 Supti Saha Mou1
Supti Saha Mou1 Miranda J. Haus
Miranda J. Haus Zachary D. Hayden
Zachary D. Hayden Eric L. Patterson
Eric L. Patterson Debalina Saha
Debalina Saha
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Agron., 04 March 2025
Sec. Weed Management
Volume 7 - 2025 | https://doi.org/10.3389/fagro.2025.1556418
Climate change, driven by rising atmospheric carbon dioxide (CO2), increasing temperatures, and shifting precipitation patterns, is profoundly impacting agricultural systems worldwide. These environmental changes significantly affect weed growth, distribution, and management, posing challenges across agronomic, horticultural, and ornamental crops. This review explores the impacts of climate change on weeds, focusing on the differential responses of C3 and C4 weed species to elevated CO2, higher temperatures, and drought stress. It also examines how these climatic factors influence weed management practices, particularly herbicide efficacy. While much research has focused on agronomic crops, ornamental crop productions remain underexplored, despite their unique challenges. Ornamental production systems often involve diverse plant species grown in confined spaces, making weed management more complex and sensitive to herbicide residues. These challenges are compounded by the adaptability and invasiveness of weeds under changing climatic conditions. The review highlights critical knowledge gaps, particularly the limited understanding of how climatic factors impact weed physiology and herbicide performance in ornamental settings. Addressing these gaps is essential to develop climate-resilient strategies for sustainable weed management across diverse agricultural systems.
The ornamental crop industry in the United States, which includes Christmas tree production, nurseries, greenhouse cultivation, and landscape management, is a billion-dollars industry. In 2018, a survey conducted by the Green Industry Research Consortium among 1,727 participants reported total annual sales of USD 2.392 billion (Khachatryan et al., 2020). Over the past 15 years, the container production segment within the nursery industry has been growing rapidly and is expected to continue expanding. However, a major issue in container-grown nurseries is weed infestations. Since resources like water, nutrients, and soil air are limited within containers, weeds compete with the main plants for these essentials (Case et al., 2005). Nursery growers estimate that manually removing weeds from containers costs between USD 500 to USD 4000 per acre (USD 1235 to USD 9880 per hectare). The economic damage caused by weed infestations can be as high as USD 7000 per acre (USD 17290 per hectare) (Mathers, 2003). So, efficient weed control is critical, as the impact of weeds is often underestimated (Case et al., 2005). If weed management practices are improved to reduce costs, it could significantly benefit the industry (Adams et al., 1998).
The increase in carbon dioxide (CO2) and other greenhouse gases, primarily due to human activities like land use changes and fossil fuel burning, is a major cause of global warming. Higher CO2 levels generally boost the growth and yield of most plants by enhancing photosynthesis and water use efficiency (Rogers and Dahlman, 1993; Amthor, 1995). However, these changes can also affect the growth rates of both crops and weeds, potentially leading to shifts in vegetation patterns and altering regional ecosystems (Pagare et al., 2017). Climate change impacts species’ geographic ranges, life cycles, population dynamics, and interactions between crops and weeds. Plants with C3 photosynthetic pathways, which include most food crops, are likely to benefit more from CO2 increases than C4 plants. However, rising temperatures could give C4 plants, many of which are common weeds, a competitive advantage over C3 plants. This shift may complicate weed management, as higher CO2 levels can also increase the growth of some weed species and make them harder to control. Additionally, warmer temperatures may speed up the life cycles of certain plants, causing weeds to mature and decay more quickly (Singh et al., 2011).
Given these challenges, understanding how weeds respond to elevated CO2 levels, rising temperatures, and drought is essential for managing their impact on agricultural systems. Climate change influences weed physiology, growth, and competitive interactions, while also altering the efficacy of herbicides and other management practices. This review synthesizes current research on the impacts of climate change on weeds across agronomic, horticultural, and ornamental cropping systems. Particular attention is given to the physiological and morphological responses of C3 and C4 weed species to changing climatic factors, as well as the effects of these changes on herbicide performance and broader weed management strategies. By consolidating findings across multiple disciplines, this review aims to provide a comprehensive understanding of how climate change shapes weed dynamics and management approaches.
Agriculture is considered as the most vulnerable sector to climate change because of to its huge size and sensitivity to weather parameters which can cause significant economic impacts (Mendelsohn, 2009). The impact of increasing temperatures, precipitation variation, and CO2 fertilization differs according to the crop, location, and magnitude of change in the parameters (Adams et al., 1998). Higher temperatures are likely to shorten the time for crop growth, reducing their overall yield (Mahato, 2014). If both temperate and tropical regions warm by 2°C, it’s expected that wheat, rice, and maize production will decrease (Challinor et al., 2014). Climate change projections suggest more frequent droughts in many parts of the world and the area which would be affected by droughts expected to increase from 15.4% to 44.0% by 2100. Major crop yields in drought-prone areas could drop by more than 50% by 2050 and nearly 90% by 2100 (Li et al., 2009). As atmospheric CO2 levels continue to rise, it’s expected that crop yields will increase, although the extent of this increase will vary depending on the type of crop. Particularly, C3 crops are likely to produce greater yields, and both C3 and C4 crops may require less water under less stressful conditions (DaMatta et al., 2010). The levels of nutrients like nitrogen, iron, zinc, and sulfur, primarily found in proteins, decrease in non-leguminous C3 crops with higher CO2 levels (Uddling et al., 2018). With increased CO2, rice crops exhibit enhanced vegetative and reproductive growth and higher seed yields at air temperature of 29°C; however, seed set decreases with rising temperatures (Madan et al., 2012). Elevated CO2 levels lead to decreases in zinc and iron content and the protein concentrations in C3 crops and legumes while C4 plants remain unaffected by increased CO2 levels (Myers et al., 2014). Furthermore, higher CO2 levels and rising temperatures are escalating the threat of potato late blight, rice blast, and sheath blight, which could severely threaten global food security (Gautam et al., 2013).
Changes in climate are likely to affect the development and survival of pathogens in humid and warmer regions (Rosenzweig et al., 2001; Elad and Pertot, 2014). Changes in an area’s climate or weather patterns are predicted to increase a crop’s susceptibility to various pests, diseases, and weeds (Rosenzweig et al., 2001). However, there are projections that a one-degree increase in temperature could lead to a 10–25% increase in losses due to insect pest infestations (Shrestha, 2019). Statistical evidence shows that increased rainfall and temperatures have raised pesticide costs for crops such as corn, potatoes, and soybeans, while reducing costs for wheat in the USA (Chen and McCarl, 2001).
Climate change also affect the microbial populations in soil and their enzymatic activities. Experiments using temperature gradient tunnels, with temperatures 4–5°C higher than field conditions, revealed significantly higher microbial populations. Nitrogen-fixing bacteria, P-solubilizers, fungi, and enzymatic activities were notably elevated across a range of temperatures, with optimal conditions yielding the highest parameters (Kaur et al., 2014). Additionally, warmer, and more humid regions are conducive to increased prevalence of insect pests and diseases (Rosenzweig and Liverman, 1992). On the other hand, the growth of endophytic fungi and plant growth-promoting bacteria can have positive, negative, or neutral effects, depending on the temperature range (Compant et al., 2010).
Climate change significantly impacts horticultural crop production, particularly affecting the yields of fruits and vegetables. The variability in rainfall and rising temperatures, primary indicators of climate change, are major challenges that disrupt the growth, development, and productivity of horticultural crops (Datta, 2013). Unpredictable high temperatures and altered rainfall patterns lead to heat stress and water scarcity, resulting in shorter growing periods and decreased production of fruits and vegetables. These changes not only reduce water availability essential for crop growth but also negatively impact the photosynthesis process, crucial for crop development (Malhotra and Srivastva, 2014; Malhotra, 2015). Specifically, fruit crops such as bananas may suffer from heat stress, soil moisture stress, and flooding due to climate change (Kumar and Kumar, 2007). Similarly, apples, apricots, cherries, and litchis face increased risks of sunburn and fruit cracking due to high temperatures during the maturation phase. The changes in climatic conditions also affect the chilling requirements for pome and stone fruits, altering the timing of dormancy breaking (Malhotra, 2017).
In the case of vegetables, significant losses are observed in crops like tomatoes, where high temperatures lead to reduced fruit set, smaller fruits, and lower quality. This is due to a range of heat-induced reproductive failures, including bud drop, abnormal flower development, poor pollen production, and ovule abortion (Hazra et al., 2007). Peppers also experience inhibited fruit set under high post-pollination temperatures, highlighting the sensitivity of fertilization processes to thermal stress. Additionally, vegetables like tomato, watermelon, potato, squash, soybeans, and carrots are highly susceptible to damage from air pollution, with yields potentially reduced by 5-15% when ozone levels exceed 50 ppb (Narayan, 2009).
The U.S. Green Industry, encompassing horticultural nurseries, landscape designers, Christmas tree producers and related retail and distribution sectors, continues to grow robustly despite economic fluctuations. In 2020, this sector saw a 9% increase in the wholesale value of floriculture crops from the previous year, with major contributions from states like Florida, California, and Michigan, accounting for a significant portion of the national total of $4.80 billion (Usda and National Agricultural Statistics, S, 2021). In Michigan, the ornamental and landscape industry have a substantial economic footprint, generating a total impact of $1.26 billion, including related sectors (Michigan Department of and Rural, 2019). The industry directly employs over 16,000 people, highlighting its importance to the state’s economy (Knudson, 2018). Climate change can create unprecedented challenges in this industry, which vary across different types of ornamental crops.
In the United States, the Christmas tree industry is a profitable sector, encompassing approximately 15,000 farms across nearly 350,000 acres, generating around $250 million in annual sales and providing employment for over 100,000 individuals (National Christmas Tree, A, 2021). However, this area is facing significant challenges due to climate change and weed management issues. Rising fall temperatures, a consequence of climate change, are prompting farmers to harvest trees earlier than usual, often shifting from late November to mid-October. This adjustment is primarily to meet market demands but results in the trees experiencing insufficient cold weather necessary for optimal needle retention. Consequently, this warmer weather and altered harvest schedules are compromising the quality of Christmas trees, as evidenced by studies highlighting the importance of proper cold acclimation for needle retention (Mitcham-Butler et al., 1988; MacDonald, 2010; Thiagarajan, 2012; MacDonald et al., 2014). Additionally, weed infestation poses a severe problem throughout all stages of Christmas tree production. Often grown in sandy soils where water resources are scarce, these trees face increased competition from weeds for these limited resources, elevating their risk of drought stress. Weeds that overshadow young saplings can block essential sunlight, hindering photosynthesis, which is crucial for the development of leaves and overall tree growth (Roberts and Long, 1992). Effective weed management, particularly during the critical first three years of a tree’s life, is essential for the establishment of robust root systems, which are vital for enduring later drought conditions (Zandstra and O’Donnell, 2018). Unmanaged weeds not only threaten the physical growth of Christmas trees but also provide shelter for harmful wildlife like field mice, rabbits, and deer, further jeopardizing the trees. Furthermore, weeds compete with Christmas trees for nutrients, adversely affecting the aesthetic qualities of the trees, such as needle size and color quality (Oregon State University, 1981).
Climate change is threatening the growth of the ornamental crop industry and is affecting other ornamental crops in various ways. Temperature increases lead to physiological changes in plants, affecting growth, development, and productivity. An increase in temperature can lead to a variety of physiological changes in plants, including increased respiration rates, varied rates of photosynthesis, and the redistribution of photosynthesis to economically important parts of the plant. These changes can also alter plant phenology, reduce the overall crop growing period, and accelerate the days to flowering and fruiting, as well as speed up fruit maturity, ripening, and senescence. Additionally, temperature increases might not be evenly distributed between day and night or across different seasons (Srinivasa Rao et al., 2010). The blooming of various ornamental plants like Rhododendron, Orchid, Tulipa, Alstroemeria, Magnolia, Saussurea, Impatiens, and Narcissus may be adversely affected by decreasing chilling periods. Some species may become endangered, while others may experience reduced or less vibrant flowering. Flowering plant species, particularly those grown in open fields, will require high humidity and ample water to survive under challenging conditions. For instance, Chrysanthemum, a short-day plant, cannot bloom year-round in open fields. Similarly, Jasmine ceases to flower at temperatures below 19°C, resulting in smaller flowers. In tropical orchids, temperatures below 15°C prevent flowers from fully opening, while temperatures exceeding 35°C can cause buds to drop and spikes to become unmarketable (Hirpo and Gebeyehu, 2019).
