
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Agron. , 13 October 2022
Sec. Weed Management
Volume 4 - 2022 | https://doi.org/10.3389/fagro.2022.985067
 Muhammadu Tajudeen Salaudeen1*
Muhammadu Tajudeen Salaudeen1* Emmanuel Daniya1
Emmanuel Daniya1 Olayemi Mikail Olaniyi2
Olayemi Mikail Olaniyi2 Taliha Abiodun Folorunso3
Taliha Abiodun Folorunso3 Jibril Abdullahi Bala3
Jibril Abdullahi Bala3 Ibrahim Mohammed Abdullahi2
Ibrahim Mohammed Abdullahi2 Bello Kontagora Nuhu2
Bello Kontagora Nuhu2 Adeyinka Peace Adedigba3
Adeyinka Peace Adedigba3 Blessing Israel Oluwole1
Blessing Israel Oluwole1 Abdullah Oreoluwa Bankole3
Abdullah Oreoluwa Bankole3 Odunayo Moses Macarthy3
Odunayo Moses Macarthy3Maize is a food crop for millions of people in sub-Saharan Africa. However, severe weed infestation might cause significant yield loss. This study investigated weed composition, abundance and distribution in maize-based cropping systems in the Southern Guinea Savanna of Nigeria. Fields were surveyed between February and March 2022. The Importance Value Index (IVI) of each weed species was determined using relative frequency, relative density, and relative abundance. A total of 29 weed species from 15 families were identified. Poaceae (34.9 %) was the most prevalent weed family, followed by Cyperaceae (26.9 %) and Portulacaceae (12.8 %). The IVI showed that Cyperus rotundus (38.6), Portulaca oleraceae (29.4), Digitaria horizontalis (25.5), Brachiaria deflexa (24.2), Senna obtusifolia (17.6), Ageratum conyzoides (16.0), Cynodon dactylon (12.6), Phyllanthus niruri (11.1) and Eragrostis sp. (10.6) were the most dominant. Principal Component Analysis (PCA) ordination biplot revealed that Setaria pumila was strongly associated with sole maize cultivation, as was Desmodium scorpiurus with maize/rice intercropping, C. dactylon with maize/pepper, Euphorbia hirta with maize/cassava, and Cleome viscosa with maize/amaranthus intercropping. Special attention to these weed species is required when making an informed decision on the choice of weed control measures.to reduce yield losses in endemic areas.
Sub-Saharan Africa's countries will add more than one billion people by 2050, accounting for more than half of the global population increase (UNDESAPPD (United Nations, Department of Economic and Social Affairs, Population Division), 2019). Unfortunately, the current food production levels are insufficient to feed the predicted population, and addressing this need is a significant problem (Ranganathan et al., 2018). According to a Food and Agriculture Organisation report (FAO et al., 2019), millions of people in sub-Saharan Africa are food insecure. These factors adversely impact healthy living and long-term development. As a result, adopting new high-intensity cropping methods is critical for enhancing agricultural output in this situation (Waha et al., 2020). Maize (Zea mays L.) production contributes to food security and income for smallholder farmers in sub-Saharan Africa. It is also an important raw material in the livestock industry for feed formulation. Although it is often grown during the rainy season, it can also be cultivated in a lowland ecosystem during the dry season if dependable irrigation facilities are available. Despite its numerous applications, maize productivity is threatened by weed invasion at various stages of growth (Colbach et al., 2020).
Weeds absorb moisture, nutrients, and sunlight instead of agricultural plants, lowering crop yield and quality (Sabanci and Aydin, 2017). Some weeds also act as alternate hosts for insect pests and pathogens. They are more economically damaging than insects, fungi, or other crop pests (Gharde et al., 2018), causing approximately 40 % of crop output loss. Weeds vary from location to location and season to season, and the level of infestation primarily depends on the species, density and diversity in a crop community (Korav et al., 2018). The morphological architecture of maize plants makes them poor competitors to weeds. Therefore, an infestation of weeds during the prime growth period may result in unacceptable losses (Chauhan, 2020). Weeds are diverse and spread across a vast geographical area. Some species are better suited to highland or lowland environments, while others can thrive in both.
The ability of a weed to dominate specific farmland has significance for its survival and geographical spread. Oldenlandia corymbosa, for example, is known to rapidly establish and dominate in a location (Global Invasive Species Database, 2022). The weed produces a large number of seeds, which can be dispersed by farm machinery, wind, and rain. As a result, it becomes increasingly difficult to manage. Ageratum conyzoides also produces a large number of seeds and is resistant to a wide range of germination temperatures and light conditions (Yuan and Wen, 2018). Allelopathy is another mechanism of survival and field colonization. For example, some weeds, such as A. conyzoides and Portulaca oleracea, can release compounds that impair the growth of agricultural plants in their vicinity (Rashidi et al., 2021; Paul et al., 2022).
Weed management is critical for achieving maximum crop production and will play a key role in determining whether we can fulfil future food demands (Korav et al., 2018). Crop production is unattractive to teeming unemployed youths due to the numerous obstacles associated with weed management. Manual weed control is time-consuming, and labour is scarce during the cropping season. Herbicide application provides an immediate response to weed infestations, but it is costly, hazardous to humans and livestock, contaminates soil and groundwater, and escalates production costs. Smallholder farmers in sub-Saharan Africa use various combinations of maize intercropping systems involving cowpea, groundnut, or soyabean for weed control. These combinations can provide some control, but those including sorghum (Sorghum bicolor L.), sugarcane (Saccharum officinarum L.), rice (Oryza sativa L.), pepper (Capsicum spp), cassava (Manihot esculenta Crantz), and millet (Pennisetum spp.) worsen weed infestations.
Weed abundance and distribution are critical determinants of field invasion and crop yield.
Changes in these variables are the result of selective pressures applied by farmers during crop production (Nkoa et al., 2015). Tillage methods, sowing dates, inter and intra-row spacing, herbicide, fertiliser, and cropping system selection are just a few examples. Information on the weed population that causes the most damage to an agricultural crop is crucial for creating long-term weed management strategies. It would also help to increase crop output and harvest quality. Typically, this information is obtained through phytosociological surveys, which include indices such as relative frequency, relative density, relative abundance, and relative importance (Silva et al., 2017). From this perspective, the most impacted component (frequency, density, and abundance) may provide evidence of how environmental agents interact with the weed population (Silva et al., 2017). Therefore, this study investigated weed composition, abundance, and distribution in a lowland environment with maize-based cropping systems.
The study was conducted in the four Local Government Areas (LGAs) of Niger State, Southern Guinea Savanna agro-ecological zone of Nigeria. The selected LGAs were Bosso, Gbako, Katcha, and Lapai (Figure 1). The State lies between 08o−11o30' N and 03o−07o40' E. The dry and rainy seasons are distinct, with annual rainfall ranging from 1,100 mm in the North to 1,600 mm in the South. The highest temperatures are frequently reported between March and June, while the lowest occur between December and January. In the northern zone of the State, the rainy season lasts for 120 days, whereas, in the southern part, it is approximately 150 days. The soils are Alfisols, with a high water-holding capacity derived from basement complex rocks.
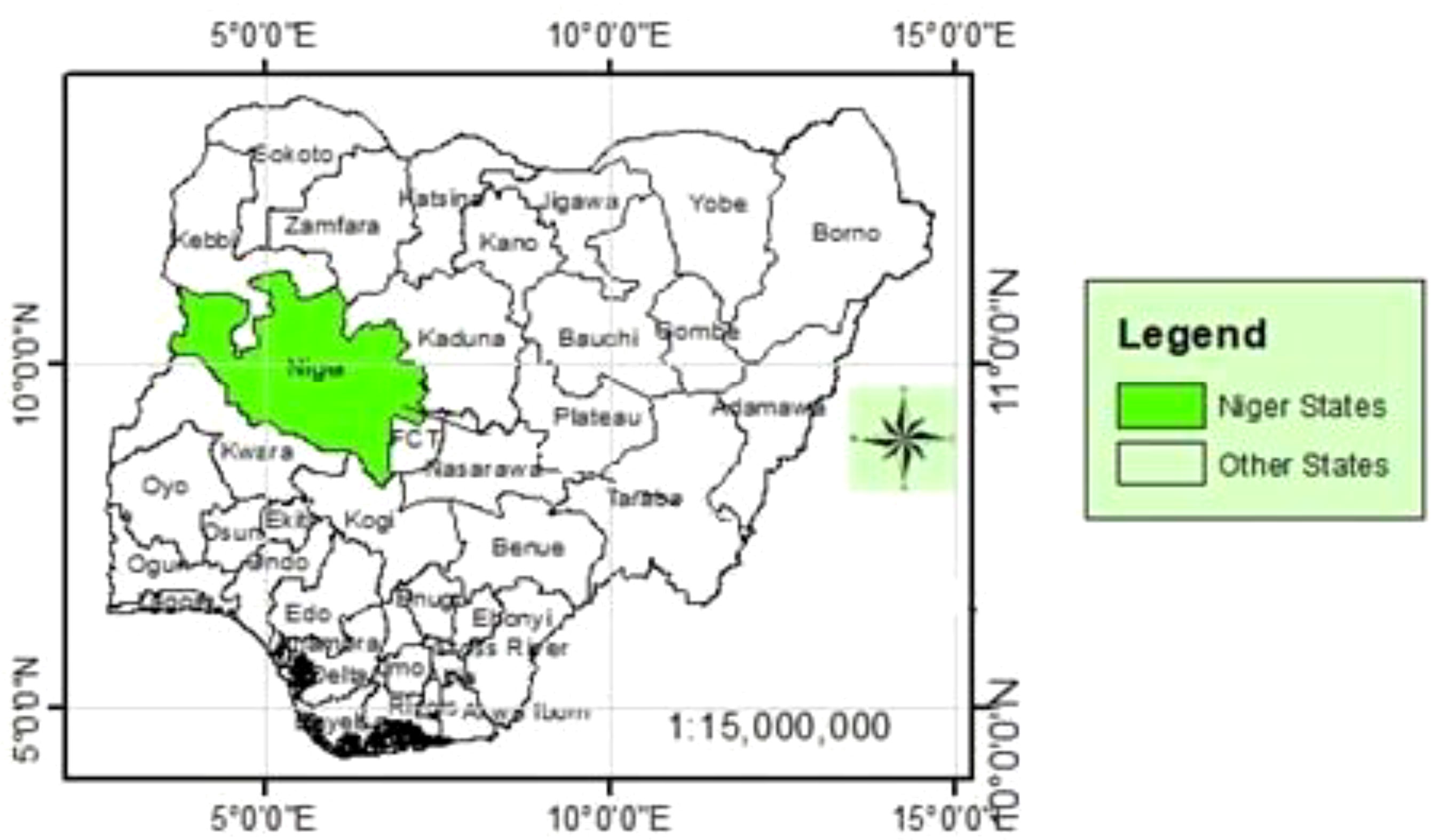
Figure 1 Map of Nigeria showing Niger State surveyed for weeds associated with different maize-based cropping systems.
