 Adrian Matysek
Adrian Matysek Sumudu Perera Kimmantudawage2
Sumudu Perera Kimmantudawage2 Andrea B. Maier
Andrea B. Maier- 1Department of Human Genetics, University of Amsterdam, Amsterdam UMC, University Medical Centers, Amsterdam, Netherlands
- 2Department of Medicine and Aged Care, Royal Melbourne Hospital, Faculty of Medicine, Dentistry and Health Sciences, University of Melbourne, Melbourne, VIC, Australia
- 3Department of Psychological Medicine, Yong Loo Lin School of Medicine, National University of Singapore, Singapore, Singapore
- 4Healthy Longevity Translational Research Programme, Yong Loo Lin School of Medicine, National University of Singapore, Singapore, Singapore
- 5Centre for Healthy Longevity, National University Health System, Singapore, Singapore
- 6Department of Human Movement Sciences, Faculty of Behavioral and Movement Sciences, Amsterdam Movement Sciences, Vrije Universiteit, Amsterdam, Netherlands
Background: Dementia is a global challenge with 10 million individuals being diagnosed every year. Currently, there are no established disease-modifying treatments for dementia. Impaired nutrient sensing has been implicated in the pathogenesis of dementia. Compounds that inhibit the glycogen synthase kinase-3 (GSK3) pathway have been investigated as a possible treatment to attenuate the progression of the disease, particularly the suppression of the hyper-phosphorylation process of the tau protein.
Aims: Systematically summarizing compounds which have been tested to inhibit the GSK3 pathway to treat cognitive impairment and dementia.
Methods: PubMed, Embase and Web of Science databases were searched from inception until 28 July 2021 for articles published in English. Interventional animal studies inhibiting the GSK3 pathway in Alzheimer’s disease (AD), Parkinson’s dementia, Lewy body dementia, vascular dementia, mild cognitive impairment (MCI) and normal cognitive ageing investigating the change in cognition as the outcome were included. The Systematic Review Centre for Laboratory animal Experimentation’s risk of bias tool for animal studies was applied.
Results: Out of 4,154 articles, 29 described compounds inhibiting the GSK3 pathway. All studies were based on animal models of MCI, AD or normal cognitive ageing. Thirteen out of 21 natural compounds and five out of nine synthetic compounds tested in MCI and dementia animal models showed an overall positive effect on cognition. No articles reported human studies. The risk of bias was largely unclear.
Conclusion: Novel therapeutics involved in the modulation of the GSK3 nutrient sensing pathway have the potential to improve cognitive function. Overall, there is a clear lack of translation from animal models to humans.
1 Introduction
The number of people aged 60 years and older is increasing globally, with a similar trend being seen in those affected by neurodegenerative diseases. In 2020, globally over 50 million people were living with dementia, of which 60–70% with Alzheimer’s disease (AD). Dementia has multiple causes and is characterized by impaired cognitive function. The neurodegenerative process of dementia begins a long time before the presentation of clinical symptoms (Katsuno et al., 2018). Factors contributing to the neurodegenerative process include metabolic disorders such as hyperglycaemia (Gale et al., 2018). As such it has been hypothesized that targeting ageing pathways may be a viable therapeutic option for treating dementia. One such pathway is the nutrient sensing pathway such as glycogen synthase kinase-3 (GSK3) which has been linked to the onset of dementias (Efeyan et al., 2015).
Impaired nutrient sensing is defined as dysregulation in processing nutrients for mammalian cells (de Lucia et al., 2020). Hyperglycemia such as in diabetes can lead to inflammation and apoptosis associated with high GSK3 activity. Hyperlipidemia and obesity can also increase GSK3 activity (Liu and Yao, 2016). The regulation of GSK3, one of the main molecules involved in insulin signalling (Jolivalt et al., 2008), plays an important role in this process. GSK3 is a protein kinase and present in many processes in organisms such as cell signalling and cellular transport (Souder and Anderson, 2019). It has two isoforms: alpha and beta. Both take part in glycogen metabolism and the phosphorylation process of over hundred substrates (Beurel et al., 2015), which are important for memory formation (Takashima, 2012). Impaired GSK3, when phosphorylated, induces hyper-phosphorylation of the tau protein, and subsequently lead to the formation of neurofibrillary tangles (NFTs) (Sayas and Ávila, 2021). Impaired GSK3 also contributes to the formation of amyloid beta (Aβ) plaques through the amyloid precursor protein (APP) cleavage pathway (Giese, 2009). Higher levels of GSK3 are associated with neuronal loss in Huntington’s disease and AD (L’Episcopo et al., 2016; Gale et al., 2018). Therefore, inhibition of GSK3 by therapeutic compounds could be a promising approach in managing dementia (Jope and Roh, 2006; Bhat et al., 2018).
This systematic review summarizes novel compounds inhibiting the GSK3 pathway in the context of preventing and treating cognitive impairment and dementia.
2 Methods
2.1 Selection of Articles
The protocol of this systematic review was registered at PROSPERO International prospective register of systematic reviews (Reg #: CRD42018091645). PubMed, Web of Science and Embase databases were searched until the 28 July 2021. Key search terms included Vascular Dementia (VD); AD; Lewy Body Dementia (LBD); Parkinson’s Disease (PD); cognitive ageing; autophagy; lysosome; ubiquitin; proteasome endopeptidase complex; molecular chaperone; unfolded protein response; insulin; mTOR; GSK3; protein kinase B (PKB)/Ak strain transforming (Akt); phosphoinositide 3-kinase (PI3K); 5′ AMP-activated protein kinase (AMPK); sirtuin; sirolimus; everolimus; temsirolimus; rapamycin; metformin; dipeptidyl peptidase 4 (DPP-4); glucagon-like peptide-1 (GLP-1); nicotinamide; nicotinamide adenine dinucleotide (NAD); spermidine; matinib; imatinib; nilotinib; dasatinib; bosutinib; ponatinib; bafetinib; lithium; heat-shock protein; caloric restriction; carbohydrate restricted diet; protein restricted diet; cognition. Additional relevant articles were identified by screening the references of included articles. After removing duplicates, remaining articles were screened for inclusion using the Covidence systematic review software (Veritas Health Innovation, Melbourne, Australia).
2.1.1 Eligibility Criteria
Articles included in this review met the following inclusion criteria: 1) study population: animals or humans; normal cognitive ageing or dementia (AD, VD, LBD, PD). Populations likely to have a faster pace of cognitive ageing such as those with type 2 diabetes mellitus, insulin-resistance and obesity, were also included. For animals, normal ageing was defined as a strain not at a greater propensity to develop dementia and not manipulated to mimic dementia. Dementia models were defined as strains at a greater propensity to develop dementia compared to normal ageing strains. 2) Interventional study design with comparators, including randomised controlled trial, quasi-randomised controlled trials, and pre/post studies. 3) Intervention: compounds targeting cognition by inhibiting the GSK3 nutrient sensing pathway. 4) Outcome: cognitive function measured using standardized cognitive tests.
Articles were excluded if they met one of the following exclusion criteria: in vitro data only, conference abstract, review, editorial, or letter to the editor, or published in a language other than English.
2.2 Study Selection and Data Extraction
Two review authors (AM and SP) independently screened the titles and abstracts and subsequently the full text articles of potentially relevant studies against the inclusion and exclusion criteria. A third reviewer (ABM or LF) resolved any disagreements between the two review authors. All studies were divided into two groups: studies testing natural compounds (containing substances produced naturally by living organisms (Ouyang et al., 2014) and studies testing synthetic compounds.
The following variables were independently extracted for the included studies by two reviewers (AM and SP): author, year of publication, intervention, species/animal model, sample size (treatment group, control group), age, sex, duration of intervention, dose of therapeutic and type of model (dementia or normal ageing). For binary outcomes, the number of events, percentage of events or odds ratios with 95% confidence intervals were extracted. For continuous outcomes, the mean or median value, standard deviation, standard error, 95% confidence intervals or interquartile range, mean difference, and p-values were extracted.
2.3 Data Analysis
Interventions were considered preventative if the intervention was administered prior to the onset of dementia, with a delayed onset of dementia or reduced incidence rate after administration. Interventions were considered therapeutic if the intervention was administered after the dementia onset and subsequently slowed its progression or improved cognitive function.
An overall positive effect of the administered compound on cognitive performance was defined as positive finding on primary cognitive outcome, or >50% of the cognitive tests demonstrating a statistically significant improvement in the treatment group compared to the comparator group. A moderately-positive result was defined as ≥20% of the cognitive tests demonstrating a statistically significant improvement. A finding was considered negative where <20% of the cognitive outcomes were positive in the treatment group compared to the comparator group.
