 Nolwenn Kergosien
Nolwenn Kergosien Valérie Stiger-Pouvreau
Valérie Stiger-Pouvreau Solène Connan
Solène Connan Franck Hennequart2
Franck Hennequart2- 1Univ Brest, CNRS, IRD, Ifremer, LEMAR, Plouzane, France
- 2ALGAIA, R&D Center, Saint-Lô, France
The worldwide growing food demand and the excessive use of synthetic and chemical inputs compel the agricultural sector to find innovative and sustainable solutions to enhance or at least maintain crop yields in times of increased abiotic stresses levels linked to global change. Currently, great research efforts are carried out on brown seaweeds as their environment lead them to produce a broad range of compounds, with osmoregulatory, antioxidant, pro-bacterial, and plant-growth promoting activities. Indeed, numerous studies are looking at different combinations of algal species, extraction processes, plant species and environments of plant culture to highlight the various effects of algal extracts on plant growth and development, and resistance to abiotic stresses. Consequently, a wide variety of novel commercial products are emerging, presenting diversified chemical compositions, formulations and means of application. Such products allow the biostimulation of plants and soil by alleviating abiotic stresses such as drought, frost, and salt. The action of brown macroalgal extracts on plant and soil health has been repeatedly demonstrated, yet the precise relation between the extract chemical composition and its subsequent effect is still to be elucidated, as molecular synergy is suspected to play an important role. In this review, we present the interest of using brown macroalgal extracts to produce biostimulants with beneficial action on soil health, plant growth and development, as well as resistance against abiotic stresses, in relation to the molecular changes occurring.
1 Introduction
The upcoming years will see the agriculture sector facing several threats that could hinder its capacity to ensure global food safety. Indeed, world population is estimated to reach 9.7 billion by 2050 (UN DESA, 2017), considerably increasing crop production need for direct human consumption and livestock farming. Meanwhile, global change is altering biotic and abiotic factors (Shrivastava and Kumar, 2015; Raza et al., 2019), negatively impacting crop development and yield (European Environment Agency, 2017; FAO, 2021). Hence, the crop production demand will rise in an increasingly hostile environment for agricultural cultivation, which calls for a modification of current agricultural practices. Additionally, as ecological awareness is gaining momentum, being durable and safe is one of the leading challenges of modern agriculture (Sujeeth et al., 2022). During the last decades, intensive agriculture has rendered the once-fertile arable soils into impoverished lands, both in terms of nutrients and physical structure (Patle et al., 2019; Mandal et al., 2020). Non-degradable synthetic inputs have tenacious persistence in the environment and are responsible for eutrophication from run-offs and water pollution. These chemicals also have harmful consequences on human and animal health, stemming from their strong toxicity (Ali et al., 2021a). Consequently, more and more legislations are being passed to manage nutrient excess and reduce the range of harmful synthetic agricultural products, which have resulted in an increase in fertiliser prices (European Commission, 2021; Baffes and Chian Koh, 2022). As a result, the agricultural industry attempts to reduce synthetic inputs in crops and is pressured to look for efficient natural alternatives.
Biostimulants are currently gaining attention in this regard. According to the last European regulation on fertilisers EU2019/1009, a biostimulant is a product of natural origin, that “stimulates plant nutrition processes independently of the product’s nutrient content with the sole aim of improving one or more of the following characteristics of the plant or the plant rhizosphere: (a) nutrient use efficiency, (b) tolerance to abiotic stress, (c) quality traits, (d) availability of confined nutrients in soil or rhizosphere” (European Parliament and Council, 2019). Overall, the use of biostimulants on crops lead to an increase of the parameters sought by the agri-food sector, i.e. food product yield, quality, and shelf-life (EBIC, 2021). Their utilization, in synergy with organic and inorganic fertilisers, enhances the effects of the latter, and confers abiotic stress resilience (Rouphael and Colla, 2018). It allows the reduction of synthetic inputs, and indirectly their impact on the environment by limiting leachates. The biostimulant market is ever-growing, with a worldwide double-digit growth, showing a will of the agricultural sector to shift their practices (Smiglak-Krajewska and Wojciechowska-Solis, 2021; Moolla, 2022). This rapid growth is also linked to numerous actors and products already present on the market, sometimes creating confusion on terminologies and claimed product effects (Caradonia et al., 2019; Moolla, 2022). To harmonize the European market, a new regulation frames the registration of biostimulants and the authorization to commercialize under this nomenclature. It encompasses several criteria, including evidences of biostimulant effects by independent studies, and microbiological and chemical safety, notably regarding heavy metals contamination (European Parliament and Council, 2019).
Biostimulants are categorized according to their raw material source, their principal bioactive compound, or their mode of action (du Jardin, 2015). Among them, one of the most promising classes is brown macroalgal extracts (BME). They do not naturally possess enough fertilizing power to be classified as fertilisers, but they can stimulate plant internal responses for growth and defense, thus having tremendous potential for the agricultural sector (Samuels et al., 2022). Overall, BME application results in increases in production levels, health and quality of crops (Nanda et al., 2022), and is considered harmless for humans, animals and the environment (Renaut et al., 2019). Finally, as brown macroalgae are growing at sea, they do not enter in land competition with crop production (Deolu-Ajayi et al., 2021) and do not require water nor fertilisers supply to grow. Therefore, BME are rapidly developing in the biostimulant market, with the emergence of numerous commercial products (El Boukhari et al., 2020; Critchley et al., 2021; EBIC, 2021). Brown macroalgae, or Phaeophyceae (Chromista, Ochrophyta), are the second largest group of macroscopic algae, with ~2,100 identified species (Guiry and Guiry, 2023). They all contain a specific xanthophyll pigment, fucoxanthin, responsible for their characteristic brown-yellow color (Lee, 1999). They also produce a broad range of bioactive compounds, of which a large number have positive effects on plant growth and development, and stress alleviation (Sujeeth et al., 2022).
The present review aims to highlight the potential of BME as biostimulants for agrochemical alternatives. Various brown macroalgal compounds displaying plant promotion and stress alleviation activities are first presented. Then, biostimulant activities are displayed in respect to macroalgal species, plant species and potential compounds involved. Though biocontrol-like activities have sometimes been reported for biostimulant products, this review only focuses on biostimulant effects i.e., under abiotic stresses. Finally, the potential molecular mechanisms implicated in the various activities are presented. The conceptual diagram in Figure 1 summarizes the full potential of BME regarding plant global health.
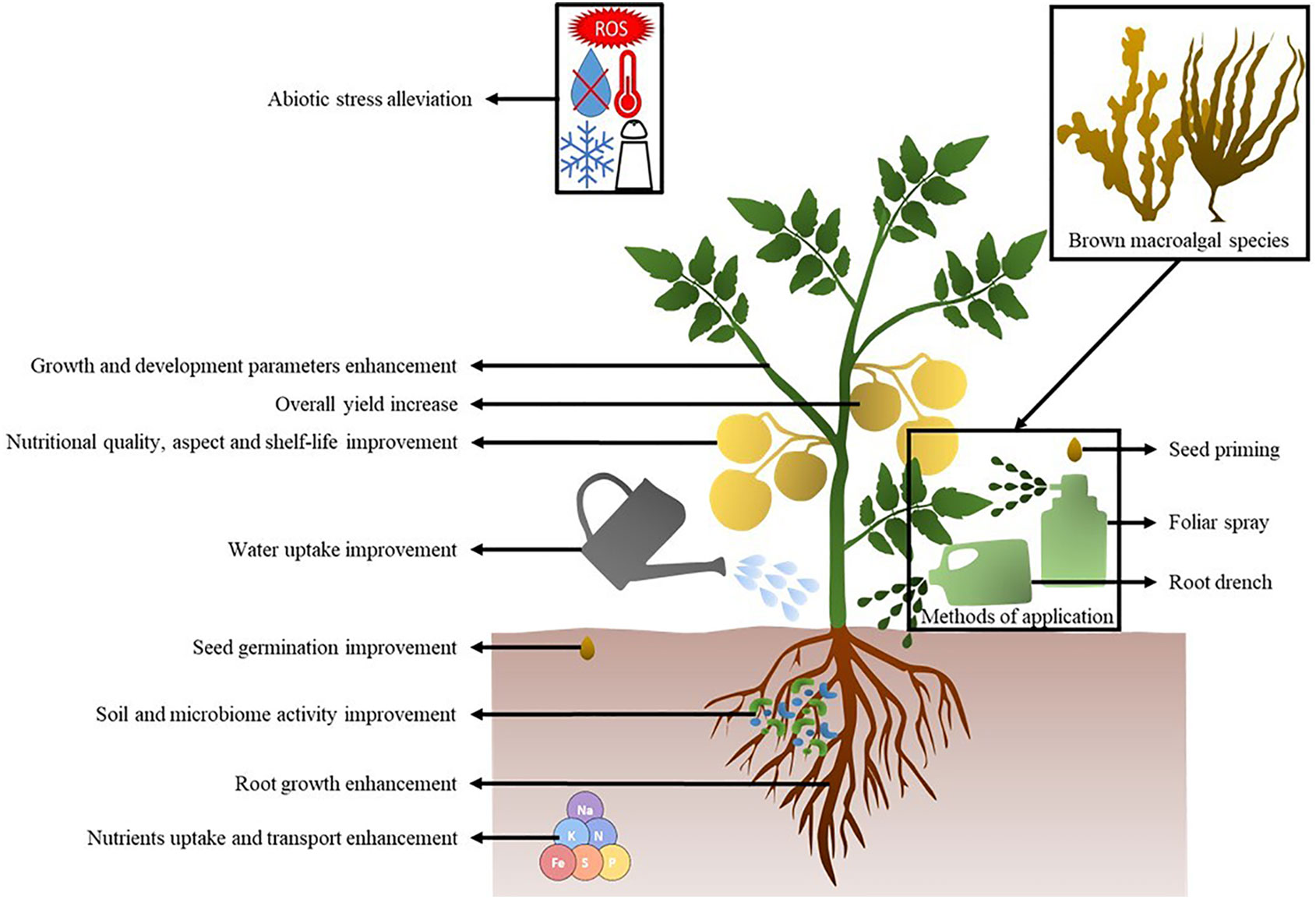
Figure 1 Conceptual diagram of the beneficiary effects of brown macroalgal extracts on plants.
2 Brown macroalgal metabolites beneficial to plant health and growth
Brown macroalgae biochemical composition is modulated by their stressful living environment, which involves both biotic and abiotic parameters. In response, they produce a wide range of bioactive metabolites that modulate their resilience to stress factors (Connan and Stengel, 2011; Stengel et al., 2011; Lalegerie et al., 2020). These metabolites, i.e. minerals, carbohydrates, proteins, lipids, phlorotannins, pigments, vitamins, have applications in numerous sectors, including plant health and growth (Sujeeth et al., 2022).
Brown macroalgae have strong absorption and accumulation capacities of surrounding salts and minerals. As a result, they usually have a high mineral content, between 9 and 44% of the algal dry weight (DW). This content varies depending on species, seasonality, and surrounding seawater mineral concentration (Circuncisão et al., 2018). Minerals are involved in various plant cellular metabolism pathways and can trigger and/or directly enhance plant growth and defense mechanisms, leading to enhanced crop yields and quality (Kolbert et al., 2022; Lešková et al., 2022). Indeed, metal elements such as copper (Van Nguyen et al., 2022), iron (Rout and Sahoo, 2015; Murgia et al., 2022), manganese (Alejandro et al., 2020), molybdenum (Huang et al., 2022), nickel (Tanveer, 2020), and zinc (Thiébaut and Hanikenne, 2022) are essential plant micronutrients, involved in energy metabolism, photosynthesis, respiration, reproduction, gene expression regulation, and stress tolerance (D’Oria et al., 2021). Conversely, mineral deficiencies can reduce nutrient uptake and metabolism, which in turn negatively affect growth, yield and quality of the harvested product (Courbet et al., 2019; Thiébaut and Hanikenne, 2022).
Brown macroalgae are especially rich in carbohydrates, i.e. alginates, fucoidans, laminarans, mannitol and cellulose (Percival and McDowell, 1967; Stiger-Pouvreau et al., 2016), reaching up to 80% DW in some species; this proportion varying greatly according to species, geographical location, maturation degree and seasonality (Stiger-Pouvreau et al., 2016; Mohd Fauziee et al., 2021). Carbohydrates have several functions in soil health and crop enhancement, notably as metabolic triggers signals for physiological defense responses such as the production of phytochemical compounds (Laporte et al., 2007; Briceño-Domínguez et al., 2014). These effects were notably observed in cabbage (Di Stasio et al., 2017b), tobacco (Chandía and Matsuhiro, 2008), tomato (Mzibra et al., 2021), and wheat (Chandía et al., 2004; Zou et al., 2019; Zou et al., 2021).
