 Julian Winkler1*
Julian Winkler1* Stephan M. Junge2†
Stephan M. Junge2† Abozar Nasirahmadi1†Oliver Hensel1
Abozar Nasirahmadi1†Oliver Hensel1 Maria R. Finckh2
Maria R. Finckh2 Sascha M. Kirchner1
Sascha M. Kirchner1- 1Department of Agricultural and Biosystems Engineering, University of Kassel, Witzenhausen, Germany
- 2Department of Organic Plant Protection, University of Kassel, Witzenhausen, Germany
Introduction: The Colorado potato beetle (Leptinotarsa decemlineata) (CPB) is a major threat to potato crops in Europe, Asia, and America. In Europe, it is expected that climate change will cause the species to expand its range northwards and allow for an additional generation per year in regions where it is already established. The effectiveness of insecticides, a common method of control, is decreasing due to the emergence of resistance to several substances. Therefore, research into alternative measures is becoming increasingly necessary. One preventive measure that has already been shown to have great effects in past studies is the use of organic mulch. For organic mulching, conserved or freshly cut plant material is spread on the field usually when the potato emerges. Previous research has mainly used cereal straw for this purpose in plots that are relatively small compared to actual agricultural fields.
Materials and methods: To address the lack of large-scale field trials in the literature, a series of on-station and on-farm field trials were conducted in this study. Organic, legume-rich mulches were used primarily, which so far have not been extensively investigated. Over a period of four years and in a total of six field trials, CPB and larval infestations were assessed.
Results and discussion: Our results show effects on the initial infestation of young plants by adult CPB and on the emergence time of the first two larval stages. By the time of flowering, when the feeding effect of CPB on yield is known to be greatest, L3 and L4 larvae were reduced by about 30% in the on-station trials and by about 40% in the on-farm trials. In addition, when CPB larval stages were used to calculate a weighted daily leaf consumption per stage, total calculated leaf consumption was on average about 40% lower in the mulch treatments across all sampling dates and all trials. The recorded yields were equal or higher in the mulched treatments. In conclusion, organic mulching can effectively reduce CPB damage even on a large scale and can have a positive impact on yield. Further agronomic benefits of mulching, as well as drawbacks and putative mechanisms of action, are discussed.
1 Introduction
Potatoes are an important crop in Europe, contributing 3.1% to total agricultural monetary output and being grown on 1.66 million hectares. Germany accounts for 273 thousand hectares (Eurostat, 2021), of which around 10 thousand hectares are grown organically (BLE, 2023). Potatoes are typically planted in Germany around April and have an average yield of approximately 40 and 20 tons per hectare in the conventional and organic sector, respectively (BLE, 2023). Yields are especially dependent on water availability and nutrients (Koch et al., 2020), but also on pest pressure. Among the pests Phytophthora infestans is considered to be the most economically important (Dupuis et al., 2024). Besides this pest, the Colorado potato beetle Say (CPB) (Leptinotarsa decemlineata, Coleoptera, Chrysomelidae) is a widespread pest in America, Europe, and Asia (EPPO, 2022) causing significant yield losses or even complete crop failure (Hare, 1990; Alyokhin et al., 2022). In Central Europe it usually completes one to two generations per year (Pulatov et al., 2016). Climate change could intensify the challenges by favouring CPB expansion into northern regions (Wang et al., 2017), and by allowing for additional generations within a given season (Pulatov et al., 2016; Junge et al., 2022). CPB overwinter in the soil in or near the potato field where they develop (Weber and Ferro, 1993). After overwintering, most CPB adults only migrate short distances (Voss and Ferro, 1990; Ferro et al., 1991). Egg laying typically occurs in May and early June, depending primarily on temperature (Racca et al., 2014). An effective method to control CPB is to plant potatoes at large distances from the previous year’s fields (Weisz et al., 1994, 1996). However, it is not always feasible for farmers to do so. Currently, CPB is largely controlled with insecticides. As their effectiveness is increasingly limited due to the growing number of resistant populations to several insecticidal active ingredients (Mota-Sanchez and Wise, 2022), new approaches are needed that focus on prevention rather than control.
A promising preventive method is the use of organic mulches, i.e. plant material applied to the soil at the time of potato plant emergence. This approach has been shown to have the potential to reduce CPB in small-scale field trials. The organic mulch material used in previous studies in North America was cereal straw (Zehnder and Hough-Goldstein, 1990; Stoner, 1993; Brust, 1994, 1996; Stoner et al., 1996; Johnson et al., 2004; Genger et al., 2018). Other studies conducted in Europe have tested fresh plant materials such as grass mulch (Dvořák et al., 2012) or rye/pea, triticale/vetch, and grass/clover mulch (Junge et al., 2022). All these studies found a reduction of CPB larvae in some trials. In contrast to straw, legume-rich mulch materials such as those used in Junge et al. (2022) provide an additional fertilizing effect and can contribute to humus formation (Tripolskaja et al., 2014). In organic farming, legume-rich green manures are a typical component of crop rotations and are often available in surplus in stockless operations in Germany. Mulching organic materials is also an element of regenerative agriculture, a European trend in agriculture that focuses on soil health and biodiversity (Schreefel et al., 2020) and has recently been shown to enrich the soil bacterial community (Singh et al., 2023).
So far, the actual mechanisms leading to CPB reduction by mulch are unclear. Hypotheses include increased predation due to 1) higher numbers of predators, i.e. carabids (Brust, 1994) or ladybirds (Szendrei and Weber, 2009), 2) impeded CPB movement due to plant materials on the ground (Szendrei et al., 2009), 3) a less favourable microclimate (Johnson et al., 2004) or 4) a change in the mineral status of the plants to the disadvantage of CPB (Alyokhin and Atlihan, 2005). Difficulties of CPB in finding its host due to a change in colour contrasts caused by organic mulches (5) could also be responsible for the lower numbers of CPB, as it was shown for other phytophagous insects (Saucke and Döring, 2004; Döring and Röhrig, 2016). With these uncertainties in mind, it remains unclear, whether the effects previously found in small-scale trials also apply to agricultural scales.
In order to evaluate other organic mulch materials than straw and to address the lack of larger field trials in the literature, a number of field trials, including on-farm trials, were conducted to answer the questions: “Can the results of previous studies with straw mulched potatoes be confirmed with other mulch materials?” and “Can the effect of organic mulching found in previous studies in small-scale trials be applied to a large scale?”. We concentrated mainly on legume-rich organic mulches, such as triticale/vetch and grass/clover, but also used straw mulch. In addition to assessing the number of the developmental stages of CPB, we have calculated the expected consumption by the beetles based on the fractions of larval stages present at a given time according to their feeding rate to better visualise the effect of mulching.
2 Materials and methods
2.1 On-station trials
Field trials consisted of a total of three on-station trials conducted from 2019 to 2021, and three on-farm trials conducted in 2021 and 2022. On-station trials were conducted at the experimental farm of the University of Kassel in Neu-Eichenberg, Germany (51°22’48”N 9°54’41”E, 223 m ASL). The soil at the University of Kassel’s experimental farm in Neu-Eichenberg is characterized as silt loam (Haplic Luvisol). The trials were set up as randomized complete block designs with four blocks. In 2019, each block comprised the two treatments no mulch (control) and mulch (triticale/vetch) replicated twice, resulting in a total of sixteen plots of 90m2 each. Soil cultivation was carried out in autumn with a plough. Additional information on previous crops and dates of planting, mulching and harvest are given in Table 1. The CPB data from the 2019 trial have been presented in part in Junge et al. (2022), but are analysed here in detail, particularly with regard to the individual larval stages. Both the 2020 and 2021 trials were integrated into a long-term split plot experiment with reduced tillage since 2016. The four blocks in these two trials comprised the three treatments unmulched (control), triticale/vetch mulched, and grass/clover mulched, resulting in twelve plots with a plot size of 60 m2. In these two trials, in contrast to all other trials, a compensatory fertilisation with hair meal pellets corresponding to 100 kg N/ha was applied in the unmulched control. Based on measurements of the nitrogen content of the trticale/vetch mulch material in past trials, approximately this amount of nitrogen was assumed to be contained in the quantity of mulch material applied. In addition to CPB, the trials also investigated aphid abundance and virus transmission. This was not an objective of this study. CPB-specific insecticides were used on the dates shown in Table 1 to prevent excessive impact of CPB damage to other aspects of the trial. The insecticide Novodor FC (Biofa, Münsingen, Germany) contains Bacillus thuringiensis var. tenebrionis (Btt) at a concentration of 1 mil. IU/ha and was commercially available during the relevant time period in Germany and regularly used in organic potato production. In all on-station trials potatoes of the variety Laura were planted at a distance of 75 cm between rows and a distance of 33 cm within rows, which corresponds to four tubers per m2.

