 Marie-Josée Simard*
Marie-Josée Simard* Lydia Maheux
Lydia Maheux- Agriculture and Agri-Food Canada, Saint-Jean-sur-Richelieu Research and Development Centre, Saint-Jean-sur-Richelieu, QC, Canada
Roadsides are vectors of spread for invasive and other non-native plants. Therefore, fields located along roadsides could harbour more weeds and less native species compared to more isolated fields. To determine if field edges that are close to roadsides have different floras compared to more isolated fields, we surveyed 26 field pairs (52 fields) located in the province of Québec, Canada. For each pair, one field could be directly accessed by a major paved road (AD fields) while the other field, located on the same farm, was more isolated (IS fields) and could only be accessed via a secondary farm road. Two borders of these fields (IS) were sampled as well as the parallel (AD-pa) and the perpendicular border (AD-pe) of A fields and the adjacent roadside (RO). Plant species present along these field borders were recorded and classified (e.g. non-native, native, monocot, dicot, annual, perennial) in 0.5 m2 quadrats located every 20 m. The number of common ragweed (Ambrosia artemisiifolia) plants was also noted. Analyses included linear mixed models, generalized estimating equations models and multiple correspondence analyses. All border types had equivalent species richness. Roadsides had higher densities of common ragweed and lower occurrences of native and perennial species compared to field edges. All analyses indicate roadside floras are different from field edge floras but field edges close to roadsides were similar to those of more isolated fields. Results do not support a simple diffusive spread of roadside plants into field crop edges since field edges located along roadsides did not harbour more common ragweed plants or more roadside species.
Introduction
Roadsides generally harbour more invasive and non-native species than the adjacent landscapes (Tyser and Worley, 1992; Hansen and Clevenger, 2005; Barbosa et al., 2010). Species are dispersed during road construction, maintenance and use as propagules travel on and in vehicles and are dispersed by vehicle airflow (Taylor et al., 2012; Ansong and Pickering, 2013; von der Lippe et al., 2013; Rauschert et al., 2017; Khan et al., 2018). Plants also spread without human assistance along the edge of these corridors where light conditions, disturbance and soil properties are altered (Forman and Alexander, 1998; Spellerberg, 1998). Roads can therefore facilitate the spread of invasive and other non-native species including weeds into the adjacent landscape (Tyser and Worley, 1992; Gelbard and Belnap, 2003; Joly et al., 2011; Meunier and Lavoie, 2012; Follak et al., 2018; Speziale et al., 2018). These species can reduce biodiversity and modify ecosystems (Gaertner et al., 2009). Although there are multiple cases of increased species richness by invasive and non-native plants (Sax et al., 2002), especially in agricultural landscapes (Lázaro-Lobo and Ervin, 2019), these additions are not necessarily beneficial (Benvenuti, 2007). Although the impact of road networks on natural landscapes and grasslands has been evaluated (Tyser and Worley, 1992; Irl et al., 2014), few studies have evaluated the effect of the proximity of paved roads on the composition of intensively managed field crop borders.
In the province of Québec, Canada, common ragweed (Ambrosia artemisiifolia L., Asteraceae) is particularly abundant along roadsides, especially along paved roads where densities can average 16 plants m-2 (Simard and Benoit, 2010; Joly et al., 2011). The species is native to central North America and invasive in European countries such as Austria, France, Hungary, Italy and Switzerland as well as China and Australia (Bassett and Crompton, 1975; Gaudeul et al., 2011; Afonin et al., 2018). In Austria, 153 to 728 seeds m-2 are reported to be representative of roadside densities (Milakovic et al., 2014). The species is still increasing in abundance in Eastern Canada (Lavoie et al., 2007) and abundant roadside populations could potentially increase the likelihood of finding the species in adjacent field borders (Simard and Benoit, 2010). Common ragweed seeds do not have specialised dispersal structures, such as plumes and wings, and are mainly spread by passive transport on soil stuck on vehicles or as a contaminant of manure, seed and feed (Bassett and Crompton, 1975; Frick et al., 2011; Chauvel et al., 2012). If seeds on roadsides get stuck on tractor wheels or other equipment, most of these will disperse in the first field the tractor goes through. Water drained from roadsides can also flood into adjacent fields and smaller common ragweed seeds do float (Fumanal et al., 2007). Moreover, roadside mowing has been shown to spread common ragweed seeds (Vitalos and Karrer, 2009). The species is also very competitive and resistant to various environmental factors prevailing along roadsides such as high de-icing salt concentrations (DiTommaso, 2004) and heavy metal contaminants (Bae et al., 2016).
Our goal was to determine if field borders that are adjacent to roadsides had higher densities of common ragweed compared to fields that are more isolated and if a higher occurrence of non-native species was observed along with a lower account of native species in these adjacent field borders. Our main hypotheses were that ragweed density would be higher in field borders adjacent to roadsides and that these borders have more non-native and less native species compared to those of more isolated fields.
Materials and methods
From 2008 to 2012, 26 field pairs located along the agricultural area of the province of Québec, Canada were selected and surveyed once (Figure 1). Each field pair had to be on the same farm and as similar as possible (crop and soil type). One field had to be adjacent to a paved public road (field AD) and the other had to be accessible only by farm roads and be surrounded by other fields or forested land (field IS) (Figure 2). For each field, we recorded the crop grown, the herbicides applied, the presence of water in the ditches, the type of habitat on the other side of the border (forest, crop type, pasture etc.). A tabular view of the variables is presented in the Supplementary Table 1. The distance between the field border closest to the roadside and the roadside boundary was also recorded. For each field pair, all species present were recorded and the density of Ambrosia artemisiifolia was evaluated in 0.5 m x 1 m quadrats (longer quadrat edge perpendicular to road or field border) located every 20 m along 1-the roadside section bordering the adjacent field (RO), 2-the edge of the adjacent field parallel to the roadside (AD-pa), 3-the edge of the adjacent field perpendicular to the roadside (AD-pe), 4-a field edge of the isolated field (IS) (Figure 2). All sampling was done in July-August after herbicide applications. All the species recorded were classified either as native or non-native based on the Database of Vascular Plants of Canada (VASCAN) (Brouillet et al., 2010) or as a crop species. All species were also classified as monocotyledonous, dicotyledonous or other (non-flowering plants) and as annual, perennial or biennial based on available life-cycle descriptions in published Canadian floras. Some plants were recorded at the gender level when structures needed for species level identification (often flowers) were absent. Species richness was based on the sum of all species present. The number of unique species present in two consecutive quadrats was computed in order to present species richness per square meter.