Proteins are not usually abundant in brown macroalgae, ranging from 3 to 15% DW, owing to species, seasonality and environmental conditions such as temperature variations (Fleurence, 1999). Brown macroalgae are rich in aspartic and glutamic acids (Pangestuti and Kim, 2015), which are involved in the biosynthesis of nitrogen-containing compounds such as proline (El-Metwally et al., 2022), improve nitrogen uptake (Colla et al., 2015; El-Metwally et al., 2022), have a positive effect on photosynthetic activity (El-Metwally et al., 2022), and reduce physiological damages induced by oxidative stress (Rizwan et al., 2017; El-Metwally et al., 2022).
Brown macroalgae have a low lipid content, ranging from 0.3 to 6% DW (Nugroho Jati et al., 2019; Alghazeer et al., 2022). Lipophilic compounds play roles in plant growth and stress resilience processes, regulating cell membrane permeability and increasing the tolerance to freezing stress (Rayorath et al., 2008b; Muthuramalingam et al., 2022).
Phlorotannins, specific phenolic compounds of brown macroalgae (Stengel et al., 2011; Stiger-Pouvreau et al., 2014), range from 0.1 to 18% DW (Connan et al., 2004; Stiger-Pouvreau et al., 2014) depending on localization, season, light, and algal species and age (Stiger-Pouvreau et al., 2014). They have a broad range of bioactivities, including antioxidant, antibacterial, antifungal, anthelmintic and nematicide to alleviate biotic and abiotic stresses in plants (Zubia et al., 2015), and enhance the nutritional quality of food products (Teklić et al., 2021).
Finally, pigments or vitamins could have biostimulant properties, but few studies have been made to evaluate their effects on plants (Khan et al., 2009; Ali et al., 2021a; Teklić et al., 2021).
3 Beneficiary effects of BME on plants
BME have multiple beneficiary effects on plants (Figure 1; Table 1). Several parameters influence the ability to obtain these desired outcomes. Indeed, various combinations of BME (species, methods of extraction, qualitative composition…), plant species and methods of application (dose rate, foliar or root drench application…) induce a wide variability of end results (Caradonia et al., 2022).
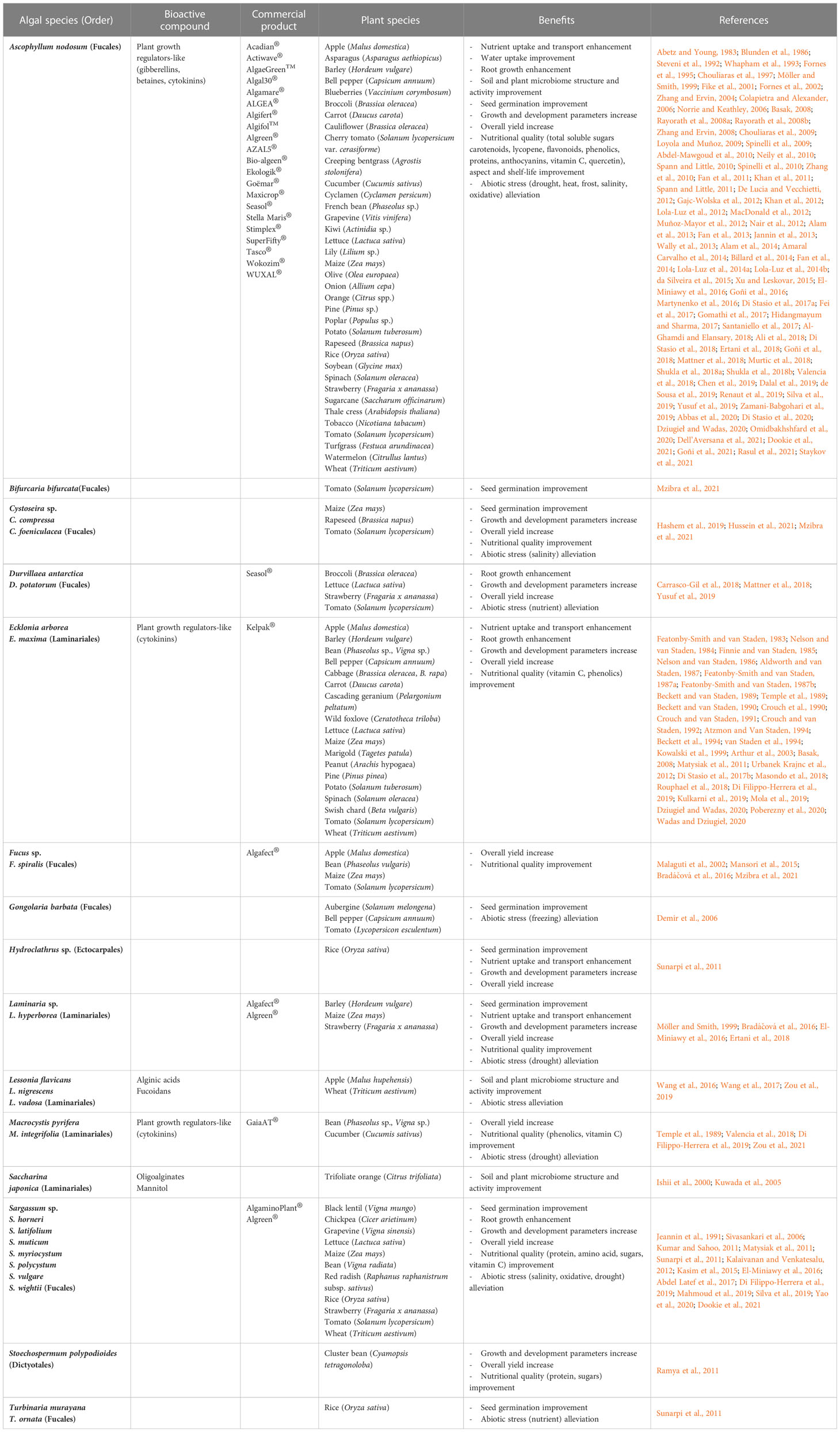
Table 1 Brown macroalgal species, bioactive compounds and associated biostimulant commercial products, regarding their activities on various plants.
3.1 Soil structure and quality
A healthy soil is essential for plant productivity. BME can improve soil physical, chemical, and biological properties (Samuels et al., 2022). Indeed, they contain several chelating agents, such as alginates, that can combine with metallic ions to form high molecular weight aggregates (Kholssi et al., 2022). These aggregates promote water absorption and retention, as well as soil stability and aeration through improved pores capillary activity (Illera-Vives et al., 2015; Shukla et al., 2019). Furthermore, BME improve nutrients availability by stimulating soil microbial activity for nutrient mineralization (Renaut et al., 2019), and as a direct source of organic matter and trace elements (Di Filippo-Herrera et al., 2019). BME can also function as biosorption agents for polluted soils bioremediation. Their polyanionic abilities allow the entrapment of pollutants, notably heavy metals, to eliminate them from the environment (Amador-Castro et al., 2021).
3.2 Root growth and nutrients uptake
BME can stimulate plant roots growth and development, in particular when applied on early growth stages (Samuels et al., 2022). Growth and elongation stimulatory activity on tomato roots was reported for the commercial Ecklonia maxima extract at lower concentrations (0.038-0.057g/L), while a more concentrated extract (0.230g/L) inhibited root growth (Finnie and van Staden, 1985). The authors related the activity to endogenous plant hormones and found an equivalent activity with 0.1 nM cytokinin zeatin. Root growth and development parameters enhancement were also assessed both in lettuce treated by a Durvillaea commercial mixture (Yusuf et al., 2019), and strawberry treated by Ascophyllum nodosum (AN), Sargassum sp. and Laminaria sp. extracts under nutrient stress (El-Miniawy et al., 2016). Improvements in the root system allow a better nutrients and water absorption and use, by acting on genes involved in nutrient root uptake. Indeed, nitrogen, phosphorus, potassium and sulphur uptakes were enhanced after the application of the commercial AN extract, following the improvement of rapeseed root growth (Billard et al., 2014). Similarly, a Laminaria sp. acid extract improved boron, calcium, copper, iron, manganese, magnesium, molybdenum, sulphur and zinc uptake in nutrient-stressed maize (Ertani et al., 2018). The root stimulatory effect was lost when the extract was reduced to ash, suggesting an organic origin of the bioactive compounds (Zhang and Ervin, 2004).
3.3 Soil and plant microbiome
Leaves, roots and surrounding soil are fully colonized by microorganisms, allowing active microbe-plant interactions. These surfaces secrete soluble compounds such as carbohydrates, amino and organic acids, that can be used as an carbon source by a wide range of beneficial microorganisms for their proliferation (Haichar et al., 2008). Microorganisms have an important role in soil health and healthy plant growth and development, through biofilm formation for particles aggregation, water retention, nutrient cycling and transport, degradation of toxic substances, and control of plant illnesses (Renaut et al., 2019).
The action mechanism of BME on rhizosphere and phyllosphere microbiota and the precise interactions between microbiota and plants are still unknown. Extracts characterization, observations of their impacts on microbial communities and resulting plant-microbe interactions can help to understand the links between algal extract composition, microbial structure and plant growth enhancement (Renaut et al., 2019). The modification of the plant and soil secretions by BME can influence the structure and activity of the microbial population, which in turn impact the plant growth and development process (Shukla et al., 2019). In this sense, an increase in root and soil microbial α-diversities and a low but significant positive impact on microbial β-diversity following the application of an AN commercial extract on pepper and tomato were observed (Renaut et al., 2019). The rhizosphere microbial α-diversity of strawberry was also increased after the application of another AN commercial extract rich in betaine, alginic acid and caidrine (Spinelli et al., 2010). Both studies suggest at least a partial implication of the extracts on the structure of the microbial communities. BME can also prompt beneficial soil microbial growth and activity, as well as soil conditioning compounds production by these microorganisms (Deolu-Ajayi et al., 2021).
3.4 Seed priming
Plant seedlings can be subjected to various abiotic stresses, including drought and salinity, threatening their germination and establishment. These stresses can trigger a physiological dormant state that prevents germination under unfavorable conditions. Application of BME can induce a “primed” physiological state of seedlings by inducing various biochemical changes that lead to dormancy break and trigger germination under stressful conditions (Masondo et al., 2018). Furthermore, seed priming also provides seedlings with additional sources of minerals, amino acids and soluble sugars to enhance germination and improve vigor (Silva et al., 2019). As a result, BME induce early seed germination and increase germination rate and establishment in many crops, for many algal species (Kumar and Sahoo, 2011; Lola-Luz et al., 2012; Masondo et al., 2018; Mzibra et al., 2021). For example, seedlings of aubergine, pepper and tomato subjected to cold temperatures, treated with a Gongolaria barbata extract had enhanced germination rates linked to improved vigour (Demir et al., 2006). Additionally, tomato seedlings treated with Cystoseira foeniculacea, Fucus spiralis and Bifurcaria bifurcata extracts increased germination rate and percentage (Mzibra et al., 2021). Seed priming also grants the grown plant faster and/or stronger responses to abiotic stresses (Sujeeth et al., 2022). Extract drenching is the main method of application for seed priming, with caution paid to extract composition and concentration, as high concentrations of extracts can lead to a germination inhibition, due to the presence of several growth regulators compounds (Silva et al., 2019).
3.5 Plant growth and development
BME have a positive impact on overall plant growth and development, demonstrated by an increase in root and shoot length, surface area and volume, stem size, and overall fresh and dry weight. For example, improvements in all physical and biochemical growth parameters of bean were observed after the application of a low concentrated Stoechospermum polypodioides extract (Ramya et al., 2011). Meanwhile, higher concentrations application resulted in growth inhibition. In addition, growth stimulatory activities of commercial AN and Durvillaea extracts were tested on nutrient-stressed lettuce, leading to root biomass increase (Yusuf et al., 2019). The impact of an AN commercial extract on lettuce, melon, pepper and tomato was studied under drought stress (Neily et al., 2010). In each case, the treatment allowed an improvement of growth parameters, as well as an early development of root and shoot. BME can be applied directly to soil and roots or as a foliar spray, and effects on plant growth are concentration-dependent (El Boukhari et al., 2020). The growth-promoting properties of the extracts could be attributed to the presence of oligosaccharides, amino acids and vitamins (Briceño-Domínguez et al., 2014; Renaut et al., 2019).
BME can increase the chlorophyll content of treated plant leaves for an improved photosynthesis. Indeed, a higher leaf chlorophyll content in nutrient-stressed lettuce was measured after AN commercial alkali extract application (Yusuf et al., 2019). The extract reduced chlorophyll degradation in chloroplasts, linked to plant endogenous betaines synthesis enhancement induced by BME (Blunden et al., 1996). Furthermore, antioxidant compounds, like phlorotannins and carotenoids, can protect the photosynthetic apparatus against photooxidative damages occurring during abiotic stress by scavenging reactive oxygen species (ROS) thus preventing chlorophyll degradation and increasing leaf chlorophyll content (Mansori et al., 2015). Extracts can also trigger early flowering and fruiting in bean (El-Yazied et al., 2012), pepper (Ali et al., 2021b), marigold (van Staden et al., 1994), strawberry (El-Miniawy et al., 2016) and tomato (Crouch and van Staden, 1992; Ali et al., 2021b). It is suspected that BME trigger the biosynthesis of endogenous cytokinins, which influence flowering (Samuels et al., 2022).