Table 1 Details of on-station trials.
2.2 On-farm trials
On-farm trials were conducted on commercial organic potato fields located within a 15 km radius of the experimental farm (Table 2). The soil texture of the corresponding field was sandy loam and loam, respectively. The trials were set up as randomized complete block designs with four replicates in 2021a and 2022 and three replicates in trial 2021b. Each block comprised the three treatments unmulched (control), triticale/vetch mulched, and straw or grass silage (in 2022) mulched. This resulted in twelve or nine (in 2021a) plots, each with an area of 500 m2. The 2021b and 2022 trials were conducted on the same farm, which had chisel ploughed its fields in autumn and planted the variety Belana. The 2021a trial was carried out on a farm that used a plough and planted the variety Laura. Table 2 lists the dates for planting, mulching, Btt application, and harvest, as well as previous crops. The planting distance between rows and within rows was consistent with the on-station trials, at 75 cm and 33 cm respectively.
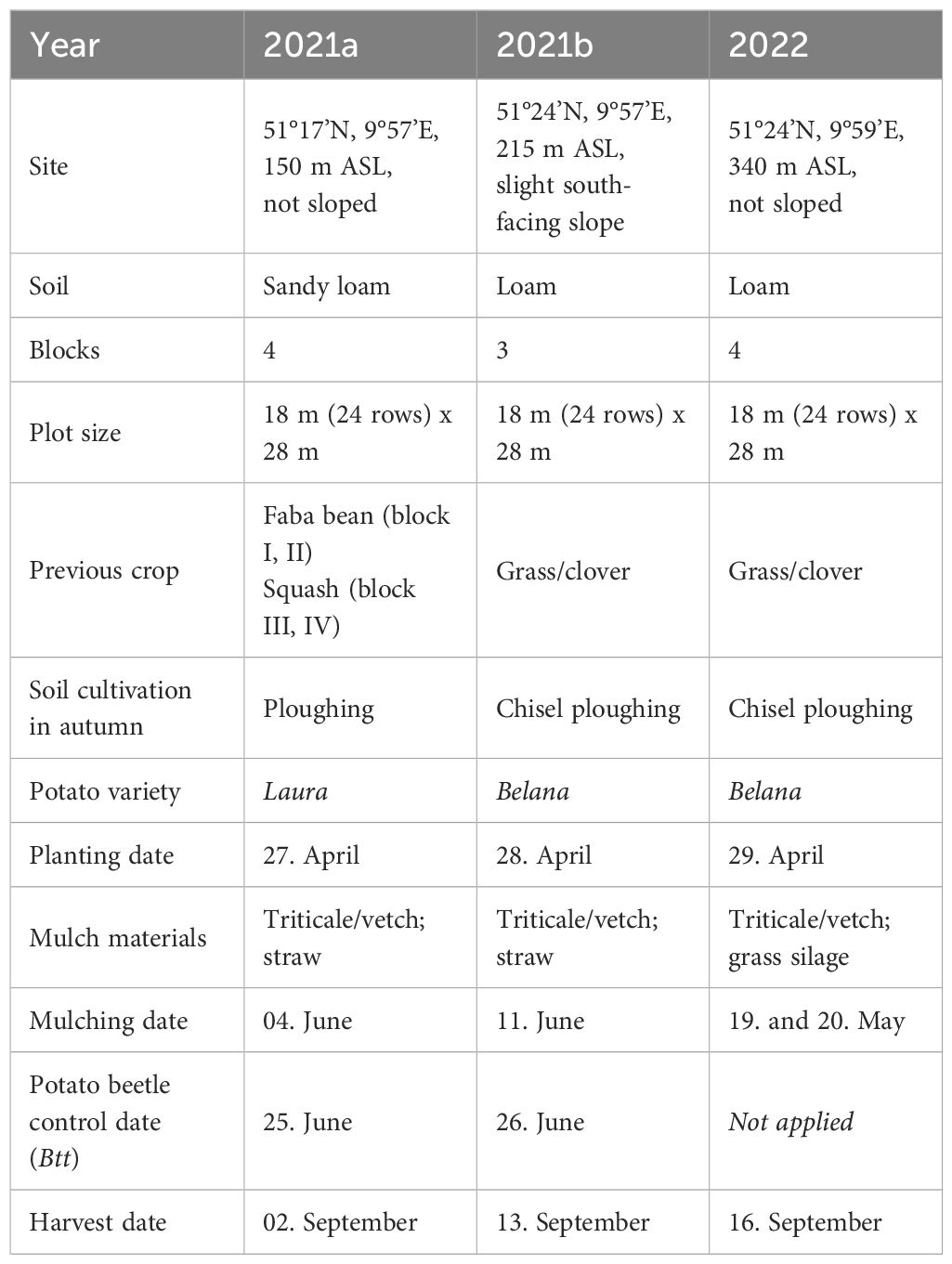
Table 2 Details of on-farm trials.
2.3 Weather conditions
Table 3 shows wheather data during the conduction of the field trials. The trials were carried out under temperate climatic conditions with a long-term (1990–2020) mean temperature of 9.3°C and a mean annual precipitation of 663 mm. No wheather data are available for the farm sites but these were nearby with very similar climatic conditions.
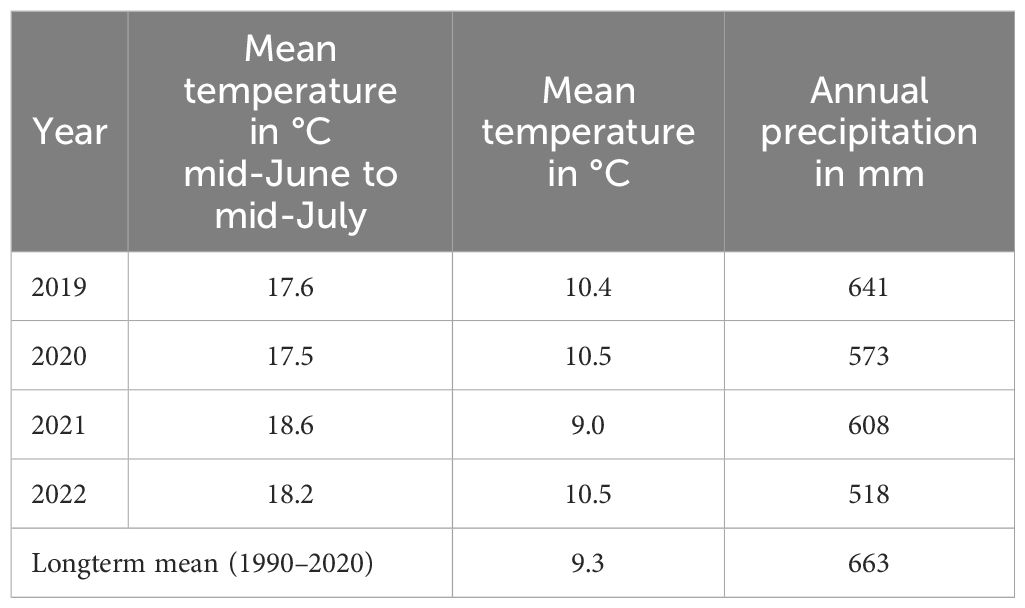
Table 3 Weather data on the experimental farm of the University of Kassel.