To investigate if the cognitive effect of compounds is dependent on study characteristics, studies were grouped based on study outcome (positive, moderately positive, negative), study population (MCI/dementia, normal ageing), the duration of intervention and the dosage of the compound.
2.4 Registered Clinical Trials in Humans
To identify ongoing and unpublished, completed human trials investigating GSK3 inhibitors, the ClinicalTrials.gov database (ClinicalTrials.gov, 2022) was screened for trials registered before 19 February 2022. Trials were included if GSK3 inhibition was mentioned, or compounds summarized in this review were utilized in combination with cognitive function, dementia, MCI, AD, VD, PD or LBD.
2.5 Risk of Bias
Two reviewers assessed the risk of bias (AM and SP). The Systematic Review Centre for Laboratory animal Experimentation’s risk of bias tool (SYRCLE) was used for animal studies (Hooijmans et al., 2014), which includes eight sources of bias: sequence generation, baseline characteristics, allocation concealment, random housing, blinding of personnel, random outcome assessment, incomplete outcome data, and selective outcome reporting. Results were denoted green for low risk, yellow for unclear risk, and red for high risk.
3 Results
3.1 Study Selection and Characteristics
The literature searches and selection process are illustrated in Figure 1. After exclusion of 4,154 duplicates, 3,517 articles were screened for title and abstracts of which 844 underwent full text screening. Thirty-two studies from 29 articles were included, investigating impaired nutrient sensing using compounds interfering with the GSK3 pathway on cognition in mice (19 studies), rats (12 studies) and zebrafish [one study (Koehler et al., 2019)] (Table 1). The Wistar rat was used most often to study dementia (8 out of 32 studies), whereas the C57BL/6 mice were used to study normal ageing [two studies (Jiang et al., 2020; Zhou et al., 2020)]. The sample size per treatment group varied between six (Xu et al., 2018; Lin et al., 2021) to 20 animals (Yang et al., 2013; Ibrahim et al., 2020). The majority of used animal dementia models were transgenic, overexpressing or producing mutant products of human genes such as amyloid precursor protein (APP) [Jiang et al., 2020; Zhou et al., 2020; Lin et al., 2021) and tau (Zhang et al., 2018)]. Dementia was also induced by administration of intracerebroventricular Aβ (Huang et al., 2019; Akhtar et al., 2020; Fan et al., 2020; Sun et al., 2020; Yan et al., 2021) or streptozotocin (Tang et al., 2018; Wang et al., 2018; Zhu et al., 2019; Akhtar et al., 2020). Cognitive tests used in the trials included Morris water maze, open field test, Barnes maze task, elevated plus maze, locomotor activity, Y-maze test, passive avoidance test, passive avoidance learning and novel object recognition test. Details of the tests are summarized in Supplementary Table 1. GSK3 inhibition was studied at different stages of the nutrient sensing pathway, which is illustrated in Figure 2.
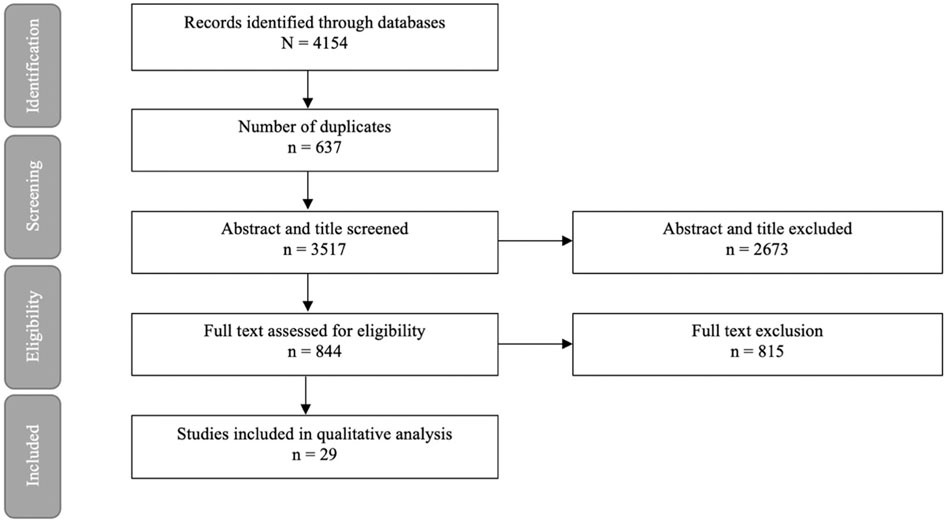
FIGURE 1. Article selection process.
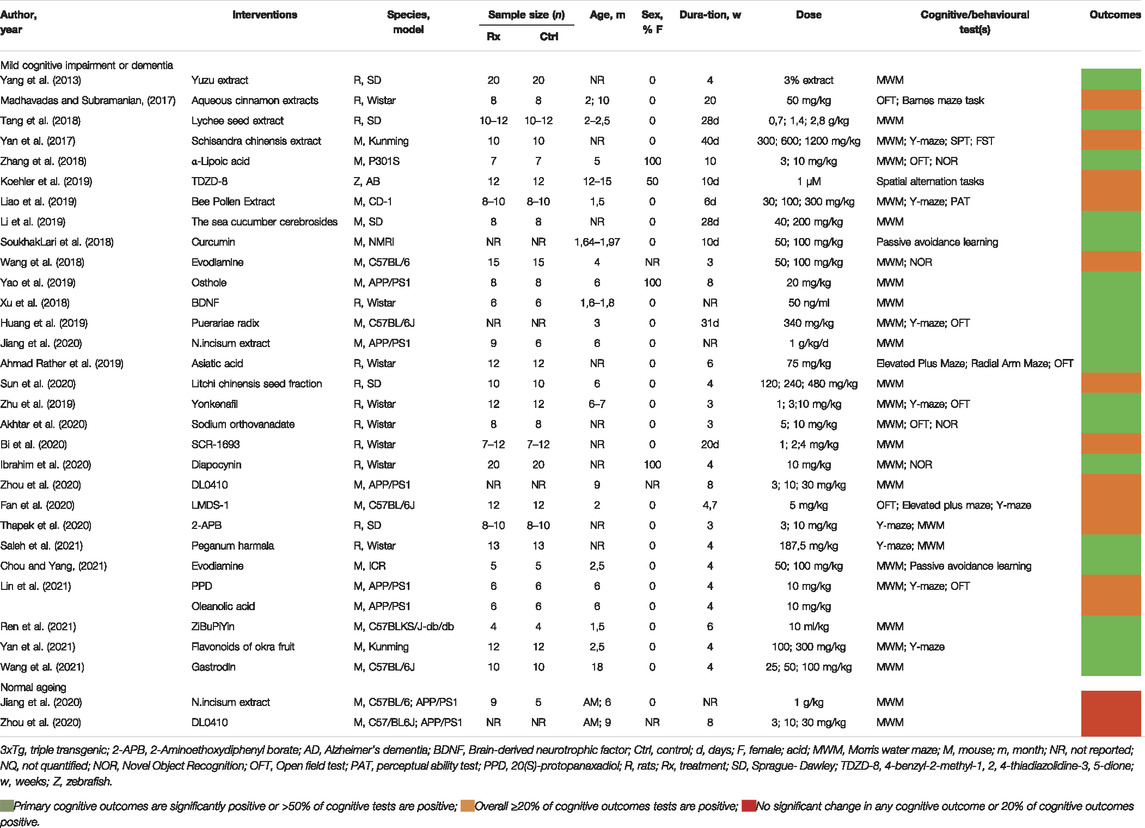
TABLE 1. Characteristics of studies testing the effect of compounds interfering with the GSK3 pathway on cognition.