3.6 Crop yield and quality
Improvements of the above-mentioned parameters by BME result into enhanced crops yields, as observed in barley (Blunden et al., 1996), bean (El-Yazied et al., 2012), broccoli (Gajc-Wolska et al., 2012), cabbage (Di Stasio et al., 2017b), grapevine (Taskos et al., 2019), lettuce (Mola et al., 2019), maize (Hussein et al., 2021), onion (Hidangmayum and Sharma, 2017), pepper (Arthur et al., 2003), potato (Dziugieł and Wadas, 2020), rapeseed (Jannin et al., 2013), spinach (Rouphael et al., 2018), tomato (Yao et al., 2020), watermelon (Abdel-Mawgoud et al., 2010) and wheat (Amaral Carvalho et al., 2014). Additionally, an AN commercial extract had a positive impact on apple’s fruit set, number, size and weight, when applied at low concentrations (de Sousa et al., 2019). An increase was measured in maize ear weight of 37-42% and 48-50% after E. maxima and Sargassum sp. commercial extracts application, respectively (Matysiak et al., 2011). Increases in fruit size and harvest volume of blueberry (Loyola and Muñoz, 2009), grapevine (Norrie and Keathley, 2006; Khan et al., 2012), kiwi (Chouliaras et al., 1997), pepper (Arthur et al., 2003), potato (Dziugieł and Wadas, 2020) and tomato (Dookie et al., 2021) were also observed. Nevertheless, as commercial biostimulants are sometimes supplemented with growth enhancers including synthetic plant hormones to boost biostimulation effects, increases in yield cannot be solely attributed to BME.
3.7 Biofortification
The application of BME enhances the nutritional value of the crops. Indeed, the nutritional qualities of broccoli (Lola-Luz et al., 2014b), carrot (Poberezny et al., 2020), cucumber (Valencia et al., 2018), grapevine (Frioni et al., 2018), onion (Lola-Luz et al., 2014a), potato (Lola-Luz et al., 2014a), spinach (Fan et al., 2011) and strawberry (Kapur et al., 2018) were improved upon the addition of BME, such as AN or Macrocystis sp. In the same way, after BME application, the fruits accumulate higher amounts of healthy and taste-beneficial compounds, such as soluble carbohydrates (fructose and sucrose), phenols, flavonoids, anthocyanins, quercetin and vitamin C (Di Stasio et al., 2018; Frioni et al., 2018; Kapur et al., 2018; Valencia et al., 2018). Shelf-life seems to be extended by AN commercial extracts treatment on grapes (Norrie and Keathley, 2006) and spinach (Fan et al., 2014). On the contrary, AN and E. maxima commercial extracts application decreased preservation duration of apple fruits (Basak, 2008).
3.8 Resilience to abiotic stresses
Field-cultivated crops are regularly exposed to multiple abiotic stresses usually occurring simultaneously. They originate from environmental conditions variation, and usually encompass oxidative, hydric, frost, heat, and saline stresses (Samuels et al., 2022). BME contain compounds that can elicit several defense systems in plants by triggering stress signals involved in transduction pathways, and heighten resistance and resilience to these stresses to a certain extent (Larsen et al., 2003). Indeed, an AN commercial extract applied on cherry tomato under saline and drought stresses limited yield loss to 15% (Murtic et al., 2018).
Oxidative stress induces damaging free radicals and ROS production in the plant. In Arabidospis, pepper and tomato under oxidative stress, AN extract application induced the accumulation of cytokinins and phenolic compounds, which have ROS-scavenging capabilities (Omidbakhshfard et al., 2020; Staykov et al., 2021). Salinity and drought stresses can be similarly mitigated by a reduction of osmotic potential to avoid excessive water loss. This involves osmoprotectants accumulation, such as proline, carbohydrates, organic acids and proteins, and stimulation of stomatal closure, which can be enhanced by BME, such as AN extracts on asparagus (Al-Ghamdi and Elansary, 2018) and spinach (Xu and Leskovar, 2015), or Cystoseira sp. extract on rapeseed (Hashem et al., 2019). For example, long-term dehydration effect prevention consecutive to the application of AN extracts on drought-stressed Arabidopsis was observed (Santaniello et al., 2017; Rasul et al., 2021). Conversely, heat stress is mitigated by an increase in transpiration to improve thermoregulation (Franzoni et al., 2022). Freezing stress protection is enhanced by the accumulation of proline, soluble sugars and lipophilic compounds such as unsaturated fatty acids, as observed in freezing-stressed Arabidopsis treated by an AN lipophilic extract (Nair et al., 2012). Stress mitigation can also be enhanced by the application of several extracts from different macroalgal species at once, to handle a wider variety of stresses (Deolu-Ajayi et al., 2021).
4 Molecular modes of action of BME in crops
Currently, the precise mechanisms activated and regulated by BME application are still not fully understood, as the intricate molecular processes responsible for plant growth responses and stress alleviation induced by BME are complex. Extensive composition analysis of BME and improved genomic techniques to oversee resulting effects on plant physiology are key elements to unveil the processes in which the extracts constituents improve plant growth, health and vigour (Sujeeth et al., 2022). It is suspected that these compounds act as signaling mechanisms for specific metabolic and hormonal pathways, to modulate gene expression and induce metabolic changes in the plant (Nanda et al., 2022; EBIC, 2023; Table 2). Under stress conditions, these pathways can be amplified to improve plant adaptation and survival, or delay stress effects. Furthermore, some of these compounds present antioxidant activities, which directly reduce the deteriorating effects of free radicals produced by plants under stress conditions (Franzoni et al., 2022). Small variations in extract composition can lead to different molecular and cellular end-results, which suggest the activation of distinct underlying pathways. Furthermore, a single extract can concurrently trigger and/or restrain several metabolic or hormonal pathways. Thus, no generalities can be made on the mode of action and the subsequent benefits solely based on extract composition (Deolu-Ajayi et al., 2021).
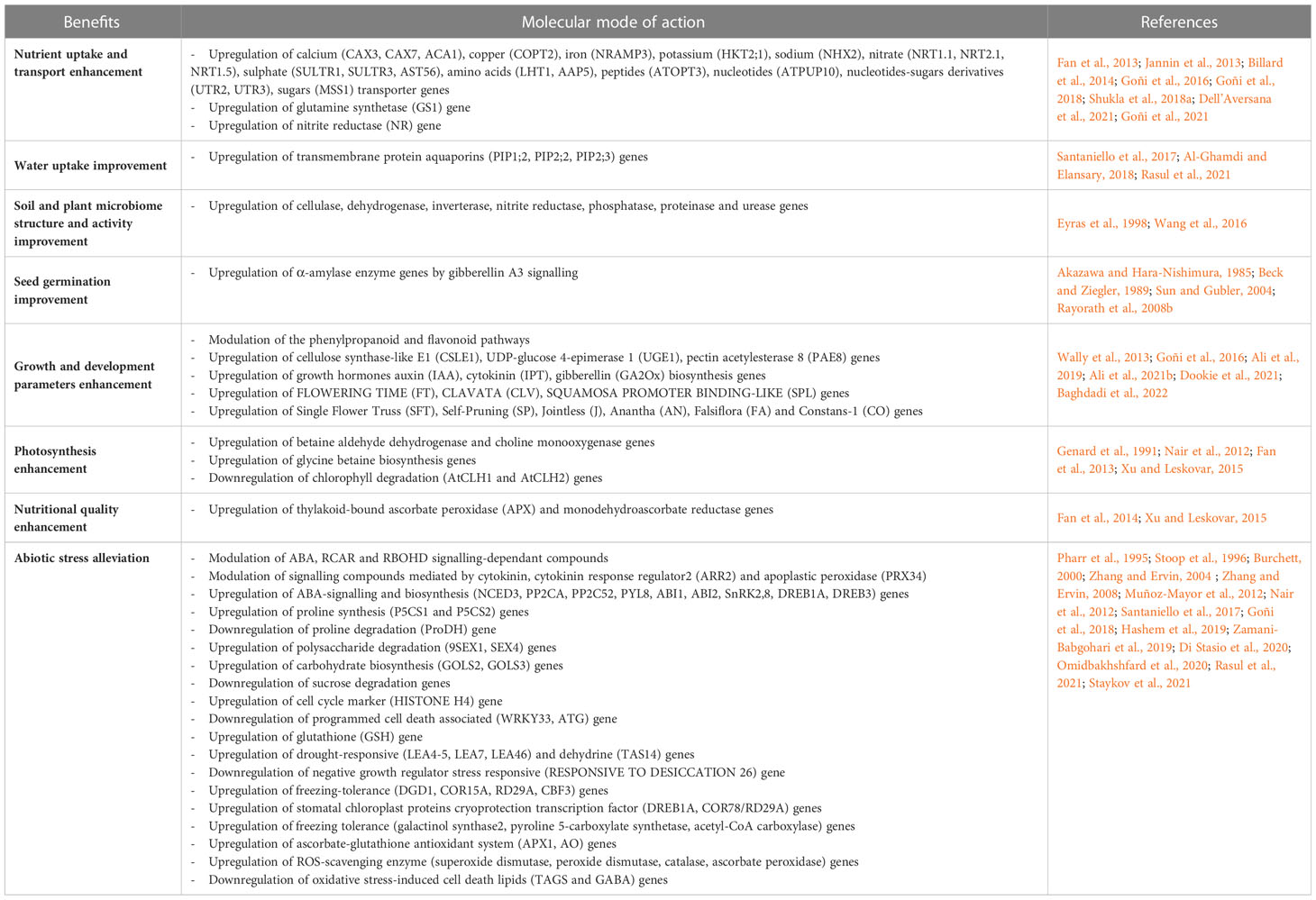
Table 2 Benefits induced by brown macroalgal extracts on plants, and their suspected molecular mode of action in plants.
4.1 Nutrient and water uptake and transport enhancement
Nutrients uptake and transport are stimulated through the upregulation of several genes responsible for transcription of nutrient transporters in the plant membrane, which can be induced by BME application. For example, AN extracts application on Arabidopsis and barley upregulated calcium, copper, nitrate, sulphate, amino acids, peptides, nucleotides, sugar, nucleotide-sugar derivatives transporters genes (Goñi et al., 2016; Goñi et al., 2021). Meanwhile, AN commercial extracts on spinach upregulate the glutamine synthetase gene that catalyses the inorganic nitrogen form, ammonium, into the organic form, glutamine, and the nitrate reductase gene responsible for the nitrate-to-nitrite reduction towards the assimilation of nitrate (Fan et al., 2013). BME application can also trigger the synthesis of membrane proteins aquaporins to facilitate water transmembrane exchanges (Santaniello et al., 2017; Al-Ghamdi and Elansary, 2018; Rasul et al., 2021).
4.2 Soil and plant microbiome structure and activity improvement
BME differentially trigger several enzyme genes involved in compounds degradation into assimilable nutrients for the microbial community development. Depending on the extract composition, microorganism genes expression varies differently, impacting encoded enzyme activity, such as cellulase, invertase, nitrite reductase, phosphatase, proteinase, and urease, for cellulose, carbohydrates, nitrite, phosphate, proteins, and urea degradation respectively (Eyras et al., 1998). Assimilable nutrients availability is thus altered, and differentially favors microbial communities. This positive modification of the microbiome structure was observed in apple cultivars following the application of a Lessonia extract (Wang et al., 2016).
4.3 Seed priming and germination improvement
The priming action of BME to enhance the seed resistance to stresses is modulated through epigenetic and chromatin pathways (Lämke and Bäurle, 2017). The acquired resistance is then retained by the plant from a few days to several weeks (Liu et al., 2022). Beside seed resistance improvement, BME compounds, found for example in AN, can trigger the biosynthesis of the hormone gibberellin A3 that functions as a signal for α-amylase enzyme gene activation. The enzyme then participates in starch hydrolysis into carbohydrates, which is used as an energy source by the seed for germination (Akazawa and Hara-Nishimura, 1985; Beck and Ziegler, 1989; Sun and Gubler, 2004; Rayorath et al., 2008b). Other compounds can also be responsible for α-amylase enzyme activation, as it was also elicited by a GA3-deprived AN extract (Rayorath et al., 2008b).