2.4 Organic mulch application
Mulch materials were applied around the time of potato emergence, i.e. three to six weeks after planting (see Tables 1 and 2 for exact dates). The application was carried out using an adapted manure spreader with a spreading width of 1.5 m covering two rows of potatoes (Figures 1, 2). The amounts of triticale/vetch and grass/clover used were 50 t/ha in the first year and were increased to 60 t/ha in subsequent trials (Table 4) to achieve a more opaque ground cover, with a particular focus on weed suppression. An exception was on-farm trial 2021a where a reduced amount of only 35 t/ha was tried to test the possibility of a smaller, and therefore cheaper and easier to obtain, quantity. A quantity of 45 tons/ha was used for the grass silage. This amount was regarded as similar in opacity as the material was drier. The amount of straw used (4–6 t/ha) was based on Kirchner et al. (2014) who found that this amount had a significant effect on aphid infestation and virus transmission. Tricitale/vetch and grass/clover mulches were obtained from nearby fields and cut and chopped one day before application with a forage harvester. Straw and grass silage was obtained from one of the farmers and was produced the year before use.

Figure 1 Mulching of on-farm trial 2021b with a manure spreader showing the three treatments triticale/vetch mulch (front left), control (front right) and straw mulch (back left). Photo: Julian Winkler.

Figure 2 On-station trial 2019 at flowering showing the control (left) and triticale/vetch mulch treatment (right). Photo: Stephan M. Junge.
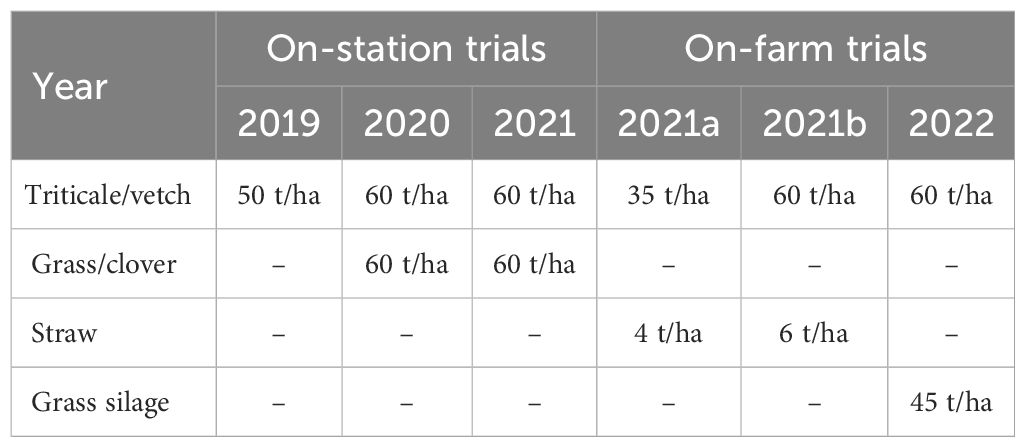
Table 4 Fresh weight of organic mulch applied in field trials.
2.5 Assessment of Colorado potato beetles
The experimental farm and the two commercial farms involved in this study have a history of growing potatoes, resulting in a natural CPB pressure. The distance between the field trials and the potato fields from the previous year was less than 100 m on the experimental farm and a few hundred meters to about 1.5 km on the commercial farms. After mulching, a few plants per plot were checked twice per week for CPB infestation. Actual assessments began in the week the first eggs were observed in these preliminary examinations. In the actual assessment, CPB and larvae, categorized by instar, were counted visually on fifteen whole plants in the on-station trials and eighteen plants per plot in the on-farm trials, three to four times at intervals of 7 to 15 days (exact dates see results). The fifteen plants in the smaller on-station trials were located in the centre row of the plots and did not change between assessments. The eighteen plants in the on-farm trials consisted of six plant triplets, which were randomly distributed over the plot, excluding a 3 m edge, and reselected at each new assessment date. Each selected plant was searched and all leaves were gently turned over to record all individuals and their respective life stages. These comprised eggs (not shown), larval stages 1 to 4 and imagines.
2.6 Yield
Harvesting was done by hand in all experiments. In the on-station trials, two 10 m rows of potatoes with a total area of 15 m2 in the center of each plot were harvested. In the on-farm trials four rows of 3 m, randomly distributed over each plot excluding a 3 m edge, were harvested, resulting in an area of 9 m2 per plot. The potatoes were stored in wooden boxes for two months after harvesting. They were then weighed plot by plot on a scale with a reading accuracy and reproducibility of 1 g. The yield included all potato tubers, except those that were heavily rotted. The yields of all trials were then converted into tons per hectare.
2.7 Data handling, graphing and statistical analysis
All data analyses were carried out in R, version 4.2.1 (R Core Team, 2022). Data processing was done using the tidyverse environment (Wickham et al., 2019) and diagrams were created using the package ggplot2 (Wickham, 2016). The model selection process applied largely followed the guidelines of Zuur and Ieno (2021). Differences in the occurrence of CPB were analyzed trial by trial using generalized linear mixed models with a distribution that fitted the specific set of data. Therefore, the models for each set of data were first fitted with glmmTMB (Brooks et al., 2017) using the Poisson, Negative Binomial and Generalized Poisson distributions. The specific error structure including blocks (all experiments) and plots within blocks (on-station trial 2019) was taken into account in the random structure. If zero-inflation was detected in the Poisson model, a Bernoulli part was added. Zero-inflation tests and all other model validations were performed using the DHARMa package (Hartig, 2022). These included a Kolmogorow-Smirnow test, a dispersion test, an outlier test, a Levene test for homogeneity, and a visual check of a qq-plot (observed vs. expected), all based on DHARMa residuals from 1000 simulations. Models were only selected if they showed no conspicuousness in any of these tests. An additional inclusion of the Akaike information criteria led to the model of choice. As an example, the model formulation selected for the L4-larvae at the last date in the 2021 on-station experiment with a zero-inflated negative binomial distribution was as follows: glmmTMB(L4 ~ treatment + (1 | block), family = “nbinom2”, ziformula = ~ 1, data = on_station_2020_07_13). To avoid data dredging, only L3 and L4 larvae were tested as these consume by far the most leaf tissue (Ferro et al., 1985). In addition, only the last assessment date was tested (the last two dates in the 2021 on-farm trials) as this corresponds to the stage of potato flowering, which is the period of greatest yield effects (Hare, 1980). Where significant differences were found, a Dunnett post-hoc test was performed using the multcomp package (Hothorn et al., 2008). Differences in yield were analyzed using linear mixed models fitted with glmmTMB and a Gaussian distribution. Likewise, model validation was performed using the DHARMa package and the above-mentioned tests with residuals from 1000 simulations and was followed by a Dunnett post-hoc test if applicable.
To calculate total leaf consumption per plant, the number of individuals at each stage was multiplied by the feeding rates determined by Ferro et al. (1985) at 20°C, as this was the temperature investigated by Ferro et al. (1985) that most closely corresponded to the temperatures prevailing in the field trials. The study described the mean values for the completion of developmental stages L1, L2, L3, and L4 at 20°C as 3.7, 3.8, 2.5, and 6.6 days, respectively. The corresponding consumption rates measured at 20°C were 0.9, 1.9, 5.1, and 27.4 cm2 of foliage consumed during the respective stages’ lifespan. This results in daily feeding rates of 0.24, 0.50, 2.04, and 4.15 cm2 of foliage per day. The feeding rate of adult CPB over 10 days was reported to be 50.6, resulting in a daily consumption of 5.06 cm2 foliage per day. The daily consumption values were multiplied by the mean number of individuals counted for each stage, treatment, and date. For instance, on July 10th, 2019, the 0.83 L4 larvae in the control multiplied by the feeding rate of 0.50 cm2 of foliage per day resulted in 0.415 cm2 foliage consumed by L4 larvae. The results were presented in stacked form for each date and each treatment.