Amid these climatic challenges, the issue of weed infestation in nursery container production becomes even more critical. Improvements in weed management could significantly boost profitability for nursery operators, positively affecting the extensive nursery market in Michigan. Globally, the ornamental and cut flower market sees the U.S. holding a 12.5% share, trailing behind China and Europe (Darras, 2020). Unlike field production where direct soil sowing allows for effective post-germination weed control via herbicides or mechanical means, container production needs preventive measures against weeds due to the impracticality of direct herbicide application on containerized plants (Altland, 2003).
Weeds pose a significant threat to ornamental crop production by depriving ornamental plants from essential resources, ultimately reducing plant growth and quality. Unlike agronomic crops, ornamental plants are not grown for a single season and harvested, but rather remain in production for several years, making traditional weed control strategies less effective (Beeson, 1991). In many agronomic crops, guidelines help growers deciding the best time to control weeds by comparing the cost of weed management to the expected increase in yield (Smith, 1968; Cousens, 1987; O’Donovan, 2005; Knezevic, 2014; Swanton et al., 2020). However, this approach doesn’t work well in ornamental plant production because of the wide variety of plant species and the different ways they are grown. Since ornamental plants are valued for their appearance rather than yield, even a small amount of weed competition can reduce their marketability, making weed management more complex (Beeson, 1991). In nursery environments, weed infestations lead to biomass reductions and production delays, regardless of species or container size. As weed density increases, competition effects initially develop slowly but intensify over time, severely impacting plant growth. For instance, shoot dry weight reductions of 18%–53% in Japanese holly (Ilex crenata) and 12%–56% in ligustrum (Ligustrum vulgare) have been observed when grown under varying levels of weed competition in different container sizes (Khamare et al., 2023). Presence of a single redroot pigweed (Amaranthus retroflexus) or large crabgrass (Digitaria sanguinalis) plant in a 2.4 L container reduced Japanese holly dry weight by up to 60% (Fretz, 1972). Higher weed densities, such as five weeds per container of barnyardgrass (Echinochloa crus-galli), Digitaria sanguinalis, or Chinese foxtail (Setaria faberi), competing for 83 days, resulted in a 72% reduction in Bailey’s redosier dogwood (Cornus sericea) shoot dry weight (Walker and Williams, 1988). Other common weeds, including coffeeweed (Senna obtusifolia), cocklebur (Xanthium strumarium), and eclipta (Eclipta prostrata), have been documented to negatively affect numerous container-grown ornamentals (Walker and Williams, 1988; Norcini and Stamps, 1994; Fain et al., 2003; Khamare et al., 2020). Additionally, weed competition leads to reduced leaf size, lower flower production, and poor plant vigor, with bush cinquefoil (Potentilla fruticosa ‘Gold Drop’) experiencing a 99% decrease in flower production due to Digitaria sanguinalis (Walker and Williams, 1989).
In Christmas tree production, weeds can hinder the growth of Christmas trees and reduce tree survival rates. During the critical establishment phase, particularly the first three years after transplanting, weed competition for moisture can result in drought stress, stunted growth, and even tree mortality (Cui and Smith, 1991; Knowe and Stein, 1995; NeSmith and Lindstrom, 1996; Schneider et al., 1998; Harper et al., 2005; Zandstra and O’Donnell, 2018). This issue is more severe in sandy soils, where limited water availability is further depleted by weed competition, making trees highly susceptible to drought. Additionally, weed shading reduces photosynthetic activity, limiting leaf area development and overall tree vigor (Roberts and Long, 1992). As trees mature, weeds interfere with cultural practices such as pruning and pesticide applications and can shade lower branches, affecting uniformity and quality (Brown et al., 1991; Zandstra and O’Donnell, 2018). Several broadleaf and grassy weeds, including field bindweed (Convolvulus arvensis), horseweed (Erigeron canadensis), common ragweed (Ambrosia artemisiifolia), giant foxtail (Setaria faberi), and large crabgrass (Digitaria sanguinalis), invade tree branches, making removal difficult. Vining weeds, such as Virginia creeper (Parthenocissus quinquefolia), poison ivy (Toxicodendron radicans), and wild grape (Vitis spp.), entangle trees, preventing effective management without risking herbicide damage (Zandstra and O’Donnell, 2018). Additionally, weed abrasion on lower branches can cause needle browning and drop, reducing tree quality and marketability (Willoughby and Palmer, 1997).
Managing weeds in ornamental crop production requires careful herbicide selection since different species react differently to chemical applications (Figure 1, Table 1). Several herbicides that are safe for established ornamentals, can cause damage to newly planted crops, making weed control especially difficult during propagation (Fausey, 2003). Additionally, because ornamentals vary widely in sensitivity, herbicide manufacturers must test each species under different conditions before including them on product labels, limiting available options for growers (Mervosh and Ahrens, 1998).

Figure 1. Injuries in ornamental plants due to herbicide application. (A) phytotoxicity symptoms of Christmas tree (Fraser fir) included burning and curling of tips. (B) browning and discoloration of needles of Christmas tree (Fraser fir). (C) browning and discoloration of leaves and flowers of Azalea. (D) leaf chlorosis and discoloration of Petunia.
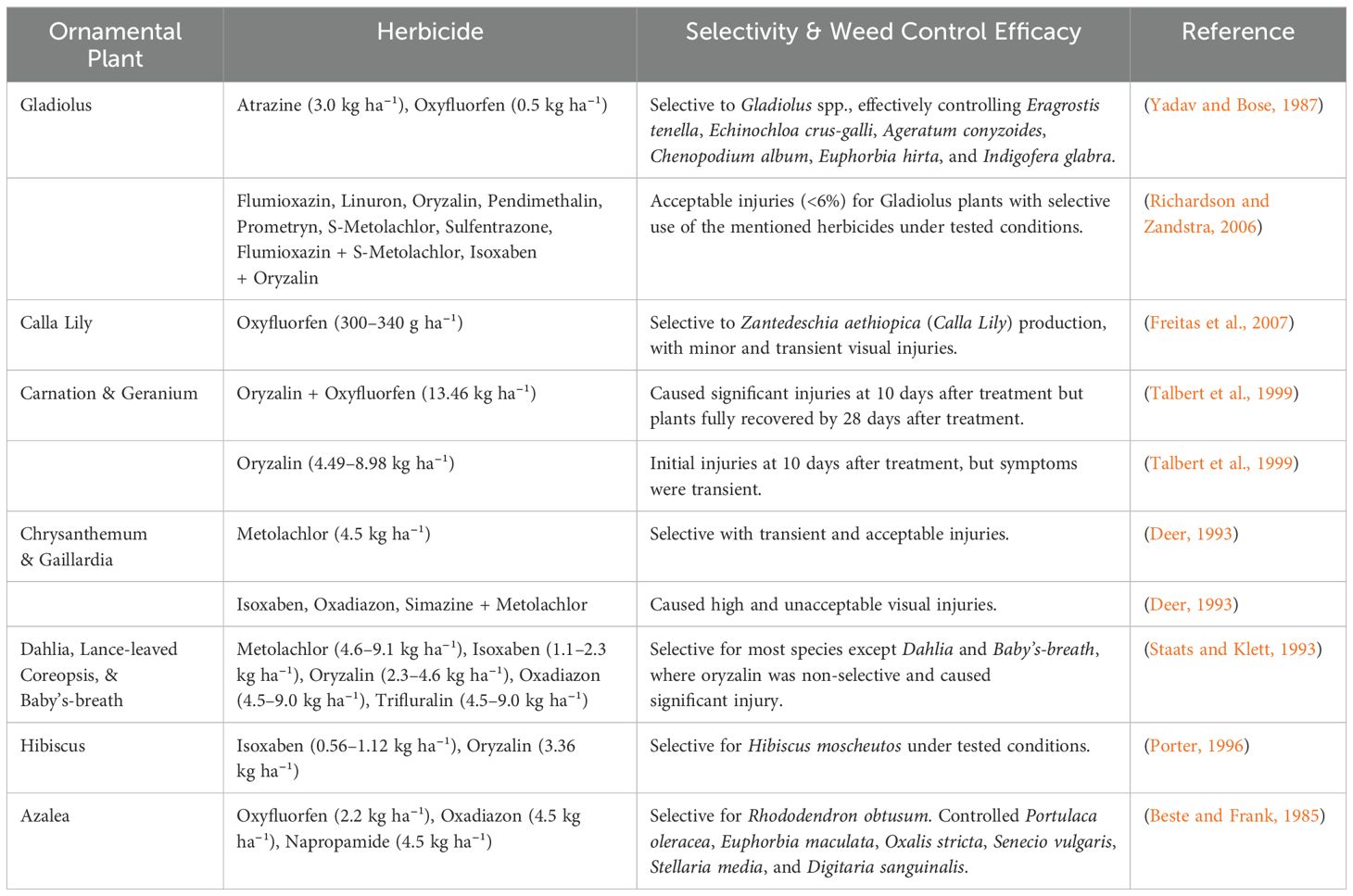
Table 1. Herbicide selectivity of ornamental crops.
Greenhouse production adds another layer of complexity, as very few herbicides are labeled for indoor use due to concerns about volatility and crop damage. Herbicide vapors can accumulate in enclosed spaces and injure crops, while spray drift from ventilation systems can cause unintended exposure (Smith, 2019). (Norcini et al., 1996) noted that herbicide use is lowest in greenhouses compared to field and container-grown ornamentals, forcing growers to rely more on alternative weed management strategies.
While herbicides are an important tool in floriculture and landscaping, their use must be carefully managed to prevent damage to desirable plants. Poor herbicide selectivity can affect plant aesthetics, reducing their value in ornamental landscapes (Seixas et al., 2011; Queiroz et al., 2016).
Elevated CO2 levels are also significantly altering the physiology and morphology of weeds, impacting their growth, reproductive strategies, and competitive interactions in ecosystems. The current levels of atmospheric CO2 are suboptimal for the photosynthesis process in C3 plants, which lose carbon through photorespiration process, making their photosynthesis less efficient than in C4 plants. C4 plants have a special mechanism that concentrates CO2 directly at the site of carboxylation, effectively reducing the carbon loss seen in photorespiration (Acock, 1990; Naidu, 2013). With higher CO2 levels, C3 plants experience an increase in photosynthesis because there’s more CO2 available inside their leaves and CO2 lost by photorespiration will be less (Manisankar and Ramesh, 2019). This leads to greater growth and leaf area in C3 plants compared to C4 plants when CO2 levels rise. For instance, the study of Ziska et al. (1999) showed that the C3 weed Chenopodium album showed increased photosynthesis and reduced stomatal conductance at higher CO2 levels, but these changes were not observed in the C4 weed Amaranthus retroflexus. Additionally, C3 weeds such as parthenium (Parthenium hysterophorus L.) and chromolaena (Chromolaena odorata L.), were expected to become more robust and aggressive under higher CO2 conditions (Chandrasena, 2009).
O’Donnell and Adkins (O’Donnell and Adkins, 2001) found that when the C3 weed wild oat grown at elevated CO2 levels (480 ppm), produced 44% more seed than at lower CO2 levels (357 ppm). Similarly, increased CO2 led to more growth and biomass in the C4 weed Amaranthus viridis (Naidu, 2013). Increased CO2 not only promotes greater branching and leaf area but also affects other characteristics detrimental to human health, such as increased pollen production in ragweed (Ambrosia artemisiifolia), which exacerbates allergies, and a rise in spine production in Cirsium arvense, which can affect animals grazing in infested areas (Ziska and Caulfield, 2000; Ziska, 2002). However, the response to CO2 can vary significantly within and between species, depending on environmental factors such as temperature, light, moisture, and nutrient availability (Patterson, 1995).
The rising levels of atmospheric CO2 have significantly influenced geographical distribution. This change has the potential to make certain weeds more problematic and to allow them to expand into new areas (Malarkodi et al., 2017). Weeds that reproduce vegetatively, such as through roots and stolons, have shown particularly strong growth responses to the recent increases in CO2, underscoring the profound impact of elevated CO2 on invasive species (Ziska and George, 2004). For instance, doubling CO2 levels can enhance the growth of many invasive plants by 46%. These invasive plants have grown more quickly than native plants due to a 30% rise in CO2 levels over the last century (Altland, 2003; Ziska, 2003). Ziska et al. (2011) suggest that higher CO2 levels could alter the biology of invasive weeds, enhancing their growth and potentially increasing their seed dispersal through wind by making plants taller or larger. This could affect invasive species like Cirsium arvense L., Sonchus arvensis L., Sonchus oleraceus L., and Carduus nutans L., known for their wind-dispersed seeds.
