 Xurui Jin
Xurui Jin Tingxi Long1†
Tingxi Long1† Yi Zeng
Yi Zeng Xian Zhang
Xian Zhang Lijing Yan
Lijing Yan Chenkai Wu
Chenkai Wu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Aging Neurosci. , 25 February 2022
Sec. Neurocognitive Aging and Behavior
Volume 13 - 2021 | https://doi.org/10.3389/fnagi.2021.710966
This article is part of the Research Topic Genes and Aging: From Bench Side to Bed side View all 10 articles
Recent literature suggested that ALDH2 mutation is associated with alcohol metabolism, and ethanol intake might jointly increase the risk of Alzheimer’s disease (AD) in mice. However, it is unclear whether this synergistic effect exists among humans. We examined the associations of four single nucleotide polymorphisms (SNPs) on aldehyde dehydrogenase (ALDH) and alcohol dehydrogenase (ADH) genes (i.e., ALDH2 rs671, ADH1B rs1229984, ADH1B rs1042026, and ADH1C rs1693482) and cognitive impairment among the oldest-old. We also investigated whether this association was modified by ethanol intake from alcohol consumption. Data were from the Chinese Longitudinal Healthy Longevity Survey genetic sub-study, including 1,949 participants aged over 90 years. Participants with a Mini-Mental State Examination (MMSE) score of < 18 were considered cognitively impaired. Alcohol consumption was categorized as heavy, moderate, or never drinkers. With the dominant model, carrying A allele on rs671, C allele on rs1229984, and T allele on rs1042026 was associated with 33% (95% confidence interval [CI]: 5%, 69%), 33% (95% CI: 2%, 75%), and 29% (95% CI: 3%, 62%) higher odds of cognitive impairment in the multivariable-adjusted logistic model, respectively. We did not observe a significant interaction between those SNPs and alcohol consumption. Among the oldest-old, carrying ALDH2 rs671 mutation was associated with higher odds of cognitive impairment independent of alcohol consumption.
Aging is a major contributor to cognitive decline and dementia across the world, and it is becoming a major public health concern. Previous studies have discussed demographic or socioeconomic risk factors for dementia among old adults, with limited attention on genetic risk factors using large samples. Most of the genetic research on cognitive function focused on candidate genes which have been demonstrated to be associated with Alzheimer’s disease (AD) or identified by genome-wide association studies (GWAS). Those candidate genes include apolipoprotein E (APOE), catechol-O-methyltransferase (COMT) (Komulainen et al., 2008), brain-derived neurotrophic factor (BDNF) (Bray et al., 2005), and dystrobrevin-binding protein 1 (DTNBP1) (Wray et al., 2008), with the APOE ε4 allele being by far the strongest genetic risk factor and accounts for about 5% of the variance in lifetime cognitive change and 4% of the variance in AD. In addition to those genetic factors, some recent works have also indicated the key role of some alcohol metabolism-related genes as enzymes involved in the detoxification of the ethanol metabolism in the pathology of AD.
Alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) are the two major alcohol-metabolizing enzymes (Chen et al., 2014). The mutation of several single nucleotide polymorphisms (SNPs) on those two genes results in the change of enzymatic activity, such as rs671 in ALDH, rs1693482 in ADH1C, and rs1229984 and rs1042026 in ADH1B (Hurley and Edenberg, 2012). Among the Asian population, the prevalence of ADH1B, ADH1C, and ALDH2 mutation is performed. It is estimated that nearly 30% of people in Asia (∼8% of the world population) carry the genetic variants of the ALDH2 A allele by rs671 (Eng et al., 2007). Previous literature has indicated that a reduced activity of ADH and ALDH would lead to an excess of acetaldehyde and result in oxidative stress and mitochondrial dysfunctions which have been identified in both familial and sporadic AD (Zhang and Ren, 2011; Chen et al., 2012; Swerdlow, 2018). At the population level, ADH and ALDH genes have been shown to be associated with alcohol dependence (Macgregor et al., 2009)—a leading cause of dementia (Koch et al., 2019). At present, the associations between ALDH2 and ADH genetic polymorphisms with cognitive function or AD were inconclusive. Some studies suggested that ALDH2 (rs671 polymorphism) is a risk factor for AD in Japanese (Kamino et al., 2000), whereas others reported no association. A recent study including 339 AD patients and 168 healthy controls investigated the association of several SNPs on ADH and ALDH with AD and found a suggestive association between ADH1C rs2241894 and AD among women (Wu et al., 2021). However, some other genetic studies have found that the negative impact of genetic factors may be cumulative by age, such as APOE gene on cognitive function, and there is a fewer study of ADH and ALDH genes among the oldest-old population.
A recent experimental study suggested that chronic excessive ethanol intake and the ALDH2 gene mutation might jointly increase the risk of AD in mice (Joshi et al., 2019). Alcohol consumption, as one of the major risk factors of brain damage, is associated with the incidence of dementia. There is a J-shaped association of alcohol consumption with dementia, where excessive alcohol intake or abstinence increased dementia risk, compared with consuming 9–112 g/week (Topiwala and Ebmeier, 2018). In a large cross-sectional study from Southern China, occasional rather than moderate alcohol use was found to be associated with better cognitive function (Au Yeung et al., 2011), suggesting that the observed effects could be driven by a complex interaction between alcohol and other factors. In systematic reviews, drinking patterns are associated with AD and also cognitive function. It is unclear whether ALDH2 mutation is associated with cognitive function and whether this mutation and ethanol intake synergistically contribute to the development of AD among humans.
