 Adnane Bargaz
Adnane Bargaz Joshua Nasielski
Joshua Nasielski Marney E. Isaac
Marney E. Isaac Erik S. Jensen2
Erik S. Jensen2 Georg Carlsson
Georg Carlsson- 1Plant-Microbe Interactions Lab, Agrobiosciences, Mohammed VI Polytechnic University, Bengurir, Morocco
- 2Department of Biosystems and Technology, Swedish University of Agricultural Sciences, Alnarp, Sweden
- 3Department of Plant Agriculture, University of Guelph, Guelph, ON, Canada
- 4Department of Physical and Environmental Sciences, University of Toronto Scarborough, Toronto, ON, Canada
Commercial legume varieties vary in terms of their drought tolerance when grown as sole crops, though relatively little is known about how legume variety selection affects cereal–legume intercrop performance under drought conditions. This study aims to test the hypothesis that positive rhizosphere interactions in faba bean–wheat intercrops will confer a “buffering capacity” on faba bean and wheat performance under water stress and that this effect will (i) depend on faba bean varietal selection and (ii) be enhanced with increasing faba bean varietal diversity. In a greenhouse experiment, three commercial faba bean (Vicia faba L.) varieties [Gloria (G), Alexia (A), Julia (J)] were grown in sole crop or intercropped with spring wheat (Triticum aestivum L.) under well-watered or water-stress conditions. Under intercropping, either one, two, or all three faba bean varieties were grown together with wheat to test the effect of intraspecific diversity on a cereal–legume intercrop performance. Consistent with the proposed hypothesis, we found that, under well-watered and water-stress conditions, wheat and faba bean shoot biomass production and nitrogen (N) acquisition improved with intercropping and that faba bean variety and variety mixture strongly modulated the intercropping effect. Interestingly, in both well-watered and water-stress conditions, wheat dry biomass and N accumulation were greatest in intercrops containing Gloria, while nodule number, nodule weight, and N accumulation in faba bean were greatest for intercrops containing Alexia and Julia (AJ). The effect of varietal diversity was inconsistent. Intercrops with two faba bean varieties tended to have positive or neutral effects on measured wheat and faba bean variables. However, overall performance under intercropping was generally reduced when all three faba bean varieties were planted with wheat. The effect of faba bean species diversity can buffer faba bean–wheat intercrop performance against water stress, and intercropping tended to have positive or neutral effects on the measured wheat and faba bean variables, notably with two-varietal faba bean mixtures.
Introduction
Intercropping is an agroecological land management practice where at least two crop species are grown on the same field at the same time (Wezel et al., 2014). Traditionally used by smallholder farmers to increase crop production, mitigate risks, and use land more efficiently, intercrops also hold great potential to increase the sustainability of modern crop production (Ghosh, 2004; Li et al., 2007; Bybee-Finley and Ryan, 2018). Intercropping cereals with grain legumes (i.e., pulses) are particularly promising because of (i) their socio-economic importance and key role in many human diets and (ii) the well-known complementarity of cereal–legume intercrops (Bedoussac et al., 2015). The greater land productivity attributed to cereal–legume intercrops is mainly due to the positive rhizosphere interactions between intercropped grasses and legumes, which enhances the efficiency of soil resource use, particularly nitrogen (N) (Hauggaard-Nielsen et al., 2010; Raseduzzaman and Jensen, 2017; Jensen et al., 2020). Symbiotic nitrogen fixation (SNF), performed by rhizobial bacteria in organs on legume roots called nodules, is a key agroecosystem service provided by legumes. Cereals can outcompete legumes for soil mineral N, thus reducing soil N availability and subsequently stimulating SNF of the legume (Unkovich and Pate, 2000). This has been shown to result in higher total N uptake per unit of land in cereal–legume intercrops compared to sole crops (Bulson et al., 1997; Hauggaard-Nielsen et al., 2008, 2009; Li et al., 2009; Lithourgidis and Dordas, 2010), particularly in perennial intercrops.
Nevertheless, the complementarity of soil resource use in cereal–legume intercrops is challenged by the fact that many grain legumes are sensitive to a broad spectrum of environmental constraints that can cause both spatial and temporal variation in legume growth and SNF (Androsoff et al., 1995; Carlsson and Huss-Danell, 2003). Water limitation is a major driver of legume yield instability (Hungria and Vargas, 2000; Hauggaard-Nielsen et al., 2008; Nasielski et al., 2015; Farooq et al., 2017) and nodule performance (Marino et al., 2007; Abdelhamid et al., 2011). However, rhizosphere interactions in diversified root systems of cereal–legume intercrops species have been shown to alleviate the impact of soil nutrient limitations (Li et al., 2007; Zhang et al., 2011; Bargaz et al., 2017; Bukovsky-Reyes et al., 2019). For example, legume SNF is also strongly dependent on phosphorus (P) availability, and in P-deficient soils, facilitation and complementarity effects on P use have been found in several types of legume–cereal intercrops (Hinsinger et al., 2011; Isaac et al., 2012; Shen et al., 2013). Improved rhizosphere nutrient availability is one of many advantages associated with cereals intercropped with nodulated legumes; however, possible links between rhizosphere interactions and SNF or yield stability under drought in cereal–legume intercrops involving faba bean (Vicia faba) have so far not been fully explored.
While broad trends in abiotic drivers of legume performance have been well-explored, the varietal breadth and its drivers of performance under intercropping of faba bean, a cool-season grain legume grown as a food and feed crop on an estimated 2.6 million ha worldwide (Jensen et al., 2010), remain less explored. Faba bean production is concentrated in China and Ethiopia, where smallholder farmers commonly intercrop faba bean with wheat (Triticum aestivum L.), a combination that tends to generate higher yield than when each crop is grown separately (Xiao et al., 2004; Agegnehu et al., 2008; Wang et al., 2017). Faba bean is of particular research interest because of its relatively high reliance on SNF as a source of N (Unkovich and Pate, 2000; Jensen et al., 2010). However, faba bean is considered more sensitive to water deficit than other grain legumes (McDonald and Paulsen, 1997; Amede et al., 2003; Khan et al., 2010). There exists considerable genotypic variation in drought tolerance among commercial faba bean varieties (Karkanis et al., 2018). For example, Alghamdi et al. (2015) found a large variation in water use efficiency among 20 faba bean cultivars from Egypt, the Kingdom of Saudi Arabia, Sudan, and Spain. In a study of 16 faba bean cultivars from across Europe, Zhao et al. (2018) identified large differences in root morphological response to an artificially imposed drought between the cultivars originating from northern (Estonia, Latvia, and Sweden) and southern (Portugal) Europe. The performance of faba bean–wheat intercrops in terms of rhizosphere complementarity, SNF, and N use under drought is probably modulated by cultivar selection.
In this study, we investigate the role of legume varietal diversity on legume and cereal performance under intercropping. Given enhanced rhizosphere heterogeneity with increasing intraspecific (varietal) and interspecific (intercropping) diversity, we expect stimulated SNF, N accumulation, and root biomass. We hypothesize that faba bean–wheat intercrops, and specifically, increasing faba bean varietal diversity will buffer biomass production and N acquisition against the negative effects of water stress. To test this hypothesis, a greenhouse experiment was conducted, with wheat and three varieties of faba bean grown as sole crops, single-variety faba bean–wheat intercrops, and multiple-variety faba bean–wheat intercrops. While extrapolations from the greenhouse to the field can only be made with caution, the study provides initial data of varietal influence on intercrop performance, which could justify more intensive field trials. Addressing the question of how intraspecific and interspecific diversity affects SNF in faba bean under drought will offer new insights for improved faba bean–wheat intercropping in regions where water limitations are common.