Surveys were conducted on 18 maize-based farms in the selected LGAs between February and March 2022. Two of the farms were located in Bosso LGA (Anguwan-Shaba _01 and Anguwan-Shaba _02), seven in Gbako LGA (Sabon-Gida_01− Sabon-Gida_07), seven in Katcha LGA (Katcha_01− Katcha_07) and two in Lapai LGA (Lapai GGSS-Day_01 and Lapai GGSS-Day_02). Structured questionnaires were administered to 90 maize farmers (respondents) to obtain information on the cropping history (farm size, maize-based cropping systems, weed control methods and farmers’ knowledge of allelopathic weeds). The coordinates of each field were taken using a Global Positioning System (GPS- 4300; Ethrex Garmin, Taiwan) (John et al., 2020). Weeds were sampled when the maize plants were between the vegetative and grain-filling stages. Twenty 0.25 m2 quadrats were randomly placed along the transect in the “W” guided pattern mapped out in each field for weed species collection (Sintayehu, 2019). The weed species in each quadrat were counted and identified using a standard weed manual (Akobundu et al., 2016).
The following phytosociological parameters were calculated for each species: frequency, relative frequency, density, relative density, abundance, relative abundance, and Importance Value Index (IVI) (Lopes et al., 2021).
The data on the number of weed species from the 18 locations were subjected to Principal Component Analysis (PCA) to determine the relationships among the weed species and between the weed species and maize cropping systems (sole maize, maize/amaranthus, maize/cassava, maize/pepper, and maize/rice). The PCA was based on a correlation matrix of the weed species (variables) (Restuccia et al., 2020). Data were analysed using Statistical Analysis System software version 9.2 (SAS Institute Inc., Cary, NC).
Regardless of location, the farms were less than 1 ha in size (Figure 2). Seventy per cent of farmers in Bosso LGA intercropped maize with rice, while the remaining 30 % intercropped with cassava (Figure 3). Gbako LGA had slightly different results: 60 % of farmers intercropped maize with rice, followed by 25.7 % who used pepper, and the remaining 14.3 % intercropped maize with amaranthus. In Katcha, 60 % of farmers intercropped maize with rice, 25.7 % cultivated maize alone, and 14.3 % intercropped with pepper. Ninety per cent of farmers in Lapai LGA intercropped maize with rice, and the remaining 10 % intercropped with amaranthus (Figure 3). In Bosso LGA, 70 % of the farmers used herbicide in combination with hand weeding, and the remaining 30 % used hand weeding alone (Figure 4). In Gbako LGA, 71.4 % of respondents used hand weeding combined with herbicide for weed control, and 28.6 % used hand weeding alone. In Katcha LGA, 65.7 % of farmers used both hand weeding and herbicide, while 34.3 % used only hand weeding. In Lapai LGA, 60 % of farmers mixed hand weeding with herbicide application, and 40 % depended primarily on hand weeding. Based on the oral interview, farmers stopped applying herbicides when they no longer worked. In such cases, herbicide combinations were utilised. None of the farmers knew allelopathic weeds in Bosso, whereas 14.3 %, 11.4 %, and 10 % were aware in Gbako, Katcha, and Lapai LGA, respectively (Figure 5).
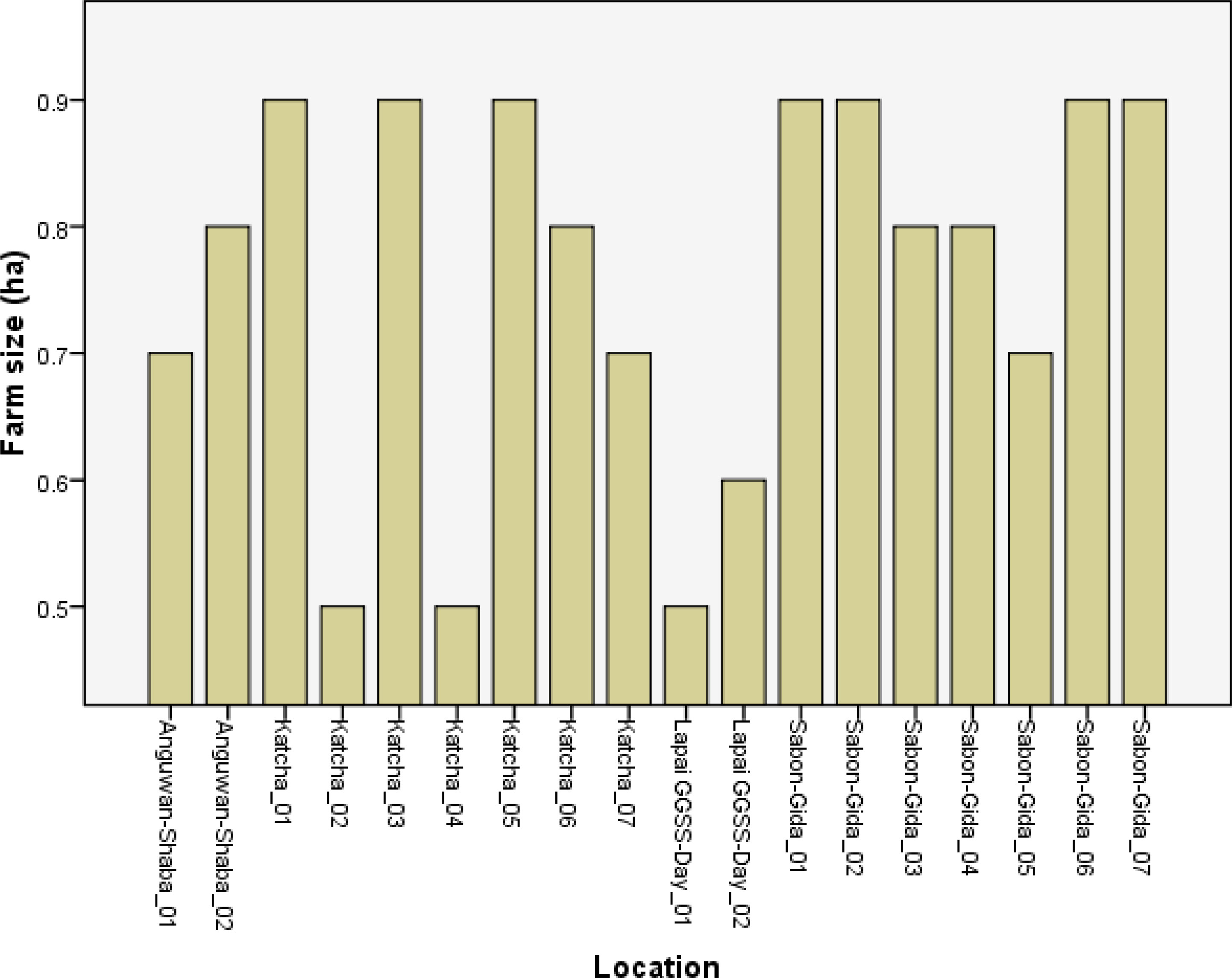
Figure 2 Farm size in different locations surveyed for maize-based cropping systems in Southern Guinea Savanna of Nigeria.