Studies have also highlighted how elevated CO2 affects crop-weed competition, with weeds like velvetleaf (Abutilon theophrasti) showing increased biomass, thereby reducing the yield and biomass of competing crops such as sorghum. This interaction is further complicated when common cocklebur (Xanthium strumarium) is present, as it diminishes sorghum’s growth and potential yield under higher CO2 levels (Ziska et al., 2004; Ziska and Runion, 2006). The adaptive responses of weeds to increased CO2, such as enhanced growth and reproductive capacity in Parthenium hysterophorus (Naidu and Paroha, 2008), indicate a shifting dynamic that could make managing these species more challenging under future climate scenarios.
Furthermore, elevated CO2 has been shown to increase the rate and final percentage of weed seed germination. (Ziska and Bunce, 1993) found that doubling atmospheric CO2 concentration (from 350 to 700 μl l⁻¹) enhanced germination in Amaranthus hybridus and Chenopodium album. A separate field experiment demonstrated a significant increase in the total number of weed seedlings three weeks after tilling under elevated CO2 conditions, indicating that rising atmospheric CO2 may promote greater weed establishment.
Temperature plays a crucial role in determining the distribution, growth, development, and phenology of plants, making specific species more suited to certain temperature zones. Plant processes like photosynthesis, respiration, and transpiration are all highly dependent on temperature, and any changes can significantly impact plant growth. The reproductive stage of plants is particularly sensitive to temperature fluctuations. Weeds, like other plants, react to temperature changes and develop strategies to adapt or migrate to more suitable conditions (Metz et al., 2007).
With the expected rise in global temperatures, it is likely that C4 weeds, which thrive under warmer conditions, will outcompete C3 weeds, altering existing weed-crop interactions (Tubiello et al., 2007). Moreover, warmer winters are predicted to enhance the survival rates of winter annual weeds, and increased summer temperatures could lead to more vigorous growth of thermophilic summer annuals, pushing their geographical range further north (Walck et al., 2011; Hanzlik and Gerowitt, 2012). Elevated temperatures can also trigger various physiological changes in weeds, including shortened vegetative phases and life cycles, reduced tillering, decreased seed filling duration and rate, and increased oxidative damage (Jagadish et al., 2011). Studies have shown that weeds like wild oat develop faster in higher temperatures, which can lead to a denser soil seed bank, giving them a competitive advantage over crops (O’Donnell and Adkins, 2001). Additionally, certain summer weed species, such as Sida spinosa (prickly sida), exhibit increased growth and higher root-to-shoot ratios under elevated temperatures (Tungate et al., 2007). It has been also found that climate change will facilitate the spread of weeds like Datura stramonium L., which grows more robustly at higher temperatures (Cavero et al., 1999). Moreover, Hieracium aurantiacum L. has been observed to expand rapidly in Australia due to favorable warm conditions that accelerate its growth and reproductive cycles (Brinkley and Bomford, 2002).
Temperature have influence on the seed germination of weed seeds. The study of (Bandara et al., 2017) shows that elevated temperatures up to 35°C increased germination, while temperatures beyond 35°C caused a significant decline. Grasses and broadleaf weeds showed increased cumulative seed germination (CSG) at 35°C, whereas sedges showed no significant response. Echinochloa crus-galli germination increased with rising temperatures, while Leptochloa chinensis, Lindernia rotundifolia, and Monochoria vaginalis had higher germination at 35°C but declined beyond this point. In contrast, Cyperus deformis germination decreased up to 30°C and then increased at higher temperatures.
Studies have indicated that increased CO2 concentrations may enhance a plant’s ability to endure both hotter and colder temperatures. These adaptabilities are often dependent on how moisture availability is altered through changing rainfall patterns or other environmental factors such as nitrogen levels (Luo and Mooney, 1999; Bunce and Ziska, 2000). Weeds, with their broad ecological tolerance, are expected to take advantage of these changes, outcompeting more specialized, less adaptable species. The warming climate is likely to facilitate the northward migration of tropical and subtropical C4 species, introducing them into temperate zones where they may become aggressive new competitors to the existing flora (Parry, 1998). This emerging scenario highlights the need for innovative and adaptable weed management strategies to address the challenges posed by an evolving weed landscape under climate change.
Increasing temperatures are leading to higher evaporation rates, contributing to drier conditions in monsoon regions, with a projected 5–8% increase in areas vulnerable to drought (Giannini et al., 2008; Rodenburg et al., 2011). Predictions for future rainfall patterns indicate that they will likely become more unpredictable, with droughts and floods becoming more frequent. Such climatic shifts will complicate the control and spread of weeds in agricultural ecosystems, particularly affecting the management of weeds in spring-sown crops during summer droughts (Peters and Gerowitt, 2014). Prolonged drought conditions are expected to favor the growth of C4 and parasitic weeds like Striga hermonthica, while excessive moisture will benefit weeds like Rhamphicarpa fistulosa (Hochst.) Benth (Rodenburg et al., 2010).
Weeds exhibit various adaptive strategies to cope with drought, demonstrating significant plasticity in their response to water stress. For instance, Abutilon theophrasti (velvetleaf) conserves water by shedding its oldest leaves, prioritizing resources for younger leaves to support growth and reproduction under drought conditions (Schmidt et al., 2011). This species also responds to drought by reducing its overall leaf area to maintain higher leaf water potential, a strategy observed in other weeds as well (Ward et al., 1999).
Weeds adapt to high temperatures, which often accompany drought, by developing physical barriers such as leaf hairs or by changing leaf orientation to reduce solar exposure (Hasanuzzaman et al., 2013). Additionally, A. theophrasti has been noted to adjust leaf blade angles to minimize direct sunlight impact, further aiding in temperature regulation (Karkanis et al., 2011). The lifecycle of this weed is also shortened under drought, with earlier flowering times to ensure reproductive success despite adverse conditions (Volis et al., 2004; Volis, 2009; Karkanis et al., 2011).
Root architecture plays a crucial role in drought tolerance. Many weeds increase their root-to-shoot ratios, enhancing their ability to access deeper water sources during dry periods. This is particularly evident in species like Sida spinosa and Convolvulus arvensis, which develop extensive root systems to maximize water uptake (Mekki, 2007; Sosnoskie and Hanson, 2016). Enhanced root growth is supported by biochemical adaptations such as increased solute allocation to roots, which helps plants recover quickly from water stress (Gilgen and Feller, 2013; 2014).
Moreover, weeds often modify their anatomical structures to minimize water loss. Abutilon theophrasti increases leaf epicuticular wax deposition under drought, which helps retain moisture (Hatterman-Valenti et al., 2011). Some species, like Ecballium elaterium and Salsola elaeagnifolium, develop denser leaf tissues with multiple layers of palisade cells, reducing the space for water vapor to escape and helping maintain hydration (Christodoulakis et al., 2009, 2011). These physiological and biochemical responses not only enhance the survival and competitiveness of weeds under drought conditions.
Opportunistic weed species can adapt to climate change effectively due to their advanced dispersal and superior adaptation abilities (Chapin et al., 1996; Bergmann et al., 2010; Pautasso et al., 2010). The dispersion of these species is often facilitated by human activities, such as transportation and trade, which can act as catalysts. For example, the spread of Ambrosia artemisiifolia is not always clearly distinguishable from climate-driven mechanisms (Von der Lippe et al., 2013; Milakovic et al., 2014). The range extension of maize weeds of the genus Setaria and A. theophrasti to Northern America may result from the interaction between human activities and climatic changes (Andersen et al., 1985; Douglas et al., 1985; Warwick and Black, 1986; Wang and Dekker, 1995).
Climate change is anticipated to significantly influence the composition of weed species in arable lands. The pace of predicted climate change surpasses any climate variations recorded over the past 420,000 years (Petit et al., 1999; Loss et al., 2011), positioning it as a potentially major factor in shaping the distribution of arable weeds (Pautasso et al., 2010). Changes in climate result in modifications to environmental conditions, including temperature and precipitation, which directly impact arable weeds. Additionally, these climate changes also indirectly impact weed dynamics by forcing farmers to adjust their practices, including choosing different crops and changing the timing of planting and harvesting (Fleming and Vanclay, 2010).
Higher temperatures influence seed emergence, potentially making some dormant weeds invasive and altering species distribution. Increased aridity and elevated CO2 levels particularly boost the growth of C4 weeds under stress conditions. These changes enhance the competitive advantage of weeds over crops, posing increased challenges for agriculture and ecosystem management. Elevated CO2 levels enhance the growth of C3 plants more than C4 plants, with C3 plants showing a 41% growth increase compared to 22% for C4 plants when CO2 concentration is doubled (Poorter, 1993). This is due to decreased photorespiration and increased net photosynthesis in C3 plants, improving their overall carbon balance and water use efficiency.
One important example of climate influence is the potential northward migration of Pueraria lobata (kudzu), an invasive species often found across the United States. About 15 years ago, kudzu was predominantly found up to the Ohio Valley and the Mason-Dixon line, restricted by colder winter temperatures (Sasek and Strain, 1990). However, recent trends have shown an increase in kudzu populations linked to rising winter temperatures. Additionally, the expected rise in nighttime temperatures relative to daytime temperatures due to global warming could more significantly reduce seed production in crops compared to weeds, such as in the case of Vigna unguiculata (Ahmed et al., 1993), thereby impacting competitive dynamics. As the climate warms, many weeds that thrive in warm-season conditions, typically originating from tropical areas, are likely to extend their range northward.
Mechanical methods are predominantly used in developed and developing countries to manage weed populations. This approach is globally recognized as a fundamental weed control strategy in agricultural systems. However, climatic changes, particularly an increase in CO2 levels, could complicate this method by enhancing underground carbon storage and consequently boosting the growth of roots or rhizomes in perennial weeds (Rogers et al., 1994). Such increases in root ratios and subterranean growth can make it challenging to eradicate certain perennial weeds, like Canada thistle (Cirsium arvense), that can regenerate from root fragments left in the soil after tillage (Ziska et al., 2004). Similar responses may be seen in weeds such as skeletonweed (Chondrilla juncea) and silverleaf nightshade (Solanum elaeagnifolium) (Kriticos et al., 2010).
Moreover, climate change can also influence the effectiveness of cover crops as a non-chemical weed management method by altering their establishment, biomass production, and competitive interactions with weeds. (DuPre et al., 2022) found that rising temperatures and decreasing moisture availability affected early- and mid-season cover crop mixtures differently, leading to shifts in weed biomass and species composition. In semi-arid regions, cover crops must balance weed suppression with soil moisture conservation (Kumar et al., 2020). The early-season five-species mixture, including oat, brassicas, and legumes, produced the highest biomass and maintained cooler soil temperatures, improving weed suppression. However, it significantly reduced soil moisture, which could limit subsequent crop yields, a challenge that may intensify under warming conditions (Whitlock et al., 2017). In contrast, the mid-season seven-species mixture, dominated by brassicas, showed stable biomass production and increased brassica abundance, suggesting brassicas may be more resilient for weed suppression in warming and drying climates (Franco et al., 2021). Species composition, planting time, and environmental conditions play key roles in determining cover crop success in weed management (Carr et al., 2004; Florence and McGuire, 2020; Smith et al., 2020).
Furthermore, climate change impacts on soil temperature, moisture availability, and weed seed emergence patterns can significantly influence the effectiveness of mulching as a weed management strategy. Noted by (Singh et al., 2015) that weed seeds present at different depths respond to environmental factors such as temperature fluctuations, soil moisture levels, light availability, and predator activity. Tillage practices influence weed seed distribution, with minimum tillage (MT) and zero tillage (ZT) systems concentrating most weed seeds near the soil surface, whereas conventional tillage (CT) distributes them more uniformly throughout the tilled layer (Chauhan and Johnson, 2009). As climate change alters rainfall patterns and soil temperature, these differences in weed seed placement and emergence become more pronounced.