Materials and Methods
Plant Materials and Growth Conditions
The experiments were conducted in a greenhouse at the Swedish University of Agricultural Sciences (SLU), Alnarp, Sweden during spring and summer 2014 under natural illumination with day/night temperatures of 28/20°C, 16 h photoperiod, and 60% relative humidity. Seeds of three faba bean (V. faba) varieties, namely Alexia (A), Gloria (G), and Julia (J), and the spring wheat cultivar Dacke (T. aestivum) were used in this study. The faba bean varieties are registered by Saatzucht Gleisdorf (www.saatzuchtgleisdorf.at) and are among six official market Austrian spring faba bean varieties in Austria, Sweden, Germany, and Baltic countries. These faba bean varieties were selected to be consistent with previous research (Dhamala, 2012), and there was no a priori reason to believe that either faba bean varieties differed in terms of performance under drought and/or intercropped conditions. Both faba bean and wheat seeds were germinated in a sterilized peat germination medium, and the seedlings were transplanted into free drained pots that were filled with 2.5 kg of mixed air-dried soil (60% volume) and sand (40% volume). The clay sandy soil (pH: 7.9, electrical conductivity: 0.08 dS m−1; total N: 0.21%; total C: 2.66%; Olsen-P: 2.9 mg kg−1) used in this study was collected from the top 20 cm soil layer in an agricultural field at SLU Alnarp, Sweden (55°13′N, 13°4′E) and sieved (2 mm) before being used for growing plants in the greenhouse. For 15 days after sowing, the potted soil was kept moistened to 80% water-holding capacity (WHC).
The experiment was arranged as an 11-by-2 full factorial design, with 11 crop treatments and two water treatments. The crop treatments had three faba bean varieties and wheat grown as follows:
• Four pure stands: four sole crop treatments; A, G, J, and W.
• Seven intercropping combinations: varietal mixtures in intercropping with wheat (W) in one- (WA, WG, and WJ), two- (WAG, WAJ, and WGJ), and three-faba bean (WAGJ) mixtures.
• Water treatments correspond to two soil moisture levels: 80% WHC (sufficient water) and 30% WHC (water limitation). The water volume necessary to maintain 80 and 30% WHC was determined by weighing the pots twice per week during watering. Soil moisture treatments were imposed from day 15 to 49 post-transplantation.
The experiment was performed in a replacement design in which plant density remained constant (four seedlings per pot) across sole cropping and intercropping treatments, a common approach to assessing legume–cereal intercropping interactions (Hauggaard-Nielsen et al., 2008). In sole crop treatments, faba bean and wheat cultivars were grown as sole crops with four seedlings per pot per water regime. Intercropping treatments also contained four seedlings per pot per water regime but with two different species proportions depending on faba bean varietal composition. Single faba bean varieties and two faba bean varietal mixtures contained two faba bean seedlings (either the same variety or one variety each) and two wheat seedlings per pot, while the three faba bean varietal mixture contained three faba bean seedlings (one of each variety) and one wheat seedling per pot.
Measurement of Plant Growth, Nodulation, and N Concentration in Shoots and Nodules
Plants were harvested at 49 days after transplantation (flowering stage for faba bean and terminal spikelet initiation for wheat) and separated into shoots, roots, and nodules. Roots were obtained by carefully separating nodulated roots from the loosely adhering soil. The rhizosphere soil was removed from the nodulated roots that were washed through a sieve, and the nodules were detached and counted. The dry weight (DW) of shoots (SDW), nodules (NDW), and roots (RDW) were determined before they were ground to a fine powder to determine P and N contents. The dry weight of SDW and NDW were also used to determine the efficiency of use in rhizobial symbiosis (EURS). This parameter was estimated as the slope of the regression model (y = ax+b) of plant dry biomass (y) as a function of nodule dry biomass (x), where “b” corresponds to the plant biomass production without nodules (g SDW0) and “a” corresponds to the EURS as (g SDW – g SDW0) g−1 NDW (Zaman-Allah et al., 2007, Bargaz et al., 2012). The N content per plant of finely ground dry samples of faba bean nodules and shoots (faba bean and wheat) was determined using a CHN Element Analyzer (Thermo Scientific Fisons - EA-1108, Waltham, MA, United States).
Leaf Relative Water Content (RWC) and Drought Sensitivity Index
Relative water content in leaves was measured according to the method proposed by Krouma (2010). Fresh samples were weighed (FW) and then incubated by immersion in distilled water overnight at ~20°C and turgid weight (TW) was determined. After incubation, the samples were blotted dry and weighed prior to oven drying at 80°C for 48 h for dry weight determination. The RWC was calculated as follows:
Drought intensity index (DII) was calculated based on plant growth using DW as an indicator (Krouma, 2010) of water-limited (DWs) vs. well-watered (DWc) plants as follows:
Statistical Analysis
The experiment was performed in a completely randomized design with six replicates corresponding to six pots for each crop × water treatment. Two-way ANOVA and comparison of means ± SE were performed using Fisher's least significant difference test at 5% probability. Relationships between nodule and shoot DW were tested by regression analysis of which the significance of the correlation was determined.
Results
Wheat and Faba Bean Biomass Accumulation in Shoots and Roots
Both cropping and water treatments influenced biomass dry matter accumulation in wheat and faba bean shoots (Figures 1A, 2A). In both well-watered and water stress treatments, intercropping wheat with faba bean typically increased wheat shoot DW per plant over that achieved in wheat sole crop (Figure 1A), while water stress significantly decreased wheat and faba bean shoot DW per plant irrespective of the cropping treatment. Wheat shoot DW response to water and intercropping treatments was modulated by the faba bean variety used (Figure 1A). In both well-watered and water-stress conditions, wheat intercropped with the faba bean Alexia variety, and to a lesser extent with Julia, had greater shoot DW than that achieved in the wheat sole crop, but wheat shoot DW was much less affected when intercropped with Alexia (Figure 1A). Under water stress, the decreases observed in wheat shoot DW were reduced when intercropped with Gloria in the two-variety intercrops containing Alexia-Gloria and Gloria-Julia (WAG and WGJ) compared to Alexia-Julia (WAJ) (Figure 1A, Supplementary Figure 1). When water was not limiting, the lowest faba bean shoot DW occurred in the three-variety mixture Alexia-Gloria-Julia (WAGJ) (Figure 2A). Wheat intercropping also stimulated faba bean shoot DW compared to faba bean sole crop, for all three varieties tested. But as opposed to wheat, faba bean shoot DW was not affected by intraspecific diversity under water stress, and the various mixtures had no significant effect on faba bean shoot DW (Figure 2A).
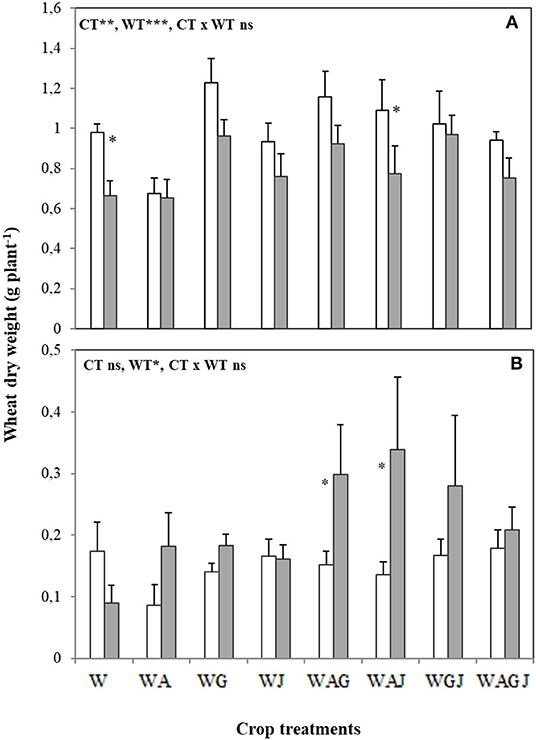
Figure 1. Dry weight of shoot (A) and root (B) of spring wheat (Triticum aestivum L. cv Dack) grown as a sole crop or intercropped with the three faba bean varieties Alexia (A), Gloria (G), and Julia (J) under adequate watering (80% field capacity; open bars) vs. water-stressed (30% field capacity, filled bars) conditions. Data are means and SE of six replicates harvested at 49 days post-transplantation. *Significant difference at p < 0.05.