4.4 Growth, photosynthesis, and nutritional quality enhancement
The stimulation of plant growth by BME is modulated through several plant hormonal pathways, namely phenylpropanoid and flavonoid (Sujeeth et al., 2022). BME compounds are able to act as signals for the endogenous plant growth hormones auxin, cytokinin and gibberellin biosynthesis through the upregulation of their respective genes (Wally et al., 2013; Ali et al., 2019; Shukla et al., 2019; Ali et al., 2021b; Baghdadi et al., 2022). Moreover, accumulation of plant growth hormones was correlated with growth parameters improvement of tomato and bell pepper, as well as on Arabidopsis consecutive to the application of AN and Sargassum vulgare extracts (Wally et al., 2013; Ali et al., 2019; Ali et al., 2021b; Baghdadi et al., 2022). AN and Sargassum extracts applied on tomato upregulated the genes responsible for flowering, which is regarded as an index of yield (Dookie et al., 2021; Baghdadi et al., 2022). BME also elicit the upregulation of cellulose synthase-like E1, UDP-glucose 4-epimerase 1 and pectin acetylesterase 8 genes, responsible for the development and strengthening of plant cell walls (Goñi et al., 2016).
BME can improve the plant photosynthetic system by upregulating betaine aldehyde dehydrogenase and choline monooxygenase genes involved in the photosynthetic process (Xu and Leskovar, 2015). They also trigger the upregulation of glycine betaine biosynthesis genes that prevent chlorophyll degradation and delay photosynthetic activity loss, and the downregulation of chlorophyll degradation genes (Genard et al., 1991; Nair et al., 2012). Both processes resulted in increased leaf chlorophyll content and enhanced photosynthetic activity, as observed after AN extract application on spinach (Fan et al., 2013; Xu and Leskovar, 2015).
Quality and shelf-life of post-harvest food products are improved upon BME applications. Indeed, an AN commercial alkali extract elicited the upregulation of thylakoid-bound ascorbate peroxidase and monodehydroascorbate reductase genes involved in the accumulation of carotenoids, flavonoids, phenols in spinach (Fan et al., 2014; Xu and Leskovar, 2015).
4.5 Abiotic stress alleviation
Plants cultivated in open fields are frequently subjected to oxidative, hydric, saline, heat, and frost stresses. Those stresses and their impact on plant health and yield can be alleviated by BME through several metabolic pathways. For example, alginates of AN can act as signals for the accumulation of abscisic acid through the activation of its biosynthesis and signaling genes (Santaniello et al., 2017; Rasul et al., 2021). Accumulation of abscisic acid in plants is one key factor in multiple stress resilience. Furthermore, osmotic balance maintenance is a crucial parameter to ensure the plant survival, as it allows efficient water and ion exchange through cell membranes. When a plant is subjected to saline, hydric or frost stress, application of an AN lipophilic extract can trigger the accumulation of proline, an osmoprotectant, through the upregulation of proline synthesis and the downregulation of proline degradation genes (Nair et al., 2012). Similarly, the accumulation of mannitol is induced by the upregulation of genes encoding for mannitol-1-P-dehydrogenase and mannose-6-P-reductase enzymes, responsible for the reduction of fructose-6-P and mannose-6-P into mannitol-1-P, respectively, while the gene encoding for mannitol dehydrogenase, responsible for the oxidation of mannitol into carbohydrate monomers, is downregulated (Pharr et al., 1995; Stoop et al., 1996).
Oxidative stress generates ROS responsible for several damages in plants. BME application on crops under oxidative stress can upregulate the ROS-scavenging enzymes genes to inhibit ROS production and prevent harm (Fike et al., 2001; Abdel Latef et al., 2017; Goñi et al., 2018; Hussein et al., 2021). Indeed, AN extract application on oxidative-stressed Arabidopsis upregulated genes associated to ROS detoxification ascorbate-glutathione antioxidant system, and downregulated programmed cell death-associated and lipid cell death-associated genes, which suggest a reduction of damages caused by oxidative stress (Omidbakhshfard et al., 2020; Rasul et al., 2021; Staykov et al., 2021). Saline and hydric stresses are both mainly mitigated through osmotic pathways, including the accumulation of osmoprotectants. Stressed Arabidopsis treated with an AN commercial alkali extract modulated cytokinin response regulator2 and apoplastic peroxidase activities, both involved in stomatal closure (Rasul et al., 2021). Stomatal closure enhancement allows for a reduction of transpiration and an increase of water retention. AN extracts also elicited the upregulation of drought-responsive and dehydrin genes (Muñoz-Mayor et al., 2012; Goñi et al., 2018; Rasul et al., 2021), and downregulated the negative growth regulator stress responsive gene in Arabidopsis (Rasul et al., 2021). Frost stress is alleviated through the upregulation of freezing-tolerance genes, the upregulation of stomatal chloroplast proteins cryoprotection transcription factor genes and the upregulation of genes coding for enzymes involved in the synthesis of freezing tolerance compounds (Nair et al., 2012; Zamani-Babgohari et al., 2019). Protection against frost is also improved through the accumulation of several compounds, including proline and unsaturated fatty acids, involved in cell protection against freezing-induced damages, as well as carbohydrates, to increase the energy stock for continued metabolism performance. Carbohydrates accumulation is mediated through the upregulation of polysaccharide degradation genes, carbohydrate biosynthesis genes and the downregulation of sucrose degradation genes (Nair et al., 2012).
5 Conclusion and future directions
As the agricultural sector is actively looking for agrochemical alternatives to enhance crop yield and prevent abiotic stress threats, brown macroalgal biostimulants are gaining momentum. The wide range of bioactive compounds of brown macroalgae make them prime candidates for such applications. Encouraging results have already been highlighted regarding soil structure and stability, seed priming, nutrient and water uptake, root and plant growth and development, overall yield and nutritional quality, and abiotic stress alleviation. Nevertheless, the precise mode of action of extracts are still unclear, as several metabolic pathways are involved, and the extracts complex composition associated with compounds’ synergy do not allow a clear understanding of the mechanisms involved. Improvements in genomic techniques and extensive characterization of BME could allow, in the future, for a better comprehension of plant mechanisms, which in turn would allow for extract optimization to further enhance the activities. In addition, these findings will allow commercial BME producers to develop and standardize more systematically their extracts which will guarantee more homogeneous products with sufficient bioactivity to meet targeted results. Currently, brown macroalgae are still considered an underexploited resource, but are gaining more interest from scientists and industries. Indeed, their application fields are numerous, and their offshore growth neither requires water and fertilisers supply nor competes with crop production for arable lands. Brown macroalgae are a renewable resource, thanks to their high recovery rate, but harvest from wild populations must be tightly regulated to avoid biomass overexploitation and surrounding marine ecosystem disruption. In this regard, cultivation could ensure a continuous supply of algal biomass without threatening wild communities.
Author contributions
Conceptualization: NK, VS-P, SC, FH, and JB. Methodology and investigation: NK, VS-P, SC, FH, and JB. Bibliographic research and writing of the original draft: NK. Supervision: VS-P, SC, FH, and JB. Reviewing and editing: NK, VS-P, SC, FH and JB. Funding acquisition: VS-P, FH, and JB. All authors contributed to the article and approved the submitted version.
Funding
This research was funded through the project SAVE-C (Fundamental and applied research, Grant No: (ANR-19-SARG-0008) which is an International Partnership funded by the ANR under the call SARGASSUM Program 2019.
Acknowledgments
All authors thank Reviewers for their constructive criticisms which helped to improve this manuscript. This review is part of a PhD project carried out by the first author at the Laboratoire des Sciences de l’Environnement Marin (LEMAR UMR6539) in the IUEM (UBO) and the company ALGAIA, under the supervision of the 4 others co-authors.
Conflict of interest
Authors NK, FH, JB were employed by company ALGAIA.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbas M., Anwar J., Zafar-Ul-Hye M., Khan R. I., Saleem M., Rahi A. A., et al. (2020). Effect of seaweed extract on productivity and quality attributes of four onion cultivars. Horticulturae 6, 28–42. doi: 10.3390/horticulturae6020028
Abdel Latef A. A. H., Srivastava A. K., Saber H., Alwaleed E. A., Tran L. S. P. (2017). Sargassum muticum and Jania rubens regulate amino acid metabolism to improve growth and alleviate salinity in chickpea. Sci. Rep. 7, 1–12. doi: 10.1038/s41598-017-07692-w
Abdel-Mawgoud A. M., Tantaway A., Hafez M. M., Habib H. A. M. (2010). Seaweed extract improves growth, yield and quality of different watermelon hybrids. Res. J. Agric. Biol. Sci. 6, 161–168.
Abetz P., Young C. L. (1983). The effect of seaweed extract sprays derived from Ascophyllum nodosum on lettuce and cauliflower crops. Botanica Marina 26, 487–492. doi: 10.1515/botm.1983.26.10.487
Akazawa T., Hara-Nishimura I. (1985). Topographic aspects of biosynthesis, extracellular secretion, and intracellular storage of proteins in plant cells. Annu. Rev. Plant Physiol. 36, 441–472. doi: 10.1146/annurev.pp.36.060185.002301
Alam M. Z., Braun G., Norrie J., Hodges D. M. (2013). Effect of Ascophyllum extract application on plant growth, fruit yield and soil microbial communities of strawberry. Can. J. Plant Sci. 93, 23–36. doi: 10.4141/CJPS2011-260
Alam M. Z., Braun G., Norrie J., Hodges D. M. (2014). Ascophyllum extract application can promote plant growth and root yield in carrot associated with increased root-zone soil microbial activity. Can. J. Plant Sci. 94, 337–348. doi: 10.4141/CJPS2013-135
Aldworth S. J., van Staden J. (1987). The effect of seaweed concentrate on seedling transplants. South Afr. J. Bot. 53, 187–189. doi: 10.1016/s0254-6299(16)31428-4
Alejandro S., Höller S., Meier B., Peiter E. (2020). Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00300
Al-Ghamdi A. A., Elansary H. O. (2018). Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Biochem. 129, 273–284. doi: 10.1016/j.plaphy.2018.06.008
Alghazeer R., El Fatah H., Azwai S., Elghmasi S., Sidati M., El Fituri A., et al. (2022). Nutritional and nonnutritional content of underexploited edible seaweeds. Aquac. Nutr. 2022, 1–8. doi: 10.1155/2022/8422414
Ali O., Ramsubhag A., Jayaraman J. (2018). Ascophyllum nodosum (Linnaeus ) le jolis seaweed extract improves seed germination in tomato and sweet pepper under NaCl-induced salt stress. Trop. Agric. 95, 141–148.
Ali O., Ramsubhag A., Jayaraman J. (2019). Biostimulatory activities of Ascophyllum nodosum extract in tomato and sweet pepper crops in a tropical environment. PLoS One 14, e0216710. doi: 10.1371/journal.pone.0216710
Ali O., Ramsubhag A., Jayaraman J. (2021a). Biostimulant properties of seaweed extracts in plants: Implications towards sustainable crop production. Plants 10, 1–27. doi: 10.3390/plants10030531
Ali O., Ramsubhag A., Jayaraman J. (2021b). Phytoelicitor activity of Sargassum vulgare and Acanthophora spicifera extracts and their prospects for use in vegetable crops for sustainable crop production. J. Appl. Phycol. 33, 639–651. doi: 10.1007/s10811-020-02309-8
Amador-Castro F., García-Cayuela T., Alper H. S., Rodriguez-Martinez V., Carrillo-Nieves D. (2021). Valorization of pelagic Sargassum biomass into sustainable applications: Current trends and challenges. J. Environ. Manage. 283, 112013–112025. doi: 10.1016/j.jenvman.2021.112013
Amaral Carvalho M. E., de Camargo e Castro P. R., Gallo L. A., de Castro Ferraz Junior M. V. (2014). Seaweed extract provides development and production of wheat. Rev. Agrarian 7, 166–170. doi: 10.5281/zenodo.51607
Arthur G. D., Stirk W. A., van Staden J. (2003). Effect of a seaweed concentrate on the growth and yield of three varieties of Capsicum annuum. South Afr. J. Bot. 69, 207–211. doi: 10.1016/S0254-6299(15)30348-3
Atzmon N., Van Staden J. (1994). The effect of seaweed concentrate on the growth of Pinus pinea seedlings. New Forests 8, 279–288. doi: 10.1007/BF00025373
Baffes J., Chian Koh W. (2022)Fertilizer prices expected to remain higher for longer. In: World bank blogs. Available at: https://blogs.worldbank.org/opendata/fertilizer-prices-expected-remain-higher-longer (Accessed January 18, 2023).