3 Results
During the years of the study, CPB first appeared in the trials by about the end of May and began laying eggs by the beginning of June. L1 larvae were not observed before the middle or end of June, depending on the year and location. Only one generation could be observed in all years of the study. The number of larvae per plant generally varied greatly between the years and locations (Figures 3, 4).
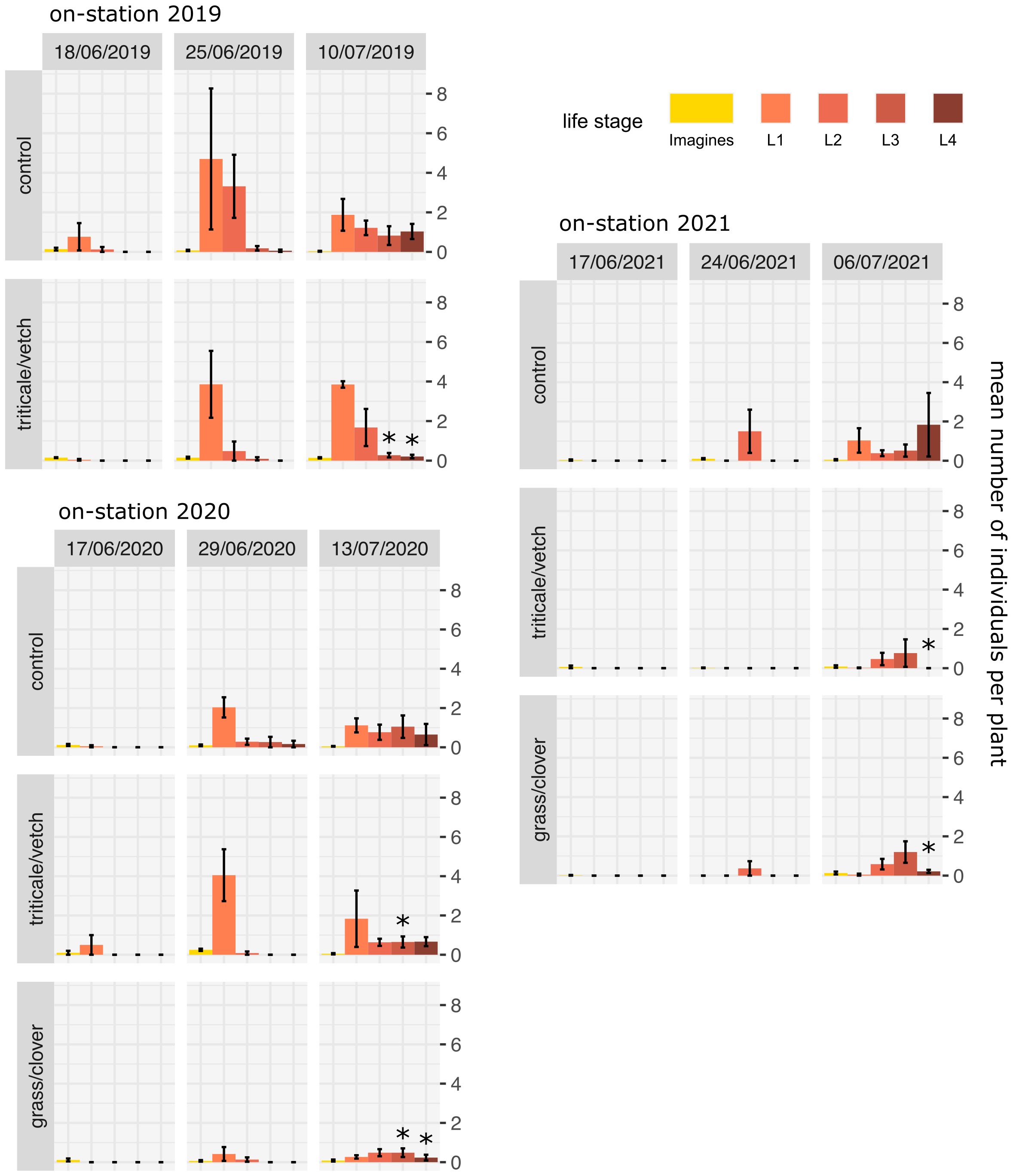
Figure 3 Mean number of CPB individuals per plant in on-station trials (± standard error). Stars indicate significant differences between the control and a specific treatment according to generalized linear mixed models and Dunnett post-hoc tests. * p<0.05.
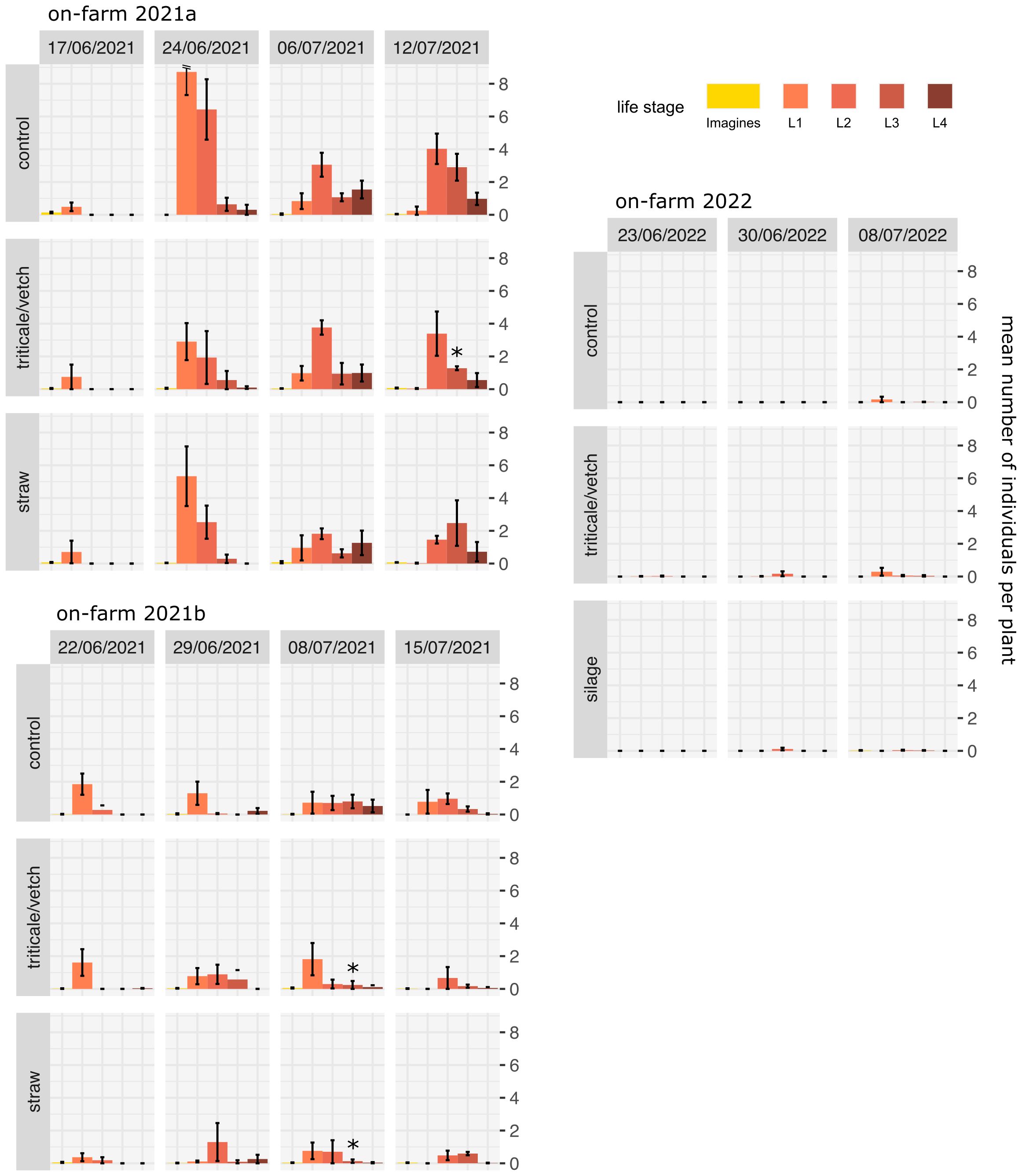
Figure 4 Mean number of CPB individuals per plant in on-farm trials (± standard error). Stars indicate significant differences between the control and a specific treatment according to generalized linear mixed models and Dunnett post-hoc tests. * p<0.05.