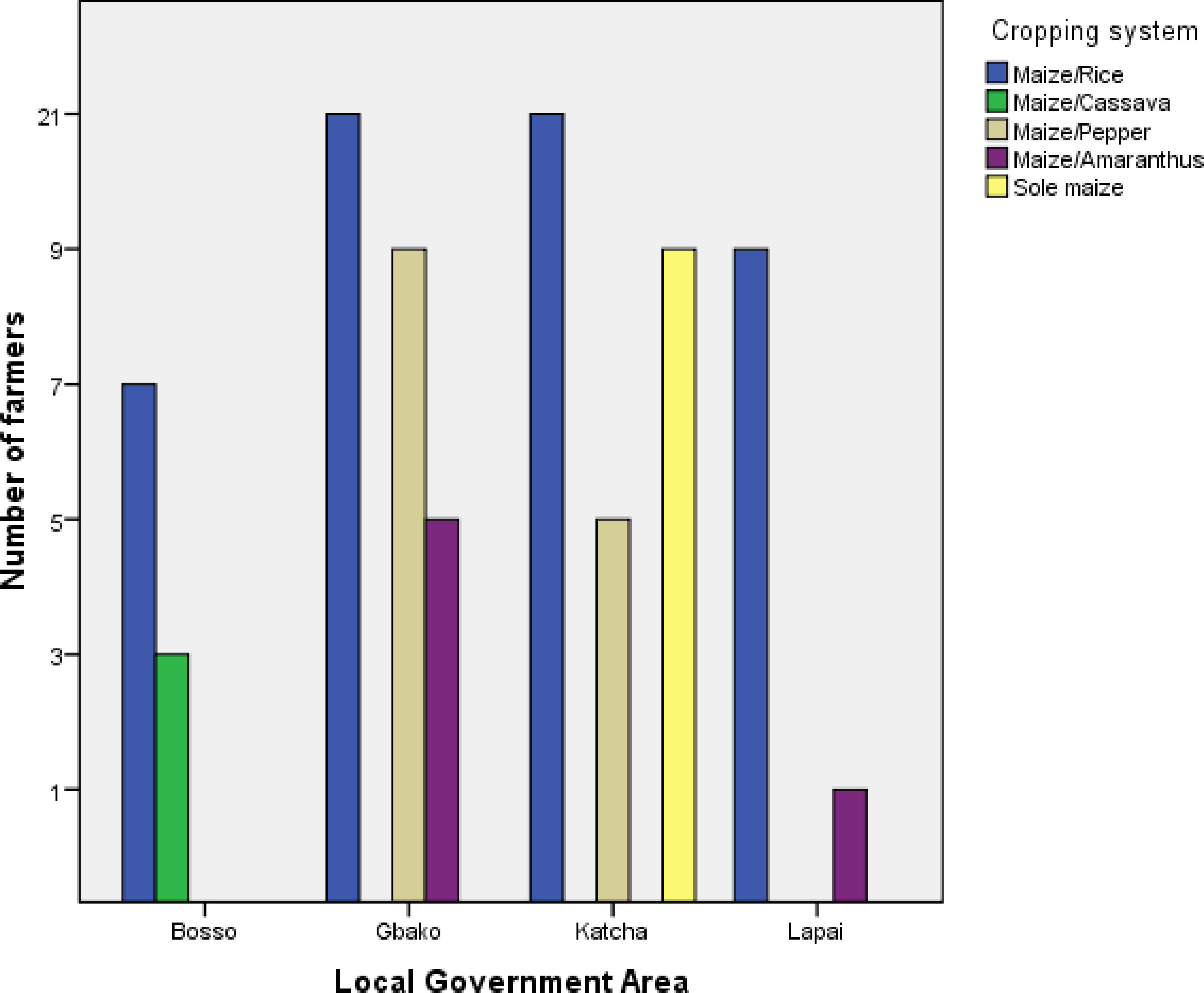
Figure 3 Frequency distribution of the farmers involved in maize-based cropping systems in Southern Guinea Savanna of Nigeria.
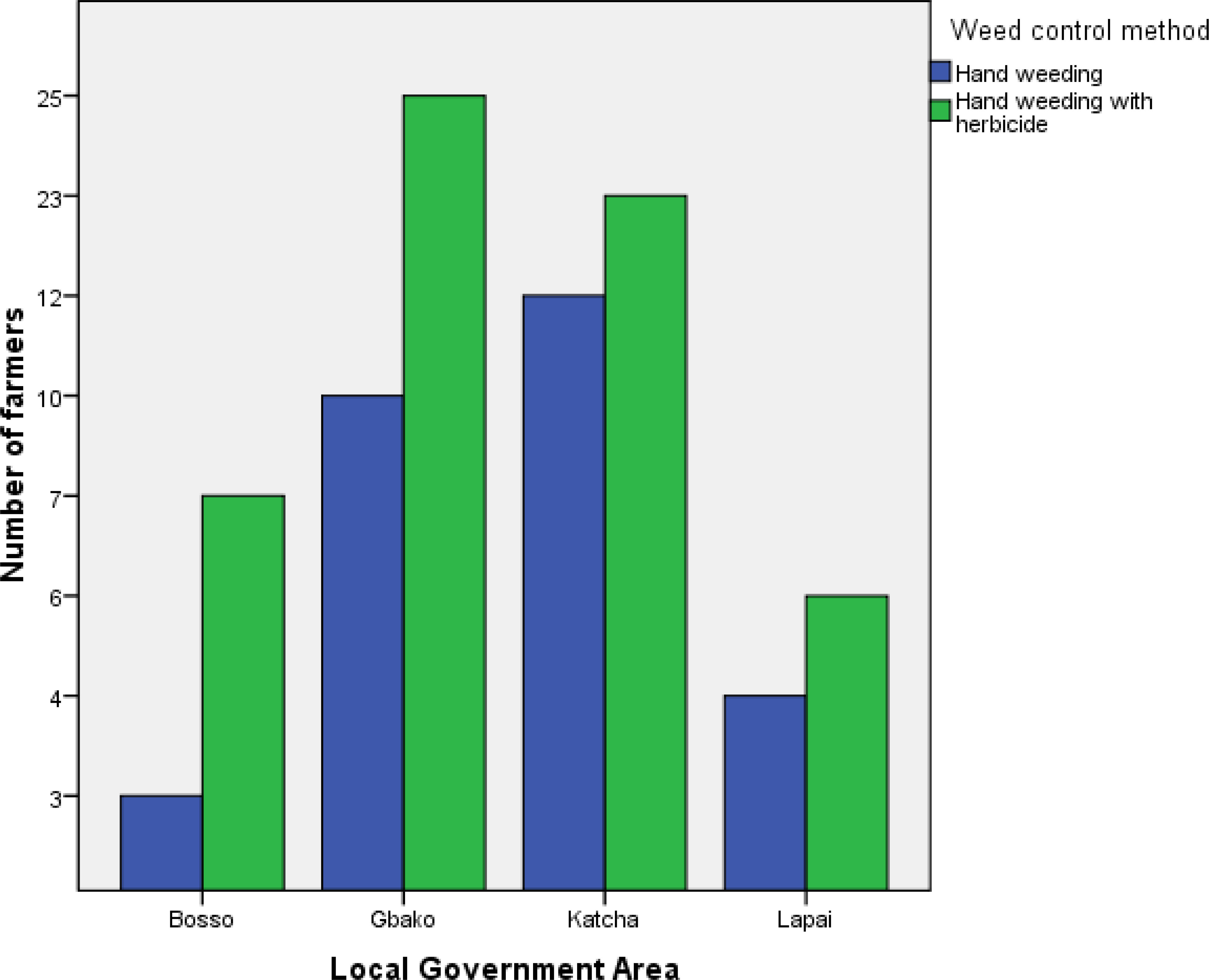
Figure 4 Frequency distribution of the farmers adopting different weed control methods under maize-based cropping systems in Southern Guinea Savanna of Nigeria.