Baghdadi A., Della Lucia M. C., Borella M., Bertoldo G., Ravi S., Zegada-Lizarazu W., et al. (2022). A dual-omics approach for profiling plant responses to biostimulant applications under controlled and field conditions. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.983772
Basak A. (2008). Effect of preharvest treatment with seaweed products, kelpak® and goëmar BM 86®, on fruit quality in apple. Int. J. Fruit Sci. 8, 1–14. doi: 10.1080/15538360802365251
Beck E., Ziegler P. (1989). Biosynthesis and degradation of starch in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 95–117. doi: 10.1146/annurev.pp.40.060189.000523
Beckett R. P., Mathegka A. D. M., van Staden J. (1994). Effect of seaweed concentrate on yield of nutrient-stressed tepary bean (Phaseolus acutifolius gray). J. Appl. Phycol. 6, 429–430. doi: 10.1007/BF02182161
Beckett R. P., van Staden J. (1989). The effect of seaweed concentrate on the growth and yield of potassium stressed wheat. Plant Soil 116, 29–36. doi: 10.1007/BF02327254
Beckett R. P., van Staden J. (1990). The effect of seaweed concentrate on the uptake of foliar-applied Cu, Mn and zn by tomato seedlings. South Afr. J. Bot. 56, 389–392. doi: 10.1016/s0254-6299(16)31068-7
Billard V., Etienne P., Jannin L., Garnica M., Cruz F., Garcia-Mina J. M., et al. (2014). Two biostimulants derived from algae or humic acid induce similar responses in the mineral content and gene expression of winter oilseed rape (Brassica napus l.). J. Plant Growth Regul. 33, 305–316. doi: 10.1007/s00344-013-9372-2
Blunden G., Cripps A. L., Gordon S. M., Mason T. G., Turnei C. H. (1986). The characterisation and quantitative estimation of betaines in commercial seaweed extracts. Botanica Marina 29, 155–160. doi: 10.1515/botm.1986.29.2.155
Blunden G., Jenkins T., Liu Y. W. (1996). Enhanced leaf chlorophyll levels in plants treated with seaweed extract. J. Appl. Phycol. 8, 535–543. doi: 10.1007/BF02186333
Bradáčová K., Weber N. F., Morad-Talab N., Asim M., Imran M., Weinmann M., et al. (2016). Micronutrients (Zn/Mn), seaweed extracts, and plant growth-promoting bacteria as cold-stress protectants in maize. Chem. Biol. Technol. Agric. 3, 1–10. doi: 10.1186/s40538-016-0069-1
Briceño-Domínguez D., Hernández-Carmona G., Moyo M., Stirk W., van Staden J. (2014). Plant growth promoting activity of seaweed liquid extracts produced from Macrocystis pyrifera under different pH and temperature conditions. J. Appl. Phycol. 26, 2203–2210. doi: 10.1007/s10811-014-0237-2
Burchett S. (2000). The mechanism of action of liquid seaweed extracts in the manipulation of frost resistance in winter barley (Hordeum vulgare. l). Univ. Plymouth Res. Theses, 1–242.
Caradonia F., Battaglia V., Righi L., Pascali G., La Torre A. (2019). Plant biostimulant regulatory framework: prospects in Europe and current situation at international level. J. Plant Growth Regul. 38, 438–448. doi: 10.1007/s00344-018-9853-4
Caradonia F., Ronga D., Tava A., Francia E. (2022). Plant biostimulants in sustainable potato production: An overview. Potato Res. 65, 83–104. doi: 10.1007/s11540-021-09510-3
Carrasco-Gil S., Hernandez-Apaolaza L., Lucena J. J. (2018). Effect of several commercial seaweed extracts in the mitigation of iron chlorosis of tomato plants (Solanum lycopersicum l.). Plant Growth Regul. 86, 401–411. doi: 10.1007/s10725-018-0438-9
Chandía N. P., Matsuhiro B. (2008). Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 42, 235–240. doi: 10.1016/j.ijbiomac.2007.10.023
Chandía N. P., Matsuhiro B., Mejías E., Moenne A. (2004). Alginic acids in Lessonia vadosa: Partial hydrolysis and elicitor properties of the polymannuronic acid fraction. J. Appl. Phycol. 16, 127–133. doi: 10.1023/B:JAPH.0000044778.44193.a8
Chen D., Huang Y., Shen D., Zhou W., AO J., Jiang Y., et al. (2019). Effects of seaweed extracts on promoting growth and improving stress resistance in sugarcane. Asian Agric. Res. 5, 69–76. doi: 10.22004/AG.ECON.292416
Chouliaras V., Gerascapoulos D., Lionakis S. (1997). Effects of seaweed extract on fruit growth, weight and maturation of “hayward” kiwifruit. Acta Hortic. 2, 485–492. doi: 10.17660/actahortic.1997.444.74
Chouliaras V., Tasioula M., Chatzissavvidis C., Therios I., Tsabolatidou E. (2009). The effects of a seaweed extract in addition to nitrogen and boron fertilization on productivity, fruit maturation, leaf nutritional status and oil quality of the olive (Olea europaea l.) cultivar koroneiki. J. Sci. Food Agric. 89, 984–988. doi: 10.1002/jsfa.3543
Circuncisão A. R., Catarino M. D., Cardoso S. M., Silva A. M. S. (2018). Minerals from macroalgae origin: Health benefits and risks for consumers. Mar. Drugs 16, 400–430. doi: 10.3390/md16110400
Colapietra M., Alexander A. (2006). Effect of foliar fertilization on yield and quality of table grapes. Acta Hortic. 721, 213–218. doi: 10.17660/ActaHortic.2006.721.28
Colla G., Nardi S., Cardarelli M., Ertani A., Lucini L., Canaguier R., et al. (2015). Protein hydrolysates as biostimulants in horticulture. Scientia Hortic. 196, 28–38. doi: 10.1016/j.scienta.2015.08.037
Connan S., Goulard F., Stiger V., Deslandes E., Ar Gall E. (2004). Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Botanica Marina 47, 410–416. doi: 10.1515/BOT.2004.057
Connan S., Stengel D. B. (2011). Impacts of ambient salinity and copper on brown algae: 2. interactive effects on phenolic pool and assessment of metal binding capacity of phlorotannin. Aquat. Toxicol. 104, 1–13. doi: 10.1016/j.aquatox.2011.03.016
Courbet G., Gallardo K., Vigani G., Brunel-Muguet S., Trouverie J., Salon C., et al. (2019). Disentangling the complexity and diversity of crosstalk between sulfur and other mineral nutrients in cultivated plants. J. Exp. Bot. 70, 4183–4196. doi: 10.1093/jxb/erz214
Critchley A. T., Critchley J. S. C., Norrie J., Gupta S., van Staden J. (2021). “Perspectives on the global biostimulant market: Applications, volumes, and values 2016 Data and projections to 2022,” in Biostimulants for crops from seed germination to plant development: A practical approach. Eds. Gupta S., Van Staden J. (Cambridge, Massachusetts: Academic Press), 289–296. doi: 10.1016/B978-0-12-823048-0.00012-5
Crouch I. J., Beckett R. P., van Staden J. (1990). Effect of seaweed concentrate on the growth and mineral nutrition of nutrient-stressed lettuce. J. Appl. Phycol. 2, 269–272. doi: 10.1007/BF02179784
Crouch I. J., van Staden J. (1991). Evidence for rooting factors in a seaweed concentrated prepared from Ecklonia maxima. J. Plant Physiol. 137, 319–322. doi: 10.1016/S0176-1617(11)80138-0
Crouch I. J., van Staden J. (1992). Effect of seaweed concentrate on the establishment and yield of greenhouse tomato plants. J. Appl. Phycol. 4, 291–296. doi: 10.1007/BF02185785
Dalal A., Bourstein R., Haish N., Shenhar I., Wallach R., Moshelion M. (2019). Dynamic physiological phenotyping of drought-stressed pepper plants treated with “productivity-enhancing” and “survivability-enhancing” biostimulants. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00905
da Silveira L. C. I., Mattos P., Mógor Á.F., Daros E., de Oliveira Bettini M., Norrie J. (2015). Effect of kelp extract on sugarcane plantlets biomass accumulation. Idesia 33, 31–33. doi: 10.4067/S0718-34292015000300005
Dell’Aversana E., Cirillo V., Van Oosten M. J., Di Stasio E., Saiano K., Woodrow P., et al. (2021). Ascophyllum nodosum based extracts counteract salinity stress in tomato by remodeling leaf nitrogen metabolism. Plants 10, 1044–1066. doi: 10.3390/plants10061044
De Lucia B., Vecchietti L. (2012). Type of bio-stimulant and application method effects on stem quality and root system growth in L.A. lily. Eur. J. Hortic. Sci. 77, 10–15.
Demir N., Dural B., Yildirim K. (2006). Effect of seaweed suspensions on seed germination of tomato, pepper and aubergine. J. Biol. Sci. 6, 1130–1133. doi: 10.3923/jbs.2006.1130.1133
Deolu-Ajayi A., van der Meer I., van der Werf A., Karlova R. (2021). The power of seaweeds as plant biostimulants to boost crop production under abiotic stress. Plant Cell Environ. 45, 2537–2553. doi: 10.1111/PCE.14391
de Sousa A. M., Ayub R. A., Viencz T., Botelho R. V. (2019). Fruit set and yield of apple trees cv. gala treated with seaweed extract of Ascophyllum nodosum and thidiazuron. Rev. Bras. Fruticultura 41, 72–84. doi: 10.1590/0100-29452019072
Di Filippo-Herrera D. A., Muñoz-Ochoa M., Hernández-Herrera R. M., Hernández-Carmona G. (2019). Biostimulant activity of individual and blended seaweed extracts on the germination and growth of the mung bean. J. Appl. Phycol. 31, 2025–2037. doi: 10.1007/s10811-018-1680-2
Di Stasio E., Cirillo V., Raimondi G., Giordano M., Esposito M., Maggio A. (2020). Osmo-priming with seaweed extracts enhances yield of salt-stressed tomato plants. Agronomy 10, 1559–1572. doi: 10.3390/agronomy10101559
Di Stasio E., Raimondi G., Van Oosten M., Maggio A. (2017a). “Algal derivatives may protect crops from residual soil salinity: a case study on a tomato-wheat rotation,” in European Geosciences union general assembly (Vienna, Austria:Geophysical Research Abstracts), 19408.
Di Stasio E., Rouphael Y., Colla G., Raimondi G., Giordano M., Pannico A., et al. (2017b). The influence of Ecklonia maxima seaweed extract on growth, photosynthetic activity and mineral composition of Brassica rapa l. subsp. sylvestris under nutrient stress conditions. Eur. J. Hortic. Sci. 82, 286–293. doi: 10.17660/eJHS.2017/82.6.3
Di Stasio E., Van Oosten M. J., Silletti S., Raimondi G., Dell’Aversana E., Carillo P., et al. (2018). Ascophyllum nodosum-based algal extracts act as enhancers of growth, fruit quality, and adaptation to stress in salinized tomato plants. J. Appl. Phycol. 30, 2675–2686. doi: 10.1007/s10811-018-1439-9
Dookie M., Ali O., Ramsubhag A., Jayaraman J. (2021). Flowering gene regulation in tomato plants treated with brown seaweed extracts. Scientia Hortic. 276, 109715–109723. doi: 10.1016/j.scienta.2020.109715
D’Oria A., Courbet G., Lornac A., Pluchon S., Arkoun M., Maillard A., et al. (2021). Specificity and plasticity of the functional ionome of Brassica napus and Triticum aestivum exposed to micronutrient or beneficial nutrient deprivation and predictive sensitivity of the ionomic signatures. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.641678
du Jardin P. (2015). Plant biostimulants: Definition, concept, main categories and regulation. Scientia Hortic. 196, 3–14. doi: 10.1016/j.scienta.2015.09.021
Dziugieł T., Wadas W. (2020). Possibility of increasing early crop potato yield with foliar application of seaweed extracts and humic acids. J. Cent. Eur. Agric. 21, 300–310. doi: 10.5513/JCEA01/21.2.2576
EBIC (2021) The European biostimulants industry. Available at: https://biostimulants.eu/ (Accessed November 25, 2022).
EBIC (2023) Recent insights into the mode of action of seaweed-based plant biostimulants. Available at: https://biostimulants.eu/publications/ (Accessed February 15, 2023).
El Boukhari M. E. M., Barakate M., Bouhia Y., Lyamlouli K. (2020). Trends in seaweed extract based biostimulants : Manufacturing process and beneficial effect on soil-plant systems. Plants 9, 1–23. doi: 10.3390/plants9030359
El-Metwally I. M., Sadak M. S., Saudy H. S. (2022). Stimulation effects of glutamic and 5-aminolevulinic acids on photosynthetic pigments, physio-biochemical constituents, antioxidant activity, and yield of peanut. Gesunde Pflanzen 74, 915–924. doi: 10.1007/s10343-022-00663-w
El-Miniawy S. M., Ragab M. E., Youssef S. M., Metwall A. A. (2016). Influence of foliar spraying of seaweed extract on growth, yield and quality of strawberry plants. J. Appl. Sci. Res. 10, 88–94.
El-Yazied A., El-Gizawy A. M., Ragab M. I., Hamed E. S. (2012). Effect of seaweed extract and compost treatments on growth, yield and quality of snap bean. J. Am. Sci. 8, 1545–1003.
Ertani A., Francioso O., Tinti A., Schiavon M., Pizzeghello D., Nardi S. (2018). Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays l. using a combination of chemical, biochemical and morphological approaches. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00428
European Commission (2021) Report from the commission to the council and the European parliament on the implementation of council directive 91/676/EEC concerning the protection of waters against pollution caused by nitrates from agricultural sources based on member state reports fo (Brussels, Belgium). Available at: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=COM%3A2021%3A1000%3AFIN&qid=1633953687154 (Accessed February 15, 2023).
