, specifically. Recognition of Bsal as an aquatic invasive species might be considered for jurisdictions in North America, to increase focal attention on relevant biosecurity measures to forestall injurious pathogen-species transmission (e.g., Table A3.2 in Seybold et al., 2021).
Another expected challenge is the ability to implement a response after an initial detection. Although preparation for a response to a Bsal detection can be guided by the Rapid Response Template (Bsal Task Force, 2022), conceptual advances garnered from scenario exercises, decision-support studies, applied research results, species-vulnerability assessments, lessons learned from the actions taken in Spain (Martel et al., 2020), the Bd-generated portfolio of possible actions (Garner et al., 2016; see also Mendelson et al., 2019), and expert opinion has unearthed substantial nuance with species, environmental, and jurisdictional constraints. Owing to these complex interlaced factors, an abbreviated version of the Rapid Response Template is being generated to highlight main steps along a time trajectory to better prepare managers ahead of a Bsal detection, and to help expedite initial phases of a reactive management approach. Priority aims need to be clear, such as assessment of whether containment if not eradication of the pathogen is feasible during initial detections in North America, and if rescue of at-risk or other species with Bsal-chytridiomycosis susceptibility is possible (Mendelson et al., 2019). Also missing from the current Rapid Response Template is guidance for management agencies interested in proactive activities, prior to first detection of Bsal. Proactive management is expected to be superior to reactive management, but the Rapid Response Template is only concerned with the key sequence of main actions after an initial detection; as contexts, uncertainties, ethics, and capacity to respond to a Bsal detection are weighed, the way forward may not be clear. The Decision Science working group has the expertise to help address these complex uncertainties.
Although forecasting and prediction for wildlife diseases is always challenging, the no-analog environmental and amphibian-diversity conditions in North America make predicting the outcome of a Bsal introduction particularly challenging. One approach to deal with this extreme uncertainty that the Bsal Task Force has advanced through the efforts of the Decision Science WG is to frame the disease-management problem as decisions faced by management and regulatory agencies (Bernard and Grant, 2019). Through this approach, researchers can identify the ‘critical’ uncertainties: those which, if resolved, would lead to the selection of different management actions (Canessa et al., 2020). This requires the identification and engagement of authorities who are responsible for management of the amphibian disease risk, as well as populations and habitats. The fragmented jurisdictions and authorities that are responsible for wildlife habitat and population management in North America are a principal challenge (e.g., in the US: Olson and Pilliod, 2022). For example, within the US, States have authority over wildlife management within their boundaries, and each of the 50 States has different Departments and/or personnel that oversee wildlife health and conservation activities, and States may have limited amphibian expertise if their internal priorities are more focused on higher-profile bird and mammal species. However, this is more complex, as within States: Tribes are independent nations and have authority over indigenous tribal lands; National Parks have authority on lands they manage under the US Department of Interior; and the US Department of Fish and Wildlife has authority over National Wildlife Refuges and US Endangered Species. More specifically, if potential loss of Endangered species is at stake from a novel Bsal outbreak in the US, the US Fish and Wildlife Service in partnership with State authorities, their established partners for animal rescue operations, such as regional-to-national zoos and aquariums, and amphibian- and Bsal-specific experts would be assembled for consultation and rapid action. Furthermore, management actions may involve permissions required by the US Environmental Protection Agency or State Department of Environmental Quality for chemical applications, or if on US Department of Agriculture lands or for use of federal funds, the permit process of the National Environmental Protection Act may be triggered for proposals of ground-disturbing activities, such as deployment of quarantine fences for exclusion of people or animals or drift fences for catching animals for biosecurity purposes. Who the members of an Incident Command Structure (Figure 2) for a first Bsal detection would be very context dependent, and likely include local, State, federal, and international consultants.
Finding ‘rules of thumb’ for proactive management, or for reacting to a Bsal detection, is complicated by these multiple dimensions of governance in decision making but may be developed into programmatic approaches by continuing to work with multiple management agencies at regional scales to solve their separate disease-management decision problems. This is an ongoing adaptive learning trajectory that will likely follow leadership of the Decision Science WG specialists. In addition to that WG being integral to consultations if Bsal were to be detected anywhere in North America, the larger Bsal Task Force partnerships across national jurisdictions of Canada, US, and Mexico also are likely to be available for consultation.
Ongoing Bsal surveillance in North America is another challenge. As for many conservation programs, the sustainability of surveillance programs can be an important barrier to implementation, with funding resources being a common limiting factor to program execution (Olson, 2007). Although each Bsal Task Force WG relies on funding to sustain efforts, surveillance efforts like SNAPS are more critically dependent upon funds for expenses such as analyzing field samples for Bsal at diagnostic laboratories. Despite the early successes and continued growth of SNAPS across North America, the current funding model is not sustainable. Under this, SNAPS can only support 25 institutions over the long term, and this is insufficient to sample all high-risk locations. A priority is to expand SNAPS into additional high-risk US states that are not currently sampled (e.g., Connecticut, Louisiana, New Jersey, and Oregon; Figure 4; Yap et al., 2017; Gray et al., 2023b). Expanding the currently limited surveillance in Mexico is also a priority. Additionally, the WG activities are currently performed by volunteers, which is a workforce-capacity challenge for maintaining and expanding the working-group activities. Preliminary assessment results demonstrate high satisfaction among both students and instructors, and many reported a greater appreciation for nature and wildlife after participating in SNAPS. Despite the success of SNAPS, there remains a need for a more permanent surveillance plan for Bsal that includes a sustainable funding model.
As above for SNAPS being particularly reliant on economic capital for program capability, the Bsal Task Force, overall, is especially reliant on human capital (Olson, 2007) which when overstretched, reduces the potential effectiveness of the conservation program, and perhaps is an element to consider for proactive conservation efforts, in particular. Fluctuating personnel time reduces capacity to proactively tackle gritty problems as a volunteer organization, such as time available for individual participants to become involved in multiple WGs or across geographies to network skillsets and common aims. In working with partner agencies, natural-resource managers additionally may be constrained in engaging in sustained long-term proactive amphibian conservation, as a potential disease threat such as Bsal comes secondary to current priorities that also may be understaffed and underfunded (Olson and Pilliod, 2022).
Lastly, gaps in amphibian-disease research have been recently identified (Grant et al., 2018; Bienentreu and Lesbarrères, 2020; Grant et al., 2023), many of which are relevant to Bsal. For example, Bienentreu and Lesbarrères (2020) recommended three areas likely to be of high relevance for Bsal management planning and implementation: 1) characterization of the host community assemblage (e.g., see community effects of Bd characterized by Harjoe et al., 2022); 2) life-stage and species-specific effects of a disease; and 3) assessment of environmental factors contributing to pathogen emergence and disease outcomes in hosts. Independently, Grant et al. (2023) developed 25 emerging priority research needs for amphibian conservation, with 13 having relevance to Bsal chytridiomycosis, although some stem from ongoing Bd-chytridiomycosis concerns. We have clustered these by priorities within five thematic areas of Bsal relevance (Table 4), some of which echo gaps and challenges mentioned above. Five priorities in Grant et al. (2023) address the thematic area of the need for more information on habitat or species management to increase disease resistance, three priorities address the theme of how to forestall threat effects for species of conservation concern, and two priorities address reducing transmission of disease-causing pathogens (i.e., elevated biosecurity and healthy-trade practices) and knowledge of system status. The one priority highlighted in Grant et al. (2023) that we have not considered here relates to correlated socioeconomic trends and the interaction with climate-change projections.