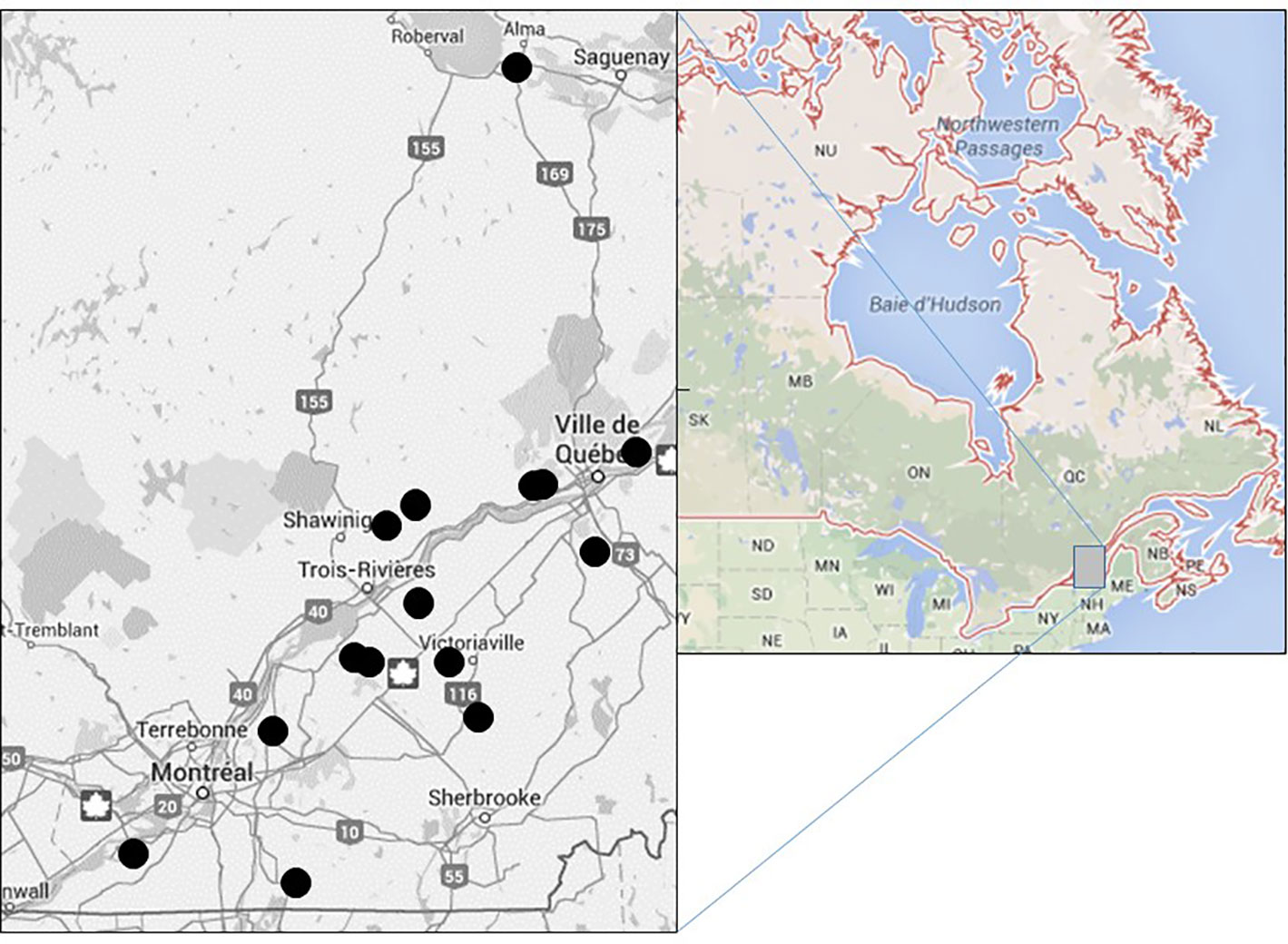
Figure 1 Location of field pairs (black circles, left map) and sampling area in Canada (grey square, right map).
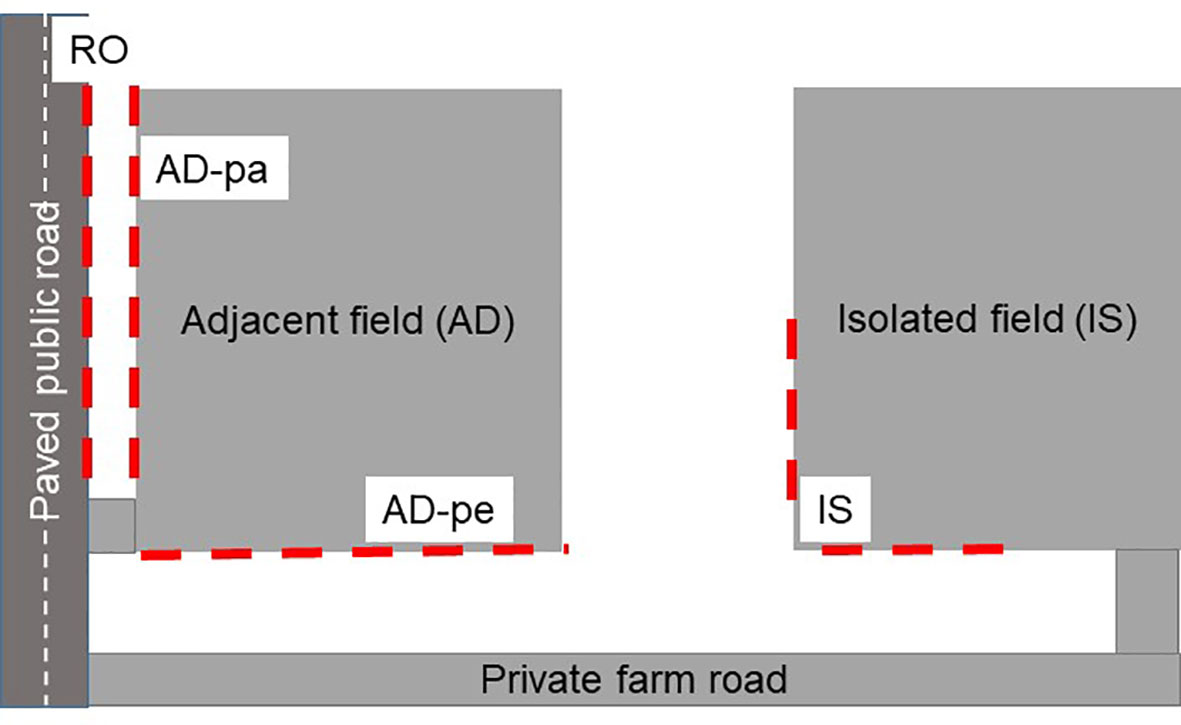
Figure 2 Schematic representation of field pairs (adjacent field and isolated field) and sampling transects (red dashed lines).