European Environment Agency (2017). “Climate change, impacts and vulnerability in Europe 2016,” in An indicator-based report (Copenhagen, Denmark). doi: 10.2800/534806
European Parliament and Council (2019)Regulation (EU) 2019/1009 of the European parliament and of the council of 5 June 2019 laying down rules on the making available on the market of EU fertilising products and amending regulations (EC) no 1069/2009 and (EC) no 1107/2009 and repealing regula. In: Official journal of the European union. Available at: https://eur-lex.europa.eu/eli/reg/2019/1009/oj (Accessed September 11, 2022).
Eyras M. C., Rostagno C. M., Defossé G. E. (1998). Biological evaluation of seaweed composting. Compost Sci. Utilization 6, 74–81. doi: 10.1080/1065657X.1998.10701943
Fan D., Hodges D. M., Critchley A. T., Prithiviraj B. (2013). A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 44, 1873–1884. doi: 10.1080/00103624.2013.790404
Fan D., Hodges D. M., Zhang J., Kirby C. W., Ji X., Locke S. J., et al. (2011). Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea l.) which protects Caenorhabditis elegans against oxidative and thermal stress. Food Chem. 124, 195–202. doi: 10.1016/j.foodchem.2010.06.008
Fan D., Kandasamy S., Hodges D. M., Critchley A. T., Prithiviraj B. (2014). Pre-harvest treatment of spinach with Ascophyllum nodosum extract improves post-harvest storage and quality. Scientia Hortic. 170, 70–74. doi: 10.1016/j.scienta.2014.02.038
FAO (2021). The impact of disasters and crises on agriculture and food security: 2021 (Rome, Italy:FAO). doi: 10.4060/cb3673en
Featonby-Smith B. C., van Staden J. (1983). The effect of seaweed concentrate and fertilizer on the growth of Beta vulgaris. Z. für Pflanzenphysiologie 112, 155–162. doi: 10.1016/s0044-328x(83)80030-0
Featonby-Smith B. C., van Staden J. (1987a). Effect of seaweed concentrate on yield and seed quality of Arachis hypogaea. South Afr. J. Bot. 53, 190–193. doi: 10.1016/s0254-6299(16)31429-6
Featonby-Smith B. C., van Staden J. (1987b). Effects of seaweed concentrate on grain yield in barley. South Afr. J. Bot. 53, 125–128. doi: 10.1016/s0254-6299(16)31446-6
Fei H., Crouse M., Papadopoulos Y., Vessey J. K. (2017). Enhancing the productivity of hybrid poplar (Populus × hybrid) and switchgrass (Panicum virgatum l.) by the application of beneficial soil microbes and a seaweed extract. Biomass Bioenergy 107, 122–134. doi: 10.1016/j.biombioe.2017.09.022
Fike J. H., Allen V. G., Schmidt R. E., Zhang X., Fontenot J. P., Bagley C. P., et al. (2001). Tasco-forage: I. influence of a seaweed extract on antioxidant activity in tall fescue and in ruminants. J. Anim. Sci. 79, 1011–1021. doi: 10.2527/2001.7941011x
Finnie J. F., van Staden J. (1985). Effect of seaweed concentrate and applied hormones on in vitro cultured tomato roots. J. Plant Physiol. 120, 215–222. doi: 10.1016/S0176-1617(85)80108-5
Fleurence J. (1999). Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 10, 25–28. doi: 10.1016/S0924-2244(99)00015-1
Fornes F., Sánchez-Perales M., Guardiola J. L. (1995). Effect of a seaweed extract on citrus fruit maturation. Acta Hortic. 379, 75–82. doi: 10.17660/actahortic.1995.379.6
Fornes F., Sánchez-Perales M., Guardiola J. L. (2002). Effect of a seaweed extract on the productivity of “de nules” clementine mandarin and navelina orange. Botanica Marina 45, 486–489. doi: 10.1515/BOT.2002.051
Franzoni G., Cocetta G., Prinsi B., Ferrante A., Espen L. (2022). Biostimulants on crops: their impact under abiotic stress conditions. Horticulturae 8, 189–208. doi: 10.3390/horticulturae8030189
Frioni T., Sabbatini P., Tombesi S., Norrie J., Poni S., Gatti M., et al. (2018). Effects of a biostimulant derived from the brown seaweed Ascophyllum nodosum on ripening dynamics and fruit quality of grapevines. Scientia Hortic. 232, 97–106. doi: 10.1016/j.scienta.2017.12.054
Gajc-Wolska J., Spiewski T., Grabowska A. (2012). The effect of seaweed extracts on the yield and quality parameters of broccoli (Brassica oleracea var. cymosa l.) in open field production. Acta Hortic. 1009, 83–90. doi: 10.17660/actahortic.2013.1009.9
Genard H., Lesaos J., Billard J. P., Tremolieres A., Boucaud J. (1991). Effect of salinity on lipid composition, glycine betaine content and photosynthetic activity in chloroplasts of Suaeda maritima. Plant Physiol. Biochem. 29, 421–427.
Gomathi R., Kohila S., Ramachandiran K. (2017). Evaluating the effect of seaweed formulations on the quality and yield of sugarcane. Madras Agric. J. 104, 161–165. doi: 10.29321/maj.04.000423
Goñi O., Fort A., Quille P., McKeown P. C., Spillane C., O’Connell S. (2016). Comparative transcriptome analysis of two ascophyllum nodosum extract biostimulants: same seaweed but different. J. Agric. Food Chem. 64, 2980–2989. doi: 10.1021/acs.jafc.6b00621
Goñi O., Łangowski Ł., Feeney E., Quille P., O’Connell S. (2021). Reducing nitrogen input in barley crops while maintaining yields using an engineered biostimulant derived from ascophyllum nodosum to enhance nitrogen use efficiency. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.664682
Goñi O., Quille P., O’Connell S. (2018). Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 126, 63–73. doi: 10.1016/j.plaphy.2018.02.024
Guiry M. D., Guiry G. M. (2023) AlgaeBase. world-wide electronic publication, national university of irlande, Galway. Available at: http://www.algaebase.org (Accessed March 3, 2023).
Haichar F. E. Z., Marol C., Berge O., Rangel-Castro J. I., Prosser J. I., Balesdent J., et al. (2008). Plant host habitat and root exudates shape soil bacterial community structure. ISME J. 2, 1221–1230. doi: 10.1038/ismej.2008.80
Hashem H. A., Mansour H. A., El-Khawas S. A., Hassanein R. A. (2019). The potentiality of marine macro-algae as bio-fertilizers to improve the productivity and salt stress tolerance of canola (Brassica napus l.) plants. Agronomy 9, 146–163. doi: 10.3390/agronomy9030146
Hidangmayum A., Sharma R. (2017). Effect of different concentration of commercial seaweed liquid extract of Ascophyllum nodosum on germination of onion (Allium cepa l.). Int. J. Sci. Res. 6, 1488–1491. doi: 10.21275/art20175686
Huang X. Y., Hu D. W., Zhao F. J. (2022). Molybdenum: More than an essential element. J. Exp. Bot. 73, 1766–1774. doi: 10.1093/jxb/erab534
Hussein M. H., Eltanahy E., Al Bakry A. F., Elsafty N., Elshamy M. M. (2021). Seaweed extracts as prospective plant growth bio-stimulant and salinity stress alleviator for Vigna sinensis and Zea mays. J. Appl. Phycol. 33, 1273–1291. doi: 10.1007/s10811-020-02330-x
Illera-Vives M., López-Fabal A., López-Mosquera M. E., Ribeiro H. M. (2015). Mineralization dynamics in soil fertilized with seaweed–fish waste compost. J. Sci. Food Agric. 95, 3047–3054. doi: 10.1002/JSFA.7207
Ishii T., Aikawa J., Kirino S., Kitabayashi H., Matsumoto I., Kadoya K. (2000). Effects of alginate oligosaccharide and polyamines on hyphal growth of vesicular-arbuscular mycorrhizal fungi and their infectivity of citrus roots. Proceeding Int. Soc. Citriculture 73, 529–533. doi: 10.2503/JJSHS.73.529
Jannin L., Arkoun M., Etienne P., Laîné P., Goux D., Garnica M., et al. (2013). Brassica napus growth is promoted by Ascophyllum nodosum (L.) le jolis seaweed extract: Microarray analysis and physiological characterization of n, c, and s metabolisms. J. Plant Growth Regul. 32, 31–52. doi: 10.1007/s00344-012-9273-9
Jeannin I., Lescure J. C., Morot-Gaudry J. F. (1991). The effects of aqueous seaweed sprays on the growth of maize. Botanica Marina 34, 469–474. doi: 10.1515/botm.1991.34.6.469
Kalaivanan C., Venkatesalu V. (2012). Utilization of seaweed Sargassum myriocystum extracts as a stimulant of seedlings of Vigna mungo (L.) hepper. Spanish J. Agric. Res. 10, 466–470. doi: 10.5424/sjar/2012102-507-10
Kapur B., Sarıdaş M. A., Çeliktopuz E., Kafkas E., Paydaş Kargı S. (2018). Health and taste related compounds in strawberries under various irrigation regimes and bio-stimulant application. Food Chem. 263, 67–73. doi: 10.1016/j.foodchem.2018.04.108
Kasim W. A., Hamada E. A. M., El-Din N. G. S., Eskander S. (2015). Influence of seaweed extracts on the growth, some metabolic activities and yield of wheat grown under drought stress. Int. J. Agron. Agric. Res. 7, 173–189.
Khan A. S., Ahmad B., Jaskani M. J., Ahmad R., Malik A. U. (2012). Foliar application of mixture of amino acids and seaweed (Ascophyllum nodosum) extract improve growth and physicochemical properties of grapes. Int. J. Agric. Biol. 14, 383–388.
Khan W., Hiltz D., Critchley A. T., Prithiviraj B. (2011). Bioassay to detect Ascophyllum nodosum extract-induced cytokinin-like activity in Arabidopsis thaliana. J. Appl. Phycol. 23, 409–414. doi: 10.1007/s10811-010-9583-x
Khan W., Rayirath U. P., Subramanian S., Jithesh M. N., Rayorath P., Hodges D. M., et al. (2009). Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 28, 386–399. doi: 10.1007/s00344-009-9103-x
Kholssi R., Lougraimzi H., Grina F., Lorentz J. F., Silva I., Castaño-Sánchez O., et al. (2022). Green agriculture: A review of the application of micro- and macroalgae and their impact on crop production on soil quality. J. Soil Sci. Plant Nutr. 22, 1–15. doi: 10.1007/s42729-022-00944-3
Kolbert Z., Cuypers A., Verbruggen N. (2022). Essential trace metals: Micronutrients with large impact. J. Exp. Bot. 73, 1685–1687. doi: 10.1093/jxb/erac025
Kowalski B., Jäger A. K., Van Staden J. (1999). The effect of a seaweed concentrate on the in vitro growth and acclimatization of potato plantlets. Potato Res. 42, 131–139. doi: 10.1007/bf02358403
Kulkarni M. G., Rengasamy K. R. R., Pendota S. C., Gruz J., Plačková L., Novák O., et al. (2019). Bioactive molecules derived from smoke and seaweed Ecklonia maxima showing phytohormone-like activity in Spinacia oleracea l. New Biotechnol. 48, 83–89. doi: 10.1016/j.nbt.2018.08.004
Kumar G., Sahoo D. (2011). Effect of seaweed liquid extract on growth and yield of Triticum aestivum var. pusa gold. J. Appl. Phycol. 23, 251–255. doi: 10.1007/s10811-011-9660-9
Kuwada K., Kuramoto M., Utamura M., Matsushita I., Shibata Y., Ishii T. (2005). Effect of mannitol from Laminaria japonica, other sugar alcohols, and marine alga polysaccharides on in vitro hyphal growth of Gigaspora margarita and root colonization of trifoliate orange. Plant Soil 276, 279–286. doi: 10.1007/s11104-005-4985-2
Lalegerie F., Gager L., Stiger-Pouvreau V., Connan S. (2020). The stressful life of red and brown seaweeds on the temperate intertidal zone: effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. Adv. Botanical Res. 95, 247–287. doi: 10.1016/bs.abr.2019.11.007
Lämke J., Bäurle I. (2017). Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 18, 1–11. doi: 10.1186/s13059-017-1263-6
Laporte D., Vera J., Chandía N. P., Zúñiga E. A., Matsuhiro B., Moenne A. (2007). Structurally unrelated algal oligosaccharides differentially stimulate growth and defense against tobacco mosaic virus in tobacco plants. J. Appl. Phycol. 19, 79–88. doi: 10.1007/s10811-006-9114-y
Larsen B., Salem D. M. S. A., Sallam M. A. E., Mishrikey M. M., Beltagy A. I. (2003). Characterization of the alginates from algae harvested at the Egyptian red Sea coast. Carbohydr. Res. 338, 2325–2336. doi: 10.1016/S0008-6215(03)00378-1
Lešková A., Javot H., Giehl R. F. H. (2022). Metal crossroads in plants: modulation of nutrient acquisition and root development by essential trace metals. J. Exp. Bot. 73, 1751–1765. doi: 10.1093/jxb/erab483
Liu H., Able A. J., Able J. A. (2022). Priming crops for the future: rewiring stress memory. Trends Plant Sci. 27, 699–716. doi: 10.1016/j.tplants.2021.11.015
Lola-Luz T., Hennequart F., Gaffney M. (2012). “Effect of a novel cold process seaweed extract on the germination of vegetable seeds and flowering of cyclamen,” in Agricultural Forum 2012, (Tullamore, Ireland:Agricultural Forum) Tullamore.