Table 4 Questions with relevance for Bsal chytridiomycosis knowledge gaps that were recently listed as top amphibian conservation research priorities in the Anthropocene by a world assembly of experts (from Grant et al., 2023).
The Bsal Task Force, along with other efforts worldwide, has advanced highly relevant knowledge of Bsal in North America, even though it is not known to occur on the continent at this time. For example, in 2014, when it was fully realized that some North American salamanders were likely susceptible to Bsal chytridiomycosis, we did not know if the pathogen was already on the continent. Now, owing to the thousands of specimen samples examined to date from across North America with no detections resulting from these broad surveillance activities (Waddle et al., 2020), we are more certain that Bsal is not already present (data accessible at amphibiandisease.org; Koo et al., 2021). Import restrictions have reduced the potential for human-mediated transmission through international trade markets, and new science findings of Bsal-susceptible species have been provided to inform adaptive processes of import restrictions. Research-and-development of Bsal diagnostics approaches has highlighted laboratory-result uncertainties, which have led to additional testing of analysis sensitivity per laboratory (round-robin laboratory test described above), and development of new more-reliable diagnostic approaches (Ossiboff et al., 2019). To ensure accurate Bsal-positive laboratory results are reported, two qPCR samples analyzed in different laboratories with laboratory-positive results are currently required to confirm Bsal detection in North America, with histopathology analyses being an additional requirement in Europe. Biosecurity efforts for field work have been disseminated (e.g., Gray et al., 2017; Julian et al., 2020; Olson et al., 2021a; Bletz et al., 2023a; Bletz et al., 2023b), and the Healthy Trade working group is making significant progress to bridge common Bsal biosecurity objectives between field and captive contexts. These efforts have somewhat reduced our uncertainty whether Bsal occurs in North America, and have supported the implementation of some mitigation strategies, though uncertainty remains and the threat of Bsal is not eliminated.
As such, the relevance of the Rapid Response Plan developed by the Bsal Task Force is heightened. If Bsal were to be suspected to be in North America, the Bsal Task Force has facilitated coordination of laboratories that can confirm the presence of Bsal, followed by the assembly of an Incident Command Response team (Figure 2) to guide an initial response. The activities of the Bsal Task Force and its partners have effectively fostered a collaborative among scientists, managers, and policy makers, sharing increased knowledge of the Bsal threat that can improve initial responses to discovery of Bsal in North America. Scenario exercises have helped map out likely early decisions and constraints in responses; principal concerns include the ability to contain or eradicate Bsal from an area and the potential non-target effects of any management actions. Learning and adaptive management are expected to improve future responses, and applied management approaches suggested for Bd (Garner et al., 2016; Mendelson et al., 2019) can be considered for Bsal.
The Bsal Task Force has made significant progress in planning for the expected invasion of Bsal in North America. Although this work is not yet completed after a decade of Bsal Task Force activities, the preparation for a Bsal invasion is far ahead of past experience with other amphibian-disease threats (Garner et al., 2016; Mendelson et al., 2019; Martel et al., 2020). The lessons learned we outline in this paper can serve as a model for other strategies to manage wildlife disease (e.g., marine wildlife disease: Glidden et al., 2022).
This work represents the composite accomplishments and challenges of the North American Bsal Task Force and its partners over about a 10-year time span, with potential application to other novel species threat-management contexts.
DO: Conceptualization, Investigation, Project administration, Visualization, Writing – original draft, Writing – review & editing. EG: Conceptualization, Project administration, Writing – review & editing, Visualization. MB: Conceptualization, Visualization, Writing – review & editing. JP: Conceptualization, Data curation, Writing – review & editing. DL: Visualization, Writing – review & editing, Data curation, Resources. JK: Conceptualization, Resources, Writing – review & editing. MA: Conceptualization, Writing – review & editing. MB: Writing – review & editing. MC: Writing – review & editing. MF: Writing – review & editing, Conceptualization, Project administration, Resources. MG: Conceptualization, Writing – review & editing, Resources. AH: Conceptualization, Writing – review & editing, Project administration. MK: Writing – review & editing, Conceptualization, Project administration. OM: Writing – review & editing. EAR: Writing – review & editing. LR: Conceptualization, Resources, Writing – review & editing. MS: Writing – review & editing. AS: Writing – review & editing, Conceptualization. LSh: Writing – review & editing. LSp: Writing – review & editing. JW: Writing – review & editing. AW: Writing – review & editing. BM: Conceptualization, Visualization, Writing – original draft, Project administration.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. MG was supported by NSF Division of Environmental Biology grant 1814520. This is U.S. Geological Survey publication: #8 of the Disease Decision Analysis and Research Group (DDAR), and contribution #905 of the Amphibian and Research and Monitoring Initiative (ARMI). A 2020 grant from the Wildlife Disease Association for publication of the North American Bsal Strategic Plan and related Bsal Task Force outreach materials aided conceptualization of this paper.
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Surveys described in the Healthy Trade section of this paper were organized and implemented by University partners and were not conducted on behalf of the U.S. Geological Survey. We thank Tiffany Yap and Dan Grear for use of their risk maps, Kathryn Ronnenberg and Jennifer Rowe for assistance with figure graphics, and Kathryn Ronnenberg for editorial assistance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer UOG declared a shared affiliation with the author EAR to the handling editor at the time of review.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/famrs.2024.1347541/full#supplementary-material
Adams M. J., Harris R. N., Grant E. H. C., Gray M. J., Hopkins M. C., Iverson S. A., et al. (2018). Prepublication communication of research results. EcoHealth 15 (3), 478–481. doi: 10.1007/s10393-018-1352-3
Alford R. A., Richards S. J. (1999). Global amphibian declines: a problem in applied ecology. Ann. Rev. Ecol. Syst. 30, 133–165. doi: 10.1146/annurev.ecolsys.30.1.133
AmphibianArk (2023) Amphibian Ark: keeping threatened amphibian species afloat. Available at: https://www.amphibianark.org/about-us/ (Accessed 29 November 2023).
Barnhart K., Bletz M. C., LaBumbard B., Tokash-Peters A., Gabor C. R., Woodhams D. C. (2020). Batrachochytrium salamandrivorans elicits acute stress response in spotted salamanders but not infection or mortality. Anim. Conserv. 23, 533–546. doi: 10.1111/acv.12565
Basanta M. D., Avila-Akerberg V., Byrne A. Q., Castellanos-Morales G., Martínez T. M. G., Maldonado-López Y., et al. (2022). The fungal pathogen Batrachochytrium salamandrivorans is not detected in wild and captive amphibians from Mexico. PeerJ 10, e14117. doi: 10.7717/peerj.14117
Basanta M. D., Rebollar E. A., Parra-Olea G. (2019). Potential risk of Batrachochytrium salamandrivorans in Mexico. PloS One 14 (2), e0211960. doi: 10.1371/journal.pone.0211960
Bernard R. F., Grant E. H. C. (2019). Identifying common decision problem elements for the management of emerging fungal diseases of wildlife. Soc Natur. Resour. 32, 1040–1055. doi: 10.1080/08941920.2019.1610820
Bernard R. F., Grant E. H. C. (2021). Rapid assessment indicates context-dependent mitigation for amphibian disease risk. Wildlife Soc Bull. 45, 290–299. doi: 10.1002/wsb.1198
Bienentreu J.-F., Lesbarrères D. (2020). Amphibian disease ecology: are we just scratching the surface? Herpetologica 76, 153–166. doi: 10.1655/0018-0831-76.2.153
Bishop C. A. (1992). “The effects of pesticides on amphibians and the implications for determining causes of declines in amphibian populations,” in Declines in Canadian Amphibian Populations: Designing a National Monitoring Strategy. Eds. Bishop C. A., Pettit K. E. (Occas. Pap. No. 76, Can. Wildlife Serv), 67–70.