Statistical analyses
Database management and descriptive statistical analyses were performed using JMP Professional 15.0 Statistical Analysis Hardware (SAS Institute Inc., Cary, NC). Explanatory variables such as the type of bordering habitat on the other side of the transect or the herbicide applied had to be discarded due to the high number of different categories or low frequency of some categories. Common ragweed density and species counts [total and by type (weedy, native, perennial etc)] as a function of location (roadside, field border parallel to roadside, field border perpendicular to roadside, field border of field with no access to a public road) were evaluated using linear mixed models (proc MIXED) in SAS Version 9.4 (SAS Institute Inc., Cary, NC). Fixed effects included location and random effects included producer, year, field and quadrat. Homogeneity of variance and normality of residuals were verified. Heterogeneous variances were modeled according to group when needed, logarithmic transformation was applied to common ragweed densities to meet these criteria, and means on raw data are presented. Statistical significance between multiple comparisons were adjusted with Bonferonni correction. For presence/absence data, a generalized estimating equations model was used with a logit link and binomial distribution to evaluate the effect of location on the frequency of species in different groups (classified as native, non-native, perennial etc., as above). To identify locations with similar weed frequency profiles and associations between weed frequencies we performed a multiple correspondence analysis (MCA) on the species with a frequency of at least 5%. The Multiple correspondence analysis (MCA) is a generalization of principal component analysis when the variables to be analyzed are categorical (Abdi and Williams, 2010). We use weed category (species and presence/absence) as active variables and location as supplementary variables. We lused the R package FactoMineR for MCA analysis and the factoextra R package to help in interpretation and visualization. To support the classification generated by the multiple correspondence analysis, we also undertook a cluster analysis using the R package ClustOfVar (31).
Results