When comparing the different mulch treatments with the controls across all trials, a reduction in L1 larvae ranging from 20% in the on-station trial 2019 to >80% in the on-station trial 2021 was observed at the end of June (Figures 3, 4). Exceptions were the on-station trial 2020 (no reduction) and the on-farm trial 2022 (no L1 larvae found). In addition, there was a general trend that larvae of a subsequent larval stages were already present in the control at a given time. Thus, by the end of June, more L2 larvae and, to a lower extent, L3 and L4 larvae were already present in the control treatments and the the number of L1 larvae was correspondingly lower. By beginning to mid July, L3 and L4 larvae were significantly reduced in at least one mulch treatment per trial compared to the unmulched control. Taken together, L3 and L4 were reduced by about 30% on the last date in on-station trials and by about 40% on the last two dates in the 2021 on-farm trials. No reduction was observed in the 2022 trial, where the number of CPB was overall negligible.
Daily leaf consumption, calculated by multiplying the numbers of each life stage by its respective feeding rate, generally increased from mid-June until potato flowering in July (Figure 5). The calculated consumption was lower in the mulch treatments on almost all dates in all trials. In all on-station trials and the on-farm trial 2021a the percentage of reduction increased until the last date, where the reduction was on average about 40%. In on-farm trial 2021b, the reduction only increased to the second to last date. Total leaf consumption was negligible in the 2022 on-farm trial. Despite their low numbers, L3 and L4 larvae account for most of the consumption.
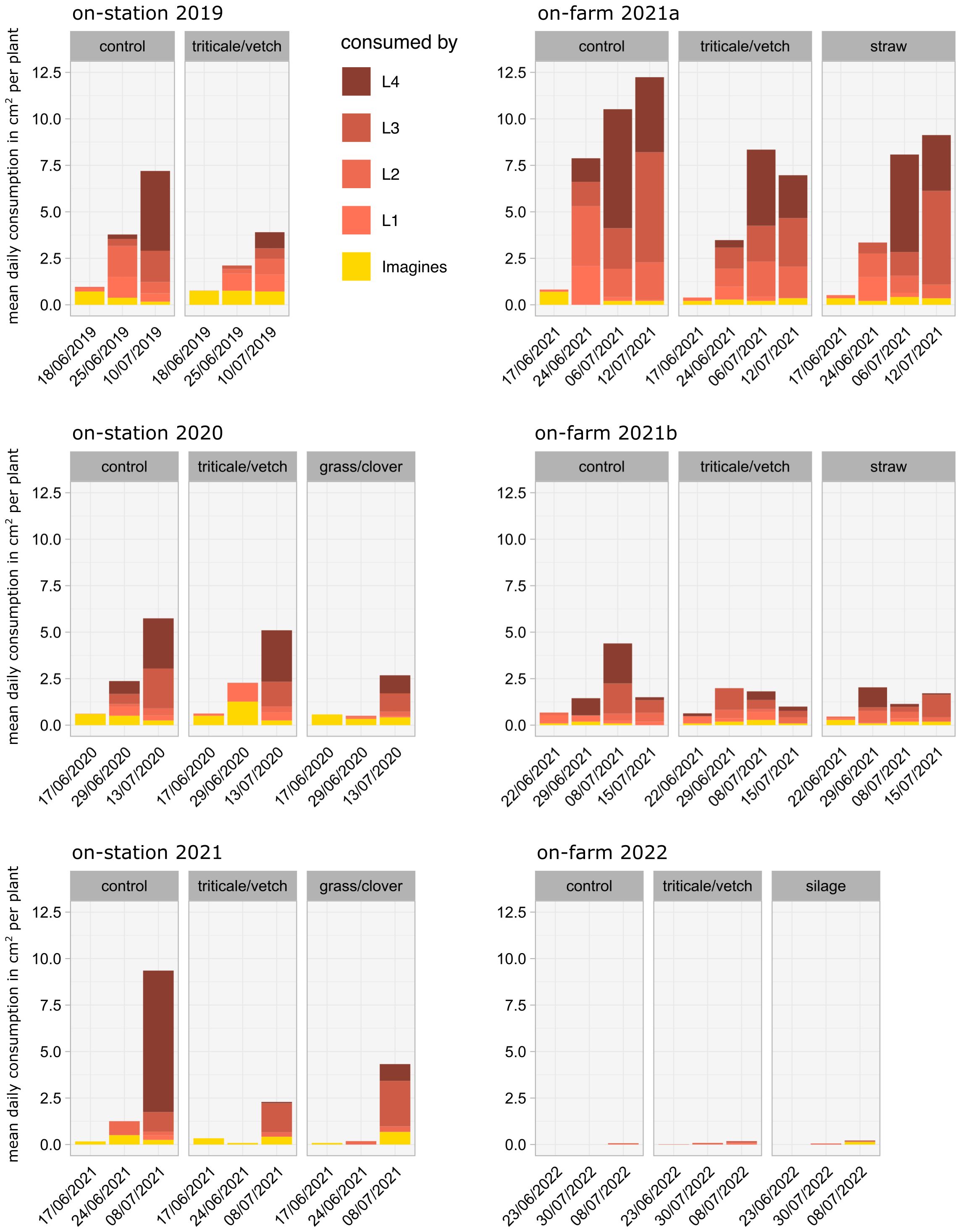
Figure 5 Mean daily leaf tissue consumption in cm2 per plant in on-station and on-farm trials, calculated from the number of individuals multiplied by the specific feeding rates (from Ferro et al., 1985) according to their life stage.
Total yields were mostly between 20 and 30 t/ha with the exception of on-farm trials 2021b and 2022 which yielded <20 t/ha and >30 t/ha, respectively (Figure 6). Yields of treatments with triticale/vetch mulch in on-station trial 2019 (p <0.001), on-farm trial 2021b (p <0.01), and grass silage mulch in on-farm trial 2022 (p <0.05) were significantly higher than in the unmulched controls. All other mulch treatments did not differ significantly from the controls, not even in the on-station trial in 2021 where the controls had received 100 kg/ha additional N fertilizer.