Blaustein A. R., Kiesecker J. M., Chivers D. P., Anthony R. G. (1997). Ambient UV-B radiation causes deformities in amphibian embryos. Proc. Nat. Acad. Sci. U.S.A. 94, 137735–113737. doi: 10.1073/pnas.94.25.13735
Blaustein A. R., Wake D. B. (1990). Declining amphibian populations: a global phenomenon? Trends Ecol. Evol. 5 (7), 203–204. doi: 10.1016/0169-5347(90)90129-2
Bletz M. C., Grant E. H. C. (2022). Bsal basics: better together. In: Webinar (Partners in Amphibian and Reptile Conservation, Disease Task Team). Available at: https://www.youtube.com/watch?v=ahXtnr0KIzE (Accessed 29 November 2023).
Bletz M. C., Julian J. T., Kirchgessner M., Drasher J. M., Henry P. F. P., Jewell S. D., et al. (2023a). Disinfection protocols for herpetofaunal pathogens. Herpetol. Rev. 54 (2), 200–203.
Bletz M. C., Kelly M., Sabino-Pinto J., Bales E., Van Praet S., Bert W., et al. (2018). Disruption of skin microbiota contributes to salamander disease. P. R. Soc Lond. B Bio. 285, 20180758. doi: 10.1098/rspb.2018.0758
Bletz M. C., LaBumbard B. C., Le Sage E. H., Woodhams D. C. (2021). Extraction-free detection of amphibian pathogens from water baths. Dis. Aquat. Organ. 146, 81–89. doi: 10.3354/dao03621
Bletz M. C., Palmisano J., Julian J. T., Shender L., Olson D. H. (2023b). Amping up biosecurity for herps: simpler, stricter practices can help protect reptiles and amphibians from disease. Wildlife Prof. 17 (6), 46–49. Available at: https://parcplace.org/wp-content/uploads/2023/12/Amping-up-Biosecurity-for-Herps.pdf (Accessed 2 February 2024).
Blooi M., Pasmans F., Longcore J. E., Spitzen-van der Sluijs A., Vercammen F., Martel A. (2013). Duplex real-time PCR for rapid simultaneous detection of Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans in amphibian samples. J. Clin. Microbiol. 51, 4173–4177. doi: 10.1128/JCM.02313-13
Bosch J., Martinez-Solano I., Garcia-Paris M. (2001). Evidence of a chytrid fungus infection involved in the decline of the common midwife toad (Alytes obstetricans) in protected areas of central Spain. Biol. Conserv. 97, 331–337. doi: 10.1016/S0006-3207(00)00132-4
Brunner J. L. (2020). Pooled samples and eDNA-based detection can facilitate the “clean trade” of aquatic animals. Sci. Rep.-UK 10, 10280. doi: 10.1038/s41598-020-66280-7
Bsal Task Force (2022) A North American strategic plan to prevent and control invasions of the lethal salamander pathogen Batrachochytrium salamandrivorans. Available at: https://www.salamanderfungus.org/wp-content/uploads/2022/03/Bsal-Strategic-Plan_March-2022_FINAL.pdf (Accessed 29 November 2023).
Bsal Task Force (2023a) 2022 Annual Report (North American Bsal Task Force). Available at: https://www.salamanderfungus.org/wp-content/uploads/2023/06/Bsal-Annual-Report-2022-FINAL-2.0v.pdf (Accessed 29 November 2023).
Bsal Task Force (2023b) Diagnostics: QA/QC Program and Reports. Available at: https://www.diagnostics.salamanderfungus.org/qaqc (Accessed 29 November 2023).
Canessa S., Spitzen-van der Sluijs A., Stark T., Allen B. E., Bishop P. J., Bletz M., et al. (2020). Conservation decisions under pressure: lessons from an exercise in rapid response to wildlife disease. Conserv. Sci. Pract. 2 (1), e141. doi: 10.1111/csp2.141
Carey C., Bryant C. J. (1995). Possible interrelationships among environmental toxicants, amphibian development, and decline of amphibian populations. Environ. Health Perspect. 103 (Suppl. 4), 13–17. doi: 10.2307/3432406
Carter E. D., Bletz M. C., Le Sage M., LaBumbard B., Rollins-Smith L. A., Woodhams D. C., et al. (2021). Winter is coming–temperature affects immune defenses and susceptibility to Batrachochytrium salamandrivorans. PloS Pathog. 17 (2), e1009234. doi: 10.1371/journal.ppat.1009234
Carter E. D., Miller D. L., Peterson A. C., Sutton W. C. J., Cusaac P. W., Spatz J. A., et al. (2020). Conservation risk of Batrachochytrium salamandrivorans to endemic lungless salamanders. Conserv. Lett. 13, e12675. doi: 10.1111/conl.12675
Carvalho T., Becker C. G., Toledo L. F. (2017). Historical amphibian declines and extinctions in Brazil linked to chytridiomycosis. Proc. R. Acad. Soc B. 284, 20162254. doi: 10.1098/rspb.2016.2254
Castro Monzon F., Rödel M.-O., Ruland F., Parra-Olea G., Jeschke J. M. (2022). Batrachochytrium salamandrivorans amphibian host species and invasion range. EcoHealth 19, 475–486. doi: 10.1007/s10393-022-01620-9
Cavasos K., Adhikari R., Poudyal N. C., Brunner J. L., Warwick A., Gray M. J. (2023a). Understanding the demand for and value of pathogen-free amphibians to US pet owners. Conserv. Sci. Pract. 5 (9), e12995. doi: 10.1111/csp2.12995
Cavasos K., Adhikari R. K., Poudyal N. C., Warwick A. R., Gray M. J. (2023d). Natural area visitors’ potential role in preventing pathogen threats to amphibian biodiversity. Environ. Conserv. 50, 142–147. doi: 10.1017/S0376892923000048
Cavasos K., Poudyal N. C., Brunner J. L., Warwick A. R., Jones J., Moherman N., et al. (2023b). Attitudes and behavioral intentions of pet amphibian owners about biosecurity practices. EcoHealth 20, 194–207. doi: 10.1007/s10393-023-01645-8
Cavasos K., Poudyal N. C., Brunner J. L., Warwick A. R., Jones J., Moherman N., et al. (2023c). Exploring business stakeholder engagement in sustainable business practices: evidence from the US pet amphibian industry. Bus. Strateg. Environ. 32 (8), 5909–5921. doi: 10.1002/bse.3455
CBSA (Canada Border Services Agency) (2018) Environment and Climate Change Canada (ECCC)’s Import Restrictions on Salamanders. Customs Notice 17–17. Available at: https://www.cbsa-asfc.gc.ca/publications/cn-ad/cn17-17-eng.html (Accessed 29 November 2023).