Additionally, biological control methods, which involve introducing host-specific natural enemies such as insects, fungi, or herbivores to target weeds, are expected to be impacted by changes in climate. These environmental changes can potentially affect both the biological control agents and the weeds they target, leading to diminished efficacy of biocontrol measures. Climatic changes might disrupt established biocontrol systems by altering the morphological development and reproductive cycles of the biocontrol agents. Increased CO2 levels could also change the chemical properties of weeds, influencing their interactions with herbivores. This could involve alterations in the carbon-to-nitrogen ratio in plants, affecting the feeding preferences and growth rates of herbivorous pests (Singh et al., 2011, 2016). The changes brought by global warming might even offer new opportunities for employing certain insect species, like Ophraella communa, as biocontrol agents against specific weed infestations (Cardarelli et al., 2018).
Furthermore, elevated CO2 levels may alter the secondary compound profiles of weeds, affecting weed-herbivore interactions and potentially complicating biocontrol strategies. Increased atmospheric CO2 could lead to changes in the feeding habits and growth rates of herbivores, impacting the dynamics of biocontrol across various ecosystems (Ziska et al., 2005; Malarkodi et al., 2017). As greenhouse gas concentrations rise, the relationship between plants and herbivores may evolve, affecting the efficacy of biocontrol at both spatial and temporal scales. Factors such as drought may also increase levels of certain insect-resistance chemicals in weeds, which could render biocontrol methods less effective, particularly in drier conditions (Gerard et al., 2010). Moreover, temperature increases might influence the distribution and viability of both biocontrol agents and their target hosts. For instance, elevated temperatures could facilitate the migration of biocontrol agents from subtropical to temperate areas, impacting the overall effectiveness of biocontrol systems. Faster life cycles of both biocontrol agents and weeds due to higher temperatures could further complicate biocontrol strategies (Kriticos et al., 2010).
These complex interactions underscore the challenges and uncertainties in predicting how climate change will ultimately affect the effectiveness of both mechanical and biological weed control methods. As Patterson (Patterson, 1995) noted, warming could extend the overwintering range of insect populations, potentially altering their geographical spread. Additionally, elevated CO2 may change plant nutrient content, which, along with shifts in temperature, could influence the activity of insects and the phenology of both biocontrol agents and weeds (Reeves, 2017). The differential capacities of weeds and their bioagents to adapt to these changing conditions make it difficult to predict the outcomes of such climatic shifts (Holt and Hochberg, 1997).
Changes in temperature, wind speed, soil moisture, and atmospheric humidity can influence the effectiveness of herbicides (Muzik, 1976). The efficiency of certain herbicides can be altered by the increasing CO2 levels and warmer and wetter conditions by influencing the physiology of plants. Glyphosate uptake in plants like Desmodium tortuosum (Sw.) DC., a C3 weed, is influenced by atmospheric temperature (Sharma and Singh, 2001). Similarly, increases in temperature or humidity have been shown to triple the effectiveness of mesotrione on weeds such as Xanthium strumarium and A. theophrastii (Johnson and Young, 2002). On the other hand, the herbicide pyrithiobac, part of the pyrimidinylthiobenzoic acid group, shows diminished effectiveness on Amaranthus palmeri L. when temperatures stray from the 20-34°C range (Mahan et al., 2004). Additionally, Anderson et al. (1993) found that the effectiveness of glufosinate-ammonium depends on humidity, as it influences the moisture level of the plant’s surface (cuticle). Ramsey et al. (2005) reported that relative humidity impacts the rate at which droplets dry on plant surfaces.
With the increase of the CO2 concentration, the effectiveness of herbicides may decrease. Increased CO2 levels cause morpho-physiological and anatomical changes in plants, impacting the uptake and movement of herbicides within them (Ziska and Teasdale, 2000; Manea et al., 2011). In C3 plants, there is a reduction in the number and conductance of stomata, as well as an increase in leaf thickness, which can interfere with the absorption of herbicides through the foliage (Nowak et al., 2004; Ainsworth and Long, 2005). Additionally, higher starch accumulation on the leaf surface can occur (Patterson, 1995). Perennial weeds such as rhodes grass (Chloris gayana), weeping lovegrass (Eragostis curvula), and dallisgrass (Paspalum dilatatum) might become more problematic if their vegetative growth is stimulated by enhanced photosynthesis due to elevated CO2 levels. These changes are expected to reduce the effectiveness of commonly used herbicides like glyphosate, possibly due to a dilution effect, although the exact mechanism leading to increased glyphosate tolerance is still unclear (Manea et al., 2011). One potential reason could be reduced herbicide translocation as the root system becomes more robust, and an increased root-to-shoot ratio may also play a crucial role in herbicide efficacy (Ziska et al., 2004). Common lambsquarters (Chenopodium album), a C3 plant has demonstrated a higher tolerance to glyphosate because increased CO2 promotes its growth and biomass (Ziska et al., 1999). Furthermore, higher concentrations of CO2 tend to boost the growth of rhizomes or tubers in many perennial weeds more than their aboveground parts, potentially complicating the control of these weeds with herbicides (Patterson et al., 1999). For example, research on Canada thistle (Cirsium arvense) has shown that elevated CO2 increases the root-to-shoot ratio in field conditions, leading to a dilution effect that reduces glyphosate’s effectiveness due to enhanced below-ground growth (Ziska et al., 2004).
In weed control, the timing of application could also be affected and decrease the chemical susceptibility of the weeds in the seedling stage under the elevated CO2 condition. For perennial weeds, increasing CO2 could stimulate greater below ground growth (rhizomes, tubers, and roots), diluting the active ingredient and making chemical control more difficult and costly. At the biochemical level, CO2 could alter herbicide-specific chemistry in such a way as to directly reduce the efficacy of the active ingredient (Bowes, 1996). Rising CO2 levels can decrease the chemical efficacy in the control of annual and perennial weeds (Ziska and Reunion, 2007). Increased spraying frequency could overcome CO2-induced reductions in efficacy but could increase residual effects within the environment (Ziska et al., 2004). Increased use of herbicides for control of weeds carries further risks for human and animal health because it could increase the presence of these chemicals in the environment. Mechanical tillage may lead to additional plant propagation in a higher CO2 environment, with increased asexual reproduction from belowground structures and negative effects on weed control (e.g., Canada thistle) (Malarkodi et al., 2017).
Temperature significantly impacts herbicide effectiveness in both direct and indirect ways. Temperature influences the physical properties of herbicides, affecting their diffusion rates, the viscosity of plant cuticle waxes, and the properties of the spray solutions themselves (Price, 1983). For instance, higher temperatures can reduce the viscosity of cuticular lipids and enhance the herbicide permeability and diffusion which is evidenced by increased uptake and translocation of 14C-glyphosate in Desmodium tortuosum at higher temperatures (Sharma and Singh, 2001). Similarly, the efficacy of Flumiclorac on common lambsquarters and redroot pigweed significantly increased with rising temperatures, from 10°C to 40°C (Fausey and Renner, 2001). However, while higher air temperatures generally accelerate the absorption and movement of most foliar-applied herbicides, they can also trigger faster herbicide metabolism in plants, which might reduce the effectiveness of the herbicide (Kells et al., 1984; Johnson and Young, 2002). For example, although the absorption and translocation of mesotrione in velvetleaf and common cocklebur tripled at 32°C, its efficacy decreased markedly on common waterhemp and large crabgrass under the same conditions (Johnson and Young, 2002).
The effectiveness of specific herbicide is dependent on temperature. The effectiveness and selectivity of both pre-emergence and post-emergence herbicides can be reduced due to prolonged high temperature (Madafiglio et al., 2000; Medd et al., 2001). This effect can also cause a selective herbicide to behave non-selectively when temperatures exceed the optimal range (Amare, 2016). Furthermore, Bailey (Bailey, 2003) reported that high temperatures accelerate the degradation of herbicides, and this less persistence complicates weed management during colder seasons and leading to problems with herbicide carry-over in warmer seasons. Increased temperatures also reduce protein synthesis in plants, which decreases their demand for amino acids necessary for the action of enzyme inhibitor herbicides such as glyphosate [(N-phosphonomethyl) glycine], thereby reducing their efficacy (Varanasi et al., 2015). Conversely, Patterson et al. (1999) found that higher temperatures might enhance the uptake and translocation of many herbicides, potentially increasing their effectiveness. This is evident in the performance of flumetsulam against Raphanus raphanistrum and clodinafop-propargyl against Avena spp., which both show improved efficacy at higher temperatures (Madafiglio et al., 2000; Medd et al., 2001).
Furthermore, high soil temperatures can diminish the efficacy of soil-applied herbicides by increasing their volatility and the rate of microbial degradation. A study by Atienza et al. (2001) demonstrated that the volatilization of triallate herbicide from soils increased significantly with temperature, highlighting substantial losses in both sandy and loamy soils as temperatures rose from 5°C to 25°C.
Enhanced frequency and intensity of rainfall can significantly impair the effectiveness of soil-applied herbicides by adversely affecting their uptake, retention, and overall activity in the soil (Bailey, 2004; Rodenburg et al., 2011). This can lead to a decreased ability of these chemicals to control weed populations effectively. Additionally, environmental condition such as drought can cause plants to develop physiological changes, such as increased cuticle thickness and more pronounced leaf pubescence. These adaptations serve as barriers, making it more difficult for herbicides to penetrate into the leaves and perform effectively (Patterson, 1995). Such changes not only impact the efficacy of weed control methods but also influence the recovery and subsequent growth of both crops and weeds after herbicide application.
Furthermore, as aridity and drought conditions become more common, there is an increased risk of herbicides volatilizing into the atmosphere, which reduces their availability to act on target weeds. On the other hand, frequent and heavy rainfalls can disrupt the timing of herbicide applications by shortening the “rain safe periods,” the critical windows during which herbicides can be applied effectively without being washed off by rain. This complicates scheduling and reduces the flexibility of managing weed control within agricultural systems.
Intense rainfall events or consistent heavy rains throughout the year can exacerbate the leaching of soil-applied herbicides, potentially causing them to percolate through the soil profile and contaminate groundwater sources (Froud-Williams, 1996). This not only poses environmental risks but also leads to inefficiencies in herbicide usage.
To illustrate the impacts of elevated CO2, temperature, and other climatic factors, including soil moisture, on herbicide efficacy for both C3 and C4 weeds, Tables 2, 3 summarize key findings from recent studies.
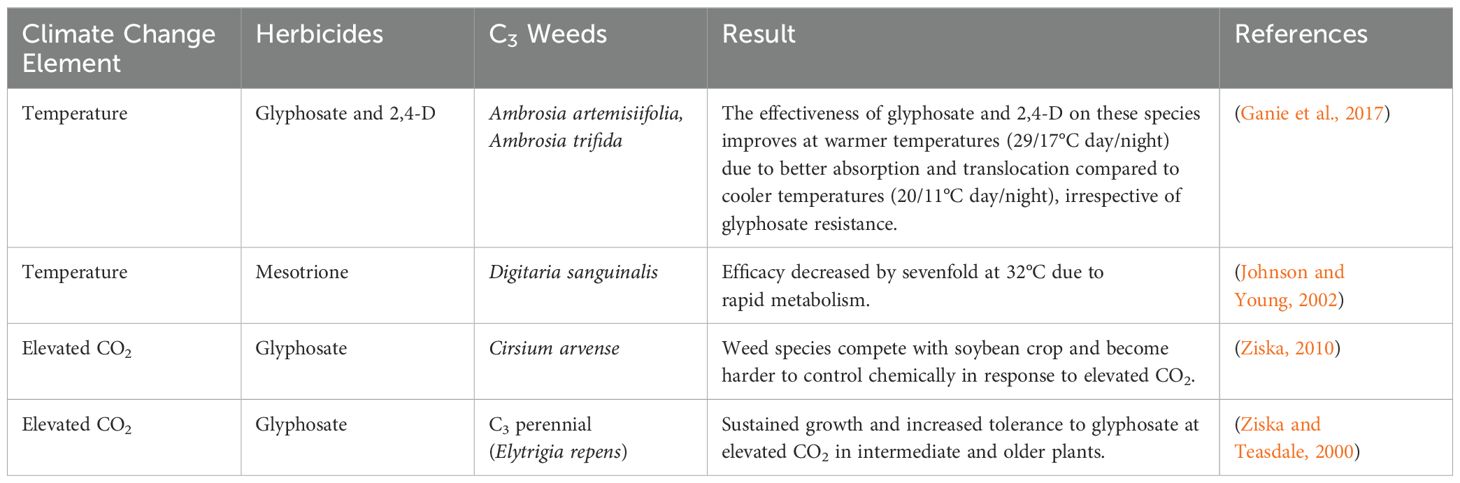
Table 2. Herbicides efficacy on C3 weeds under different climate change elements.