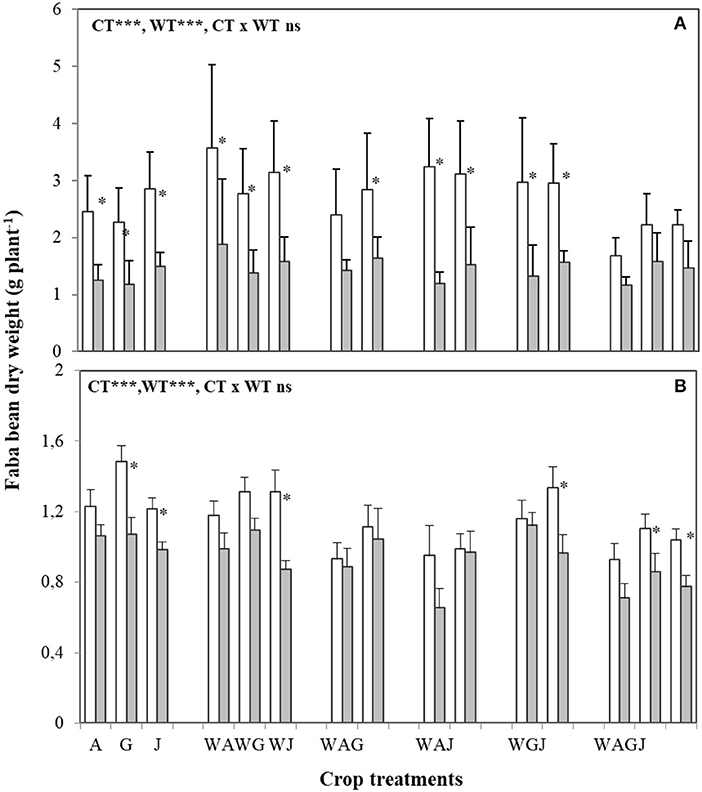
Figure 2. Dry weight of shoot (A) and root (B) of the three faba bean varieties A, G, and J grown as sole crops (A, G, and J) or intercropped with wheat (W) under adequate watering (80% field capacity; open bars) vs. water-stressed (30% field capacity, filled bars) conditions. Data are means and SE of six replicates harvested at 49 days post-transplantation. An asterisk * denotes a significant difference at p < 0.05.
Wheat and faba bean root DW were also affected by the water treatments (Figures 1B, 2B). The effect of faba bean intercropping on wheat root DW was most notable under water stress. Wheat root DW generally increased under water stress, particularly when intercropped with WAG and WAJ, where root DW increased by 50% compared to the well-watered treatment (Figure 1B, Supplementary Figure 1). Conversely, faba bean root DW tended to decrease under water stress (Figure 2B). However, this effect was dependent on faba bean variety, as root DW in Alexia was much more resistant to changes invoked by the water stress treatment (Figure 2B, Supplementary Figure 2).
Faba Bean Nodulation and the Relationship Between Nodulation and Shoot Dry Weight (SDW)
Nodule weight and number per plant were affected by both water and cropping treatments, with no interaction between water and cropping treatment effects observed. Water stress consistently reduced nodule weight and number, while intercropping faba bean with wheat consistently increased nodule DW and number irrespective of the water treatment (Figure 3). Yet, the effect of intercropping and water stress on nodule parameters was dependent on faba bean variety and faba bean intraspecific diversity. Evaluated as individual faba bean varieties, Alexia was most resistant to changes in nodule number and DW caused by water stress, and faba bean–wheat intercrops which included Alexia were also more resistant to the effects of water stress on nodule parameters. Nodule number per plant was not affected by water stress in the intercrops containing Alexia, Alexia-Gloria, and Alexia-Gloria-Julia (WA, WAG, and WAGJ), while the decrease in nodule DW was reduced in the intercrops containing Alexia, Alexia-Julia, and Alexia-Gloria (WA, WAJ, and WAG) (Figure 3). Intraspecific diversity did not always confer benefits to nodule parameters, as the worse-performing intercrop in terms of nodule DW was a three-variety faba bean mixture (WAGJ) under both well-watered and water-stress conditions (Figure 3).
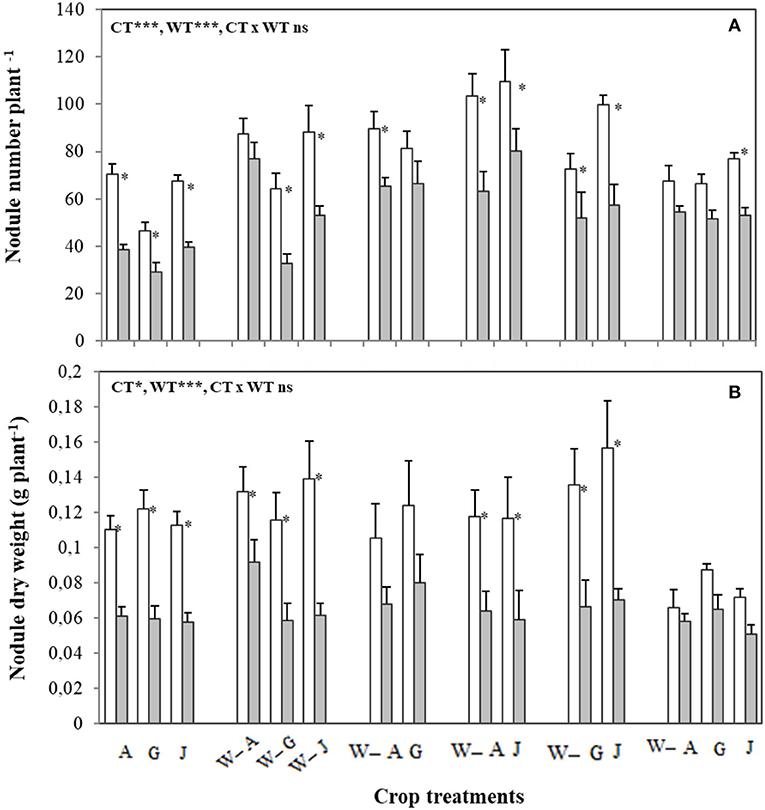
Figure 3. Number (A) and dry weight (B) of nodules of the three faba bean varieties, A, G, and J, grown as sole crops (A, G, and J) or intercropped with W under adequate watering (80% field capacity; open bars) vs. water-stressed (30% field capacity, filled bars) conditions. Data are means and SE of six replicates harvested at 49 days post-transplantation. An asterisk * denotes a significant difference at p < 0.05.
Under well-watered conditions, a significant relationship between shoot DW and both number and DW of nodules was observed for all faba bean varieties grown as a sole crop or in combination with wheat, although this relationship weakened under water-stress conditions, particularly for Julia (Supplementary Table 1). The relationship between nodulation and shoot DW was stronger under water stress for Alexia and Gloria (WAG) (when grown as a sole crop or as an intercrop with wheat) and in the two-variety intercrop that included both Alexia and Gloria (WAG) (Supplementary Table 1). The slope of the relationship between nodule DW and shoot DW, which measures the EURS, was positive and significant for all water and cropping treatment combinations, indicating that aboveground growth was always correlated to nodulation (Figure 4). EURS appeared stronger and significant (p < 0.001) when faba bean was intercropped with wheat, as indicated by the greater slope and R2 values (Figure 4B). Within crop treatments, water stress did not strongly affect EURS, except for the intercrops with two faba bean varieties (Figure 4C), where water stress did reduce the relationship between nodule DW and shoot DW. Conversely, in the intercrop with three faba bean varieties (WAGJ) (Figure 4D), water stress increased (p < 0.01) EURS (Figure 4D), as opposed to well-watered conditions, where nodule DW was weakly related (p < 0.05) to shoot DW (Table 1), leading to a lower EURS (Figure 4D).