Cheng T. L., Reichard J. D., Coleman T. H., Weller T. J., Thogmartin W. E., Reichert B. E., et al. (2021). The scope and severity of white-nose syndrome on hibernating bats in North America. Conserv. Biol. 35, 1586–1597. doi: 10.1111/cobi.13739
Commonwealth of Australia (2016) Threat abatement plan for infection of amphibians with chytrid fungus resulting in chytridiomycosis (Australian Government, Department of the Environment and Energy). Available at: https://www.dcceew.gov.au/sites/default/files/documents/tap-chytrid-fungus-2016.pdf (Accessed 29 November 2023).
Cook C. N., Mascia M. B., Schwartz M. W., Possingham H. P., Fuller R. A. (2013). Achieving conservation science that bridges the knowledge-action boundary. Conserv. Biol. 27, 669–678. doi: 10.1111/cobi.12050
Crawshaw L., Buchanan T., Shirose L., Palahnuk A., Cai H. Y., Bennett A. M., et al. (2022). Widespread occurrence of Batrachochytrium dendrobatidis in Ontario, Canada, and predicted habitat suitability for the emerging Batrachocytrium salamandrivorans. Ecol. Evol. 12 (4), e8798. doi: 10.1002/ece3.8798
Cunningham A. A., Beckmann K., Perkins M., Fitzpatrick L., Cromie R., Redbond J., et al. (2017). Emerging disease in UK amphibians. Vet. Rec. 176, 468. doi: 10.1136/vr.h2264
DiRenzo G. V., Longo A. V., Muletz-Wolz C. R., Pessier A. P., Goodheart J. A., Lips K. R. (2021). Plethodontid salamanders show variable disease dynamics in response to Batrachochytrium salamandrivorans chytridiomycosis. Biol. Invasions 23, 2797–2815. doi: 10.1007/s10530-021-02536-1
Fieschi-Méric L., Denoël M., Lesbarrères D. (2023). No detection of Ranaviruses nor Chytrids among salamanders and newts in Algonquin Provincial Park, Ontario, Canada. Herpetol. Rev. 54 (2), 204–207.
Finn C., Grattarola F., Pincheira-Donoso D. (2023). More losers than winners: investigating Anthropocene defaunation through the diversity of population trends. Biol. Rev. 98 (5), 1732–1748. doi: 10.1111/brv.12974
Fitzpatrick L. D., Pasmans F., Martel A., Cunningham. A. A. (2018). Epidemiological tracing of Batrachochytrium salamandrivorans identifies widespread infection and associated mortalities in private amphibian collections. Sci. Rep–UK 8, 13845. doi: 10.1038/s41598-018-31800-z
Friday B., Holzheuser C., Lips K. R., Longo A. V. (2020). Preparing for invasion: assessing risk of infection by chytrid fungi in southeastern plethodontid salamanders. J. Exper. Zool. 333, 829–840. doi: 10.1002/jez.2427
Gagliardo R., Crump P., Griffith E., Mendelson J., Ross H., Zippel K. (2009). The principles of rapid response for amphibian conservation using the programmes in Panama as an example. Internat. Zoo Yearbook 42, 125–135. doi: 10.1111/j.1748-1090.2008.00043.x
García-Rodríguez A., Basanta M. D., García-Castillo M. G., Zumbado-Ulate H., Neam K., Rovito S., et al. (2022). Anticipating the potential impacts of Batrachochytrium salamandrivorans on Neotropical salamander diversity. Biotropica 54, 157–169. doi: 10.1111/btp.13042
Garner T. W. J., Schmidt B. R., Martel A., Pasmans F., Muths E., Cunningham A. A., et al. (2016). Mitigating amphibian chytridiomycosis in nature. Phil. Trans. R. Soc Lond. B Bio. 371, 20160207. doi: 10.1098/rstb.2016.0207
Ghose S. L., Yap T. A., Byrne A. Q., Sulaeman H., Rosenblum E. B., Chan-Alvarado A., et al. (2023). Continent-wide recent emergence of a global pathogen in African amphibians. Front. Conserv. Sci. 4, 1069490. doi: 10.3389/fcosc.2023.1069490
Glidden C. K., Field L. C., Bachhuber S., Hennessey S. M., Cates R., Cohen L., et al. (2022). Strategies for managing marine disease. Ecol. Appl. 32, e2643. doi: 10.1002/eap.2643
Gluesenkamp A. G., Muscher-Hodges B. J., Lee M. M., Sandoval N. M., Fenolio D. B. (2018). Sampling for Batrachochytrium dendrobatidis and B. salamandrivorans in the Texas blind salamander (Eurycea rathbuni). Herpetol. Rev. 49, 44–46.
Govindarajulu P., Matthews E., Ovaska K. (2017). Swabbing for Batrachochytrium salamandrivorans on wild rough-skinned newts (Taricha granulosa) and pet-traded amphibians on Southern Vancouver Island. Herpetol. Rev. 48, 564–568.
Grant E. H. C., Adams M. J., Fisher R. N., Grear D. A., Halstead B. J., Hossack B. R., et al. (2018). Identifying management-relevant research priorities for responding to disease-associated amphibian declines. Global Ecol. Conserv. 16, e00441. doi: 10.1016/j.gecco.2018.e00441
Grant E. H. C., Amburgey S. M., Gratwicke B., Chaves V. A., Belasen A. M., Bickford D., et al. (2023). Priority research needs to inform amphibian conservation in the Anthropocene. Conserv. Sci. Pract. 2023, e12988. doi: 10.1111/csp2.12988
Grant E. H. C., Miller D. A., Schmidt B. R., Adams M. J., Amburgey S. M., Chambert T., et al. (2016a). Quantitative evidence for the effects of multiple drivers on continental-scale amphibian declines. Sci. Rep.–UK 6, 25625. doi: 10.1038/srep25625
Grant E. H. C., Muths E., Katz R. A., Canessa S., Adams M. J., Ballard J. R., et al. (2016b). Salamander chytrid fungus (Batrachochytrium salamandrivorans) in the United States – developing research, monitoring, and management strategies. USGS Open-File Report 2015-1233 (Reston, VA: U.S. Geological Survey).
Grant E. H. C., Muths E., Katz R. A., Canessa S., Adams M. J., Ballard J. R., et al. (2017). Using decision analysis to support proactive management of emerging infectious wildlife diseases. Front. Ecol. Environ. 15, 214–221. doi: 10.1002/fee.1481
Gray M. J., Allender M. C., Haman K. H., Harris R. N., Olson D. H. (2018). Facilitating early detection and rapid response: an alert system to combat emerging herpetofaunal diseases. Wildlife Prof. 12 (5), 33–35. Available at: https://parcplace.org/species/herpdiseasealert/ (Accessed 2 February 2024).