The aim of the present study was twofold. First, we investigated the associations of four SNPs (i.e., ALDH2 rs671, ADH1B rs1229984, ADH1B rs1042026, and ADH1C rs1693482) and cognitive function among nearly 2,000 Chinese oldest-old from a population-based cohort study. Considering the high prevalence of ADH and ALDH gene mutation in the Chinese population and their unique characterization, we then investigated whether this association would be modified by the level of alcohol consumption.
The present study used data from the CLHLS, which is an ongoing longitudinal study that began in 1998 with follow-up surveys for every 2–3 years. The CLHLS is a Chinese nationwide survey conducted in randomly selected counties and cities in 22 of 30 provinces covering 85% of the population of China. All centenarians from the selected areas who agreed to participate were included in the study. Based on sex and place of residence (i.e., living in the same street, village, city, or county) for a given centenarian, randomly selected octogenarians and non-agenarians were also sampled. More details about the sampling procedure and quality of data of this survey have been published elsewhere (Zeng et al., 2017). Ethical approval was obtained from the Research Ethics Committees of Peking University and Duke University (IRB00001052-13074). All participants or their legal representatives signed written consent forms in the baseline and follow-up surveys. In this cross-sectional study, we derived the data from the baseline survey of each participant. The analyses were based on the CLHLS genetic dataset, comprising 1,949 adults aged over 90 years.
The cognitive function of CLHLS participants was assessed by the Chinese version of the Mini-Mental State Examination (MMSE) through a home-based interview, which includes 24 items, covering 7 subscales including orientation (4 points for time orientation and 1 point for place orientation); naming foods (naming as many kinds of food as possible in 1 min, 7 points); registration of 3 words (3 points); attention and calculation (mentally subtracting 3 iteratively from 20, 5 points); copy a figure (1 point); recall (delayed recall of the 3 words mentioned above, 3 points); and language (2 points for naming objectives, 1 point for repeating a sentence, and 3 points for listening and following directions). The MMSE score ranges from 0 to 30. Higher scores represent a better cognitive function. The validity and reliability of this Chinese MMSE have been verified in several previous studies (Zhang, 2006; An and Liu, 2016). Consistent with previous studies (Zhang et al., 2019), because a high proportion of our participants did not have formal education (∼70%), cognitive impairment was defined as an MMSE score of < 18.
During each interview, the interviewers measured a range of demographic, behavioral, and socioeconomic covariates. Alcohol consumption was self-reported and categorized as heavy, moderate, or never drinkers. Current alcohol users who consumed >25 g of alcohol per day (for men) and > 15 g (for women) were considered heavy drinkers. Current alcohol users who consumed ≤25 g (for men) and ≤15 g (for women) were considered moderate drinkers. Participants who reported they never drank were considered never drinkers. All variables were measured at baseline (indicate year). All self-reported information was collected through face-to-face home interview by trained research staff members with more than 12 years of education.
Following the previous studies, we included the following variables as confounders: age (years), sex, residence (rural vs. urban), education years, smoking (never, former, and current), regular physical activity (yes vs. no), dietary pattern, leisure activity score, and self-reported chronic diseases [e.g., hypertension, diabetes, heart disease, stroke, and chronic obstructive pulmonary disease (COPD)]. Dietary pattern was categorized as unfavorable, intermediate, or favorable by a simplified healthy eating index based on the intake frequency of nine food categories, including fruits, vegetables, fish, bean products, tea, garlic, eggs, sugar, and salt-preserved vegetables. A 3-point scale question was used to measure the current intake frequency of each food group: “always or almost every day,” “sometimes or occasionally,” or “rarely or never.” Those 3 terms received the scores of 2, 1, or 0, respectively, with higher scores indicating a higher level of consumption. Two of those food groups, i.e., sugar and salt-preserved vegetables, were received the scores of 0, 1, and 2, respectively, for some evidence of the negative impact of the high consumption of those two food groups. The leisure activity score was summarized from the following eight activities: visiting neighbors, shopping, cooking, washing clothes, walking 1 km, lifting 5 kg, crouching and standing up three times, and taking public transportation. We scored each activity as 1 for “never,” 2 for “sometimes,” and 3 for “almost every day.” The score ranged from 5 to 21 with a higher score, indicating more leisure activities. All self-reported information was collected through face-to-face home interview by trained research staff members at the baseline survey. Interviewees were encouraged to answer as many questions as possible. If they were unable to answer questions, a close family member or another proxy, such as a primary caregiver, provided answers (Zeng, 2012).
The CLHLS collected DNA samples from parts of participants in 1998, 2000, 2002, 2005, 2008–2009, and 2011–2012 waves of the survey. Genotyping of DNA samples was produced by the Beijing Genomics Institute (BGI), and the BGI genotyping quality control procedures of the CLHLS genetic study have been published elsewhere (Zeng et al., 2016). We extracted four SNPs associated with alcohol metabolism from the GWAS data (i.e., ALDH2 rs671, ADH1B rs1229984, ADH1B rs1042026, and ADH1C rs1693482).