Lola-Luz T., Hennequart F., Gaffney M. (2014a). Effect on health promoting phytochemicals following seaweed application, in potato and onion crops grown under a low input agricultural system. Scientia Hortic. 170, 224–227. doi: 10.1016/j.scienta.2014.03.022
Lola-Luz T., Hennequart F., Gaffney M. (2014b). Effect on yield, total phenolic, total flavonoid and total isothiocyanate content of two broccoli cultivars (Brassica oleraceae var. italica) following the application of a commercial brown seaweed extract (Ascophyllum nodosum). Agric. Food Sci. 23, 28–37. doi: 10.23986/afsci.8832
Loyola N., Muñoz C. (2009). Effect of the biostimulant foliar addition of marine algae on “O’Neal” production. Acta Hortic. 810, 709–722. doi: 10.17660/ActaHortic.2009.810.94
MacDonald J. E., Hacking J., Weng Y., Norrie J. (2012). Root growth of containerized lodgepole pine seedlings in response to Ascophyllum nodosum extract application during nursery culture. Can. J. Plant Sci. 92, 1207–1212. doi: 10.4141/CJPS2011-279
Mahmoud S. H., Salama D. M., El-Tanahy A. M. M., Abd El-Samad E. H. (2019). Utilization of seaweed (Sargassum vulgare) extract to enhance growth, yield and nutritional quality of red radish plants. Ann. Agric. Sci. 64, 167–175. doi: 10.1016/j.aoas.2019.11.002
Malaguti D., Rombolà A. D., Gerin M., Simoni G., Tagliavini M., Marangoni B. (2002). Effect of seaweed extracts-based leaf sprays on the mineral status, yield and fruit quality of apple. Acta Hortic. 594, 357–359. doi: 10.17660/ActaHortic.2002.594.44
Mandal A., Sarkar B., Mandal S., Vithanage M., Patra A. K., Manna M. C. (2020). “Chapter 7 - impact of agrochemicals on soil health,” in Agrochemicals detection, treatment and remediation, (Oxford, United Kingdom:Butterworth-Heinemann) 161–187. doi: 10.1016/b978-0-08-103017-2.00007-6
Mansori M., Chernane H., Latique S., Benaliat A., Hsissou D., El Kaoua M. (2015). Seaweed extract effect on water deficit and antioxidative mechanisms in bean plants (Phaseolus vulgaris l.). J. Appl. Phycol. 27, 1689–1698. doi: 10.1007/s10811-014-0455-7
Martynenko A., Shotton K., Astatkie T., Petrash G., Fowler C., Neily W., et al. (2016). Thermal imaging of soybean response to drought stress: the effect of Ascophyllum nodosum seaweed extract. SpringerPlus 5, 1–14. doi: 10.1186/s40064-016-3019-2
Masondo N. A., Kulkarni M. G., Finnie J. F., van Staden J. (2018). Influence of biostimulants-seed-priming on Ceratotheca triloba germination and seedling growth under low temperatures, low osmotic potential and salinity stress. Ecotoxicology Environ. Saf. 147, 43–48. doi: 10.1016/j.ecoenv.2017.08.017
Mattner S. W., Milinkovic M., Arioli T. (2018). Increased growth response of strawberry roots to a commercial extract from Durvillaea potatorum and Ascophyllum nodosum. J. Appl. Phycology 30, 2943–2951. doi: 10.1007/s10811-017-1387-9
Matysiak K., Kaczmarek S., Krawczyk R. (2011). Influence of seaweed extracts and mixture of humic and fulvic acids on germination and growth of Zea mays l. Acta Scientiarum Polonorum 10, 33–45.
Mohd Fauziee N. A., Chang L. S., Wan Mustapha W. A., Md Nor A. R., Lim S. J. (2021). Functional polysaccharides of fucoidan, laminaran and alginate from Malaysian brown seaweeds (Sargassum polycystum, Turbinaria ornata and Padina boryana). Int. J. Biol. Macromol. 167, 1135–1145. doi: 10.1016/j.ijbiomac.2020.11.067
Mola I., Cozzolino E., Ottaiano L., Giordano M., Rouphael Y., Colla G., et al. (2019). Effect of vegetal- and seaweed extract-based biostimulants on agronomical and leaf quality traits of plastic tunnel-grown baby lettuce under four regimes of nitrogen fertilization. Agronomy 9, 571–586. doi: 10.3390/agronomy9100571
Möller M., Smith M. L. (1999). The effects of priming treatments using seaweed suspensions on the water sensitivity of barley (Hordeum vulgare l.) caryopses. Ann. Appl. Biol. 135, 515–521. doi: 10.1111/j.1744-7348.1999.tb00882.x
Moolla A. (2022). “Key trends in biostimulants, a Kline presentation,” in ACI European biostimulant summit (Seville, Spain:Active Communications International).
Muñoz-Mayor A., Pineda B., Garcia-Abellán J. O., Antón T., Garcia-Sogo B., Sanchez-Bel P., et al. (2012). Overexpression of dehydrin tas14 gene improves the osmotic stress imposed by drought and salinity in tomato. J. Plant Physiol. 169, 459–468. doi: 10.1016/j.jplph.2011.11.018
Murgia I., Marzorati F., Vigani G., Morandini P. (2022). Plant iron nutrition: The long road from soil to seeds. J. Exp. Bot. 73, 1809–1824. doi: 10.1093/jxb/erab531
Murtic S., Oljaca R., Murtic M. S., Vranac A., Koleska I., Karic L. (2018). Effects of seaweed extract on the growth, yield and quality of cherry tomato under different growth conditions. Acta Agriculturae Slovenica 111, 315–325. doi: 10.14720/aas.2018.111.2.07
Muthuramalingam P., Shin H., Adarshan S., Jeyasri R., Priya A., Chen J. T., et al. (2022). Molecular insights into freezing stress in peach based on multi-omics and biotechnology: An overview. Plants 11, 812–827. doi: 10.3390/plants11060812
Mzibra A., Aasfar A., Benhima R., Khouloud M., Boulif R., Douira A., et al. (2021). Biostimulants derived from moroccan seaweeds: Seed germination metabolomics and growth promotion of tomato plant. J. Plant Growth Regul. 40, 353–370. doi: 10.1007/s00344-020-10104-5
Nair P., Kandasamy S., Zhang J., Ji X., Kirby C., Benkel B., et al. (2012). Transcriptional and metabolomic analysis of Ascophyllum nodosum mediated freezing tolerance in Arabidopsis thaliana. BMC Genomics 13, 1–23. doi: 10.1186/1471-2164-13-643
Nanda S., Kumar G., Hussain S. (2022). Utilization of seaweed-based biostimulants in improving plant and soil health: current updates and future prospective. Int. J. Environ. Sci. Technol. 19, 12839–12852. doi: 10.1007/s13762-021-03568-9
Neily W., Shishkov L., Nickerson S., Titus D., Norrie J. (2010). “Commercial extract from the brown seaweed ascophyllum nodosum (Acadian) improves early establishment and helps resist water stress in vegetable and flower seedlings,” in Horticultural science, vol. 105. (Palm Desert, Califonia: American Society for Horticultural Science).
Nelson W. R., van Staden J. (1984). The effect of seaweed concentrate on wheat culms. J. Plant Physiol. 115, 433–437. doi: 10.1016/S0176-1617(84)80042-5
Nelson W. R., van Staden J. (1986). Effect of seaweed concentrate on the growth of wheat. South Afr. J. Sci. 82, 199–200.
Norrie J., Keathley J. P. (2006). Benefits of Ascophyllum nodosum marine-plant extract applications to “Thompson seedless” grape production. Acta Hortic. 727, 243–247. doi: 10.17660/actahortic.2006.727.27
Nugroho Jati B., Nuraeni C., Yunilawati R., Oktarina E. (2019). Phytochemical screening and total lipid content of marine macroalgae from binuangeun beach. J. Physics: Conf. Ser., 1317, 1–7. doi: 10.1088/1742-6596/1317/1/012103
Omidbakhshfard M. A., Sujeeth N., Gupta S., Omranian N., Guinan K. J., Brotman Y., et al. (2020). A biostimulant obtained from the seaweed Ascophyllum nodosum protects Arabidopsis thaliana from severe oxidative stress. Int. J. Mol. Sci. 21, 474–499. doi: 10.3390/ijms21020474
Pangestuti R., Kim S. (2015). “Chapter 6 - seaweed proteins, peptides, and amino acids,” in Seaweed sustainability (Cambridge, Massachusetts: Elsevier Inc), 125–140. doi: 10.1016/B978-0-12-418697-2.00006-4
Patle P. N., Kadu P. R., Gabhane A. R., Pharande A. L., Bhagat A. P., Bhoyar S. M., et al. (2019). Consequences provoked due to excess application of agrochemical on soil health deterioration - a review for sustainable agriculture. J. Pharmacogn. Phytochem. 8, 63–66.
Percival E., McDowell R. H. (1967). Chemistry and enzymology of marine algal polysaccharides (London: Academic Press).
Pharr D. M., Stoop J. M. H., Feusi M. E. S., Williamson J. D., Massel M. O., Conkling M. A. (1995). The dual role of mannitol as osmoprotectant and photoassimilate in celery. HortScience 30, 1182–1188. doi: 10.21273/HORTSCI.30.6.1182
Poberezny J., Szczepanek M., Wszelaczynska E., Prus P. (2020). The quality of carrot after field biostimulant application and after storage. Sustainability 12, 1386–1399. doi: 10.3390/su12041386
Ramya S. S., Nagaraj S., Vijayanand N. (2011). Influence of seaweed liquid extracts on growth, biochemical and yield characteristics of Cyamopsis tetragonoloba (L.) taub. J. Phytol. 3, 37–41.
Rasul F., Gupta S., Olas J. J., Gechev T., Sujeeth N., Mueller-Roeber B. (2021). Priming with a seaweed extract strongly improves drought tolerance in Arabidopsis. Int. J. Mol. Sci. 22, 1–28. doi: 10.3390/ijms22031469
Rayorath P., Jithesh M. N., Farid A., Khan W., Palanisamy R., Hankins S. D., et al. (2008a). Rapid bioassays to evaluate the plant growth promoting activity of Ascophyllum nodosum (L.) le jol. using a model plant, Arabidopsis thaliana (L.) heynh. J. Appl. Phycol. 20, 423–429. doi: 10.1007/s10811-007-9280-6
Rayorath P., Khan W., Palanisamy R., Mackinnon S. L., Stefanova R., Hankins S. D., et al. (2008b). Extracts of the brown seaweed Ascophyllum nodosum induce gibberellic acid (GA3)-independent amylase activity in barley. J. Plant Growth Regul. 27, 370–379. doi: 10.1007/s00344-008-9063-6
Raza A., Razzaq A., Mehmood S. S., Zou X., Zhang X., Lv Y., et al. (2019). Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 8, 34–63. doi: 10.3390/plants8020034
Renaut S., Masse J., Norrie J. P., Blal B., Hijri M. (2019). A commercial seaweed extract structured microbial communities associated with tomato and pepper roots and significantly increased crop yield. Microbial Biotechnol. 12, 1346–1358. doi: 10.1111/1751-7915.13473
Rizwan M., Ali S., Zaheer Akbar M., Shakoor M. B., Mahmood A., Ishaque W., et al. (2017). Foliar application of aspartic acid lowers cadmium uptake and cd-induced oxidative stress in rice under cd stress. Environ. Sci. Pollut. Res. 24, 21938–21947. doi: 10.1007/s11356-017-9860-1
Rouphael Y., Colla G. (2018). Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01655
Rouphael Y., Giordano M., Cardarelli M., Cozzolino E., Mori M., Kyriacou M. C., et al. (2018). Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 8, 126–141. doi: 10.3390/agronomy8070126
Rout G. R., Sahoo S. (2015). Role of iron in plant growth and metabolism. Rev. Agric. Sci. 3, 1–24. doi: 10.7831/ras.3.1
Samuels L. J., Setati M. E., Blancquaert E. H. (2022). Towards a better understanding of the potential benefits of seaweed based biostimulants in Vitis vinifera l. cultivars. Plants 11, 348–363. doi: 10.3390/plants11030348
Santaniello A., Scartazza A., Gresta F., Loreti E., Biasone A., Di Tommaso D., et al. (2017). Ascophyllum nodosum seaweed extract alleviates drought stress in Arabidopsis by affecting photosynthetic performance and related gene expression. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01362
Shrivastava P., Kumar R. (2015). Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 22, 123–131. doi: 10.1016/j.sjbs.2014.12.001
Shukla P. S., Borza T., Critchley A. T., Hiltz D., Norrie J., Prithiviraj B. (2018a). Ascophyllum nodosum extract mitigates salinity stress in Arabidopsis thaliana by modulating the expression of miRNA involved in stress tolerance and nutrient acquisition. PloS One 13, 1–25. doi: 10.1371/journal.pone.0206221
Shukla P. S., Mantin E. G., Adil M., Bajpai S., Critchley A. T., Prithiviraj B. (2019). Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00655
Shukla P. S., Shotton K., Norman E., Neily W., Critchley A. T., Prithiviraj B. (2018b). Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB Plants 10, 1–8. doi: 10.1093/aobpla/plx051
Silva L. D., Bahcevandziev K., Pereira L. (2019). Production of bio-fertilizer from Ascophyllum nodosum and Sargassum muticum (Phaeophyceae). J. Oceanology Limnology 37, 918–927. doi: 10.1007/s00343-019-8109-x
Sivasankari S., Venkatesalu V., Anantharaj M., Chandrasekaran M. (2006). Effect of seaweed extracts on the growth and biochemical constituents of Vigna sinensis. Bioresource Technol. 97, 1745–1751. doi: 10.1016/j.biortech.2005.06.016
Smiglak-Krajewska M., Wojciechowska-Solis J. (2021). Environmental awareness of farmers vs. agricultural sustainability. Eur. Res. Stud. J. 24, 161–172. doi: 10.35808/ersj/1956
Spann T. M., Little H. A. (2010). Effect of stimplex ® crop biostimulant on drought tolerance of ‘ Hamlin ’ sweet orange. Proc. Florida State Hortic. Soc. 123, 100–104.