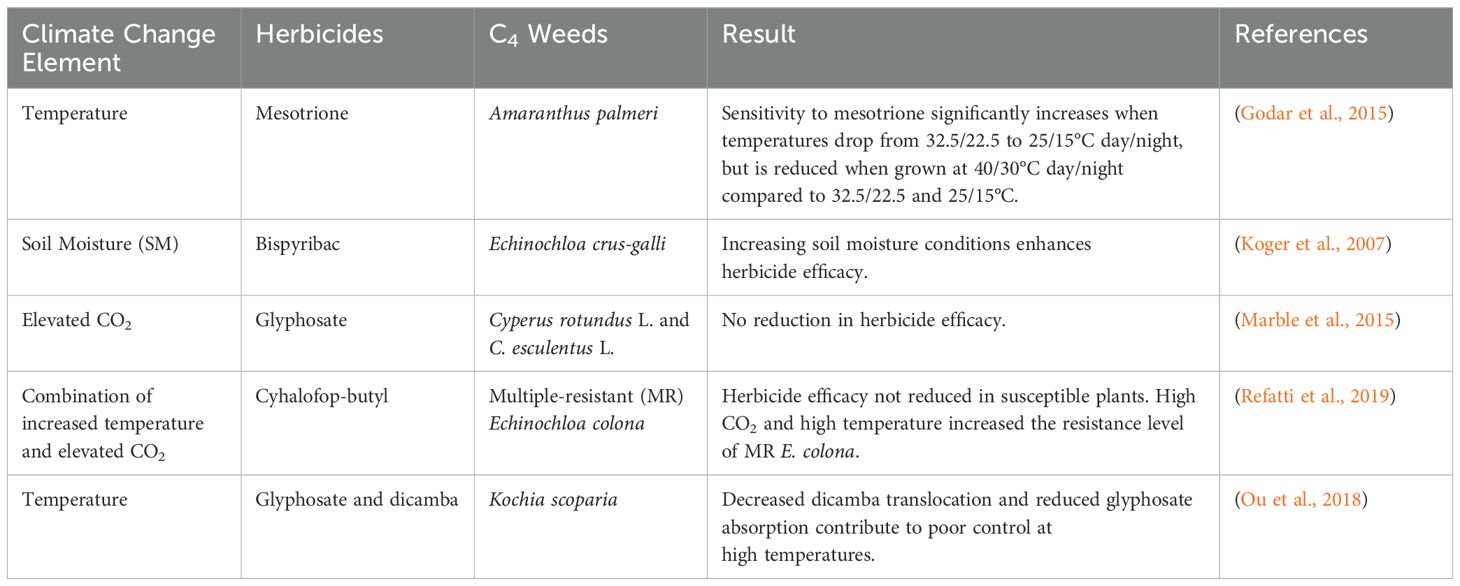
Table 3. Herbicides efficacy on C4 weeds under different climate change elements.
Climate change intensifies herbicide resistance by promoting the dissemination and evolution of resistant biotypes. Elevated CO2 levels have been shown to increase glyphosate tolerance in both C4 species, such as redroot pigweed (Amaranthus retroflexus), and C3 species, such as common lambsquarters (Chenopodium album) (Ziska et al., 1999). In Cirsium arvense (Canada thistle), reduced glyphosate efficacy under elevated CO2 was attributed to increased root biomass and a dilution effect (Ziska et al., 2004). Similarly, Elytrigia repens (quackgrass) exhibited greater herbicide resistance under CO2-enriched environments due to faster growth and a shortened seedling stage, which is typically the most vulnerable phase (Ziska and Teasdale, 2000).
The durability of herbicide management in genetically modified (GM) crops is closely linked to climate stability. Climate variations can affect the movement of genes and the spread of resistance traits between GM crops and their related weed species, depending on their genetic compatibility (Ziska et al., 2019). (Ziska et al., 2012) observed greater floral synchrony and higher outcrossing rates between herbicide-resistant rice (Clearfield™ 161) and weedy red rice (StgS) under elevated CO2 levels (300, 400, and 600 ppm), leading to an increase in herbicide-resistant hybrid progeny. However, the impact of elevated temperatures on gene flow and resistance evolution remains uncertain.
Dissemination of resistant biotypes is also driven by spatial mechanisms, such as wind dispersal or transportation through farm machinery, and temporal mechanisms, including persistent seed and bud banks, particularly in perennial weeds (Ott et al., 2019). These processes increase the prevalence of resistant weeds locally and regionally, compounding management challenges. For instance, Bassia scoparia (kochia) has become a globally significant weed due to its widespread resistance to glyphosate, facilitated by both herbicide selection pressures and climate change (Kaur et al., 2024).
Additionally, herbicide-resistant crops could alter weed communities, leading to reduced herbicide efficacy under changing climatic conditions. These findings underscore the urgent need to develop adaptive weed management strategies to mitigate the compounded effects of herbicide resistance and climate change (Ziska and Teasdale, 2000; Singh et al., 2011).
The findings from this review indicate that climate change will significantly impact ornamental crop production, weed management practices, and the growth and distribution of weeds. Understanding how weed species respond to elevated CO2, rising temperatures, and drought stress is critical, particularly given the unique challenges of ornamental production systems. Unlike agronomic crops, ornamental crops often involve multiple species grown in close proximity, where aesthetic qualities are prioritized over yield, complicating weed management. Limited research has addressed how climate change affects herbicide efficacy in ornamental settings, such as Christmas tree production, due to funding constraints and research focus on agronomic systems.
There is an urgent need to investigate how increasing temperatures and CO2 levels alter the morphology of weeds, making them more adaptive to these changing conditions. Additionally, drought stress and shifting precipitation patterns may influence weed management practices and herbicide performance in nurseries and greenhouses. Future research should address these gaps to develop sustainable, climate-resilient weed management strategies tailored to ornamental crop production.
SM: Investigation, Validation, Writing – original draft. MH: Writing – review & editing. ZH: Writing – review & editing. EP: Writing – review & editing. DS: Conceptualization, Funding acquisition, Investigation, Resources, Supervision, Validation, Visualization, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The author(s) declare financial support was received for the research, authorship, and/or publication of this article. United States Department of Agriculture (USDA) National Institute of Food and Agriculture, Hatch Project Number MICL02670.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acock B. (1990). “Effects of carbon dioxide on photosynthesis, plant growth, and other processes,” in Impact of Carbon Dioxide, Trace Gases, and Climate Change on Global Agriculture. Eds. Bruce A., Chair K., Rosenberg N. J., Allen L. H. Jr. (Madison, WI, USA: American Society of Agronomy, Inc.; Crop Science Society of America, Inc.; Soil Science Society of America, Inc), 45–60.
Adams R. M., Hurd B. H., Lenhart S., Leary N. (1998). Effects of global climate change on agriculture: An interpretative review. Climate Res. 11, 19–30. doi: 10.3354/cr011019
Ahmed F. E., Hall A. E., Madore M. A. (1993). Interactive effect of high temperature and elevated carbon dioxide concentration on cowpea. Plant Cell Environ. 16, 835–842. doi: 10.1111/j.1365-3040.1993.tb00505.x
Ainsworth E. A., Long S. P. (2005). What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties, and plant production to rising CO2. New Phytol. 165, 351–372. doi: 10.1111/j.1469-8137.2004.01224.x
Altland J. E. (2003). Weed Control in Container Crops: A Guide to Effective Weed Management through Preventive Measures (Corvallis, OR, USA: Oregon State University Extension Service).
Amare T. (2016). Review on impact of climate change on weed and their management. J. Agric. Biol. Environ. Stat 2, 21–27. doi: 10.11648/j.ajbes.20160203.12
Amthor J. S. (1995). Terrestrial higher-plant response to increasing atmospheric CO2 in relation to the global carbon cycle. Global Change Biol. 1, 243–274. doi: 10.1111/j.1365-
Andersen R. N., Menges R. M., Conn J. S. (1985). Variability in velvetleaf (Abutilon theophrasti) and reproduction beyond its current range in North America. Weed Sci. 33, 507–512. doi: 10.1017/S0043174500082746
Anderson D. M., Swanton C. J., Hall J. C., Mersey B. G. (1993). The influence of temperature and relative humidity on the efficacy of glufosinate-ammonium. Weed Res. 33, 139–147. doi: 10.1111/j.1365-3180.1993.tb01927.x
Atienza J., Tabernero M. T., Álvarez-Benedí J., Sanz M. (2001). Volatilisation of triallate as affected by soil texture and air velocity. Chemosphere 42, 257–261. doi: 10.1016/S0045-6535(00)00075-8
Bailey S. W. (2003). Climate change and decreasing herbicide persistence. Pest Manage. Sci. 60, 158–162. doi: 10.1002/ps.v60:2
Bailey S. W. (2004). Climate change and decreasing herbicide persistence. Pest Manage. Sci. 60, 158–162. doi: 10.1002/ps.v60:2
Bandara R. M. U. S., Ilangakoon T. K., Dissanayaka H.M.M.K.K.H., de Silva Y.M.S.H.I.U., Piyasiri C. H., Dissanayaka D. M. C. B. (2017). Effect of elevated temperature on weed seed germination in paddy soil seed bank. Proceedings of the 4th International Conference on Agriculture and Forestry 3, 16–18. doi: 10.17501/icoaf.2017.3103
Beeson R. C. (1991). Scheduling woody plants for production and harvest. HortTechnology 1, 30–35. doi: 10.21273/HORTTECH.1.1.30
Bergmann J., Pompe S., Ohlemüller R. (2010). The Iberian Peninsula as a potential source for the plant species pool in Germany under projected climate change. Plant Ecol. 207, 191–201. doi: 10.1007/s11258-009-9664-6
Beste C. E., Frank J. R. (1985). Weed control in newly planted azaleas. J. Environ. Horticulture 3, 12–14. doi: 10.24266/0738-2898-3.1.12
Bowes G. (1996). “Photosynthetic responses to changing atmospheric carbon dioxide concentration,” in Photosynthesis and the Environment. Ed. Baker N. R. (Kluwer Publishing, Dordrecht, The Netherlands), 387–407.
Brinkley T. R., Bomford M. (2002). Agricultural sleeper weeds: What is the potential threat? (Kingston, ACT: Bureau of Rural Sciences).
Brown J. H., Cowen W. F. Jr., Heiligmann R. B. (1991). Ohio Christmas tree producers manual. (Columbus, OH, USA: Ohio State University Extension Publications). doi: 10.1079/9780851994390.0333
Bunce J. A., Ziska L. H. (2000). Crop ecosystem responses to climatic change: Crop/weed interactions. Climate Change Global Crop Productivity 333-348.
Cardarelli E., Musacchio A., Montagnani C., Bogliani G., Citterio S., Gentili R. (2018). Ambrosia artemisiifolia control in agricultural areas: Effect of grassland seeding and herbivory by the exotic leaf beetle Ophraella communa. NeoBiota 38, 1–22. doi: 10.3897/neobiota.38.23562
Carr P. M., Horsley R. D., Poland W. W. (2004). Barley, oat, and cereal-pea mixtures as dryland forages in the Northern Great Plains. Agron. J. 96, 677–684. doi: 10.2134/agronj2004.0677
Case L., Mathers H., Senesac A. (2005). A review of weed control practices in container nurseries. HortTechnology 15, 535–545. doi: 10.21273/HORTTECH.15.3.0535
Cavero J., Zaragoza C., Suso M. L., Pardo A. (1999). Competition between maize and Datura stramonium in an irrigated field under semi-arid conditions. Weed Res. 39, 225–240. doi: 10.1046/j.1365-3180.1999.00140.x
Challinor A. J., Watson J., Lobell D. B., Howden S. M., Smith D. R., Chhetri N. (2014). A meta-analysis of crop yield under climate change and adaptation. Nat. Climate Change 4, 287–291. doi: 10.1038/nclimate2153
Chandrasena N. (2009). How will weed management change under climate change? Some perspectives. J. Crop Weed 5, 95–105. https://www.cropandweed.com/vol5issue2/pdf/25.pdf.