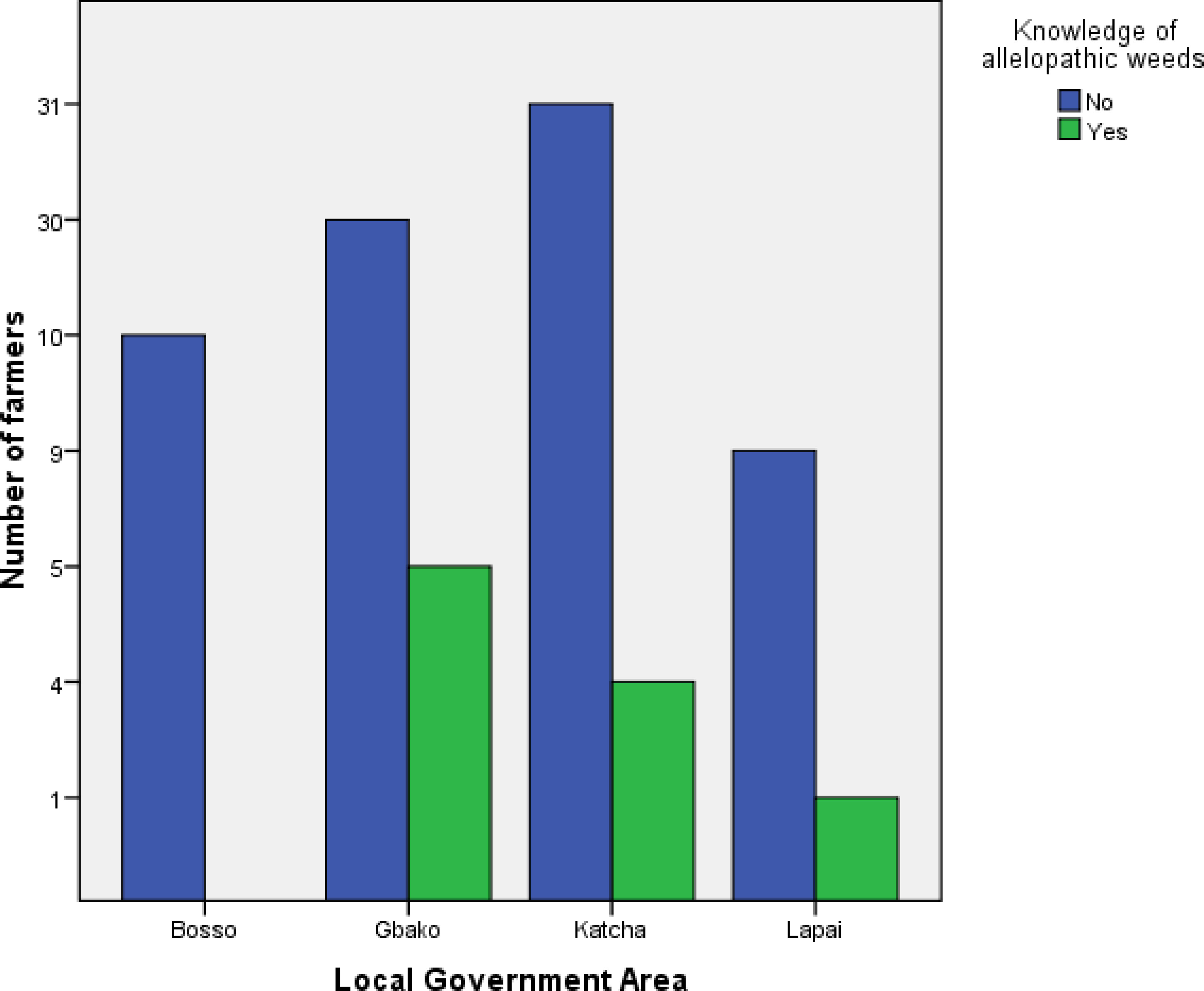
Figure 5 Farmers’ knowledge of allelopathic weeds under maize-based cropping systems in Southern Guinea Savanna of Nigeria.
Across the investigated sites, a total of 2, 041 individuals of 29 weed species belonging to 15 families were identified (Table 1). Cyperus rotundus L. accounted for 26.9 % of the total individuals, followed by Portulaca oleracea L. (12.8 %), Digitaria horizontalis Willd (12.1 %), and Brachiaria deflexa (Schumach.) C. E. Hubb with 11.9 %. Poaceae (34.9 %) were the most abundant, followed by Cyperaceae (26.9 %), Portulacaceae (12.8 %), Asteraceae (9.6 %), and Euphorbiaceae (6.2 %). The families Rubiaceae, Acanthaceae, and Malvaceae accounted for 2.3 %, 2.1 %, and 1.6 % of all weeds, respectively. However, the Fabaceae and Solanaceae families were both present in identical proportions (1.2 %). The remaining families had abysmally few members, namely Convolvulaceae (0.4 %), Capparidaceae (0.3 %), Commelinaceae (0.3 %), Leguminosae (0.2 %), and Loganiaceae (0.2 %). Broadleaf weeds accounted for 38.2 % of all weed species in terms of morphology (Table 1). With a relative frequency of 16.3 %, 12.9 %, and 11.7 %, respectively, P. oleracea, D. horizontalis, and B. deflexa ranked the highest among the weed species associated with maize fields. Cyperus rotundus (26.9 %), P. oleracea (12.8 %), D. horizontalis (12.1 %), and B. deflexa (11.9 %) were the most frequent and diversely populated weed species based on relative density. Senna obtusifolia (L.) Irwin & Bar had the highest relative abundance (16.2 %), followed by Dactyloctenum aegyptium (L.) P. Beauv, Ipomoea triloba, and Setaria pumila (Poir) Roem & Schult, all with 7.6 %. Eragrostis sp., Spermacoce ocymoides Burm F., and Cleome viscosa L. had a relative abundance of 6.9 %, 6.8 %, and 6.7 %, respectively. Based on IVI, the most common and prevailing weed species in the maize fields was C. rotundus with an IVI of 38.6. The other important and troublesome species were P. oleracea with an IVI of 29.4, followed by D. horizontalis (IVI = 25.5), B. deflexa (IVI = 24.2), S. obtusifolia (IVI = 17.6), A. conyzoides (IVI = 16.0), Cynodon dactylon (L.) Pers (IVI = 12.6), Phyllanthus niruri (IVI = 11.1), and Eragrostis sp. (IVI = 10.6) (Table 1). These weed species belong to the Portulacaceae (broadleaf), Poaceae (grass), Cyperaceae (sedge), Fabaceae (broadleaf), Convolvulaceae (broadleaf), Rubiaceae (broadleaf), and Capparidaceae (broadleaf) (Table 1).
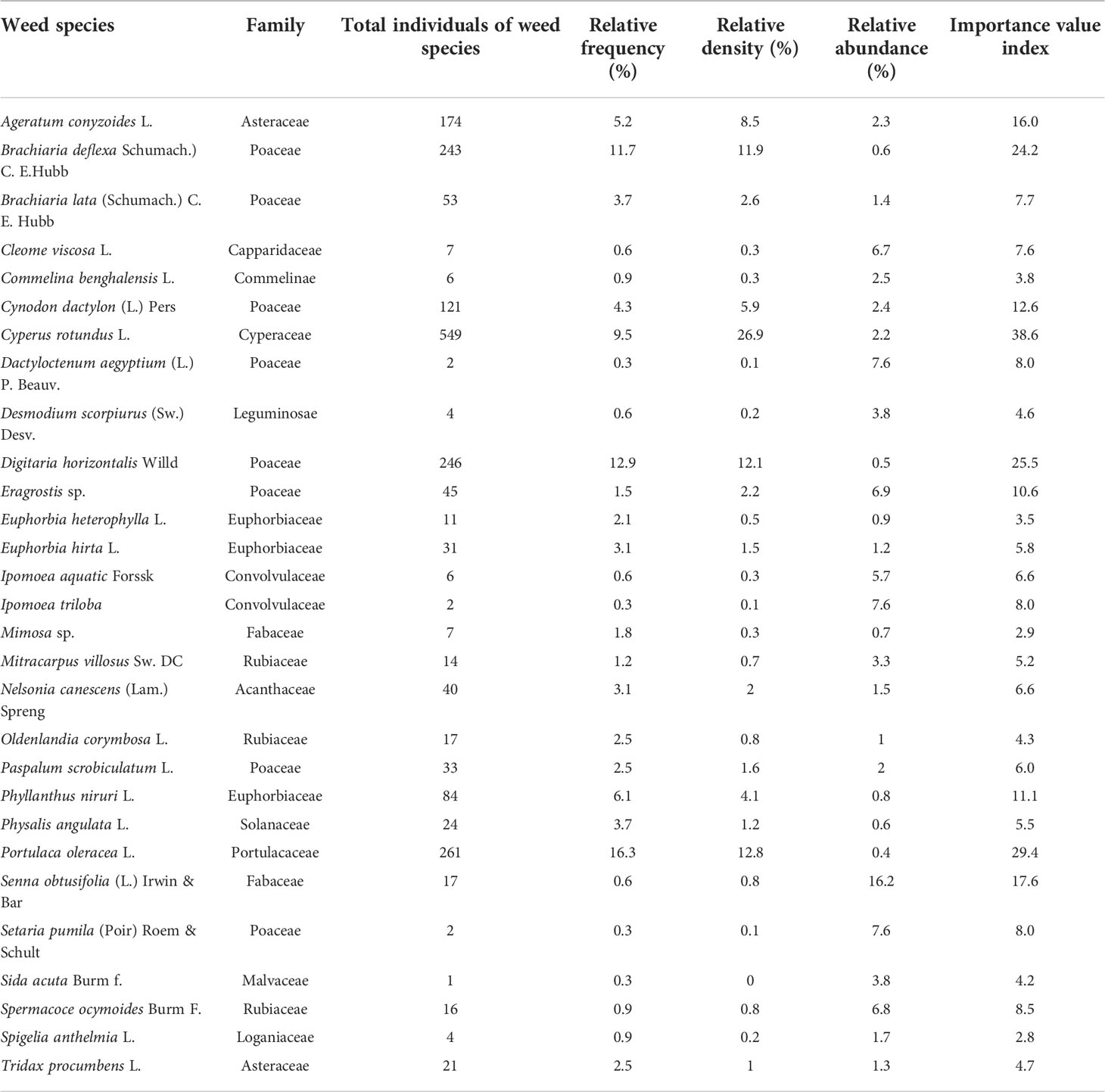
Table 1 Phytosociological parameters of the weed species from the survey of irrigated maize-based cropping systems in Southern Guinea Savanna of Nigeria.