A total of 1350 quadrats were sampled. Fields were cultivated in corn (39%), forage crops (26%), soybean (16%) or cereal (16%). The field border closest to the roadside was at an average distance of 8.58 m ±0.16 (STE). A total of 167 species were recorded. The most frequently recorded species along field borders were Taraxacum officinale (present in 54% of quadrats), followed by Equisetum arvense (44%), Trifolium repens (29%) and Vicia cracca (27%). On roadsides, the most frequently recorded species were Ambrosia artemisiifolia (61%), followed by grasses [(Elymus repens (42%) and Poa sp. (30%)], Trifolium arvense (26%), Plantago major (26%) and Matricaria sp. (23%). All the species recorded are listed in the Supplementary Table 2.
Ragweed density
The type of border habitat had a significant effect on common ragweed abundance (p < 0.001). Common ragweed abundance was 11 times higher along paved roads than along field borders (68.79 plants vs 6.23 plants m-2) but all field borders had equivalent numbers regardless of their location in an isolated field or in a field border close to a paved road, whether parallel or perpendicular to the road (Figure 3).
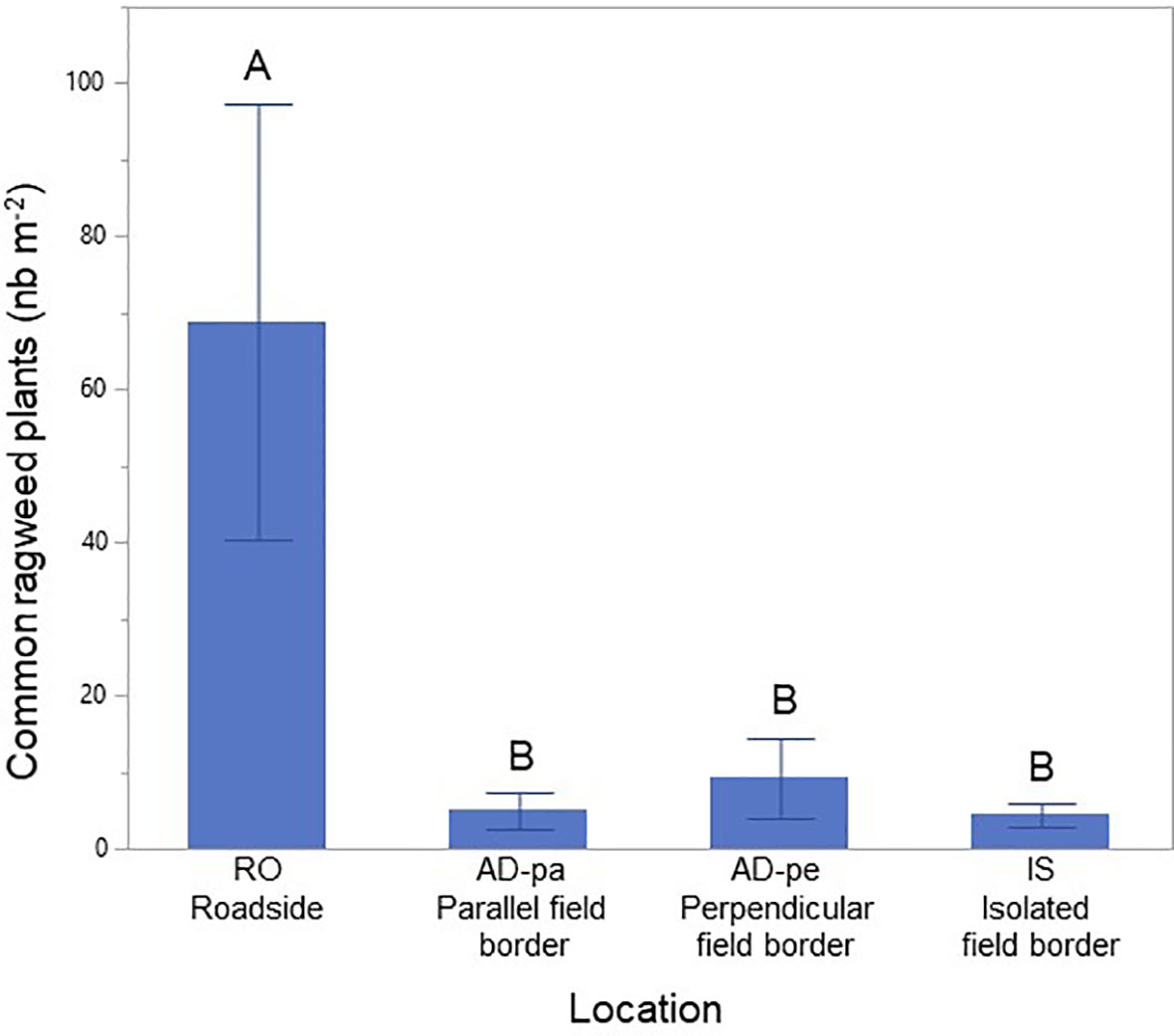
Figure 3 Average common ragweed (Ambrosia artemisiifolia) density along paved roadsides and field borders. Different letters indicate significant differences at alpha=0.05, Error bars indicate ±STE.
Species richness and composition
Species richness was equivalent between habitats (p=0.253), averaging 9.88 ± 0.28 (STE) (Figure 4). The frequency of non-native species was also equivalent between all habitats (p=0.280), averaging 7.03 ± 0.11 (STE) per square meter. At an average of 2.13 ± 0.13 (STE), the frequency of native species was lower along roadsides (p=0.023) compared to field edges and all field edges had equivalent numbers of native species, averaging 2.53 ± 0.07 (STE). At an average of 5.18 ± 0.23, perennial species (p=0.001) were also less recorded along roadsides compared to field habitats who averaged at 6.29 ± 0.14 (STE) (Figure 5). MCA analyses done on a subsample of 29 species (species with frequencies lower than 5% in the entire dataset were excluded) also indicate the species composition of paved roads was different from that of field borders with a stronger association along the second axe (Figure 6), supported by the cluster analysis (not shown), with the presence of Matricaria sp., Trifolium agrarium, Poa sp., Ambriosia artemisiifolia and Polygonum aviculare compared to the field borders. The field borders could not be clearly separated although isolated fields had a weak cluster of Oxalis stricta, Leucanthemum vulgare, Fragaria sp. and Cerastium fontanum (details not shown).
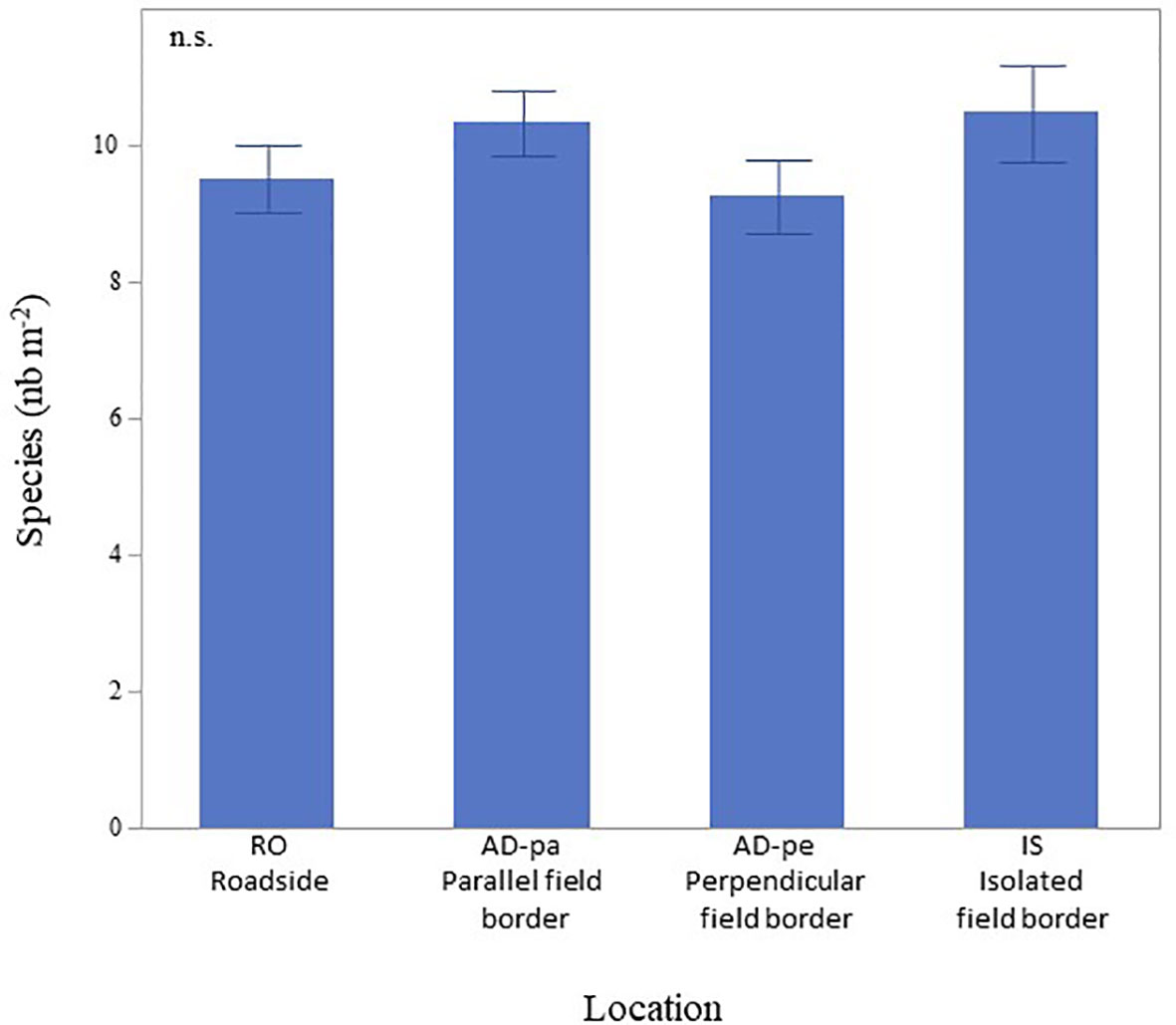
Figure 4 Species richness along roadsides and field borders. Values are not significantly different (n.s. p=0.3285). Error bars indicate ±STE.