Gray M. J., Carter E. D., Piovia-Scott J., Cusaac J. P. W., Peterson A. C., Whetstone R. D., et al. (2023b). Broad host susceptibility of North American amphibian species to Batrachochytrium salamandrivorans suggests high invasion potential and biodiversity risk. Nat. Commun. 14, 3270. doi: 10.1038/s41467-023-38979-4
Gray M. J., Duffus A. L. J., Haman K. H., Harris R. N., Allender M. C., Thompson T. A., et al. (2017). Pathogen surveillance in herpetofaunal populations: guidance on study design, sample collection, biosecurity, and intervention strategies. Herpetol. Rev. 48, 334–351.
Gray M. J., Lewis J. P., Nanjappa P., Klocke B., Pasmans F., Martel A. N., et al. (2015). Batrachochytrium salamandrivorans: the North American response and a call for action. PloS Pathog. 11, e1005251. doi: 10.1371/journal.ppat.1005251
Gray M. J., Ossiboff R. J., Berger L., Bletz M. C., Carter E. D., DeMarchi J. A., et al. (2023a). One Health approach to globalizing, accelerating, and focusing amphibian and reptile disease research – Reflections and opinions from the First Global Amphibian and Reptile Disease Conference. Emerg. Infect. Dis. 29, e1–e7. doi: 10.3201/eid2910.221899
Grear D. A., Mosher B. A., Richgels K. L. D., Grant E. H. C. (2021). Evaluation of regulatory action and surveillance as preventive risk-mitigation to an emerging global amphibian pathogen Batrachochytrium salamandrivorans (Bsal). Biol. Conserv. 260, 109222. doi: 10.1016/j.biocon.2021.109222
Grisnik M., Gray M. J., Piovia-Scott J., Carter E. D., Sutton W. B. (2023). Incorporating caudate species susceptibilities and climate change into models of Batrachochytrium salamandrivorans risk in the United States of America. Biol. Conserv. 284, 110181. doi: 10.1016/j.biocon.2023.110181
Haman K. H., Henry P. F. P. (2022)Field biosecurity 101. In: Webinar (Partners in Amphibian and Reptile Conservation, Disease Task Team). Available at: https://www.youtube.com/watch?v=L3wfg9kXx_c&t=2308s (Accessed 29 November 2023).
Hardman R. H., Reinert L. K., Irwin K. J., Oziminski K., Rollins-Smith L., Miller D. L. (2023). Disease state associated with chronic toe lesions in hellbenders may alter anti-chytrid skin defenses. Sci. Rep.–UK 13, 1982. doi: 10.1038/s41598-023-28334-4
Harjoe C. C., Buck J. C., Rohr J. R., Olson D. H., Blaustein A. R. (2022). Pathogenic fungus causes density-and trait-mediated trophic cascades in an aquatic community. Ecosphere 13, e4043. doi: 10.1002/ecs2.4043
Hill A. J., Hardman R. H., Sutton W. B., Grisnik M. S., Gunderson J. H., Walker D. M. (2021). Absence of Batrachochytrium salamandrivorans in a global hotspot for salamander biodiversity. J. Wildlife Dis. 57 (3), 553–560. doi: 10.7589/JWD-D-20-00218
Hopkins M. C., Adams M. J., Super P. E., Olson D. H., Hickman C. R., English P., et al. (2018). Batrachochytrium salamandriovrans (Bsal) in Appalachia—Using scenario building to proactively prepare for a wildlife disease outbreak caused by an invasive amphibian chytrid fungus. USGS Open-File Report 2018-1150 (Reston, VA: US Geological Survey).
Isidoro-Ayza M., Grear D. A., Chambouvet A. (2019). Pathology and case definition of severe Perkinsea infection of frogs. Vet. Pathol. 56 (1), 133–142. doi: 10.1177/0300985818798132
Isidoro-Ayza M., Lorch J. M., Grear D. A., Winzeler M., Calhoun D. L., Barichivich W. J. (2017). Pathogenic lineage of Perkinsea associated with mass mortality of frogs across the United States. Sci. Rep.–UK 7 (1), 10288. doi: 10.1038/s41598-017-10456-1
Islam M. R., Gray M. J., Peace A. (2021). “Identifying the dominant transmission pathway in a multi-stage infection model of the emerging fungal pathogen Batrachochytrium salamandrivorans on the eastern newt,” in Infectious Diseases and Our Planet. Eds. Teboh-Ewungkem M. I., Akumah Ngwa G. (Springer), 193–216.
IUCN (2023) The IUCN Red List of Threatened Species. Version 2022-2 (International Union for the Conservation of Nature). Available at: https://www.iucnredlist.org (Accessed 29 November 2023).
Iwanowicz D. D., Schill W. B., Olson D. H., Adams M. J., Densmore C., Coprnman R. S., et al. (2017). Potential concerns with analytical methods used for the detection of Batrachochytrium salamandrivorans from archived DNA of amphibian swab samples, Oregon, USA. Herpetol. Rev. 48, 352–355.
Julian J. T., Henry P. F. P., Drasher J. M., Jewell S. D., Michell K., Oxenrider K. J., et al. (2020). Minimizing the spread of herpetofaunal pathogens in aquatic habitats by decontaminating construction equipment. Herpetol. Rev. 51, 472–483.
Klocke B., Becker M., Lewis J., Fleischer R. C., Muletz-Wolz C. R., Rockwood L., et al. (2017). Batrachochytrium salamandrivorans not detected in US survey of pet salamanders. Sci. Rep.–UK 7, 13132. doi: 10.1038/s41598-017-13500-2
Kolby J. E., Smith K. M., Berger L., Karesh W. B., Preston A., Pessier A. P., et al. (2014). First evidence of amphibian chytrid fungus Batrachochytrium dendrobatidis) and Ranavirus in Hong Kong amphibian trade. PloS One 9 (3), e90750. doi: 10.1371/journal.pone.0090750
Koo M. S., Vredenburg V. T., Deck J. B., Olson D. H., Ronnenberg K. L., Wake D. B. (2021). Tracking, synthesizing, and sharing global Batrachochytrium data at AmphibianDisease. org. Front. Vet. Sci. 8, 728232. doi: 10.3389/fvets.2021.728232
Kumar R., Malagon D. A., Carter E. D., Miller D. L., Bohanon M. L., Cusaac J. P. W., et al. (2020). Experimental methodologies can affect pathogenicity of Batrachochytrium salamandrivorans infections. PloS One 15, e0235370. doi: 10.1371/journal.pone.0235370
Leopold B. D., Cummins J. L., Kessler W. B. (2018). “Development of federal, state, and provincial laws. Chapter 6,” in North American Wildlife Policy and Law. Eds. Leopold B. D., Kessler W. B., Cummins J. L. (Missoula, MT: Boone and Crockett Club, Missoula), 53–63.