The genotypes were defined by following the additive and dominant models. In a dominant model, any genotype that contains one or two copies of the minor allele is coded as one; otherwise, the genotype that does not contain any copy of the minor allele is coded as zero. In an additive model, carrying two copies of the minor allele was coded as three. One copy of the minor allele with one copy of the major allele and two major alleles were coded as two and one, respectively. We mainly used the dominant models in the GxE analysis to define the genotype because it can distinguish the genotype carriers and non-carriers, but the additive model cannot. In addition, further grouping the samples in the additive model would result in many more GxE interaction terms in the regressions and would, in turn, negatively affect the estimates and complicate the discussions.
The characteristics of participants were compared according to cognitive function (with vs. without impairment). Means and standard deviations (SDs) were calculated for continuous variables; counts and percentages were calculated for categorical variables. We used both logistic regression (binary outcome: cognitive impairment) and linear regression (continuous outcome: MMSE score) models to examine the unadjusted and adjusted associations between the four polymorphisms and cognitive impairment, respectively. We build two regression models, namely, partly adjusted: adjusted for age at baseline in years, sex, residency, and education years; and fully adjusted: additionally adjusted for smoking status, alcohol consumption, current physical activity, dietary pattern and leisure activity score, and five kinds of self-reported diseases on the basement of the partly adjusted model.
To examine whether the association between the four polymorphisms and cognitive impairment was modified by alcohol consumption, we first ran a logistic regression model with an interaction term between those four polymorphisms and alcohol consumption, respectively, and then conducted the regression analysis among persons with different levels of alcohol consumption (e.g., heavy, moderate, and never drinkers), separately.
All statistical analyses were conducted using STATA version 14.0 (Stata Corp., College Station, TX, United States).
Demographic and the four polymorphisms information are detailed by cognitive function in Table 1. A total of 1,949 participants were included after excluding 101 participants who lacked cognitive assessment (n = 64) and aged below 90 years (n = 37). Of 1,949 participants aged over 90 years, the average age was 101.3 ± 3.3 years; 76.3% were women. Participants with cognitive impairment are more likely to be older, women, with lower education level, not smoking, with unfavorable dietary pattern, and with lower leisure activities (Table 1, p-values < 0.05).
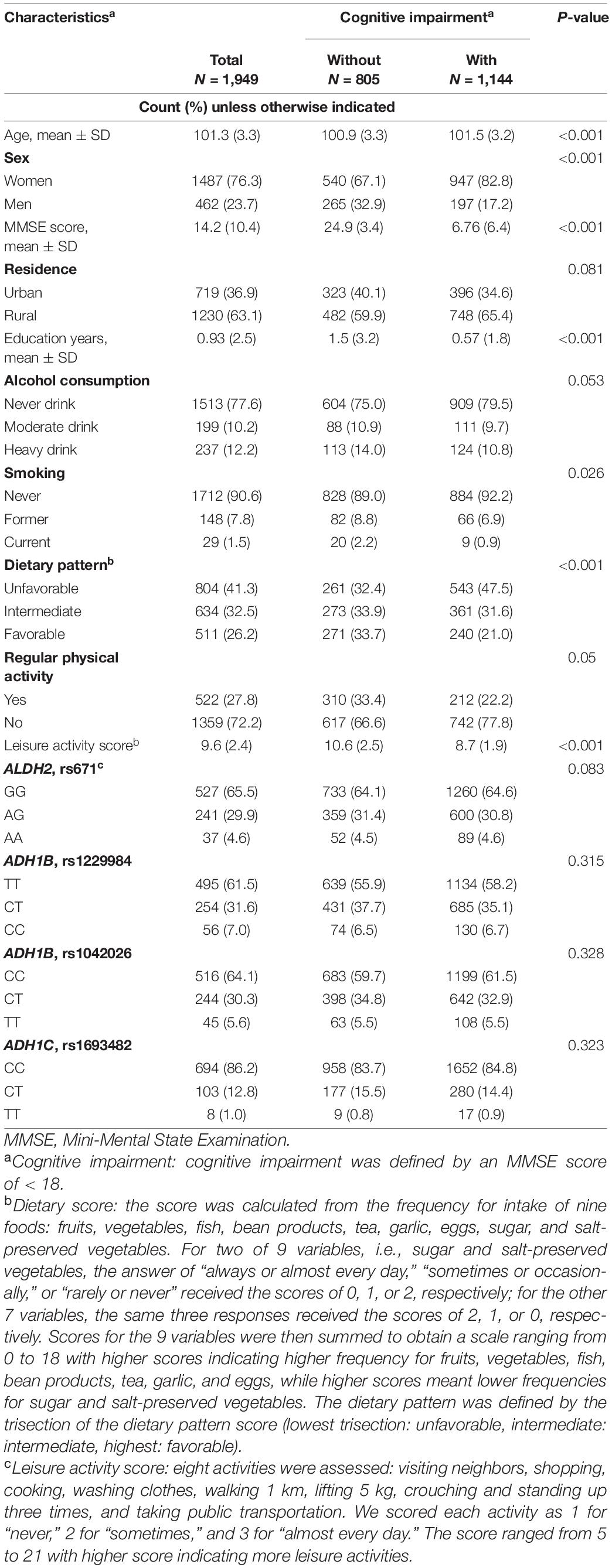
Table 1. Selected characteristics of the participants by cognitive function.