The abundance and distribution of weeds varied according to the cultivation practices used in the study area (Table 2). The farms under maize/rice intercropping included Anguwan-Shaba_02, Katcha_01, Katcha_05, Katcha_06, Lapai GGSS-Day_01, Sabon-Gida_01, Sabon-Gida_03, Sabon-Gida_05, and Sabon-Gida_06 (Table 2). The difference in the grass and broadleaf abundance was not conspicuous in Anguwan-Shaba_02. Of the 87 individuals, sedges were the most abundant with 37 individuals followed by grasses with 27 individuals and broadleaf species with 23 individuals. In Katcha_01, grass weeds predominated with 100 individuals, whereas 46 individuals of broadleaf species were observed. Of the 104 individuals found in Katcha_05, sedges (58 individuals) were the most prevalent, followed by grasses (32 individuals) and broadleaves (14 individuals). Moreover, in Katcha_06, grass weeds constituted 36 individuals with a comparable number of broadleaf species (31 individuals), but only one individual of sedges was found (Table 2). Weed abundance varied substantially in Lapai GGSS-Day_01, where sedges predominated (33 individuals) against 13 individuals of broadleaf species and just one individual of grasses. Sabon-Gida_01 constituted 173 individuals, with sedges being the most abundant (158 individuals), followed by broadleaves with 8 individuals and grasses with 7 individuals. Sedge weeds dominated at Sabon-Gida_03 with 54 individuals, followed by broadleaves (19 individuals) and grass weeds (6 individuals). Sabon-Gida_05 had only grass (38 individuals) and broadleaf (28 individuals) weeds. Sabon-Gida_06, on the other hand, contained 98 individuals with 37 grasses, 33 broadleaves, and 28 sedges (Table 2).
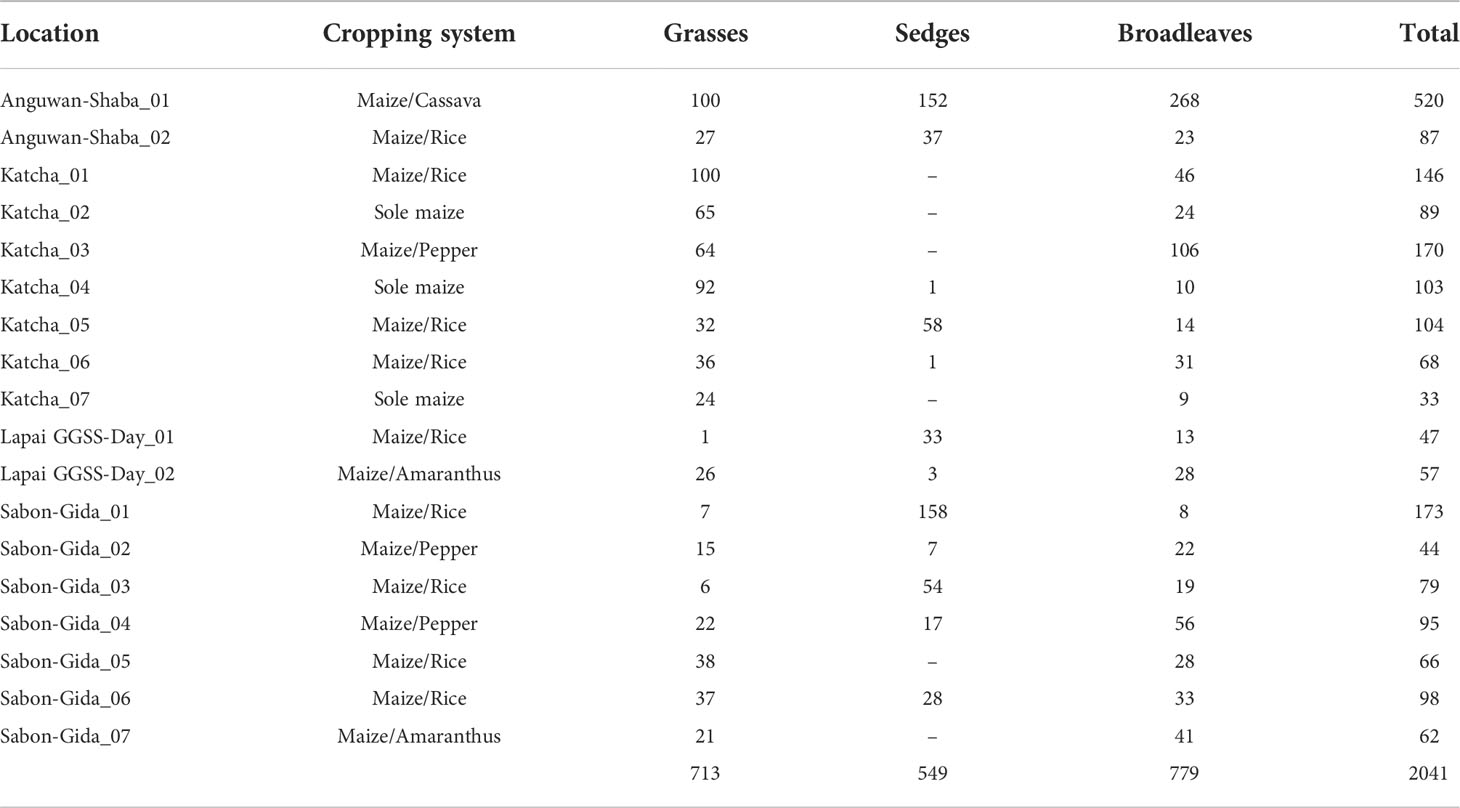
Table 2 Total individuals of grasses, sedges and broadleaf weeds under different maize-based cropping systems in Southern Guinea Savanna of Nigeria.
The farms supporting only maize cultivation included Katcha_02, Katcha_04, and Katcha_07 (Table 2). Weeds found at Katcha_02 were grasses and broadleaves, with grasses dominating (65 individuals) over the broadleaves (24 individuals). In Katcha_04, grass weeds predominated (92 individuals), followed by broadleaves (10 individuals), and only 1 individual of sedges. There were no sedges in Katcha_07, although there were 24 and 9 individuals of grass and broadleaf species, respectively (Table 2). Maize/pepper cultivation was witnessed in Katcha_03, where only broadleaf (106 individuals) and grass (64 individuals) species were found. In Sabon-Gida_02, broadleaf weeds were the most abundant (22 individuals), followed by grasses (15 individuals) and sedge weeds (7 individuals). In Sabon-Gida_04, broadleaves were represented by 56 individuals, followed by grass weeds with 22 individuals and sedges with 17 individuals (Table 2). Maize/amaranthus intercropping in Lapai GGSS-Day_02 resulted in comparable numbers of grass (26 individuals) and broadleaf (28 individuals) weeds but only a limited number of sedges (3 individuals) was found. In Sabon-Gida_07 it resulted in 41 broadleaf and 21 grass individuals but no sedges (Table 2). Maize/cassava intercropping was seen only in Anguwan-Shaba_01, where there was a total of 520 individuals with the maximum number of broadleaf species (268 individuals), followed by sedges (152 individuals) and grass weeds (100 individuals) (Table 2).
The PCA of the weed species from the 18 locations showed that the first three PCs accounted for 86.8 % of the cumulative variance (Table 3). Principal Component 1 (PC 1) explained 43.3 % of the total variance and correlated significantly with Mimosa sp., P. niruri, Eragrostis sp., A. conyzoides, Tridax procumbens L. Euphorbia heterophylla L., Oldenlandia corymbosa L., E. hirta, D. horizontalis, C. rotundus, Physalis angulata L., and P. oleracea (Table 3). Principal Component 2 (PC 2) explained 23.9 % of the variance and correlated positively withB.deflexa,Desmodium scorpiurus(Sw.) Desv., I. triloba, Mitracarpus villosus Sw. DC, and Spigelia anthelmia L., and negatively with C. benghalensis, B. lata, C.viscosa, and S. ocymoides. Furthermore, PC 3 added 19.6 % variance, with Dactyloctenum aegyptium (L.) P. Beauv., Ipomoea aquatica Forssk, Sida acuta Burm f., and S. obtusifolia contributing positively (Table 3). The PCA ordination biplot (Figure 6) revealed that S. pumila was the only species associated with sole maize cultivation, as were D. scorpiurus, I. triloba, S. anthelmia, M. villosus, and B. deflexa with maize and rice intercropping, C. dactylon, I. aquatica, D. aegyptium, S. obtusifolia, and S. acuta with maize and pepper cultivation, E. hirta and P. oleracea with maize and cassava intercropping, and C. viscosa, C. benghalensis, and B. lata with maize and amaranthus cultivation (Figure 6).