Chapin F. S. III, Bret-Harte M. S., Hobbie S. E., Zhong H. (1996). Plant functional types as predictors of transient responses of arctic vegetation to global change. J. Vegetation Sci. 7, 347–358. doi: 10.2307/3236278
Chauhan B. S., Johnson D. E. (2009). Influence of tillage systems on weed seedling emergence pattern in rainfed rice. Soil Tillage Res. 106, 15–21. doi: 10.1016/j.still.2009.10.004
Chen C. C., McCarl B. A. (2001). An investigation of the relationship between pesticide usage and climate change. Climatic Change 50, 475–487. doi: 10.1023/A:1010655503471
Christodoulakis N. C., Lampri P. N., Fasseas C. (2009). Structural and cytochemical investigation of the leaf of silverleaf nightshade (Solanum elaeagnifolium), a drought-resistant alien weed of the Greek flora. Aust. J. Bot. 57, 432–438. doi: 10.1071/BT08210
Christodoulakis N. S., Kollia K., Fasseas C. (2011). Leaf structure and histochemistry of Ecballium elaterium (L.) A. Rich. (squirting cucumber). Flora – Morphology Distribution Funct. Ecol. Plants 206, 191–197. doi: 10.1016/j.flora.2010.03.004
Compant S., van der Heijden M. G. A., Sessitsch A. (2010). Climate change effects on beneficial plant-microorganism interactions. FEMS Microbiol. Ecol. 73, 197–214. doi: 10.1111/j.1574-6941.2010.00900.x
Cui M., Smith W. K. (1991). Photosynthesis, water relations, and mortality in Abies lasiocarpa seedlings during natural establishment. Tree Physiol. 8, 37–46. doi: 10.1093/treephys/8.1.37
DaMatta F. M., Grandis A., Arenque B. C., Buckeridge M. S. (2010). Impacts of climate changes on crop physiology and food quality. Food Res. Int. 43, 1814–1823. doi: 10.1016/j.foodres.2009.11.001
Darras A. I. (2020). Implementation of sustainable practices to ornamental plant cultivation worldwide: A critical review. Agronomy 10, 1570. doi: 10.3390/agronomy10101570
Datta S. (2013). Impact of climate change in Indian horticulture - A review. Int. J. Science Environ. Technol. 2, 661–671.
Deer J. F. (1993). Wildflower tolerance to metolachlor and metolachlor combined with other broadleaf herbicides. HortScience 10, 1023–1026. doi: 10.21273/HORTSCI.28.10.1023
Douglas B. J., Thomas A. G., Morrison I. N., Maw M. G. (1985). The biology of Canadian weeds. 70. Setaria viridis (L.) Beauv. Can. J. Plant Sci. 65, 669–690. doi: 10.4141/cjps85-089
DuPre M. E., Seipel T., Bourgault M., Boss D. L., Menalled F. D. (2022). Predicted climate conditions and cover crop composition modify weed communities in semiarid agroecosystems. Weed Res. 62, 38–48. doi: 10.1111/wre.12514
Elad Y., Pertot I. (2014). Climate change impact on plant pathogens and plant diseases. J. Crop Improvement 28, 99–139. doi: 10.1080/15427528.2014.865412
Fain G. B., Knight P. R., Gilliam C. H., Olive J. W. (2003). Effect of fertilizer placement on prostrate spurge growth in container production. J. Environ. Horticulture 21, 177–180. doi: 10.24266/0738-2898-21.4.177
Fausey J. C. (2003). Controlling liverwort and moss now and in the future. HortTechnology 13, 35–38. doi: 10.21273/HORTTECH.13.1.0035
Fausey J. C., Renner K. A. (2001). Environmental effects on CGA248757 and efficacy/soybean flumiclorac tolerance. Weed Sci. 49, 668–674. doi: 10.1614/0043-1745(2001)049[0668:EEOCAF]2.0.CO;2
Fleming A., Vanclay F. (2010). Farmer responses to climate change and sustainable agriculture: A review. Agron. Sustain. Dev. 30, 11–19. doi: 10.1051/agro/2009028
Florence A. M., McGuire A. M. (2020). Do diverse cover crop mixtures perform better than monocultures? A systematic review. Agron. J. 112, 3513–3534. doi: 10.1002/agj2.20443
Franco J. G., Gramig G. G., Beamer K. P., Hendrickson J. R. (2021). Cover crop mixtures enhance stability but not productivity in a semi-arid climate. Agron. J. 113, 2264–2680. doi: 10.1002/agj2.20695
Freitas F. C. L., Grossi J. A. S., Barros A. F., Mesquita E. R., Ferreira F. A. (2007). Weed control in ornamental plant seedling production. Planta Daninha 25, 595–601. doi: 10.1590/S0100-83582007000300020
Fretz T. A. (1972). Weed competition in container-grown Japanese holly. HortScience 7, 485–486. doi: 10.21273/HORTSCI.7.6.485
Froud-Williams R. J. (1996). Weeds and climate change: implications for their ecology and control. Aspects Appl. Biol. 45, 187–196.
Ganie Z. A., Jugulam M., Jhala A. J. (2017). Temperature influences efficacy, absorption, and translocation of 2,4-D or glyphosate in glyphosate-resistant and glyphosate-susceptible common ragweed (Ambrosia artemisiifolia) and giant ragweed (Ambrosia trifida). Weed Sci. 65, 588–602. doi: 10.1017/wsc.2017.32
Gautam H. R., Bhardwaj M. L., Kumar R. (2013). Climate change and its impact on plant diseases. Curr. Sci. 105, 1685–1691.
Gerard P. J., Kean J. M., Phillips C. B., Fowler S. V., Walker G. P., Charles J. G. (2010). Possible impacts of climate change on biocontrol systems in New Zealand (New Zealand: AgResearch).
Giannini A., Biasutti M., Held I. M., Sobel A. H. (2008). A global perspective on African climate. Climatic Change 90, 359–383. doi: 10.1007/s10584-008-9396-y
Gilgen A. K., Feller U. (2013). Drought stress alters solute allocation in broadleaf dock (Rumex obtusifolius). Weed Sci. 61, 104–108. doi: 10.1614/WS-D-12-00074.1
Gilgen A. K., Feller U. (2014). Effects of drought and subsequent rewatering on Rumex obtusifolius leaves of different ages: Reversible and irreversible damages. J. Plant Interact. 9, 75–81. doi: 10.1080/17429145.2013.765043
Godar A. S., Varanasi V. K., Nakka S., Prasad P. V., Thompson C. R., Mithila J. (2015). Physiological and molecular mechanisms of differential sensitivity of Palmer amaranth (Amaranthus palmeri) to mesotrione at varying growth temperatures. PloS One 10. doi: 10.1371/journal.pone.0126731
Hanzlik K., Gerowitt B. (2012). Occurrence and distribution of important weed species in German winter oilseed rape fields. J. Plant Dis. Prot. 119, 107–120. doi: 10.1007/BF03356429
Harper G. J., Comeau P. G., Biring B. S. (2005). A comparison of herbicide and mulch mat treatments for reducing grass, herb, and shrub competition in the BC interior Douglas-fir zone—Ten-year results. Western J. Appl. Forestry 20, 167–176. doi: 10.1093/wjaf/20.3.167
Hasanuzzaman M., Nahar K., Alam M., Roychowdhury R., Fujita M. (2013). Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 14, 9643–9684. doi: 10.3390/ijms14059643
Hatterman-Valenti H., Pitty A., Owen M. (2011). Environmental effects on velvetleaf (Abutilon theophrasti) epicuticular wax deposition and herbicide absorption. Weed Sci. 59, 14–21. doi: 10.1614/WS-D-10-00061.1
Hazra P., Samsul H. A., Sikder D., Peter K. V. (2007). Breeding tomato (Lycopersicon esculentum Mill) resistant to high temperature stress. Int. J. Plant Breed. 1, 31–40.
Hirpo F. H., Gebeyehu M. N. (2019). Review on the effects of climate change variability on horticultural productivity. Int. J. Environ. Sci. Natural Resour. 17, 555968. doi: 10.19080/IJESNR.2019.17.555969
Holt R. D., Hochberg M. E. (1997). When is biological control evolutionarily stable (or is it)? Ecology 78, 1673–1683. doi: 10.1890/0012-9658(1997)078[1673:WIBCES]2.0.CO;2
Jagadish K. S., Cairns J. E., Kumar A., Somayanda I. M., Craufurd P. Q. (2011). Does susceptibility to heat stress confound screening for drought tolerance in rice. Funct. Plant Biol. 38, 261–269. doi: 10.1071/FP10224
Johnson B. C., Young B. G. (2002). Influence of temperature and relative humidity on the foliar activity of mesotrione. Weed Sci. 50, 157–161. doi: 10.1614/0043-1745(2002)050[0157:IOTARH]2.0.CO;2
Karkanis A., Bilalis D., Efthimiadou A. (2011). Architectural plasticity, photosynthesis and growth responses of velvetleaf (Abutilon theophrasti Medicus) plants to water stress in a semi-arid environment. Aust. J. Crop Sci. 5, 369–374.
Kaur J., Gosal S. K., Kaur P. (2014). Effects of climate change on plant associated microbial communities and enzyme activities. Afr. J. Microbiol. Res. 8, 3087–3093. doi: 10.5897/AJMR2014.6750
Kaur R., Kumar S., Ali S. A., Kumar S., Ezing U. M. (2024). Impacts of climate change on crop-weed dynamics: Challenges and strategies for weed management in a changing climate. Open J. Environ. Biol. 9, 15–21. doi: 10.17352/ojeb.000042
Kells J. J., Meggitt W. F., Penner D. (1984). Absorption, translocation, and activity of fluazifop-butyl as influenced by plant growth stage and environment. Weed Sci. 32, 143–149. doi: 10.1017/S0043174500058689
Khachatryan H., Hodges A., Hall C., Palma M. (2020). Production and marketing practices and trade flows in the United States green industr. South. Cooperative Ser. Bull. 421, 2020–2021.
Khamare Y., Marble S. C., Chandler A. (2020). Fertilizer placement effects on eclipta (Eclipta prostrata) growth and competition with container-grown ornamentals. Weed Sci. 68, 496–502. doi: 10.1017/wsc.2020.43
Khamare Y., Marble S. C., Pearson B. J., Chen J., Devkota P. (2023). Effect of weed competition on growth of container-grown ornamental plants in four different container sizes. Horticulturae 9, 317. doi: 10.3390/horticulturae9030317
Knezevic S. Z. (2014). “Integrated weed management in soybean,” in Recent advances in weed management. Eds. Chauhan B. S., Mahajan G. (Springer, Switzerland: Springer), 223–237.
Knowe S. A., Stein W. I. (1995). Predicting the effects of site preparation and protection on the development of young Douglas-fir plantations. Can. J. For. Res. 25, 1538–1547. doi: 10.1139/x95-167
Knudson W. (2018). The Economic Impact of Michigan’s Food and Agriculture System (East Lansing, MI, USA: Michigan State University Product Center).
Koger C. H., Dodds D. M., Reynolds D. B. (2007). Effect of adjuvants and urea ammonium nitrate on bispyribac efficacy, absorption, and translocation in barnyardgrass (Echinochloa crus-galli). I. Efficacy, rainfastness, and soil moisture. Weed Sci. 55, 399–405. doi: 10.1614/WS-06-146.1
Kriticos D. J., Crossman N. D., Ota N., Scott J. K. (2010). Climate Change and Invasive Plants in South Australia (Canberra, Australia: National Research Flagship Climatic Adaptation).
Kumar R., Kumar K. K. (2007). Managing physiological disorders in litchi. Indian Horticulture 52, 22–24.
Kumar V., Obour A., Jha P., Liu R., Manuchehri M. R., Dille J. A., et al. (2020). Integrating cover crops for weed management in the semi-arid U.S. Great Plains: Opportunities and challenges. Weed Sci. 68, 311–323. doi: 10.1017/wsc.2020.26
Li Y., Ye W., Wang M., Yan X. (2009). Climate change and drought: A risk assessment of crop-yield impacts. Climate Res. 39, 31–46. doi: 10.3354/cr00797
Loss S. R., Terwilliger L. A., Peterson A. C. (2011). Assisted colonization: integrating conservation strategies in the face of climate change. Biol. Conserv. 144, 92–100. doi: 10.1016/j.biocon.2010.11.016
Luo Y., Mooney H. A. (1999). Carbon dioxide and environmental stress (San Diego, CA, USA: Academic Press).