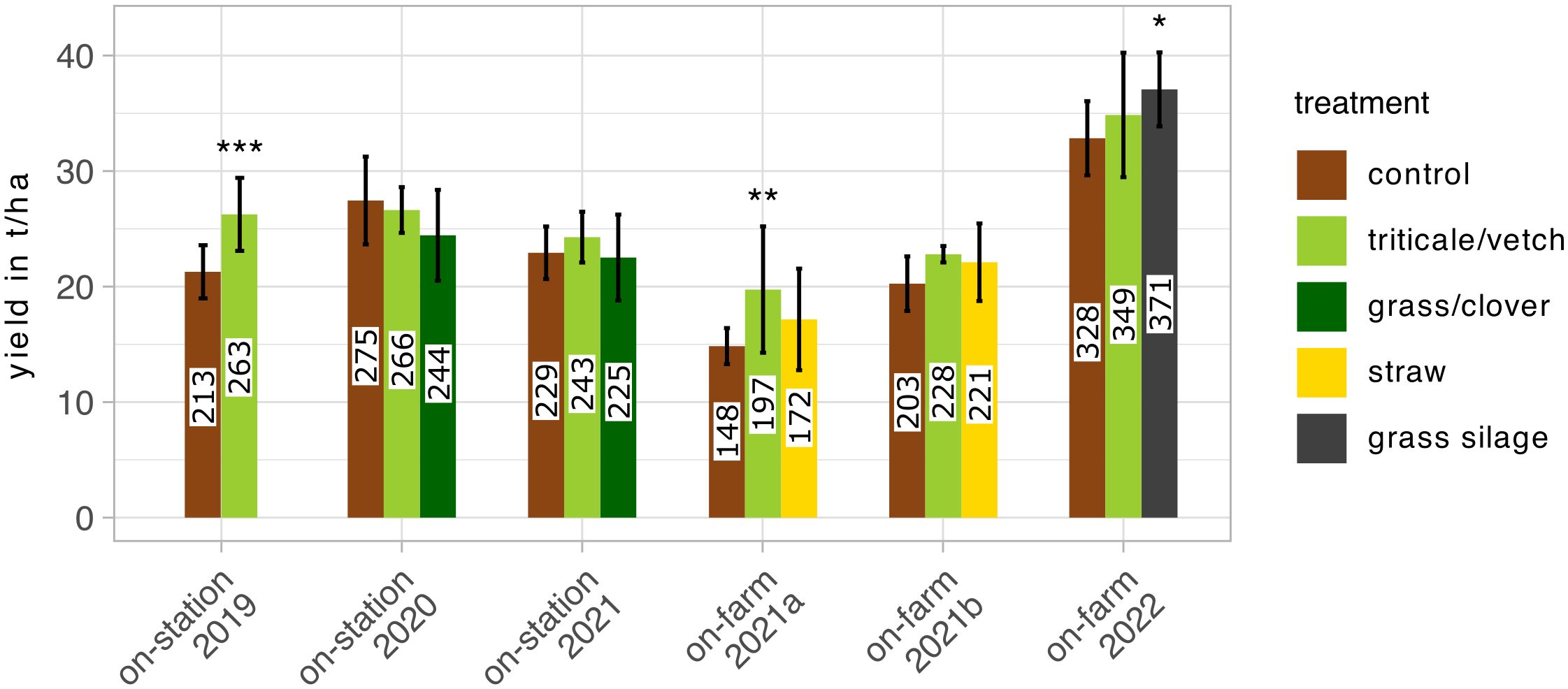
Figure 6 Mean total potato yield in on-station and on-farm trials (± standard deviation). Stars indicate significant differences between the control and a specific treatment according to linear mixed models and Dunnett post-hoc tests. * p<0.05, ** p<0.01, *** p<0.001.
4 Discussion
Natural colonization by CPB was moderate to high in on-station trials and in both the 2021 on-farm trials, but was relatively low in the 2022 trial. Therefore, only in the former trials can a reliable statement be made about the effectiveness of mulching. The low pressure observed in 2022 may have been due to the distance of more than 1 km from the nearest plot of the previous year, which was the 2021b trial plot. In addition to this important factor, infestation is generally influenced by annual variations, especially temperature (Racca et al., 2014). Taken together, however, our results confirm that organic mulching can be effective in reducing CPB infestation where CPB pressure is substantial. This has been shown for several mulch materials and for the first time in considerably larger-scale on-farm trials. The reduction is indicated both by a lower number of larvae of a given stage at a given time and by the fact that the larvae in the mulched treatments were, on average, at preceding larval stages. This results in lower total numbers of larvae early in the season and higher numbers of L3 and L4 larvae later in the season in the mulched plots compared to the unmulched plots. The shift to subesquent larval stages could especially be observed in the triticale/vetch treatment in the on-station trial 2020, where the number of L1 larvae was lower in the control than in the triticale/vetch treatment, but was accompanied by the presence of L2, L3 and L4 larvae, which were not yet present in the mulch treatments at that time.
The effect of mulching was most apparent when the feeding rates and subsequent leaf consumption of each stage were taken into account (Figure 5). On the last and penultimate sampling dates, when the potato plants were in flower and thus the effect of leaf consumption on yield is greatest (Hare, 1980), the L3 and L4 larvae account for a large proportion of the differences in the stacked bars due to their high feeding rate. The effect of mulching, which may be partially obscured in Figures 3 and 4, is emphasised here graphically.
Our results are in line with other studies (Zehnder and Hough-Goldstein, 1990; Stoner, 1993; Brust, 1994, 1996; Stoner et al., 1996; Johnson et al., 2004; Dvořák et al., 2012; Genger et al., 2018; Junge et al., 2022), which also reported a reduction of CPB and/or defoliation activity with the use of straw, vetch, rye, rye/pea, or triticale/vetch. Junge et al. (2022) conducted a study using the same 2019 dataset in a summarized form as this study and reported slightly reduced or even increased total numbers of larvae in mulched plots later in the season; however, their results do not contradict our findings. Our study found a reduction in subsequent larval stages later in the season, but we found partly increased abundances of preceding stages. Of the aforementioned studies, only Brust (1994, 1996) recorded individual larval stages. He also found that the reducing effects occurred mainly at subsequent larval stages. The reduction in potato leaf consumption we calculated is consistent with field assessments made by others who reported changes in defoliation in the field due to mulching, particularly during the potato flowering period (Zehnder and Hough-Goldstein, 1990; Stoner, 1993; Brust, 1994, 1996; Stoner et al., 1996; Junge et al., 2022).
Yields were generally higher in the treatments with mulch albeit often not statistically significant. In previous studies, high CPB reduction was often paired with higher yields. In our study, a comparison of CPB reductions with yields suggests that differences in CPB infestation may also contribute to the higher yields. However, in two of the farm-scale trials, it was observed that CPB infestation was rather low and yields were increased under mulch. This suggests that CPB was not the main factor responsible for the observed yield differences in these two trials. Nonetheless, it is also possible that a change in CPB abundance occurred after the last recording date, as recording was not continued after flowering. Other factors that can affect the yield of the mulch treatment include improved water availability, as found by (Edwards et al., 2000), and the addition of nutrients. In the on-station trials conducted in 2020 and 2021, the controls were fertilized with 100 kg N/ha to compensate for the nutrient input from the mulch treatment. In these trials, the yields of mulched and unmulched potatoes did not differ significantly, indicating that the mulch can be a substantial source of nutrition for the potatoes and as such can compensate for external fertilization.
Organic mulching can play an important role in the preventive regulation of CPB infestation but should not be considered an exhaustive measure. Probably the most effective measure that can lead to a practical absence of CPB is to cultivate a field at least 1.5 km away from the potato fields of the previous year (Weisz et al., 1994, 1996). Where this is not possible, at least maximizing the distance can be combined with other measures, of which organic mulching can be an important one. Other classically recommended preventive measures are the removal of potato volunteers, pre-sprouting of seed potatoes and the use of early and tolerant varieties. However, these are not always considered to be very effective (Kühne, 2021). As curative measures, insecticides are also available. However, due to increasing failure and the constant need for new agents, Alyokhin et al. (2015) have already considered the case of CPB as an “insecticide treadmill”.
From an economic point of view, the application of organic mulches can save crop protection measures, but this alone does not justify the rather expensive application of mulches. However, organic mulching is associated with a number of other beneficial properties. These include considerable reduction of potato late blight (caused by Phytophthora infestans) under temperate (Finckh et al., 2018) as well as tropical (Nazareno et al., 2020) conditions, the reduction of evaporation and the maintenance of soil water contents (Edwards et al., 2000; Johnson et al., 2004; Farzi et al., 2017), a strong reduction in soil erosion on sloped fields (Edwards et al., 2000; Döring et al., 2005) and the suppression of weeds (Teasdale and Mohler, 2000; Genger et al., 2018). In addition, the use of straw has been shown to be far superior to insecticides in the control of aphid vectors transmitting non-persistent viruses in seed potato production (Kirchner et al., 2014), with the bonus of not harming beneficial insects. Some studies even indicated a direct promotion of arthropod predators when mulch is used (Brust, 1994; Dudas et al., 2016; Genger et al., 2018). Plant materials for organic mulching can originate from by-products of the harvest, green manure, or meadows and can considerably contriburte to plant nutrition as can be seen when comparing the yields under mulch versus 100 kg N fertilizer in the on-station trials in 2021 (Figure 6). These can thus be used as fertiliser in livestock-reduced or livestock-free agriculture, a form of agriculture that is considered an essential component of the future global food security (Röös et al., 2020).