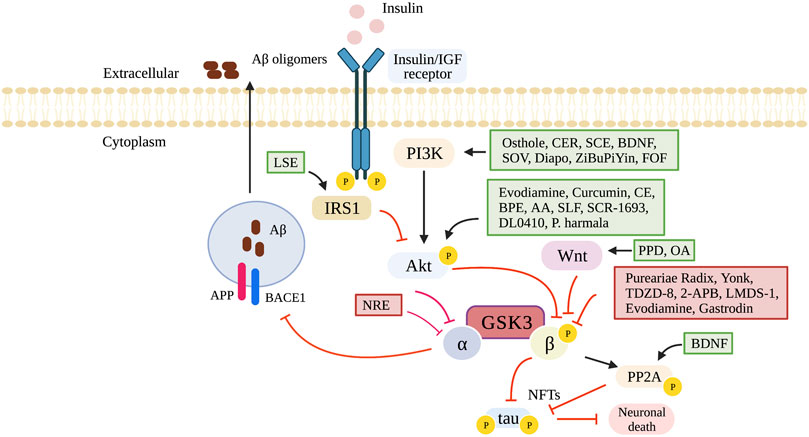
FIGURE 2. GSK3 pathway and its modulating compounds. 2-APB, 2-Aminoethoxydiphenyl borate; AA, Asiatic acid; Akt, Protein kinase-B (serine/threonine kinase); APP, amyloid precursor protein; BACE1, beta-site APP cleaving enzyme 1; BDNF, brain-derived neurotrophic factor; BPE, bee pollen extract; CE, cinnamon extract; CER, sea cucumber cerebrosides; Diapo, diapocynin; FOF, flavonoids of okra fruit; GSK3, glycogen synthase kinase-3; Insulin/IGF receptor, insulin-like growth factor 1 receptor; IRS1, insulin receptor substrate 1; NFTs, neurofibrillary tangles; NRE, N.incisum extract; OA, Oleanolic acid; P, phosphorylation; P. harmala, Peganum harmala; PPD, 20(S)-protopanaxadiol; PI3K, phosphoinositide 3-kinase; PP2A, protein phosphatase 2 (alpha isoform); SCE, Schisandra chinensis extract; SLF, seed of litchi chinensis fraction; SOV, sodium orthovanodate; Yonk, Yonkenafil. Created in BioRender.com.
3.2 Natural Compounds
Twenty-two out of 32 studies tested naturally derived compounds, they include: compounds rich in flavonoids such as yuzu extract (Yang et al., 2013), bee pollen extract (Liao et al., 2019), N. incisum extract (Jiang et al., 2020) and flavonoids of okra fruit (Yan et al., 2021); compounds containing alkaloids, such as Puerariae radix (Huang et al., 2019), ZiBuPiYin (Ren et al., 2021), evodiamine (Wang et al., 2018; Chou and Yang, 2021) and Peganum harmala (Saleh et al., 2021); aquesous cinnamon extracts (Madhavadas and Subramanian, 2017), which contain type A linked procyanidin trimer; schisandra chinensis extract (Yan et al., 2017), which contains ligand diabenzo [a, c] cyclooctadiene; sea cucumber cerebrosides (Li et al., 2019), which is a class of neural glycospinolipids; osthole (Yao et al., 2019) which is a derivative of coumarin; curcumin (SoukhakLari et al., 2018), which is the main ingredient of turmeric; litchi chinensis seed fraction (Sun et al., 2020), which entails phenolic compounds and glycosides; Asiatic acid (Ahmad Rather et al., 2019), which is a pentacyclic triterpenoid; Yonkenafil (Zhu et al., 2019), which is a novel phosphodiesterase type 5 inhibitor and an analogue of sildenafil; 20(S)-protopanaxadiol, oleanolic acid (Lin et al., 2021), which is an active compound found in ginseng and a Chinese herb gastrodin (Wang et al., 2021).
Thirteen studies using natural compounds showed an overall positive effect (Yang et al., 2013; SoukhakLari et al., 2018; Tang et al., 2018; Ahmad Rather et al., 2019; Huang et al., 2019; Li et al., 2019; Yao et al., 2019; Jiang et al., 2020; Chou and Yang, 2021; Ren et al., 2021; Saleh et al., 2021; Wang et al., 2021; Yan et al., 2021) on cognition, seven studies a moderately positive (Madhavadas and Subramanian, 2017; Yan et al., 2017; Wang et al., 2018; Liao et al., 2019; Zhu et al., 2019; Sun et al., 2020; Lin et al., 2021) and one a negative effect (Jiang et al., 2020). The effect of natural compounds on cognition were tested in dementia animal models in rat (n = 7 out of 21 studies); mice (n = 14 out of 21 studies), and in normal ageing animal model in mice [n = 1 out of 1 study (Jiang et al., 2020)]. A dose-response relationship was examined in ten studies, of which seven studies (Yan et al., 2017, 2021; Wang et al., 2018; Liao et al., 2019; Zhu et al., 2019; Sun et al., 2020; Chou and Yang, 2021) showed that a higher dose led to better cognitive outcomes. The duration of administered natural compounds ranged from 6 days (Liao et al., 2019) to 20 weeks (Madhavadas and Subramanian, 2017) and did not affect the outcome.
The following compounds improved spatial memory: seed of litchi chinensis fraction, sea cucumber cerebrosides, evodiamine, osthole, Pureariae Radix, N.incisum extract, Asiatic acid, seed of litchi chinensis fraction, Yonkenafil, P. harmala (Tang et al., 2018; Wang et al., 2018; Ahmad Rather et al., 2019; Huang et al., 2019; Li et al., 2019; Yao et al., 2019; Zhu et al., 2019; Jiang et al., 2020; Sun et al., 2020; Saleh et al., 2021). Pureariae Radix showed improvement in recognition memory (Huang et al., 2019). Short-term memory was improved after administration of yuzu, schisandra chinensis extract, Pureariae Radix (Yang et al., 2013; Yan et al., 2017; Huang et al., 2019) and bee pollen extract showed improvement in long-term memory (Liao et al., 2019). Compound yuzu did not show any improvement in long-term memory (Yang et al., 2013). Motor activity did not improve after administration of schisandra chinensis extract and Yonkenafil, flavonoids of okra fruit (Yan et al., 2017, 2021; Zhu et al., 2019).
3.3 Synthetic Compounds
Ten out of 32 studies tested synthetic compounds including: SCR-1693 (Bi et al., 2020), an acetylcholinesterase inhibitor and calcium channel blocker; brain-derived neurotrophic factor (BDNF) (Xu et al., 2018); α-Lipoic acid (Zhang et al., 2018); diapocynin (Ibrahim et al., 2020), a NOX inhibitor, sodium orthovanadate (Akhtar et al., 2020), a tyrosine phosphatase inhibitor; DL0410 (Zhou et al., 2020) cholinesterase inhibitor; 2-APB (Thapak et al., 2020), which inhibit both inositol trisphosphate (IP3) receptors and transient receptor potential (TRP) channels; LMDS-1 (Fan et al., 2020), a potential TrkB receptor agonist and TDZD-8 (Koehler et al., 2019), a selective non-ATP competitive inhibitor of glycogen synthase kinase 3 beta (GSK3β).
Four studies using synthetic compounds showed an overall positive effect on cognition (Xu et al., 2018; Zhang et al., 2018; Akhtar et al., 2020; Ibrahim et al., 2020), five a moderately positive (Koehler et al., 2019; Bi et al., 2020; Fan et al., 2020; Thapak et al., 2020; Zhou et al., 2020) and one a negative effect (Zhou et al., 2020). The effect of synthetic compounds on cognition were studied in dementia animal models in rat [n = 5 out of 9 studies (Xu et al., 2018; Akhtar et al., 2020; Bi et al., 2020; Ibrahim et al., 2020; Thapak et al., 2020)], mice [n = 3 out of 9 studies (Zhang et al., 2018; Fan et al., 2020; Zhou et al., 2020)], zebrafish [n = 1 out of 9 studies (Koehler et al., 2019)] and in normal animal ageing model in mice [n = 1 out of 1 study (Zhou et al., 2020)].
Six studies (Zhang et al., 2018; Akhtar et al., 2020; Bi et al., 2020; Thapak et al., 2020; Zhou et al., 2020; Wang et al., 2021) investigated a dose-response relationship. For SCR-1693 (Bi et al., 2020), a higher dose gave better cognitive outcomes. The duration of administered compounds ranged from 10 days (Koehler et al., 2019) to 10 weeks (Zhang et al., 2018). No effect of the duration of the administrated compounds on cognition was found.
The following compounds improved spatial memory: α-Lipoic acid, BDNF, sodium orthovanadate, SCR-1693, diapocynin, DL0410, 2-APB (Xu et al., 2018; Zhang et al., 2018; Akhtar et al., 2020; Bi et al., 2020; Ibrahim et al., 2020; Thapak et al., 2020; Zhou et al., 2020). Sodium orthovanadate showed improvement in recognition memory (Akhtar et al., 2020). LMDS-1 improved short-term memory but not long-term memory (Fan et al., 2020). Motor activity did not improve after administration of the following compounds: α-Lipoic acid, sodium orthovanodate, diapocynin (Zhang et al., 2018; Akhtar et al., 2020; Ibrahim et al., 2020).