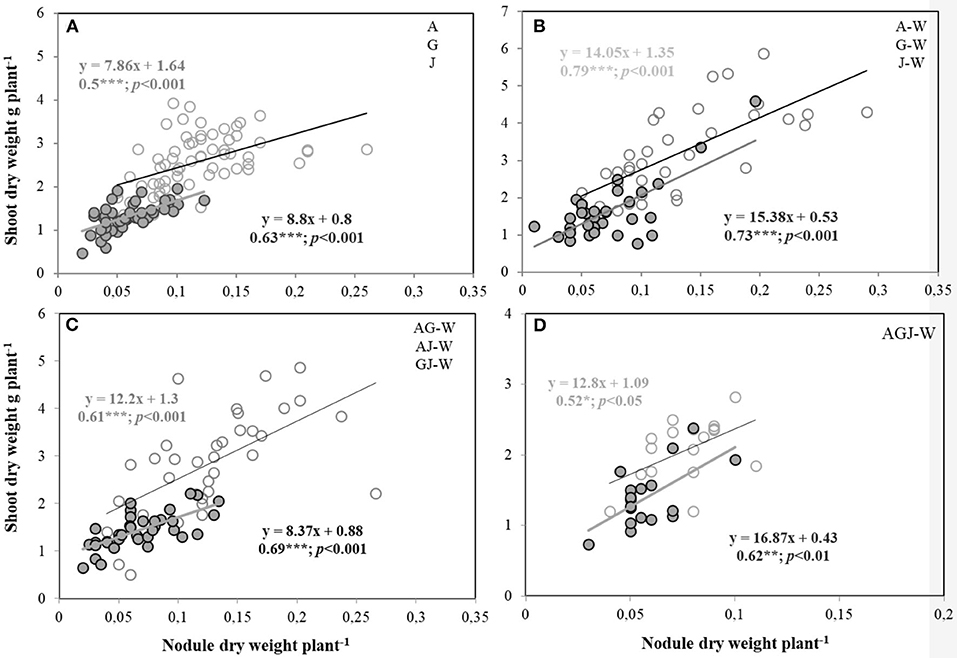
Figure 4. Efficiency in the use of the rhizobial symbiosis (EURS) for growth of the three faba bean varieties, A, G, and J and W, grown as sole crops [A, G, and J in (A)] or intercropped with W in three different levels: wheat intercropped with a single faba bean variety [(B) WA, WG, and WJ], wheat intercropped with two different faba bean varieties [(C) WAG, WAJ, and WGJ], and wheat intercropped with three different faba bean varieties [WAGJ in (D)]. Plants were grown under water sufficiency (80% field capacity; open circles) vs. water limitation (30% field capacity; filled circles). Asterisks *, **, and *** denote significant difference at p < 0.05, p < 0.01, and p < 0.001.
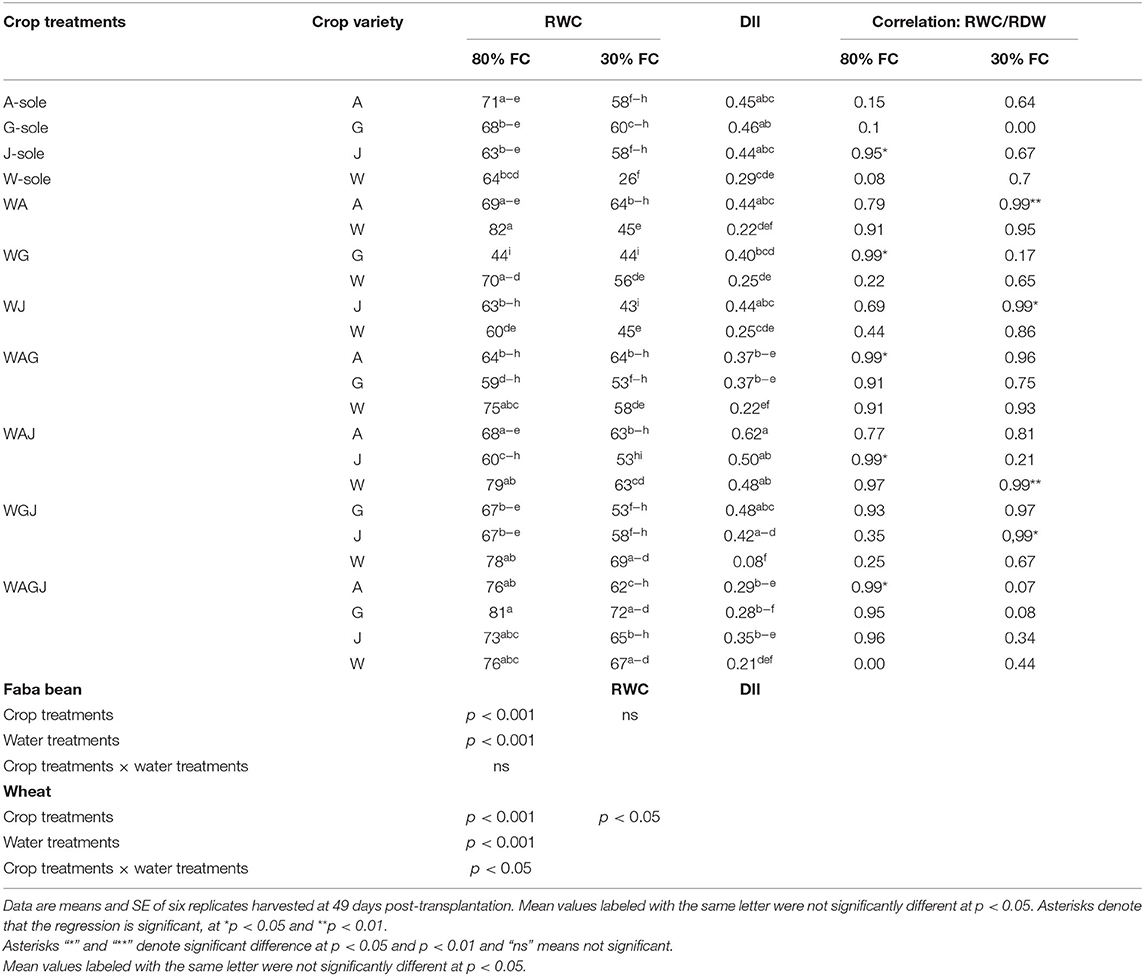
Table 1. Leaf relative water content (RWC), drought intensity index (DII), and correlation coefficient (R2 as assessed by correlation analysis) between RWC and root dry weight (RDW) parameters of three faba bean varieties, Alexia (A), Gloria (G), and Julia (J), grown as sole crops (A-, G-, and J-sole) or under intercropping with wheat (W) (WA, WG, WJ, WAG, WAJ, WGJ, and WAGJ) under water sufficiency (80% field capacity) and water limitation (30% field capacity).