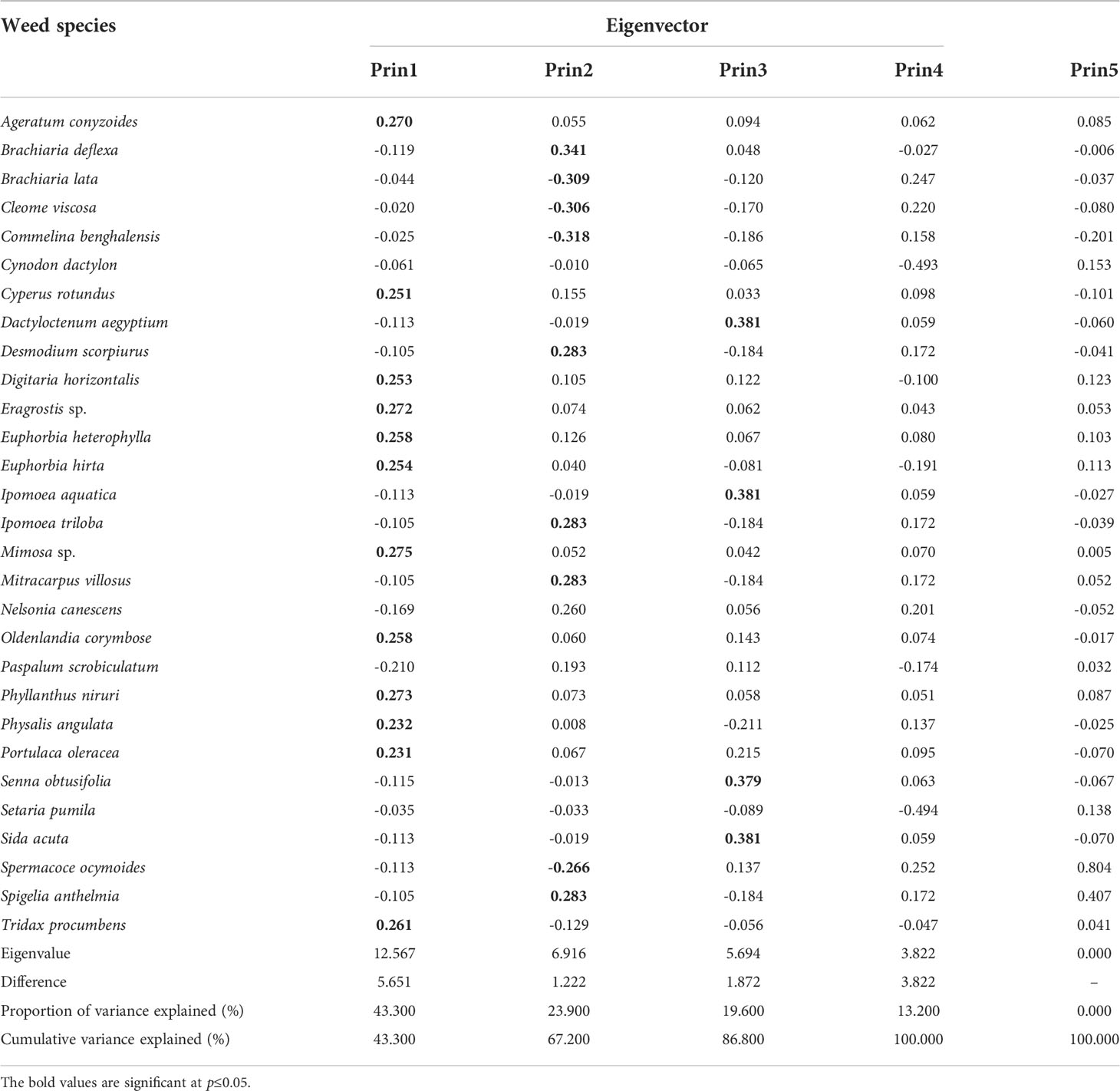
Table 3 Eigenvectors of the principal components from the weeds associated with maize-based cropping systems in Southern Guinea Savanna of Nigeria.
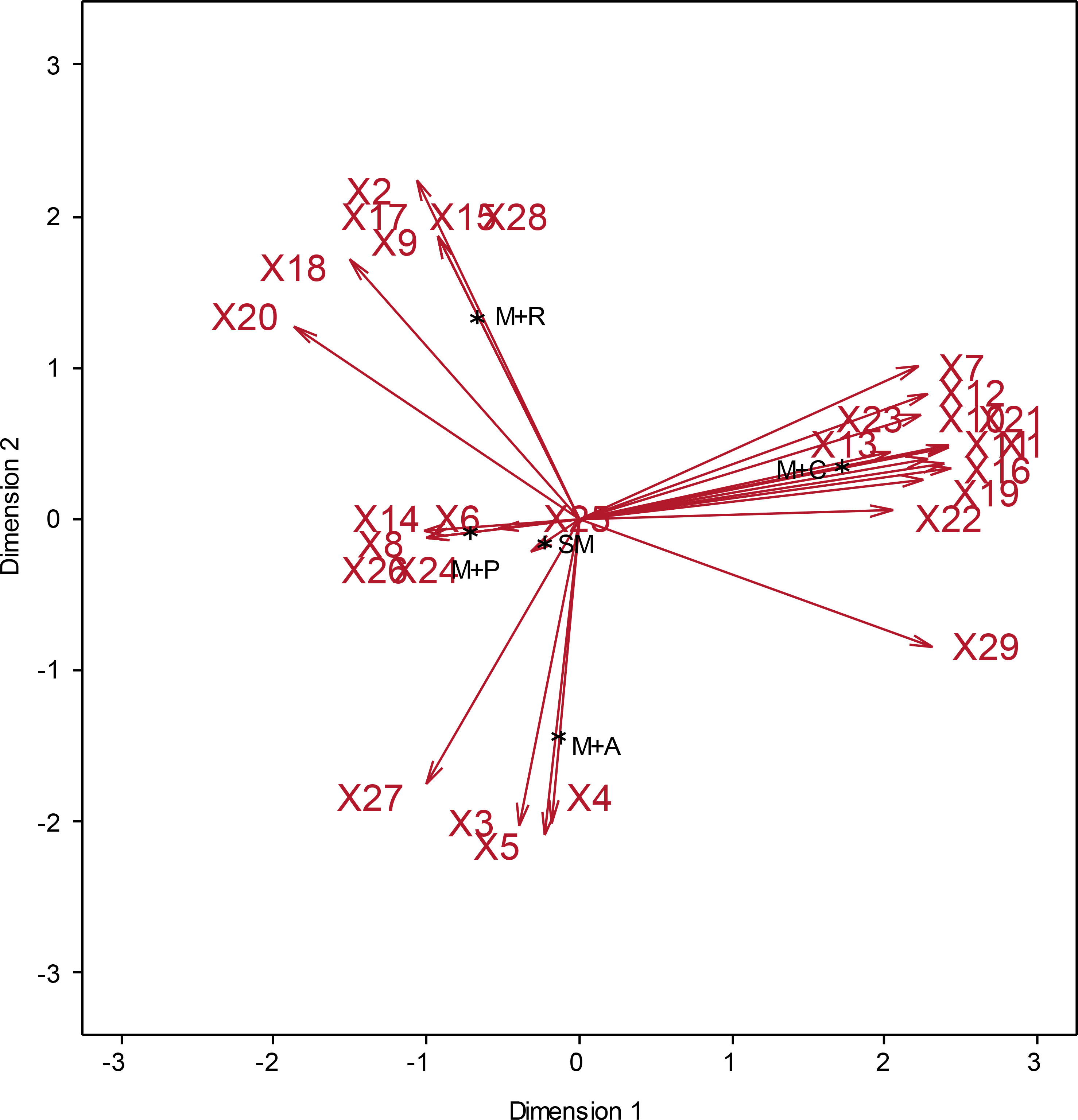
Figure 6 Principal component analysis ordination biplot of the weed species associated with maize-based cropping systems in Southern Guinea Savanna of Nigeria. X1, Ageratum conyzoides; X2, Brachiaria deflexa; X3, Brachiaria lata; X4, Cleome viscosa; X5, Commelina benghalensis; X6, Cynodon dactylon; X7, Cyperus rotundus; X8, Dactyloctenum aegyptium; X9, Desmodium scorpiurus; X10, Digitaria horizontalis; X11, Eragrostis sp.; X12, Euphorbia heterophylla; X13, Euphorbia hirta; X14, Ipomoea aquatica; X15, Ipomoea triloba; X16, Mimosa sp.; X17, Mitracarpus villosus; X18, Nelsonia canescens; X19, Oldenlandia corymbose; X20, Paspalum scrobiculatum; X21, Phyllanthus niruri; X22, Physalis angulate; X23, Portulaca oleracea; X24, Senna obtusifolia; X25, Setaria pumila; X26, Sida acuta; X27, Spermacoce ocymoides; X28, Spigelia anthelmia; X29, Tridax procumbens; M+A, Maize/Amaranthus intercropping; M+C, Maize/Cassava intercropping; M+P, Maize/Pepper intercropping; M+R, Maize/Rice intercropping; SM, Sole Maize.