3.4 Human Registered Trials
No trials were found mentioning the inhibition of the GSK3 pathway and cognitive outcomes. Based on the compounds being described in this review, three human trials were found testing curcumin to improve cognition. One active, not recruiting phase 2 trial investigates curcumin (with aerobic yoga exercise) on levels of blood biomarkers and cognitive function in individuals with MCI or subjective cognitive impairment (NCT01811381). A completed phase 2 trials investigate the effect of curcumin on physical function and cognition in older adults (NCT03085680), and another phase 2 trial the effect of curcumin on mental capacity and amyloid-beta blood concentrations in AD patients (NCT01001637).
3.5 Risk of Bias Across Studies
Table 2 shows the SYRCLE risk of bias ratings for animal studies. All studies showed an unclear risk of bias on allocation concealment, random housing, blinding of personnel and random outcome assessment. A low risk was found across the sequence generation, baseline characteristics, incomplete outcome data and selective outcome reporting. Eleven studies showed unclear risk in the sequence generation criterion (Yang et al., 2013; Madhavadas and Subramanian, 2017; Tang et al., 2018; Wang et al., 2018; Ahmad Rather et al., 2019; Huang et al., 2019; Koehler et al., 2019; Liao et al., 2019; Akhtar et al., 2020; Thapak et al., 2020; Saleh et al., 2021). Unclear risk was mainly due to the lack of reported data.
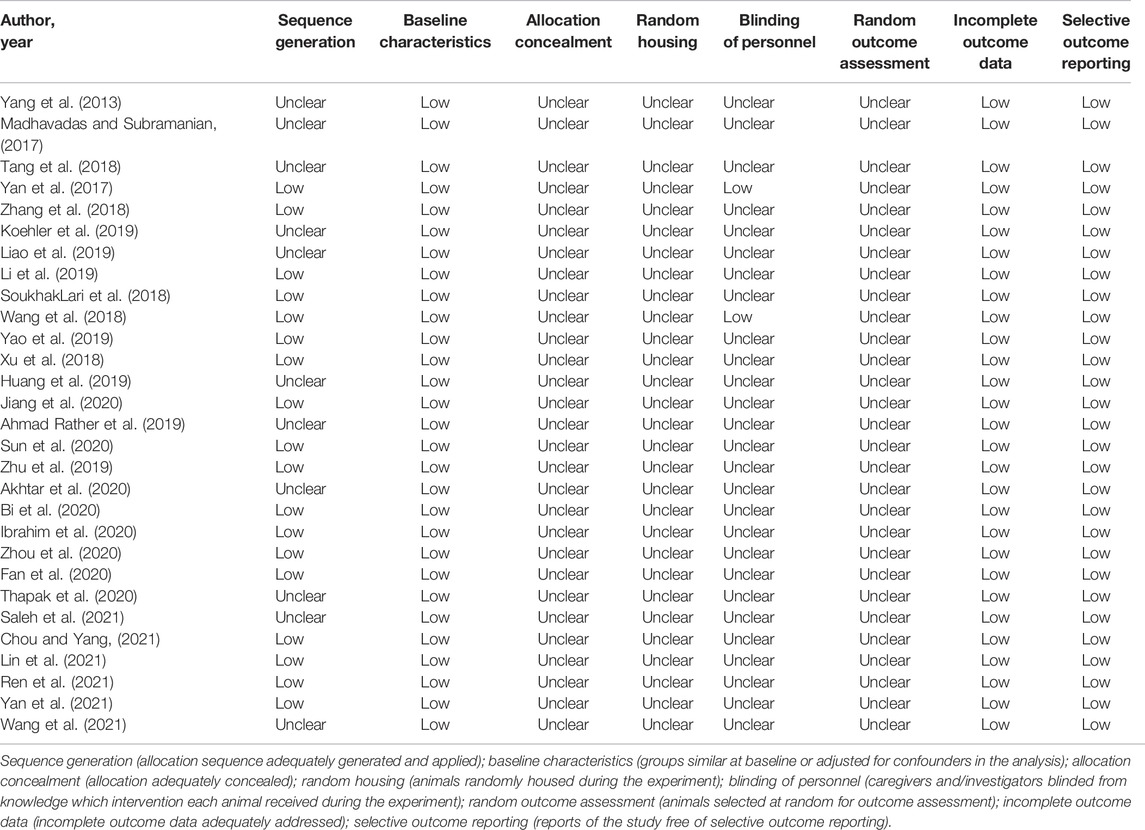
TABLE 2. SYRCLE risk of bias for animal studies.
4 Discussion
Compounds inhibiting the GSK3 pathway may improve cognition in animals with cognitive impairment, but there is limited evidence for delaying cognitive decline or dementia onset in ageing animal models.
The GSK3 pathway can be inhibited at different stages of the nutrient sensing pathway. For the compounds osthole, sea cucumber cerebrosides, schisandra chinensis extract, brain-derived neurotrophic factor, sodium orthovanodate, diapocynin, ZiBuPiYin and flavonoids of okra fruit, GSK3β is inhibited by activation of the PI3K, and subsequent activation of Akt impaired GSK3 phosphorylation (Yan et al., 2017, 2021; Xu et al., 2018; Li et al., 2019; Yao et al., 2019; Akhtar et al., 2020; Ibrahim et al., 2020; Ren et al., 2021). The PI3K/Akt pathway is a key for regulating cell survival, growth and metabolism, which is important for cognitive function such as learning and memory (Shu et al., 2013). It plays an important role in apoptosis and autophagy in the nervous system, but also in reducing neuronal and nerve damage (Li et al., 2016). There is evidence that the insulin-PI3K/Akt pathway is reduced in AD, which makes compounds activating this pathway promising for the treatment of cognitive decline (Kong et al., 2013). Of the aforementioned compounds, osthole has been shown to inhibit the PI3K/Akt pathway via the activation of PTEN (Zhu et al., 2018). Brain-derived neurotrophic factor and diapocynin inhibits PI3K and AKT directly. Sodium orthovanadate inhibits PI3K and AKT downstream via the inhibition of PTP (Takada et al., 2004). The exact mechanism for sea cucumber cerebroside, and schisandra chinensis inhibition of this pathway remains uncertain.
The compounds evodiamine, curcumin, cinnamon extract, bee pollen extract, Asiatic acid, seed of litchi chinensis fraction, SCR-1693, DL0410 and P.harmala activate Akt, which leads to GSK3 phosphorylation impairment (Madhavadas and Subramanian, 2017; SoukhakLari et al., 2018; Tang et al., 2018; Wang et al., 2018; Ahmad Rather et al., 2019; Liao et al., 2019; Bi et al., 2020; Zhou et al., 2020; Chou and Yang, 2021; Saleh et al., 2021). Akt is important in protecting injured neurons (Huang et al., 2017) and promoting neurite outgrowth (Park et al., 2012). In early stages of neurofibrillary pathology as well as in hippocampal neurons in type 2 diabetes animal models, the levels of phosphorylated Akt are downregulated (Griffin et al., 2005; Xiang et al., 2015). Asiatic acid reduces oxidant mediated neuronal apoptosis, which leads to the inhibition of GSK3β (Ahmad Rather et al., 2019).
The compounds Pureariae Radix, Yonkenafil, TDZD-8, 2-APB, evodiamine, LMDS-1 and gastrodin directly modulate GSK3β (Wang et al., 2018; Huang et al., 2019; Koehler et al., 2019; Zhu et al., 2019; Fan et al., 2020; Thapak et al., 2020; Chou and Yang, 2021; Wang et al., 2021). Pureariae Radix prevents the decrease of inactive GSK3β Ser9 phosphorylation (Huang et al., 2019). Yonkenafil activates the phosphorylation of insulin receptor substrate 1 (IRS-1), leading to the phosphorylation of GSK3 (Zhu et al., 2019). TDZD-8 decreases the ratio of active/inactive GSK3β, and subsequently decreases pTau (Ser199) (Koehler et al., 2019). 20(S)-protopanaxadiol and oleanolic acid inhibit GSK3β via Wnt/GSK3β/β-catenin pathway activation (Lin et al., 2021).