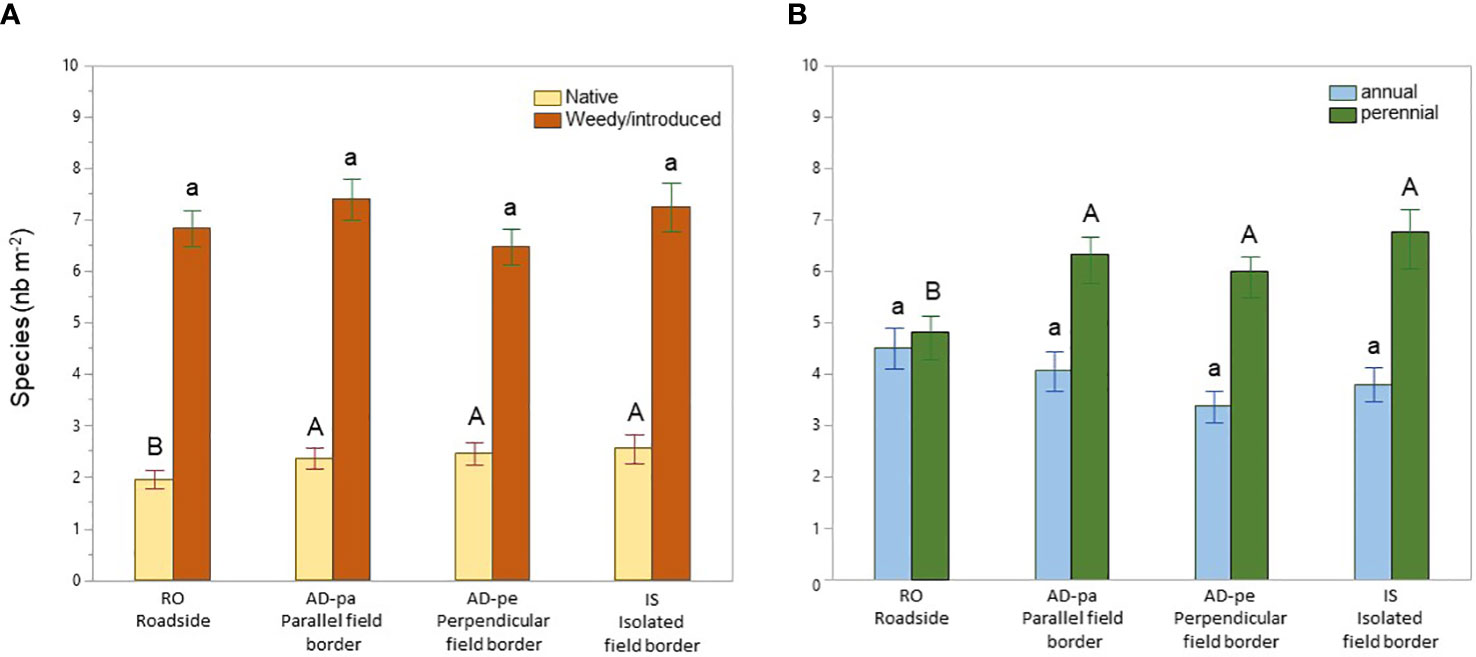
Figure 5 Species richness of native and weedy/introduced species (A) as well as annual and perennial species (B) along roadsides and field borders. Different letters indicate significant differences at alpha=0.05 within plant type, Error bars indicate ±STE.