Le Sage E. H., LaBumbard B. C., Reinert L. K., Miller B. T., Richards-Zawacki C. L., Woodhams D. C., et al. (2021). Preparatory immunity: Seasonality of mucosal skin defences and Batrachochytrium infections in southern leopard frogs. J. Anim. Ecol. 90, 542–554. doi: 10.1111/1365-2656.13386
Lesbarrères D., Balseiro A., Brunner J., Chinchar V. G., Duffus A., Kerby J., et al. (2012). Ranavirus: past, present and future. Biol. Lett. 8, 481–483. doi: 10.1098/rsbl.2011.0951
Lips K. R., Diffendorfer J., Mendelson J. R. III, Sears M. W. (2008). Riding the wave: reconciling the roles of disease and climate change in amphibian declines. PloS Biol. 6, 441–454. doi: 10.1371/journal.pbio.0060072
Longcore J. E., Pessier A. P., Nichols D. K. (1999). Batrachochytrium dendrobatidis gen. et sp. Nov., a chytrid pathogenic to amphibians. Mycologia 91, 219–227. doi: 10.1080/00275514.1999.12061011
Longo A. V., Fleischer R. C., Lips K. R. (2019). Double trouble: co-infections of chytrid fungi will severely impact widely distributed newts. Biol. Invasions 21, 2233–2245. doi: 10.1007/s10530-019-01973-3
Lopes P. C., French S. S., Woodhams D. C., Binning S. A. (2021). Sickness behaviors across vertebrate taxa: proximate and ultimate mechanisms. J. Exper. Biol. 224, jeb225847. doi: 10.1242/jeb.225847
Luedtke J. A., Chanson J., Neam K., Hobin L., Maciel A. O., Catenazzi A., et al. (2023). Status of the world’s amphibians: emerging threats and ongoing declines. Nature 622, 308–314. doi: 10.1038/s41586-023-06578-4
Malagon D. A., Melara L. A., Proper O. F., Lenhart S., Carter E. D., Fordyce J. A., et al. (2020). Host density and habitat structure influence host contact rates and Batrachochytrium salamandrivorans transmission. Sci. Rep.-UK 10, 5584. doi: 10.1038/s41598-020-62351-x
Martel A., Blooi M., Adriaensen C., Van Rooij P., Beukema W., Fisher M. C., et al. (2014). Wildlife disease. Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science 346, 630–631. doi: 10.1126/science.1258268
Martel A., Spitzen-van der Sluijs A., Blooi M., Bert W., Ducatelle R., Fisher M. C., et al. (2013). Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proc. Nat. Acad. Sci. U.S.A. 110, 15325–15329. doi: 10.1073/pnas.1307356110
Martel A., Vila-Escale M., Fernandez-Giberteau D., Martinez-Silvestre A., Canessa S., Van Praet S., et al. (2020). Integral chain management of wildlife diseases. Conserv. Lett. 13, e12707. doi: 10.1111/conl.12707
Mendelson J. R., Whitfield S. M., Sredl M. J. (2019). A recovery engine strategy for amphibian conservation in the context of disease. Biol. Conserv. 236, 188–191. doi: 10.1016/j.biocon.2019.05.025
Mosher B. A., Bernard R. F., Lorch J. M., Richgels K. L. D., White C. L., Miller D. A., et al. (2020). Successful molecular detection studies require clear communication among diverse research partners. Front. Ecol. Environ. 18, 43–51. doi: 10.1002/fee.2141
Mosher B. A., Brand A. B., Wiewel A. N. M., Miller D. A. W., Gray M. J., Miller D. L., et al. (2019). Estimating occurrence, prevalence, and detection of amphibian pathogens: insights from occupancy models. J. Wildlife Dis. 55, 563–575. doi: 10.7589/2018-02-042
Mosher B. A., Huyvaert K. P., Bailey L. L. (2018). Beyond the swab: using ecosystem monitoring to understand the distribution of an amphibian pathogen. Oecologia 188, 319–330. doi: 10.1007/s00442-018-4167-6
Mosher B. A., Huyvaert K. P., Chestnut T., Kerby J. L., Madison J. D., Bailey L. L. (2017). Design- and model-based strategies for detecting and quantifying an amphibian pathogen in environmental samples. Ecol. Evol. 7, 10952–10962. doi: 10.1002/ece3.3616
Moubarak M., Fischhoff I. R., Han B. A., Castellanos A. A. (2022). A spatially explicit risk assessment of salamander populations to Batrachochytrium salamandrivorans in the United States. Divers. Distrib. 28, 2316–2329. doi: 10.1111/ddi.13627
Nguyen T. T., Nguyen T. V., Ziegler T., Pasmans F., Martel A. (2017). Trade in wild anurans vectors the urodelan pathogen Batrachochytrium salmandrivorans into Europe. Amphibia-Reptilia 38, 554–556. doi: 10.1163/15685381-00003125
O’Hanlon S. J., Rieux A., Farrer R. A., Rosa G. M., Waldman B., Bataille A., et al. (2018). Recent Asian origin of chytrid fungi causing global amphibian declines. Science 360 (6389), 621–627. doi: 10.1126/science.aar1965
Olson D. H. (2007). “Implementation considerations”, Chapter 11,” in Conservation of Rare or Little-Known Species: Biological, Social, and Economic Considerations. Eds. Raphael M. G., Molina R. (Washington, DC/Covelo, CA: Island Press), 303–333.
Olson D. H. (2015) Batrachochytrium salamandrivorans: North America Call to Action (International Reptile Conservation Federation magazine, Reptiles & Amphibians). Available at: https://parcplace.org/wp-content/uploads/2017/08/BsalBrief.pdf (Accessed 29 November 2023). Froglog 115, 11.; 2).
Olson D. H., Aanensen D. M., Ronnenberg K. L., Powell C. I., Walker S. F., Bielby J., et al. (2013). Mapping the global emergence of Batrachochytrium dendrobatidis, the amphibian chytrid fungus. PloS One 8 (2), e56802. doi: 10.1371/journal.pone.0056802
Olson D. H., Gray M., Pasmans F., Grayfer L., Wilber M. Q., Carter E. D., et al. (2022). The rising tide of herpetological disease science and management. Herpetol. Rev. 53, 416–417.
Olson D. H., Haman K. H., Gray M., Harris R., Thompson T., Iredale M., et al. (2021a). Enhanced between-site biosecurity to minimize herpetofaunal disease-causing pathogen transmission. Herpetol. Rev. 52 (1), 29–39.
Olson D. H., Pilliod D. S. (2022). Elevating the human dimension of amphibian and reptile conservation, a USA perspective. Conserv. Sci. Pract. 4 (6), e12685. doi: 10.1111/csp2.12685
Olson D. H., Pilliod D. (2024). “Amphibian and reptile conservation in the United States of America,” Chapter 22,” in Strategies for Conservation Success in Herpetology. Eds. Walls S. C., O’Donnell K. M. (University Heights, OH: Society for the Study of Amphibians and Reptiles).
Olson D. H., Ronnenberg K. L., Glidden C. K., Christiansen K. R., Blaustein A. R. (2021b). Global patterns of the fungal pathogen Batrachochytrium dendrobatidis support conservation urgency. Front.Vet. Sci. 8, 685877. doi: 10.3389/fvets.2021.685877
Ossiboff R. J., Towe A. E., Brown M. A., Longo A. V., Lips K. R., Miller D. L., et al. (2019). Differentiating Batrachochytrium dendrobatidis and B. salamandrivorans in amphibian chytridiomycosis using RNAScope® in situ hybridization. Front. Vet. Sci. 6, 304. doi: 10.3389/fvets.2019.00304
PARC (2023a) Save Our Amphibians & Reptiles: How YOU can Reduce the Spread of Disease. Available at: https://parcplace.org/species/herpetofaunal-disease-resources/ (Accessed 30 November 2023).
PARC (2023b) Amphibian Week 2023. Available at: https://parcplace.org/education/amphibianweek2023/ (Accessed 29 November 2023).