In the partially adjusted logistic regression model, carrying A allele on rs671, C allele on rs1229984, T allele on rs1042026, and T allele on rs1693482 was associated with higher odds of cognitive impairment [rs671: odds ratio (OR): 1.19, 95% CI: 0.97, 1.47; rs1229984: OR: 1.27, 95% CI: 1.00, 1.61; rs1042026: OR: 1.26, 95% CI: 1.04, 1.55; rs1693482: OR: 1.19, 95% CI: 0.91, 1.57; Table 2]. In the fully adjusted logistic regression model, the associations of rs671, rs1229984, and rs1042026 with cognitive impairment persisted to be significant. In the partially adjusted linear regression model, carrying T allele on rs1042026 and T allele on rs1693482 was significantly associated with a lower MMSE score of 0.90 (95% CI: 0.01, 1.80). In the fully adjusted linear regression model, carrying A on rs671 was associated with a lower MMSE score of 1.05 (95% CI: −2.03, −0.09).
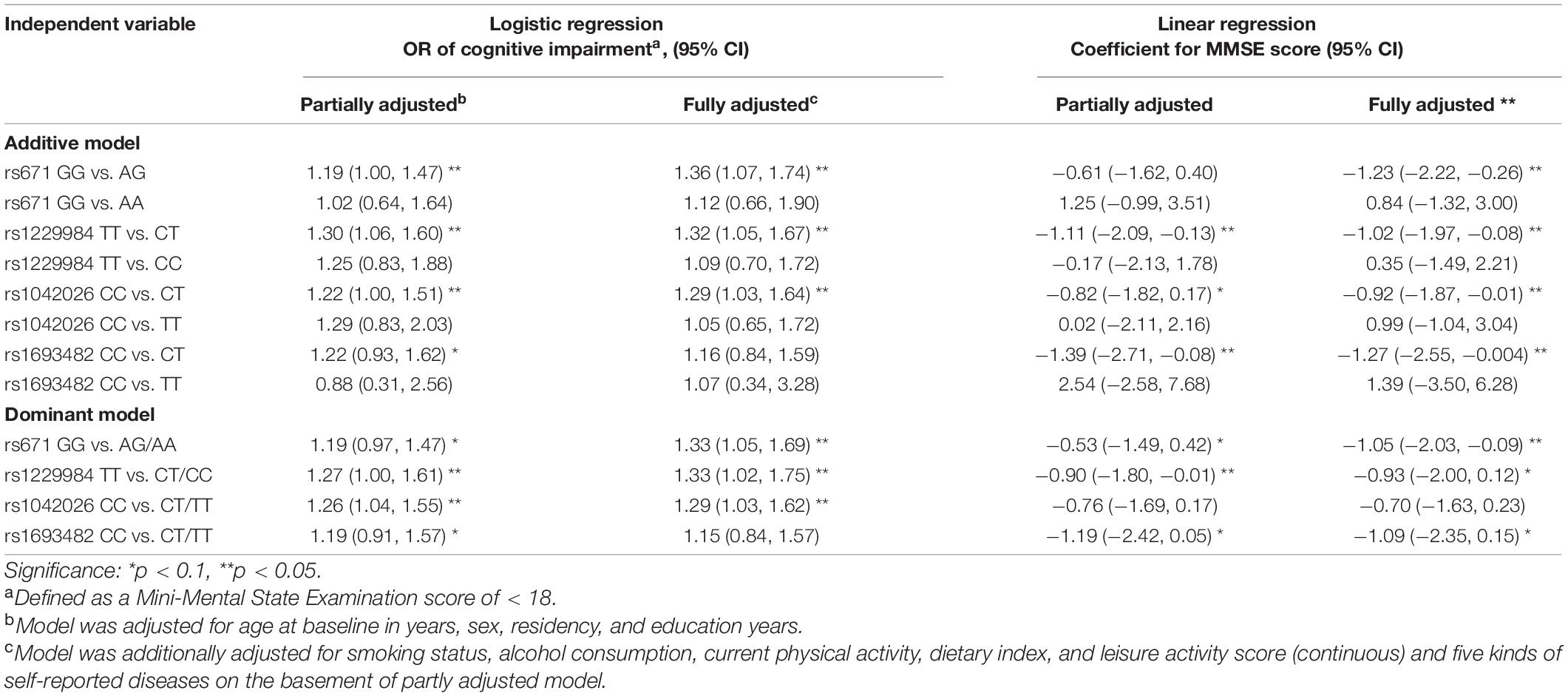
Table 2. Association of ALDH2 rs671, ADH1B rs1229984, ADH1B rs1042026, and ADH1C rs1693482 polymorphisms with cognitive impairment.a
In the additive model, the one major allele with one minor allele genotypes of rs671, rs1229984, and rs1042026 was significantly associated with lower MMSE scores and higher odds of cognitive impairment compared with the two major allele genotypes (Table 2). However, the two minor allele genotypes of those four SNPs were not significantly associated with MMSE score or cognitive impairment in the fully or partially adjusted model.
The association between alcohol consumption and cognitive impairment was analyzed in the partially adjusted model and fully adjusted model (Supplementary Table 1). In the fully adjusted linear regression model, moderate alcohol use and heavy alcohol use were significantly associated with a lower MMSE score (moderate: −2.52, 95% CI: −2.49, −2.55; heavy: −2.83, 95% CI: −2.36, −3.30) compared with participants who never consumed alcohol.
Table 3 presents the associations between four SNPs (those specified here) with the dominant model and cognitive impairment stratified by alcohol consumption. In the subgroup analyses, there is no evidence that the association of each of four SNPs with cognitive impairment differed by alcohol consumption (p-values for interactions > 0.05).