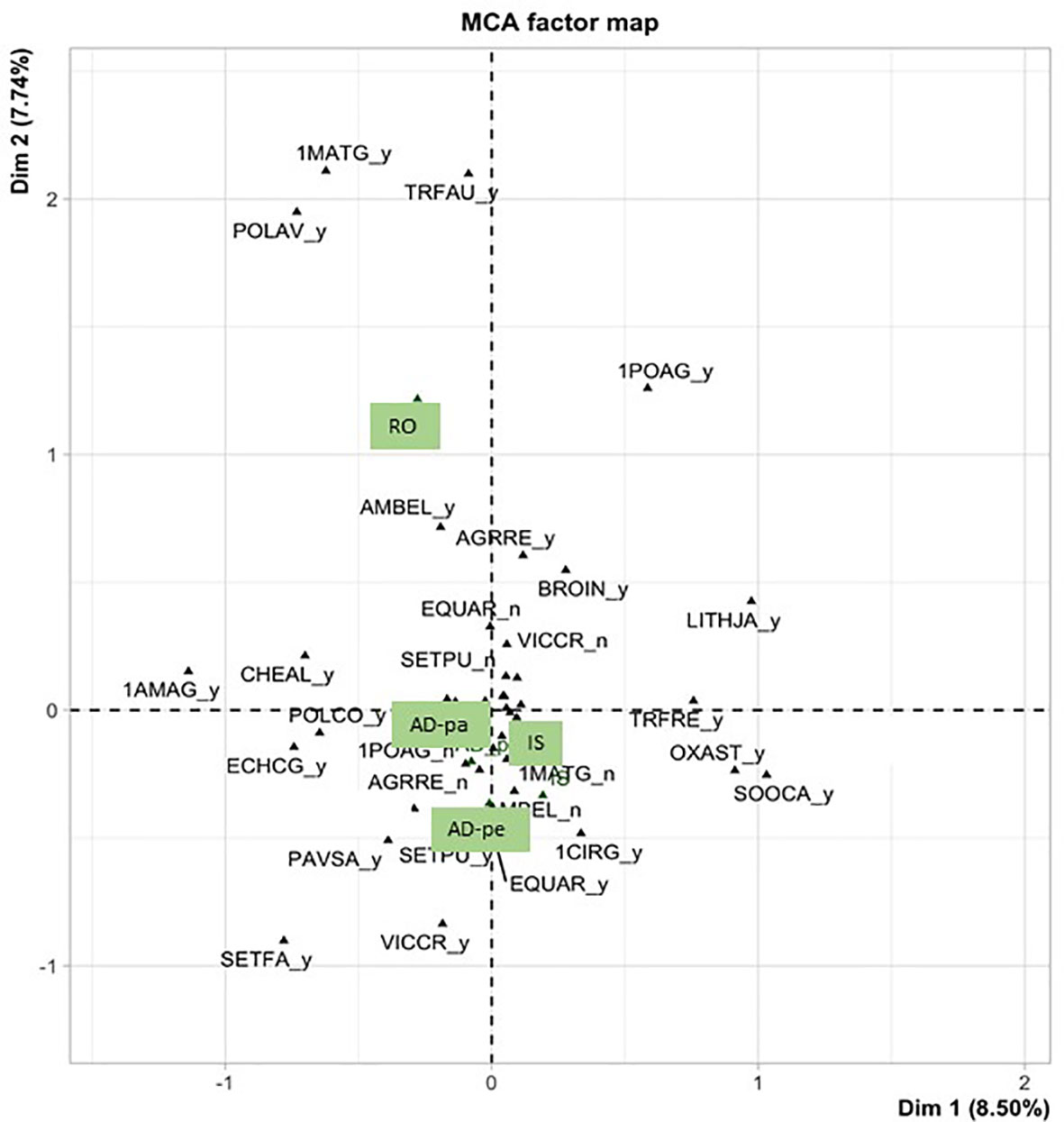
Figure 6 Biplot of the two first components of the multiple correspondence analysis (MCA) carried out on the presence (_y) or absence (_n) of the most frequent species observed along paved roadsides (RO) and field borders (AD-pa, Ad-pe and IS, see Figure 2). 1AMAG, Amaranthus sp; 1CIRG, Cirsium sp; 1MATG. Matricaria sp.; 1POAG, Poa sp; AMBEL, Ambrosia artemisiifolia; AGGRE, Elymus repens; BROIN,Bromus inermis; CHEAL, Chenopodium album; CHYLE, Leucanthemum vulgare; ECHCG, Echinochloa crus-galli, EQUAR, Equisetum arvense; LITHJA, Lathyrus japonicus; OXAST, Oxalis stricta; PAVSA,Pastinaca sativa; POLCO, Fallopia convolvulus; POLAV, Polygonum aviculare; SETFA, Setaria faberii; SETPU, Setaria pumila; SOOCA, Solidago Canadensis; TRFAU, Trifolium arvense; TRFRE, Trifolium repens; VICCR, Vicia cracca.
Discussion
Roadsides
Paved roadsides had equivalent species richness compared to field edges but could be distinguished from field borders by a ten fold higher density of common ragweed plants and a lower prevalence of native species and of perennial species. Higher common ragweed density along roadsides compared to field borders was expected (Simard and Benoit, 2010), especially since only paved roads were surveyed to increase the potential of finding evidence of non-native spread into field borders. Paved roads have been shown to harbour more non-native species than non-paved roads (Gelbard and Belnap, 2003) and more common ragweed (Joly et al., 2011). Roadsides floras are often not a subset of the surrounding vegetation (Corcos et al., 2020) as they generate disturbed habitats where the establishment of perennial species is impeded (Albert et al., 2013) and early successional species such as common ragweed establish and grow. Common ragweed plants can also germinate in high concentrations of roadside contaminants (DiTommaso, 2004; Bae et al., 2016). Car traffic and roadside mowing after seed formation will further increase the spread of roadside species (Christen and Matlack, 2006), as evidenced by empirical studies on common ragweed dispersal (Vitalos and Karrer, 2009; von der Lippe et al., 2013; Lemke et al., 2021). Although non-native diversity along roadsides is possibly higher than in the surrounding vegetation (Tyser and Worley, 1992) and as been found to be negatively associated with native plant diversity (Peltzer and MacLeod, 2014), non-native diversity was not higher along the surveyed roadsides than in field crop borders. This could be explained by the fact that most weedy species in Canada are non-native (Darbyshire, 2003) and that most fields were grown with annual crops (74% of surveyed fields), which are subject to higher disturbance regimes and open conditions facilitating weed growth compared to perennial crops. Roadsides are also planted with non-native and native turf-grass species that include Poa pratensis and perennial legumes such as Trifolium arvense (Bae et al., 2015), thus explaining their association with these species compared to field borders.
Field borders
Field borders, whether parallel or perpendicular to an adjacent roadside or along fields that were more isolated had equivalent species richness. Contrary to our hypothesis, field edges close to paved roadsides did not harbour higher densities of common ragweed or more non-native and less native species compared to those of more isolated fields and the orientation of the border to the roadside did not influence results. Moreover, field borders closer to roadsides were not clearly associated with different plant species compared to borders surrounding more isolated fields. Common ragweed densities and field edge species composition are probably modulated more by past and present weed control strategies in fields, or field edge characteristics, than the presence of bordering roadside populations. Additionally, weed populations have been shown not to follow simple diffusive dispersal patterns of spread because of stochasticity from human mediated dispersal (Sullivan et al., 2009; Barnaud et al., 2013), especially given the long history of introduction of weedy species in the surveyed area (Bassett and Crompton, 1975; Lavoie et al., 2007). Results do not support a simple diffusive spread of roadside plants into field crop edges.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
M-JS, funding acquisition, project administration, conceptualisation, data curation, partial analyses, and writing; LM data curation, review and editing. Both authors contributed to the article and approved the submitted version.