Nitrogen Accumulation in Wheat and Faba Bean
Wheat N accumulation decreased under water stress when planted as a sole crop, but the negative effect of water stress on wheat N accumulation was generally averted, or even ameliorated, when intercropped with faba bean (Figure 5). The magnitude and direction of the effect of faba bean on wheat N content per plant under water stress were strongly dependent on faba bean variety. Irrespective of water treatment, wheat N accumulation was greatest when intercropped with the variety Gloria, indicating that faba bean variety influences the synergistic effects of faba bean–wheat intercrops, particularly under water stress (Figure 5). Wheat shoot N content increased by 20% under water stress in the intercropping treatment containing both Gloria and Julia (WGJ) but decreased 38% in wheat intercropped with both Alexia and Julia (WAJ). As opposed to wheat, faba bean N accumulation was consistently reduced by water stress, with the shoot and nodule N accumulation showing similar trends in variation across water and cropping treatments (Figures 6A,B). Shoot N accumulation varied across the faba bean varieties and intercropping treatments under water stress, from 28 mg N plant−1 in Gloria planted as a sole crop to 50 mg N plant−1 in Julia planted in the two-variety (WGJ) intercrop (Figure 6A). Under well-watered conditions, intercropping with wheat increased faba bean nodule and shoot N accumulation, while the effect of intraspecific faba bean diversity on N accumulation was not consistent. The two-variety intercrops comprising the variety Julia (wheat-Alexia-Julia “WAJ” and wheat-Gloria-Julia “WGJ”) had greater nodule and shoot N content per plant than wheat-Alexia-Gloria (WAG), while N accumulation was consistently reduced in the three-variety faba bean intercrop. Interestingly, when grown in a single faba bean variety intercrop with wheat, the variety Alexia accumulated the greatest amount of shoot N (90.08 mg N plant−1).
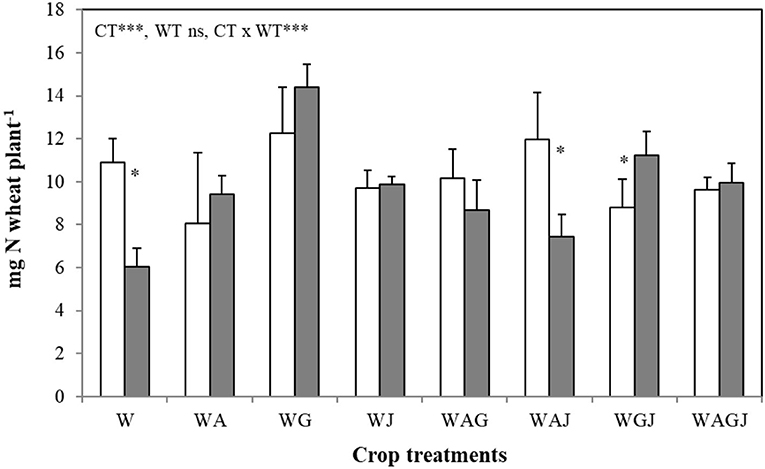
Figure 5. Nitrogen content in shoots of spring wheat (Triticum aestivum L. cv Dack) grown as a sole crop or intercropped with faba bean under adequate watering (80% field capacity; open bars) vs. water-stressed (30% field capacity, filled bars) conditions. Data are means and SE of three replicates harvested at 49 days post-transplantation. An asterisk * denotes a significant difference at p < 0.05.
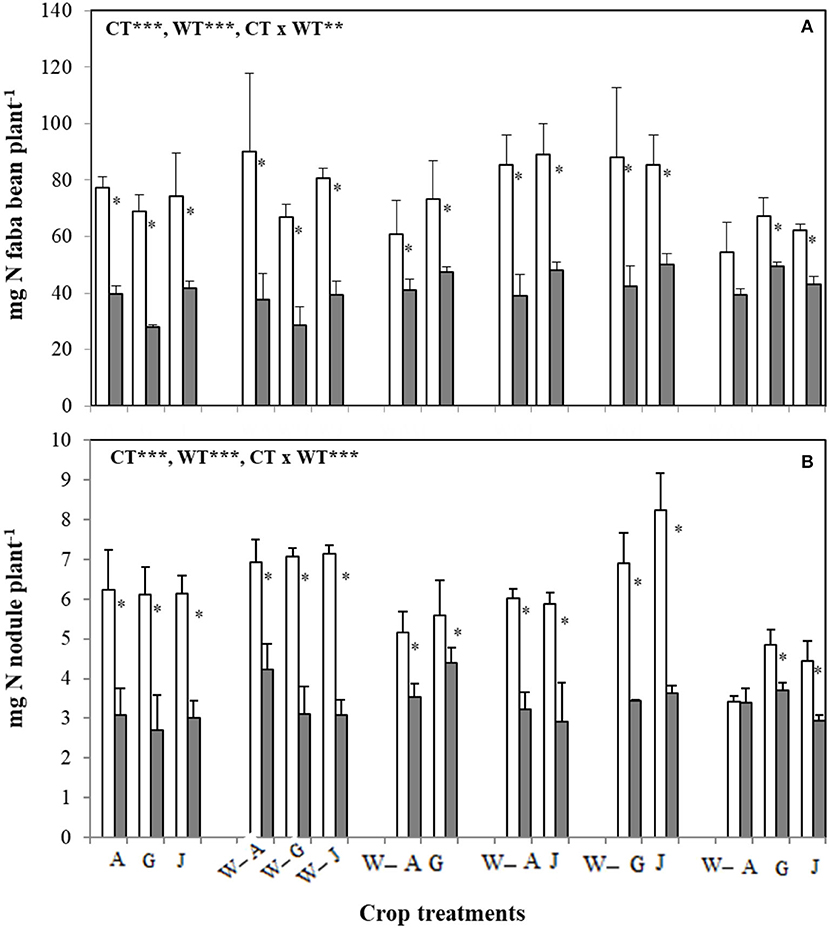
Figure 6. Nitrogen content in shoots (A) and nodules (B) of the three faba bean varieties, A, G, and J, grown as sole crops (A, G, and J) or intercropped with W under adequate watering (80% field capacity; open bars) vs. water-stressed (30% field capacity, filled bars) conditions. Data are means and SE of three replicates harvested at 49 days post-transplantation. An asterisk * denotes a significant difference at p < 0.05.
Leaf RWC and DII
For both faba bean and wheat, leaf RWC appeared to be significantly affected by crop and water treatments, and the interaction of these factors was significant in wheat only (Table 1). While leaf RWC decreased under water limitation in all crop species, the decrease was mostly significant in wheat either as sole crops or as intercrops with WG, WA, WG, WAG, and WAJ. In contrast, leaf RWC in faba bean did not substantially vary in response to water limitation, except for the varieties Julia, Gloria, and Alexia in the crop combinations wheat-Julia (WJ), WGJ, and the three-faba bean varietal mixtures in intercropping with wheat (WAGJ), respectively.
The DII significantly varies in wheat (p < 0.05) comparatively to faba bean varieties with only Alexia having the highest DII within the crop combination WAJ. Correlations between leaf RWC and RDW, as indicative of root importance in water use, were significant in several faba bean crop treatments compared to those in wheat, as the only statistically significant correlation (R2 = 0.99**) was noted in WAJ under water limitation. Under water limitation, the correlations between leaf RWC and RDW were less consistent in all sole cropped faba bean varieties; meanwhile, these parameters were positively and significantly correlated in the one-variety intercrops comprising Alexia and Julia (R2 = 0.99**) and the two-variety intercrops comprising Julia (WGJ) (Table 1).
Discussion
The present study demonstrates, in a controlled greenhouse environment, that benefits of varietal diversity are apparent in intercrop situations with and without water limitation, although the observed effects of intraspecific diversity on intercrop performance were somewhat inconsistent. We found that the faba bean variety influences several important measures of intercrop performance, supporting the results of studies conducted on oat–pea intercrops (Peltonen-Sainio et al., 2017), maize–common bean intercrops (Isaacs et al., 2016), and maize–cowpea intercrop (Abera et al., 2017). In the present study, variations in above- and below-ground parameters related to plant growth, nodulation, and nutrient acquisition demonstrated that faba bean–wheat intercrops buffered biomass dry matter accumulation, nodulation, and N accumulation in water-limited faba bean (in intercrops containing the varieties WAJ) and wheat (in intercrops containing the variety Gloria). This effect depends on the legume variety and the varietal diversity level, which likely modulated the intercrop effect. For instance, intercrops containing the variety Alexia improved nodule number, weight, and N accumulation in faba bean, while wheat biomass and N accumulation were greatest in intercrops containing the variety Gloria in both limited- and sufficient-water treatments. These results are consistent with the findings of Dhamala (2012), who studied the effect of diversity in faba bean–wheat intercrops in the field. In an organically managed cropping system, significantly higher N utilization efficiency was found in faba bean–wheat intercrops containing the varieties Alexia and Gloria. They found, on a per-plant basis, that intercropped varietal mixtures increased biomass yield and N2 fixation (Dhamala, 2012), which is consistent with the finding of this study that intercrops with two faba bean varieties tended to have positive or neutral effects on measured wheat and faba bean variables. However, caution is needed to disaggregate the effects of intraspecific from interspecific competition exerted by different faba bean varieties intercropped with wheat at different seeding ratios under limited vs. sufficient water conditions.