MacDonald M. T. (2010). Physiological significance of ethylene in needle abscission of root-detached balsam fir (Abies balsamea (L.) Mill.). Québec, Canada: Université Laval. Doctoral dissertation.
MacDonald M. T., Lada R. R., Veitch R. S., Thiagarajan A., Adams A. D. (2014). Postharvest needle abscission resistance of balsam fir (Abies balsamea L.) is modified by harvest date. Can. J. For. Res. 44, 1394–1401. doi: 10.1139/cjfr-2014-0199
Madafiglio G. P., Medd R. W., Cornish P. S., Van de Ven R. (2000). Temperature mediated responses of flumetsulam and metosulam on Raphanus raphanistrum. Weed Res. 40, 387–395. doi: 10.1046/j.1365-3180.2000.00200.x
Madan P., Jagadish S. V. K., Craufurd P. Q., Fitzgerald M., Lafarge T., Wheeler T. R. (2012). Effect of elevated CO2 and high temperature on seed-set and grain quality of rice. J. Exp. Bot. 63, 3843–3852. doi: 10.1093/jxb/ers077
Mahan J. R., Dotray P. A., Light G. G. (2004). Thermal dependence of enzyme function and inhibition; implications for herbicide efficacy and tolerance. Physiologia Plantarum 20, 187–195. doi: 10.1111/j.0031-9317.2004.0255.x
Mahato A. (2014). Climate change and its impact on agriculture. Int. J. Sci. Res. Publications 4, 1–6.
Malarkodi N., Manikandan N., Ramaraj A. P. (2017). Impact of climate change on weeds and weed management—A review. J. Innovative Agric. 4, 1–7.
Malhotra S. K. (2015). Hydroponic in horticulture, an overview. Soilless Gardening India Magazine 11, 6–8.
Malhotra S. K., Srivastva A. K. (2014). “Climate smart horticulture for addressing food, nutritional security and climate challenges,” in Shodh Chintan: Scientific Articles. Ed. Srivastava A. K. (New Delhi, India: ASM Foundation), 83–97. doi: 10.56093/ijas.v87i1.67138
Manea A., Leishman M. R., Downey P. O. (2011). Exotic C4 grasses have increased tolerance to glyphosate under elevated carbon dioxide. Weed Sci. 59, 28–36. doi: 10.1614/WS-D-10-00080.1
Manisankar G., Ramesh T. (2019). Response of weeds under elevated CO2 and temperature: A review. J. Pharmacognosy Phytochem. SP2, 427–431.
Marble S. C., Prior S. A., Runion G. B., Torbert H. A. (2015). Control of yellow and purple nutsedge in elevated CO2 environments with glyphosate and halosulfuron. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00001
Mathers H. M. (2003). Novel methods of weed control in containers. HortTechnology 13, 28–34. doi: 10.21273/HORTTECH.13.1.0028
Medd R. W., Van de Ven R., Pickering D. I., Nordblom T. (2001). Determination of environment specific dose response relationships for clodinafop-propargyl on Avena spp. Weed Res. 41, 351–368. doi: 10.1046/j.1365-3180.2001.00243.x
Mekki M. (2007). Biology, distribution and impacts of silverleaf nightshade (Solanum elaeagnifolium Cav.). Bull. OEPP/EPPO Bull. 37, 114–118. doi: 10.1111/j.1365-2338.2007.01094.x
Mendelsohn R. (2009). The impact of climate change on agriculture in developing countries. J. Natural Resour. Policy Res. 1, 5–19. doi: 10.1080/19390450802495882
Mervosh T. L., Ahrens J. F (1998). Preemergence herbicides for container-grown perennials. Proceedings of the Annual Meeting of the Northeastern Weed Science Society. USA: Northeastern Weed Science Society (Vol. 52, pp. 131–131).
Metz B., Davidson O., Bosch P., Dave R., Mayer L. A. (2007). “Climate change,” in Synthesis Report: Summary for Policymakers (IPCC, Geneva, Switzerland).
Michigan Department of A., Rural D. (2019). Michigan agriculture facts & Figures. (Lansing, MI, USA: Michigan Department of Agriculture and Rural Development (MDARD))
Milakovic I., Fiedler K., Karrer G. (2014). Management of roadside populations of invasive Ambrosia artemisiifolia by mowing. Weed Res. 54, 256–264. doi: 10.1111/wre.12074
Mitcham-Butler E. J., Hinesley L. E., Pharr D. M. (1988). Effect of harvest date, storage temperature, and moisture status on postharvest needle retention of Fraser fir. J. Environ. Horticulture 6, 1–4. doi: 10.24266/0738-2898-6.1.1
Muzik T. J. (1976). “Influence of environmental factors on toxicity to plants,” in Herbicides: Physiology, Biochemistry, Ecology. Ed. Audus L. J. (Academic Press, New York), 203–247.
Myers S. S., Zanobetti A., Kloog I., Huybers P., Leakey A. D. B., Bloom A. J., et al. (2014). Increasing CO2 threatens human nutrition. Nature 510, 139–142. doi: 10.1038/nature13179
Naidu V. S. (2013). Invasive potential of C3-C4 intermediate Parthenium hysterophorus under elevated CO2. Indian J. Agric. Sci. 83, 176–179.
Naidu V. S. G. R., Paroha S. (2008). Growth and biomass partitioning in two weed species, Parthenium hysterophorus (C3) and Amaranthus viridis (C4) under elevated CO2. Ecology Environ. Conserv. 14, 9–12.
Narayan R. (2009). “Air pollution a threat in vegetable productions,” in International Conference on Horticulture: Horticulture for Livelihood Security and Economic Growth. Eds. Sulladmath U. V., Swamy K. R. M. (Bangalore, India: PNASF (Prem Nath Agricultural Science Foundation)), 158–159.
National Christmas Tree, A (2021). Quick tree facts. (Chesterfield, MO, USA: National Christmas Tree Association).
NeSmith D. S., Lindstrom O. M. (1996). Vegetation management of Leyland cypress grown for Christmas trees. J. Environ. Horticulture 14, 42–43. doi: 10.24266/0738-2898-14.1.42
Norcini J. G., Garber M. P., Hudson W. G., Jones R. K., Chase A. R., Bondari K. (1996). Pest management in the United States greenhouse and nursery industry: IV. Weed control. HortTechnology 6, 211–216. doi: 10.21273/HORTTECH.6.3.211
Norcini J. G., Stamps R. H. (1994). Container nursery weed control (University of Florida: Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences).
Nowak R. S., Ellsworth D. S., Smith S. D. (2004). Functional responses of plants to elevated atmospheric CO2: Do photosynthetic and productivity data from FACE experiments support early predictions? New Phytol. 162, 253–280. doi: 10.1111/j.1469-8137.2004.01033.x
O’Donnell C. C., Adkins S. W. (2001). Wild oat and climate change: The effect of CO2 concentration, temperature, and water deficit on the growth and development of wild oat in monoculture. Weed Sci. 49, 694–702. doi: 10.1614/0043-1745(2001)049[0694:WOACCT]2.0.CO;2
O’Donovan J. T. (2005). Weed economic thresholds: Useful agronomic tool or pipe dream? Phytoprotection 77, 13–28. doi: 10.7202/706205ar
Oregon State University E. (1981). Weed control in Christmas trees (Corvallis, OR, USA; Oregon State University Extension Service).
Ott J. P., Klimešová J., Hartnett D. C. (2019). The ecology and significance of below-ground bud banks in plants. Ann. Bot. 123, 1099–1118. doi: 10.1093/aob/mcz051
Ou J., Stahlman P. W., Jugulam M. (2018). Reduced absorption of glyphosate and decreased translocation of dicamba contribute to poor control of kochia (Kochia scoparia) at high temperature. Pest Manage. Sci. 74, 1134–1142. doi: 10.1002/ps.4840
Pagare S. A., Bhatia M. A., Tripathi N., Pagare S., Kumar B. H. (2017). Weeds: An introduction and their responses to climate change. Trends Biosci. 10, 2123–2129.
Parry M. L. (1998). The impact of climate change on European agriculture The Bawden Memorial Lectures 1973-1998, Silver Jubilee Edition (Surrey, U.K: British Crop Protection Council).
Patterson D. T. (1995). Weeds in a changing climate. Weed Sci. 43, 685–701. doi: 10.1017/S0043174500081832
Patterson D. T., Westbrook J. K., Joyce R. J. V., Lingren P. D., Rogasik J. (1999). Weeds, insects, and diseases. Climate Change 43, 711–727. doi: 10.1023/A:1005549400875
Pautasso M., Dehnen-Schmutz K., Holdenrieder O. (2010). Plant health and global change—some implications for landscape management. Biol. Rev. 85, 729–755. doi: 10.1111/j.1469-185X.2010.00123.x
Peters K., Gerowitt B. (2014). Important maize weeds profit in growth and reproduction from climate change conditions represented by higher temperatures and reduced humidity. J. Appl. Bot. Food Qual. 87, 234–242. doi: 10.5073/JABFQ.2014.087.033
Petit J. R., Jouzel J., Raynaud D. (1999). Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 399, 429–436. doi: 10.1038/20859
Poorter H. (1993). Interspecific variation in the growth response of plants to an elevated ambient CO2 concentration CO2 and biosphere. (Dordrecht, Netherlands: Springer Nature)
Porter W. C. (1996). Isoxaben and isoxaben combinations for weed control in container-grown herbaceous flowering perennials. J. Environ. Horticulture 14, 27–30. doi: 10.24266/0738-2898-14.1.27
Price C. E. (1983). “The effect of environment on foliage uptake and translocation of herbicides,” in Aspects of Applied Biology, 4, Influence of Environmental Factors on Herbicide Performance and Crop and Weed Biology (Warwick, UK: The Association of Applied Biologists, Warwick), 157–169. ed. A.O.A. Biologists.
Queiroz J. R. G., Silva A. C. Jr., Martins D. (2016). Herbicide selectivity in tropical ornamental species. Planta Daninha 34, 795–802. doi: 10.1590/s0100-83582016340400020
Ramsey R. J. L., Stephenson G. R., Hall J. C. (2005). A review of the effects of humidity, humectants, and surfactant composition on the absorption and efficacy of highly water-soluble herbicides. Pesticide Biochem. Physiol. 82, 162–175. doi: 10.1016/j.pestbp.2005.02.005
Reeves J. L. (2017). Climate change effects on biological control of invasive plants by insects. CAB Rev. 12, 1–8. doi: 10.1079/PAVSNNR201712001
Refatti J. P., Avila L. A., Camargo E. R., Ziska L. H., Oliveira C., Salas-Perez R., et al. (2019). High [CO2] and temperature increase resistance to cyhalofop-butyl in multiple-resistant Echinochloa colona. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00529
Richardson R. J., Zandstra B. H. (2006). Evaluation of flumioxazin and other herbicides for weed control in gladiolus. Weed Technol. 20, 394–398. doi: 10.1614/WT-05-031R2.1
Roberts S. D., Long J. N. (1992). Production efficiency in Abies lasiocarpa: Influence of vertical distribution of leaf area. Can. J. For. Res. 22, 1230–1234. doi: 10.1139/x92-164
Rodenburg J., Meinke H., Johnson D. E. (2011). Challenges for weed management in African rice systems in a changing climate. J. Agric. Sci. 149, 427–435. doi: 10.1017/S0021859611000207
Rodenburg J., Riches C. R., Kayeke J. M. (2010). Addressing current and future problems of parasitic weeds in rice. Crop Prot. 29, 210–221. doi: 10.1016/j.cropro.2009.10.015
Rogers H. H., Dahlman R. C. (1993). Crop responses to CO2 enrichment. Vegetatio. 14, 117–131. doi: 10.1007/BF00048148
Rogers H. H., Runion G. B., Krupa S. V. (1994). Plant responses to atmospheric CO2 enrichment, with emphasis on roots and the rhizosphere. Environ. pollut. 83, 155–189. doi: 10.1016/0269-7491(94)90034-5
Rosenzweig C., Iglesius A., Yang X. B., Epstein P. R., Chivian E. (2001). Climate change and extreme weather events—Implications for food production, plant diseases, and pests. Global Change Hum. Health 2, 90–104. doi: 10.1023/A:1015086831467
Rosenzweig C., Liverman D. (1992). “Predicted effects of climate change on agriculture: A comparison of temperate and tropical regions,” in Global Climate Change: Implications, Challenges, and Mitigation Measures. Ed. Majumdar S. K. (Easton, PA, USA: Pennsylvania Academy of Science), 342–361.