In conclusion, organic mulching can be a valuable preventive component in on-farm CPB control. Due to its positive impact on yield and many other benefits, the use of mulch is also economically attractive. Further elucidation of the still largely unknown mechanism could in future help to refine the method and enhance the effect of mulch on CPB.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: http://dx.doi.org/10.48662/daks-28; DaKS – Datenrepository der Universität Kassel.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
JW: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. SJ: Writing – review & editing. AN: Writing – review & editing. OH: Supervision, Writing – review & editing. MF: Writing – review & editing. SK: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was mainly funded by the European Union’s Horizon 2020 Research and Innovation programme as part of the project EcoStack (Grant Agreement no. 773554). Part of the work was also funded by the European Union FP7 Project (no. 289277), “Optimizing Subsidiary Crop Application in Rotations” (OSCAR) and the German Federal Ministry of Food and Agriculture (BMEL) under a resolution of the German Bundestag through the Federal Agency for Agriculture and Food (BLE) under the Federal Program for Organic Farming and Other Forms of Sustainable Agriculture, grant number 2818OE016 (VORAN – Improving Organic Crop Rotations with Transfer Mulch for Regenerative Appropriate Nutrient Management).
Acknowledgments
We thank Rainer Wedemeyer, Mario Plass, and Joachim Deckers, as well as the farmers for managing the potato trials at the Experimental Farm of the University of Kassel and both commercial farms. We are also grateful to Inger Deilke, Simeon Leisch-Waskönig, Lara Schubert, Vera Stolle-Brüers, Sarah Bender and Felix Rudolph for assisting with sampling.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alyokhin A., Atlihan R. (2005). Reduced fitness of the Colorado potato beetle (Coleoptera: Chrysomelidae) on potato plants grown in manure-amended soil. Environ. Entomol. 34, 963–968. doi: 10.1603/0046–225X-34.4.963
Alyokhin A., Mota-Sanchez D., Baker M., Snyder W. E., Menasha S., Whalon M., et al. (2015). The Red Queen in a potato field: integrated pest management versus chemical dependency in Colorado potato beetle control. Pest Manage. Sci. 71, 343–356. doi: 10.1002/ps.3826
Alyokhin A., Rondon S. I., Gao Y. (2022). Insect pests of potato: global perspectives on biology and management (San Diego: Academic Press).
BLE. (2023). Bericht zur Markt- und Versorgungslage Kartoffeln. Bundesanstalt für Landwirtschaft und Ernährung. Available online at: https://www.ble.de/SharedDocs/Downloads/DE/BZL/Daten-Berichte/Kartoffeln/2023BerichtKartoffeln.pdf?:blob=publicationFile&v=2.
Brooks M. E., Kristensen K., Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R. J. 9, 378–400. doi: 10.32614/RJ-2017–066
Brust G. E. (1994). Natural enemies in straw-mulch reduce Colorado potato beetle populations and damage in potato. Biol. Control. 4, 163–169. doi: 10.1006/bcon.1994.1026
Brust G. E. (1996). Interaction of mulch and Bacillus thuringiensis subsp. tenebrionis on Colorado potato beetle (Coleoptera: Chrysomelidae) populations and damage in potato. J. Econ. Entomol. 89, 467–474. doi: 10.1093/jee/89.2.467
Döring T. F., Brandt M., Heß J., Finckh M. R., Saucke H. (2005). Effects of straw mulch on soil nitrate dynamics, weeds, yield and soil erosion in organically grown potatoes. Field Crops Res. 94, 238–249. doi: 10.1016/j.fcr.2005.01.006
Döring T. F., Röhrig K. (2016). Behavioural response of winged aphids to visual contrasts in the field. Ann. Appl. Biol. 168, 421–434. doi: 10.1111/aab.12273
Dudas P., Menyhart L., Gedeon C., Ambrus G., Toth F. (2016). The effect of hay mulching on soil temperature and the abundance and diversity of soil-dwelling arthropods in potato fields. Eur. J. Entomol. 113, 456–461. doi: 10.14411/eje.2016.059
Dupuis B., Nkuriyingoma P., Ballmer T. (2024). Economic impact of potato virus Y (PVY) in europe. Potato. Res. 67, 55–72. doi: 10.1007/s11540–023-09623-x
Dvořák P., Tomášek J., Kuchtová P., Hamouz K., Hajšlová J., Schulzová V. (2012). Effect of mulching materials on potato production in different soil-climatic conditions. Roman. Agric. Res. 29, 201–209.
Edwards L., Burney J. R., Richter G., MacRae A. H. (2000). Evaluation of compost and straw mulching on soil-loss characteristics in erosion plots of potatoes in Prince Edward Island, Canada. Agricult. Ecosyst. Environ. 81, 217–222. doi: 10.1016/S0167–8809(00)00162–6
EPPO. (2022). Leptinotarsa decemlineata (LPTNDE) (EPPO Global Database). Available online at: https://gd.eppo.int/taxon/LPTNDE (Accessed November 25, 2022).
Eurostat. (2021). The EU potato sector - statistics on production, prices and trade. Available online at: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=The_EU_potato_sector_-_statistics_on_production,_prices_and_trade (Accessed March 30, 2024).
Farzi R., Gholami M., Baninasab B., Gheysari M. (2017). Evaluation of different mulch materials for reducing soil surface evaporation in semi-arid region. Soil Use Manage. 33, 120–128. doi: 10.1111/sum.12325
Ferro D. N., Logan J. A., Voss R. H., Elkinton J. S. (1985). Colorado potato beetle (Coleoptera: Chrysomelidae) temperature-dependent growth and feeding rates. Environ. Entomol. 14, 343–348. doi: 10.1093/ee/14.3.343
Ferro D. N., Tuttle A. F., Weber D. C. (1991). Ovipositional and flight behavior of overwintered Colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 20, 1309–1314. doi: 10.1093/ee/20.5.1309
Finckh M., Junge S., Schmidt J., Weedon O. (2018). “Disease and pest management in organic farming: a case for applied agroecology,” Improving organic crop cultivation, Köpke U. (London: Burleigh Dodds Science Publishing), 271–301. doi: 10.1201/9781351114578
Genger R. K., Rouse D. I., Charkowski A. O. (2018). Straw mulch increases potato yield and suppresses weeds in an organic production system. Biol. Agric. Horticult. 34, 53–69. doi: 10.1080/01448765.2017.1371077
Hare D. J. (1980). Impact of defoliation by the Colorado potato beetle on potato yields. J. Econ. Entomol. 73, 369–373. doi: 10.1093/jee/73.3.369
Hare J. D. (1990). Ecology and management of the Colorado potato beetle. Annu. Rev. Entomol. 35, 81–100. doi: 10.1146/annurev.en.35.010190.000501
Hartig F. (2022). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Available online at: https://CRAN.R-project.org/package=DHARMa.