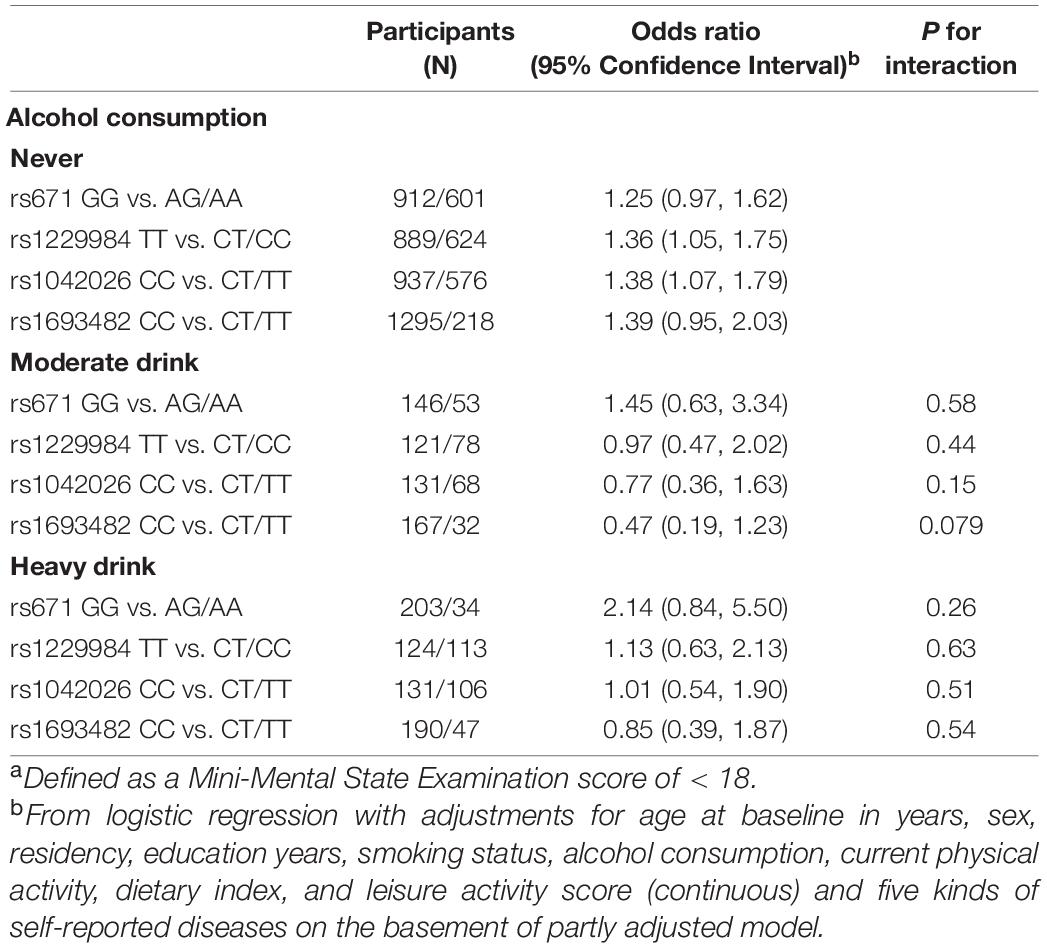
Table 3. The association of ALDH2 rs671 polymorphism with cognitive impairment stratified by alcohol consumption and fresh fruit consumption.a
Using data from nearly 2,000 Chinese adults aged over 90 years, we found that the SNPs on ADH and ALDH2 genes were associated with higher odds of cognitive impairment. In addition, the results showed that the associations between those polymorphisms and cognitive impairments were not modified by alcohol consumption, which is different from the findings of a recent experimental study (Joshi et al., 2019).
The results indicated that carrying the ALDH2 and ADH mutation was associated with higher odds of cognitive impairment among the oldest-old population. Its mutation results in a reduction of the ALDH2 enzymatic activity, which is widely mutated in the Asian population (∼30%). One possible explanation was that those two mutations have a negative impact on mitochondrial functions and the metabolism of 4-hydroxy-2-nonenal (4-NHE). ALDH2 and ADH are involved in the detoxification of the ethanol metabolism and other aldehydes, including 4-HNE. It has been shown that 4HNE concentration increased in the brain tissue of the ALDH2*2 transgenic mice in an age-dependent manner. Such increases correlated with neurodegeneration memory loss and AD-like pathological changes in these ALDH2*2 transgenic mice (Ohsawa et al., 2008). In addition, 4HNE levels are higher in the hippocampus of postmortem samples from patients with AD (Williams et al., 2006). Among the population level, previous studies reported the inconsistent results on this association. A cross-sectional case-control study has shown that ALDH2 genotype is associated with cognitive function among 139 Chinese patients with Parkinson’s disease (mean age: 63.0 years) (Yu et al., 2016), and another study has demonstrated its association among 411 Chinese with an average age of 77.4 years (Wang et al., 2008). However, in another study including 690 Korean community residents (mean age: 72.8 years) (Kim et al., 2004), the association between ALDH2 genotype and cognitive functions was not significant. This paradox finding may be due to the mean age difference of study samples. It is plausible that the genetic risk of carrying the ALDH2 mutation is cumulative (Licher et al., 2019). Researchers found that APOE as well as other common genetic variants could have a cumulative risk on the progression of dementia and AD as age advances (van der Lee et al., 2018). To be more specific, with the negative influence of those mutations on neuronal functioning, the carriers may have a higher speed of neuronal cell loss which is irreversible. Accordingly, during the early life stage, the difference in cognitive function between the carriers and non-carriers may not be significant, while as people age, the harmful effects of genetic variants may build up, and the carriers may have a lower average cognitive function compared with non-carriers of the same age due to the cumulative damage on neurons (Bai and Mei, 2011). In addition to rs671 on ALDH2, we also found that rs1229984 and rs1042026 on ADH1B gene were associated with the cognitive function among the oldest-old population. The ADH1B rs1229984 T allele encodes a super active enzyme subunit that could accelerate the conversion from ethanol into acetaldehyde (Lin et al., 2021). ADH1C rs1693482 is found to be associated with a twofold difference in ADH Vmax in vitro (Birley et al., 2009). One experimental study has found that in the animal model of AD, the overexpressing of ADH1B suppresses the β-amyloid-induced neuron apoptosis (Wang et al., 2019). Corresponding the results, it may indicate that the genes associated with alcohol metabolism might be important in the progress of cognitive impairment or AD. To note, these results also determined the potential role of mitochondrial dysfunctions in the pathology of AD.