The brain-derived neurotrophic factor activates protein phosphatase 2 (alpha isoform) (PP2A), which leads to a decrease in p-Tau. The result is a dephosphorylation of active GSK3β and an enhancement of inactive GSK3β Ser9 (Xu et al., 2018). Compound N. incisum extract modulates extracellular Aß production caused by unphosphorylated GSK3β. Specifically, it suppresses the expression of the cleaving enzyme beta-site APP cleaving enzyme 1 (BACE1), preventing the cleavage of GSK3 substrate APP. The result is a decrease in Aß production and subsequently the decrease of p-Tau and neuronal death (Jiang et al., 2020). Compound lychee seed extract reduces the deposit of Aß associated with insulin dysfunction and resistance (Tang et al., 2018). IRS-1 is activated by catechin, procyanidin A1 and procyanidin A2 derived from lychee seed extract. The result is the inhibition in the activity of GSK3β in the PI3K/Akt/GSK3 pathway (Xiong et al., 2020).
The GSK3 pathway is one of the multi-aetiological factors implicated in insulin resistance (Leng et al., 2010), which might be of importance as more than 80% of patients with AD have either type 2 diabetes mellitus or an impaired fasting glucose (Barbagallo, 2014). Disturbed glucose metabolism has also been shown to increase the risk of developing dementia by 50% (Biessels et al., 2014).
Inflammation is one of the main features observed in both type 2 diabetes mellitus (Tsalamandris et al., 2019) and AD as well as other neurodegenerative diseases (Guzman-Martinez et al., 2019). Yang et al. (2013), Ahmad Rather et al. (2019), Huang et al. (2019), Yao et al. (2019), Zhu et al. (2019), Ibrahim et al. (2020), Jiang et al. (2020), Sun et al. (2020), Zhou et al. (2020), Chou and Yang (2021), Yan et al. (2021) of the investigated compounds have anti-inflammatory properties in additional to their GSK3 inhibitory properties. Low grade systematic chronic inflammation is a hallmark of human ageing and an impaired immune response. Therewith, anti-inflammatory properties could be another important pathway contributing to the cognitive benefits of such compounds (Tangestani Fard and Stough, 2019).
Four studies investigated N. incisum extract, Yonkenafil and DL0410 showed that these compounds decrease necrosis factor alpha (TNF alpha) or interleukin 6 (IL-6). It is noted that levels of these two cytokines are elevated in AD and type 2 diabetes mellitus (55). TNF alpha activates the c-Jun kinase (JNK), which leads to the inhibition of serine phosphorylation of IRS and blockade of insulin signalling (Gao and Ye, 2012). Interestingly, high levels of insulin lead to increasing levels of TNF alpha and IL-6. Hyperinsulinemia accelerates inflammation and increases Aß levels, leading to AD pathogenesis (Fishel et al., 2005; Chatterjee and Mudher, 2018). Schisandra chinensis extract, diapocynin and sodium orthovanadate promote the expression of brain-derived neurotrophic factor gene which promotes neurogenesis, modulates neuronal activity and may downregulate the production of Aß (Buchman et al., 2016). Lychee seed extract, evodiamine and SCR-1693 promotes significant decrease in acetylcholinesterase (AChE) activity. It is known that AChE inhibition is an efficient therapeutic for dementia through the rescue of cholinergic deficit (Eldufani and Blaise, 2019).
Impaired spatial memory is an early clinical sign of AD (Zhu et al., 2017). Compounds seed of litchi chinensis fraction, α-Lipoic acid, sea cucumber cerebrosides, evodiamine, osthole, brain-derived neurotrophic factor, Pureariae Radix, N.incisum extract, Asiatic acid, seed of litchi chinensis fraction, Yonkenafil, sodium orthovanodate, SCR-1693, diapocynin, DL0410, 2-APB, P. harmala (Tang et al., 2018; Wang et al., 2018; Xu et al., 2018; Zhang et al., 2018; Ahmad Rather et al., 2019; Huang et al., 2019; Li et al., 2019; Yao et al., 2019; Zhu et al., 2019; Akhtar et al., 2020; Bi et al., 2020; Ibrahim et al., 2020; Jiang et al., 2020; Sun et al., 2020; Thapak et al., 2020; Zhou et al., 2020; Saleh et al., 2021) showed improvement in spatial memory, which is consistent with the effects of other GSK3 inhibitors such as SB 216763 (Nguyen et al., 2018) or lithium, the first FDA (Food and Drug Administration) approved GSK3 inhibitor (Liu et al., 2020). Compounds yuzu, schisandra chinensis extract, Pureariae Radix and LMDS-1 (Yang et al., 2013; Yan et al., 2017; Huang et al., 2019; Fan et al., 2020) improved short-term memory which is commonly impaired in AD (Della Sala et al., 2012). Loss of long-term memory is also characteristic for AD (Parra et al., 2009) and bee pollen extract showed improvement in long-term memory (Liao et al., 2019).
GSK-3β can phosphorylate tau protein at various sites in vitro and in cell culture models and the epitopes are consistent with those found to be hyperphosphorylated in AD brains. Consequently, activation of GSK-3β leads to aggregation of the tau protein (Lei et al., 2011). Activated GSK-3β is also involved in Aβ formation and accumulation in human AD brains because it modulates the cleavage of APP (Lauretti et al., 2020). These mechanisms could be potentially targeted by GSK3 inhibitors. There has consistently been a poor translation of successful therapeutics of pre-clinical animal dementia models to successful interventions in human dementia clinical trials (Franco and Cedazo-Minguez, 2014).Only two compounds included in this review, α-Lipoic acid and curcumin were studied in human clinical trials. Results from one of the trials concluded that α-Lipoic acid can slow cognitive decline (Shinto et al., 2013). Three trials concluded that the administration of curcumin can disaggregate the Aß deposit in the brain (Baum et al., 2008), prevent Aß aggregation (Begum et al., 2008) and improve memory in healthy adults (Small et al., 2018).
Compounds tested in normal ageing animal models did not show cognitive improvement (Jiang et al., 2020; Zhou et al., 2020). Most of the animal models rely on the utilization of genetic mutations associated with familial AD, while the most common form of AD is sporadic AD (LaFerla and Green, 2012). Furthermore, publication bias cannot be ruled out, particularly in animal studies which are unlikely to be registered and may be less likely to be published if results are negative (van der Naald et al., 2020). It was not possible to perform formal statistical analysis due to small sample sizes of studies examining a number of compounds and the conclusion was based on reported p-values. The significance of p-values is influenced by the sample size, which is often small in animal studies. The search included studies, which investigated the modulation of the GSK3 pathway. Therefore, compounds potentially inhibiting the GSK3 pathway without mentioning the pathway could have been missed. Finally, information on baseline characteristics, outcome assessment, blinding of personnel, housing, allocation concealment, and sequence generation, were often unclear. Future animal trials should consider following the SYRCLE guidelines (Hooijmans et al., 2014).
5 Conclusion
The results of this systematic review suggest that the investigated compounds can improve cognitive function in MCI and dementia animal models. Further studies are required to fully elucidate the potential of GSK3 inhibitors in MCI/dementia as well as initiation of clinical trials in humans with compounds inhibiting the GSK3 pathway.
Author Contributions
ABM designed the article, AM, SK, LF and ABM analysed and interpreted the data, and AM and SK drafted the article and LF and ABM revised it critically for important intellectual content, and all authors approved the version to be published, and all authors agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Funding
This work was funded by the University of Melbourne and the Medical Research Future Fund via the Melbourne Academic Centre for Health. The funder has no role in the design and conduct of the review, the analysis and interpretation of the data.