Sasek T. W., Strain B. R. (1990). Implications of atmospheric CO2 enrichment and climatic change for the geographical distribution of two introduced vines in the USA. Climate Change 16, 31–51. doi: 10.1007/BF00137345
Schmidt J. J., Blankenship E. E., Lindquist J. L. (2011). Corn and velvetleaf (Abutilon theophrasti) transpiration in response to drying soil. Weed Sci. 59, 50–54. doi: 10.1614/WS-D-10-00078.1
Schneider W. G., Knowe S. A., Harrington T. B. (1998). Predicting survival of planted Douglas-fir and ponderosa pine seedlings on dry, low elevation sites in southwestern Oregon. New Forests 15, 139–159. doi: 10.1023/A:1006549427671
Seixas A. L., Alves P. L. C. A., Carvalho L. B. (2011). Pendimethalin aplicado à casca de arroz e serragem para o controle de plantas daninhas em Ixora chinensis Lam. Ciec. e Agrotecnologia 35, 718–727. doi: 10.1590/S1413-70542011000400010
Sharma S. D., Singh M. (2001). Environmental factors affecting absorption and bio-efficacy of glyphosate in Florida beggar weed (Desmodium tortuosum). Crop Prot. 20, 511–516. doi: 10.1016/S0261-2194(01)00065-5
Shrestha S. (2019). Effects of climate change in agricultural insect pest. Acta Scientiarum Agriculturae 3, 74–80. doi: 10.31080/ASAG.2019.03.0727
Singh M., Bhullar M. S., Chauhan B. S. (2015). Seed bank dynamics and emergence pattern of weeds as affected by tillage systems in dry direct-seeded rice. Crop Prot. 67, 168–177. doi: 10.1016/j.cropro.2014.10.015
Singh M. C., Dubey S. C., Yaduraju N. T. (2016). Climate change and its possible impacts on weeds. Int. J. Environ. Sci. Technol. 5, 1530–1539.
Singh R. P., Singh R. K., Singh M. K. (2011). Impact of climate and carbon dioxide change on weeds and their management—a review. Indian J. Weed Sci. 43, 1–11.
Smith R. J. Jr. (1968). Weed competition in rice. Weed Sci. 16, 252–255. doi: 10.1017/S0043174500031461
Smith T. (2019). Managing weeds in and around the greenhouse. (Amherst, MA, USA: University of Massachusetts Amherst Extension).
Smith R. G., Warren N. D., Cordeau S. (2020). Are cover crop mixtures better at suppressing weeds than cover crop monocultures? Weed Sci. 68, 186–194. doi: 10.1017/wsc.2019.69
Sosnoskie L. M., Hanson B. D. (2016). Field bindweed (Convolvulus arvensis) control in early and late-planted processing tomatoes. Weed Technol. 30, 708–716. doi: 10.1614/WT-D-15-00160.1
Srinivasa Rao N. K., Laxman R. H., Bhatt R. M. (2010). “Impacts of climate change on vegetable crops,” in Challenges of Climate Change in INDIAn Horticulture. Eds. Singh H. P., Singh J. P., Lal S. S. (New Delhi, India: Westville Publications House), 113–123.
Staats D., Klett J. E. (1993). Evaluation of weed control and phytotoxicity of preemergence herbicides applied to container-grown herbaceous and woody plants. J. Environ. Horticulture 11, 78–80. doi: 10.24266/0738-2898-11.2.78
Swanton C. J., Weaver S., Cowan P., Van Acker R., Deen W., Shreshta A. (2020). Weed thresholds: Theory and applicability. Expanded Context Weed Manage. 2, 9–29.
Talbert R. E., Schmidt L. A., Burgos N. R., Johnson J. A., Curless J. K., Norsworthy J. K. (1999). “Field evaluation of herbicides on small fruit, vegetable, and ornamental crops,” in Arkansas Agricultural Experiment Station, Division of Agriculture, vol. 461. (Arkansas Agricultural Experiment Station: University of Arkansas). Research series.
Thiagarajan A. (2012). Physiology of low temperature-modulated postharvest needle senescence and abscission in balsam fir (Abies balsamea L.) (Quebec City, QC, Canada: Université Laval).
Tubiello F. N., Soussana J. F., Howden S. M. (2007). Crop and pasture response to climate change. Proc. Natl. Acad. Sci. United States America 104, 19686–19690. doi: 10.1073/pnas.0701728104
Tungate K. D., Israel D. W., Watson D. M., Rufty T. W. (2007). Potential changes in weed competitiveness in an agroecological system with elevated temperatures. Environ. Exp. Bot. 60, 42–49. doi: 10.1016/j.envexpbot.2006.06.001
Uddling J., Broberg M. C., Feng Z., Pleijel H. (2018). Crop quality under rising atmospheric CO2. Curr. Opin. Plant Biol. 45, 262–267. doi: 10.1016/j.pbi.2018.06.001
Usda, National Agricultural Statistics, S (2021). Floriculture crops 2020 summary. (Washington, D.C., USA: U.S. Department of Agriculture (USDA), National Agricultural Statistics Service (NASS)).
Varanasi A., Prasad P. V. V., Jugulam M. (2015). Impact of climate change factors on weeds and herbicide efficacy. Adv. Agron. 135, 107–146. doi: 10.1016/bs.agron.2015.09.002
Volis S. (2009). Plasticity, its cost, and phenotypic selection under water and nutrient stress in two annual grasses. Biol. J. Linn. Soc. 97, 581–593. doi: 10.1111/j.1095-8312.2009.01239.x
Volis S., Verhoeven K. J. F., Mendlinger S., Ward D. (2004). Phenotypic selection and regulation of reproduction in different environments in wild barley. J. Evolutionary Biol. 17, 1121–1131. doi: 10.1111/j.1420-9101.2004.00738.x
Von der Lippe M., Bullock J. M., Kowarik I. (2013). Human-mediated dispersal of seeds by the airflow of vehicles. PloS One 8, e52733. doi: 10.1371/journal.pone.0052733
Walck J. L., Hidayati S. N., Dixon K. W., Thompson K., Poschlod P. (2011). Climate change and plant regeneration from seed. Global Change Biol. 17, 2145–2161. doi: 10.1111/j.1365-2486.2010.02368.x
Walker K. L., Williams D. J. (1988). Grass interference in container-grown Bailey’s redosier dogwood (Cornus × Baileyi). Weed Sci. 36, 621–624. doi: 10.1017/S0043174500089196
Walker K. L., Williams D. J. (1989). Annual grass interference in container-grown bush cinquefoil (Potentilla fruticosa). Weed Sci. 37, 73–75. doi: 10.1017/S0043174500055880
Wang R. L., Dekker J. (1995). Weedy adaptation in Setaria spp.: III. Variation in herbicide resistance in Setaria spp. Pesticide Biochem. Physiol. 51, 99–116. doi: 10.1006/pest.1995.1011
Ward J. K., Tissue D. T., Thomas R. B., Strain B. R. (1999). Comparative responses of model C3 and C4 plants to drought in low and elevated CO2. Global Change Biol. 5, 857–867. doi: 10.1046/j.1365-2486.1999.00270.x
Warwick S. I., Black L. D. (1986). Genecological variation in recently established populations of Abutilon theophrasti (velvetleaf). Can. J. Bot. 64, 1632–1643. doi: 10.1139/b86-219
Whitlock C. W., Maxwell B., Silverman N., Wade A. A. (2017). Montana climate assessment (Bozeman and Missoula, MT, USA: Montana State University and University of Montana).
Willoughby I., Palmer C. (1997). Weed control in Christmas tree plantations (London, UK: The Stationery Office).
Yadav L. P., Bose T. K. (1987). Chemical weed control in tuberose and gladiolus. Acta Hortic. 205, 177–185. doi: 10.17660/ActaHortic.1987.205.26
Zandstra B., O’Donnell J. (2018). Weed control in Christmas trees (E3237, pp. 1–12) (East Lansing, MI, USA: Michigan State University Extension Bulletin).
Ziska L. H. (2002). Influence of rising atmospheric CO2 since 1900 on early growth and photosynthetic response of a noxious invasive weed, Canada thistle (Cirsium arvense). Funct. Plant Biol. 29, 1387–1392. doi: 10.1071/FP02052
Ziska L. H. (2003). Evaluation of yield loss in field sorghum from a C3 and C4 weed with increasing CO2. Weed Sci. 51, 914–918. doi: 10.1614/WS-03-002R
Ziska L. H. (2010). Elevated carbon dioxide alters chemical management of Canada thistle in no-till soybean. Field Crop Res. 119, 299–303. doi: 10.1016/j.fcr.2010.07.002
Ziska L. H., Blumenthal D. M., Franks S. J. (2019). Understanding the nexus of rising CO2, climate change, and evolution in weed biology. Invasive Plant Sci. Manage. 12, 79–88. doi: 10.1017/inp.2019.12
Ziska L. H., Blumenthal D. M., Runion G. B., Hunt E. R. Jr., Diaz-Soltero H. (2011). Invasive species and climate change: an agronomic perspective. Climatic Change 105, 13–42. doi: 10.1371/journal.pone.0081510
Ziska L. H., Bunce J. A. (1993). The influence of elevated CO2 and temperature on seed germination and emergence from soil. Field Crops Res. 34, 147–157. doi: 10.1016/0378-4290(93)90003-6
Ziska L. H., Caulfield F. A. (2000). Rising CO2 and pollen production of common ragweed (Ambrosia artemisiifolia), a known allergy inducing species: implications for public health. Aust. J. Plant Physiol. 27, 893–898. doi: 10.1071/PP00032
Ziska L. H., Emche S. D., Johnson E. L., George K., Reed D. R., Sicher R. C. (2005). Alterations in the production and concentration of selected alkaloids as a function of rising atmospheric carbon dioxide and air temperature: Implications for ethnopharmacology. Global Change Biol. 11, 1798–1807. doi: 10.1111/j.1365-2486.2005.001029.x
Ziska L. H., Faulkner S. S., Lydon J. (2004). Changes in biomass and root: Shoot ratio of field-grown Canada thistle (Cirsium arvense), a noxious, invasive weed, with elevated CO2: Implications for control with glyphosate. Weed Sci. 52, 584–588. doi: 10.1614/WS-03-161R
Ziska L. H., Gealy D. R., Tomecek M. B., Jackson A. K., Black H. L. (2012). Recent and projected increases in atmospheric CO2 concentration can enhance gene flow between wild and genetically altered rice (Oryza sativa). PloS One 7, e37522. doi: 10.1371/journal.pone.0037522
Ziska L. H., George K. (2004). Rising carbon dioxide and invasive, noxious plants: potential threats and consequences. World Resource Rev. 16, 427–447.
Ziska L. H., Reunion G. B. (2007). “Future weed, pest and disease problems for plants,” in Agroecosystems in a changing climate. Eds. Newton P. C. D., Carran A., Edwards G. R., Niklaus P. A. (CRC, Boston), 262–279.
Ziska L. H., Runion G. B. (2006). “Future weed, pest, and disease problems for plants,” in Agroecosystems in a Changing Climate. Eds. Newton P., Carman A., Edwards G., Niklaus P. (Boca Raton, FL, USA: CRC Press), 262–287.
Ziska L. H., Teasdale J. R. (2000). Sustained growth and increased tolerance to glyphosate observed in a C3 perennial weed, quackgrass (Elytrigia repens), grown at elevated carbon dioxide. Aust. J. Plant Physiol. 27, 159–164. doi: 10.1071/PP99099
Keywords: elevated temperature, increased carbon dioxide, drought, ornamental crops, weed control, herbicide efficacy
Citation: Mou SS, Haus MJ, Hayden ZD, Patterson EL and Saha D (2025) Climate-driven challenges in weed management for ornamental crop production in the United States: a review. Front. Agron. 7:1556418. doi: 10.3389/fagro.2025.1556418
Received: 06 January 2025; Accepted: 14 February 2025;
Published: 04 March 2025.
Edited by:
Simerjeet Kaur, Punjab Agricultural University, IndiaReviewed by:
Stephen Christopher Marble, University of Florida, United StatesCopyright © 2025 Mou, Haus, Hayden, Patterson and Saha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Debalina Saha, c2FoYWRlYjJAbXN1LmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.