On the other hand, the effect of intraspecific faba bean diversity was also inconsistent and generally reduced when all three faba bean varieties were planted together and intercropped with wheat. A negative effect was observed, plausibly due to the exposure of plants to a combination of water limitation and gradually decreasing soil nutrient availability, as no nutrients were added during the 49-day experiment. Despite the unfavorable growth conditions, intercropping of wheat with faba bean (especially with WG and WAG) typically increased wheat shoot biomass in both well-watered and water-stress treatments and so more increased for wheat root biomass in intercropping with Gloria-Julia and Alexia-Julia. This may provide evidence for a shift or enhanced niche complementarity that can result from enhanced genetic diversity (Brooker et al., 2016) that improved water-limited wheat performance, although a rigorous test of this hypothesis would require additional measurements of root stratification pattern along with nodule spatial variation (Bargaz et al., 2016). This agrees with several studies that belowground niche complementarity in legume–cereal intercrops may optimize the rhizosphere biological functionalities with positive consequences on resource use and adaptation to environmental constraints (De Vallavieille-Pope, 2004; Ghosh, 2004; Li et al., 2007; Bedoussac and Justes, 2011; Hinsinger et al., 2011). However, the agroecological advantages of legume–cereal intercropping, which combines interspecific or intraspecific species diversity, are still poorly understood, notably under water limitation.
Increased root biomass allocation under water limitation for both wheat and faba bean in intercropping (notably with Alexia-Gloria, Alexia-Julia, and Gloria-Julia, Supplementary Figures 1, 2) indicates increased intercropping performance, which may presumably be due to wheat intercrop whose root allocation was also higher and positively correlated with leaf water content under water limitation (Table 1). This finding suggests greater water uptake and/or water availability, possibly due to improvements in soil hydraulic conductivity caused by changes in rooting patterns (Prieto et al., 2012; Pang et al., 2013). Improved hydraulic conductivity and hydraulic lift, although not measured in this study, is likely a mechanism contributing to enhanced root biomass, root depth, avoidance of nodule dehydration (surviving longer in the upper soil layers), and improved overall performance of intercropped faba bean under water limitation conditions (Bargaz et al., 2016). Coherently, increased RWC was associated with a lower drought severity index, which may have resulted from positive facilitative interactions, such as in several faba bean–wheat intercrops (containing Alexia, Gloria, Julia, Alexia-Julia, and Gloria-Julia) (Table 1) that expressed positive correlations in terms of RWC and biomass of roots whose morphological traits are paramount in optimizing water and nutrient acquisition (Ho et al., 2005), which presumably is due to niche differentiation and complementarity reducing negative competitive interactions and niche overlap (Brooker et al., 2016).
The significant correlations between biomasses of both nodules and shoots (Figure 4, Supplementary Table 1) indicated close dependency of below- and above-ground biomass and that the EURS is likely to increase (based on the slope of the regression) in all intercropped faba bean varietal mixtures compared to faba bean sole cropping under both water treatments. Notably, intercropped faba bean containing one variety or all three varieties expressed positive and significant correlations between belowground and aboveground biomass regardless of water treatments. Such a positive effect of both intercropping and intraspecific diversity on faba bean nodulation and plant growth is, thus, likely a consequence of increased competition for soil N that led to the optimization of EURS rather than a direct stimulation of nodulation and N2 fixation. This stimulation has largely been reported for legume N acquisition under intercropping (Li et al., 2001, 2009; Knudsen et al., 2004; Banik et al., 2006; Vasileva et al., 2011, Bargaz et al., 2016), meanwhile little is known as to how EURS varies in response to increased intraspecific faba bean diversity and water limitation. In addition to improved EURS, increasing faba bean intraspecific diversity (notably in intercrops containing Alexia-Gloria, Alexia-Julia, and Gloria-Julia) was also beneficial for wheat growth, whose root biomass significantly increased under water limitation. Moreover, faba bean mixture containing the variety Gloria had a slight advantage on wheat N concentration under both water treatments. Conversely, a field study by Dhamala (2012) found that the growth and the development of wheat were highly suppressed in the faba bean mixture (containing Gloria), while both crop harvest index and N balance were highest in faba bean mixture varieties intercropped with wheat.
In a recent meta-analysis, Reiss and Drinkwater (2018) found that planting multiple varieties of the same crop species (i.e., increased intraspecific diversity) resulted in modest yield improvements over sole crops, but the magnitude of this intraspecific diversity effect was larger when biotic or abiotic stresses were present. The authors concluded that varietal mixtures were more advantageous under stress because of greater functional trait diversity, similar to the findings of another literature review (Adu-Gyamfi et al., 2015). Such findings are in line with this study, indicating that faba bean variety selection will influence faba bean–wheat intercrop productivity under water-limited conditions. The initial evidence of this study elucidating, for the first time, a certain level of drought tolerance under increased faba bean varietal diversity and wheat intercropping requires more investigations; mainly field trials are warranted to investigate whether the effects found in this greenhouse study persists in more realistic conditions.