Conflict of Interest
AM is a scientific co-founder of Cognescence Ltd. and Immunidex Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank Patrick Condron from the Brownless Biomedical Library, University of Melbourne, for his assistance with the search strategy. We thank Daniel S. Heard and Camilla S. L. Tuttle for their help in conducting the literature search.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fragi.2022.898853/full#supplementary-material
References
Ahmad Rather, M., Justin-Thenmozhi, A., Manivasagam, T., Saravanababu, C., Guillemin, G. J., and Essa, M. M. (2019). Asiatic Acid Attenuated Aluminum Chloride-Induced Tau Pathology, Oxidative Stress and Apoptosis via AKT/GSK-3β Signaling Pathway in Wistar Rats. Neurotox. Res. 35, 955–968. doi:10.1007/s12640-019-9999-2
Akhtar, A., Bishnoi, M., and Sah, S. P. (2020). Sodium Orthovanadate Improves Learning and Memory in Intracerebroventricular-Streptozotocin Rat Model of Alzheimer’s Disease through Modulation of Brain Insulin Resistance Induced Tau Pathology. Brain Res. Bull. 164, 83–97. doi:10.1016/j.brainresbull.2020.08.001
Barbagallo, M. (2014). Type 2 Diabetes Mellitus and Alzheimer’s Disease. World J. Diabetes 5, 889. doi:10.4239/wjd.v5.i6.889
Baum, L., Lam, C. W. K., Cheung, S. K.-K., Kwok, T., Lui, V., Tsoh, J., et al. (2008). Six-Month Randomized, Placebo-Controlled, Double-Blind, Pilot Clinical Trial of Curcumin in Patients with Alzheimer Disease. J. Clin. Psychopharmacol. 28, 110–113. doi:10.1097/jcp.0b013e318160862c
Begum, A. N., Jones, M. R., Lim, G. P., Morihara, T., Kim, P., Heath, D. D., et al. (2008). Curcumin Structure-Function, Bioavailability, and Efficacy in Models of Neuroinflammation and Alzheimer’s Disease. J. Pharmacol. Exp. Ther. 326, 196–208. doi:10.1124/jpet.108.137455
Beurel, E., Grieco, S. F., and Jope, R. S. (2015). Glycogen Synthase Kinase-3 (GSK3): Regulation, Actions, and Diseases. Pharmacol. Ther. 148, 114–131. doi:10.1016/j.pharmthera.2014.11.016
Bhat, R. V., Andersson, U., Andersson, S., Knerr, L., Bauer, U., and Sundgren-Andersson, A. K. (2018). The Conundrum of GSK3 Inhibitors: Is it the Dawn of a New Beginning? J. Alzheimers Dis. 64, S547–S554. doi:10.3233/JAD-179934
Bi, A., An, W., Wang, C., Hua, Y., Fang, F., Dong, X., et al. (2020). SCR-1693 Inhibits Tau Phosphorylation and Improves Insulin Resistance Associated Cognitive Deficits. Neuropharmacology 168, 108027. doi:10.1016/j.neuropharm.2020.108027
Biessels, G. J., Strachan, M. W. J., Visseren, F. L. J., Kappelle, L. J., and Whitmer, R. A. (2014). Dementia and Cognitive Decline in Type 2 Diabetes and Prediabetic Stages: towards Targeted Interventions. Lancet Diabetes Endocrinol. 2, 246–255. doi:10.1016/S2213-8587(13)70088-3
Buchman, A. S., Yu, L., Boyle, P. A., Schneider, J. A., De Jager, P. L., and Bennett, D. A. (2016). Higher Brain BDNF Gene Expression Is Associated with Slower Cognitive Decline in Older Adults. Neurology 86, 735–741. doi:10.1212/WNL.0000000000002387
Chatterjee, S., and Mudher, A. (2018). Alzheimer’s Disease and Type 2 Diabetes: A Critical Assessment of the Shared Pathological Traits. Front. Neurosci. 12, 383. doi:10.3389/fnins.2018.00383
Chou, C.-H., and Yang, C.-R. (2021). Neuroprotective Studies of Evodiamine in an Okadaic Acid-Induced Neurotoxicity. Ijms 22, 5347. doi:10.3390/ijms22105347
de Lucia, C., Murphy, T., Steves, C. J., Dobson, R. J. B., Proitsi, P., and Thuret, S. (2020). Lifestyle Mediates the Role of Nutrient-Sensing Pathways in Cognitive Aging: Cellular and Epidemiological Evidence. Commun. Biol. 3, 157. doi:10.1038/s42003-020-0844-1
Della Sala, S., Parra, M. A., Fabi, K., Luzzi, S., and Abrahams, S. (2012). Short-term Memory Binding Is Impaired in AD but Not in Non-AD Dementias. Neuropsychologia 50, 833–840. doi:10.1016/j.neuropsychologia.2012.01.018
Efeyan, A., Comb, W. C., and Sabatini, D. M. (2015). Nutrient-sensing Mechanisms and Pathways. Nature 517, 302–310. doi:10.1038/nature14190
Eldufani, J., and Blaise, G. (2019). The Role of Acetylcholinesterase Inhibitors Such as Neostigmine and Rivastigmine on Chronic Pain and Cognitive Function in Aging: A Review of Recent Clinical Applications. Alzheimers Dement (N Y) 5, 175–183. doi:10.1016/j.trci.2019.03.004
Fan, C.-H., Lin, C.-W., Huang, H.-J., Lee-Chen, G.-J., Sun, Y.-C., Lin, W., et al. (2020). LMDS-1, a Potential TrkB Receptor Agonist Provides a Safe and Neurotrophic Effect for Early-phase Alzheimer’s Disease. Psychopharmacology 237, 3173–3190. doi:10.1007/s00213-020-05602-z
Fishel, M. A., Watson, G. S., Montine, T. J., Wang, Q., Green, P. S., Kulstad, J. J., et al. (2005). Hyperinsulinemia Provokes Synchronous Increases in Central Inflammation and β-Amyloid in Normal Adults. Arch. Neurol. 62, 1539–44. doi:10.1001/archneur.62.10.noc50112
Franco, R., and Cedazo-Minguez, A. (2014). Successful Therapies for Alzheimer’s Disease: Why So Many in Animal Models and None in Humans? Front. Pharmacol. 5, 146. doi:10.3389/fphar.2014.00146
Gale, S. A., Acar, D., and Daffner, K. R. (2018). Dementia. Am. J. Med. 131, 1161–1169. doi:10.1016/j.amjmed.2018.01.022
Gao, Z.-g., and Ye, J.-p. (2012). Why Do Anti-inflammatory Therapies Fail to Improve Insulin Sensitivity? Acta Pharmacol. Sin. 33, 182–188. doi:10.1038/aps.2011.131
Giese, K. P. (2009). GSK-3: A Key Player in Neurodegeneration and Memory. IUBMB Life 61, 516–521. doi:10.1002/iub.187
Griffin, R. J., Moloney, A., Kelliher, M., Johnston, J. A., Ravid, R., Dockery, P., et al. (2005). Activation of Akt/PKB, Increased Phosphorylation of Akt Substrates and Loss and Altered Distribution of Akt and PTEN Are Features of Alzheimer’s Disease Pathology. J. Neurochem. 93, 105–117. doi:10.1111/j.1471-4159.2004.02949.x
Guzman-Martinez, L., Maccioni, R. B., Andrade, V., Navarrete, L. P., Pastor, M. G., and Ramos-Escobar, N. (2019). Neuroinflammation as a Common Feature of Neurodegenerative Disorders. Front. Pharmacol. 10, 1008. doi:10.3389/fphar.2019.01008
Hooijmans, C. R., Rovers, M. M., de Vries, R. B., Leenaars, M., Ritskes-Hoitinga, M., and Langendam, M. W. (2014). SYRCLE’s Risk of Bias Tool for Animal Studies. BMC Med. Res. Methodol. 14, 43. doi:10.1186/1471-2288-14-43
Huang, H.-J., Huang, C.-Y., Lee, M., Lin, J.-Y., and Hsieh-Li, H. M. (2019). Puerariae Radix Prevents Anxiety and Cognitive Deficits in Mice under Oligomeric Aβ-Induced Stress. Am. J. Chin. Med. 47, 1459–1481. doi:10.1142/S0192415X19500757
Huang, H., Liu, H., Yan, R., and Hu, M. (2017). PI3K/Akt and ERK/MAPK Signaling Promote Different Aspects of Neuron Survival and Axonal Regrowth Following Rat Facial Nerve Axotomy. Neurochem. Res. 42, 3515–3524. doi:10.1007/s11064-017-2399-1