Spann T. M., Little H. A. (2011). Applications of a commercial extract of the brown seaweed Ascophyllum nodosum increases drought tolerance in container-grown “hamlin” sweet orange nursery trees. HortScience 46, 577–582. doi: 10.21273/hortsci.46.4.577
Spinelli F., Fiori G., Noferini M., Sprocatti M., Costa G. (2009). Perspectives on the use of a seaweed extract to moderate the negative effects of alternate bearing in apple trees. J. Hortic. Sci. Biotechnol. 84, 131–137. doi: 10.1080/14620316.2009.11512610
Spinelli F., Fiori G., Noferini M., Sprocatti M., Costa G. (2010). A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Scientia Hortic. 125, 263–269. doi: 10.1016/j.scienta.2010.03.011
Staykov N. S., Angelov M., Petrov V., Minkov P., Kanojia A., Guinan K. J., et al. (2021). An Ascophyllum nodosum-derived biostimulant protects model and crop plants from oxidative stress. Metabolites 11, 1–20. doi: 10.3390/metabo11010024
Stengel D. B., Connan S., Popper Z. A. (2011). Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 29, 483–501. doi: 10.1016/j.biotechadv.2011.05.016
Steveni C. M., Norrington-Davies J., Hankins S. D. (1992). Effect of seaweed concentrate on hydroponically grown spring barley. J. Appl. Phycol. 4, 173–180. doi: 10.1007/BF02442466
Stiger-Pouvreau V., Bourgougnon N., Deslandes E. (2016). “Chapter 8 - carbohydrates from seaweeds,” in Seaweed in health and disease prevention. Eds. Fleurence J., Levine I. (London: Academic Press), 223–274. doi: 10.1016/B978-0-12-802772-1.00008-7
Stiger-Pouvreau V., Jégou C., Cérantola S., Guérard F., Le Lann K. (2014). “Chapter 13 - phlorotannins in sargassaceae species from Brittany (France): Interesting molecules for ecophysiological and valorisation purposes,” in Advances in botanical research. (Cambridge, Massachusetts: Academic Press). Ed. Bourgougnon N., 379–411. doi: 10.1016/B978-0-12-408062-1.00013-5
Stoop J. M. H., Williamson J. D., Pharr D. M. (1996). Mannitol metabolism in plants: A method for coping with stress. Trends Plant Sci. 1, 139–144. doi: 10.1016/s1360-1385(96)80048-3
Sujeeth N., Petrov V., Guinan K. J., Rasul F., O’sullivan J. T., Gechev T. S. (2022). Current insights into the molecular mode of action of seaweed-based biostimulants and the sustainability of seaweeds as raw material resources. Int. J. Mol. Sci. 23, 7654–7677. doi: 10.3390/ijms23147654
Sun T. P., Gubler F. (2004). Molecular mechanism of gibberellin signalling in plants. Annu. Rev. Plant Biol. 55, 197–223. doi: 10.1146/annurev.arplant.55.031903.141753
Sunarpi S., Jupri A., Kurnianingsih R., Julisaniah N. I., Nikmatullah A. (2011). Effect of seaweed extracts on growth and yield of rice plants. Nusantara Bioscience 8, 73–77. doi: 10.13057/nusbiosci/n020204
Tanveer M. (2020). “An overview of nickel toxicity in plants,” in Metal toxicology handbook. Eds. Bagchi D., Bagchi M. (Boca Raton, Florida: CRC Press), 465–474. doi: 10.1201/9780429438004-34
Taskos D., Stamatiadis S., Yvin J. C., Jamois F. (2019). Effects of an Ascophyllum nodosum (L.) le jol. extract on grapevine yield and berry composition of a merlot vineyard. Scientia Hortic. 250, 27–32. doi: 10.1016/j.scienta.2019.02.030
Teklić T., Parađiković N., Špoljarević M., Zeljković S., Lončarić Z., Lisjak M. (2021). Linking abiotic stress, plant metabolites, biostimulants and functional food. Ann. Appl. Biol. 178, 169–191. doi: 10.1111/aab.12651
Temple W. D., Bomke A. A., Radley R. A., Holl F. B. (1989). Effects of kelp (Macrocystis integrifolia and Ecklonia maxima) foliar applications on bean crop growth and nitrogen nutrition under varying soil moisture regimes. Plant Soil 117, 75–83. doi: 10.1007/BF02206259
Thiébaut N., Hanikenne M. (2022). Zinc deficiency responses: Bridging the gap between Arabidopsis and dicotyledonous crops. J. Exp. Bot. 73, 1699–1716. doi: 10.1093/jxb/erab491
UN DESA (2017)World population prospects - population division. In: World population prospects: 2017 revision. Available at: https://population.un.org/wpp/Download/Standard/Population/ (Accessed November 25, 2022).
Urbanek Krajnc A., Ivanuš A., Kristl J., Šušek A. (2012). Seaweed extract elicits the metabolic responses in leaves and enhances growth of Pelargonium cuttings. Eur. J. Hortic. Sci. 77, 170–181.
Valencia R. T., Acosta L. S., Hernández M. F., Rangel P. P., Robles M.Á.G., Cruz R., et al. (2018). Effect of seaweed aqueous extracts and compost on vegetative growth, yield, and nutraceutical quality of cucumber (Cucumis sativus l.) fruit. Agronomy 8, 264–277. doi: 10.3390/agronomy8110264
Van Nguyen D., Nguyen H. M., Le N. T., Nguyen K. H., Nguyen H. T., Le H. M., et al. (2022). Copper nanoparticle application enhances plant growth and grain yield in maize under drought stress conditions. J. Plant Growth Regul. 41, 364–375. doi: 10.1007/s00344-021-10301-w
van Staden J., Upfold S. J., Drewes F. E. (1994). Effect of seaweed concentrate on growth and development of the marigold Tagetes patula. J. Appl. Phycol. 6, 427–428. doi: 10.1007/BF02182160
Wadas W., Dziugieł T. (2020). Changes in assimilation area and chlorophyll content of very early potato (Solanum tuberosum l.) cultivars as influenced by biostimulants. Agronomy 10, 387–397. doi: 10.3390/agronomy10030387
Wally O. S. D., Critchley A. T., Hiltz D., Craigie J. S., Han X., Zaharia L. I., et al. (2013). Regulation of phytohormone biosynthesis and accumulation in Arabidopsis following treatment with commercial extract from the marine macroalga Ascophyllum nodosum. J. Plant Growth Regul. 32, 324–339. doi: 10.1007/s00344-012-9301-9
Wang Y., Fu F., Li J., Wang G., Wu M., Zhan J., et al. (2016). Effects of seaweed fertilizer on the growth of Malus hupehensis rehd. seedlings, soil enzyme activities and fungal communities under replant condition. Eur. J. Soil Biol. 75, 1–7. doi: 10.1016/j.ejsobi.2016.04.003
Wang Y., Xiang L., Wang S., Wang X., Chen X., Mao Z. (2017). Effects of seaweed fertilizer on the Malus hupehensis rehd. seedlings growth and soil microbial numbers under continue cropping. Acta Ecologica Sin. 37, 180–186. doi: 10.1016/j.chnaes.2017.01.004
Whapham C. A., Blunden G., Jenkins T., Hankins S. D. (1993). Significance of betaines in the increased chlorophyll content of plants treated with seaweed extract. J. Appl. Phycol. 5, 231–234. doi: 10.1007/BF00004023
Xu C., Leskovar D. I. (2015). Effects of A. nodosum seaweed extracts on spinach growth, physiology and nutrition value under drought stress. Scientia Hortic. 183, 39–47. doi: 10.1016/j.scienta.2014.12.004
Yao Y., Wang X., Chen B., Zhang M., Ma J. (2020). Seaweed extract improved yields, leaf photosynthesis, ripening time, and net returns of tomato (Solanum lycopersicum mill.). ACS Omega 5, 4242–4249. doi: 10.1021/acsomega.9b04155
Yusuf R., Kristianse P., Warwick N. (2019). Effect of two seaweed products and equivalent mineral treatments on lettuce (Lactuca sativa l.) growth. J. Agron. 18, 100–106. doi: 10.3923/ja.2019.100.106
Zamani-Babgohari M., Critchley A. T., Norrie J., Prithiviraj B. (2019). Increased freezing stress tolerance of Nicotiana tabacum l. cv. bright yellow-2 cell cultures with the medium addition of Ascophyllum nodosum (L.) le jolis extract. In Vitro Cell. Dev. Biol. - Plant 55, 321–333. doi: 10.1007/S11627-019-09972-8
Zhang X., Ervin E. H. (2004). Cytokinin-containing seaweed and humic acid extracts associated with creeping bentgrass leaf cytokinins and drought resistance. Crop Sci. 44, 1737–1745. doi: 10.2135/cropsci2004.1737
Zhang X., Ervin E. H. (2008). Impact of seaweed extract-based cytokinins and zeatin riboside on creeping bentgrass heat tolerance. Crop Sci. 48, 364–370. doi: 10.2135/cropsci2007.05.0262
Zhang X., Wang K., Ervin E. H. (2010). Optimizing dosages of seaweed extract-based cytokinins and zeatin riboside for improving creeping bentgrass heat tolerance. Crop Sci. 50, 316–320. doi: 10.2135/cropsci2009.02.0090
Zou P., Lu X., Zhao H., Yuan Y., Meng L., Zhang C., et al. (2019). Polysaccharides derived from the brown algae Lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00048
Zou P., Yang X., Yuan Y., Jing C., Cao J., Wang Y., et al. (2021). Purification and characterization of a fucoidan from the brown algae Macrocystis pyrifera and the activity of enhancing salt-stress tolerance of wheat seedlings. Int. J. Biol. Macromol. 180, 547–558. doi: 10.1016/j.ijbiomac.2021.03.039
Keywords: Phaeophyceae, seaweeds extracts, primary and secondary metabolites, plant physiology, agricultural biostimulants, commercial products, abiotic stress tolerance, mechanisms of action
Citation: Kergosien N, Stiger-Pouvreau V, Connan S, Hennequart F and Brébion J (2023) Mini-Review: brown macroalgae as a promising raw material to produce biostimulants for the agriculture sector. Front. Agron. 5:1109989. doi: 10.3389/fagro.2023.1109989
Received: 28 November 2022; Accepted: 29 March 2023;
Published: 16 May 2023.
Edited by:
Rachid Bouharroud, National Institute of Agronomic Research, MoroccoReviewed by:
Mohammed Antar, McGill University, Macdonald Campus, CanadaCopyright © 2023 Kergosien, Stiger-Pouvreau, Connan, Hennequart and Brébion. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nolwenn Kergosien, bm9sd2Vubi5rZXJnb3NpZW5AYWxnYWlhLmNvbQ==; bm9sd2Vubi5rZXJnb3NpZW5AZ21haWwuY29t