PARC [Partners in Amphibian and Reptile Conservation] (2022) Bsal Chytridiomycosis. Factsheet (Partners in Amphibian and Reptile Conservation, Wildlife Futures Program, US Fish and Wildlife Service National Wildlife Refuge System). Available at: https://parcplace.org/wp-content/uploads/2022/06/WFP-Fact-Sheet%E2%80%93Bsal_2.pdf (Accessed 29 November 2023).
Pereira K. E., Crother B. I., Sever D. M., Fontenot C. L. Jr., Pojman J. A. Sr., Wilburn D. B., et al. (2018). Skin glands of an aquatic salamander vary in size and distribution and release antimicrobial secretions effective against chytrid fungal pathogens. J. Exper. Biol. 221, jeb183707. doi: 10.1242/jeb.183707
Pereira K. E., Woodley S. K. (2021). Skin defenses of North American salamanders against a deadly salamander fungus. Anim. Conserv. 24 (4), 552–567. doi: 10.1111/acv.12666
Pessier A., Mendelson J. R. III (2017) A Manual for Control of Infectious Diseases in Amphibian Survival Assurance Colonies and Reintroduction Programs. Version 2.0 (Apple Valley, MN: IUCN/SSC Conservation Breeding Specialist Group). Available at: https://www.cbsg.org/disease-manual-amphibians-update-2017 (Accessed 29 November 2023).
Pet Product News (2015) PIJAC recommends immediate moratorium on firebelly and paddletail newt imports. Available at: https://www.petproductnews.com/news/pijac-recommends-immediate-moratorium-on-firebelly-and-paddletail-newt-imports/article_90cda85e-6ec9-5a40-a0ad-f3a9bd1b6274.html (Accessed 29 November 2023).
PIJAC (Pet Industry Joint Advisory Council) Canada (2016) Environment Canada helps pet stores take precautions against Bsal disease. Available at: https://pijacCanada.blogspot.com/2016/05/environment-Canada-helps-pet-stores.html (Accessed 29 November 2023).
Rawien J., Jairam-Doerga S. (2022). Predicted Batrachochytrium dendrobatidis infection sites in Guyana, Suriname, and French Guiana using the species distribution model maxent. PloS One 17, e0270134. doi: 10.1371/journal.pone.0270134
Richgels K. L., Russell R. E., Adams M. J., White C. L., Grant E. H. C. (2016). Spatial variation in risk and consequence of Batrachochytrium salamandrivorans introduction in the USA. R. Soc Open Sci. 3, 150616. doi: 10.1098/rsos.150616
Robinson K. A., Dunn M., Hussey S. P., Fritz-Laylin L. K. (2020a). Identification of antibiotics for use in selection of the chytrid fungi Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans. PloS One 15, e0240480. doi: 10.1371/journal.pone.0240480
Robinson K. A., Pereira K. E., Bletz M. C., Carter E. D., Gray M. J., Piovia-Scott J., et al. (2020b). Isolation and maintenance of Batrachochytrium salamandrivorans cultures. Dis. Aquat. Org. 140, 1–11. doi: 10.3354/dao03488
Rollins-Smith L. A. (2020). Global amphibian declines, disease, and the ongoing battle between Batrachochytrium fungi and the immune system. Herpetologica 76, 178–188. doi: 10.1655/0018-0831-76.2.178
Rollins-Smith L. A., Le Sage E. H. (2021). Batrachochytrium fungi: stealth invaders in amphibian skin. Curr. Opin. Microbiol. 61, 124–132. doi: 10.1016/j.mib.2021.04.002
Rollins-Smith L. A., Reinert L. K., Le Sage M., Linney K. N., Gillard B. M., Umile T. P., et al. (2022). Lymphocyte inhibition by the salamander-killing chytrid fungus, Batrachochytrium salamandrivorans. Infect. Immun. 90 (3), e00020–e00022. doi: 10.1128/iai.00020-22
Ruggs E., Hebl M. (2012)Diversity, inclusion, and cultural awareness for classroom and outreach education. In: Apply Research to Practice (ARP) Resources (Literature Overview). Available at: https://teach.ufl.edu/wp-content/uploads/2016/07/ARP_DiversityInclusionCulturalAwareness_Overview.pdf (Accessed 29 November 2023).
Russell R. E., Katz R. A., Richgels K. L. D., Walsh D. P., Grant E. H. C. (2017). A framework for modeling emerging diseases to inform management. Emerg. Infect. Dis. 23, 1–6. doi: 10.3201/eid2301.161452
Sadinski W. J., Dunson W. A. (1992). A multilevel study of effects of low pH on amphibians of temporary ponds. J. Herpetol. 26, 413–422. doi: 10.2307/1565117
Schloegel L. M., Picco A. M., Kilpatrick A. M., Davies A. J., Hyatt A. D., Daszak P. (2009). Magnitude of the US trade in amphibians and presence of Batrachochytrium dendrobatidis and ranavirus infection in imported North American bullfrogs (Rana catesbeiana). Biol. Conserv. 142, 1420–1426. doi: 10.1016/j.biocon.2009.02.007
Seybold S. J., Frankel S. J., Olson D. H., Flitcroft R., Kerns B. K., Lee J. C., et al. (2021)“Northwest region,” in “Appendix: regional summaries”. In: Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector (Cham, Switzerland: Springer Verlag) (Accessed 29 November 2023).
Sheley W. C., Gray M. J., Wilber J. G., Cray C., Carter E. D., Miller D. L. (2023). Electrolyte imbalances and dehydration play a key role in Batrachochytrium salamandrivorans chytridiomycosis. Front. Vet. Sci. 9, 1055153. doi: 10.3389/fvets.2022.1055153
Skerratt L. F., Berger L., Hines H. B., McDonald K. R., Mendez D., Speare R. (2008). Survey protocol for detecting chytridiomycosis in All Australian frog populations. Dis. Aquat. Org. 80, 85–94. doi: 10.3354/dao01923
Skerratt L. F., Berger L., Speare R., Cashins S., McDonald K. R., Phillott A. D., et al. (2007). Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. EcoHealth 4, 125–134. doi: 10.1007/s10393-007-0093-5
Smith H. K., Pasmans F., Dhaenens M., Deforce D., Bonte D., Verheyen K., et al. (2018). Skin mucosome activity as an indicator of Batrachochytrium salamandrivorans susceptibility in salamanders. PloS One 13 (7), e0199295. doi: 10.1371/journal.pone.0199295
Standish I., Leis E., Schmitz N., Credico J., Erickson S., Bailey J., et al. (2018). Optimizing, validating, and field testing a multiplex qPCR for the detection of amphibian pathogens. Dis. Aquat. Organ. 129, 1–13. doi: 10.3354/dao03230
Stegen G., Pasmans F., Schmidt B. R., Rouffaer L. O., Van Praet S., Schaub M., et al. (2017). Drivers of salamander extirpation mediated by Batrachochytrium salamandrivorans. Nature 544, 353–356. doi: 10.1038/nature22059
Stuart S. N., Chanson J. S., Cox N. A., Young B. E., Rodrigues A. S., Fischman D. L., et al. (2004). Status and trends of amphibian declines and extinctions worldwide. Science 306, 1783–1786. doi: 10.1126/science.1103538
Thomas V., Blooi M., Rooij P. V., Praet S. V., Verbrugghe E., Grasselli E., et al. (2018). Recommendations on diagnostic tools for Batrachochytrium salamandrivorans. Transbound Emerg. Dis. 65, e478–e488. doi: 10.1111/tbed.12787
Tompros A., Dean A. D., Fenton A., Wilber M. Q., Carter E. D., Gray M. J. (2022a). Frequency-dependent transmission of Batrachochytrium salamandrivorans in eastern newts. Transbound Emerg. Dis. 69, 731–741. doi: 10.1111/tbed.14043
Tompros A., Wilber M. Q., Fenton A., Carter E. D., Gray M. J. (2022b). Efficacy of plant-derived fungicides at inhibiting Batrachochytrium salamandrivorans growth. J. Fungi 8, 1025. doi: 10.3390/jof8101025
Towe A. E., Gray M. J., Carter E. D., Wilber M. Q., Ossiboff R. J., Ash K., et al. (2021). Batrachochytrium salamandrivorans can devour more than salamanders. J. Wildlife Dis. 57, 942–948. doi: 10.7589/JWD-D-20-00214
U.S. Department of Agriculture [USDA] (2023) By the Secretary of Agriculture of the United States of America: A Proclamation. National Amphibian Week, May 7–13, 2023. Available at: https://parcplace.org/wp-content/uploads/2023/05/Amphibian-Week-National-Proclamation-by-USDA-2023.pdf (Accessed 29 November 2023).