Conclusions
When intercropped with wheat, faba bean varietal diversity had significant effects on several plant traits important to intercrop performance in both well-watered and water-limited conditions. However, the effect of intraspecific diversity itself was inconsistent across the different plant traits, and intercropping tended to have positive or neutral effects on the measured wheat and faba bean variables, notably with two-varietal faba bean mixtures. But intercrop performance was generally reduced when all the three faba bean varieties were planted together, which suggests that faba bean varietal mixture level has a significant impact on the performance of faba bean–wheat intercrops and may be a critical management decision for farmers planting intercrops, particularly in drought-prone areas. The hypothesis that increased faba bean species diversity can buffer faba bean–wheat intercrop performance against water limitation via positive rhizosphere interactions was not conclusively rejected. Likely, multiple experimental variables, such as plant density, intercrop species, the type of stress, and its intensity and duration, will influence the relationship between intraspecific diversity in intercrops and performance under drought stress.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by the Swedish Research Council Formas within the framework of the project N° 229-2012-814 granted to AB (mobility starting grant for young researchers) and the Stiftelsen Oscar och Lili Lamms Minne granted to GC.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2021.655973/full#supplementary-material
References
Abdelhamid, M. T., Palta, J. A., Veneklaas, E. J., Atkins, C., Turner, N. C., and Siddique, K. H. (2011). Drying the surface soil reduces the nitrogen content of faba bean (Vicia faba L.) through a reduction in nitrogen fixation. Plant Soil 339, 351–362. doi: 10.1007/s11104-010-0586-9
Abera, R., Worku, W., and Beyene, S. (2017). Performance variation among improved common bean (Phaseolus vulgaris L.) genotypes under sole and intercropping with maize (Zea mays L.). Afr. J. Agric. Res. 2, 397–405. doi: 10.5897/AJAR2016.11794
Adu-Gyamfi, P., Mahmood, T., and Trethowan, R. (2015). Can wheat varietal mixtures buffer the impacts of water deficit? Crop Past. Sci. 66, 757–769. doi: 10.1071/CP14177
Agegnehu, G., Ghizaw, A., and Sinebo, W. (2008). Yield potential and land-use efficiency of wheat and faba bean mixed intercropping. Agr. Sustain. Dev. 28, 257–263. doi: 10.1051/agro:2008012
Alghamdi, S. S., Al-Shameri, A. M., Migdadi, H. M., Ammar, M. H., El-Harty, E. H., Khan, M. A., et al. (2015). Physiological and molecular characterization of faba bean (Vicia faba L.) genotypes for adaptation to drought stress. J. Agr. Crop Sci. 201, 401–409. doi: 10.1111/jac.12110
Amede, T., Schubert, S., and Stahr, K. (2003). Mechanisms of drought resistance in seed legumes. I. – Osmotic adjustment. Ethiop. J. Sci. 26, 37–46. doi: 10.4314/sinet.v26i1.18198
Androsoff, G. L., Van Kessel, C., and Pennock, D. J. (1995). Landscape-scale estimates of dinitrogen fixation by Pisum sativum by nitrogen-15 natural abundance and enriched isotope dilution. Biol. Fertil. Soils 20, 33–40. doi: 10.1007/BF00307838
Banik, P. A., Midya, B. K., Sarkar, S., and Ghose, S. (2006). Wheat and chickpea intercropping systems in an additive series experiment: advantages and weed smothering. Eur. J. Agron. 24, 325–332. doi: 10.1016/j.eja.2005.10.010
Bargaz, A., Ghoulam, C., Amenc, L., Lazali, M., Faghire, M., and Drevon, J. J. (2012). Phosphoenol pyruvate phosphatase is induced in the root nodule cortex of Phaseolus vulgaris under phosphorus deficiency. J. Exp. Bot. 63, 4723–4730. doi: 10.1093/jxb/ers151
Bargaz, A., Isaac, M. E., Jensen, E. S., and Carlsson, G. (2016). Nodulation and root growth increase in lower soil layers of water-limited faba bean intercropped with wheat. J. Plant Nutr. Soil Sci. 179, 537–546. doi: 10.1002/jpln.201500533
Bargaz, A., Noyce, G. L., Genevieve, F. R., Carlsson, C., Furze, J. R., Jensen, E. J., et al. (2017). Species interactions enhance root allocation, microbial diversity and P acquisition in intercropped wheat and soybean under P deficiency. Appl. Soil Ecol. 120, 179–188. doi: 10.1016/j.apsoil.2017.08.011
Bedoussac, L., Journet, E., Hauggaard-Nielsen, H., Naudin, C., Corre-Hellou, G., Jensen, E. S., et al. (2015). Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron. Sustain. Dev. 35, 911–935. doi: 10.1007/s13593-014-0277-7
Bedoussac, L., and Justes, E. (2011). A comparison of commonly used indices for evaluating species interactions and intercrop efficiency: application to durum wheat–winter pea intercrops. Field Crop Res. 124, 25–36. doi: 10.1016/j.fcr.2011.05.025
Brooker, R. W., Karley, A. J., Newton, A. C., Pakeman, R. J., and Schöb, C. (2016). Facilitation and sustainable agriculture: a mechanistic approach to reconciling crop production and conservation. Func. Ecol. 30, 98–107. doi: 10.1111/1365-2435.12496
Bukovsky-Reyes, S., Isaac, M. E., and Blesh, B. (2019). Effects of intercropping and soil properties on root functional traits of cover crops. Agric. Ecosyst. Environ. 285:106614. doi: 10.1016/j.agee.2019.106614
Bulson, H. A. J., Snaydon, R. W., and Stopes, C. E. (1997). Effects of plant density on intercropped wheat and field beans in an organic farming system. J. Agric. Sci. 128, 59–71. doi: 10.1017/S0021859696003759
Bybee-Finley, K., and Ryan, M. R. (2018). Advancing intercropping research and practices in industrialized agricultural landscapes. Agriculture 8:80. doi: 10.3390/agriculture8060080
Carlsson, G., and Huss-Danell, K. (2003). Nitrogen fixation in perennial forage legumes in the field. Plant Soil 253, 353–372. doi: 10.1023/A:1024847017371
De Vallavieille-Pope, C. (2004). Management of disease resistance diversity of cultivars of a species in single fields: controlling epidemics. C. R. Biol. 327, 611–620. doi: 10.1016/j.crvi.2003.11.014
Dhamala, N. R. (2012). Effect of varietal and species diversity on nitrogen fixation and nitrogen cycling in an organic faba bean spring wheat intercropping system. Second cycle, A2E. Alnarp: SLU, Department of Work Science, Business Economics, and Environmental Psychology. Available online at: https://stud.epsilon.slu.se/4369/
Farooq, M., Gogoi, N., Barthakur, S., Baroowa, B., Bharadwaj, N., Alghamdi, S. S., et al. (2017). Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 203, 81–102. doi: 10.1111/jac.12169
Ghosh, P. K. (2004). Growth, yield, competition and economics of groundnut/cereal fodder intercropping systems in the semi-arid tropics of India. Field Crops Res. 88, 227–237. doi: 10.1016/j.fcr.2004.01.015
Hauggaard-Nielsen, H., Gooding, M., Ambus, P., Corre-Hellou, G., Crozat, Y., Dahlmann, C., et al. (2009). Pea–barley intercropping for efficient symbiotic N2-fixation, soil N acquisition and use of other nutrients in European organic cropping systems. Field Crops Res. 113, 64–71. doi: 10.1016/j.fcr.2009.04.009
Hauggaard-Nielsen, H., Holdensen, L., Wulfsohn, D., and Jensen, E. S. (2010). Spatial variation of N2-fixation in field pea (Pisum sativum L.) at the field scale determined by the 15N natural abundance method. Plant Soil 327, 167–184. doi: 10.1007/s11104-009-0043-9
Hauggaard-Nielsen, H., Jørnsgaard, B., Kinane, J., and Jensen, E. S. (2008). Grain legume–cereal intercropping: the practical application of diversity, competition and facilitation in arable and organic cropping systems. Renew. Agric. Food Syst. 23, 3–12. doi: 10.1017/S1742170507002025
Hinsinger, P., Betencourt, E., Bernard, L., Brauman, A., Plassard, C., Shen, J., et al. (2011). P for two, sharing a scarce resource: soil phosphorus acquisition in the rhizosphere of intercropped species. Plant Physiol. 156, 1078–1086. doi: 10.1104/pp.111.175331