Funding
This study was funded by Agriculture and Agri-Food Canada grant E.1211.NZ.
Acknowledgments
We thank Geneviève Bégin, Aline Philibert and all the students and producers who participated in the survey.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2022.1005093/full#supplementary-material
References
Abdi H., Williams L. J. (2010). Principal component analysis. Wiley interdisciplinary reviews. Comput. Stat. 2 (4), 433–459. doi: 10.1002/wics.101
Afonin A. N., Luneva N. N., Fedorova Y. A., Kletchkovskiy Y. E., Chebanovskaya A. F. (2018). History of introduction and distribution of common ragweed (Ambrosia artemisiifolia l.) in the European part of the Russian federation and in the Ukraine. Bull. OEPP 48 (2), 266–273. doi: 10.1111/epp.12484
Albert A., Brisson J., Dubé J., Lavoie C. (2013). Do woody plants prevent the establishment of common reed along highways? insights from southern Quebec. Invasive Plant Sci. Manage. 6 (4), 585–592. doi: 10.1614/IPSM-D-13-00025.1
Ansong M., Pickering C. (2013). Are weeds hitchhiking a ride on your car? a systematic review of seed dispersal on cars. PloS One 8 (11), e80275. doi: 10.1371/journal.pone.0080275
Bae J., Benoit D. L., Watson A. K. (2016). Effect of heavy metals on seed germination and seedling growth of common ragweed and roadside ground cover legumes. Environ. pollut. 213, 112–118. doi: 10.1016/j.envpol.2015.11.041
Bae J., Byun C., Watson A. K., Benoit D. L. (2015). Ground cover species selection to manage common ragweed (Ambrosia artemisiifolia l.) in roadside edge of highway. Plant Ecol. 216, 263–271. doi: 10.1007/s11258-014-0433-9
Barbosa N. P. U., Wilson Fernandes G., Carneiro M. A. A., Lélis A. C. Jr. (2010). Distribution of non-native invasive species and soil properties in proximity to paved roads and unpaved roads in a quartzitic mountainous grassland of southeastern Brazil (rupestrian fields). Biol. Invasion. 12, 3745–3755. doi: 10.1007/s10530-010-9767-y
Barnaud A., Kalwij J. M., Berthouly-Salazar C., McGeoch M. A., Jansen van Vuuren B. (2013). Are road verges corridors for weed invasion? insights from the fine-scale spatial genetic structure of Raphanus raphanistrum. Weed Res. 53, 362–369. doi: 10.1111/wre.12033
Bassett I. J., Crompton C. W. (1975). The biology of Canadian weeds. 11. Ambrosia artemisiifolia l. and A. psilostachya DC. Can. J. Plant Sci. 55, 463–476. doi: 10.4141/cjps75-072
Benvenuti S. (2007). Weed seed movement and dispersal strategies in the agricultural environment. Weed Biol. Manage. 7, 141–154. doi: 10.1111/j.1445-6664.2007.00249.x
Brouillet L., Desmet P., Coursol F., Meades S. J., Favreau M., Anions M., et al. (2010) Database of vascular plants of Canada (VASCAN). Available at: http://data.canadensys.net/vascanhttp://www.gbif.org/dataset/3f8a1297-3259-4700-91fc-acc4170b27ce (Accessed July 20, 2022).
Chauvel B., Martinez Q., Guillemin J. P. (2012). “Importance of seeds in the process of common ragweed invasion,” in International Symposium on Current Trends in Plant Protection IZBIS Institut za zaštitu bilja I životnu sredinu. (Belgrade: Institute for Plant Protection and Environment).
Christen D., Matlack G. (2006). The role of roadsides in plant invasions: a demographic approach. Conserv. Biol. 20, 385–391. doi: 10.1111/j.1523-1739.2006.00315.x
Corcos D., Nascimbene J., Campesan M., Donadello D., Segat V., Marini L. (2020). Establishment dynamics of native and exotic plants after disturbance along roadsides. Appl. Veg. Sci. 23, 277–284. doi: 10.1111/avsc.12481
Darbyshire S. J. (2003). Inventory of Canadian agricultural weeds (Ottawa, Canada: Agriculture and Agri-Food Canada, Research Branch).
DiTommaso A. (2004). Germination behavior of common ragweed (Ambrosia artemisiifolia) populations across a range of salinities. Weed. Sci. 52, 1002–1009. doi: 10.1614/WS-04-030R1
Follak S., Schleicher C., Schwarz M. (2018). Roads support the spread of invasive Asclepias syriaca in Austria. Die Bodenkultur: J. Land Manage. Food Environ. 69 (4), 257–265. doi: 10.2478/boku-2018-0022
Forman R. T. T., Alexander L. E. (1998). Roads and their major ecological effects. Ann. Rev. Ecol. Syst. 29 (1), 207–231. doi: 10.1146/annurev.ecolsys.29.1.207
Frick G., Boschung H., Schulz-Schroeder G., Russ G., Ujčič-Vrhovnik I., Jakovac-Strajn B., et al. (2011). Ragweed (Ambrosia sp.) seeds in bird feed. Biotechnol. Agron. Soc Environ. 15 (S1), 39–44.
Fumanal B., Chauvel B., Sabatier A., Bretagnolle F. (2007). Variability and cryptic heteromorphism of ambrosia artemisiifolia seeds: What consequences for its invasion in France? Ann. Bot. 100 (2), 305–313. doi: 10.1093/aob/mcm108
Gaertner M., Den Breeyen A., Hui C., Richardson D. M. (2009). Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: a meta-analysis. Progr. Phys. Geogr. 33 (3), 319–338. doi: 10.1177/0309133309341607
Gaudeul M., Giraud T., Kiss L., Shykoff J. A. (2011). Nuclear and chloroplast microsatellites show multiple introductions in the worldwide invasion history of common ragweed, Ambrosia artemisiifolia. PloS One 6 (3), e17658.