Ibrahim, W. W., Ismail, H. M., Khattab, M. M., and Abdelkader, N. F. (2020). Cognitive Enhancing Effect of Diapocynin in D-Galactose-Ovariectomy-Induced Alzheimer’s-like Disease in Rats: Role of ERK, GSK-3β, and JNK Signaling. Toxicol. Appl. Pharmacol. 398, 115028. doi:10.1016/j.taap.2020.115028
Jiang, X.-W., Liu, W.-W., Wu, Y.-T., Wu, Q., Lu, H.-Y., Xu, Z.-H., et al. (2020). Notopterygium Incisum Extract (NRE) Rescues Cognitive Deficits in APP/PS1 Alzhneimer’s Disease Mice by Attenuating Amyloid-Beta, Tau, and Neuroinflammation Pathology. J. Ethnopharmacol. 249, 112433. doi:10.1016/j.jep.2019.112433
Jolivalt, C. G., Lee, C. A., Beiswenger, K. K., Smith, J. L., Orlov, M., Torrance, M. A., et al. (2008). Defective Insulin Signaling Pathway and Increased Glycogen Synthase Kinase-3 Activity in the Brain of Diabetic Mice: Parallels with Alzheimer’s Disease and Correction by Insulin. J. Neurosci. Res. 86, 3265–3274. doi:10.1002/jnr.21787
Jope, R., and Roh, M.-S. (2006). Glycogen Synthase Kinase-3 (GSK3) in Psychiatric Diseases and Therapeutic Interventions. Curr. Drug Targets 7, 1421–1434. doi:10.2174/1389450110607011421
Katsuno, M., Sahashi, K., Iguchi, Y., and Hashizume, A. (2018). Preclinical Progression of Neurodegenerative Diseases. Nagoya J. Med. Sci. 80, 289–298. doi:10.18999/nagjms.80.3.289
Koehler, D., Shah, Z. A., and Williams, F. E. (2019). The GSK3β Inhibitor, TDZD-8, Rescues Cognition in a Zebrafish Model of Okadaic Acid-Induced Alzheimer’s Disease. Neurochem. Int. 122, 31–37. doi:10.1016/j.neuint.2018.10.022
Kong, J., Ren, G., Jia, N., Wang, Y., Zhang, H., Zhang, W., et al. (2013). Effects of Nicorandil in Neuroprotective Activation of PI3K/AKT Pathways in a Cellular Model of Alzheimer’s Disease. Eur. Neurol. 70, 233–241. doi:10.1159/000351247
L'Episcopo, F., Drouin-Ouellet, J., Tirolo, C., Pulvirenti, A., Giugno, R., Testa, N., et al. (2016). GSK-3β-induced Tau Pathology Drives Hippocampal Neuronal Cell Death in Huntington’s Disease: Involvement of Astrocyte-Neuron Interactions. Cell Death Dis. 7, e2206. doi:10.1038/cddis.2016.104
LaFerla, F. M., and Green, K. N. (2012). Animal Models of Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2, a006320. doi:10.1101/cshperspect.a006320
Lauretti, E., Dincer, O., and Praticò, D. (2020). Glycogen Synthase Kinase-3 Signaling in Alzheimer’s Disease. Biochim. Biophys. Acta (BBA) - Mol. Cell Res. 1867, 118664. doi:10.1016/j.bbamcr.2020.118664
Lei, P., Ayton, S., Bush, A. I., and Adlard, P. A. (2011). GSK-3 in Neurodegenerative Diseases. Int. J. Alzheimer's Dis. 2011, 1–9. doi:10.4061/2011/189246
Leng, S., Zhang, W., Zheng, Y., Liberman, Z., Rhodes, C. J., Eldar-Finkelman, H., et al. (2010). Glycogen Synthase Kinase 3β Mediates High Glucose-Induced Ubiquitination and Proteasome Degradation of Insulin Receptor Substrate 1. J. Endocrinol. 206, 171–181. doi:10.1677/JOE-09-0456
Li, Q., Che, H. X., Wang, C. C., Zhang, L. Y., Ding, L., Xue, C. H., et al. (2019). Cerebrosides from Sea Cucumber Improved Aβ1-42 -Induced Cognitive Deficiency in a Rat Model of Alzheimer’s Disease. Mol. Nutr. Food Res. 63, 1800707. doi:10.1002/mnfr.201800707
Li, X., Qi, Z., Zhao, L., and Yu, Z. (2016). Astaxanthin Reduces Type 2 Diabetic-Associated Cognitive Decline in Rats via Activation of PI3K/Akt and Attenuation of Oxidative Stress. Mol. Med. Rep. 13, 973–979. doi:10.3892/mmr.2015.4615
Liao, Y., Bae, H. J., Zhang, J., Kwon, Y., Koo, B., Jung, I. H., et al. (2019). The Ameliorating Effects of Bee Pollen on Scopolamine-Induced Cognitive Impairment in Mice. Biol. Pharm. Bull. 42, 379–388. doi:10.1248/bpb.b18-00552
Lin, K., Sze, S. C.-W., Liu, B., Zhang, Z., Zhang, Z., Zhu, P., et al. (2021). 20(S)-protopanaxadiol and Oleanolic Acid Ameliorate Cognitive Deficits in APP/PS1 Transgenic Mice by Enhancing Hippocampal Neurogenesis. J. Ginseng Res. 45, 325–333. doi:10.1016/j.jgr.2020.07.003
Liu, M., Qian, T., Zhou, W., Tao, X., Sang, S., and Zhao, L. (2020). Beneficial Effects of Low-Dose Lithium on Cognitive Ability and Pathological Alteration of Alzheimer’s Disease Transgenic Mice Model. NeuroReport 31, 943–951. doi:10.1097/WNR.0000000000001499
Liu, X., and Yao, Z. (2016). Chronic Over-nutrition and Dysregulation of GSK3 in Diseases. Nutr. Metab. (Lond) 13, 49. doi:10.1186/s12986-016-0108-8
Madhavadas, S., and Subramanian, S. (2017). Cognition Enhancing Effect of the Aqueous Extract of Cinnamomum Zeylanicum on Non-transgenic Alzheimer’s Disease Rat Model: Biochemical, Histological, and Behavioural Studies. Nutr. Neurosci. 20, 526–537. doi:10.1080/1028415X.2016.1194593
Nguyen, T., Fan, T., George, S. R., and Perreault, M. L. (2018). Disparate Effects of Lithium and a GSK-3 Inhibitor on Neuronal Oscillatory Activity in Prefrontal Cortex and Hippocampus. Front. Aging Neurosci. 9, 434. doi:10.3389/fnagi.2017.00434
Ouyang, L., Luo, Y., Tian, M., Zhang, S.-Y., Lu, R., Wang, J.-H., et al. (2014). Plant Natural Products: from Traditional Compounds to New Emerging Drugs in Cancer Therapy. Cell Prolif. 47, 506–515. doi:10.1111/cpr.12143
Park, J.-H., Lee, S.-B., Lee, K.-H., and Ahn, J.-Y. (2012). Nuclear Akt Promotes Neurite Outgrowth in the Early Stage of Neuritogenesis. BMB Rep. 45, 521–525. doi:10.5483/BMBRep.2012.45.9.114
Parra, M. A., Abrahams, S., Fabi, K., Logie, R., Luzzi, S., and Sala, S. D. (2009). Short-term Memory Binding Deficits in Alzheimer's Disease. Brain 132, 1057–1066. doi:10.1093/brain/awp036
Ren, W.-m., Weng, Z.-b., Li, X., and Zhan, L.-b. (2021). Neuroprotective Effects of ZiBuPiYin Recipe on Db/db Mice via PI3K-Akt Signaling Pathway by Activating Grb2. Neural Plast. 2021, 1–10. doi:10.1155/2021/8825698
Saleh, R. A., Eissa, T. F., Abdallah, D. M., Saad, M. A., and El-Abhar, H. S. (2021). Peganum Harmala Enhanced GLP-1 and Restored Insulin Signaling to Alleviate AlCl3-Induced Alzheimer-like Pathology Model. Sci. Rep. 11, 12040. doi:10.1038/s41598-021-90545-4
Sayas, C. L., and Ávila, J. (2021). GSK-3 and Tau: A Key Duet in Alzheimer’s Disease. Cells 10, 721. doi:10.3390/cells10040721
Shinto, L., Quinn, J., Montine, T., Dodge, H. H., Woodward, W., Baldauf-Wagner, S., et al. (2013). A Randomized Placebo-Controlled Pilot Trial of Omega-3 Fatty Acids and Alpha Lipoic Acid in Alzheimer’s Disease. J. Alzheimers Dis. 38, 111–120. doi:10.3233/JAD-130722
Shu, Y., Zhang, H., Kang, T., Zhang, J.-j., Yang, Y., Liu, H., et al. (2013). PI3K/Akt Signal Pathway Involved in the Cognitive Impairment Caused by Chronic Cerebral Hypoperfusion in Rats. PLoS ONE 8, e81901. doi:10.1371/journal.pone.0081901
Small, G. W., Siddarth, P., Li, Z., Miller, K. J., Ercoli, L., Emerson, N. D., et al. (2018). Memory and Brain Amyloid and Tau Effects of a Bioavailable Form of Curcumin in Non-Demented Adults: A Double-Blind, Placebo-Controlled 18-Month Trial. Am. J. Geriatr. Psychiatry 26, 266–277. doi:10.1016/j.jagp.2017.10.010