Maize cultivation during the dry season in the study locations showed the potential of irrigated agriculture to boost food and nutrition security. The cultivation of maize on less than 1 ha revealed that crop production in the study locations was primarily at the subsistence level. This corroborates the findings of Bjornlund et al. (2020), who reported that agricultural production in sub-Saharan Africa is primarily for domestic consumption. In addition, the low land hectares cultivated with maize could be attributed to the fact that it was grown outside of its natural environment (upland). Although maize does not perform well in a lowland environment, some farmers grow it during the dry season to avoid waterlogging. The widespread maize intercropping systems observed in the study locations were due to farmers’ preferences for different types of food. Maize intercropping with rice was the most prevalent because lowland rice cultivars are adapted to hydromorphic farmland. As reported by WBGU (2020), crop diversity has the potential to improve sustainability while also preserving a variety of ecosystem functions. Therefore, intercropping maize with other crops would enhance the number of harvests while reducing the environmental impacts that agricultural production might have (Grote et al., 2021). It is laudable that none of the respondents relied primarily on herbicides for weed control. Excessive herbicide use might contribute to the ineffectiveness of some herbicides and the spread of weeds in Bosso, Gbako, and Katcha LGAs. According to Schütte et al. (2017), indiscriminate herbicide application promotes herbicide resistance and weed persistence. This is worrisome due to the negative impacts of herbicides on farmers, the environment, and biodiversity. The prevalence of farmers engaged in hand weeding is consistent with the findings of Sims et al. (2018). Hand weeding, frequently employed for weed management, may be attributable to a long-standing cultural tradition. It is cheap but tedious, making crop production unappealing to young people. In addition, farmers’ responses to allelopathic weed knowledge revealed a general lack of awareness in the study area.
Ecological analysis of the weed data showed that C. rotundus, P. oleracea, D. horizontalis, B. deflexa, S. obtusifolia, A. conyzoides, C. dactylon, P. niruri and Eragrostis sp. were the major weeds associated with the irrigated maize fields in this study. This indicates that these weed species are ecologically significant in the surveyed areas. Infestation of these weed species may increase competition with maize for growth resources, resulting in severe yield losses if not effectively controlled. Cyperus rotundus is a major concern since it is difficult to control due to its hardy underground stems. Travlos et al. (2018) revealed that weed species with high relative densities and importance values can compete better in an agricultural field than other weed species. The predominance of Poaceae among the identified families showed its importance in the maize ecosystem. The species of this family produce abundant seeds and exhibit aggressive growth and efficient seed dispersal (Silva et al., 2015). These qualities enable its members to occupy ecological niches in environments favourable for plant growth. The highest importance value exhibited by C. rotundus refers to its damaging effect on the maize crop. According to Silva et al. (2015), C. rotundus thrives well in a humid, shaded or open place and can form an obnoxious weed community, competing with the crop for space and nutrients. The propensity to regenerate via seeds, rhizomes, tubers, and basal bulbs also contributed to its widespread distribution. Its propagules can remain dormant for extended periods, allowing it to survive periodic flooding or dry seasons. These characteristics give it an edge over other weed species in a specific location (Henry et al., 2021).
The high importance values in P. oleracea, D. horizontalis, B. deflexa, S. obtusifolia, A. conyzoides, C. dactylon. P. niruri and Eragrostis sp. suggest that they are highly aggressive weeds in the maize crop cycle and must be eradicated. In the phytosociological weed surveys carried out in some maize fields, Khan et al. (2012) and Khatam et al. (2013) also reported that some of these weeds exhibited high importance values in the weed community. Moreover, the coexistence and abundance of Poaceae and Asteraceae weeds in some fields support the findings of Štefanić et al. (2019) from a survey of arable areas on the Istria Peninsula. Some species such as A. conyzoides, O. corymbosa, and P. oleracea have a short life cycle (annual) that allows them to reproduce quickly, giving them the potential for a very high intrinsic rate of increase (Global Invasive Species Database, 2022). In addition, A. conyzoides seeds germinate at varying temperatures to ensure survival and persistence (Paul et al., 2022). Therefore, its remarkable physiological adaptability has also contributed to its prevalence in agricultural farmlands. Although O. corymbosa and P. oleracea are annual plants, they can act as short-lived perennials under favourable conditions (India Biodiversity Portal, 2018). In a study, Mercado and Lapitan (2018) reported abundant A. conyzoides and P. oleracea in maize and rice intercropped fields, similar to what we found in this study. The ability of these weeds to produce and disperse thousands of seeds over a long distance makes them successful in infested farmlands. Spermacoce ocymoides and P. niruri, like O. corymbosa and P. oleracea, produce numerous seeds (Mercado and Lapitan, 2018; Tropical Plants Database, 2022). At the onset of the wet season, the seeds germinate and emerge quickly. Furthermore, they are well-suited to hydromorphic soils, and mature plants may withstand floods for several years without harm.
The uneven distribution of grass, sedge and broadleaf weeds throughout the surveyed sites was caused partly by different weed management strategies. Furthermore, broadleaf weeds were more prevalent than grass and sedge species due to their smothering effect on grass and sedge species. The positive association of PCs with weed species demonstrated that increasing the abundance of one of them increases the prevalence of the others. This is consistent with the findings of Ciaccia et al. (2019) in a study with C. rotundus and P. oleracea. Although crop combinations aimed at weed management should consider soil fertility sustainability, none of the crops intercropped with maize possessed this attribute. The composition and distribution of identified weed species demonstrated the impact of maize farmers’ cropping systems. Setaria pumila’s significant association with sole maize cultivation, for example, suggested that weeds in the same family as a crop could be damaging to its development and production performance.
Because it belongs to the same family as the cassava component, the appearance of E. hirta in maize and cassava intercropped fields could be attributed to the same reason. Furthermore, the unique interaction of S. pumila with sole maize farmlands may be related to seed dispersal ease, long-term longevity, and fast germination and emergence (Dowsett et al., 2018). Portulaca oleracea, on the other hand, belongs to a different family (Portulacaceae) yet has been identified as a significant maize weed. This is consistent with the findings of Mendes et al. (2018), who reported it as a prominent maize weed. Furthermore, P. oleracea produces allelopathic compounds that inhibit the growth of other plants in its vicinity. In another study, Rashidi et al. (2021) demonstrated that P. oleracea reduced the germination of common bean (Phaseolus vulgaris L.) and onion seeds (Allium cepa L.). Desmodium scorpiurus, I. triloba, S. anthelmia, and M. villosus, on the other hand, produce a large number of seeds as a mechanism of dissemination and perpetuation in maize and rice intercropped fields. Consequently, the shading effect of their broad leaves limits the proportion of sunlight required for grass weed growth and development. The same reason likely explains the abundance of broadleaf weeds in maize and amaranthus intercropped, and maize and pepper cultivation systems. Cynodon. dactylon reproduces through seeds (Ngondya et al., 2019). It has a root system that can grow deep into the soil under drought conditions. These characteristics most likely contributed to its survival and abundance under the maize and pepper intercropping system. The maize and rice intercropping system resulted in mild grass infestation (B. deflexa), most likely due to the preponderance of broadleaf weeds in the invaded fields. This contradicts the findings of Henry et al. (2021), who found that intercropping maize with rice increased the incidence of grass weeds.
In conclusion, C. rotundus, P. oleracea, D. horizontalis, B. deflexa, S. obtusifolia, A. conyzoides, C. dactylon, P. niruri, and Eragrostis sp. were the most important weed species in the investigated locations. Poaceae was the family with the most members. According to morphology, broadleaf species ac-counted for 38.2 % of the total species studied. Anguwan-Shaba_01 and Katcha_01 had the most grass weeds (100 individuals), while Sabon-Gida_01 and Anguwan-Shaba_01 had the most sedge (158 individuals) and broadleaf (268 individuals) weeds, respectively. Setaria pumila was strongly associated with sole maize cultivation, as was D. scorpiurus with maize and rice inter-cropping, C. dactylon with maize and pepper cultivation, E. hirta with maize and cassava inter-cropping, and C. viscosa with maize and amaranthus cultivation. These weed species are trouble-some and have the potential to impair maize productivity. Therefore, site-specific and weed-specific control measures should be prioritized when developing a sustainable weed management plan for maize production. Farmers should desist from excessive herbicide application to avoid deleterious impacts on humans, livestock and the environment.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
MS, ED, OO, TF, and JB contributed equally to undertaking this project. All authors contributed to the article and approved the submitted version.
The authors are grateful to the Tertiary Education Trust Fund (TETFund), Federal Government of Nigeria, for funding this project (Ref: TETF/ES/DR&D-CE/NRF2020/SETI/26/VOL.1).
The management of the Federal University of Technology, Minna is appreciated for the enabling environment to undertake the study. We also thank all the farmers who allowed us to collect weeds from their farms.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Akobundu I. O., Ekeleme F., Agyakwa C. W., Ogazie C. A. (2016). A handbook of West African weeds. 3rd ed. (Ibadan- Nigeria: IITA), 521pp.