Gelbard J. L., Belnap J. (2003). Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv. Biol. 17, 420–432. doi: 10.1046/j.1523–1739.2003.01408.x
Hansen M. J., Clevenger A. P. (2005). The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biol. Conserv. 125, 249–259. doi: 10.1016/j.biocon.2005.03.024
Irl S. D. H., Steinbauer M. J., Epperlein L., Harter D. E. V., Jentsch A., Patz S., et al. (2014). The hitchhiker’s guide to island endemism: biodiversity and endemic perennial plant species in roadside and surrounding vegetation. Biodivers. Conserv. 23, 2273–2287. doi: 10.1007/s10531-014-0722-6
Joly M., Bertrand P., Gbangou R. Y., White M.-C., Dubé J., Lavoie C. (2011). Paving the way for invasive species: Road type and the spread of common ragweed (Ambrosia artemisiifolia). Environ. Manage. 48, 514–522. doi: 10.1007/s00267-011-9711-7
Khan I., Navie S., George D., O’Donnell C., Adkins S. W. (2018). Alien and native plant seed dispersal by vehicles. Austral Ecol. 43, 76–88. doi: 10.1111/aec.12545
Lavoie C., Jodoin Y., Gourseaud de Merlis A. (2007). How does common ragweed (Ambrosia artemisiifolia l.) spread? a historical analysis using herbarium records. J. Biogeogr. 34, 1751–1761. doi: 10.1111/j.1365-2699.2007.01730.x
Lázaro-Lobo A., Ervin G. N. (2019). A global examination on the differential impacts of roadsides on native vs. exotic and weedy plant species. Global Ecol. Conserv. 17, e00555. doi: 10.1016/j.gecco.2019.e00555
Lemke A., Buchholz S., Kowarik I., Starfinger U., von der Lippe M. (2021). Interaction of traffic intensity and habitat features shape invasion dynamics of an invasive alien species (Ambrosia artemisiifolia) in a regional road network. NeoBiota 64, 55–175. doi: 10.3897/neobiota.64.58775
Meunier G., Lavoie C. (2012). Roads as corridors for invasive plant species: New evidence from smooth bedstraw (Galium mollugo). Invasive Plant Sci. Manage. 5 (1), 92–100. doi: 10.1614/IPSM-D-11-00049.1
Milakovic I., Fiedler K., Karrer G. (2014). Fine-tuning of a mowing regime, a method for the management of the invasive plant, Ambrosia artemisiifolia, at different population densities. Weed Biol. Manage. 14, 232–241. doi: 10.1111/wbm.12051
Peltzer D. A., MacLeod C. J. (2014). Weed syndromes on marginal lands. Austral Ecol. 39, 39–49. doi: 10.1111/aec.12043
Rauschert E. S. J., Mortensen D. A., Bloser S. M. (2017). Human-mediated dispersal via rural road maintenance can move invasive propagules. Biol. Invasion. 19, 2047–2058. doi: 10.1007/s10530-017-1416-2
Sax D. F., Gaines S. D., Brown J. H. (2002). Species invasions exceed extinctions on islands worldwide: a comparative study of plants and birds. Am. Nat. 160, 766–783. doi: 10.1086/343877
Simard M.-J., Benoit D. L. (2010). Distribution and abundance of an allergenic weed, common ragweed (Ambrosia artemisiifolia l.), in rural settings of southern québec, Canada. Can. J. Plant Sci. 90, 549–557. doi: 10.4141/CJPS09174
Spellerberg I. (1998). Ecological effects of roads and traffic: a literature review. global. Ecol. Biogeogr. Lett. 7, 317–333. doi: 10.1046/j.1466-822x.1998.00308.x
Speziale K. L., di Virgilio A., Lescano M. N., Pirk G., Franzese J. (2018). Synergy between roads and disturbance favour bromus tectorum l. invasion. PeerJ 6, e5529. doi: 10.7717/peerj.5529
Sullivan J. J., Williams P. A., Timmins S. M., Smale M. C. (2009). Distribution and spread of environmental weeds along new Zealand roadsides. New Z. J. Ecol. 33, 190–204.
Taylor K., Brummer T., Taper M. L., Wing A., Rew L. J. (2012). Human-mediated long-distance dispersal: an empirical evaluation of seed dispersal by vehicles. Divers. Distrib. 18, 942–951. doi: 10.1111/j.1472-4642.2012.00926.x
Tyser R. W., Worley C. A. (1992). Alien flora in grasslands adjacent to road and trail corridors in glacier national park, Montana (U.S.A.). Conserv. Biol. 6 (2), 253–262. doi: 10.1046/j.1523-1739.1992.620253.x
Vitalos M., Karrer G. (2009). “Dispersal of ambrosia artemisiifolia seeds along roads: the contribution of traffic and mowing machines,” in Biological invasions: Towards a synthesis, vol. 8 . Eds. Pyšek P., Pergl J. (Prague: Neobiota) 8, 53–60.
Keywords: field crop border, native plant, paved road, species richness, ragweed (Ambrosia artemisiifolia)
Citation: Simard M-J and Maheux L (2022) Roadsides and neighboring field edges harbor different weed compositions. Front. Agron. 4:1005093. doi: 10.3389/fagro.2022.1005093
Received: 27 July 2022; Accepted: 26 September 2022;
Published: 10 October 2022.
Edited by:
Simerjeet Kaur, Punjab Agricultural University, IndiaReviewed by:
Aleksandra Savic, Institute for Plant Protection and Environment (IZBIS), SerbiaAhmet Uludag, Çanakkale Onsekiz Mart University, Turkey
Copyright © 2022 Simard and Maheux. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie-Josée Simard, bWFyaWUtam9zZWUuc2ltYXJkQGFnci5nYy5jYQ==