We found that those associations were not modified by alcohol consumption. One experimental study found that the ALDH2*2 mutation increases the damage by sustained ethanol exposure in mouse brain, and they found that ALDH2*2 deficiency increases ethanol-induced neuroinflammation in vivo (Joshi et al., 2019). However, we found that on the population level, such interactive effect was not significant. It may indicate that the damage of alcohol to the brain is stable and consistent which may not be varied by genotype. In addition, another possible explanation was that the consumption of stored fruits and vegetables, as well as fruit juice, would bring ethanol into the human body and the total ethanol intake is more difficult to measure.
Methodological strengths of this study include a large sample size of the oldest old, and we have included multiple genes related to alcohol metabolism. This study also has several limitations: (1) it has a cross-sectional design and cannot evaluate changes in alcohol consumption or establish causality. However, the results were robust to adjustment of a number of indicators such as diseases. Nevertheless, prospective studies on the incidence of cognitive impairment are warranted; (2) the alcohol consumption was self-reported, thus non-specific (Muntner et al., 2014). This possibility of recall bias is a common concern in the longitudinal cohort studies, although CLHLS data have been validated as being reliable in previous studies (Zeng et al., 2008). In the CLHLS study, all self-reported information was collected through face-to-face home interview by trained research staff members. Interviewees were encouraged to answer as many questions as possible. If they were unable to answer questions, a close family member or another proxy, such as a primary caregiver, provided answers (Zeng et al., 2008). (3) We used the Chinese version of MMSE to measure cognition, which is not a clinical diagnosis for cognitive impairment (Zhang, 1993; Chou, 2003). However, it is a validated instrument in population-based studies. (4) As the study included only participants who aged over 90 years and the average lifespan of women is longer than men, a higher proportion of women were taken into the analysis sample which may lead to bias. (5) The percentage of drinker is 20% and the results need to be validated within a larger cohort.
We found that some SNPs associated with alcohol metabolism were associated with higher odds of cognitive impairment among the Chinese oldest-old and those associations were independent of alcohol consumption. Due to its mutation can be easily observed even without genotyping, it may have more public health implications especially for the Asian population on the AD risk stratification and alcohol control. Further studies on how to modify the genetic risk for cognitive impairment associated with gene mutation associated with alcohol metabolism are warranted.
Publicly available datasets were analyzed in this study. This data can be found here: https://sites.duke.edu/centerforaging/programs/chinese-longitudinal-healthy-longevity-survey-clhls.
The studies involving human participants were reviewed and approved by the Biomedical Ethics Review Committee of Peking University (IRB00001052-11015). The patients/participants provided their written informed consent to participate in this study.
All authors listed have made a substantial, direct, and intellectual contribution to the work, and approved it for publication.
This study was supported by the National Natural Science Foundation of China (71110107025, 71233001, and 71490732) to YZ, the National Institute on Aging/National Institutes of Health (R01 AG023627) to YZ, and 2020YFC2005600 from the Chinese Ministry of Science and Technology to CW.
XJ and XZ were employed by MindRank AI Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2021.710966/full#supplementary-material
An, R., and Liu, G. G. (2016). Cognitive impairment and mortality among the oldest-old Chinese. Int. J. Geriatr. Psychiatry 31, 1345–1353. doi: 10.1002/gps.4442
Au Yeung, S. L., Leung, G. M., Chan, W. M., Hui, Y. F., Lam, T. H., Schooling, C. M., et al. (2011). Moderate alcohol use and cognitive function in an elderly Chinese cohort. J. Am. Geriatr. Soc. 59, 172–174. doi: 10.1111/j.1532-5415.2010.03195.x
Bai, J., and Mei, Y. (2011). Overexpression of aldehyde dehydrogenase-2 attenuates neurotoxicity induced by 4-hydroxynonenal in cultured primary hippocampal neurons. Neurotox. Res. 19, 412–422. doi: 10.1007/s12640-010-9183-1
Birley, A. J., James, M. R., Dickson, P. A., Montgomery, G. W., Heath, A. C., Martin, N. G., et al. (2009). ADH single nucleotide polymorphism associations with alcohol metabolism in vivo. Hum. Mol. Genet. 18, 1533–1542. doi: 10.1093/hmg/ddp060
Bray, N. J., Preece, A., Williams, N. M., Moskvina, V., Buckland, P. R., Owen, M. J., et al. (2005). Haplotypes at the dystrobrevin binding protein 1 (DTNBP1) gene locus mediate risk for schizophrenia through reduced DTNBP1 expression. Hum. Mol. Genet. 14, 1947–1954. doi: 10.1093/hmg/ddi199
Chen, C. H., Ferreira, J. C., Gross, E. R., and Mochly-Rosen, D. (2014). Targeting aldehyde dehydrogenase 2: new therapeutic opportunities. Physiol. Rev. 94, 1–34. doi: 10.1152/physrev.00017.2013
Chen, X., Guo, C., and Kong, J. (2012). Oxidative stress in neurodegenerative diseases. Neural. Regen. Res. 7, 376–385. doi: 10.3969/j.issn.1673-5374.2012.05.009
Chou, K.-L. (2003). Correlates of everyday competence in Chinese older adults. Aging Ment. Health 7, 308–315.