Ho, D. M., Juan, C. R., Brown, K. M., and Lynch, J. P. (2005). Root architectural tradeoffs for water and phosphorus acquisition. Func. Plant Biol. 32, 737–748. doi: 10.1071/FP05043
Hungria, M., and Vargas, M. A. (2000). Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res. 65, 151–164. doi: 10.1016/S0378-4290(99)00084-2
Isaac, M. E., Hinsinger, P., and Harmand, J. M. (2012). Nitrogen and phosphorus economy of a legume tree-cereal intercropping system under controlled conditions. Sci. Total Environ. 434, 71–78. doi: 10.1016/j.scitotenv.2011.12.071
Isaacs, K. B., Snapp, S. S., Kelly, J. D., and Chung, K. R. (2016). Farmer knowledge identifies a competitive bean ideotype for maize–bean intercrop systems in Rwanda. Agric. Food Secur. 5, 1–18. doi: 10.1186/s40066-016-0062-8
Jensen, E. S., Carlsson, G., and Hauggaard-Nielsen, H. (2020). Intercropping of grain legumes and cereals improves the use of soil N resources and reduces the requirement for synthetic fertilizer N: a global-scale analysis. Agron. Sustain. Dev. 40, 1–9. doi: 10.1007/s13593-020-0607-x
Jensen, E. S., Peoples, M. B., and Hauggaard-Nielsen, H. (2010). Faba bean in cropping systems. Field Crops Res. 115, 203–216. doi: 10.1016/j.fcr.2009.10.008
Karkanis, A., Ntatsi, G. N., Lepse, L., Fernández, J. A., Vågen, I. M., Rewald, B., et al. (2018). Faba bean cultivation—revealing novel managing practices for more sustainable and competitive European cropping systems. Front. Plant Sci. 9, 1–14. doi: 10.3389/fpls.2018.01115
Khan, H. R., Paull, J. G., Siddique, K. H. M., and Stoddard, F. L. (2010). Faba bean breeding for drought-affected environments: a physiological and agronomic perspective. Field Crops Res. 115, 279–286. doi: 10.1016/j.fcr.2009.09.003
Knudsen, M. T., Hauggaard-Nielsen, H., and Jensen, E. S. (2004). Pea, faba bean and lupin intercropped with spring barley—performance and N2 fixation at two temperate locations. J. Agric. Sci. 142, 617–627. doi: 10.1017/S0021859604004745
Krouma, A. (2010). Plant water relations and photosynthetic activity in three Tunisian chickpea (Cicer arietinum L.) genotypes subjected to drought. Turk. J. Agric. Forestry 34, 257–264. doi: 10.3906/tar-0904-1
Li, L., Li, S. M., Sun, J. H., Zhou, L. L., Bao, X. G., Zhang, H. G., et al. (2007). Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci. U. S. A. 104, 11192–11196. doi: 10.1073/pnas.0704591104
Li, L., Sun, J., Zhang, F., Li, X., Yang, S., and Rengel, Z. (2001). Wheat/maize or wheat/soybean strip intercropping. I. Yield advantage and interspecific interactions on nutrients. Field Crop Res. 71, 123–137. doi: 10.1016/S0378-4290(01)00156-3
Li, Y.-Y., Yu, C.-B., Cheng, X., Li, C.-J., Sun, J.-H., Zhang, F.-S., et al. (2009). Intercropping alleviates the inhibitory effect of N fertilization on nodulation and symbiotic N2 fixation of faba bean. Plant Soil 323, 295–308. doi: 10.1007/s11104-009-9938-8
Lithourgidis, A. S., and Dordas, C. A. (2010). Forage yield, growth rate, and nitrogen uptake of faba bean intercrops with wheat, barley, and rye in three seeding ratios. Crop Sci. 50, 2148–2158. doi: 10.2135/cropsci2009.12.0735
Marino, D., Frendo, P., Ladrera, R., Zabalza, A., Puppo, A., Arrese-Igor, C., et al. (2007). Nitrogen fixation control under drought stress. Localized or systemic? Plant Physiol. 143, 1968–1974. doi: 10.1104/pp.106.097139
McDonald, G. K., and Paulsen, G. M. (1997). High temperature effects on photosynthesis and water relations of grain legumes. Plant Soil 196, 47–58. doi: 10.1023/A:1004249200050
Nasielski, J., Furze, J. R., Tan, J., Bargaz, A., Thevathasan, N. V., and Isaac, M. E. (2015). Agroforestry promotes soybean yield stability and N2-fixation under water stress. Agron. Sustain. Dev. 35, 1541–1549. doi: 10.1007/s13593-015-0330-1
Pang, J., Wang, Y., Lambers, H., Tibbett, M., Siddique, K. H. M., and Ryan, M. H. (2013). Commensalism in an agroecosystem: hydraulic redistribution by deep-rooted legumes improves survival of a droughted shallow-rooted legume companion. Physiol. Plant. 149, 79–90. doi: 10.1111/ppl.12020
Peltonen-Sainio, P., Kontturi, M., Laine, A., Niskanen, M., Hurme, T., and Jauhiainen, L. (2017). Characterizing an outperforming pea cultivar for intercropping with oat at high latitudes. Agric. Food Sci. 26, 138–147. doi: 10.23986/afsci.61076
Prieto, I., Armas, C., and Pugnaire, F. I. (2012). Water release through plant roots: new insights into its consequences at the plant and ecosystem level. New Phytol. 193, 830–841. doi: 10.1111/j.1469-8137.2011.04039.x
Raseduzzaman, M., and Jensen, E. S. (2017). Does intercropping enhance yield stability in arable crop production? A meta-analysis. Eur. J. Agron. 91, 25–33. doi: 10.1016/j.eja.2017.09.009
Reiss, E. R., and Drinkwater, L. E. (2018). Cultivar mixtures: a meta-analysis of the effect of intraspecific diversity on crop yield. Ecol. Appl. 28, 62–77. doi: 10.1002/eap.1629
Shen, J., Li, C., Mi, G., Li, L., Yuan, L., Jiang, R., et al. (2013). Maximizing root/rhizosphere efficiency to improve crop productivity and nutrient use efficiency in intensive agriculture of China. J. Exp. Bot. 64, 1181–1192. doi: 10.1093/jxb/ers342
Unkovich, M. J., and Pate, J. S. (2000). An appraisal of recent field measurements of symbiotic N2 fixation by annual legumes. Field Crops Res. 65, 211–228. doi: 10.1016/S0378-4290(99)00088-X
Vasileva, V., Kostov, O., Vasilev, E., and Athar, M. (2011). Effect of mineral nitrogen fertilization on growth characteristics of lucerne under induced water deficiency stress. Pakistan J. Bot. 43, 2925–2928. Available online at: http://www.pakbs.org/pjbot/PDFs/43(6)/46.pdf
Wang, G. Z., Li, H. G., Christie, P., Zhang, F. S., Zhang, J. L., and Bever, J. D. (2017). Plant-soil feedback contributes to intercropping overyielding by reducing the negative effect of take-all on wheat and compensating the growth of faba bean. Plant Soil 415, 1–12. doi: 10.1007/s11104-016-3139-z
Wezel, A., Casagrande, M., Celette, F., Vian, J.-F., Ferrer, A., Peigné, J., et al. (2014). Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 34, 1–20. doi: 10.1007/s13593-013-0180-7
Xiao, Y., Li, L., and Zhang, F. (2004). Effect of root contact on interspecific competition and N transfer between wheat and faba bean using direct and indirect 15 N techniques. Plant Soil 262, 45–54. doi: 10.1023/B:PLSO.0000037019.34719.0d
Zaman-Allah, M., Sifi, B., L'Taief, B., El-Aouni, M. H., and Drevon, J. J. (2007). Symbiotic response to low phosphorus supply in two common bean (Phaseolus vulgaris L.) genotypes. Symbiosis 44, 109–113. Available online at: https://dalspace.library.dal.ca/handle/10222/78298
Zhang, F. S., Cui, Z. L., Fan, M. S., Zhang, W. F., Chen, X. P., and Jiang, R. F. (2011). Integrated soil–crop system management: reducing environmental risk while increasing crop productivity and improving nutrient use efficiency in China. J. Environ. Qual. 40, 1051–1057. doi: 10.2134/jeq2010.0292
Keywords: cereal, drought, intercropping, legumes, nitrogen, nodulation, Vicia faba
Citation: Bargaz A, Nasielski J, Isaac ME, Jensen ES and Carlsson G (2021) Faba Bean Variety Mixture Can Modulate Faba Bean–Wheat Intercrop Performance Under Water Limitation. Front. Agron. 3:655973. doi: 10.3389/fagro.2021.655973
Received: 19 January 2021; Accepted: 12 May 2021;
Published: 08 June 2021.
Edited by:
Mohammad Bagher Hassanpouraghdam, University of Maragheh, IranReviewed by:
Michele Monti, Mediterranea University of Reggio Calabria, ItalyEric Justes, Centre de Coopération Internationale en Recherche Agronomique pour le Développement (CIRAD), France
Muhammad Ali Raza, Sichuan Agricultural University, China
Copyright © 2021 Bargaz, Nasielski, Isaac, Jensen and Carlsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Adnane Bargaz, YWRuYW5lLmJhcmdhekB1bTZwLm1h