Bjornlund V., Bjornlund H., Van Rooyen A. F. (2020). Why agricultural production in sub-Saharan Africa remains low compared to the rest of the world – a historical perspective. Int. J. Water Resour. Dev. 36, S20–S53. doi: 10.1080/07900627.2020.1739512
Chauhan B. S. (2020). Grand challenges in weed management. Front. Agron 1. doi: 10.3389/fagro.2019.00003
Ciaccia C., La Torre A., Ferlito F., Testani E., Battaglia V., Salvati L., et al. (2019). Agroecological practices and agrobiodiversity: A case study on organic orange in southern Italy. Agron 9, 85. doi: 10.3390/agronomy9020085
Colbach N., Petit S., Chauvel B., Deytieux V., Lechenet M., Munier-Jolain N., et al. (2020). The pitfalls of relating weeds, herbicide use, and crop yield: Don't fall into the trap! a critical review. Front. Agron. 2. doi: 10.3389/fagro.2020.615470
Dowsett C., James T. K., Buddenhagen C. E. (2018). Yellow bristle grass (Setaria pumila) germination. N. Z. Plant Prot 71, 72–80. doi: 10.30843/nzpp.2018.71.154
FAO, IFAD, UNICEF, WFP, WHO (2019). The state of food security and nutrition in the world 2019. safeguarding against economic slowdowns and downturns (Rome: FAO).
Gharde Y., Singh P. K., Dubey R. P., Gupta P. K. (2018). Assessment of yield and economic losses in agriculture due to weeds in India. Crop Prot. 107, 12–18. doi: 10.1016/j.cropro.2018.01.007
Global Invasive Species Database (2022) Ageratum conyzoides. Available at: http://www.iucngisd.org/gisd/species.php?sc=1493 (Accessed March 29, 2022).
Grote U., Fasse A., Nguyen T. T., Erenstein O. (2021). Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 4. doi: 10.3389/fsufs.2020.617009
Henry G. M., Elmore M. T., Gannon T. W. (2021). Cyperus esculentus and Cyperus rotundus. In: Biology and Management of Problematic Crop Weed Species, Chauhan B. S. ed, (Massachusetts: Academic Press) pp 151 – 172.
India Biodiversity Portal (2018) Biodiversity. Available at: http://indiabiodiversity.org/species/list (Accessed March 29, 2022).
John K. N., Valentin V., Abdullah B., Bayat M., Kargar M. H., Zargar M. (2020). Weed mapping technologies in discerning and managing weed infestation levels of farming systems. Res. Crops 21, 93–98. doi: 10.31830/2348-7542.2020.015
Khan N. W, Khan N., Khan I. A. (2012). Integration of nitrogen fertilizer and herbicides for efficient weed management in maize crop. Sarhad J. Agric 28, 457–463.
Khatam A., Nawab K., Ahmad W., Khan M. Z., Mian I. A. (2013). Effect of various herbicides and manual control on yield, yield components and weeds of maize. Pak. J. Weed Sci. Res 19, 209–216. doi: 10.28941/pjwsr.v19i2.498
Korav S., Dhaka A. K., Singh R., Premaradhya N., Reddy G. C (2018). A study on crop weed competition in field crops. J. Pharmacogn. Phytochem. 7, 3235–3240. doi: 10.22271/phyto
Lopes C. C., Fontes C. L. O., Lazzarini L. E. S., Freitas F. C. L., Filho J. H. C., Sousa E. R. (2021). Phytosociological survey of weed plants in soybean culture in the gurguéia valley. Paranaensis -Sci Agrar. 20, 75–80. doi: 10.18188/sap.v20i1.25964
Mendes W. S., Filho L. O. V., Pereira N. A., Boechat C. L., Mielezrski F. (2018). Phytosociology and behavior of weeds in maize as influenced by spatial arrangements. J. Agric. Sci. 10, 199–208. doi: 10.5539/jas.v10n9p199
Mercado M. J. T., Lapitan V. C. (2018). Interaction of weed composition and species abund ance under different crop rotation pattern. J. Bio. Env. Sci. 13, 59–70. doi: 10.13140/RG.2.2.28498.40641
Ngondya I. B., Treydte A. C., Ndakidemi P. A., Munishi L. K. (2019). Can Cynodon dactylon suppress the growth and development of the invasive weeds Tagetes minuta and Gutenbergia cordifolia? Plants 8, 576. doi: 10.3390/plants8120576
Nkoa R., Owen M. D. K., Swanton C. J. (2015). Weed abundance, distribution, diversity, and community analyses. Weed Sci. 63, 64–90. doi: 10.1614/WS-D-13-00075.1
Paul S., Datta B. K., Ratnaparkhe M. B., Dholakia B. B. (2022). Turning waste into beneficial resource: Implication of Ageratum conyzoides l. @ in sustainable agriculture, environment and biopharma sectors. Mol. Biotechnol. 64, 221–244. doi: 10.1007/s12033-021-00409-5
Ranganathan J., Waite R., Searchinger T., Craig Hanson C. (2018) How to sustainably feed 10 billion people by 2050, in 21 charts. Available at: https://www.wri.org/insights/how-sustainably-feed-10-billion-people-2050-21-charts (Accessed March 30, 2022).
Rashidi S., Yousefi A. R., Goicoechea N., Pouryousef M., Moradi P., Vitalini S., et al. (2021). Allelopathic interactions between seeds of Portulaca oleracea l. and various crop species. Appl. Sci. 11, 3539– 3549. doi: 10.3390/app11083539
Restuccia A., Scavo A., Lombardo S., Pandino G., Fontanazza S., Anastasi U., et al. (2020). Long-term effect of cover crops on species abundance and diversity of weed flora. Plants 9, 1506–1520. doi: 10.3390/plants9111506
Sabanci K., Aydin C. (2017). Smart robotic weed control system for sugar beet. J. Agr Sci. Tech. 19, 73–83. doi: 20.1001.1.16807073.2017.19.1.6.1
Schütte G., Eckerstorfer M., Rastelli V., Reichenbecher. W., Restrepo−Vassalli S., Ruohonen−Lehto M., et al. (2017). Herbicide resistance and biodiversity: agronomic and environmental aspects of genetically modified herbicide−resistant plants. Environ. Sci. Eur. 29, 1–12. doi: 10.1186/s12302-016-0100-y
Silva M. R. M., Cantanheda. J. D., Correa M. J. P., Mesquita M. L. R. (2015). Phytosociology and interference of weeds in upland rice in maranhao state, northeastern Brazil. Afr. J. Agric. Res. 10, 3412–3420. doi: 10.5897/AJAR2015.9916
Silva M. R. M., Costa E. A., Correa M. J. P., Rodrigues A. A. C., Mesquita M. L. R. (2017). Floristic and phytosociology of weeds in upland rice in the humid tropics. Planta Daninha 35, 1–11. doi: 10.1590/s0100-83582017350100083
Sims B., Corsi S., Gbehounou G., Kienzle J., Taguchi M., Theodor F. T. (2018). Sustainable weed management for conservation agriculture: Options for smallholder farmers. Agriculture 8, 118. doi: 10.3390/agriculture8080118
Sintayehu A. (2019). Weed flora survey in field crops of northwestern Ethiopia. Afr. J. Agric. Res. 14, 749–758. doi: 10.5897/AJAR2019.13947
Štefanić E., Kovačević V., Antunović S., Japundžić-Palenkić B., Zima D., Turalija A., et al. (2019). Floristic biodiversity of weed communities in arable lands of istria peninsula (from 2005 to 2017). Ekol. Bratisl. 38, 166–177. doi: 10.2478/eko-2019-0013
Travlos I. S., Cheimona N., Roussis I., Bilalis D. J. (2018). Weed-species abundance and diversity indices in relation to tillage systems and fertilization. Front. Environ. Sci. 6. doi: 10.3389/fenvs.2018.00011
Tropical Plants Database (2022) Spermacoce ocymoides. Available at: https://tropical.theferns.info/viewtropical.php?id=Spermacoce+ocymoides (Accessed April 30, 2022).
UNDESAPPD (United Nations, Department of Economic and Social Affairs, Population Division) (2019). World population prospects 2019: Highlights (ST/ESA/SER.A/423). (New York: United Nations).
Waha K., Dietrich J. P., Portmann F. T., Siebert S., Thornton P. K., Bondeau A., et al. (2020). Multiple cropping systems of the world and the potential for increasing cropping intensity. Glob. Environ. Change 64, 1–13. doi: 10.1016/j.gloenvcha.2020.102131
WBGU (2020) Rethinking land in the anthropocene: From separation to integration (Berlin: Advisory Council on Global Change (WBGU). Available at: https://www.wbgu.de/en/publications/publication/landshift (Accessed April 30, 2022).
Keywords: allelopathy, density, dominance, importance value, principal component, cropping system
Citation: Salaudeen MT, Daniya E, Olaniyi OM, Folorunso TA, Bala JA, Abdullahi IM, Nuhu BK, Adedigba AP, Oluwole BI, Bankole AO and Macarthy OM (2022) Phytosociological survey of weeds in irrigated maize fields in a Southern Guinea Savanna of Nigeria. Front. Agron. 4:985067. doi: 10.3389/fagro.2022.985067
Received: 03 July 2022; Accepted: 27 September 2022;
Published: 13 October 2022.
Edited by:
Daizy Rani Batish, Panjab University, IndiaReviewed by:
Amarpreet Kaur, Panjab University, IndiaCopyright © 2022 Salaudeen, Daniya, Olaniyi, Folorunso, Bala, Abdullahi, Nuhu, Adedigba, Oluwole, Bankole and Macarthy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammadu Tajudeen Salaudeen, bXRzYWxhdWRlZW5AZnV0bWlubmEuZWR1Lm5n
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.