Eng, M. Y., Luczak, S. E., and Wall, T. L. (2007). ALDH2, ADH1B, and ADH1C genotypes in Asians: a literature review. Alcohol Res. Health 30, 22–27.
Hurley, T. D., and Edenberg, H. J. (2012). Genes encoding enzymes involved in ethanol metabolism. Alcohol Res. 34, 339–344.
Joshi, A. U., Van Wassenhove, L. D., Logas, K. R., Minhas, P. S., Andreasson, K. I., Weinberg, K. I., et al. (2019). Aldehyde dehydrogenase 2 activity and aldehydic load contribute to neuroinflammation and Alzheimer’s disease related pathology. Acta Neuropathol. Commun. 7:190. doi: 10.1186/s40478-019-0839-7
Kamino, K., Nagasaka, K., Imagawa, M., Yamamoto, H., Yoneda, H., Ueki, A., et al. (2000). Deficiency in mitochondrial aldehyde dehydrogenase increases the risk for late-onset Alzheimer’s disease in the Japanese population. Biochem. Biophys. Res. Commun. 273, 192–196. doi: 10.1006/bbrc.2000.2923
Kim, J. M., Stewart, R., Shin, I. S., Jung, J. S., and Yoon, J. S. (2004). Assessment of association between mitochondrial aldehyde dehydrogenase polymorphism and Alzheimer’s disease in an older Korean population. Neurobiol. Aging 25, 295–301. doi: 10.1016/S0197-4580(03)00114-3
Koch, M., Fitzpatrick, A. L., Rapp, S. R., Nahin, R. L., Williamson, J. D., Lopez, O. L., et al. (2019). Alcohol Consumption and Risk of Dementia and Cognitive Decline Among Older Adults With or Without Mild Cognitive Impairment. JAMA Netw. Open 2:e1910319. doi: 10.1001/jamanetworkopen.2019.10319
Komulainen, P., Pedersen, M., Hänninen, T., Bruunsgaard, H., Lakka, T. A., Kivipelto, M., et al. (2008). BDNF is a novel marker of cognitive function in ageing women: the DR’s EXTRA Study. Neurobiol. Learn. Mem. 90, 596–603. doi: 10.1016/j.nlm.2008.07.014
Licher, S., Ahmad, S., Karamujic-Comic, H., Voortman, T., Leening, M. J. G., Ikram, M. A., et al. (2019). Genetic predisposition, modifiable-risk-factor profile and long-term dementia risk in the general population. Nat. Med. 25, 1364–1369. doi: 10.1038/s41591-019-0547-7
Lin, C. H., Nfor, O. N., Ho, C. C., Hsu, S. Y., Tantoh, D. M., Liaw, Y. C., et al. (2021). Association of ADH1B polymorphism and alcohol consumption with increased risk of intracerebral hemorrhagic stroke. J. Transl. Med. 19:227. doi: 10.1186/s12967-021-02904-4
Macgregor, S., Lind, P. A., Bucholz, K. K., Hansell, N. K., Madden, P. A., Richter, M. M., et al. (2009). Associations of ADH and ALDH2 gene variation with self report alcohol reactions, consumption and dependence: an integrated analysis. Hum. Mol. Genet. 18, 580–593. doi: 10.1093/hmg/ddn372
Muntner, P., Colantonio, L. D., Cushman, M., Colantonio, L. D., Cushman, M., and Goff, D. C. Jr., et al. (2014). Validation of the atherosclerotic cardiovascular disease Pooled Cohort risk equations. JAMA 311, 1406–1415. doi: 10.1001/jama.2014.2630
Ohsawa, I., Nishimaki, K., Murakami, Y., Suzuki, Y., Ishikawa, M., Ohta, S., et al. (2008). Age-dependent neurodegeneration accompanying memory loss in transgenic mice defective in mitochondrial aldehyde dehydrogenase 2 activity. J. Neurosci. 24, 6239–6249. doi: 10.1523/JNEUROSCI.4956-07.2008
Swerdlow, R. H. (2018). Mitochondria and Mitochondrial Cascades in Alzheimer’s Disease. J. Alzheimer’s Dis. 62, 1403–1416. doi: 10.3233/JAD-170585
Topiwala, A., and Ebmeier, K. P. (2018). Effects of drinking on late-life brain and cognition. Evid. Based Ment. Health 21, 12–15. doi: 10.1136/eb-2017-102820
van der Lee, S. J., Wolters, F. J., Ikram, M. K., Hofman, A., Ikram, M. A., Amin, N., et al. (2018). The effect of APOE and other common genetic variants on the onset of Alzheimer’s disease and dementia. Lancet Neurol. 17, 434–444. doi: 10.1016/S1474-4422(18)30053-X
Wang, B., Wang, J., Zhou, S., Tan, S., He, X., Yang, Z., et al. (2008). The association of mitochondrial aldehyde dehydrogenase gene (ALDH2) polymorphism with susceptibility to late-onset Alzheimer’s disease in Chinese. J. Neurol. Sci. 268, 172–175. doi: 10.1016/j.jns.2007.12.006