U.S. Department of the Interior [USDI] (2021) By the Secretary of the Interior of the UNITED STATES of America: A Proclamation. National Amphibian Week, May 2–8, 2021. Available at: https://d9-wret.s3.us-west-2.amazonaws.com/assets/palladium/production/s3fs-public/atoms/files/2021Proclamation.pdf (Accessed 29 November 2023).
U.S. Fish & Wildlife Service (2007) Chiricahua Leopard Frog (Rana chiricahuensis) Final Recovery Plan (New Mexico, USA: U.S. Fish a& Wildlife Service, Southwest Region, Albuquerque). Available at: https://www.amphibians.org/wp-content/uploads/2019/04/Chiricahua-leopard-frog-Recovery-Plan.pdf (Accessed 29 November 2023).
U.S. Fish & Wildlife Service (2016) Injurious wildlife species; Listing salamanders due to risk of salamander Chytrid Fungus. A rule by the Fish and Wildlife Service on 01/13/2016. Available at: https://www.federalregister.gov/documents/2016/01/13/2016-00452/injurious-wildlife-species-listing-salamanders-due-to-risk-of-salamander-chytrid-fungus (Accessed 29 November 2023).
Van Rooij P., Pasmans F., Coen Y., Martel A. (2017). Efficacy of chemical disinfectants for the containment of the salamander chytrid fungus Batrachochytrium salamandrivorans. PloS One 12, e0186269. doi: 10.1371/journal.pone.0186269
Waddle J. H., Grear D. A., Mosher B. A., Grant E. H. C., Adams M. J., Backlin A. R., et al. (2020). Batrachochytrium salamandrivorans (Bsal) not detected in an intensive survey of wild North American amphibians. Sci. Rep.–UK 10, 1–7. doi: 10.1038/s41598-020-69486-x
Wake D. B., Vredenburg V. T. (2008). Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Nat. Acad. Sci. U.S.A. 105, 11466–11473. doi: 10.1073/pnas.0801921105
White C. L., Forzán M. J., Pessier A. P., Allender M. C., Ballard J. R., Catenazzi A., et al. (2016). Amphibian: a case definition and diagnostic criteria for Batrachochytrium salamandrivorans chytridiomycosis. Herpetol. Rev. 47, 207–209.
Wilber M. Q., Carter E. D., Gray M. J., Briggs C. J. (2021). Putative resistance and tolerance mechanisms have little impact on disease progression for an emerging salamander pathogen. Funct. Ecol. 35, 847–859. doi: 10.1111/1365-2435.13754
WOAH (World Organisation for Animal Health) (2023) Chytridiomycosis (Batrachochytrium salamandrivorans). Available at: https://www.woah.org/en/disease/chytridiomycosis-batrachochytrium-salmandrivorans/ (Accessed 29 November 2023).
Woodhams D. C., Barnhart K. L., Bletz M. C., Campos A. J., Ganem S. J., Hertz A., et al. (2018a)Batrachochytrium: biology and management of amphibian chytridiomycosis. In: Encyclopedia of Life Sciences (Wiley) (Accessed 29 November 2023).
Woodhams D. C., LaBumbard B. C., Barnhart K. L., Becker M. H., Bletz M. C., Escobar L. A., et al. (2018b). Prodigiosin, violacein, and volatile organic compounds produced by widespread cutaneous bacteria of amphibians can inhibit two Batrachochytrium fungal pathogens. Microbial Ecol. 75, 1049–1062. doi: 10.1007/s00248-017-1095-7
Wright A. D., Bernard R. F., Mosher B. A., O’Donnell K. M., Braunagel T., DiRenzo G. V., et al. (2020). Moving from decision to action in conservation science. Biol. Conserv. 249, 108698. doi: 10.1016/j.biocon.2020.108698
Yap T. A., Koo M. S., Ambrose R. F., Wake D. B., Vredenburg V. T. (2015). Averting a North American biodiversity crisis. Science 349, 481–482.
Yap T. A., Nguyen N. T., Serr M., Shepack A., Vredenburg V. T. (2017). Batrachochytrium salamandrivorans and the risk of a second amphibian pandemic. EcoHealth 14, 851–864. doi: 10.1007/s10393-017-1278-1
Yuan Z., Martel A., Wu J., Van Praet S., Canessa S., Pasmans F. (2018). Widespread occurrence of an emerging fungal pathogen in heavily traded Chinese urodelan species. Conserv. Lett. 11, e12436. doi: 10.1111/conl.12436
Keywords: chytridiomycosis, disease, threat, mitigation, Bsal, salamander
Citation: Olson DH, Grant EHC, Bletz M, Piovia-Scott J, Lesbarrères D, Kerby JL, Adams MJ, Breitman MF, Christman MR, Forzán MJ, Gray MJ, Hill AJ, Koo MS, Milenkaya O, Rebollar EA, Rollins-Smith LA, Serr M, Shepack A, Shirose L, Sprague L, Walke JB, Warwick AR and Mosher BA (2024) Preparing for a Bsal invasion into North America has improved multi-sector readiness. Front. Amphib. Reptile Sci. 2:1347541. doi: 10.3389/famrs.2024.1347541
Received: 04 December 2023; Accepted: 25 January 2024;
Published: 06 March 2024.
Edited by:
Faraham Ahmadzadeh, Shahid Beheshti University, IranReviewed by:
Kerim Çiçek, Ege University, TürkiyeCopyright © 2024 Olson, Grant, Bletz, Piovia-Scott, Lesbarrères, Kerby, Adams, Breitman, Christman, Forzán, Gray, Hill, Koo, Milenkaya, Rebollar, Rollins-Smith, Serr, Shepack, Shirose, Sprague, Walke, Warwick and Mosher. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Deanna H. Olson, ZGVhbm5hLm9sc29uQHVzZGEuZ292
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.