Hothorn T., Bretz F., Westfall P. (2008). Simultaneous inference in general parametric models. Biometrical. J. 50, 346–363. doi: 10.1002/bimj.200810425
Johnson J. M., Hough-Goldstein J. A., Vangessel M. J. (2004). Effects of straw mulch on pest insects, predators, and weeds in watermelons and potatoes. Environ. Entomol. 33, 1632–1643. doi: 10.1603/0046–225X-33.6.1632
Junge S. M., Leisch-Waskönig S., Winkler J., Kirchner S. M., Saucke H., Finckh M. R. (2022). Late to the party—Transferred mulch from green manures delays colorado potato beetle infestation in regenerative potato cropping systems. Agriculture 12, 2130. doi: 10.3390/agriculture12122130
Kirchner S. M., Hiltunen L. H., Santala J., Döring T. F., Ketola J., Kankaala A., et al. (2014). Comparison of straw mulch, insecticides, mineral oil, and birch extract for control of transmission of potato virus Y in seed potato crops. Potato. Res. 57, 59–75. doi: 10.1007/s11540–014-9254–4
Koch M., Naumann M., Pawelzik E., Gransee A., Thiel H. (2020). The importance of nutrient management for potato production part I: plant nutrition and yield. Potato. Res. 63, 97–119. doi: 10.1007/s11540–019-09431–2
Kühne S. (2021). Kartoffelkäfer (Leptinotarsa decemlineata (Say 1824)) in Deutschland – Regulierungsstrategien von 1936 bis in die Gegenwart. J. für. Kulturpflanzen. 73, 225–232. doi: 10.5073/JfK.2021.07–08.06
Mota-Sanchez D., Wise J. C. (2022). Leptinotarsa decemlineata. Arthropod Pesticide Resistance Database. Available online at: https://www.pesticideresistance.org/display.php?page=species&arId=141&deleteParam=genus&key=leptinotarsa (Accessed November 26, 2022).
Nazareno N. R. X., Finckh M. R., dos Santos Canalli L. B., Yada I. F. U., Kawakami J. (2020). Potential use of fresh mulch to curb potato late blight epidemics in Brazil. Summa. Phytopathol. 46, 191–197. doi: 10.1590/0100–5405/216539
Pulatov B., Jönsson A. M., Wilcke R. A., Linderson M.-L., Hall K., Bärring L. (2016). Evaluation of the phenological synchrony between potato crop and Colorado potato beetle under future climate in Europe. Agricult. Ecosyst. Environ. 224, 39–49. doi: 10.1016/j.agee.2016.03.027
Racca P., Tschöpe B., Falke K., Kleinhenz B., Rossberg D. (2014). “Forecasting of Colorado Potato Beetle Development with Computer Aided System SIMLEP Decision Support System,” in Integrated Pest Management. Ed. Abrol D. P. (Academic Press, San Diego), 79–91. doi: 10.1016/B978–0-12–398529–3.00006–3
R Core Team. (2022). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Röös E., Carlsson G., Ferawati F., Hefni M., Stephan A., Tidåker P., et al. (2020). Less meat, more legumes: prospects and challenges in the transition toward sustainable diets in Sweden. Renewable Agric. Food Syst. 35, 192–205. doi: 10.1017/S1742170518000443
Saucke H., Döring T. F. (2004). Potato virus Y reduction by straw mulch in organic potatoes. Ann. Appl. Biol. 144, 347–355. doi: 10.1111/j.1744–7348.2004.tb00350.x
Schreefel L., Schulte R. P. O., de Boer I. J. M., Schrijver A. P., van Zanten H. H. E. (2020). Regenerative agriculture – the soil is the base. Global Food Secur. 26, 100404. doi: 10.1016/j.gfs.2020.100404
Singh I., Hussain M., Manjunath G., Chandra N., Ravikanth G. (2023). Regenerative agriculture augments bacterial community structure for a healthier soil and agriculture. Front. Agron. 5. doi: 10.3389/fagro.2023.1134514
Stoner K. A. (1993). Effects of straw and leaf mulches and trickle irrigation on the abundance of Colorado potato beetles (Coleoptera: Chrysomelidae) on potato in Connecticut. J. Entomol. Sci. 28, 393–403. doi: 10.18474/0749–8004-28.4.393
Stoner K. A., Ferrandino F. J., Gent M. P. N., Elmer W. H., LaMondia J. A. (1996). Effects of straw mulch, spent mushroom compost, and fumigation on the density of Colorado potato beetles (Coleoptera: Chrysomelidae) in potatoes. J. Econ. Entomol. 89, 1267–1280. doi: 10.1093/jee/89.5.1267
Szendrei Z., Kramer M., Weber D. C. (2009). Habitat manipulation in potato affects Colorado potato beetle dispersal. J. Appl. Entomol. 133, 711–719. doi: 10.1111/j.1439–0418.2009.01429.x
Szendrei Z., Weber D. C. (2009). Response of predators to habitat manipulation in potato fields. Biol. Control. 50, 123–128. doi: 10.1016/j.biocontrol.2009.04.003
Teasdale J. R., Mohler C. L. (2000). The quantitative relationship between weed emergence and the physical properties of mulches. Weed. Sci. 48, 385–392. doi: 10.1614/0043–1745(2000)048[0385:TQRBWE]2.0.CO;2
Tripolskaja L., Romanovskaja D., Slepetiene A., Razukas A., Sidlauskas G. (2014). Effect of the chemical composition of green manure crops on humus formation in a Soddy-Podzolic soil. Eurasian. Soil Sc. 47, 310–318. doi: 10.1134/S1064229314040097
Voss R. H., Ferro D. N. (1990). Phenology of flight and walking by Colorado potato beetle (Coleoptera: Chrysomelidae) adults in western Massachusetts. Environ. Entomol. 19, 117–122. doi: 10.1093/ee/19.1.117
Wang C., Hawthorne D., Qin Y., Pan X., Li Z., Zhu S. (2017). Impact of climate and host availability on future distribution of Colorado potato beetle. Sci. Rep. 7, 4489. doi: 10.1038/s41598–017-04607–7
Weber D. C., Ferro D. N. (1993). Distribution of overwintering Colorado potato beetle in and near Massachusetts potato fields. Entomol. Experimentalis. Applicata. 66, 191–196. doi: 10.1111/j.1570–7458.1993.tb00708.x
Weisz R., Smilowitz Z., Christ B. (1994). Distance, rotation, and border crops affect Colorado potato beetle (Coleoptera: Chrysomelidae) colonization and population density and early blight (Alternaria solani) severity in rotated potato fields. J. Econ. Entomol. 87, 723–729. doi: 10.1093/jee/87.3.723
Weisz R., Smilowitz Z., Fleischer S. (1996). Evaluating risk of colorado potato beetle (Coleoptera: chrysomelidae) infestation as a function of migratory distance. J. Econ. Entomol. 89, 435–441. doi: 10.1093/jee/89.2.435
Wickham H. (2016). ggplot2: Elegant Graphics for Data Analysis (New York: Springer). Available at: https://ggplot2.tidyverse.org.
Wickham H., Averick M., Bryan J., Chang W., McGowan L. D., François R., et al. (2019). Welcome to the tidyverse. J. Open Source Software. 4, 1686. doi: 10.21105/joss.01686
Zehnder G. W., Hough-Goldstein J. (1990). Colorado potato beetle (Coleoptera: Chrysomelidae) population development and effects on yield of potatoes with and without straw mulch. J. Econ. Entomol. 83, 1982–1987. doi: 10.1093/jee/83.5.1982
Keywords: agroecological plant protection, biological control, green manures, integrated pest management, on-farm trial, organic agriculture, organic mulches
Citation: Winkler J, Junge SM, Nasirahmadi A, Hensel O, Finckh MR and Kirchner SM (2024) Reduction of Colorado potato beetle damage by various organic mulches. Front. Agron. 6:1335388. doi: 10.3389/fagro.2024.1335388
Received: 08 November 2023; Accepted: 04 June 2024;
Published: 19 June 2024.
Edited by:
Murray B. Isman, University of British Columbia, CanadaReviewed by:
Jozsef Kiss, Szent István University, HungarySilvia I. Rondon, Oregon State University, United States
Copyright © 2024 Winkler, Junge, Nasirahmadi, Hensel, Finckh and Kirchner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julian Winkler, anVsaWFuLndpbmtsZXJAdW5pLWthc3NlbC5kZQ==
†Present address: Stephan M. Junge, Interessensgemeinschaft Gesunder Boden, Regensburg, Germany
Abozar Nasirahmadi, Department of Energy and Technology, Swedish University of Agricultural Sciences, Uppsala, Sweden