Wang, Y., Zhang, Y., Zhang, X., Yang, T., Liu, C., and Wang, P. (2019). Alcohol Dehydrogenase 1B Suppresses β-Amyloid-Induced Neuron Apoptosis. Front. Aging Neurosci. 11:135. doi: 10.3389/fnagi.2019.00135
Williams, T. I., Lynn, B. C., Markesbery, W. R., and Lovell, M. A. (2006). Increased levels of 4-hydroxynonenal and acrolein, neurotoxic markers of lipid peroxidation, in the brain in Mild Cognitive Impairment and early Alzheimer’s disease. Neurobiol. Aging 27, 1094–1099. doi: 10.1016/j.neurobiolaging.2005.06.004
Wray, N. R., James, M. R., Handoko, H. Y., Dumenil, T., Lind, P. A., Montgomery, G. W., et al. (2008). Association study of candidate variants from brain-derived neurotrophic factor and dystrobrevin-binding protein 1 with neuroticism, anxiety, and depression. Psychiatric Genet. 18, 219–225. doi: 10.1097/YPG.0b013e3283050aee
Wu, Y. Y., Lee, Y. S., Liu, Y. L., Hsu, W. C., Ho, W. M., Huang, Y. H., et al. (2021). Association Study of Alcohol Dehydrogenase and Aldehyde Dehydrogenase Polymorphism With Alzheimer Disease in the Taiwanese Population. Front. Neurosci. 15:625885. doi: 10.3389/fnins.2021.625885
Yu, R. L., Tan, C. H., Lu, Y. C., and Wu, R. M. (2016). Aldehyde dehydrogenase 2 is associated with cognitive functions in patients with Parkinson’s disease. Sci. Rep. 6:30424. doi: 10.1038/srep30424
Zeng, Y. (2012). Towards Deeper Research and Better Policy for Healthy Aging –Using the Unique Data of Chinese Longitudinal Healthy Longevity Survey. China Eco. J. 5, 131–149. doi: 10.1080/17538963.2013.764677
Zeng, Y., Feng, Q., Gu, D., and Vaupel, J. W. (2017). Demographics, phenotypic health characteristics and genetic analysis of centenarians in China. Mech. Ageing Dev. 165, 86–97. doi: 10.1016/j.mad.2016.12.010
Zeng, Y., Nie, C., Min, J., Liu, X., Li, M., Chen, H., et al. (2016). Novel loci and pathways significantly associated with longevity. Sci. Rep. 6:21243. doi: 10.1038/srep21243
Zeng, Y., Poston, D. L., Vlosky, D. A., and Gu, D. (2008). “An exploration of the subjective well-being of the Chinese oldest-old,” in Healthy Longevity in China: Demographic, Socioeconomic, and Psychological Dimensions, eds Y. Zeng and D. L. Poston Jr., D. A. Vlosky and D. Gu (Springer: Business Media), 347–356. doi: 10.1007/978-1-4020-6752-5_21
Zhang, M. Y. (1993). Manual of Evaluation of Scales in Psychiatrics. Changsha: Hunan Science and Technology Press, 38–41.
Zhang, Q., Wu, Y., Han, T., and Liu, E. (2019). Changes in Cognitive Function and Risk Factors for Cognitive Impairment of the Elderly in China: 2005-2014. Int. J. Environ. Res. Public Health 16:2847. doi: 10.3390/ijerph16162847
Zhang, Y., and Ren, J. (2011). ALDH2 in alcoholic heart diseases: molecular mechanism and clinical implications. Pharmacol. Ther. 132, 86–95. doi: 10.1016/j.pharmthera.2011.05.008
Keywords: ethanol intake, Asian flush, alcohol metabolizing genes, Alzheimer’s disease, cognitive impairment
Citation: Jin X, Long T, Chen H, Zeng Y, Zhang X, Yan L and Wu C (2022) Associations of Alcohol Dehydrogenase and Aldehyde Dehydrogenase Polymorphism With Cognitive Impairment Among the Oldest-Old in China. Front. Aging Neurosci. 13:710966. doi: 10.3389/fnagi.2021.710966
Received: 17 May 2021; Accepted: 28 December 2021;
Published: 25 February 2022.
Edited by:
Wael M. Y. Mohamed, International Islamic University Malaysia, MalaysiaReviewed by:
Zhixiong Sun, Columbia University, United StatesCopyright © 2022 Jin, Long, Chen, Zeng, Zhang, Yan and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lijing Yan, bGlqaW5nLnlhbkBkdWtla3Vuc2hhbi5lZHUuY24=; Chenkai Wu, Y2hlbmthaS53dUBkdWtla3Vuc2hhbi5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.