Souder, D. C., and Anderson, R. M. (2019). An Expanding GSK3 Network: Implications for Aging Research. GeroScience 41, 369–382. doi:10.1007/s11357-019-00085-z
SoukhakLari, R., Moezi, L., Pirsalami, F., Ashjazadeh, N., and Moosavi, M. (2018). Curcumin Ameliorates Scopolamine-Induced Mice Memory Retrieval Deficit and Restores Hippocampal P-Akt and P-GSK-3β. Eur. J. Pharmacol. 841, 28–32. doi:10.1016/j.ejphar.2018.10.012
Sun, Y., Wu, A., Li, X., Qin, D., Jin, B., Liu, J., et al. (2020). The Seed of Litchi Chinensis Fraction Ameliorates Hippocampal Neuronal Injury in an Aβ25-35-Induced Alzheimer’s Disease Rat Model via the AKT/GSK-3β Pathway. Pharm. Biol. 58, 35–43. doi:10.1080/13880209.2019.1697298
Takada, Y., Hashimoto, M., Kasahara, J., Aihara, K., and Fukunaga, K. (2004). Cytoprotective Effect of Sodium Orthovanadate on Ischemia/Reperfusion-Induced Injury in the Rat Heart Involves Akt Activation and Inhibition of Fodrin Breakdown and Apoptosis. J. Pharmacol. Exp. Ther. 311, 1249–1255. doi:10.1124/jpet.104.070839
Takashima, A. (2012). GSK-3β and Memory Formation. Front. Mol. Neurosci. 5, 47. doi:10.3389/fnmol.2012.00047
Tang, Y., Yu, C., Wu, J., Chen, H., Zeng, Y., Wang, X., et al. (2018). Lychee Seed Extract Protects against Neuronal Injury and Improves Cognitive Function in Rats with Type II Diabetes Mellitus with Cognitive Impairment. Int. J. Mol. Med. 41, 251–263. doi:10.3892/ijmm.2017.3245
Tangestani Fard, M., and Stough, C. (2019). A Review and Hypothesized Model of the Mechanisms that Underpin the Relationship between Inflammation and Cognition in the Elderly. Front. Aging Neurosci. 11, 56. doi:10.3389/fnagi.2019.00056
Thapak, P., Khare, P., Bishnoi, M., and Sharma, S. S. (2020). Neuroprotective Effect of 2-Aminoethoxydiphenyl Borate (2-APB) in Amyloid β-Induced Memory Dysfunction: A Mechanistic Study. Cell. Mol. Neurobiol. 42, 1211–1223. doi:10.1007/s10571-020-01012-z
Tsalamandris, S., Antonopoulos, A. S., Oikonomou, E., Papamikroulis, G.-A., Vogiatzi, G., Papaioannou, S., et al. (2019). The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 14, 50–59. doi:10.15420/ecr.2018.33.1
van der Naald, M., Wenker, S., Doevendans, P. A., Wever, K. E., and Chamuleau, S. A. J. (2020). Publication Rate in Preclinical Research: a Plea for Preregistration. BMJ Open Sci. 4, e100051. doi:10.1136/bmjos-2019-100051
Wang, D., Wang, C., Liu, L., and Li, S. (2018). Protective Effects of Evodiamine in Experimental Paradigm of Alzheimer’s Disease. Cogn. Neurodyn. 12, 303–313. doi:10.1007/s11571-017-9471-z
Wang, X., Zhang, B., Li, X., Liu, X., Wang, S., Xie, Y., et al. (2021). Mechanisms Underlying Gastrodin Alleviating Vincristine-Induced Peripheral Neuropathic Pain. Front. Pharmacol. 12, 744663. doi:10.3389/fphar.2021.744663
Xiang, Q., Zhang, J., Li, C.-Y., Wang, Y., Zeng, M.-J., Cai, Z.-X., et al. (2015). Insulin Resistance-Induced Hyperglycemia Decreased the Activation of Akt/CREB in hippocampus Neurons: Molecular Evidence for Mechanism of Diabetes-Induced Cognitive Dysfunction. Neuropeptides 54, 9–15. doi:10.1016/j.npep.2015.08.009
Xiong, R., Wang, X.-L., Wu, J.-M., Tang, Y., Qiu, W.-Q., Shen, X., et al. (2020). Polyphenols Isolated from Lychee Seed Inhibit Alzheimer’s Disease-Associated Tau through Improving Insulin Resistance via the IRS-1/PI3K/Akt/GSK-3β Pathway. J. Ethnopharmacol. 251, 112548. doi:10.1016/j.jep.2020.112548
Xu, A.-H., Yang, Y., Sun, Y.-X., and Zhang, C.-D. (2018). Exogenous Brain-Derived Neurotrophic Factor Attenuates Cognitive Impairment Induced by Okadaic Acid in a Rat Model of Alzheimer’s Disease. Neural Regen. Res. 13, 2173–2181. doi:10.4103/1673-5374.241471
Yan, T., He, B., Wan, S., Xu, M., Yang, H., Xiao, F., et al. (2017). Antidepressant-like Effects and Cognitive Enhancement of Schisandra Chinensis in Chronic Unpredictable Mild Stress Mice and its Related Mechanism. Sci. Rep. 7, 6903. doi:10.1038/s41598-017-07407-1
Yan, T., Liu, B., Wang, N., Liao, Z., Wu, B., He, B., et al. (2021). The Flavonoids of Okra Insulates against Oxidative Stress, Neuroinflammation and Restores BDNF Levels in Aβ1–42 Induced Mouse Model of Alzheimer’s Disease. Exp. Gerontol. 147, 111263. doi:10.1016/j.exger.2021.111263
Yang, H. J., Hwang, J. T., Kwon, D. Y., Kim, M. J., Kang, S., Moon, N. R., et al. (2013). Yuzu Extract Prevents Cognitive Decline and Impaired Glucose Homeostasis in β-Amyloid–Infused Rats. J. Nutr. 143, 1093–1099. doi:10.3945/jn.112.173401
Yao, Y., Wang, Y., Kong, L., Chen, Y., and Yang, J. (2019). Osthole Decreases Tau Protein Phosphorylation via PI3K/AKT/GSK-3β Signaling Pathway in Alzheimer’s Disease. Life Sci. 217, 16–24. doi:10.1016/j.lfs.2018.11.038
Zhang, Y.-H., Wang, D.-W., Xu, S.-F., Zhang, S., Fan, Y.-G., Yang, Y.-Y., et al. (2018). α-Lipoic Acid Improves Abnormal Behavior by Mitigation of Oxidative Stress, Inflammation, Ferroptosis, and Tauopathy in P301S Tau Transgenic Mice. Redox Biol. 14, 535–548. doi:10.1016/j.redox.2017.11.001
Zhou, W., Lian, W.-w., Yan, R., Jia, H., Xu, L.-j., Wang, L., et al. (2020). DL0410 Ameliorates Cognitive Deficits in APP/PS1 Transgenic Mice by Promoting Synaptic Transmission and Reducing Neuronal Loss. Acta Pharmacol. Sin. 41, 599–611. doi:10.1038/s41401-019-0312-y
Zhu, H., Yan, H., Tang, N., Li, X., Pang, P., Li, H., et al. (2017). Impairments of Spatial Memory in an Alzheimer’s Disease Model via Degeneration of Hippocampal Cholinergic Synapses. Nat. Commun. 8, 1676. doi:10.1038/s41467-017-01943-0
Zhu, L., Zhang, Z., Hou, X.-J., Wang, Y.-F., Yang, J.-Y., and Wu, C.-F. (2019). Inhibition of PDE5 Attenuates Streptozotocin-Induced Neuroinflammation and Tau Hyperphosphorylation in a Streptozotocin-Treated Rat Model. Brain Res. 1722, 146344. doi:10.1016/j.brainres.2019.146344
Keywords: ageing, aged, Alzheiemer’s disease, dementia, cognition, glycogen syntase kinase 3, insulin resisitance
Citation: Matysek A, Kimmantudawage SP, Feng L and Maier AB (2022) Targeting Impaired Nutrient Sensing via the Glycogen Synthase Kinase-3 Pathway With Therapeutic Compounds to Prevent or Treat Dementia: A Systematic Review. Front. Aging 3:898853. doi: 10.3389/fragi.2022.898853
Received: 17 March 2022; Accepted: 27 May 2022;
Published: 18 July 2022.
Edited by:
Morten Scheibye-Knudsen, University of Copenhagen, DenmarkReviewed by:
Devin Wahl, Colorado State University, United StatesSmijin K. Soman, University of Nevada, United States
Copyright © 2022 Matysek, Kimmantudawage, Feng and Maier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea B. Maier, YS5iLm1haWVyQHZ1Lm5s